
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Public Health , 04 April 2024
Sec. Infectious Diseases: Epidemiology and Prevention
Volume 12 - 2024 | https://doi.org/10.3389/fpubh.2024.1337357
This article is part of the Research Topic World TB Day 2023: Yes! We can end TB View all 30 articles
 Bernardo Castro-Rodriguez1
Bernardo Castro-Rodriguez1 Greta Franco-Sotomayor2,3Jose Manuel Benitez-Medina4Greta Cardenas-Franco1Natalia Jiménez-Pizarro4Camilo Cardenas-Franco3Juan Luis Aguirre-Martinez3Solon Alberto Orlando2,5Javier Hermoso de Mendoza4
Greta Franco-Sotomayor2,3Jose Manuel Benitez-Medina4Greta Cardenas-Franco1Natalia Jiménez-Pizarro4Camilo Cardenas-Franco3Juan Luis Aguirre-Martinez3Solon Alberto Orlando2,5Javier Hermoso de Mendoza4 Miguel Angel Garcia-Bereguiain1*
Miguel Angel Garcia-Bereguiain1*Introduction: A major sublineage within the Mycobacterium tuberculosis (MTB) LAM family characterized by a new in-frame fusion gene Rv3346c/55c was discovered in Rio de Janeiro (Brazil) in 2007, called RDRio, associated to drug resistance. The few studies about prevalence of MTB RDRio strains in Latin America reported values ranging from 3% in Chile to 69.8% in Venezuela, although no information is available for countries like Ecuador.
Methods: A total of 814 MTB isolates from years 2012 to 2016 were screened by multiplex PCR for RDRio identification, followed by 24-loci MIRU-VNTR and spoligotyping.
Results: A total number of 17 MTB RDRio strains were identified, representing an overall prevalence of 2.09% among MTB strains in Ecuador. While 10.9% of the MTB isolates included in the study were multidrug resistance (MDR), 29.4% (5/17) of the RDRio strains were MDR.
Discussion: This is the first report of the prevalence of MTB RDRio in Ecuador, where a strong association with MDR was found, but also a very low prevalence compared to other countries in Latin America. It is important to improve molecular epidemiology tools as a part of MTB surveillance programs in Latin America to track the transmission of potentially dangerous MTB stains associated to MDR TB like MTB RDRio.
Tuberculosis (TB) is one of the leading causes of death from a single infectious agent, ranking just behind the coronavirus (COVID-19) and above HIV-AIDS (1). The disease is caused by members of the Mycobacterium tuberculosis Complex (MTBC) which are intracellular, rod-shaped, aerobic bacteria with a lipid-rich cell wall structured with mycolic acids that confer specific microbiological characteristics, like resistance to detergents, slow growth, and reaction to acid-fast stain methods. The main mechanism of spreading occurs by aerosols, affecting the lungs (pulmonary TB) and other organs (extrapulmonary TB) (1–3).
Mycobacterium tuberculosis comprehends a group of seven highly related lineages that infect mainly humans. Global distribution of these lineages differs greatly (4). In Latin America, the most frequent MTBC strains belong to the Euro-American lineage 4, which includes the Latin American-Mediterranean sublineage (LAM) (5–21). In Ecuador, there is a single report about population structure of MTBC showing that LAM sublineage is predominant (6). Moreover, LAM clonal complex comprised by Ecuadorian and Colombian strains were reported, suggesting transnational transmission of TB (6).
The genetic diversity in the MTBC species finds its origins in genomic alterations in targeted mycobacterial segments like region-of-difference (RD) loci that are used as specific markers for M. tuberculosis lineages (22, 23). In 2007, Lazzarini and collaborators applied a deletion-based PCR protocol to a collection of MTBC samples from Rio de Janeiro, Brazil, and identified a new in-frame fusion gene Rv3346c/55c that marks a major sublineage within the LAM family. This long sequence polymorphism results from a homologous recombination deletion between Rv3346c and Rv355c genes located 26.3 kb apart (22). This LAM variant, called MTB RDRio is associated with multidrug resistance and distributed worldwide (22, 24–27). The prevalence of MTB RDRio varies in Latin America: 30 to 51.9% in Brazil (22, 28), 69.8% in Venezuela (19), 37% in Colombia (17, 21, 29), 31% in Peru (21, 30), 20% in Argentina (21), 10% in Paraguay (18) and 3% in Chile (21). However, there are no reports of the RDRio in Ecuador.
The aim of this study was to address the prevalence, drug resistance profile and population structure the MTB RDRio strains circulating in Ecuador.
A collection of 814 M. tuberculosis isolates from years 2012 to 2016 stored at “Centro Nacional de Referencia para Micobacterias” from “Instituto Nacional de Salud Pública e Investigación Leopoldo Izquieta Pérez” in Guayaquil (Ecuador) were included in the study (Figure 1). MTBC isolates are routinely processed at INSPI laboratories, where culture and antibiotic resistance profiling for first-and second-line drugs used in M. tuberculosis therapy is performed for MTBC cultures following Pan American Health Organization guidelines (31–33). The samples were previously inactivated and stored for research purposes following the guidelines from this government center. The access to this MTB strains collection was approved by IRBs from “Instituto Nacional de Salud Pública e Investigación Leopoldo Izquieta Pérez” and University San Francisco de Quito (code 2017-023IN), both certified by Ministry of Public Health from Ecuador following guidelines from Declaration of Helsinki. All samples were anonymized, and no data of the patients were made available.
