 Peiyan Ni
Peiyan Ni Lingyi Fan4
Lingyi Fan4 Sangmi Chung
Sangmi Chung- 1Department of Neurobiology, Affiliated Mental Health Center and Hangzhou Seventh People's Hospital, Zhejiang University School of Medicine, Hangzhou, China
- 2Liangzhu Laboratory, State Key Laboratory of Brain-Machine Intelligence, MOE Frontier Science Center for Brain Science and Brain-Machine Integration, Zhejiang University, Hangzhou, China
- 3NHC and CAMS Key Laboratory of Medical Neurobiology, Zhejiang University, Hangzhou, China
- 4The Mental Health Center and Psychiatric Laboratory, West China Hospital, Sichuan University, Chengdu, China
- 5Department of Cell Biology and Anatomy, New York Medical College, Valhalla, NY, United States
Psychiatric disorders, such as schizophrenia (SCZ) and autism spectrum disorders (ASD), represent a global health challenge with their poorly understood and complex etiologies. Cortical interneurons (cINs) are the primary inhibitory neurons in the cortex and their subtypes, especially those that are generated from the medial ganglionic emission (MGE) region, have been shown to play an important role in the pathogenesis of these psychiatric disorders. Recent advances in induced pluripotent stem cell (iPSC) technologies provide exciting opportunities to model and study these disorders using human iPSC-derived cINs. In this review, we present a comprehensive overview of various methods employed to generate MGE-type cINs from human iPSCs, which are mainly categorized into induction by signaling molecules vs. direct genetic manipulation. We discuss their advantages, limitations, and potential applications in psychiatric disorder modeling to aid researchers in choosing the appropriate methods based on their research goals. We also provide examples of how these methods have been applied to study the pathogenesis of psychiatric disorders. In addition, we discuss ongoing challenges and future directions in the field. Overall, iPSC-derived cINs provide a powerful tool to model the developmental pathogenesis of psychiatric disorders, thus aiding in uncovering disease mechanisms and potential therapeutic targets. This review article will provide valuable resources for researchers seeking to navigate the complexities of cIN generation methods and their applications in the study of psychiatric disorders.
1 Introduction
Cortical interneurons (cINs), which comprise only about 20–30% of cortical neurons, play critical roles in fine-tuning brain circuit functions; thus, their abnormalities, especially those of medial ganglionic eminence (MGE)-derived cINs, have been associated with various neurodevelopmental psychiatric disorders, such as schizophrenia (SCZ) and autism spectrum disorders (ASD). Proper interactions between inhibitory cINs and excitatory glutamatergic neurons are required to maintain a normal excitation-inhibition (E-I) balance in cortical circuits. Thus, disrupted E-I balances in cortex results in dysfunctional synchronization of neural network oscillations (1, 2), which is thought to underlie the neurodevelopmental pathogenesis of SCZ and ASD (3). However, research on these neurodevelopmental psychiatric disorders has been hampered by a lack of proper model systems that recapitulate patients’ neurodevelopmental process and disease progression since once the patients exhibit the clinical symptoms, it is impossible to go back in time to learn how neurodevelopmental dysregulation led to the pathogenesis of psychiatric disorders. Furthermore, non-neural tissue samples such as peripheral blood mononuclear cells (PBMCs) from patients are less likely to reflect functional deficits in the brain (4). Additionally, postmortem brain samples can reveal full-blown disease phenotypes but not necessarily their pathogenesis mechanisms during neurodevelopment, and moreover, are confounded by factors such as medication history and postmortem interval, etc. (5). Animal models have inherent limitations due to species differences that hinder faithful modeling of human psychiatric pathogenesis (6, 7). Considering that the divergence between human and rodent brains has caused many central nervous system therapeutics validated in rodent models to fail (8–10), it is imperative to study human developmental neurons to understand the pathogenesis mechanisms of cIN-associated neurodevelopmental psychiatric disorders.
The ability to study critical pathogenesis events in the human developmental nervous system once seemed unimaginable due to insurmountable ethical and technical barriers. However, human induced pluripotent stem cell (iPSC) technologies provide a powerful tool to study neurodevelopmental dysregulation by allowing for the generation of subject-specific and disease-relevant developmental brain cells with the same genetic makeup as subject brains in unlimited quantities, thus providing exciting opportunities to model and study neurodevelopmental disorders. Especially, several methods have been reported for the generation of cINs from iPSCs, which would be of utmost interest to scientists who study cIN-associated neurodevelopmental psychiatric disorders. Therefore, in this minireview, we will discuss different methods of cIN generation with their pros and cons, aiming to guide researchers with different questions and demands to select appropriate methods.
2 Generation of cINs from human pluripotent stem cells
The approaches to generate cINs from human pluripotent stem cells can be broadly divided into two major categories: the first method is cIN phenotype induction by signaling molecules to mimic natural developmental process (11) and the second method is direct genetic manipulation, which greatly shortens the differentiation timeline (12–14).
