 Carlos Eduardo González-Penagos1
Carlos Eduardo González-Penagos1 Jesús Alejandro Zamora-Briseño1
Jesús Alejandro Zamora-Briseño1 Daniel Cerqueda-García1
Daniel Cerqueda-García1 Monica Améndola-Pimenta1
Monica Améndola-Pimenta1 Juan Antonio Pérez-Vega1Emanuel Hernández-Nuñez1,2
Juan Antonio Pérez-Vega1Emanuel Hernández-Nuñez1,2 Rossanna Rodríguez-Canul1*
Rossanna Rodríguez-Canul1*- 1Departamento de Recursos del Mar, Centro de Investigación y de Estudios Avanzados del Instituto Politécnico Nacional-Unidad Mérida, Mérida, Mexico
- 2CONACYT – Centro de Investigación y de Estudios Avanzados del Instituto Politécnico Nacional, Mérida, Mexico
Crude oil spills have caused substantial impacts to aquatic ecosystems. Chemical dispersants are used to palliate the impact of oil spillages, but their use is polemic due to their additional potential toxic effect when mixed with oil-derived components. In this work, we used a 16S-based metagenomic approach to analyze the changes of the gut microbiota of adult zebrafish (Danio rerio) exposed to the water accommodated fraction (WAF) of a light crude oil (35° API gravity), and the chemically enhanced WAF (CEWAF), prepared with Nokomis 3-F4® dispersant. After 96 h of exposure, WAF induced an increase in the alpha and beta diversity, altering the relative abundance of Vibrio, Flavobacterium, and Novosphingobium. In contrast, CEWAF only caused an increase in the beta diversity, and an enrichment of the genus Pseudomona. Both treatments diminished the abundances of Aeromonas, Cetobacterium, Coxiella, Dinghuibacter, and Paucibacter. Moreover, the co-occurrence network among genera was more complex in WAF than in CEWAF, indicating a greater bacterial interaction in response to WAF. Our results indicate that short-term exposure to WAF and CEWAF can induce a dysbiosis in the gut microbiota of D. rerio, but these changes are specific in each treatment.
Introduction
Crude oil extraction activities in the Gulf of Mexico (GoM) are source of pollutants (1–3). Crude oil is a complex mixture of low and high molecular weight hydrocarbons, which represent up to 75% of its total composition and includes aliphatic and polycyclic aromatic hydrocarbons (PAHs) (4). In the non-hydrocarbon fraction, crude oil also contains nitrogen, sulfur, oxygen-derived compounds, as well as trace concentrations of heavy metals, such as nickel and iron (5).
Additional to oil pollutants associated to the natural release of petroleum (Spanish = chapopoteras), and those related to anthropogenic activities, hydrocarbons can enter in marine ecosystems during oil spills, as that occurred during the blowout of the Macondo well-operated by the Deepwater Horizon platform (DWH) in 2010. In that event, ~4.4 × 106 oil barrels were released into the sea for 84 days, producing a wide negative impact in the GoM at different biological levels (3).
During oil spill events, chemical dispersants (composed predominantly by surfactants and/or chemical solvents), are frequently used to reduce the tension in the surface of spilled oil by emulsifying their surface and increasing the oil-water solubility (6). For example, during the DWH disaster, ~1.5 M gallons of the chemical dispersant Corexit EC9500® were applied (6). However, the toxicity of the resulting emulsified hydrocarbons involves ecological alterations (7), and constitute a major threat to the environment, especially when they reach coastal areas (5). Another commonly used dispersant is Nokomis 3-F4® (Mar-Len Supply, Inc., Hayward, CA), which is one of the several commercially available formulations recommended for oil spill events, by the United States Environmental Protection Agency (US EPA) (8). The public available composition of this chemical dispersant is very limited, because its formulation is subjected to industrial secret protection (https://www.epa.gov/emergency-response/nokomis-3-F4), but its use is recommended in oil spills produced in fresh water or marine environments. However, some reports have addressed that Nokomis 3-F4® exposure interacts with the estrogen receptor (ER) and the androgen receptor (AR) of some marine organisms (6). This is because, Nokomis 3-F4® contains nonylphenol ethoxylate (NPE) that is degraded to 4-nonylphenol (NP), known as xenoestrogen or endocrine disruptor. This is a compound capable of acting on the endocrine system and altering the reproductive cycle (6). Also, NP can trigger oxidative stress, and it has been associated with obesity-related disorders in several animal models (9, 10).
Crude oil-derived compounds have been associated to carcinogenic, mutagenic, and teratogenic alterations in native aquatic biota (4, 5). Moreover, the exposure to these compounds can negatively impact the symbiotic interactions between host and its associated microorganisms, such as those occupying the gut space.
The gut microbiota maintains a symbiotic relationship with the host (11–13), participating in relevant functions including host metabolism, and immunity (14–16). Microbiota also regulates the function of the intestinal barrier because by having a highly specific composition (16). However, this relationship can be altered either by intrinsic factors of the host and/or by the surrounding environment (17, 18). The imbalance in the gut microbiota, lead to alteration of the host's homeostasis producing a phenomenon known as dysbiosis (17). Changes in the gut microbiota have been used as indicator of chemicals exposure (19–21), since the toxicity of xenobiotics can be modulated after bacterial metabolization (22, 23). In fish, the gut microbiota has been used to assess the effect to exposure of several pollutants, included crude oil exposure (24, 25), antibiotics (26), or PAHs (27), among others. With respect to the study of crude oil, there is still an incomplete comprehension of the effect of crude oil in interaction with chemical dispersants on the gut microbiota of fish. Thereby, we considered that the use of model fish species could help to elucidate in fine detail changes occurring in the gut microbiota in response to crude oil-derived components.
Zebrafish Danio rerio is a good candidate in toxicological studies because it possess several advantages; it can be easily manipulated because of its small size, its short generational time, its large number of offspring per laying, as well as the transparency of eggs and embryos (28, 29). It has been used as a model organism for the discovery of pharmacological targets, as well as toxicological evaluation of heavy metals, pesticides, fungicides, nanomaterials, and many other substances. In addition, the ecological dynamics of its gut microbiota communities is well-known (11, 30, 31).
For these reasons, the aim of this study was to evaluate the acute exposure to the water-accommodated fraction (WAF) of a light crude oil, and to the chemically enhanced WAF (CEWAF) with Nokomis 3-F4® on the composition of the gut microbiota of zebrafish.
Materials and Methods
Biological Material
One-year-old zebrafish (D. rerio) were obtained from the aquaculture facilities of the Center for Research and Advanced Studies of the National Polytechnic Institute-Merida Unit (CINVESTAV-IPN). Fish were kept in UV-treated freshwater in 1 L glass containers with dechlorinated tap water at 27.5 ± 0.5°C and continuous aeration. Fish were fed twice per day ad libitum with a commercial fish diet (Wardley®) and maintained at a photoperiod of 14:10 h of light: dark. Previously, this study was reviewed and approved by the Institutional Animal Care and Use Committee of the Center for Research and Advanced Studies (CICUAL-CINVESTAV, approval number: 2875). It complies with the Mexican Official Norm (NOM-062-ZOO-1999), “Technical Specifications for the Care and Use of Laboratory Animals,” as well as all applicable federal and institutional regulations.
WAF and CEWAF Preparation for Exposure Assays
Light crude oil (extracted from Campo Pool oil-well with 35°API gravity) and Nokomis 3-F4® dispersant were provided by PEMEX Exploration and Production Company. For the exposure assays, we followed the ECETOC (European Center for Ecotoxicology and Toxicology of Chemical Compounds) and the CROSERF (Chemical Response to Oil Spills -Ecological Effects Research Forum) recommendations for the use of water-accommodated fraction (WAF) and chemically enhanced WAF (CEWAF). We followed the method described by Singer et al. (32), with the adaptations proposed by Barron and Ka'aihue (33). WAF was defined as a medium that contains only a soluble fraction of oil that remains in aqueous phase (34). Also, when a dispersant is added to the crude oil-water mixture, a chemically enhanced WAF or CEWAF is obtained. In both cases, only the aqueous phase is used (34).
The WAF and CEWAF mixtures were prepared at the same time. For WAF preparation, crude oil (1 g/L) was added to the filtered, UV-light purified water and placed in a glass jar. For CEWAF preparation, crude oil (1 g/L) was added to the filtered, UV-light purified water and placed in a glass jar. Nokomis 3-F4® was immediately added in a proportion of 1:10 (v/v, dispersant:crude oil).