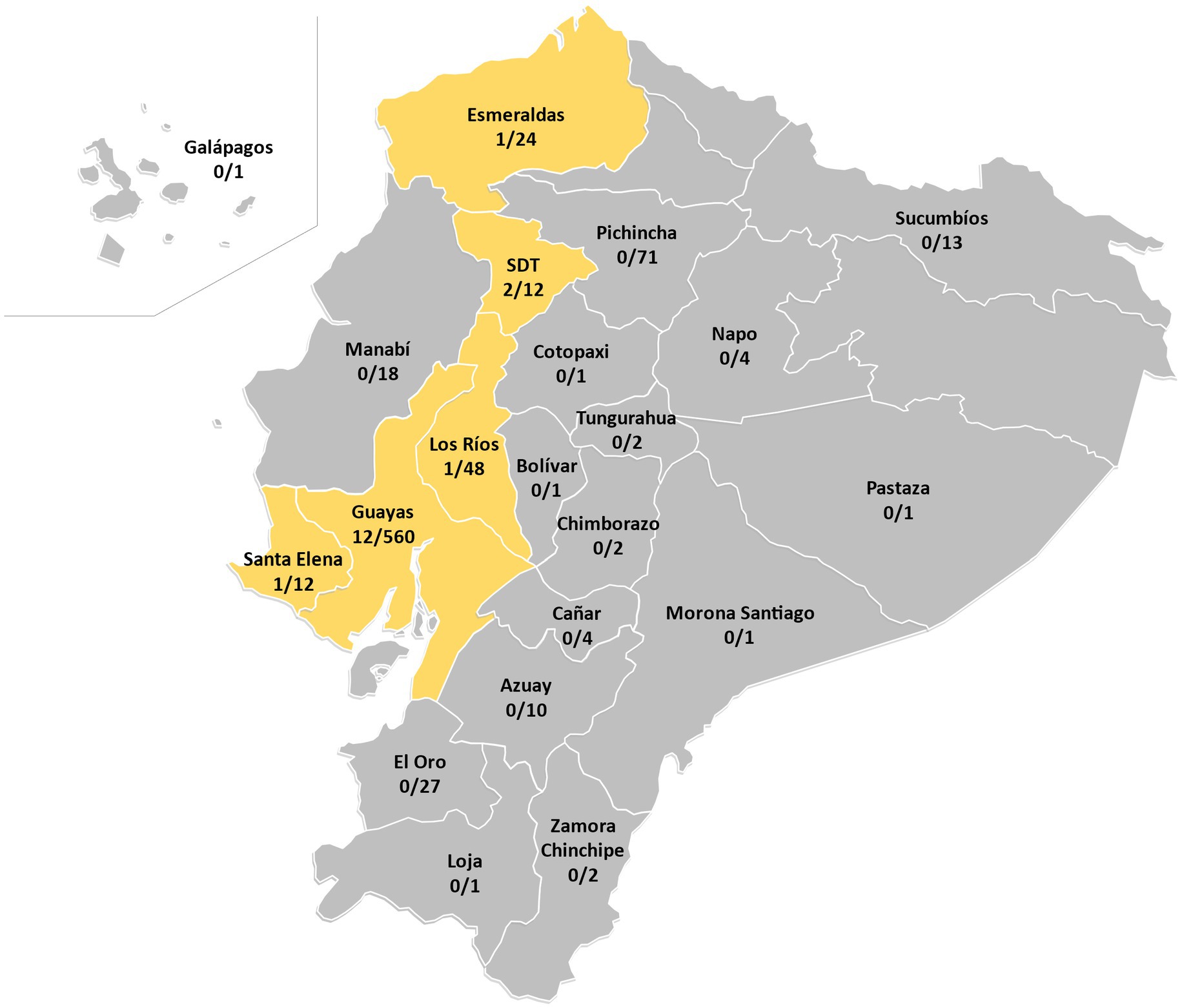
Figure 1. Ratio of RDRio isolates in each province of Ecuador (Total of RDRio isolates = 17). SDT, Santo Domingo de los Tsáchilas.
A small sample from cultures of MTBC isolates was collected and resuspended in TE buffer (10 mM Tris-HCl, 1 mM EDTA, pH 8.0), then inactivated by boiling at 95°C for 45 min. After this process, samples were centrifuged for 5 min at 10,000 g and the supernatant was directly used for genotyping, as reported elsewhere (34, 35).
A multiplex PCR developed for the detection of the RDRio pattern was performed using a two set of primers (Supplementary Figure S1): (1) a set of primers that target the IS1561’ fragment, located inside the region between the genes Rv3346c and Rv3355c, and used as a marker for wild type M. tuberculosis (530 bp); (2) another set of primers is used to flank the RDRio locus and bridge the deletion in RDRio strains (1175 bp) (36). The PCR reaction was prepared as follows: 12.5 μL of GoTaq® Green Mastermix (Promega, Wisconsin, United States) (1X), 0.5 μL of each primer (0.2 μM), 5 μL of mycobacterial DNA and completed with Nuclease-Free water to a final volume of 25 μL. The multiplex PCR program was established as follows: initial denaturation at 94°C for 3 min, 35 cycles at 94°C for 30 s, 60°C for 30 s, 72°C for 1 min 30 s, and a final extension step of 72°C for 10 min. PCR products were run in 1.5%UltraPure™ Agarose (Invitrogen, California, United States) gels of 15 cm × 10 cm in 0.5X Tris-boric acid-EDTA (TBE) buffer at 100 V for 2 h using a ladder 100 bp Plus Opti-DNA Marker (Cat. No.: G016, Applied Biological Materials Inc., British Columbia, Canada) for size determination (22, 36).
The method is PCR-based and allows the detection of different Mycobacterial Interspersed Repetitive Units (MIRU) located at multiple loci in the MTBC genome. Each MIRU allele is identified by a number; thus generating a numerical profile which is used for genotyping studies (35, 37). The PCR reaction was prepared individually to a final volume of 15 μL: 7.5 μL of GoTaq® Green Mastermix (Promega, Wisconsin, United States) (1X), 0.75 μL of MIRU primers (0.5 μM) (38), 1 μL of mycobacterial DNA and completed with Nuclease-Free water. The following PCR program was established for all MIRUs: initial denaturation at 94°C for 5 min, 35 cycles at 94°C for 1 min, 54°C for 1 min 30 s, 72°C for 1 min 30 s, and a final extension step of 72°C for 10 min. Amplicons were run in 2% UltraPure™ Agarose (Invitrogen, California, United States) gels of 15 cm × 10 cm in 0.5X Tris-boric acid-EDTA (TBE) buffer at 100 V for 3 h using a ladder 100 bp Plus Opti-DNA Marker (Cat. No.: G016, Applied Biological Materials Inc., British Columbia, Canada) for size determination. MIRU allele identification was performed according to Supply et al. (38).
