
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
BRIEF RESEARCH REPORT article
Front. Psychiatry , 03 April 2025
Sec. Psychopathology
Volume 16 - 2025 | https://doi.org/10.3389/fpsyt.2025.1421305
 Tolou Maslahati1*
Tolou Maslahati1* Franziska Glogau1Milagros Galve Gómez1Katharina Buchholz1Lisa Dormann1
Franziska Glogau1Milagros Galve Gómez1Katharina Buchholz1Lisa Dormann1 Katja Wingenfeld1
Katja Wingenfeld1 Christian Otte1Katharina Schultebraucks1,2,3†‡
Christian Otte1Katharina Schultebraucks1,2,3†‡ Stefan Roepke1†
Stefan Roepke1†Introduction: Traumatic experiences are thought to alter memory acquisition and consolidation. Cognitive models of PTSD suggest that voluntary and involuntary memories after trauma can be independently addressed through interventions. The administration of oxytocin before exposure to a trauma film led to more intrusive (involuntary) memories than placebo. The effect of oxytocin on voluntary memory of the traumatic film, however, remains unclear. The current study aimed to assess whether intranasal oxytocin administration facilitates forced-choice recognition memory after a trauma film paradigm.
Material & methods: We performed a pooled analysis of two randomized, double-blind, placebo-controlled studies (N = 437) to assess the impact of intranasal oxytocin administration on declarative memory. Participants received 24 I.U. of oxytocin, either 40 minutes before a trauma film paradigm or immediately afterward. We applied a forced-choice recognition task seven days after the trauma film paradigm. The task comprised pre-, peri, and post-trauma film scene details.
Results: The administration of oxytocin did not affect recognition performance for any film scene (F(2, 401) = .49, p = .61). Participants remembered significantly more peri-traumatic film details compared to pre- and post-trauma details (F(1.72, 802) = 103.38, p <.001).
Discussion: Although the exogenous oxytocin administration before a trauma film has been shown to influence the acquisition of intrusive memories, it does not seem to affect the recognition memory of trauma film details. That aligns with cognitive models of PTSD, suggesting that voluntary and involuntary memory after trauma can be independently addressed through experimental interventions.
During emotionally distressing experiences, hormonal and brain systems influence the acquisition and consolidation of newly acquired memories (1). After traumatic experiences, altered (involuntary and voluntary) memories of the trauma are at the core of Posttraumatic Stress Disorder (PTSD) (2), a mental disorder that has been conceptualized as a disorder of memory (3). Two of the most influential cognitive theories of PTSD, the cognitive model of PTSD (4) and the (revised) dual-representation theory of PTSD (5), suggest that trauma information is processed and stored in two distinct memory systems: involuntary re-experiencing of trauma experiences (intrusive memories) versus voluntary declarative trauma knowledge following elaboration and integration in context and autobiographical memory. Considering the challenges posed by aspects of involuntary memory, it is unsurprising that clinical interventions often focus on mitigating the frequency and intensity of involuntary memory symptoms (6–8). Ideally, interventions target the parts of the memory that trigger emotional and physiological distress but leave the declarative content intact (9). Examining the findings of experimental psychopathology reinforces the notion that interventions may indeed achieve the intended selective impact on both involuntary and voluntary facets of trauma memory. For example, investigations using a visuospatial task in trauma film paradigm studies led to decreased intrusive memories but did not affect recognition memory (10, 11). Protecting voluntary memory of the trauma is crucial for different reasons: voluntary memories are needed to prevent revictimization. Secondly, they are essential for legal documentation and testimony, such as in civil or social claims or criminal proceedings against perpetrators, frequently linked to trauma caused by human actions (10). It remains a crucial challenge to identify vulnerability factors of PTSD to develop preventative treatment for individuals exposed to trauma and further ensure that declarative memory stays unaffected.
Oxytocin, a neuropeptide involved in social cognition, stress regulation, and memory processes, has been discussed as a potential agent for trauma prevention (12). In this context, different mechanisms have been proposed through which oxytocin may influence trauma-related and emotional memory processing.
For one, oxytocin has been shown to interact with the hypothalamic-pituitary-adrenal (HPA) axis (13), which regulates cortisol release, a key stress hormone in memory processes (1). The proposed mechanism underlying this interaction is that oxytocin acts by inhibiting the transcription of the corticotropin releasing hormone and thereby inhibits the HPA axis cascade, leading to a reduced stress response (14). Results regarding the administration of oxytocin in human studies, however, are diverse. While there are confirmatory data showing diminishing effects on the HPA axis [e.g. (15, 16)], a meta-analysis found that oxytocin does not lower HPA axis reactivity per se (17). In their meta-analysis, Cardoso et al. (17) investigated the effect of oxytocin administration on HPA axis activity during laboratory stress tasks. They found that individuals with mental disorders showed a greater diminishing effect than healthy participants. This effect was even greater when the stress task caused a greater cortisol response. Consistent with these findings, we did not observe an effect of nasal oxytocin administration on cortisol release after the trauma, as previously reported (18, 19). Possibly, this absence of an effect could be attributed to the fact that our sample consisted of healthy participants and that, although the trauma film paradigm elicited a significant HPA axis response, it may not have been intense enough to reveal oxytocin-related modulation.
Another proposed mechanism by which oxytocin influences emotional memory processing is its role in neural circuits modulating limbic structures, particularly the amygdala and hippocampus, which are critical for emotional processing and memory consolidation (20). These brain regions, along with the prefrontal cortex, are rich in oxytocin receptors (21), suggesting a direct neuromodulatory role of oxytocin in emotional and memory-related processes. Moreover, oxytocin has been shown to enhance functional connectivity between the amygdala and the prefrontal cortex, a key pathway involved in emotion regulation and top-down control of fear responses (22). However, the effects of oxytocin on memory processing and trauma appear highly context-dependent and influenced by timing and individual differences.
This is reflected in mixed findings regarding its role in trauma-related memory formation: while oxytocin has been shown to prevent PTSD symptoms after traumatic experiences (23), on the one hand, it has also been found to facilitate the acquisition of intrusive memories after a trauma film paradigm (19), on the other. This apparent contradiction is not limited to PTSD-related symptoms but extends to oxytocin’s effects on memory more generally, as research has produced highly inconsistent findings. Specifically, its impact on declarative memory after trauma has yet to be assessed.
Early animal studies led to the assumption that oxytocin generally has amnestic properties on memory processes (24–26). This generalized concept was revised by first acknowledging the influences of dosage and timing on oxytocin effects and then observing memory-enhancing effects of oxytocin [e.g., (27)]. Accordingly, data showed that low doses of oxytocin delivered within one hour post-learning improved consolidation across various memory tasks in animal models, including episodic memory consolidation and retention (28, 29). It was further shown to increase recall of traumatic memory if administered immediately after trauma (foot-shock) (30).
In humans, oxytocin effects on memory processes are also diverse. During acquisition, oxytocin has been found to have memory enhancing as well as memory impairing effects. Accordingly, it improved recall of happy but not of neutral faces (31), it improved recognition of negative social stimuli (32) but also impaired recognition of words in general, irrespective of their valence (33). When administered during consolidation oxytocin improved recognition for negative (34, 35) and neutral but not for happy faces (34). During retrieval oxytocin administration only improved the recognition of social but not of neutral stimuli (36) and also improved the recognition of positive but not negative words (37). Furthermore, oxytocin impaired the ability to identify fearful faces in one study (37) and facilitated it in another (38).
