 Ellen Rose Jacobs
Ellen Rose Jacobs Rafaela Landea-Briones2
Rafaela Landea-Briones2 Laela Sayigh
Laela Sayigh
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Remote Sens. , 30 August 2022
Sec. Acoustic Remote Sensing
Volume 3 - 2022 | https://doi.org/10.3389/frsen.2022.909309
This article is part of the Research Topic Acoustic Remote Sensing of Cetacean and Pinniped Populations View all 13 articles
Eastern South Pacific southern right whales (ESPSRW) are a subpopulation of southern right whales (Eubalaena australis) off the coasts of Peru and Chile recognized by the International Union for the Conservation of Nature (IUCN) as critically endangered as a result of heavy whaling efforts in the late 18th to 20th centuries. Most recent population estimates put their numbers around 50 individuals. To test for the efficacy of passive acoustic monitoring of this population, we recorded 5 months of continuous acoustic data (January 2012-June 2012) off the southwestern tip of Isla de Chiloé. To test for trends in occurrence, we identified 11,313 individual ESPSRW upsweep calls, which have been associated with maintaining contact with conspecifics. Call occurrence increased over the course of the deployment and peaked between April and June, indicating an increase in use of this area. A clear diel pattern in which upsweep calls were predominately detected during dusk and night hours was identified, indicating ESPSRW are likely foraging during daylight hours, as upsweep calls are inversely related to foraging behavior. We quantified noise levels in the frequency range of their communication (100 Hz third octave) to understand the change in active space whales may be experiencing. We measured noise levels from 90 dB re 1 μPa to 111 dB re 1 µPa (5th and 95th percentile), a 21 dB fluctuation that results in an order-of-magnitude decrease in active space area. We identified sources of high noise at or above the 75th percentile as predominately blue whale calls (occurring in 71.6% of total sampled minutes) and ship noise (occurring in 69.4% of total sampled minutes). Ship noise was responsible for outliers in excess of 140 dB re 1 µPa. In a population as diminished as ESPSRW, such disruptions of their communication range could result in significant barriers to maintaining contact with conspecifics. Passive acoustic monitoring is a powerful tool for monitoring populations as rarely sighted as ESPSRW. Understanding trends in presence and behavior as well as potential sources of disruption to their calling behavior is vital to determining conservation measures that will be most effective toward helping this critically endangered population.
Eastern South Pacific southern right whales (Eubalaena australis; henceforth, ESPSRW) are a subpopulation off the coasts of Peru and Chile recognized by the International Union for the Conservation of Nature (IUCN) as critically endangered. One population estimate places their numbers at no more than 50 individuals, with the number of reproductive females as low as eight as of 2014 (Galletti Vernazzani et al., 2014). ESPSRW are thought to be distinct from other nearby populations of southern right whales and do not show the same signs of recovery from heavy whaling efforts of the late 18th to 20th centuries as do these other populations (Galletti Vernazzani et al., 2014). Known to range from the Chilean Golfo de Penas, at 47°S, to the coast of Chorrillos in Lima, Peru, at 12°S (Aguayo and Torres, 1986; Orihuela and Cortegana-Arias, 2013), habitat use within that range remains largely unknown. We used passive acoustic monitoring to gain information about the presence and behavior of ESPSRW in this area, while at the same time examining changes in active space that could affect their communication patterns. Our goal was to better understand why there have been fewer than 200 total sightings of this population since the early 1960s (Galletti Vernazzani et al., 2014). Documenting and monitoring their presence and movement patterns is vital for promoting population recovery through conservation efforts.
The most common type of vocalization in the southern right whale repertoire is thought to be the upsweep call, which is most likely used to keep in contact with conspecifics (Clark, 1982), and has been used in passive acoustic monitoring of North Atlantic right whales (Eubalaena glacialis) (Mussoline et al., 2012). Knowledge of trends in calling behavior has direct application to conservation efforts, such as imposing a seasonal speed limit on the East Coast of the United States to minimize ship strikes during times that North Atlantic right whales are known to be in an area (Laist et al., 2014), as well as providing critical insight into broad-scale movements and abundance in North Pacific right whales (Eubalaena japonica) (Munger et al., 2008, 2011).
Prior to this study, it was unknown whether ESPSRW calls exhibited diel patterning. Some cetacean species are known to call more at night, such as those who call to maintain contact with conspecifics when visual cues are not available (common dolphins, Delphinus delphis: Goold, 2000; blue whales, Balaenoptera musculus: Redaelli et al., 2022; Wiggins et al., 2005), while others are known to call more during daylight hours, such as those who socialize during the day and feed during night hours (sei whales, Balaenoptera borealis: Baumgartner and Fratantoni, 2008).
On a larger temporal scale, seasonal patterning of calls can provide insight into habitat usage. Many cetacean species exhibit migrations, sometimes up to thousands of kilometers (Mate et al., 1999; Rugh et al., 2001; Rasmussen et al., 2007; Olsen et al., 2009; Stevick et al., 2011), so a habitat area might only be in use by a species at certain times of the year.
Another aspect of ESPSRW communication that is not well understood is the active space of calls—the effective range over which a communication signal can be detected by a conspecific (Marten and Marler, 1977). The active space of a given call depends on call parameters such as frequency and source level, but also the background noise through which a call travels, the extent of sound attenuation over distance, and the critical ratio of signal-to-noise necessary for a sound to be audible to an individual. Changes in background noise can have significant effects on the active space of a call, leading to order-of-magnitude changes in the distance at which a call is audible to conspecifics (e.g., Janik, 2000). Detection of an acoustic signal is dependent on the difference between the source level of the signal and the ambient noise levels in that frequency band, so increasing noise will decrease the chance of signal detection and potentially mask the signal from the receiver (Richardson, et al., 1995; Bradbury and Vehrencamp, 1998). In a population as diminished as ESPSRW, changes to active space due to variation in background noise, particularly that attributable to anthropogenic sources such as ships (Jensen et al., 2009), could have implications for the ability of individuals to communicate, as has been documented in other cetacean species (Richardson, et al., 1995; Nowacek et al., 2007).