Results obtained were analyzed using the MIRU-VNTRplus web application1 (39). Lineage designation was performed by similarity search using 24-loci MIRU-VNTR data (Supplementary Data 1) and calculation of Minimum Spanning Tree (MST) and Neighbour-joining Tree (NJT) was performed to determine the population structure of Ecuadorian M. tuberculosis isolates.
The method is based on the presence/absence of 43 DNA spacer sequences, which are interspersed between 36 conserved loci located in the Direct Repeat (DR) region (37, 40). Spoligotyping was performed according to Kamerbeek et al. (41): PCR amplification of DR loci was performed using DRa and DRb primers and products were biotinylated and hybridized to a membrane containing oligonucleotides for each spacer sequence. After hybridization, the membrane is washed and then incubated in diluted streptavidin-peroxidase conjugate (Roche, United States). Membrane was exposed to chemiluminescent Amersham ECL reagents (GE Healthcare, United Kingdom) and located in an X-Ray cassette on a Hyperfilm ECL (Merck, United States). After the reaction, the film was inserted into a film developer solution in a dark room after which it is moved to the fixer solution. Thereafter, the film is dried and ready for the interpretation of the result. After developing the film a positive/negative signal is recorded in binary or octal formats for genotyping interpretations (Supplementary Data 1). The results are compared against the Fourth International Spoligotyping Database (SpolDB4) (42).
A total of 814 M. tuberculosis isolates were analyzed by multiplex PCR for MTB RDRio identification (Figure 1). Of those, 17 strains generated a band pattern in the electrophoresis gel compatible with MTB RDRio (Figure 1; Table 1; Supplementary Figure S1), representing a 2.09% of the total MTBC population analyzed. Those MTB RDRio strains were assigned to LAM MTB subfamily by MIRU-VNTR (Table 1; Figure 2; Supplementary Data 1). Those 17 isolates were subsequently analyzed by spoligotyping, with 6 strains with undetermined lineage and 11 of them associated to LAM sublineage: five belong to LAM9, one to LAM5-LAM6, two belong to LAM3, one to LAM2 and two were identified as LAM1, according to the SpolDB4 (Table 1; Figure 2; Supplementary Data 1).
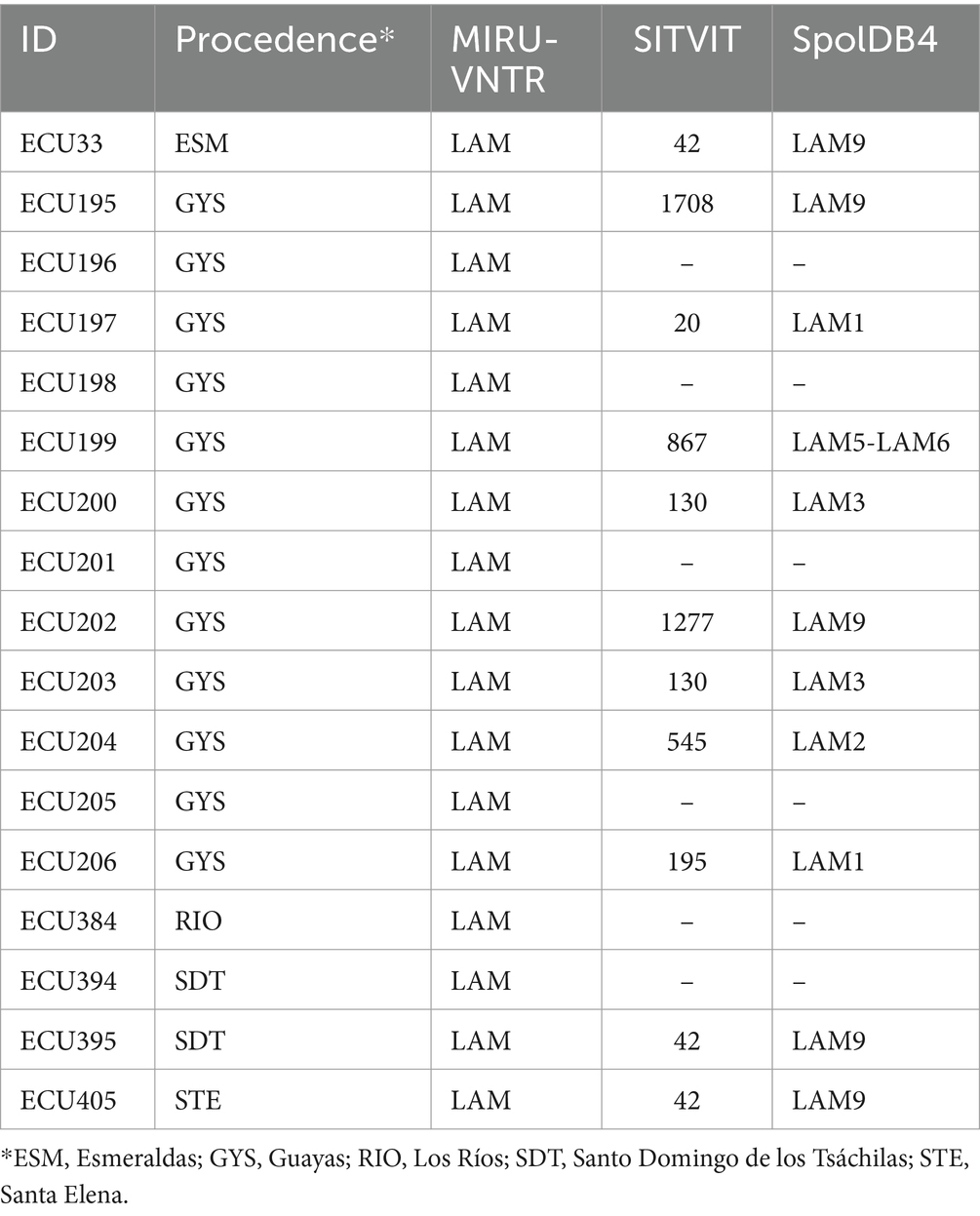
Table 1. MTB RDRio isolates in Ecuador.