The social salience hypothesis of oxytocin (39) helps to shed light on these contradictory findings. According to this framework, oxytocin does not uniformly enhance or impair memory but rather amplifies the salience of social-emotional stimuli, regardless of their valence (40, 41). As a result, its effects on declarative memory are not straightforward but depend on various factors, including the applied memory task (33, 42), context, and individual differences (43–46). This perspective suggests that oxytocin increases sensitivity to social cues in a way that is shaped by both interindividual factors and situational variables (12). Consequently, generalized statements about the direction of oxytocin’s effects on declarative memory cannot yet be made.
In previous analyses, we have shown that elevated oxytocin levels during the acquisition but not during consolidation increase the number of intrusive memories after exposure to a trauma film paradigm in healthy women (19, 47). To our knowledge, the effect of elevated oxytocin levels during encoding or consolidation of traumatic events on recognition memory has not been investigated yet. Given its role in stress regulation and memory processes, oxytocin’s effects may depend on whether it is administered before or after a traumatic experience. Before an emotional event, oxytocin is thought to enhance salience processing and amygdala reactivity, while post-event administration may facilitate memory consolidation and buffer stress responses. This distinction is particularly relevant, as encoding and consolidation represent two distinct neurocognitive processes that are thought to involve different brain mechanisms and timeframes (48).
The current study investigated the effect of exogenously increased oxytocin levels at the time of encoding (before the trauma film) or consolidation (after the trauma film) of an adverse event on recognition memory after one week in healthy women. To do so, we analyzed data of two experimental, randomized, double-blind, placebo-controlled studies (19, 47), each with a single dose of 24 I.U. nasal synthetic oxytocin (Syntocinon ®), using an established trauma film paradigm (49). Following the social salience hypothesis, we expected oxytocin administration before the trauma film to enhance the salience of trauma-related cues and, therefore, enhance memory recognition for peri-traumatic film details compared to placebo. As oxytocin has been shown to act anxiolytically when administered after traumatic events (23) and facilitate memory consolidation when administered after memory acquisition (29) or trauma (30), we also expected oxytocin administration after the trauma film to enhance memory recognition for peri-traumatic film-details.
The sample consisted of a pooled dataset of two separate studies with a similar design. The only difference between the two studies was the timing of oxytocin administration. Participants (n = 220) of the first study (19) received either 24 I.U. of nasal oxytocin or a placebo before the trauma film. Meanwhile, participants (n = 217) of the second study (35) received 24 I.U. or a placebo immediately after the trauma film. Both studies were completed at the Department of Psychiatry and Neurosciences, Campus Benjamin Franklin, Charité – Universitätsmedizin Berlin. They took place in the same room with an identical setup. Participants were limited to a single participation opportunity and were explicitly advised not to discuss the study design with other potential participants. Consequently, the probability of cohort effects and order effects occurring is minimal.
Due to the sexually dimorphic effect of oxytocin (50), only healthy women who reported female sex were included. In order to control the impact of the menstrual cycle on the endogenous oxytocin concentrations (51), subjects who did not take hormonal contraceptives were tested in the luteal phase only. The menstrual cycle phase of each participant was determined by utilizing self-reported information regarding the start date of their most recent menstruation and the duration of their cycle.
Participants were recruited via university email lists and public postings and received an expense allowance for their participation. Exclusion criteria have been published previously (19, 47). To ensure complete recovery from the trauma film, participants were contacted by phone four weeks after participation and offered psychological care in case of ongoing distress. In the second study (47),, one participant reported persistent intrusive symptoms and subsequently had six counseling sessions with a licensed psychologist. The participant’s distress and intrusive experiences resolved during the aftercare period.
Two randomized, double-blind, placebo-controlled studies with each a single dose of 24 I.U. nasal synthetic oxytocin (Syntocinon ®) were applied. The oxytocin and placebo sprays were administered by the experimenters to ensure standardized application. Syntocinon® is a commercially available and standardized nasal spray. The placebo spray contained sodium chloride (0.9%), aqua conservans, chlorobutanol, anhydrous citric acid, and glycerin, but without the active ingredient. To maintain blinding, both sprays were prepared in identical bottles with no distinguishable differences in appearance, smell, or taste. The total volume administered was 0.6 ml. The local ethics committee of Charité – Universitätsmedizin Berlin approved the study protocols of both studies (EA4/162/18; EA4/144/16). Study information was sent to participants at least 24 hours before participation, and informed consent was given upon arrival in the laboratory. Participants were asked not to smoke, do physical exercise, and not to eat or consume caffeine, alcohol, or any beverage other than water one hour before the assessment. Considering cortisol fluctuations (52), the start of every testing was set for 2 p.m.
The participants received either 24 I.U. Oxytocin (nasal spray, Syntocinon ®) or a placebo preparation (sodium chloride nasal spray) before or immediately after the trauma film. Group allocation occurred randomly in each study. The oxytocin nasal spray and the placebo preparation looked identical to ensure the double-blind design. Peak levels of oxytocin occur within 39 – 51 minutes (53); therefore, the nasal spray was given 40 minutes before the trauma film in the first study. The psychometric assessment, among other things, comprised the Beck Depression Inventory-Revised (BDI-II) (54) and the Childhood Trauma Questionnaire (CTQ) (55). We utilized the BDI-II to identify possible indicators of depressive symptoms. A BDI-II score > 13 was considered clinically relevant and led to exclusion. The CTQ was used to detect any childhood trauma that can increase the risk of PTSD after a secondary trauma in adulthood (56).
After the session, participants filled in a diary to record intrusive memories using the methodology outlined by Holmes and colleagues (57). Detailed results regarding these intrusive memories have been reported elsewhere (19, 47).
The applied trauma film is a well-established paradigm (57–59) and was presented in a dark room on a 2 x 2.5m screen. The 14-minute and 40-second clip was extracted from Gaspar Noë’s commercial film “Irreversible” (45). It portrays a distressing sequence in which a woman is subjected to a violent assault, including a brutal rape and physical beating, perpetrated by an unknown assailant in a pedestrian underpass. A trained female investigator remained in the room to monitor adherence to the instructions to ensure that the participants watched the entire scene without any visual (e.g., closing their eyes) or auditory avoidance (e.g., removing their headphones).
We adopted a forced-choice recognition task tailored to the content of the film to measure declarative memory for the trauma film. The film scenes were categorized into three phases: pre-, peri-, and post-trauma. The pre-trauma scene shows how the woman leaves a party, the peri-trauma scene shows how the woman is raped and beaten by a man, and the post-trauma scene shows how the woman’s friend sees her being transported into the ambulance. The initial test construction process is described by Rombold-Bruehl et al. (60), who at first constructed a 24-item version of the recognition test. Four correct and four incorrect statements were formulated for each of the three film sequences. Due to problems with item severity, 11 items were added to the set, resulting in a total of 35 items that participants rated in a dichotomic way. For each statement, one had to choose if the statement was correct (“correct” vs. “false”) and how certain one would be about this rating (“certain” vs. “uncertain”). Corresponding to the three film sequences, the recognition test is divided into three item groups, referring to the pre-, peri-, and post-trauma sequences. Twelve items represented the pre-trauma sequence, 12 items described the peri-trauma sequence, and 11 items the post-trauma sequence. For statistical analyses, only 17 out of the 35 items (seven pre-trauma items, four peri-trauma items, six post-trauma items) were used due to problems regarding item severity and item discrimination in our sample, which will be described in more detail in the results section of this paper.