Both mixtures were prepared in darkness conditions, mixed with a magnetic stirrer for 24 h at 5,000 x rpm to form a vortex equivalent to the 20–25% of the water column height. After this time, the mixtures were let to settle for 1 h, and the lower phase (aqueous phase) for each was collected. These phases were defined as the WAF and CEWAF stock solutions. Then, a 50% dilution (v/v) was prepared from both stock solutions (equal volumes of filtered, UV-light treated water and stock WAF/CEWAF). These dilutions were used for the WAF and CEWAF exposure bioassay, respectively. This sub-lethal concentration of 50% was chosen based on previous data (24).
WAF/CEWAF Exposure Assays
A 96 h acute static bioassay was performed using 12 male adult zebrafish. Three groups of four zebrafish with similar length and weight were placed individually in 1 L glass aquaria (WAF = 4, CEWAF = 4, and CONTROL = 4). During the bioassay, the zebrafish were kept unfed. Water quality parameters (temperature, dissolved oxygen, salinity, and pH) were measured using an YSI™ 556MPS Multi Probe System multiparameter device (Xylem Inc). In addition, nitrite and ammonium were quantified with an aquaria water quality kit (Nutrafin®). At the end of the exposure time, biometric measures were taken, and the organisms were euthanized by ice water bath immersion for five min. Each fish was surface sanitized with 70% ethanol, and intestines were dissected under aseptic conditions, fixed in five volumes of absolute ethanol, and stored at −80°C until analysis.
DNA Extraction and 16S rRNA Gene Amplification
Total genomic DNA (gDNA) extraction from the entire intestine of each fish (n = 4 per treatment) was performed using the commercial Quick-DNA™ Universal Kit (ZymoResearch©). gDNA concentration was determined with a Thermo Scientific NanoDrop™ 2000c spectrophotometer (ThermoFisher Scientific©) and integrity checked by 1% agarose gel electrophoresis.
The V3-V4 region of the 16S rRNA bacterial gene was amplified by PCR from the gDNA of each sample. For PCR amplification, we used the 16S rRNA Forward primer: 5' TCGTCGGCAGCGTCAGATGTGTATAAGAGA
CAGCCTACGGGNGGCWGCAG and 16S rRNA Reverse primer: 5' GTCTCGTGGGCTCGGAGATGTGTATAAGAGACAGGA
CTACHVGGGTATCTAATCC, which amplify a region of ~550 bp (35). PCRs were performed in a volume of 25 μL containing 12.5 μl of Dream Taq Green PCR Master Mix (2X) (Thermo Scientific©), 0.5 μL of gDNA (equivalent to 328 ± 23 ng/μl of gDNA), and 0.115 μL of each primer (125 nM). All reactions were performed in a Thermal cycler C1000 Touch™ (Bio-Rad Laboratories©) using a cycling program as follows: initial denaturation of 3 min at 95°C followed by 36 cycles of denaturation for 30 s at 95°C, annealing of 30 s at 53°C, an extension of 60 s at 72°C, and a final extension of 5 min at 72°C. PCR for no-template control was included to guarantee that no cross-tube reactions occurred.
Library Preparation and Sequencing
Library preparation and sequencing were performed in the National Sequencing and Polymorphisms Detection Unit from the National Institute of Genomic Medicine (INMEGEN), Mexico.
Amplicons clean up were carried out using AMPure XP® beads, Beckman-Coulter©. Then, amplicons were indexed with the Illumina sequencing adapters using the Nextera XT Index Kit® (Illumina©) followed by a purification step in the same conditions. Concentration of each indexed amplicon was assessed by Qubit 2.0 Fluorometer (Thermo Scientific©), while its quality was evaluated by high-resolution automated electrophoresis (Agilent Bioanalyzer 2100®). Indexed amplicons were sequenced in a paired-end (2 × 300 bp) sequencing format with a MiSeq Reagent Kit V3® (600 cycles), using the MiSeq platform (Illumina©).
Bioinformatics Analysis
Paired end reads 2 × 300 were processed with the QIIME2 pipeline (36). Demultiplexed fastq files were processed with the DADA2 plugin to resolve the amplicon sequence variants (ASVs) (37). Reads were trimmed at position 20 of the 5' end and truncating in position 280 in the 3' end for both forward and reverse reads. Chimeric sequences were removed with the “consensus” method. The taxonomy of each representative sequences of the ASVs was assigned using the QIIME plugin feature-classifier classify-consensus-vs-search (v 2.9.0) (38), using the SILVA database (version 132). The representative sequences of the ASVs were aligned with the MAFFT algorithm (39).
After the masking by positional conservation and gap filtering, a tree was built with the FastTree algorithm (40). The mitochondrial ASVs were removed and the feature table was rarefied at a sequencing depth of 9,800 reads per sample after verifying the correct sample effort by an accumulation curve. The feature table and tree were exported to the R environment v. 3.6.0 (http://www.R-project.org/) and the statistical analyses were performed with the phyloseq v. 3.6.0 (41), ggplot2 v. 3.3.0 (42), and vegan packages v. 2.5.6 (43).
First, beta diversity Permutational Analysis of Variance (PERMANOVA) test with 1,000 permutations with the weighted UniFrac distance was carried out to assess significant differences among treatments. Second, a Principal Coordinate Analysis (PCoA) on weighted UniFrac distance was calculated (44). Third, richness and alpha-diversity were calculated using observed ASVs, Shannon diversity measurements H', Simpson's metric, Chao1's metric, and abundance-based coverage estimator (ACE). Then, a linear discriminant analysis (LDA) effect size (LEfSe) (45) was performed at the ASV level to identify the microbial taxa with differential abundances among treatments, using a LDA cut-off > 2 and a Kruskal-Wallis alpha value p < 0.05. Finally, a co-occurrence network analysis was performed by pairwise comparison from the LEfSe results with a correlation analysis using the SparCC software (46) and analyzed using Cytoscape v3.7.2 (47).
Data Deposition
Raw sequences from 16S rRNA gene profiling are available in the NCBI SRA database with access numbers: SAMN13874135; SAMN13874136; SAMN13874137; SAMN13874138; SAMN13874139; SAMN13874140; SAMN13874141; SAMN13874142; SAMN13874131; SAMN13874132; SAMN13874133 and SAMN13874134; all under the NCBI BioProject PRJNA601771.
Hydrocarbon Quantification
A sample of the water column was taken for WAF and CEWAF at the beginning (0 h) and at the end (96 h) of the experiment. Total hydrocarbons, aliphatic (C10 - C40) and PAHs, including 16 US EPA priority PAHs were quantified in WAF and CEWAF following the method of Wang et al. (48). Prior to extraction, samples were enriched with 100 μl of the following standards: biphenyl d10, phenanthrene d10, chrysene d12, benzo(a)pyrene d12 (10 mg/mL), and o-terphenil (200 mg/mL). In each set of samples, a technical blank and a duplicate sample were added. Identification and quantification of the compounds was carried out with standards from Ultra Scientific® in the case of the PAHs and from Chiron© for deuterated PAHs. Total hydrocarbons were analyzed with an Agilent 7890A© gas chromatograph equipped with an FID detector. PAHs were analyzed with a Perkin-Elmer© gas chromatograph equipped with a Clarus 500® mass-selective detector using a 30 m × 0.25 mm (i.d.) x 0.25 DB-5 MS fused silica capillary column (J & W Scientific©), operating in the selected ion monitoring (SIM) mode, calibrations were verified daily, and the calibration curves were carried out for each set of samples.
Statistical Analysis
The total weigh and length of each fish were measured at the beginning and at the end of the experiment (0 and 96 h, respectively), and analyzed by a two-tailed Student's t-test with a p < 0.05. The metrics of observed ASVs, Shannon and Simpson alpha-diversity indices, Chao1 and ACE richness measurements were tested for significance differences between groups at 96 h exposure. For this, the normality of the data was tested by the Shapiro–Wilk test and a two-tailed Student's t-test was performed for parametric data using InfoStat software (49). All data are reported as a mean ± standard error (SEM) and *p < 0.05 and **p < 0.01 were considered statistically significant.
Results
General Information and D. rerio Biometrics
The acute 96 h exposure to WAF and CEWAF treatments did not alter the total length or weight of fish among groups or between times (p > 0.05) (Supplementary Table 1). Water quality parameters were constant along the experiment, including nitrites and ammonium concentrations ( = not detected, = 0.03 ± 0.0 mg/L), temperature (27 ± 0.5°C), dissolved oxygen (DO = 5.50 ± 0.25 mg/L), salinity (S = 0.0 UPS), and hydrogen potential (pH = 5.5 ± 0.0).
Sequencing Data Analyses
After sequencing the V3-V4 region of the rRNA gene, a total of 1,611,258 reads were obtained from the 12 libraries. After cleaning the raw data, 502,245 reads remained for statistics analysis (Supplementary Table 2A). The percentage of mitochondria detected in the sequencing data was 6%, while the number of chimeric sequences was 3% of the total raw reads.