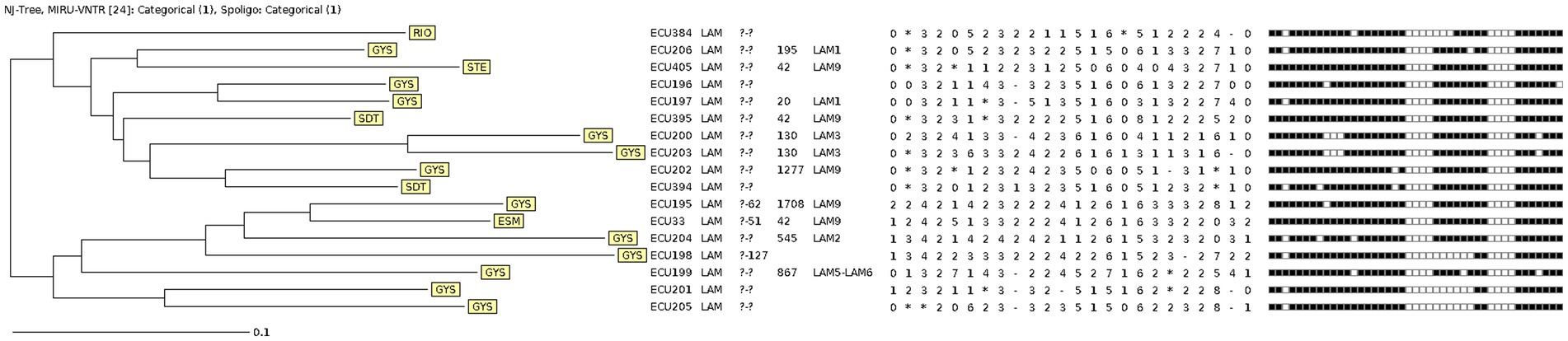
Figure 2. Neighbour-joining Tree of Ecuadorian MTB RDRio isolates using 24-loci MIRU-VNTR and Spoligotyping data. Labels use three-letter code to identify province of origin: ESM, Esmeraldas; GYS, Guayas; RIO, Los Ríos; SDT, Santo Domingo de los Tsáchilas; STE, Santa Elena.
Regarding the geographic location of the 17 MTB RDRio strains, all of them were isolated from provinces in the Coastal Region of Ecuador: Guayas, Los Rios, Santo Domingo and Santa Elena (Figure 1).
Drug susceptibility testing of MTB RDRio isolates was performed for first line antibiotics: 8/17 were resistant to isoniazid (47.1%), 1/17 was resistant to streptomycin (5.9%), 6/17 were resistant to rifampicin (35.3%) and no samples showed resistance to ethambutol and pyrazinamide; 9/17 isolates were sensitive to all first-line antibiotics (52.9%). Finally, 5/11 (29.4%) isolates were resistant to isoniazid and rifampicin, that means multi drug resistance (MDR) strains (Table 2). For the whole collection of 814 MTB strains included in the study, the MDR strains represented 10.9% (Table 3).
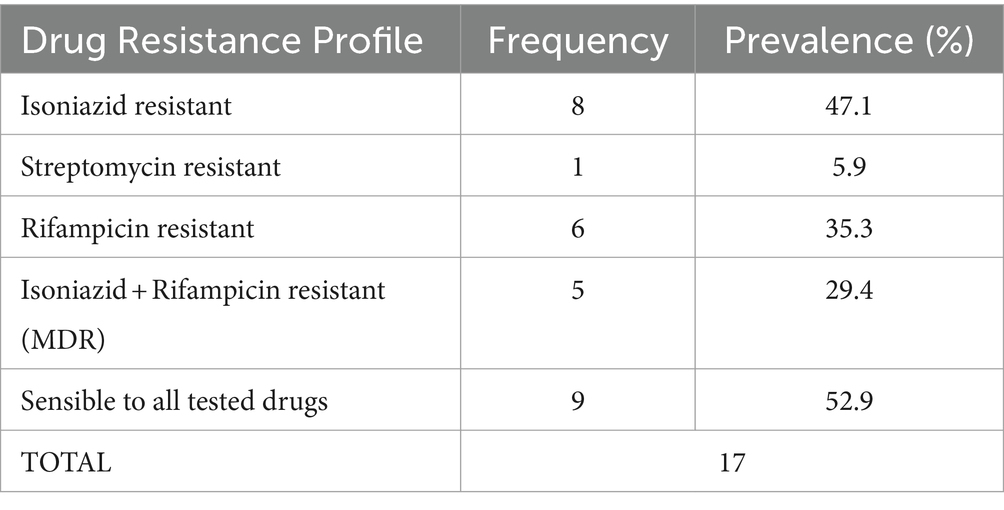
Table 2. Frequency of MTB RDRio isolates resistant to first-line therapy drugs and prevalence.
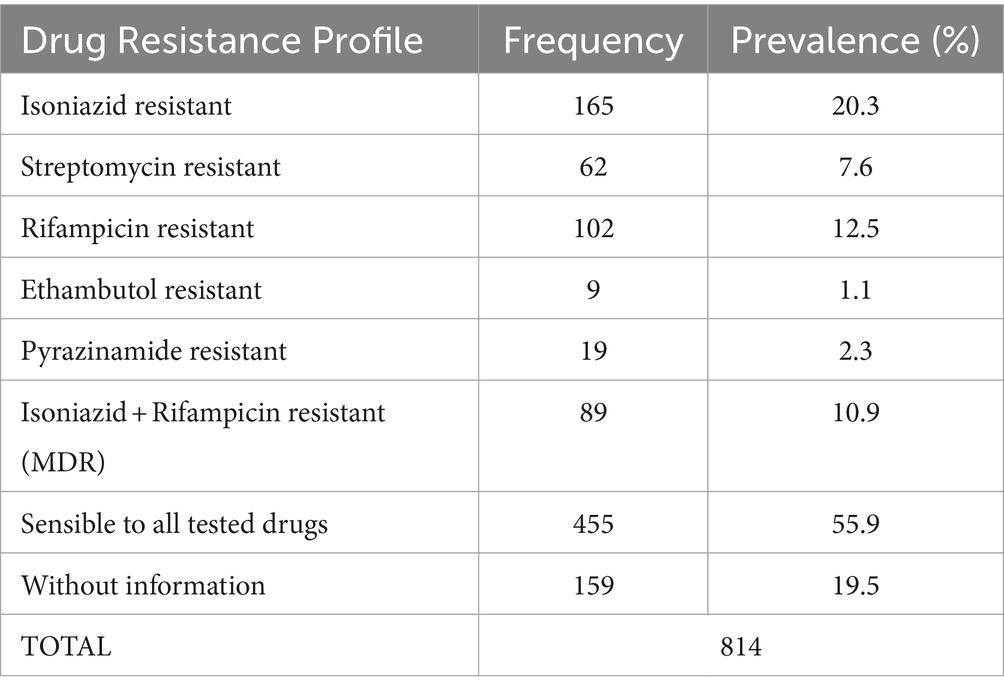
Table 3. Frequency of Ecuadorian MTB isolates resistant to first-line therapy drugs and prevalence.