Statistical analyses were conducted using SPSS Version 23.0. Statistical significance refers to a p-value < 0.05. Sample characteristics between intervention groups (oxytocin before the trauma film, oxytocin after the trauma film, pooled placebo group) were conducted using Chi-square tests for categorical data and univariate analysis of variance (ANOVA) for continuous data.
We employed a repeated measure mixed design ANOVA to investigate the impact of the intervention (between-subjects factor = oxytocin before the trauma film; oxytocin after the trauma film; placebo) and the film sequence (within-subjects variable with three levels = pre-, peri-, post-trauma scene) on the sensitivity score d´ (d - prime) for each film scene, which served as the dependent variable. According to the signal detection theory of recognition memory (61), the sensitivity score d’ determines a participant’s recognition memory performance, corrected for guessing. Hence, it distinguishes between sensitivity (the ability to answer an item correctly) and response bias (62). In the current study, the participants rated the confidence in their answers as either “certain” or “uncertain”. These ratings were used to calculate hits and false alarms. According to Maniscalco and Lau (62), a hit describes a correct answer with a confidence rating as “certain”, whereas a false alarm is an incorrect answer with “certain” confidence. To calculate d’, the false alarm rate must be subtracted from the hit rate. D’ can range from - 1 to + 1. Higher positive d’ values imply a better recognition performance.
We incorporated n = 403 out of the total n = 437 enrolled participants in the final analysis. Factors like prolonged interruptions in the experiment resulting from technical issues led to the exclusion of participants. A comprehensive report detailing the inclusion and exclusion of participants in both studies has been previously published (18, 19).
The three groups (oxytocin before the trauma film, oxytocin after the trauma film, and combined placebo group) did not differ significantly in any demographic variable or possible confounding variable presented in Table 1.
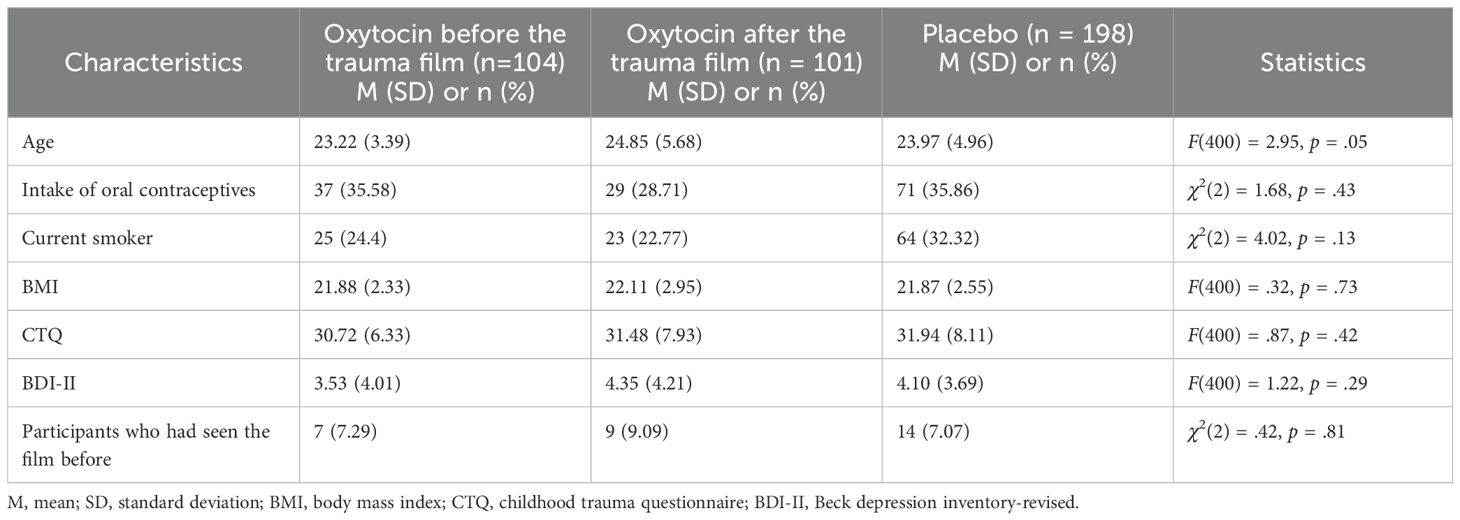
Table 1. Sample characteristics.
An analysis of the memory measure revealed that some items were insensitive, meaning they were either too easy (more than 80% correct) or too complex (less than 20% correct) to answer. Following the methods by Holmes et al. (57), we only included items correctly answered by > 20% or < 80% of the participants. Table 2 overviews the difficulty and discrimination of all 35 items. The analyses reported refer to the measure after the invariant items were removed.
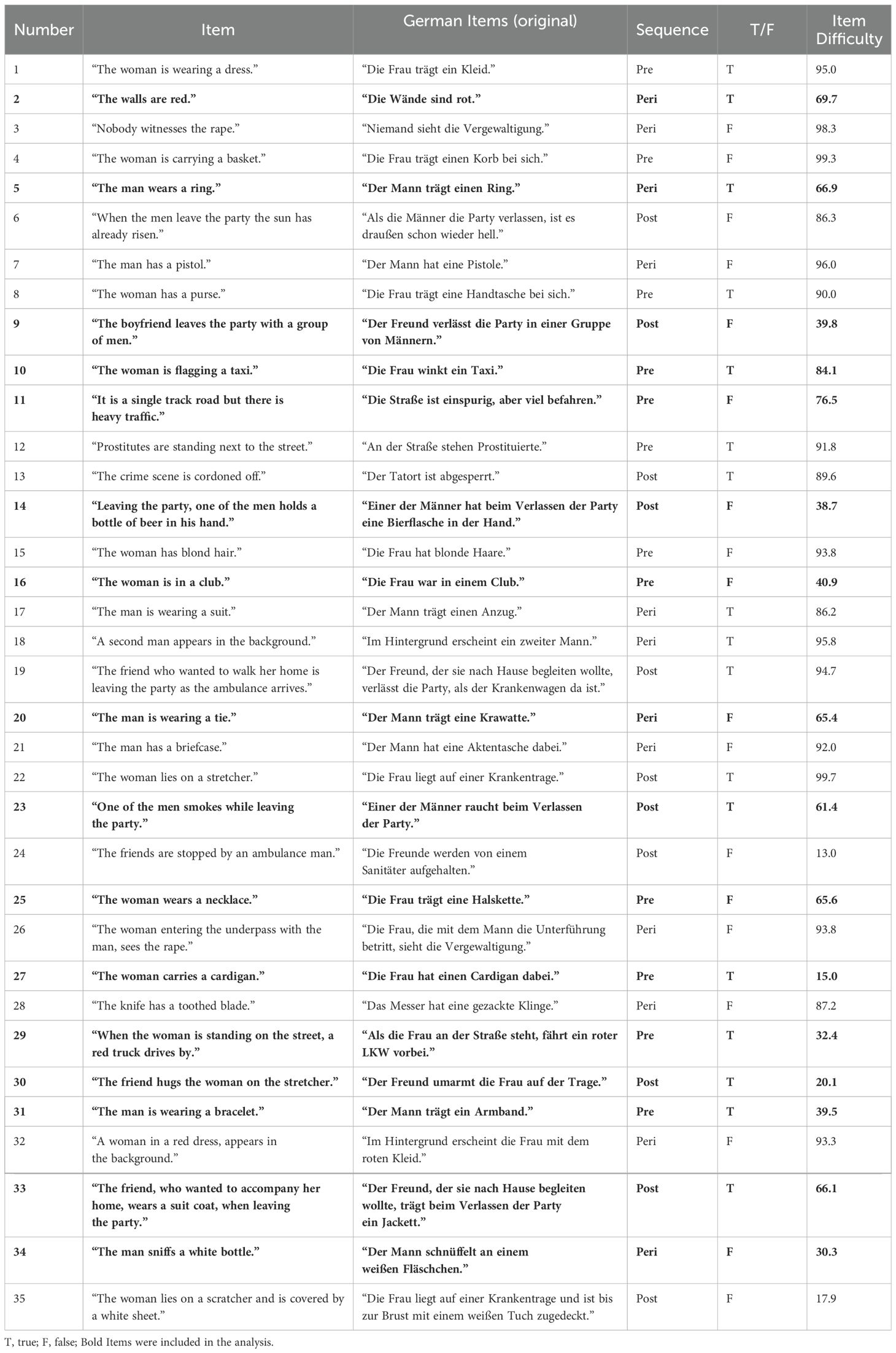
Table 2. Items of the forced-choice recognition task.