Effects of WAF and CEWAF on Richness and Alpha Diversity of Gut-Associated Microbiota of D. rerio
We used rarefaction curves to estimate the correct sampling effort. According to this analysis, all libraries possess a sufficient sequencing depth because all of them reached the plateau phase at 9,800 reads (Figure 1A). Richness and diversity of the gut-associated microbiota of zebrafish were compared among groups. To estimate richness and coverage, Chao1 and ACE indices were used, respectively. Both indices indicated a higher richness for WAF treatment in comparison with CEWAF and CONTROL, with only statistical differences between WAF and CONTROL (p < 0.05) (Table 1). In contrast, CEWAF showed a greater variability without significant differences when it was compared with the other groups (Table 1).
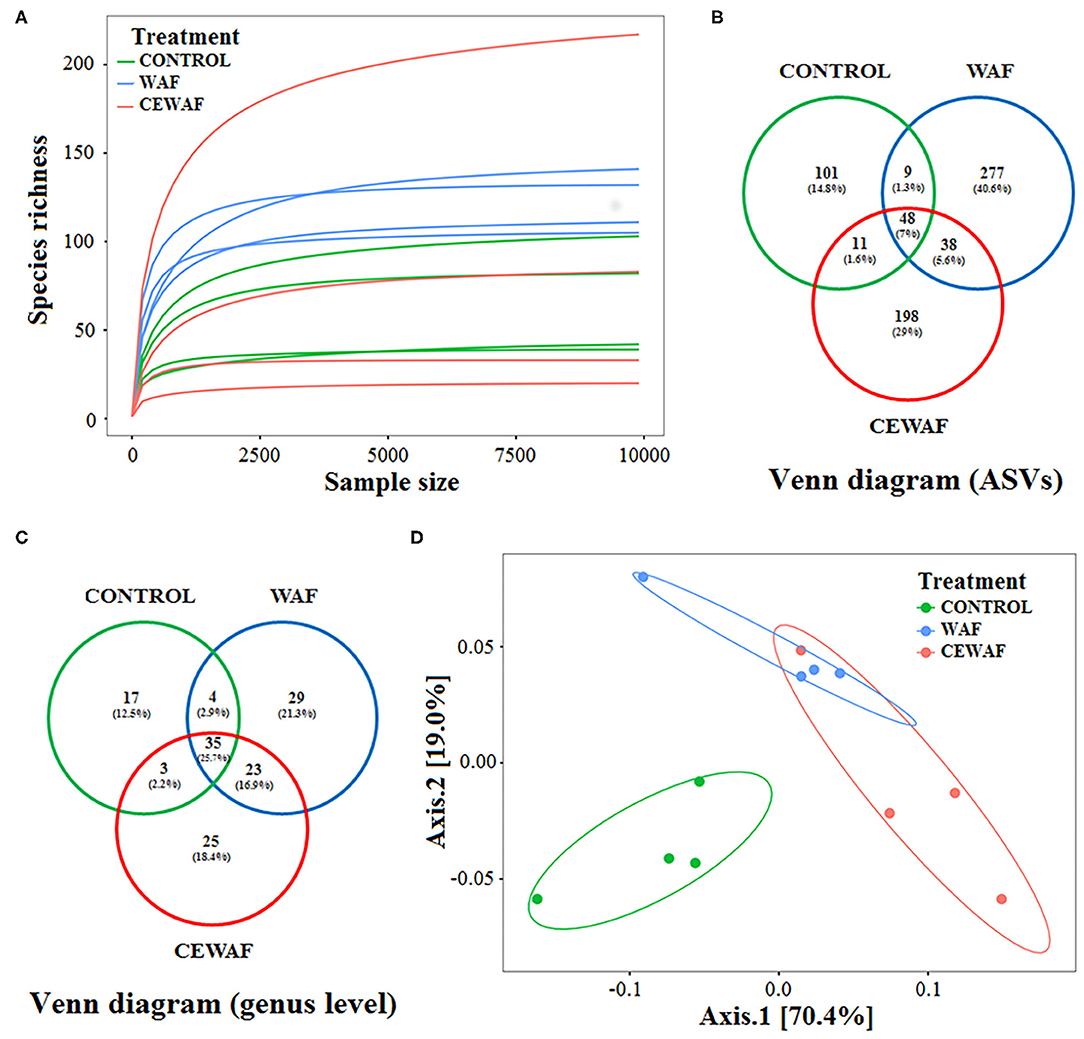
Figure 1. Main statistics of biodiversity analysis. (A) Rarefaction curves of ASVs. (B,C) Venn diagram of the ASVs that are shared among the groups. (D) UniFrac PCoA plot showing the separation among sample groups exposed to WAF (blue dots) and CEWAF (orange dots) respect to CONTROL (green dots) group.
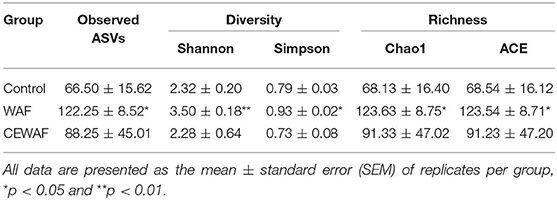
Table 1. The diversity and richness indexes of the gut microbiota in zebrafish in response to WAF, CEWAF, and control group.
A total of 682 ASVs were obtained (Figure 1B; Supplementary Table 2B). For CONTROL group, 169 ASVs (24.7%) were registered; for WAF treatment, we identified 372 (54.5%) ASVs, and for CEWAF treatment, a total of 295 (43.2%) ASVs were found (Figure 1B; Supplementary Table 2B), and the classification at genus level is shown in Figure 1C.
Effects of WAF and CEWAF on the Structure of Gut Microbiota of D. rerio
The composition of the gut microbiota of each group was compared using a PCoA analysis based on weighted UniFrac distance among samples (Figure 1D). The two axes of PCoA explained 89.4% of the total variance in bacterial composition, showing a clear separation between ASVs abundances from the gut microbiota of WAF and CEWAF treatments in relation to the CONTROL. However, PCoA did not show a clear separation between WAF and CEWAF. Paired-PERMANOVA showed differences between WAF and CONTROL (F = 13.35, R2 = 0.69, p < 0.05), and between CEWAF and CONTROL (F = 13.74, R2 = 0.69, p < 0.05). In contrast, no difference was found between WAF and CEWAF (F = 3.38, R2 = 0.36, p = 0.11).
Effects of WAF and CEWAF on the Composition of Gut Microbiota of D. rerio
A total of 32 bacterial genera with a relative abundance > 1% were identified from all the libraries (Figure 2). The information about the absolute and relative abundances of each genera is described in the Supplementary Tables 2C,D.
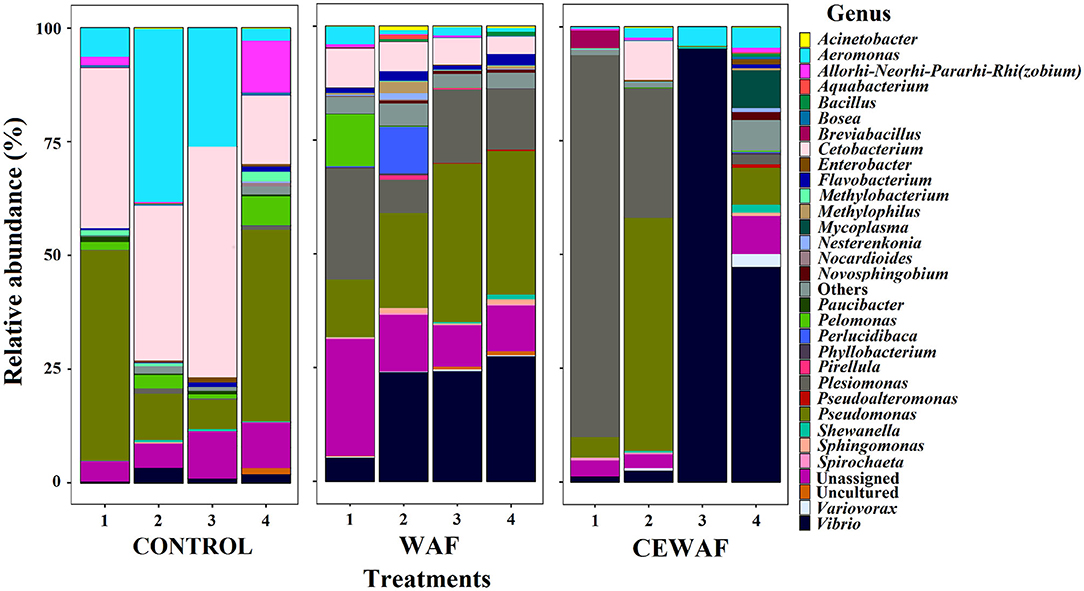
Figure 2. The relative abundance of ASVs classified at genus level (>1%).