The population structure of the MTB RDRio isolates was determined through the analysis of 24-loci MIRU-VNTR and spoligotyping using the MIRU-VNTRplus platform (Supplementary Data 1), as it is detailed in the Neighbour-Joining Tree (NJT) presented in Figure 2. Although several spoligotype patters were repeated for 2 or 3 MTB RDRio strains, none of the strains shared the same MIRU-VNTR pattern. Based on total identity in 24 loci for MIRU-VNTR, no actively transmitted MTB RDRio clones were found (Figure 2).
Additionally, a phylogenetic analysis was done using 12-loci MIRU-VNTR, including all the MTB RDRio strains reported in the bibliography (18, 43), as well as the strains identified in the current study for Ecuador. This analysis is detailed in the Minimum Spanning Tree (MST) in the Figure 3, where the maximum locus difference within a clonal complex was set at 2. MST included 17 MTB RDRio isolates from Ecuador, 8 from Argentina, 31 from Paraguay, 51 from Venezuela and 15 from Brazil. Four clonal complexes of MTB RDRio isolates are well defined in the MST: two clonal complexes comprising isolates exclusively from Ecuador, one clonal complex comprising isolates exclusively from Paraguay, and one large clonal complex including MTB RDRio isolates from Ecuador, Argentina, Brazil, Paraguay and Venezuela. Interestingly, 12 of 17 MTB RDRio strains from Ecuador are well differentiated from strains from the other South American countries.

Figure 3. Minimum Spanning Tree of Latin American RD Rio isolates isolates using 12-loci MIRU-VNTR data. Maximum locus difference within a clonal complex is 2. Labels use three-letter code to identify country of origin (ARG, Argentina; BRA, Brazil; ECU, Ecuador; PAR, Paraguay; VEN, Venezuela).
We found the lowest prevalence of MTB RDRio subfamily described for South America in Ecuador (2.09%), while neighbor countries like Peru and Colombia have an MTB RDRio prevalence over 30% (21). Although some biological advantages that improve virulence and transmissibility over other genotypes has been described for MTB RDRio (36, 44, 45), this could explain its successful transmission in some countries like Venezuela, Brazil, Peru and Colombia, but not in Ecuador (22, 46, 47). In fact, two recent studies have shown a very limited transnational transmission of MTB between Ecuador, Colombia and Peru that would also explain the low prevalence of MTB RDRio sublineage found in our study (48, 49).
The phylogenetic analysis of MTBC RDRio strains identified in Ecuador suggests a regional cluster associated to provinces in the Coastal Region but with an elevated genetic diversity and lack of active transmission chains. While a single large clonal complex includes MTB RDRio isolates from all South American countries was found, 70% of RDRio strains from Ecuador were clearly segregated in two clonal complexes. Additionally, the high genetic diversity of MTB RDRio strains in Ecuador was also confirmed by the multiple patterns of LAM spoligotypes found in our study.
The RDRio strains from Ecuador were found strongly associated to MDR TB in our study. The reported value of 29.4% MDR strains is clearly over either the 9% of MDR TB estimated for Ecuador for 2017 (6) or the 10.9% of MDR TB strains in the collection included in this study. This result points out that MTB RDRio strains should be considered of concern due to their potential to mutate to drug resistance phenotypes (21, 28, 50, 51).
Our study has two main limitations. First, we could only have access to 814 MTB strains from the national reference located in Guayaquil, so geographic bias in this collection could happened with over representation of provinces closer to this city. Second, our study was done with samples from years 2012–2016, prior to the massive Venezuelan migration that arrived to Ecuador since 2018 following this country economic collapse (52, 53). Considering that the highest prevalence of MTB RDRio in South America has been described for Venezuela (19), the current scenario for MTB RDRio prevalence in Ecuador could have changed due to importation of cases and further studies with MTB strains collection from recent years are needed.
In conclusion, we report for the first time the presence of MTB RDRio in Ecuador and its strong association to MDR-TB. We encourage the national surveillance program in Ecuador to follow up the potential expansion of this MTB strains of concern considering the TB burden associated to COVID-19 pandemic and the recent massive regional patters of migration from Venezuela where MTB RDRio is highly present.
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author.
BC-R: Conceptualization, Formal analysis, Investigation, Methodology, Writing – original draft, Writing – review & editing. GF-S: Conceptualization, Data curation, Investigation, Methodology, Resources, Supervision, Writing – review & editing. JB-M: Formal analysis, Methodology, Validation, Writing – review & editing. GC-F: Data curation, Formal analysis, Investigation, Methodology, Writing – review & editing. NJ-P: Data curation, Formal analysis, Methodology, Writing – review & editing. CC-F: Data curation, Formal analysis, Validation, Writing – review & editing. JA-M: Project administration, Resources, Supervision, Writing – review & editing. JH: Funding acquisition, Methodology, Resources, Supervision, Validation, Writing – review & editing. MG-B: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Writing – original draft. SO: Methodology, Resources, Project administration, Writing- review & editing.
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. The study was funded by Universidad de Las Américas (MED.MGB.23.01).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpubh.2024.1337357/full#supplementary-material
3. Madigan, MT, Bender, KS, Buckley, DH, Sattley, WM, and Stahl, DA. Brock biology of microorganisms. Fifteenth. New York: Pearson Education (2019).