The mean hit rates and false alarm rates for the three groups were as follows: For the oxytocin group before the trauma film, the mean hit rate was 2.92 (SD = 1.74) and the mean false alarm rate was 2.56 (SD = 1.28). For the oxytocin group after the trauma film, the mean hit rate was 3.33 (SD = 1.91) and the mean false alarm rate was 2.73 (SD = 1.61). For the placebo group, the mean hit rate was 2.89 (SD = 1.75) and the mean false alarm rate was 2.67 (SD = 1.49). The mean d´scores of the three groups (oxytocin before the trauma film, oxytocin after the trauma film, and combined placebo group) for each film scene (per-, peri-, post-trauma) are shown in Figure 1. While there was a significant within-subject factor (film-scene) effect (F(1.72, 802) = 103.38, p <.001, ηp2 = 0.20), there was no significant main effect of oxytocin F(2, 401) = .49, p = .61, ηp2 = 0.01). There was also no significant interaction effect between the treatment and film scene (F(3.45,802) = .33, p = .83, ηp2 = 0.00). Post-hoc tests revealed that peri-traumatic details were remembered significantly more than pre-trauma details (p <.001) and post-trauma details (p <.001).
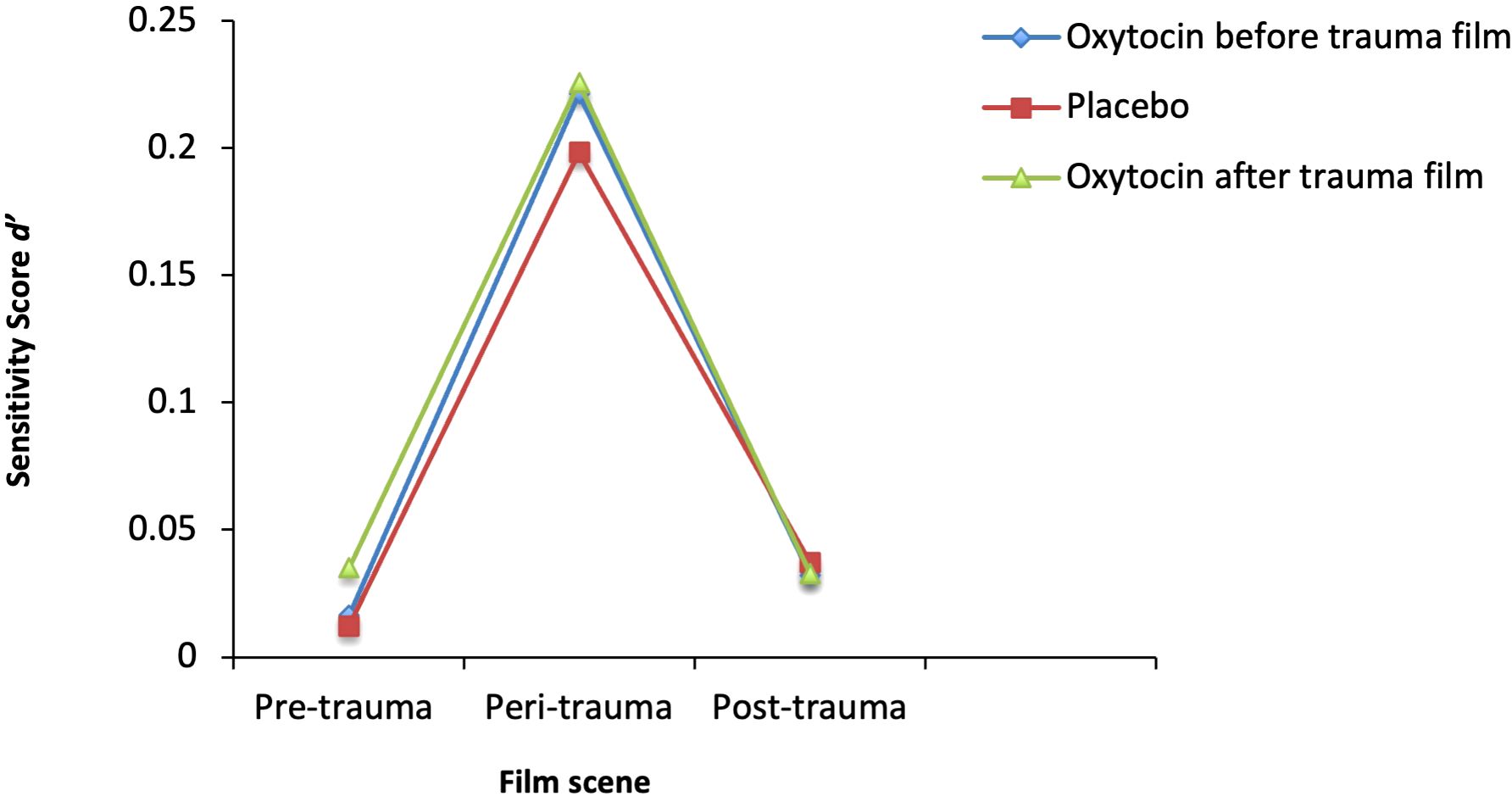
Figure 1. Recognition Performance across the Film Scenes. Mean d´-Scores in the three groups (Oxytocin before the trauma film, Oxytocin after the trauma film, Placebo) for pre-, peri-, and post-trauma film scene memory were assessed after seven days.