Jacobs et al. (2019) described the first known acoustic detections of ESPSRW upsweep calls, defining parameters of both individual calls and call bouts based on a small subset of continuous acoustic recordings. The current study built upon these data to identify temporal trends in upsweep call use both on diel and deployment-long (5 months) time scales. We then characterized the ambient noise environment of ESPSRW by quantifying noise levels in the frequency range of their calls and identifying sources of that noise. Understanding temporal trends in call production can provide important insight into the behavior of ESPSRW, allowing for more targeted conservation efforts.
Five months of continuous recordings at a sampling rate of 2 kHz were made with a Marine Autonomous Recording Unit (MARU; Cornell University) from 29 January 2012 to 17 June 2012. The MARU was placed offshore of the southwestern tip of Chiloé Island in the Corcovado Gulf, part of the Chiloense ecoregion (43.53°S, 74.44°W; Figure 1). The Chiloense Ecoregion receives both continental freshwater and oceanic subantarctic water, creating a rich ecosystem with high levels of primary productivity and calanoid copepods, which are potential prey species for right whales (Arcos, 1974; Baumgartner et al., 2011).
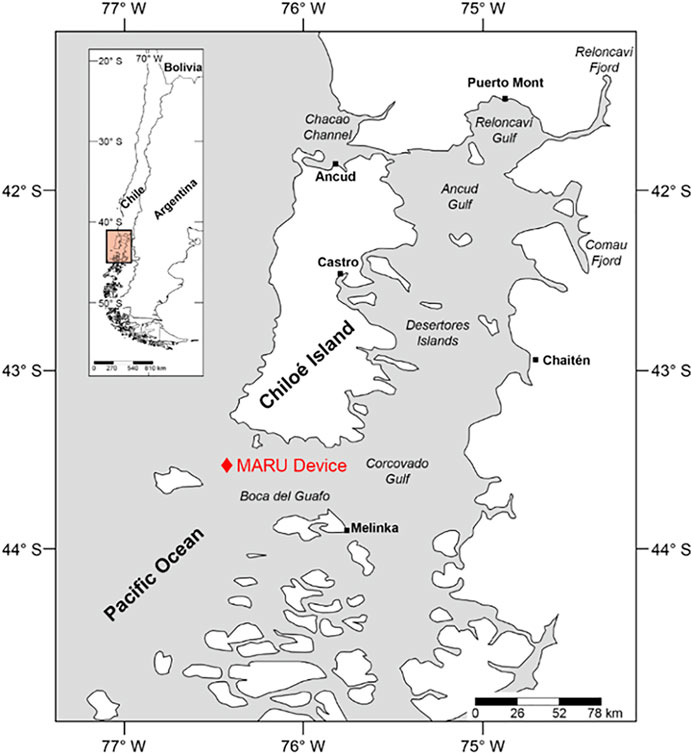
FIGURE 1. Map of Chiloé Island and the Corcovado Gulf in southwestern Chile. Red diamond indicates location of the MARU device (43.53°S, 74.44°W), deployed from 29 January to 17 June 2012.
The full dataset was visually and aurally audited for presence of right whale upsweep calls using Raven Pro 1.5 with the following display settings: 3 min time axis, 500 Hz frequency axis, 2,048 sample DFT, 1,400 sample window, and 50% overlap. A total of 11,313 individual upsweep calls were identified and exported for further analysis. Temporal trends in call detection were analyzed using R version 3.4.1 (R: R Core Team, 2020; tidyverse package: Wickham and RStudio, 2021 gridExtra package: Auguie and Antonov, 2017; viridis package: Garnier et al., 2021; ggpubr package: Kassambara, 2020).
Seasonal trends were not possible to evaluate due to the relatively short duration of the deployment. However, to look at changes in call detection over the course of the deployment, total number of calls per week was calculated and qualitatively compared over the 20 weeks of deployment.
In order to quantify the relationship between call production rates and light regime, call times were classified by light regime (dawn, day, dusk, or night as determined from daily sunrise, sunset, and nautical twilight times according to the United States Naval Observatory). On average, the dawn period lasted 1.10 (SD ± 0.04) hours, day lasted 10.52 (SD ± 1.27) hours, dusk lasted 1.08 (SD ± 0.08) hours, and night lasted 11.33 (SD ± 1.28) hours. Clock time of the start of each light phase shifted by about 3 hours over the course of the deployment (e.g., night began at 01:36 UTC on 02-02-2012 and at 22:36 UTC on 15-06-2012).
To correct for variability in call presence over time scales greater than a single day and make distribution of calls between light periods comparable between days with different overall call production rates, hourly call rates were adjusted by dividing the average hourly call rate within a light phase by the hourly call rate within that 24-h period. Higher relative call rates indicate that a larger shift in the calling rate occurred during a given light phase. This method incorporates changes in the absolute number of calls while taking into consideration variable call rates on each day.