4. Brites, D, and Gagneux, S. The nature and evolution of genomic diversity in the Mycobacterium tuberculosis complex. S Gagneux (ed) Strain variation in the Mycobacterium tuberculosis complex: its role in biology, epidemiology and control. Cham: Springer International Publishing, (2017), 1–26
5. Brynildsrud, OB, Pepperell, CS, Suffys, P, Grandjean, L, Monteserin, J, Debech, N, et al. Global expansion Mycobacterium tuberculosis lineage 4 shaped by colonial migration and local adaptation. Sci Adv. (2018) 4:5869–86. doi: 10.1126/sciadv.aat5869
6. Garzon-Chavez, D, Garcia-Bereguiain, MA, Mora-Pinargote, C, Granda-Pardo, JC, Leon-Benitez, M, Franco-Sotomayor, G, et al. Population structure and genetic diversity of Mycobacterium tuberculosis in Ecuador. Sci Rep. (2020) 10:6237. doi: 10.1038/s41598-020-62824-z
7. Grandjean, L, Iwamoto, T, Lithgow, A, Gilman, RH, Arikawa, K, Nakanishi, N, et al. The association between Mycobacterium Tuberculosis genotype and drug resistance in Peru. PLoS One. (2015) 10:e0126271. doi: 10.1371/journal.pone.0126271
8. Barletta, F, Otero, L, Collantes, J, Asto, B, de Jong, BC, Seas, C, et al. Genetic variability of Mycobacterium tuberculosis complex in patients with no known risk factors for MDR-TB in the north-eastern part of Lima, Peru. BMC Infect Dis. (2013) 13:397. doi: 10.1186/1471-2334-13-397
9. Sheen, P, Couvin, D, Grandjean, L, Zimic, M, Dominguez, M, Luna, G, et al. Genetic diversity of Mycobacterium tuberculosis in Peru and exploration of phylogenetic associations with drug resistance. PLoS One. (2013) 8:e65873. doi: 10.1371/journal.pone.0065873
10. Eldholm, V, Monteserin, J, Rieux, A, Lopez, B, Sobkowiak, B, Ritacco, V, et al. Four decades of transmission of a multidrug-resistant Mycobacterium tuberculosis outbreak strain. Nat Commun. (2015) 6:7119. doi: 10.1038/ncomms8119
11. Flores-Treviño, S, Morfín-Otero, R, Rodríguez-Noriega, E, González-Díaz, E, Pérez-Gómez, HR, Bocanegra-García, V, et al. Genetic diversity of Mycobacterium tuberculosis from Guadalajara, Mexico and identification of a rare multidrug resistant Beijing genotype. PLoS One. (2015) 10:118095. doi: 10.1371/journal.pone.0118095
12. Bocanegra-García, V, Garza-González, E, Cruz-Pulido, WL, Guevara-Molina, YL, Cantú-Ramírez, R, González, GM, et al. Molecular assessment, drug-resistant profile, and spacer oligonucleotide typing (Spoligotyping) of Mycobacterium tuberculosis strains from Tamaulipas. México J Clin Lab Anal. (2014) 28:97–103. doi: 10.1002/jcla.21650
13. Soares, RO, de Macedo, MB, von Groll, A, and da Silva, PE. Mycobacterium tuberculosis belonging to family LAM and sublineage RDRio: common strains in southern Brazil for over 10 years. Braz J Microbiol. (2013) 44:1251–5. doi: 10.1590/S1517-83822013000400032
14. von Groll, A, Martin, A, Felix, C, Prata, PFS, Honscha, GÃ, Portaels, FÃ, et al. Fitness study of the RDRio lineage and Latin Americanâ Mediterranean family Ofmycobacterium tuberculosisin the city of Rio Grande, Brazil. FEMS Microbiol Index. (2010) 58:119–27. doi: 10.1111/j.1574-695X.2009.00611.x
15. Dalla Costa, ER, Lazzarini, LCO, Perizzolo, PF, Díaz, CA, Spies, FS, Costa, LL, et al. Mycobacterium tuberculosis of the RDRio genotype is the predominant cause of tuberculosis and associated with multidrug resistance in Porto Alegre City, South Brazil. J Clin Microbiol. (2013) 51:1071–7. doi: 10.1128/JCM.01511-12
16. Vasconcellos, SEG, Acosta, CC, Gomes, LL, Conceição, EC, Lima, KV, de Araujo, MI, et al. Strain classification of Mycobacterium tuberculosis isolates in Brazil based on genotypes obtained by Spoligotyping, mycobacterial interspersed repetitive unit typing and the presence of large sequence and single nucleotide polymorphism. PLoS One. (2014) 9:e107747. doi: 10.1371/journal.pone.0107747
17. Cerezo, I, Jiménez, Y, Hernandez, J, Zozio, T, Murcia, MI, and Rastogi, N. A first insight on the population structure of Mycobacterium tuberculosis complex as studied by spoligotyping and MIRU-VNTRs in Bogotá, Colombia. Infect Genet Evol. (2012) 12:657–63. doi: 10.1016/j.meegid.2011.07.006
18. Díaz Acosta, CC, Russomando, G, Candia, N, Ritacco, V, Vasconcellos, SEG, de Berrêdo Pinho Moreira, M, et al. Exploring the Latin American Mediterranean family and the RD(Rio) lineage in Mycobacterium tuberculosis isolates from Paraguay, Argentina and Venezuela. BMC Microbiol. 19:e6. doi: 10.1186/s12866-019-1479-6
19. Méndez, MV, Abadía, E, Sequera, M, de Waard, JH, and Takiff, HE. Most LAM Mycobacterium tuberculosis strains in Venezuela, but not SIT605, belong to the RDRio subfamily. Infect Genet Evol. (2020) 84:104380. doi: 10.1016/j.meegid.2020.104380
20. Zurita, J, Espinel, N, Barba, P, Ortega-Paredes, D, Zurita-Salinas, C, Rojas, Y, et al. Genetic diversity and drug resistance of Mycobacterium tuberculosis in Ecuador. Int J Tuberculosis Lung Disease. (2019) 23:166–73. doi: 10.5588/ijtld.18.0095