Considering the relative abundance, the two major phyla were Proteobacteria (CONTROL: 64.30%, WAF: 83.89%, and CEWAF: 90.90%), and Fusobacteria (CONTROL: 33.73%, WAF: 6.16%, and CEWAF: 2.14%). Chlamydiae (0.04%) was the only phylum specifically found in CONTROL. For WAF and CEWAF, three phyla were found exclusively shared by these treatments: Spirochaetes (WAF: 0.29% and CEWAF: 0.19%), Cyanobacteria (WAF: 0.02% and CEWAF: 0.08%), and Verrucomicrobia (WAF: 0.02% and CEWAF: 0.05%). Finally, Patescibacteria phylum (0.003%) was found only in CEWAF.
The top-three of the genera with highest relative abundance per group were Cetobacterium (33.7%), Pseudomonas (26.2%) and Aeromonas (18.2%) for CONTROL; Pseudomonas (24.9%), Vibrio (20.2%) and Plesiomonas (15.2%) for WAF, and Vibrio (36.5%), Plesiomonas (28.5%) and Pseudomonas (15.9%) for CEWAF (Supplementary Table 2D). The ASVs were distributed among 136 bacterial genera (Figure 1C and Supplementary Table 2B), and the specific genera per treatment are listed in Supplementary Table 2E.
Differential Abundance Analysis on Microbial Composition of D. rerio Gut
We performed a LEfSe analysis to find the ASVs with differential abundance among the groups (Figure 3A) and by pairwise comparison (Supplementary Figures 1A,C, 2A). Using this approach, 22 representative ASVs were identified among the three conditions, 17 belonging to 9 genera (Pseudomonas, Cetobacterium, Aeromonas, Paucibacter, Flavobacterium, Dinguibacter, Coxiella, Vibrio, and Novosphingobium, while 5 ASVs were unassigned genera (Figure 3A).
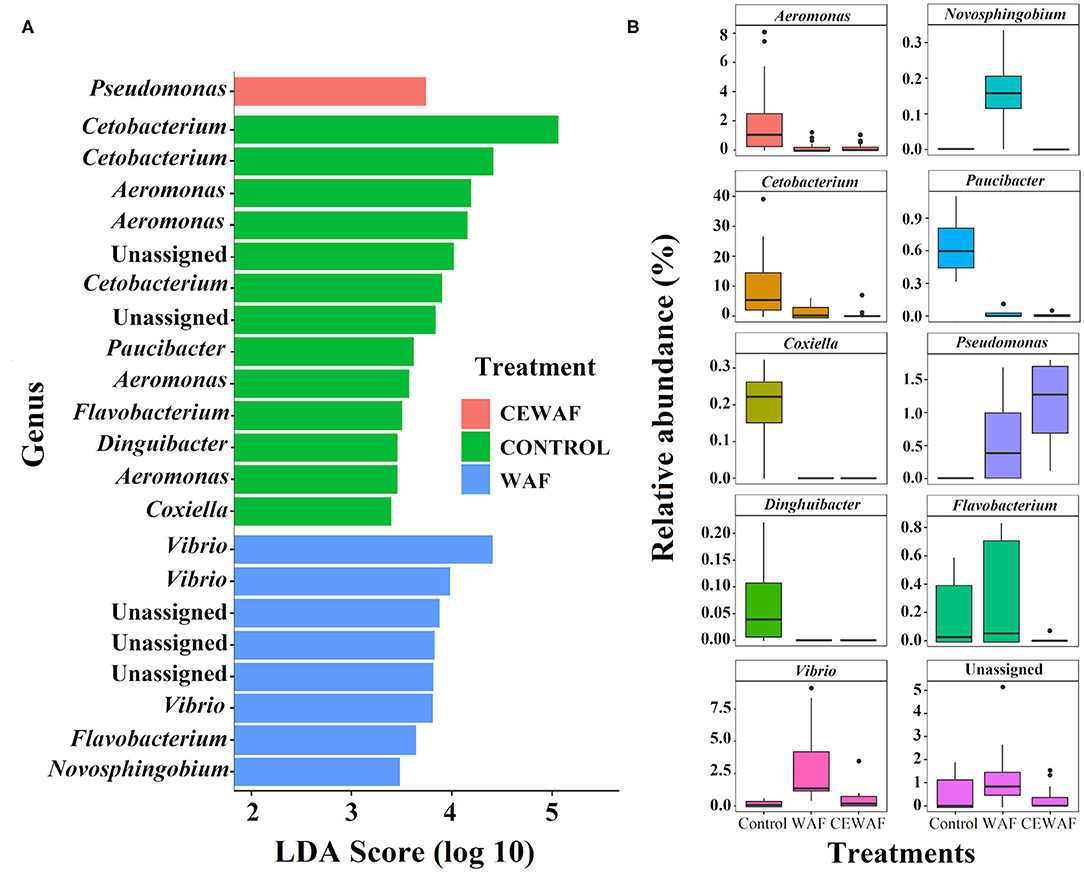
Figure 3. Statistically different phylotypes of the gut-microbiome of D. rerio according to LEfSe analysis. (A) LDA scores computed for differential ASV's compared among the three groups and represented at genus level. (B) The relative abundance of the identified differential ASVs at genus level for each group.
The changes in relative abundances of genera among groups (Figure 3B) and between groups (Supplementary Figures 1B,D, 2B) are shown, and a description of the data analysis are indicated in Supplementary Tables 2F,G,H).
Finally, the phylogenetic analysis performed with the ASVs detected by LefSe analysis between groups identified the closest related species for the ASVs with differential abundance, Supplementary Figure 3 for WAF and Supplementary Figure 4 for CEWAF.
Microbial Correlation Network Analyses
The co-occurrence networks based on the differential ASVs show the positive (co-existence) and negative (co-exclusion) interactions of the gut microbiota in response to WAF (Figure 4A) and CEWAF (Figure 4B) treatments. A total of 282 interactions were found (138 negatives and 144 positives) with 36 nodes for WAF (Figure 4A). For CEWAF, we found 100 interactions (30 negatives and 70 positives), with 24 nodes (Figure 4B). The total interactions for the bacterial communities of WAF and CEWAF treatments are shown in the Supplementary Tables 2I,J, respectively.
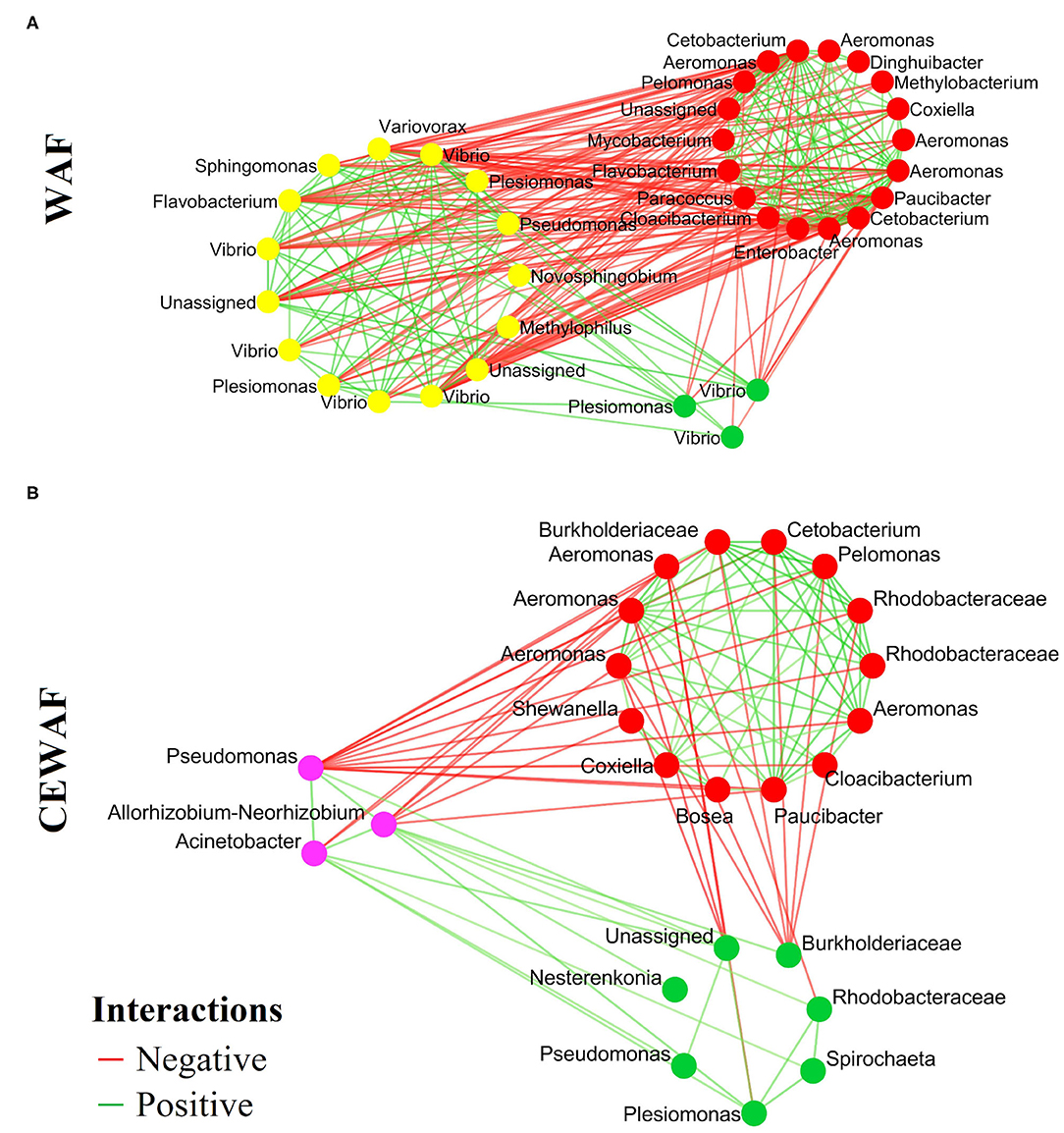
Figure 4. Co-occurrence networks with the ASVs identified in the pairwise LEfSe analysis. Each node means the statistically different genera or family for each condition. (A) Representative genera for WAF (yellow dots) and (B) CEWAF (purple dots). The red means a negative interactions and the green means a positive interaction among bacterial groups.