Ambient noise levels in the recordings were determined using a Third Octave Level analysis (TOL) in MATLAB version 2017b (MATLAB, 2017). TOLs were calculated as the RMS level over 1 minute for each third octave band over the course of the deployment (197,280 min) using a custom MATLAB script. Further analysis was done on the 100 Hz frequency band, which was identified as most relevant for the noise environment of ESPSRW, given that their upsweep calls range from 68 ± 17 Hz to 136 ± 25 Hz (Jacobs et al., 2019). 5th and 95th percentile noise levels were calculated to determine the communication range reduction experienced by ESPSRW in this area due to fluctuations in background noise.
In order to identify the loudest sources of noise in the 100 Hz frequency band, minutes were sorted from lowest to highest noise level. The noise level at the 75th percentile was identified, and minutes were then randomly subsampled to include 10% of the data at or above the 75th percentile (4,932 min subsampled from 49,320 min). These periods are hereafter referred to as “high noise.” The source of noise for each of those minutes was identified by visual inspection of spectrograms in MATLAB by a skilled observer, with the same spectrogram display settings as those used in Raven.
Noise sources were classified as either ship, blue or humpback whale, a combination of ship and blue or humpback whale occurring concurrently, or environmental sources such as rain. Percentages of the total subsample containing each noise source and their average received levels were calculated to characterize the most common sources of loud noise and determine the relative importance of each source in the soundscape experienced by ESPSRW. These minutes of high noise were then classified by the time they occurred into light regime categories using the same method as for call rates, so that the proportional amount of each light phase taken up by high noise could be calculated and the importance of each source in each light phase evaluated.
On a deployment-wide scale, there was a general upward trend in number of detections from beginning to end of the deployment, peaking between April and June (late fall to early winter; Figure 2A). Relative call production rate was significantly higher at dusk than dawn, day, or night (Figure 2B; Kruskal Wallis H = 130.22, df = 3, p = < 1e-10, pairwise-comparison test p = 8.7e-07, < 1e-10, and < 1e-10 for dusk with dawn, day, and night, respectively). Compared to an even 24-h distribution of calls, call production rates were lower than would be expected at dawn and daytime and were approximately equal to what would be expected at nighttime (Figure 2B). This trend of higher call production rates during dusk and lower call rates during the dawn and day was consistent over the course of the deployment.
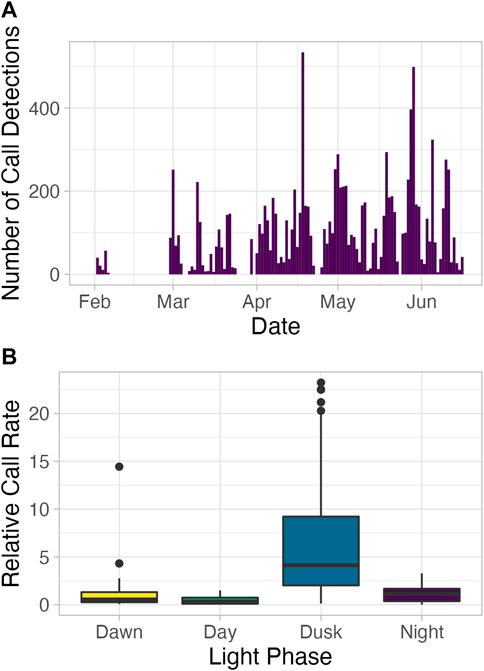
FIGURE 2. (A) Total call detections per day between 02-02-2012 and 16-06-2012. No calls were recorded between 29-01-2012, the day on which the MARU was deployed, and 02-02-2012. (B) Relative call rate by light phase. Relative call rates were calculated by dividing the light phase’s average hourly call rate by the overall day’s hourly call rate, so a higher value indicates a larger shift in the calling rate occurred in a light phase, while taking into consideration variable call rates over the course of the deployment.
TOLs calculated in the 100 Hz band for the full recording ranged from 90 dB re 1 μPa to 111 dB re 1 µPa (5th and 95th percentile), with a value of 105 dB re 1 μPa at the 75th percentile (Figure 3A). Combining this fluctuation of up to 21 dB in the noise environment over the course of the deployment with an assumption of spherical spreading, the degree to which the communication range is reduced can be calculated using Equation 1, a modification of transmission loss in the passive sonar equation (Møhl, 1981a; Urick, 1983; Jensen et al., 2009):
Where TOLhigh and TOLlow are the TOLs at the highest and lowest noise conditions, respectively. This results in an active space that is (10(111-90)/20 = ) 11 times smaller at the highest versus lowest noise conditions. Between the 25th and 75th percentiles of noise, there was a fluctuation of about 8 dB (96 dB re 1µPa to 104 dB re 1 µPa) in the 100 Hz band, resulting in an active space (10(104-96)/20 = ) 2.5 times smaller at the 75th percentile than at the 25th percentile level.
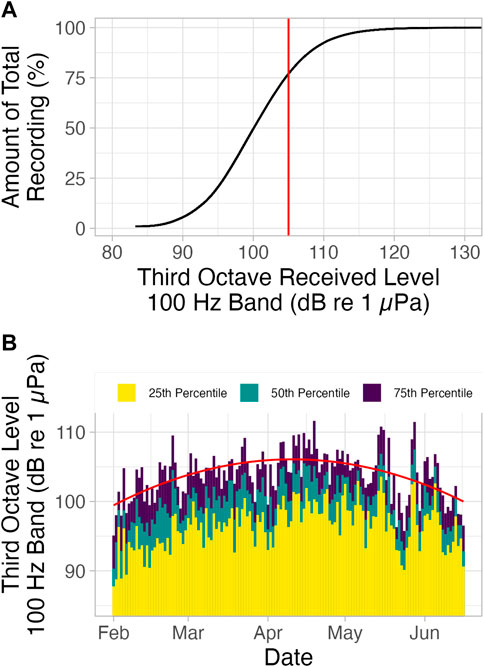
FIGURE 3. (A) Cumulative third octave received levels calculated over the full deployment in the 100 Hz band. 75% of the total third octave received levels were less than 105 dB re 1 µPa (75th percentile; indicated by red line) and all third octave received levels above this level were regarded as high noise. (B) Third octave received levels in the 100 Hz band by day, 02-02-2012 to 16-06-2012. Yellow indicates each day’s 25th percentile noise levels, green indicates 50th percentile noise levels, and purple indicates 75th percentile noise levels. Red trend line is fitted over 75th percentile noise levels.