21. Mokrousov, I, Vyazovaya, A, Iwamoto, T, Skiba, Y, Pole, I, Zhdanova, S, et al. Latin-American-Mediterranean lineage of Mycobacterium tuberculosis: human traces across pathogen’s phylogeography. Mol Phylogenet Evol. (2016) 99:133–43. doi: 10.1016/j.ympev.2016.03.020
22. Lazzarini, LCO, Huard, RC, Boechat, NL, Gomes, HM, Oelemann, MC, Kurepina, N, et al. Discovery of a novel Mycobacterium tuberculosis lineage that is a major cause of tuberculosis in Rio de Janeiro. Brazil J Clin Microbiol. (2007) 45:3891–902. doi: 10.1128/JCM.01394-07
23. Merker, M, Kohl, TA, Niemann, S, and Supply, P The evolution of strain typing in the Mycobacterium tuberculosis complex. (2017), 43–78
24. Chizimu, JY, Solo, ES, Bwalya, P, Kapalamula, TF, Akapelwa, ML, Lungu, P, et al. Genetic diversity and transmission of multidrug-resistant Mycobacterium tuberculosis strains in Lusaka. Zambia Int J Infectious Diseases. (2022) 114:142–50. doi: 10.1016/j.ijid.2021.10.044
25. Mokrousov, I, Jiao, WW, Wan, K, and Shen, A. Stranger in a strange land: Ibero-American strain of Mycobacterium tuberculosis in Tibet, China. Infect Genet Evol. (2014) 26:323–6. doi: 10.1016/j.meegid.2014.06.011
26. Bocanegra-García, V, Cortez-de-la-Fuente, LJ, Nakamura-López, Y, González, GM, Rivera, G, and Palma-Nicolás, JP. RDRio Mycobacterium tuberculosis strains associated with isoniazid resistance in northern Mexico. Enferm Infecc Microbiol Clin. (2021) 39:399–402. doi: 10.1016/j.eimce.2020.07.010
27. Huber, FD, Sánchez, A, Gomes, HM, Vasconcellos, S, Massari, V, Barreto, A, et al. Insights into the population structure of Mycobacterium tuberculosis using spoligotyping and RDRio in a southeastern Brazilian prison unit. Infect Genet Evol. (2014) 26:194–202. doi: 10.1016/j.meegid.2014.05.031
28. de Almeida, IN, Vasconcellos, SEG, de Assis Figueredo, LJ, Dantas, NGT, Augusto, CJ, Hadaad, JPA, et al. Frequency of the Mycobacterium tuberculosis RDRio genotype and its association with multidrug-resistant tuberculosis. BMC Infect Dis. (2019) 19:556. doi: 10.1186/s12879-019-4152-7
29. Puerto, G, Erazo, L, Wintaco, M, Castro, C, Ribón, W, and Guerrero, MI. Mycobacterium tuberculosis genotypes determined by spoligotyping to be circulating in Colombia between 1999 and 2012 and their possible associations with transmission and susceptibility to first-line drugs. PLoS One. (2015) 10:124308. doi: 10.1371/journal.pone.0124308
30. Taype, CA, Agapito, JC, Accinelli, RA, Espinoza, JR, Godreuil, S, Goodman, SJ, et al. Genetic diversity, population structure and drug resistance of Mycobacterium tuberculosis in Peru. Infect Genet Evol. (2012) 12:577–85. doi: 10.1016/j.meegid.2012.02.002
31. Organización Panamericana de la Salud, Organización Mundial de la Salud . Manual para el Diagnóstico Bacteriológico de la Tuberculosis. Normas y Guía Técnica-Parte 2: Cultivo, Available at: https://iris.paho.org/handle/10665.2/18616 (2008)
32. Franco-Sotomayor, G, Rivera-Olivero, IA, Leon-Benitez, M, Uruchima-Campoverde, SE, Cardenas-Franco, G, Perdomo-Castro, ME, et al. Fast, simple, and cheap: the Kudoh-Ogawa swab method as an alternative to the Petroff–Lowenstein-Jensen method for culturing of Mycobacterium tuberculosis. J Clin Microbiol. (2020) 58:e19. doi: 10.1128/JCM.01424-19
33. Garcia-Bereguiain, MA . Kudoh swab method for Mycobacterium tuberculosis culture: looking back to the 70s to fight tuberculosis. Int J Mycobacteriol. (2020) 9:229–30. doi: 10.4103/ijmy.ijmy_66_20
34. Garzon-Chavez, D, Zurita, J, Mora-Pinargote, C, Franco-Sotomayor, G, Leon-Benitez, M, Granda-Pardo, JC, et al. Prevalence, drug resistance, and genotypic diversity of the Mycobacterium tuberculosis Beijing family in Ecuador. Microb Drug Resist. (2019) 25:931–7. doi: 10.1089/mdr.2018.0429
35. Mora-Pinargote, C, Garzon-Chavez, D, Franco-Sotomayor, G, Leon-Benitez, M, Granda-Pardo, JC, Trueba, G, et al. Country-wide rapid screening for the Mycobacterium tuberculosis Beijing sublineage in Ecuador using a single-nucleotide polymorphism-polymerase chain reaction method. Int J Mycobacteriol. (2019) 8:366–70. doi: 10.4103/ijmy.ijmy_132_19
36. Gibson, AL, Huard, RC, Gey van Pittius, NC, Lazzarini, LCO, Driscoll, J, Kurepina, N, et al. Application of sensitive and specific molecular methods to uncover global dissemination of the major RDRio sublineage of the Latin American-Mediterranean Mycobacterium tuberculosis spoligotype family. J Clin Microbiol. (2008) 46:1259–67. doi: 10.1128/JCM.02231-07
37. Supply, P, Allix, C, Lesjean, S, Cardoso-Oelemann, M, Rüsch-Gerdes, S, Willery, E, et al. Proposal for standardization of optimized mycobacterial interspersed repetitive unit-variable-number tandem repeat typing of Mycobacterium tuberculosis. J Clin Microbiol. (2006) 44:4498–510. doi: 10.1128/JCM.01392-06
38. Supply, P. , Multilocus variable number tandem repeat genotyping of Mycobacterium tuberculosis technical guide, Available at: https://www.researchgate.net/publication/265990159 (2014)