Quantification of Hydrocarbons in WAF and CEWAF
The hydrocarbon quantification of WAF and CEWAF was performed at the initial time (0 h) and at the end (96 h) time of the experiment. Four PAHs (naphthalene, 2-methylnaphthalene, 1-methylnaphthalene, and perylene) were quantified for WAF at 0 h with a total concentration of 1.21 μg/L, and for CEWAF, only one PAH (perylene) was detected with a concentration of 0.18 μg/L. In contrast, these PAH were not detected at 96 h. Detailed information on the quantification of each hydrocarbon compound detected at 0 h and 96 h can be found in the Supplementary Table S3.
Discussion
In this study, we investigated if an exposure to an environmentally relevant concentration of WAF and/or CEWAF could disturb the gut microbiota of D. rerio (Supplementary Figure 5). This sort of studies demonstrate that the gut microbiota of fish can be useful in toxicological studies, additional to findings provided in metagenomics analyses from the water-column and sediments (21–23, 50). Previous studies have reported that the basal gut microbiota of D. rerio is dominated by the phyla Proteobacteria, and the prevalent presence of Firmicutes and Fusobacteria (11, 51). In this study the CONTROL group harbored the same phyla and were the most abundant. Interestingly, their relative abundance was affected by WAF and CEWAF treatments.
Here, we observed that WAF exposure caused changes in richness, abundance, alpha and beta diversity, dominance, and co-occurrence networks of the gut microbiota. In contrast, CEWAF only impacted the beta diversity, abundance, as well as the interactivity among bacterial taxa. However, both treatments generated dysbiosis. This is because the changes in the alpha diversity appears to be the most consistent indicator of intestinal dysbiosis (52). As well as the increase of the inter-individual variability in the microbiota structure (53–55), that we observed in WAF and CEWAF treated samples, respectively.
We also observed a reduction in the relative abundances of putative beneficial genera. For example, Cetobacterium is recognized as a vitamin B12 provider for its host (56). Also, Lactobacillus is considered beneficial in the regulation of intestinal functions by secreting metabolites and altering the pH to prevent the settlement of harmful bacteria (57). The reduction of these taxa such as those reported herein, could imply an important loss of resilience in the host microbiota (58, 59). In contrast, Vibrio, Acinetobacter, Streptococcus, Flavobacterium, Plesiomonas, and Pseudomonas genera are frequently considered pathogenic genera for this model (60–62). Interestingly, their relative abundance increased in both treatments, except for Flavobacterium that only increased in WAF treatment. However, Vibrio and Pseudomonas genera have been reported in the degradation of aromatic compounds (63–65). For example, after the DWH disaster, some members of the Pseudomonas genus appeared to be dominant during the oil degradation stage, when the proportions of aliphatic compounds were higher (66). In addition, previous work evaluating surfactants reported an increase in Pseudonomas in soils contaminated with hydrocarbons (67).
It is noticeable that although the exposure time was very short (96 h), the 50% WAF treatment was enough to promote an increase in the relative abundance of genera with reported hydrocarbonoclastic capabilities such as, Acinetobacter, Flavobacterium, Klebsiella, Vibrio, Staphylococcus and Shewanella (68–70). Though, Achromobacter was found only increased in response to WAF treatment, this genus has been found in the degradation of hydrocarbons such as n-alkanes and PAHs (71), while Bacosa et al. (72) described that it uses some metabolites from aromatic hydrocarbons.
Another bacterium that increased its abundance in both treatments was Bacillus, with some members capable to metabolize hydrocarbons. Bacillus is commonly found in crude oil-affected marine areas (73, 74). Likewise, Acinetobacter increased in both treatments, and several members of this genus has been proposed as a key player in PAHs degradation processes (75). Also, Sphingomonas were reported to have PAHs degradation abilities (76). Burkholderia was found only in WAF and CEWAF. This genus has been reported of having a grading capacity to degrade heavy crude oil (77). Thus, the increase of its abundance in response to light crude oil warrants further investigations because the genus Burkholderia has over 90 species reported, and it is divided into two major groups phylogenetically distant. The first group is composed of pathogenic species which highlights referred opportunistic pathogens such as Burkholderia cepacian (Bcc) complex, and the other group consists of non-pathogenic species with skills to promote plant growth and rhizoremediation (77).
On the other hand, Muricauda was found exclusively in organisms exposed to CEWAF. It has been reported as a degrader of aliphatic hydrocarbons (78), as well as Cycloclasticus and Oleiphilus, were found only in response to CEWAF. These genera have been reported as oil degraders and are frequently found in polluted marine environments (79, 80).
The co-occurrence patterns provide a new perspective to understand the structure of complex microbial communities (26, 81, 82). The bacterial interactions of the differential genera reported herein indicate that both treatments (WAF and CEWAF) can alter in different way the interactions among bacteria. This analysis suggests a greater interactivity in the gut microbiota of D. rerio exposed to WAF with respect to CEWAF. This feature in associated with a greater number of bacterial groups, specifically those with putative hydrocarbonoclastic activity, such as Flavobacterium, Pseudomonas, Novosphingobium, Sphingomonas, Vibrio, Methylophilus, Plesiomonas, and Variovorax (83–85). According to our data, it is likely that as a mechanism of defense, the gut microbiota rearranges their capabilities to enhance its hydrocarbon assimilation capabilities.
In this way, representative groups, helped to increase the presence of other groups of bacteria, triggering a positive synergy with organisms with similar function (like the intake of hydrocarbons as energy source). The alterations observed herein suggest the establishment of an ecological succession of the microbiota that use crude-oil derived compounds, as described previously (80), and have an active role for the metabolism of hydrocarbons in the gut microbiota, during the crude-oil exposure (24). Despite this, it is likely that during this process of biotransformation of hydrocarbons in the gut, the resulting metabolites also increase harmful bacteria that would negatively affect fish health (86).
Our results also indicated that the addition of Nokomis 3-F4® was able to induce a differential assembly as well as bacterial interactions in fish gut microbiota with respect to exposure to WAF. In this sense, chemical surfactants not only affect the distribution of crude oil-derived compounds in the water column, but also can trigger a differential response at the gut microbiota level. Results from this study are encouraging and future studies should focus on the evaluation of the gut metabolic process that take place in response to WAF and CEWAF at different times of exposure and concentrations.
Conclusion
This is the first study evaluating the effects of light crude oil (WAF), and its mixture with Nokomis 3-F4® (CEWAF) on the gut microbiota of zebrafish, D. rerio. We observed that both treatments caused dysbiosis. We found changes in the diversity and abundances of gut microbiota for WAF and CEWAF treated groups. Moreover, these treatments triggered an increase in the abundance of hydrocarbonoclastic genera. These findings have environmental relevance regarding the assessment of the impact of acute exposure to water soluble compounds of crude oil and its mixture with chemical dispersant.