Noise levels gradually increased from February to April, then gradually decreased from May to June (Figure 3B). Interestingly, this roughly corresponds with trends in right whale upsweep call detection (Figure 2A).
Predominant sources of noise in the high noise subsample (above the 75th percentile) were ships (21.4%), blue and humpback whales (23.6%), a combination of ships and blue and humpback whales (48.0%), and other environmental sources (6.9%; Figure 4A). Thus, ship noise was present in 69.4% of this high noise subsample. Similarly, blue and humpback whale calls were present in the communication band of right whales in 71.6% of this subsample. Although some of the large whale noise was from humpbacks, the vast majority came from blue whale calls.
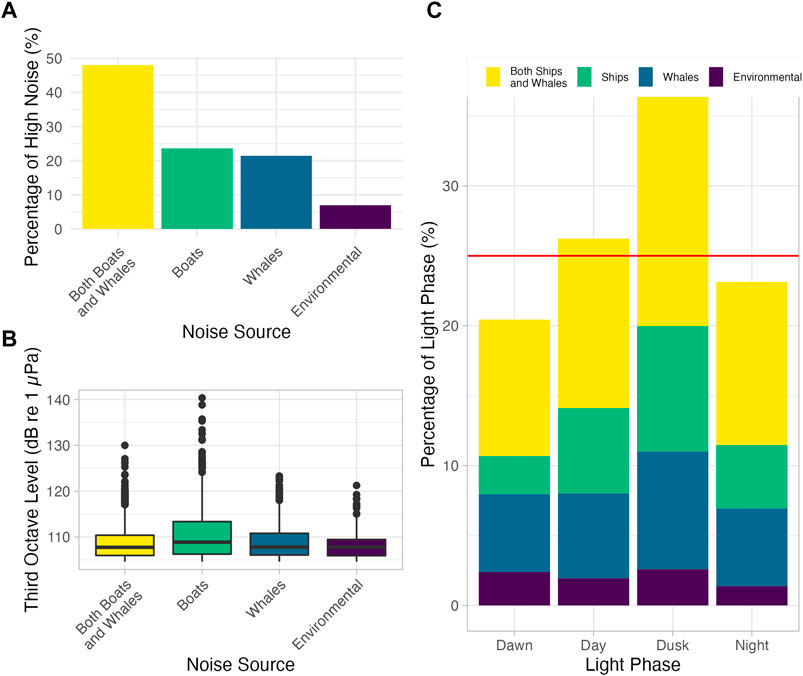
FIGURE 4. Sources of high noise. (A) Percentage of high noise (TOL above the 75th percentile, 105 dB re 1 µPa) produced by each source. Yellow indicates both ships and whales, green indicates ships, blue indicates whales, and purple indicates other environmental noise sources, such as rain. Ships and whales were present in the majority of high noise minutes. (B) TOLs of each source of high noise. Minutes containing ship noise contained the highest outliers, with some minutes over 140 dB re 1 µPa. (C) Percentage of each light phase containing high noise. Total height of each bar indicates the percentage of each light phase that contains high noise. Shading in each bar indicates the percentage of total high noise minutes that during that phase from each source. Red line at 25% indicates the level of noise expected if high noise was evenly distributed over each phase. Values greater than 25% indicate disproportionate levels of high noise, such as occurs during day and dusk, whereas lower amounts of high noise were seen during dawn and night. Ship noise and combined ship and whale noise occurred most frequently during dusk.
Median values of each category of high noise were found to be significantly different (Kruskal Wallis p = 9.03e-18; Figure 4B), with pairwise comparisons showing that only whale noise and the combination of whale and ship noise were not different (Wilcoxon pairwise comparison test p = 0.20, all others less than 0.05). Ship noise contained higher outliers than the other categories, with values in excess of 140 dB re 1 μPa; in some instances, high ship noise may have masked blue whale calls, biasing high noise towards ships as the source when blue whales may have been present.
In some light phases, a disproportionate amount of the recorded noise was classified as high noise. Of the total noise recorded during each light phase, 26% during the day and 37% during dusk were classified as high noise, whereas only 20% and 23% were classified as high noise during dawn and night respectively (Figure 4C). The level of noise expected if high noise was evenly distributed over each phase is 25%. While environmental and whale noise remained largely constant over the diel cycle, ship noise and both ship and whale noise were disproportionally present at higher levels during day and dusk than at dawn or night.
The ESPSRW subpopulation is thought to be resident to the lower South American coast, ranging from Peru to Chile, but habitat use within that range remains unclear due to the paucity of sightings (Galletti Vernazzani et al., 2014). In this study, we sought to understand trends in the detection of right whale upsweep calls and the ambient noise conditions in which they occur using passive acoustic monitoring data from the Corcovado Gulf. We found a gradual increase in call detections over the course of the February to June deployment, with a higher proportion of calls during dusk compared to dawn, day or night hours. Using a third octave level analysis, we measured ambient noise in the 100 Hz frequency band and determined that the 75th percentile noise level is 105 dB re 1 µPa. We then identified sources of noise in the upper 75th percentile range, and found that the majority of these high noise levels come from ships, with blue whale calls also contributing strongly to the ambient noise conditions.