In this study, we examined the impact of exogenous oxytocin administration before and after a distressing film on forced-choice recognition after one week in healthy women. According to the social salience hypothesis (39), we expected oxytocin administration before the trauma film to enhance trauma-related cues and, therefore, improve memory recognition for peri-traumatic film details. As previous studies about oxytocin have shown anxiolytic effects after traumatic events (23) and enhancing effects on memory consolidation when administered after trauma (30), we also expected oxytocin administration after the trauma film to enhance recognition memory for peri-traumatic film details. Contrary to our hypotheses, we found no effect of either oxytocin before or after the trauma film on recognition memory. We found significant differences in recognition performance regarding trauma film scenes. Participants in all treatment conditions showed a significantly better recognition performance for peri-traumatic film details compared to pre- or post-trauma film details. Meaning, more peri-traumatic details were answered correctly and with more certainty. While pre- and post-trauma film scenes showed mainly neutral content, the peri-trauma film scene consisted of emotional and distressing material showing rape and physical abuse of a woman. That result would align with research on the processing of emotional content and the role of the salience network (63), implying that individuals are more likely to remember information that is emotionally relevant to them (64). Research even suggests that neutral information can be particularly difficult to remember when they appear close to an emotional stimulus (65, 66). However, the result of the current study could also be explained by slightly different difficulty levels of the questions in the applied forced-choice recognition task.
In a previous study, we found that the administration of oxytocin before the trauma film led to more intrusive memories over the following four days (19). Contradictory to our hypothesis, oxytocin administration before the trauma film did not affect recognition memory. An exogenous elevation of oxytocin levels, appears to influence involuntary intrusive memories after a trauma film, while voluntary recognition memory remains unaffected. Correspondingly, a former study with a similar design by our research lab found voluntary and involuntary memory to be independently affected by the manipulation of the noradrenergic system (60, 67). While the activation of the noradrenergic system impacted involuntary memory (intrusions), it did not have an impact on voluntary (recognition) memory. The inhibition of the noradrenergic system on the other hand did only impact voluntary memory and had no effect on involuntary memory (60, 67). These results align with cognitive models of PTSD (4, 68) and research (49, 69), suggesting that voluntary and involuntary memory after trauma can be independently addressed through experimental interventions.
Also, contrary to our hypothesis, oxytocin administration after exposure to the trauma film did not affect recognition memory. That is contradictory to findings showing anxiolytic effects and decreased PTSD symptoms in patients with PTSD (23, 70). Further on, oxytocin has been shown to facilitate memory consolidation when administered after trauma (foot shock) in a rodent study (30). One hypothesis for the missing effect of oxytocin on the consolidation of memories is the administration of only a single dose of nasal oxytocin in our study. A rodent study showed that a single administration of oxytocin directly after a stressor increased contextual fear memory, but a repeated oxytocin administration post-stressor decreased fear generalization (30). Similarly to that study, the aforementioned contradicting seeming study (23) administered oxytocin repeatedly. That indicates that the effects of a single administration may differ from the effects of repeated administration. Administering oxytocin repeatedly might have influenced the outcome of the current study. Another possible hypothesis for the lack of effect in our study may be the degree of distress caused by the trauma film paradigm. The findings from the study conducted by van Zuiden and colleagues (23) suggest that oxytocin does not affect PTSD symptoms per se. Instead, it appears to be only effective in individuals who reported high levels of distress following the trauma. It seems reasonable to consider that the trauma film paradigm administered to healthy individuals may not evoke the same level of distress as actual traumatic events. Consequently, the application of oxytocin in trauma analog studies might not produce a substantial effect on memory consolidation.
The high internal validity of this study due to high experimental control, strict inclusion criteria for the participants, and the inclusion of women only can be highlighted as the study’s main strength. Further strengths of the study are assessing the oxytocin effect before and after the trauma film, examining pre-, peri-, and post-trauma film memory recognition separately, and using a placebo-controlled, double-blind design. As both the menstrual cycle and the intake of hormonal contraceptives have been shown to have an impact on endogenous oxytocin concentrations (51, 71), we considered the intake of hormonal contraceptives and made an effort to control for menstrual cycle by testing naturally cycling women in their luteal phase only. Nevertheless, the luteal phase also shows strong fluctuations in hormonal levels. Future studies may want to choose a narrower subphase of the menstrual cycle. While the homogeneity of our sample is considered a strength in terms of internal validity, it hampers generalization to more vulnerable populations, such as individuals with psychiatric disorders or with previous traumatic experiences. Results can also not be transferred to men, as oxytocin has sexually dimorphic effects potentially due to differences in oxytocin plasma concentrations (40) or different oxytocin expressions in different brain regions of women and men (72). Generalizability may further be restricted as trauma film paradigms are relatively mild stressors compared to actual traumatic events causing PTSD. It remains unclear if conclusions about memory recognition after real trauma can be drawn from the presented data. However, trauma film paradigms offer a valid examination of PTSD risk factors while being ethically justifiable (49).
Next, it is possible that keeping an intrusion diary interfered with memory processing and biased the forced choice recognition task results. Frequent diary entries can enhance memory recognition by reinforcing memory traces through repeated exposure to intrusive memories, which may lead to improved recall over time. One study, for example, showed that individuals who document intrusions experience a heightened sense of immediacy, which can enhance recognition memory performance in tasks requiring quick retrieval (73). At the same time, intrusion diaries may also introduce challenges, such as increased intrusion errors, which may lead to interference, where overlapping memories can confuse recall, potentially complicating the retrieval process and affecting overall memory accuracy (74). Therefore, the potential influence of intrusion diaries on recognition memory should be considered a limitation of the present study, as both memory enhancement and interference effects could have impacted the results. Future research is needed to investigate how intrusion diaries affect recognition memory systematically.
Finally, the applied forced-choice recognition task must be listed as a limitation. We had to exclude 18 out of 35 items to improve item discrimination. While there are valid instruments to examine intrusive memories after trauma film paradigms, there is no standardized and valid measure to examine recognition memory of film details, which has repeatedly been pointed out by different research groups (57, 60). It is, therefore, eminently necessary to construct and validate a standardized memory task for voluntary trauma memory of the applied trauma film paradigm. Different research groups could then use such a task consistently, allowing comparable results across studies. In this regard, it is essential to take various facets of recognition memory into account, given that participants’ accurate responses may be based on recollection, which entails remembering specific details, as well as familiarity, which relies only on the knowledge that an item was present without contextual information (61). It is further essential to pay attention to the type of items included in that kind of questionnaire since the use of suggestive and non-suggestive questions leads to different and sometimes opposing results (75) and to include enough items per category to be able to distinguish between correct recognition and correct rejections (62).
In summary, exogenous administration of oxytocin either during encoding or during consolidation did not affect the recognition memory after a trauma film paradigm in healthy women. Integrating the findings of previously conducted studies (19, 47), it appears that oxytocin influences the acquisition of intrusive memories after exposure to a trauma film in healthy young women but does not affect the recognition memory of trauma film details. The observation that involuntary and voluntary memory may be influenced separately holds clinical significance. It is important and, at the same time, challenging to find interventions after traumatic experiences that diminish involuntary memories while preserving voluntary memories of the trauma. The latter is crucial, especially in contexts such as legal documentation, testimony, civil or social claims, and criminal proceedings against perpetrators, often associated with trauma resulting from human actions.