Data Availability Statement
The datasets generated for this study were uploaded to the Sequence Read Archive (SRA) of the National Center for Biotechnology Information (NCBI) database with access number of the BioProject PRJNA601771 with a release date at 2020-08-03.
Ethics Statement
The animal study was reviewed and approved by The animal study was reviewed and approved by the Institutional Animal Care and Use Committee of the Center for Research and Advanced Studies (Centro de Investigación y de Estudios Avanzados del IPN) (CICUAL-Cinvestav) (APPROVAL NUMBER: 2875). And comply with the applicable Mexican Official Norm (NOM-062-ZOO-1999), Technical Specifications for the Care and Use of Laboratory Animals, as well as all applicable federal and institutional regulations.
Author Contributions
CG-P, JZ-B, and RR-C: conceptualization and experimental design. CG-P, JZ-B, and DC-G: metagenomic analysis and data curation. CG-P, JZ-B, RR-C, MA-P, DC-G, and EH-N: writing and editing. CG-P and JP-V: sampling laboratory coordination. RR-C: project administration and funding acquisition. CG-P and JZ-B: formal analysis. CG-P, JZ-B, JP-V, and MA-P: methodology. RR-C and EH-N: supervision. CG-P, JZ-B, DC-G, and MA-P: visualization. All authors contributed to the article and approved the submitted version.
Funding
This research has been funded by the Mexican National Council for Science and Technology – Mexican Ministry of Energy – Hydrocarbon Fund, project 201441. This is a contribution of the Gulf of Mexico Research Consortium (CIGoM). We acknowledge PEMEX's specific request to the Hydrocarbon Fund to address the environmental effects of oil spills in the Gulf of Mexico. CG-P is holding a doctoral scholarship (reference number 628967) provided by the National Council of Science and Technology (CONACyT). Special thanks are conveyed to Richard Mena Loria for his support in zebrafish maintenance.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpubh.2020.584953/full#supplementary-material
Supplementary Figure 1. LEfSe analysis of the ASV shown significant differential genera between conditions. (A) ASV identified at genus level with differential abundance for WAF treatment. (B) Differential abundances between WAF and CONTROL. (C) ASV at genus level with differential abundance for CEWAF treatment. (D) Differential abundances between CEWAF and CONTROL.
Supplementary Figure 2. LEfSe analysis of the ASV shown differential genera between WAF and CEWAF. (A,B) ASVs at genus level with differential abundance.
Supplementary Figure 3. Phylogenetic tree of differential, unassigned genera of WAF (yellow) and CONTROL (blue) highlighting their closest relative species.
Supplementary Figure 4. Phylogenetic tree of differential, unassigned genera of CEWAF (purple) and CONTROL (blue) highlighting their closest relative species.
Supplementary Figure 5. Conceptual map referring to gut microbiota response of zebrafish exposed to WAF and CEWAF.
References
1. Overton EB, Ashton BM, Miles MS. Historical polycyclic aromatic and petrogenic hydrocarbon loading in Northern Central Gulf of Mexico shelf sediments. Mar Pollut Bull. (2004) 49:557–63. doi: 10.1016/j.marpolbul.2004.03.012
2. Kaiser MJ, Pulsipher AG. The impact of weather and ocean forecasting on hydrocarbon production and pollution management in the Gulf of Mexico. Energy Policy. (2007) 35:966–83. doi: 10.1016/j.enpol.2006.01.026
3. Crone TJ, Tolstoy M. Magnitude of the 2010 Gulf of Mexico oil leak. J Sci. (2010) 330:634. doi: 10.1126/science.1195840
4. Perrichon P, Le Menach K, Akcha F, Cachot J, Budzinski H, Bustamante P. Toxicity assessment of water-accommodated fractions from two different oils using a zebrafish (Danio rerio) embryo-larval bioassay with a multilevel approach. Sci Total Environ. (2016) 568:952–66. doi: 10.1016/j.scitotenv.2016.04.186
5. Zhang B, Matchinski EJ, Chen B, Ye X, Jing L, Lee K. Marine Oil Spills-Oil Pollution, Sources and Effects. World Seas: an Environmental Evaluation, Academic Press. (2019). p. 391–406. doi: 10.1016/B978-0-12-805052-1.00024-3
6. Judson RS, Martin MT, Reif DM, Houck KA, Knudsen TB, Rotroff DM, et al. Analysis of eight oil spill dispersants using rapid, in vitro tests for endocrine and other biological activity. Environ Sci Technol. (2010) 44:5979–85. doi: 10.1021/es102150z
7. Rico-Martínez R, Snell TW, Shearer TL. Synergistic toxicity of macondo crude oil and dispersant Corexit 9500A® to the Brachionus plicatilis species complex (Rotifera). Environ Pollut. (2013) 173:5–10. doi: 10.1016/j.envpol.2012.09.024
8. United States Environmental Protection Agency (US EPA). National Contingency Plan Product Schedule Toxicity and Effectiveness Summaries. Available online at: https://www.epa.gov/emergency-response/national-contingency-plan-product-schedule-toxicity-and-effectiveness-summaries#Dispersants (accessed June 20, 2020).
9. Okai Y, Sato EF, Higashi-Okai K, Inoue M. Enhancing effect of the endocrine disruptor para-nonylphenol on the generation of reactive oxygen species in human blood neutrophils. Environ Health Perspect. (2004) 112:553–6. doi: 10.1289/ehp.6584
10. Hao CJ, Cheng XJ, Xia HF, Ma X. The endocrine disruptor 4-nonylphenol promotes adipocyte differentiation and induces obesity in mice. Cell Physiol Biochem. (2012) 30:382–94. doi: 10.1159/000339032
11. Roeselers G, Mittge EK, Stephens WZ, Parichy DM, Cavanaugh CM, Guillemin K, et al. Evidence for a core gut microbiota in the zebrafish. ISME J. (2011) 5:1595–608. doi: 10.1038/ismej.2011.38
12. Lozupone CA, Stombaugh JI, Gordon JI, Jansson JK, Knight R. Diversity, stability and resilience of the human gut microbiota. Nature. (2012) 489:220–30. doi: 10.1038/nature11550
13. Udayangani RM, Dananjaya SH, Nikapitiya C, Heo GJ, Lee J, De Zoysa M. Metagenomics analysis of gut microbiota and immune modulation in zebrafish (Danio rerio) fed chitosan silver nanocomposites. Fish Shellfish Immun. (2017) 66:173–84. doi: 10.1016/j.fsi.2017.05.018
14. O'Hara AM, Shanahan F. The gut flora as a forgotten organ. EMBO Rep. (2006) 7:688–93. doi: 10.1038/sj.embor.7400731
15. Clarke G, Stilling RM, Kennedy PJ, Stanton C, Cryan JF, Dinan TG. Minireview: gut microbiota: the neglected endocrine organ. J Mol Endocrinol. (2014) 28:1221–38. doi: 10.1210/me.2014-1108
16. Feng Q, Chen WD, Wang YD. Gut microbiota: an integral moderator in health and disease. Front Microbio. (2018) 9:151. doi: 10.3389/fmicb.2018.00151