Upsweep call detection rates increased over the course of the deployment, suggesting an increase in usage of this habitat by ESPSRW in late fall to early winter. Further study of this area should include year-round acoustic monitoring to determine the full duration of ESPSRW presence beyond this study’s 5-month deployment. However, knowledge that there is an increased presence of ESPSRW in the late fall to early winter could begin to inform conservation efforts.
Our observed trend of lower calling rates at dawn and daytime and higher rates at dusk is consistent with trends found for North Atlantic right whales (Mussoline et al., 2012), which are known to forage during the day, taking advantage of surface aggregations of Calanus finmarchicus (Parks et al., 2012) or deeper layers below a depth of 90 m (Baumgartner and Mate, 2003). During this active foraging time they do not vocalize (Parks et al., 2012). Diel trends in ESPSRW upsweep calling rate were also consistent with diel patterns of ‘D’ call production by sympatric Chilean blue whales (Balaenoptera musculus; Redaelli et al., 2022). Redaelli et al. (2022) found that D calls are more numerous during dusk and night than dawn or daytime hours, suggesting that the whales might be foraging at depth for denser aggregations of prey during daylight hours and producing D calls to maintain contact with conspecifics more at dusk and night.
Upsweep calls are produced mainly during twilight hours when maintaining visual contact with conspecifics may become difficult (Mussoline et al., 2012). If the observed lower call rates during dawn and daylight hours indicate that ESPSRW are spending daylight hours foraging near the surface, this could put them at increased risk of ship strike, as has been documented in North Atlantic right whales (Parks et al., 2012). This could even be a factor in the population’s inability to recover from decimation by the whaling industry.
Clark (1982) proposed that, in addition to serving as contact calls, upsweep calls were related to changes in behavioral state or when individuals join groups. After foraging during the day, whales may use upsweep calls during dusk to locate other individuals to transition to their night-time behavioral states. In small populations like ESPSRW, sparse population density could make locating conspecifics difficult. Visual contact may be even harder to maintain at dusk and during the night, so they may increasingly resort to acoustic contact to locate conspecifics.
As with any passive acoustic monitoring study, it is impossible to know whether increased numbers of calls over seasonal or diel scales come from individual whales calling more often or an increased number of calling whales, and likewise whether an absence of calls indicates an absence of vocal behavior or of whales. Previous studies comparing visual and acoustic detections of North Atlantic right whales found that vocal behavior remains consistent across time scales greater than a day (Mussoline et al., 2012), so we believe that the increasing number of calls found in later weeks of the deployment is indicative of more whales rather than an increase in calling behavior. Conversely, we assume that habitat usage remains uniform on a diel scale, so an increasing number of calls in later hours of the day indicates more vocal behavior from individual whales rather than small-scale changes in habitat use (Clark et al., 2010).
We found that changes in ambient noise in this region have the potential to greatly influence the active space of ESPSRW communication, with an order-of-magnitude change in active space between the quietest and loudest (5th and 95th percentile) ambient noise conditions at 100 Hz. Even between the 25th and 75th percentiles, the most common noise levels experienced by ESPSRW, the active space decreases by a factor of 2.5. This indicates that ESPSRW regularly experience large differences in the detectability of their calls. In a population as reduced as ESPSRW, such reductions in active space could severely jeopardize their ability to communicate with one another, decreasing the effectiveness of upsweeps as contact calls and inhibiting social activities that increase fitness. For example, even if a call is still detectable, the information content might not be properly conveyed (Jensen et al., 2009) if some features are masked by noise.
In other populations of right whales, changes in call production such as decreases in call rate have been suggested as responses to high noise from vessel traffic (Parks et al., 2007). While our study is limited by the constraints of passive acoustic data collection, it is possible that ESPSRW also exhibit behavioral responses to high noise. In our dataset, ship noise may have masked any right whale calls made during high noise periods, precluding our ability to evaluate changes in call production rate in response to high noise. Right whales have also been known to exhibit a Lombard response, increasing the amplitude of their calls in response to increased amplitude of background noise, potentially counteracting the reduction in active space in moderate noise conditions (Parks et al., 2010). However, our estimated 11-fold range reduction in high noise conditions still provides a good estimate of the degree to which the active space of these whales may be reduced by noise (Møhl, 1981b; Clark et al., 2009; Jensen et al., 2009; Hermannsen et al., 2014), regardless of any behavioral responses to mitigate masking effects. Even at the noise levels in the middle of the range, there is a reduction in active space that likely decreases their ability to communicate effectively.
Ship noise is a major contributor to the ambient noise experienced by ESPSRW. Unlike blue whale calls, ship noise is relatively new in the habitat. Ship noises encompass a wider range of frequencies and are longer lasting, so they are likely to have a more significant effect on the acoustic environment than blue whale calls (Nowacek et al., 2007). While many blue whales calling at once can be a significant addition to the acoustic environment, ship noise typically occurs without pause for greater durations. Brief pauses in loud blue whale calls may provide an opportunity for right whales to communicate, whereas pauses are less likely in persistent ship noise.