The datasets presented in this article are not readily available because the participants did not agree for the data to be made available publicly. Requests to access the datasets should be directed to TM, dG9sb3UubWFzbGFoYXRpQGNoYXJpdGUuZGU=.
The studies involving humans were approved by Ethics Committee of Charité - Universitätsmedizin Berlin. The studies were conducted in accordance with the local legislation and institutional requirements. The participants provided their written informed consent to participate in this study.
TM: Conceptualization, Data curation, Formal Analysis, Investigation, Methodology, Project administration, Software, Writing – original draft, Writing – review & editing. FG: Investigation, Writing – review & editing. MG: Investigation, Writing – review & editing. KB: Investigation, Writing – review & editing. LD: Investigation, Writing – review & editing. KW: Supervision, Writing – review & editing. CO: Supervision, Writing – review & editing. KS: Conceptualization, Methodology, Supervision, Writing – review & editing. SR: Conceptualization, Funding acquisition, Methodology, Resources, Supervision, Writing – review & editing, Project administration.
The author(s) declare that financial support was received for the research and/or publication of this article. Authors acknowledge financial support from the Open Access Publication Fund of Charité – Universi- tätsmedizin Berlin.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Roozendaal B, McGaugh JL. Memory modulation. Behav Neurosci. (2011) 125:797–824. doi: 10.1037/a0026187
2. Association AP. Diagnostic and statistical manual of mental disorders (DSM-5®). Arlington, VA: American Psychiatric Pub (2013).
3. Visser RM, Lau-Zhu A, Henson RN, Holmes EA. Multiple memory systems, multiple time points: how science can inform treatment to control the expression of unwanted emotional memories. Philos Trans R Soc B: Biol Sci. (2018) 373:20170209. doi: 10.1098/rstb.2017.0209
4. Ehlers A, Clark DM. A cognitive model of posttraumatic stress disorder. Behav Res Ther. (2000) 38:319–45. doi: 10.1016/S0005-7967(99)00123-0
5. Brewin CR, Gregory JD, Lipton M, Burgess N. Intrusive images in psychological disorders: characteristics, neural mechanisms, and treatment implications. Psychol Rev. (2010) 117:210–32. doi: 10.1037/a0018113
6. Kanstrup M, Singh L, Göransson KE, Widoff J, Taylor RS, Gamble B, et al. Reducing intrusive memories after trauma via a brief cognitive task intervention in the hospital emergency department: an exploratory pilot randomised controlled trial. Trans Psychiatry. (2021) 11:30. doi: 10.1038/s41398-020-01124-6
7. Pollmann Y, Clancy KJ, Devignes Q, Ren B, Kaufman ML, Rosso IM. Specific symptom change associated with ecological momentary assessments of intrusive trauma memories. NPP—Digital Psychiatry Neurosci. (2024) 2:1–8. doi: 10.1038/s44277-024-00019-4
8. Horsch A, Vial Y, Favrod C, Harari MM, Blackwell SE, Watson P, et al. Reducing intrusive traumatic memories after emergency caesarean section: A proof-of-principle randomized controlled study. Behav Res Ther. (2017) 94:36–47. doi: 10.1016/j.brat.2017.03.018
9. Phelps EA, Hofmann SG. Memory editing from science fiction to clinical practice. Nature. (2019) 572:43–50. doi: 10.1038/s41586-019-1433-7
10. Lau-Zhu A, Henson RN, Holmes EA. Intrusive memories and voluntary memory of a trauma film: Differential effects of a cognitive interference task after encoding. J Exp Psychol: Gen. (2019) 148:2154. doi: 10.1037/xge0000598
11. Lau-Zhu A, Henson RN, Holmes EA. Selectively interfering with intrusive but not voluntary memories of a trauma film: accounting for the role of associative memory. Clin Psychol Sci. (2021) 9:1128–43. doi: 10.1177/2167702621998315
12. Olff M, Frijling JL, Kubzansky LD, Bradley B, Ellenbogen MA, Cardoso C, et al. The role of oxytocin in social bonding, stress regulation and mental health: an update on the moderating effects of context and interindividual differences. Psychoneuroendocrinology. (2013) 38:1883–94. doi: 10.1016/j.psyneuen.2013.06.019
13. Brown CA, Cardoso C, Ellenbogen MA. A meta-analytic review of the correlation between peripheral oxytocin and cortisol concentrations. Front Neuroendocrinol. (2016) 43:19–27. doi: 10.1016/j.yfrne.2016.11.001
14. Dief AE, Sivukhina EV, Jirikowski GF. Oxytocin and stress response. Open J Endocr Metab Dis. (2018) 8:93–104. doi: 10.4236/ojemd.2018.83010
15. Ditzen B, Schaer M, Gabriel B, Bodenmann G, Ehlert U, Heinrichs M. Intranasal oxytocin increases positive communication and reduces cortisol levels during couple conflict. Biol Psychiatry. (2009) 65:728–31. doi: 10.1016/j.biopsych.2008.10.011
16. Linnen A-M, Ellenbogen MA, Cardoso C, Joober R. Intranasal oxytocin and salivary cortisol concentrations during social rejection in university students. Stress. (2012) 15:393–402. doi: 10.3109/10253890.2011.631154
17. Cardoso C, Kingdon D, Ellenbogen MA. A meta-analytic review of the impact of intranasal oxytocin administration on cortisol concentrations during laboratory tasks: moderation by method and mental health. Psychoneuroendocrinology. (2014) 49:161–70. doi: 10.1016/j.psyneuen.2014.07.014
18. Maslahati T, Wingenfeld K, Hellmann-Regen J, Kraft J, Lyu J, Keinert M, et al. Oxytocin vs. placebo effects on intrusive memory consolidation using a trauma film paradigm: a randomized, controlled experimental study in healthy women. Transl Psychiatry. (2022) 13(1):42. doi: 10.21203/rs.3.rs-2166294/v1
19. Schultebraucks K, Maslahati T, Wingenfeld K, Hellmann-Regen J, Kraft J, Kownatzki M, et al. Intranasal oxytocin administration impacts the acquisition and consolidation of trauma-associated memories: a double-blind randomized placebo-controlled experimental study in healthy women. Neuropsychopharmacology. (2021) 47(5):1046–54. doi: 10.1038/s41398-023-02339-z
20. Walia V, Wal P, Mishra S, Agrawal A, Kosey S, Patil AD. Potential role of oxytocin in the regulation of memories and treatment of memory disorders. Peptides. (2024) 177:171222. doi: 10.1016/j.peptides.2024.171222
21. Jurek B, Neumann ID. The oxytocin receptor: from intracellular signaling to behavior. Physiol Rev. (2018) 98:1805–908. doi: 10.1152/physrev.00031.2017
22. Scheele D, Lieberz J, Goertzen-Patin A, Engels C, Schneider L, Stoffel-Wagner B, et al. Trauma disclosure moderates the effects of oxytocin on intrusions and neural responses to fear. Psychother Psychosom. (2019) 88:61–3. doi: 10.1159/000496056
23. Van Zuiden M, Frijling JL, Nawijn L, Koch SB, Goslings JC, Luitse JS, et al. Intranasal oxytocin to prevent posttraumatic stress disorder symptoms: a randomized controlled trial in emergency department patients. Biol Psychiatry. (2017) 81:1030–40. doi: 10.1016/j.biopsych.2016.11.012