39. Allix-Béguec, C, Harmsen, D, Weniger, T, Supply, P, and Niemann, S. Evaluation and strategy for use of MIRU-VNTRplus, a multifunctional database for online analysis of genotyping data and phylogenetic identification of Mycobacterium tuberculosis complex isolates. J Clin Microbiol. (2008) 46:2692–9. doi: 10.1128/JCM.00540-08
40. Coll, P, and García de Viedma, D. Epidemiología molecular de la tuberculosis. Enferm Infecc Microbiol Clin. (2018) 36:233–40. doi: 10.1016/j.eimc.2018.01.001
41. Kamerbeek, J, Schouls, L, Kolk, A, van Agterveld, M, van Soolingen, D, Kuijper, S, et al. Simultaneous detection and strain differentiation of Mycobacterium tuberculosis for diagnosis and epidemiology. J Clin Microbiol. (1997) 35:907–14. doi: 10.1128/jcm.35.4.907-914.1997
42. Brudey, K, Driscoll, JR, Rigouts, L, Prodinger, WM, Gori, A, al-Hajoj, SA, et al. Mycobacterium tuberculosis complex genetic diversity: mining the fourth international spoligotyping database (SpolDB4) for classification, population genetics and epidemiology. BMC Microbiol. 6:e23. doi: 10.1186/1471-2180-6-23
43. de Almeida, AL, Scodro, RB, de Carvalho, HC, Costacurta, GF, Baldin, VP, and NCS, S. RD Rio Mycobacterium tuberculosis lineage in the Brazil/Paraguay/Argentina triple border. Tuberculosis. (2018) 110:68–78. doi: 10.1016/j.tube.2018.03.008
44. Reynaud, Y, Millet, J, and Rastogi, N. Genetic structuration, demography and evolutionary history of mycobacterium tuberculosis LAM9 sublineage in the Americas as two distinct subpopulations revealed by Bayesian analyses. PLoS One. 10:140911. doi: 10.1371/journal.pone.0140911
45. Dalla Costa, ER, Ribeiro, MO, Silva, MS, Arnold, LS, Rostirolla, DC, Cafrune, PI, et al. Correlations of mutations in katG, oxyR-ahpC and inhA genes and in vitro susceptibility in Mycobacterium tuberculosis clinical strains segregated by spoligotype families from tuberculosis prevalent countries in South America. BMC Microbiol. (2009) 9:e39. doi: 10.1186/1471-2180-9-39
46. Moraes, EB, Slompo, L, Finardi, AJ, Silveira,, Ruiz, L, Gomes, HM, et al., Tuberculosis associated factors caused by Mycobacterium tuberculosis of the RDRio genotype. Mem Inst Oswaldo Cruz (2017); 112: 182–187, doi: 10.1590/0074-02760160347
47. Mokrousov, I, Vyazovaya, A, and Narvskaya, O. Mycobacterium tuberculosis Latin American-Mediterranean family and its sublineages in the light of robust evolutionary markers. J Bacteriol. (2014) 196:1833–41. doi: 10.1128/JB.01485-13
48. Castro-Rodriguez, B, León-Ordóñez, K, Franco-Sotomayor, G, Benítez-Medina, JM, Jiménez-Pizarro, N, Cárdenas-Franco, G, et al. Population structure of Mycobacterium tuberculosis in El Oro: a first insight into Ecuador-Peru tuberculosis transmission. J Infect Public Health. (2024) 17:527–34. doi: 10.1016/j.jiph.2024.01.015
49. Castro-Rodriguez, B, Espinoza-Andrade, S, Franco-Sotomayor, G, Benítez-Medina, JM, Jiménez-Pizarro, N, Cárdenas-Franco, C, et al. A first insight into tuberculosis transmission at the border of Ecuador and Colombia: a retrospective study of the population structure of Mycobacterium tuberculosis in Esmeraldas province. Front Public Health. (2024) 12:1343350. doi: 10.3389/fpubh.2024.1343350
50. Alame Emane, AK, Guo, X, Takiff, HE, and Liu, S. Highly transmitted M. tuberculosis strains are more likely to evolve MDR/XDR and cause outbreaks, but what makes them highly transmitted? Tuberculosis. (2021) 129:102092. doi: 10.1016/j.tube.2021.102092
51. Ford, CB, Shah, RR, Maeda, MK, Gagneux, S, Murray, MB, Cohen, T, et al. Mycobacterium tuberculosis mutation rate estimates from different lineages predict substantial differences in the emergence of drug-resistant tuberculosis. Nat Genet. (2013) 45:784–90. doi: 10.1038/ng.2656
52. Arenas-Suarez, NE, Cuervo, LI, Avila, EF, Duitama-Leal, A, and Pineda-Peña, AC. The impact of immigration on tuberculosis and HIV burden between Colombia and Venezuela and across frontier regions. Cad Saude Publica. (2021) 37:e00078820. doi: 10.1590/0102-311X00078820
Keywords: Mycobacterium tuberculosis, RDRio subfamily, Spoligotyping, MIRU-VNTR, tuberculosis, Ecuador
Citation: Castro-Rodriguez B, Franco-Sotomayor G, Benitez-Medina JM, Cardenas-Franco G, Jiménez-Pizarro N, Cardenas-Franco C, Aguirre-Martinez JL, Orlando SA, Hermoso de Mendoza J and Garcia-Bereguiain MA (2024) Prevalence, drug resistance, and genotypic diversity of the RDRio subfamily of Mycobacterium tuberculosis in Ecuador: a retrospective analysis for years 2012–2016. Front. Public Health. 12:1337357. doi: 10.3389/fpubh.2024.1337357
Edited by:
Andrea Gori, University of Milano-Bicocca, ItalyReviewed by:
Richard Salvato, State Center for Health Surveillance, BrazilCopyright © 2024 Castro-Rodriguez, Franco-Sotomayor, Benitez-Medina, Cardenas-Franco, Jiménez-Pizarro, Cardenas-Franco, Aguirre-Martinez, Orlando, Hermoso de Mendoza and Garcia-Bereguiain. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Miguel Angel Garcia-Bereguiain, bWFnYmVyZWd1aWFpbkBnbWFpbC5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.