Increased levels of ship noise during day and dusk could be due to fishing activity during those light phases, with ships traveling through the area where the recording device was located during the day and potentially returning to shore at dusk. In an analysis of a larger data set from the Chiloense ecoregion that included the data analyzed for this study, Redaelli et al. (2022) found that blue whale D calls were produced primarily in dusk and night hours while songs (SEP calls) did not exhibit diel patterns. In our dataset, SEP calls were extremely common in minutes classified as high noise. With no diel pattern found for SEP calls, ships may then be the main driver of the diel pattern in the “Both Ships and Whales” noise source category. While high noise occurred most frequently during both day and dusk, right whale upsweeps occurred most frequently during dusk and night. This suggests that trends in call production are likely influenced more by behavior than by the ambient noise environment.
Ship noise is known to cause an increase in stress in North Atlantic right whales (Rolland et al., 2012). Ships accounted for 69% percent of high noise in our data set, corresponding to 17% of the overall recording time. Thus, ship noise has the potential to cause long-term negative effects on the overall health of the population. Additionally, as higher incidence of ship noise, and therefore vessel traffic, occurs during the day when ESPSRW are likely foraging near the surface, the probability of ship-strike is even greater than if whales were spending time at the surface during periods of low vessel activity. This could have significant consequences on population recovery depending on the role this habitat plays in the life histories of this population of right whales. Mother-calf pairs were identified in multiple sightings in this area, suggesting it may be used as a nursing area (Galletti Vernazzani et al., 2014), and thus high levels of stress or communication obstruction might endanger calf survival. In other populations of right whales, mother-calf pairs engage in acoustic crypsis, reducing the amplitude of communication calls and decreasing the use of long-range calls to potentially avoid detection by predators (Nielsen et al., 2019; Parks et al., 2019). Further decreases in active space due to ship noise could thus jeopardize mother-calf communication, either by masking calls or necessitating the use of higher amplitude calls that may render the pair more vulnerable to predation. In a population suffering from low population growth, the loss of even one calf can have large consequences for the population.
Our analysis is among the first to provide insight into the occurrence, vocal patterns, and communication space of ESPSRW. Knowledge of diel and seasonal trends can direct conservation efforts towards times of day and year that the habitat is in use by the population, and an understanding of the anthropogenic contributors to ambient noise levels can inform effective mitigation measures. For example, vessel speed limits could be implemented to reduce ship strikes, as has been done for North Atlantic right whales (Laist et al., 2014). Although it is unlikely that ship strikes are as frequent with this population as with their North Atlantic cousins (Kraus et al., 2005), some instances of ship strike deaths have been documented for this population (Galletti Vernazzani et al., 2014). In addition, a speed limit would decrease the magnitude of ship noise (Hermannsen et al., 2014) and thus likely also reduce stress levels and increase communication space. Overall, our study is a step toward gaining a better understanding of the acoustic environment of this critically endangered population of whales.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: WHOAS repository; Accessible at https://hdl.handle.net/1912/28280.
Ethical review and approval was not required for the animal study because passive acoustic monitoring is non-invasive and did not require direct contact with the research subjects.
ERJ and LS designed the study. LS supervised and administrated the project and curated the data. RL-B participated in and provided resources and funding for the field work expeditions. ERJ conducted the analyses and wrote the paper. All authors edited the manuscript and gave approval for submission.
Financial support for expeditions, deployments, and retrievals of MARUs, and for some data analysis, was provided by Fundacion MERI, Av. Pdte. Kennedy 5682, Vitacure, Región Metropolitana, Chile. Most data analysis for this study was carried out without external funding.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Thanks to Rodrigo Hucke-Gaete, Susannah Buchan, and the Centro Ballena Azul for their participation in the expeditions to deploy MARUs, and to Fundacion MERI for providing financial support for the deployments, as well as for some data analysis. We also thank Thomas Montt, Sociedad Pesquera y de Turismo Marítimo Los Elefantes Ltd. a., Pedro Montt 215, Dalcahue—Chiloe, Chile, for his support (and that of his crew) in cruise logistics, and for deploying and retrieving instruments, and Barbara Galletti for supplying visual sighting data that confirmed our identification of the sounds. We would also like to thank Peter T. Madsen of Aarhus University for his support and feedback on the ambient noise analyses and financial contribution to the publication cost, and Alessandro Bocconcelli of Woods Hole Oceanographic Institution for his financial and logistical contributions to the project.
Aguayo, A., and Torres, D. (1986). “Records of the Southern Right Whale, Eubalaena australis (Desmoulins 1822) from Chile between 1976 and 1982,” in Report of the International Whaling Commission, Special Issue, 10, 159–160.
Arcos, D. (1974). Los copépodos calanoídeos colectados en la región Magallánica por la Expedición Hero. Bol. La Soc. Biol. Concepción 47 (215–225), 72–4b.
Auguie, B., and Antonov, A. (2017). gridExtra: Miscellaneous Functions for “Grid” Graphics (2.3). [Computer software]. Available at: https://CRAN.R-project.org/package=gridExtra.
Baumgartner, M. F., and Fratantoni, D. M. (2008). Diel Periodicity in Both Sei Whale Vocalization Rates and the Vertical Migration of Their Copepod Prey Observed from Ocean Gliders. Limnol. Oceanogr. 53 (5part2), 2197–2209. doi:10.4319/lo.2008.53.5_part_2.2197
Baumgartner, M., Lysiak, N., Schuman, C., Urban-Rich, J., and Wenzel, F. (2011). Diel Vertical Migration Behavior of Calanus finmarchicus and its Influence on Right and Sei Whale Occurrence. Mar. Ecol. Prog. Ser. 423, 167–184. doi:10.3354/meps08931
Baumgartner, M., and Mate, B. (2003). Summertime Foraging Ecology of North Atlantic Right Whales. Mar. Ecol. Prog. Ser. 264 (3), 123–135. 10.1109/JOE.2003.813943. doi:10.3354/meps264123
Bradbury, J. W., and Vehrencamp, S. L. (1998). Principles of Animal Communication. Sunderland, Massachusetts: Sinauer.