24. Kovács GL, Bohus B, Versteeg DH, De Kloet ER, De Wied D. Effect of oxytocin and vasopressin on memory consolidation: sites of action and catecholaminergic correlates after local microinjection into limbic-midbrain structures. Brain Res. (1979) 175:303–14. doi: 10.1016/0006-8993(79)91009-6
25. Boccia M, Kopf S, Baratti CM. Effects of a single administration of oxytocin or vasopressin and their interactions with two selective receptor antagonists on memory storage in mice. Neurobiol Learn Memory. (1998) 69:136–46. doi: 10.1006/nlme.1997.3817
26. De Oliveira LF, Camboim C, Diehl F, Consiglio AR, Quillfeldt JA. Glucocorticoid-mediated effects of systemic oxytocin upon memory retrieval. Neurobiol Learn Memory. (2007) 87:67–71. doi: 10.1016/j.nlm.2006.05.006
27. Lukas M, Neumann ID. Nasal application of neuropeptide S reduces anxiety and prolongs memory in rats: social versus non-social effects. Neuropharmacology. (2012) 62:398–405. doi: 10.1016/j.neuropharm.2011.08.016
28. Chini B, Leonzino M, Braida D, Sala M. Learning about oxytocin: pharmacologic and behavioral issues. Biol Psychiatry. (2014) 76:360–6. doi: 10.1016/j.biopsych.2013.08.029
29. de Wied D, Diamant M, Fodor M. Central nervous system effects of the neurohypophyseal hormones and related peptides. Front Neuroendocrinol. (1993) 14:251–302. doi: 10.1006/frne.1993.1009
30. Janezic EM, Uppalapati S, Nagl S, Contreras M, French ED, Fellous J-M. Beneficial effects of chronic oxytocin administration and social co-housing in a rodent model of post-traumatic stress disorder. Behav Pharmacol. (2016) 27:704–17. doi: 10.1097/FBP.0000000000000270
31. Guastella AJ, Mitchell PB, Mathews F. Oxytocin enhances the encoding of positive social memories in humans. Biol Psychiatry. (2008) 64:256–8. doi: 10.1016/j.biopsych.2008.02.008
32. Weigand A, Feeser M, Gärtner M, Brandt E, Fan Y, Fuge P, et al. Effects of intranasal oxytocin prior to encoding and retrieval on recognition memory. Psychopharmacology. (2013) 227:321–9. doi: 10.1007/s00213-012-2962-z
33. Heinrichs M, Meinlschmidt G, Wippich W, Ehlert U, Hellhammer DH. Selective amnesic effects of oxytocin on human memory. Physiol Behav. (2004) 83:31–8. doi: 10.1016/S0031-9384(04)00346-4
34. Savaskan E, Ehrhardt R, Schulz A, Walter M, Schächinger H. Post-learning intranasal oxytocin modulates human memory for facial identity. Psychoneuroendocrinology. (2008) 33:368–74. doi: 10.1016/j.psyneuen.2007.12.004
35. Striepens N, Scheele D, Kendrick KM, Becker B, Schäfer L, Schwalba K, et al. Oxytocin facilitates protective responses to aversive social stimuli in males. Proc Natl Acad Sci. (2012) 109:18144–9. doi: 10.1073/pnas.1208852109
36. Rimmele U, Hediger K, Heinrichs M, Klaver P. Oxytocin makes a face in memory familiar. J Neurosci. (2009) 29:38–42. doi: 10.1523/JNEUROSCI.4260-08.2009
37. Di Simplicio M, Massey-Chase R, Cowen P, Harmer C. Oxytocin enhances processing of positive versus negative emotional information in healthy male volunteers. J Psychopharmacol. (2009) 23:241–8. doi: 10.1177/0269881108095705
38. Fischer-Shofty M, Shamay-Tsoory SG, Harari H, Levkovitz Y. The effect of intranasal administration of oxytocin on fear recognition. Neuropsychologia. (2010) 48:179–84. doi: 10.1016/j.neuropsychologia.2009.09.003
39. Shamay-Tsoory SG, Abu-Akel A. The social salience hypothesis of oxytocin. Biol Psychiatry. (2016) 79:194–202. doi: 10.1016/j.biopsych.2015.07.020
40. Macdonald K, Feifel D. Helping oxytocin deliver: considerations in the development of oxytocin-based therapeutics for brain disorders. Front Neurosci. (2013) 7:35. doi: 10.3389/fnins.2013.00035
41. Maroun M, Wagner S. Oxytocin and memory of emotional stimuli: some dance to remember, some dance to forget. Biol Psychiatry. (2016) 79:203–12. doi: 10.1016/j.biopsych.2015.07.016
42. Engelmann M, Wotjak CT, Neumann I, Ludwig M, Landgraf R. Behavioral consequences of intracerebral vasopressin and oxytocin: focus on learning and memory. Neurosci Biobehav Rev. (1996) 20:341–58. doi: 10.1016/0149-7634(95)00059-3
43. Bartz J, Simeon D, Hamilton H, Kim S, Crystal S, Braun A, et al. Oxytocin can hinder trust and cooperation in borderline personality disorder. Soc Cogn Affect Neurosci. (2011) 6:556–63. doi: 10.1093/scan/nsq085
44. Pierrehumbert B, Torrisi R, Laufer D, Halfon O, Ansermet F, Popovic MB. Oxytocin response to an experimental psychosocial challenge in adults exposed to traumatic experiences during childhood or adolescence. Neuroscience. (2010) 166:168–77. doi: 10.1016/j.neuroscience.2009.12.016
45. Hurlemann R, Scheele D. Dissecting the role of oxytocin in the formation and loss of social relationships. Biol Psychiatry. (2016) 79:185–93. doi: 10.1016/j.biopsych.2015.05.013
46. Smearman EL, Almli LM, Conneely KN, Brody GH, Sales JM, Bradley B, et al. Oxytocin receptor genetic and epigenetic variations: association with child abuse and adult psychiatric symptoms. Child Dev. (2016) 87:122–34. doi: 10.1111/cdev.2016.87.issue-1
47. Maslahati T, Wingenfeld K, Hellmann-Regen J, Kraft J, Lyu J, Keinert M, et al. Oxytocin vs. placebo effects on intrusive memory consolidation using a trauma film paradigm: a randomized, controlled experimental study in healthy women. Trans Psychiatry. (2023) 13:42. doi: 10.1038/s41398-023-02339-z
48. Crowley R, Bendor D, Javadi A-H. A review of neurobiological factors underlying the selective enhancement of memory at encoding, consolidation, and retrieval. Prog Neurobiol. (2019) 179:101615. doi: 10.1016/j.pneurobio.2019.04.004
49. James EL, Lau-Zhu A, Clark IA, Visser RM, Hagenaars MA, Holmes EA. The trauma film paradigm as an experimental psychopathology model of psychological trauma: intrusive memories and beyond. Clin Psychol Rev. (2016) 47:106–42. doi: 10.1016/j.cpr.2016.04.010
50. Ditzen B, Nater UM, Schaer M, La Marca R, Bodenmann G, Ehlert U, et al. Sex-specific effects of intranasal oxytocin on autonomic nervous system and emotional responses to couple conflict. Soc Cognit Affect Neurosci. (2013) 8:897–902. doi: 10.1093/scan/nss083
51. Engel S, Klusmann H, Ditzen B, Knaevelsrud C, Schumacher S. Menstrual cycle-related fluctuations in oxytocin concentrations: A systematic review and meta-analysis. Front Neuroendocrinol. (2019) 52:144–55. doi: 10.1016/j.yfrne.2018.11.002
52. Kirschbaum C, Hellhammer DH. Salivary cortisol in psychobiological research: an overview. Neuropsychobiology. (1989) 22:150–69. doi: 10.1159/000118611
53. Paloyelis Y, Doyle OM, Zelaya FO, Maltezos S, Williams SC, Fotopoulou A, et al. A spatiotemporal profile of in vivo cerebral blood flow changes following intranasal oxytocin in humans. Biol Psychiatry. (2016) 79:693–705. doi: 10.1016/j.biopsych.2014.10.005
54. Beck AT, Steer RA, Brown GK. Manual for the beck depression inventory-II Vol. 1. . San Antonio, TX: Psychological Corporation (1996). p. 82.