17. Rinninella E, Raoul P, Cintoni M, Franceschi F, Miggiano GAD, Gasbarrini A, et al. What is the healthy gut microbiota composition? A changing ecosystem across age, environment, diet, and diseases. Microorganisms. (2019) 7:14. doi: 10.3390/microorganisms7010014
18. Scott KP, Gratz SW, Sheridan PO, Flint HJ, Duncan SH. The influence of diet on the gut microbiota. Pharmacol Res. (2013) 69:52–60. doi: 10.1016/j.phrs.2012.10.020
19. Licht TR, Bahl MI. Impact of the gut microbiota on chemical risk assessment. Curr Opin Toxicol. (2019) 15:109–13. doi: 10.1016/j.cotox.2018.09.004
20. Claus SP, Guillou H, Ellero-Simatos S. The gut microbiota: a major player in the toxicity of environmental pollutants? Npj Biofilms Microbi. (2016) 2:1–11. doi: 10.1038/npjbiofilms.2016.3
21. Koppel N, Rekdal VM, Balskus EP. Chemical transformation of xenobiotics by the human gut microbiota. J Sci. (2017) 356:eaag2770. doi: 10.1126/science.aag2770
22. Adamovsky O, Buerger AN, Wormington AM, Ector N, Griffitt RJ, Bisesi J. The gut microbiome and aquatic toxicology: an emerging concept for environmental health. Environ Toxicol Chem. (2018) 37:2758–75. doi: 10.1002/etc.4249
23. Duperron S, Halary S, Gallet A, Marie B. Microbiome-aware ecotoxicology: relevance, pitfalls and challenges. Public Health Front. (2020) 8:407. doi: 10.3389/fpubh.2020.00407
24. Améndola-Pimenta M, Cerqueda-García D, Zamora-Briseño JA, Couoh-Puga D, Montero-Muñoz J, Árcega-Cabrera F, et al. Toxicity evaluation and microbiota response of the lined sole Achirus lineatus (chordata: achiridae) exposed to the light petroleum water-accommodated fraction (WAF). J Toxicol Env Heal A. (2020) 83:313–29. doi: 10.1080/15287394.2020.1758861
25. Cerqueda-García D, Améndola-Pimenta M, Zamora-Briseño JA, González-Penagos CE, Árcega-Cabrera F, Ceja-Moreno V, et al. Effects of chronic exposure to water accommodated fraction (WAF) of light crude oil on gut microbiota composition of the lined sole (Achirus lineatus). Mar Environ Res. (2020) 161:105116. doi: 10.1016/j.marenvres.2020.105116
26. Gaulke CA, Barton CL, Proffitt S, Tanguay RL, Sharpton TJ. Triclosan exposure is associated with rapid restructuring of the microbiome in adult zebrafish. PLoS ONE. (2016) 11:e0154632. doi: 10.1371/journal.pone.0154632
27. Brown-Peterson NJ, Krasnec MO, Lay CR, Morris JM, Griffitt RJ. Responses of juvenile southern flounder exposed to deepwater horizon oil-contaminated sediments. Environl Toxicol Chem. (2017) 36:1067–76. doi: 10.1002/etc.3629
28. Spitsbergen JM, Kent ML. The state of the art of the zebrafish model for toxicology and toxicologic pathology research—advantages and current limitations. Toxicol Pathol. (2003) 31(Suppl. 1):62–87. doi: 10.1080/01926230390174959
29. Dai YJ, Jia YF, Chen N, Bian WP, Li QK, Ma YB, et al. Zebrafish as a model system to study toxicology. Environ Toxicol Chem. (2014) 33:11–17. doi: 10.1002/etc.2406
30. Stephens WZ, Burns AR, Stagaman K, Wong S, Rawls JF, Guillemin, et al. The composition of the zebrafish intestinal microbial community varies across development. ISME J. (2016) 10:644–54. doi: 10.1038/ismej.2015.140
31. Cantas L, Sørby JRT, Aleström P, Sørum H. Culturable gut microbiota diversity in zebrafish. Zebrafish. (2012) 9:26–37. doi: 10.1089/zeb.2011.0712
32. Singer MM, Aurand D, Bragin GE, Clark JR, Coelho GM, Sowby ML, et al. Standardization of the preparation and quantitation of water-accommodated fractions of petroleum for toxicity testing. Mar Pollut Bull. (2000) 40:1007–16. doi: 10.1016/S0025-326X(00)00045-X
33. Barron MG, Ka'aihue L. Critical evaluation of CROSERF test methods for oil dispersant toxicity testing under subarctic conditions. Mar Pollut Bull. (2003) 46:1191–9. doi: 10.1016/S0025-326X(03)00125-5
34. Saco-Álvarez L, Bellas J, Nieto Ó, Bayona JM, Albaigés J, Beiras R. Toxicity and phototoxicity of water-accommodated fraction obtained from prestige fuel oil and marine fuel oil evaluated by marine bioassays. Sci Total Environ. (2008) 394:275–82. doi: 10.1016/j.scitotenv.2008.01.045
35. Klindworth A, Pruesse E, Schweer T, Peplies J, Quast C, Horn M, et al. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. (2013) 41:e1. doi: 10.1093/nar/gks808
36. Caporaso JG, Kuczynski J, Stombaugh J, Bittinger K, Bushman FD, Costello EK, et al. QIIME allows analysis of high-throughput community sequencing data. Nat Methods. (2010) 7:335–6. doi: 10.1038/nmeth.f.303
37. Callahan BJ, McMurdie PJ, Rosen MJ, Han AW, Johnson AJ, Holmes SP. DADA2: high-resolution sample inference from Illumina amplicon data. Nat Methods. (2016) 13:581–3. doi: 10.1038/nmeth.3869
38. Rognes T, Flouri T, Nichols B, Quince C, Mahé F. VSEARCH: a versatile open source tool for metagenomics. PeerJ. (2016) 4:e2584. doi: 10.7717/peerj.2584
39. Katoh K, Standley DM. MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Mol Biol Evol. (2013) 30:772–80. doi: 10.1093/molbev/mst010
40. Price MN, Dehal PS, Arkin AP. FastTree 2–approximately maximum-likelihood trees for large alignments. PLoS ONE. (2010) 5:e9490. doi: 10.1371/journal.pone.0009490
41. McMurdie PJ, Holmes S. phyloseq: an R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE. (2013) 8:e61217. doi: 10.1371/journal.pone.0061217
42. Ginestet C. ggplot2: elegant graphics for data analysis. J R Stat Soc A Stat. (2011) 174:245–6. doi: 10.1111/j.1467-985X.2010.00676_9.x
43. Oksanen J, Blanchet FG, Friendly M, Kindt R, Legendre P, McGlinn D, et al. Vegan: Community ecology package (2019). R Package Version 2:5-5.
44. Lozupone C, Knight R. UniFrac: a new phylogenetic method for comparing microbial communities. Appl Environ Microbiol. (2005) 71:8228–35. doi: 10.1128/AEM.71.12.8228-8235.2005
45. Segata N, Izard J, Waldron L, Gevers D, Miropolsky L, Garrett WS, et al. Metagenomic biomarker discovery and explanation. Genome Biol. (2011) 12:R60. doi: 10.1186/gb-2011-12-6-r60
46. Friedman J, Alm EJ. Inferring correlation networks from genomic survey data. PLoS Comput Biol. (2012) 8:e1002687. doi: 10.1371/journal.pcbi.1002687
47. Faust K, Raes J. CoNet app: inference of biological association networks using cytoscape. F1000 Res. (2016) 5:1519. doi: 10.12688/f1000research.9050.2
48. Wang Z, Fingas M, Li K. Fractionation of a light crude oil and identification and quantitation of aliphatic, aromatic, and biomarker compounds by GC-FID and GC-MS, part II. J Chromatogr Sci. (1994) 32:367–82. doi: 10.1093/chromsci/32.9.367
49. Di Rienzo JA, Casanoves F, Balzarini MG, Gonzalez L, Tablada M, Robledo CW. InfoStat Version 2018. Centro de Transferencia InfoStat, FCA: Universidad Nacional de Córdoba, Argentina.