Clark, C., Ellison, W., Southall, B., Hatch, L., Van Parijs, S., Frankel, A., et al. (2009). Acoustic Masking in Marine Ecosystems: Intuitions, Analysis, and Implication. Mar. Ecol. Prog. Ser. 395, 201–222. doi:10.3354/meps08402
Clark, C. W., Brown, M. W., and Corkeron, P. (2010). Visual and Acoustic Surveys for North Atlantic Right Whales, Eubalaena Glacialis, in Cape Cod Bay, Massachusetts, 2001-2005: Management Implications. Mar. Mammal Sci. 26 (4), 837–854. doi:10.1111/j.1748-7692.2010.00376.x
Clark, C. W. (1982). The Acoustic Repertoire of the Southern Right Whale, a Quantitative Analysis. Anim. Behav. 30 (4), 1060–1071. doi:10.1016/S0003-3472(82)80196-6
Garnier, S., Ross, N., Rudis, B., Sciaini, M., Camargo, A. P., and Scherer, C. (2021). Viridis: Colorblind-Friendly Color Maps for R (0.6.2). [Computer Software]. Available at: https://CRAN.R-project.org/package=viridis.
Goold, J. C. (2000). A Diel Pattern in Vocal Activity of Short-Beaked Common Dolphins, Delphinus delphis. Mar. Mammal. Sci. 16 (1), 240–244. doi:10.1111/j.1748-7692.2000.tb00915.x
Hermannsen, L., Beedholm, K., Tougaard, J., and Madsen, P. T. (2014). High Frequency Components of Ship Noise in Shallow Water with a Discussion of Implications for Harbor Porpoises (Phocoena Phocoena). J. Acoust. Soc. Am. 136 (4), 1640–1653. doi:10.1121/1.4893908
Jacobs, E., Duffy, M., Magolan, J., Galletti Vernazzani, B., Cabrera, E., Landea, R., et al. (2019). First Acoustic Recordings of Critically Endangered Eastern South Pacific Southern Right Whales (Eubalaena australis). Mar. Mam. Sci. 35 (1), 284–289. doi:10.1111/mms.12519
Janik, V. M. (2000). Source Levels and the Estimated Active Space of Bottlenose Dolphin (Tursiops truncatus) Whistles in the Moray Firth, Scotland. J. Comp. Physiol. A 186 (7–8), 673–680. doi:10.1007/s003590000120
Jensen, F., Bejder, L., Wahlberg, M., Aguilar de Soto, N., Johnson, M., and Madsen, P. (2009). Vessel Noise Effects on Delphinid Communication. Mar. Ecol. Prog. Ser. 395, 161–175. (Ross 1976). doi:10.3354/meps08204
Kassambara, A. (2020). Ggpubr: “Ggplot2” Based Publication Ready Plots (0.4.0). [Computer Software]. Available at: https://CRAN.R-project.org/package=ggpubr.
Kraus, S. D., Brown, M. W., Caswell, H., Clark, C. W., Fujiwara, M., Hamilton, P. K., et al. (2005). North Atlantic Right Whales in Crisis. Science 309, 561–562. doi:10.1126/science.1111200
Laist, D., Knowlton, A., and Pendleton, D. (2014). Effectiveness of Mandatory Vessel Speed Limits for Protecting North Atlantic Right Whales. Endang. Species. Res. 23 (2), 133–147. doi:10.3354/esr00586
Marten, K., and Marler, P. (1977). Sound Transmission and its Significance for Animal Vocalization. Behav. Ecol. Sociobiol. 2, 271–290. doi:10.1007/bf00299740
Mate, B. R., Lagerquist, B. A., and Calambokidis, J. (1999). Movements of North Pacific Blue Whales during the Feeding Season off Southern California and Their Southern Fall Migration1. Mar. Mammal. Sci. 15 (4), 1246–1257. doi:10.1111/j.1748-7692.1999.tb00888.x
Møhl, B. (1981a). “Masking Effects of Noise: Their Distribution in Time and Space,” in Proceedings from a Workshop on: The Question of Sound from Icebreaker Operations, 1981. Feb 23-24.
Møhl, B. (1981b). “Masking Effects of Noise: Their Distribution in Time and Space,” in The Question of Sound from Icebreaker Operations.