55. Wingenfeld K, Spitzer C, Mensebach C, Grabe HJ, Hill A, Gast U, et al. The German version of the Childhood Trauma Questionnaire (CTQ): preliminary psychometric properties. Psychother Psychosom Medizinische Psychol. (2010) 60:442–50. doi: 10.1055/s-0030-1247564
56. Breslau N, Chilcoat HD, Kessler RC, Davis GC. Previous exposure to trauma and PTSD effects of subsequent trauma: results from the Detroit Area Survey of Trauma. Am J Psychiatry. (1999) 156:902–7. doi: 10.1176/ajp.156.6.902
57. Holmes EA, Brewin CR, Hennessy RG. Trauma films, information processing, and intrusive memory development. J Exp Psychol: Gen. (2004) 133:3. doi: 10.1037/0096-3445.133.1.3
58. Holmes EA, Bourne C. Inducing and modulating intrusive emotional memories: A review of the trauma film paradigm. Acta Psychol. (2008) 127:553–66. doi: 10.1016/j.actpsy.2007.11.002
59. Weidmann A, Conradi A, Gröger K, Fehm L, Fydrich T. Using stressful films to analyze risk factors for PTSD in analogue experimental studies–which film works best? Anxiety Stress Coping. (2009) 22:549–69. doi: 10.1080/10615800802541986
60. Rombold-Bruehl F, Otte C, Renneberg B, Hellmann-Regen J, Bruch L, Wingenfeld K, et al. Impact of stress response systems on forced choice recognition in an experimental trauma film paradigm. Neurobiol Learn Memory. (2018) 156:45–52. doi: 10.1016/j.nlm.2018.10.010
61. Wixted JT. Dual-process theory and signal-detection theory of recognition memory. psychol Rev. (2007) 114:152. doi: 10.1037/0033-295X.114.1.152
62. Maniscalco B, Lau H. Signal detection theory analysis of type 1 and type 2 data: meta-d′, response-specific meta-d′, and the unequal variance SDT model. In: The cognitive neuroscience of metacognition. Heidelberg, Germany: Springer (2014). p. 25–66.
64. Holland AC, Kensinger EA. Emotion and autobiographical memory. Phys Life Rev. (2010) 7:88–131. doi: 10.1016/j.plrev.2010.01.006
65. Touryan SR, Marian DE, Shimamura AP. Effect of negative emotional pictures on associative memory for peripheral information. Memory. (2007) 15:154–66. doi: 10.1080/09658210601151310
66. Kensinger EA, Garoff-Eaton RJ, Schacter DL. Effects of emotion on memory specificity: Memory trade-offs elicited by negative visually arousing stimuli. J Memory Lang. (2007) 56:575–91. doi: 10.1016/j.jml.2006.05.004
67. Rombold F, Wingenfeld K, Renneberg B, Hellmann-Regen J, Otte C, Roepke S. Influence of the noradrenergic system on the formation of intrusive memories in women: An experimental approach with a trauma film paradigm. Psychol Med. (2016) 46(12):2523–34. doi: 10.1017/S0033291716001379
68. Brewin CR, Dalgleish T, Joseph S. A dual representation theory of posttraumatic stress disorder. psychol Rev. (1996) 103:670. doi: 10.1037/0033-295X.103.4.670
69. Soeter M, Kindt M. Dissociating response systems: erasing fear from memory. Neurobiol Learn Memory. (2010) 94:30–41. doi: 10.1016/j.nlm.2010.03.004
70. Sack M, Spieler D, Wizelman L, Epple G, Stich J, Zaba M, et al. Intranasal oxytocin reduces provoked symptoms in female patients with posttraumatic stress disorder despite exerting sympathomimetic and positive chronotropic effects in a randomized controlled trial. BMC Med. (2017) 15:1–11. doi: 10.1186/s12916-017-0801-0
71. Cyranowski JM, Hofkens TL, Frank E, Seltman H, Cai H-M, Amico JA. Evidence of dysregulated peripheral oxytocin release among depressed women. Psychosom Med. (2008) 70:967. doi: 10.1097/PSY.0b013e318188ade4
72. Dumais KM, Veenema AH. Vasopressin and oxytocin receptor systems in the brain: sex differences and sex-specific regulation of social behavior. Front Neuroendocrinol. (2016) 40:1–23. doi: 10.1016/j.yfrne.2015.04.003
73. Kleim B, Graham B, Bryant RA, Ehlers A. Capturing intrusive re-experiencing in trauma survivors’ daily lives using ecological momentary assessment. J Abnorm Psychol. (2013) 122:998–1009. doi: 10.1037/a0034957
74. Diesfeldt HFA. Analysis of intrusion errors in free recall. Tijdschr Gerontol Geriatr. (2017) 48:134–44. doi: 10.1007/s12439-017-0217-5
Keywords: PTSD, oxytocin, recognition memory, trauma, trauma film paradigm, forced choice memory
Citation: Maslahati T, Glogau F, Galve Gómez M, Buchholz K, Dormann L, Wingenfeld K, Otte C, Schultebraucks K and Roepke S (2025) Oxytocin does not impact forced-choice recognition memory in an experimental trauma film paradigm with healthy women. Front. Psychiatry 16:1421305. doi: 10.3389/fpsyt.2025.1421305
Received: 22 April 2024; Accepted: 18 March 2025;
Published: 03 April 2025.
Edited by:
Alessandra Maria Passarotti, University of Illinois Chicago, United StatesReviewed by:
Luke R. Johnson, University of Tasmania, AustraliaCopyright © 2025 Maslahati, Glogau, Galve Gómez, Buchholz, Dormann, Wingenfeld, Otte, Schultebraucks and Roepke. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Tolou Maslahati, dG9sb3UubWFzbGFoYXRpQGNoYXJpdGUuZGU=
†These authors share senior authorship
‡ORCID: Katharina Schultebraucks, orcid.org/0000-0001-5085-8249
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.