50. Jin Y, Wu S, Zeng Z, Fu Z. Effects of environmental pollutants on gut microbiota. Environ Pollut. (2017) 222:1–9. doi: 10.1016/j.envpol.2016.11.045
51. Brugman S. The zebrafish as a model to study intestinal inflammation. Dev Comp Immunol. (2016) 64:82–92. doi: 10.1016/j.dci.2016.02.020
52. Mosca A, Leclerc M, Hugot JP. Gut microbiota diversity and human diseases: should we reintroduce key predators in our ecosystem? Front Microbiol. (2016) 7:455. doi: 10.3389/fmicb.2016.00455
53. Fjellheim AJ, Playfoot KJ, Skjermo J, Vadstein O. Inter-individual variation in the dominant intestinal microbiota of reared Atlantic cod (Gadus morhua L.) larvae. Aquacult Res. (2012) 43:1499–508. doi: 10.1111/j.1365-2109.2011.02952.x
54. Givens CE, Ransom B, Bano N, Hollibaugh JT. Comparison of the gut microbiomes of 12 bony fish and 3 shark species. Mar Ecol Prog. (2015) 518:209–23. doi: 10.3354/meps11034
55. Zaneveld JR, McMinds R, Thurber RV. Stress and stability: applying the anna karenina principle to animal microbiomes. Nat Microbiol. (2017) 2:17121. doi: 10.1038/nmicrobiol.2017.121
56. Tsuchiya C, Sakata T, Sugita H. Novel ecological niche of Cetobacterium somerae, an anaerobic bacterium in the intestinal tracts of freshwater fish. Lett Appl Microbiol. (2008) 46:43–8. doi: 10.1111/j.1472-765X.2007.02258.x
57. Pérez T, Balcázar JL, Ruiz-Zarzuela I, Halaihel N, Vendrell D, De Blas I, et al. Host–microbiota interactions within the fish intestinal ecosystem. Mucosal Immunol. (2010) 3:355–60. doi: 10.1038/mi.2010.12
58. Walter J. Ecological role of lactobacilli in the gastrointestinal tract: implications for fundamental and biomedical research. Appl Environ Microb. (2008) 74:4985–96. doi: 10.1128/AEM.00753-08
59. Azad M, Kalam A, Sarker M, Li T, Yin J. Probiotic species in the modulation of gut microbiota: an overview. Biomed Res Int. (2018) 2018:9478630. doi: 10.1155/2018/9478630
60. Austin B, Zhang XH. Vibrio harveyi: a significant pathogen of marine vertebrates and invertebrates. Lett Appl Microbiol. (2006) 43:119–24. doi: 10.1111/j.1472-765X.2006.01989.x
61. Janda JM, Abbott SL, McIver CJ. Plesiomonas shigelloides revisited. Clin Microbiol. (2016) 29:349–74. doi: 10.1128/CMR.00103-15
62. Egerton S, Culloty S, Whooley J, Stanton C, Ross RP. The gut microbiota of marine fish. Front Microbiol. (2018) 9:873. doi: 10.3389/fmicb.2018.00873
63. Uhlik O, Wald J, Strejcek M, Musilova L, Ridl J, Hroudova M, et al. Identification of bacteria utilizing biphenyl, benzoate, and naphthalene in long-term contaminated soil. PLoS ONE. (2012) 7:e40653. doi: 10.1371/journal.pone.0040653
64. Guazzaroni ME, Herbst FA, Lores I, Tamames J, Peláez AI, Lopez-Cortés N, et al. Metaproteogenomic insights beyond bacterial response to naphthalene exposure and bio-stimulation. ISME J. (2013) 7:122–36. doi: 10.1038/ismej.2012.82
65. Rosano-Hernández MC, Ramírez-Saad H, Fernández-Linares L. Petroleum-influenced beach sediments of the Campeche Bank, Mexico: diversity and bacterial community structure assessment. J Environ Managt. (2012) 95:S325–31. doi: 10.1016/j.jenvman.2011.06.046
66. Hu P, Dubinsky EA, Probst AJ, Wang J, Sieber CM, Tom LM, et al. Simulation of deepwater horizon oil plume reveals substrate specialization within a complex community of hydrocarbon degraders. Proc Natl Acad Sci USA. (2017) 114:7432–7. doi: 10.1073/pnas.1703424114
67. Colores GM, Macur RE, Ward DM, Inskeep WP. Molecular analysis of surfactant-driven microbial population shifts in hydrocarbon contaminated soil. Appl Environ Microbiol. (2000) 66:2959–64. doi: 10.1128/AEM.66.7.2959-2964.2000
68. Prince RC. The microbiology of marine oil spill bioremediation. In: Ollivier B, editor. Petroleum Microbiology. Toronto, ON: Wiley (2005). p. 317–35. doi: 10.1128/9781555817589.ch16
69. Das N, Chandran P. Microbial degradation of petroleum hydrocarbon contaminants: an overview. Biotechnol Res Int. (2011) 2011:941810. doi: 10.4061/2011/941810
70. Yuan J, Lai Q, Sun F, Zheng T, Shao Z. The diversity of PAH-degrading bacteria in a deep-sea water column above the southwest Indian ridge. Front Microbiol. (2015) 6:853. doi: 10.3389/fmicb.2015.00853
71. Deng MC, Li J, Liang FR, Yi M, Xu XM, Yuan JP, et al. Isolation and characterization of a novel hydrocarbon-degrading bacterium Achromobacter spp. HZ01 from the crude oil-contaminated seawater at the Daya Bay, southern China. Mar Pollut Bull. (2014) 83:79–86. doi: 10.1016/j.marpolbul.2014.04.018
72. Bacosa HP, Suto K, Inoue C. Bacterial community dynamics during the preferential degradation of aromatic hydrocarbons by a microbial consortium. Int Biodeter Biodegr. (2012) 74109–15. doi: 10.1016/j.ibiod.2012.04.022
73. Hamzah A, Phan CW, Abu Bakar NF, Wong KK. Biodegradation of crude oil by constructed bacterial consortia and the constituent single bacteria isolated from Malaysia. Bioremediat J. (2013) 17:1–10. doi: 10.1080/10889868.2012.731447
74. Chen Q, Li J, Liu M, Sun H, Bao M. Study on the biodegradation of crude oil by free and immobilized bacterial consortium in marine environment. PLoS ONE. (2017) 12:e0174445. doi: 10.1371/journal.pone.0174445
75. Czarny J, Staninska-Pieta J, Piotrowska-Cyplik A, Juzwa W, Wolniewicz A, Marecik R. Acinetobacter spp. as the key player in diesel oil degrading community exposed to PAHs and heavy metals. J Hazard Mater. (2020) 383:121168. doi: 10.1016/j.jhazmat.2019.121168
76. Zhou L, Li H, Zhang Y, Han S, Xu H. Sphingomonas from petroleum-contaminated soils in Shenfu, China and their PAHs degradation abilities. Braz J Microbiol. (2016) 47:271–8. doi: 10.1016/j.bjm.2016.01.001
77. Okoh A, Ajisebutu S, Babalola G, Trejo-Hernandez MR. Potential of burkholderia cepacia RQ1 in the biodegradation of heavy crude oil. Int Microbiol. (2001) 4:83–87. doi: 10.1007/s101230100018
78. Jiménez N, Viñas M, Guiu-Aragonés C, Bayona JM, Albaigés J, Solanas AM. Polyphasic approach for assessing changes in an autochthonous marine bacterial community in the presence of prestige fuel oil and its biodegradation potential. Appl Microbiol Biot. (2011) 91:823–34. doi: 10.1007/s00253-011-3321-4
79. Yakimov MM, Timmis KN, Golyshin PN. Obligate oil-degrading marine bacteria. Curr Opin Biotechnol. (2007) 18:257–66. doi: 10.1016/j.copbio.2007.04.006
80. Dubinsky EA, Conrad ME, Chakraborty R, Bill M, Borglin SE, Hollibaugh JT, et al. Succession of hydrocarbon-degrading bacteria in the aftermath of the deepwater horizon oil spill in the Gulf of Mexico. Environ Sci Technol. (2013) 47:10860–7. doi: 10.1021/es401676y
81. Barberán A, Bates ST, Casamayor EO, Fierer N. Using network analysis to explore co-occurrence patterns in soil microbial communities. ISME J. (2012) 6:343–51. doi: 10.1038/ismej.2011.119
82. Comte J, Lovejoy C, Crevecoeur S, Vincent WF. Co-occurrence patterns in aquatic bacterial communities across changing permafrost landscapes. Biogeosciences. (2016) 13:175–90. doi: 10.5194/bg-13-175-2016
83. Walter JM, Bagi A, Pampanin DM. Insights into the potential of the Atlantic cod gut microbiome as biomarker of oil contamination in the marine environment. Microorganisms. (2019) 7:209. doi: 10.3390/microorganisms7070209
84. QingFeng W, ShuYing Z, Li Z, ShuGuang X. Impact of anthracene addition on microbial community structure in soil microcosms from contaminated and uncontaminated sites. Biomed Environmen Sci. (2011) 24:543–9. doi: 10.3967/0895-3988.2011.05.014
85. Posman KM, DeRito CM, Madsen EL. Benzene degradation by a Variovorax species within a coal tar-contaminated groundwater microbial community. Appl Environ Microbiol. (2017) 83:e02658–16. doi: 10.1128/AEM.02658-16
Keywords: zebrafish, gut microbiota, crude oil, WAF, CEWAF, dysbiosis
Citation: González-Penagos CE, Zamora-Briseño JA, Cerqueda-García D, Améndola-Pimenta M, Pérez-Vega JA, Hernández-Nuñez E and Rodríguez-Canul R (2020) Alterations in the Gut Microbiota of Zebrafish (Danio rerio) in Response to Water-Soluble Crude Oil Components and Its Mixture With a Chemical Dispersant. Front. Public Health 8:584953. doi: 10.3389/fpubh.2020.584953
Received: 18 July 2020; Accepted: 06 October 2020;
Published: 26 October 2020.
Edited by:
Christopher J. Martyniuk, University of Florida, United StatesReviewed by:
Robert J. Griffitt, University of Southern Mississippi, United StatesJuan M. Gonzalez, Consejo Superior de Investigaciones Científicas (CSIC), Spain
Copyright © 2020 González-Penagos, Zamora-Briseño, Cerqueda-García, Améndola-Pimenta, Pérez-Vega, Hernández-Nuñez and Rodríguez-Canul. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Rossanna Rodríguez-Canul, cm9zc2FuYS5yb2RyaWd1ZXpAY2ludmVzdGF2Lm14