Munger, L. M., Wiggins, S. M., and Hildebrand, J. A. (2011). North Pacific Right Whale Up-Call Source Levels and Propagation Distance on the Southeastern Bering Sea Shelf. J. Acoust. Soc. Am. 129 (6), 4047–4054. doi:10.1121/1.3557060
Munger, L. M., Wiggins, S. M., Moore, S. E., and Hildebrand, J. A. (2008). North Pacific Right Whale (Eubalaena Japonica) Seasonal and Diel Calling Patterns from Long-Term Acoustic Recordings in the Southeastern Bering Sea, 2000-2006. Mar. Mammal Sci. 24, 795–814. doi:10.1111/j.1748-7692.2008.00219.x
Mussoline, S., Risch, D., Clark, C., Hatch, L., Weinrich, M., Wiley, D., et al. (2012). Seasonal and Diel Variation in North Atlantic Right Whale Up-Calls: Implications for Management and Conservation in the Northwestern Atlantic Ocean. Endang. Species. Res. 17 (1), 17–26. doi:10.3354/esr00411
Nielsen, M. L. K., Bejder, L., Videsen, S. K. A., Christiansen, F., and Madsen, P. T. (2019). Acoustic Crypsis in Southern Right Whale Mother-Calf Pairs: Infrequent, Low-Output Calls to Avoid Predation? J. Exp. Biol. 222 (13), jeb190728. doi:10.1242/jeb.190728
Nowacek, D. P., Thorne, L. H., Johnston, D. W., and Tyack, P. L. (2007). Responses of Cetaceans to Anthropogenic Noise. Mammal. Rev. 37 (2), 81–115. doi:10.1111/j.1365-2907.2007.00104.x
Olsen, E., Budgell, W. P., Head, E., Kleivane, L., Nøttestad, L., Prieto, R., et al. (2009). First Satellite-Tracked Long-Distance Movement of a Sei Whale (Balaenoptera Borealis) in the North Atlantic. Aquat. Mamm. 35 (3), 313–318. doi:10.1578/AM.35.3.2009.313
Orihuela, M., and Cortegana-Arias, D. (2013). Registro de un par madre-cría de ballena franca austral (Eubalaena australis) en la costa de Lima, Perú [Record of mother-calf pair of southern right whales (Eubalaena australis) off the coast of Lima, Peru]. Rev. Peru. Biol. 20 (2), 187–188. doi:10.15381/rpb.v20i2.2685
Parks, S. E., Clark, C. W., and Tyack, P. L. (2007). Short- and Long-Term Changes in Right Whale Calling Behavior: The Potential Effects of Noise on Acoustic Communication. J. Acoust. Soc. Am. 122 (6), 3725–3731. doi:10.1121/1.2799904
Parks, S. E., Johnson, M., Nowacek, D., and Tyack, P. L. (2011). Individual Right Whales Call Louder in Increased Environmental Noise. Biol. Lett. 7 (1), 33-35. doi:10.1098/rsbl.2010.0451
Parks, S. E., Cusano, D. A., Van Parijs, S. M., and Nowacek, D. P. (2019). Acoustic Crypsis in Communication by North Atlantic Right Whale Mother-Calf Pairs on the Calving Grounds. Biol. Lett. 15 (10), 20190485. doi:10.1098/rsbl.2019.0485
Parks, S. E., Warren, J. D., Stamieszkin, K., Mayo, C. A., and Wiley, D. (2012). Dangerous Dining: Surface Foraging of North Atlantic Right Whales Increases Risk of Vessel Collisions. Biol. Lett. 8 (1), 57–60. doi:10.1098/rsbl.2011.0578
R Core Team (2020). R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing. Available at: https://www.R-project.org/.
Rasmussen, K., Palacios, D. M., Calambokidis, J., Saborío, M. T., Dalla Rosa, L., Secchi, E. R., et al. (2007). Southern Hemisphere Humpback Whales Wintering off Central America: Insights from Water Temperature into the Longest Mammalian Migration. Biol. Lett. 3 (3), 302–305. doi:10.1098/rsbl.2007.0067
Redaelli, L., Mangia Woods, S., Landea, R., and Sayigh, L. (2022). Seasonal Trends and Diel Patterns of Downsweep and SEP Calls in Chilean Blue Whales. J. Mar. Sci. Eng. 10 (3), 316. doi:10.3390/jmse10030316
Richardson, W. J., Greene, C. R., Malme, C. I., and Thomson, D. H. (1995). “Marine Mammal Sounds,” in Marine Mammals and Noise (Academic Press), 159–204. doi:10.1016/b978-0-08-057303-8.50010-0
Rolland, R. M., Parks, S. E., Hunt, K. E., Castellote, M., Corkeron, P. J., Nowacek, D. P., et al. (2012). Evidence that Ship Noise Increases Stress in Right Whales. Proc. R. Soc. B 279 (1737), 2363–2368. doi:10.1098/rspb.2011.2429
Rugh, D. J., Shelden, K. E. W., and Schulman-Janiger, A. (2001). Timing of the Gray Whale Southbound Migration. J. Cetacean Res. Manag. 3 (1), 31–39. 9.
Stevick, P. T., Neves, M. C., Johansen, F., Engel, M. H., Allen, J., Marcondes, M. C. C., et al. (2011). A Quarter of a World Away: Female Humpback Whale Moves 10 000 Km between Breeding Areas. Biol. Lett. 7 (2), 299–302. doi:10.1098/rsbl.2010.0717
Vernazzani, B. G., Cabrera, E., and Brownell, R. L. (2014). Eastern South Pacific Southern Right Whale Photo-Identification Catalog Reveals Behavior and Habitat Use Patterns. Mar. Mam. Sci. 30 (1), 389–398. doi:10.1111/mms.12030
Wickham, H., and RStudio, (2021). Tidyverse: Easily Install and Load the “Tidyverse” (1.3.1). [Computer Software]. Available at: https://CRAN.R-project.org/package=tidyverse.
Keywords: passive acoustic monitoring, southern right whales, diel trends, temporal trends, third octave levels, active space
Citation: Jacobs ER, Landea-Briones R and Sayigh L (2022) Temporal Trends and Effects of Noise on Upsweep Calls of Eastern South Pacific Southern Right Whales. Front. Remote Sens. 3:909309. doi: 10.3389/frsen.2022.909309
Received: 31 March 2022; Accepted: 10 June 2022;
Published: 30 August 2022.
Edited by:
Susan E. Parks, Syracuse University, United StatesReviewed by:
Gilberto Corso, Federal University of Rio Grande do Norte, BrazilCopyright © 2022 Jacobs, Landea-Briones and Sayigh. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ellen Rose Jacobs, ZXJqMjJAZ2VvcmdldG93bi5lZHU=, orcid.org/0000-0003-2957-3286
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.