2.1 cIN induction by signaling molecules
One of the major ways to generate cINs from human pluripotent stem cells is by providing signaling molecules during differentiation to mimic the signaling regulations that occur during normal neurodevelopment. This method well recapitulates authentic developmental processes, compared to the transcription factor-induction method that bypasses normal developmental processes. Cells can be differentiated in two dimensions (2D) as monolayer cultures or in three dimensions (3D) as spheres. Regardless of this 2D or 3D culture format, cells are treated to enrich neuroectodermal differentiation by blocking mesoderm and endoderm differentiation with dual SMAD (BMP/TGF-β pathway inhibitors) inhibitors [BMPRIA-Fc (15), Dorsomorphin (16, 17), LDN193189 (18–22), Noggin (23), or SB431542 (15–22, 24)]. Generated neuroepithelial cells can be patterned to become specific neural cells of interest. To inhibit the induction of caudal neuroepithelium phenotypes (25), neuroepithelial cells are also treated with WNT pathway inhibitors [XAV939 (20, 22), DKK1 (15), or IWP2 (16–18, 21, 26)]. Also, these rostral neuroepithelial cells are further ventralized into ganglionic eminence (GE) cell types by sonic hedgehog pathway activators [SHH (19, 20, 22–24), Purmorphamine (15, 19, 20, 22, 27, 28), or SAG (16–18, 21, 26)]. The time window of SHH treatment impacts the phenotype induction, where one group reported generation of diencephalic phenotypes with early SHH activation (20), whereas others observed MGE-phenotype induction with early SHH activation (15, 18). It is not clear what caused such discrepancies, but there was a difference in the reagents utilized for ventralization (SHH, Purmorphamine, and SAG, etc.) with different doses, along with different base media, among these protocols; thus, side by side experiments down the road could help resolve these discrepancies. As a further improvement, some protocols include FGF8 treatment to further rostralize ventral neuroepithelium into medial ganglionic eminence (MGE) phenotypes at the expense of caudal ganglionic eminence (CGE) phenotypes (18, 21), ensuring consistent MGE induction regardless of the endogenous expression level of FGF8. Overall, based on the knowledge gained on the human neurodevelopmental process, the induction of MGE-type cINs is mainly achieved by early inhibition of SMAD and WNT signaling pathways, along with the activation of the SHH signaling pathway, providing a replicable model system for the study of the neurodevelopmental pathogenetic mechanisms. The cINs induced by signaling molecules have been successfully utilized to study the mechanisms of neurodevelopmental disorders. For example, when generated developmental cINs were co-cultured with activated microglia to mimic increased developmental risks by prenatal immune activation, Park et al., found that the SCZ patient-derived cINs presented disrupted metabolic pathways and impaired mitochondrial function, arborization, synapse formation, and synaptic GABA release, which persist in SCZ cINs even after the removal of activated microglia co-culture, but not in healthy control cINs, pointing to gene×environment interactions (29).
2.2 Direct genetic manipulation
Compared to the signaling molecule-based phenotype induction method, the direct genetic manipulation method, which employs exogenous expression of transcription factors that play crucial roles in cIN differentiation, can greatly the shorten phenotype induction timeline. Taking advantage of the knowledge of the transcriptional controls during cIN differentiation, cIN-inducing candidate transcription factors were screened for their ability to induce cINs (12–14, 30). Achaete-scute complex-like homolog 1 (ASCL1, also known as MASH1), which is widely expressed in the embryonic ventral brain and plays a dominant role in determining cIN identity (31–33), was able to trigger the conversion of mouse embryonic fibroblasts into tubulin beta III (TUBB3)+ neuronal cells on its own (30). Furthermore, overexpression of a phospho-mutant form of ASCL1 (five serine residues substituted with alanine, denoted ASA), with their enhanced ability to drive neuronal differentiation (34), resulted in a production of ~2-fold more microtubule-associated protein (MAP2)+ neurons (12). Distal-less homeobox (DLX) family members are multifunctional transcription factors that promote the differentiation of progenitors into cINs and inhibit their differentiation into glial cells (35, 36). When DLX2 was expressed in conjunction with ASCL1, the iPSC-derived neurons were conferred with a cIN fate (12, 13). Likewise, DLX5/DLX6 is also found to promote cIN generation (30). LIM homeobox 6 (LHX6) is a direct target of Nkx2.1, a MGE progenitor-specific transcription factor (37, 38), and favors the fate-specification of MGE precursors into parvalbumin (PV)- or somatostatin (SST)-expressing cINs (12, 14). Combining signaling molecule activation and genetic modification together; Yuan et al. showed that the SHH pathway activation along with the LHX6 induction significantly increased the percentage of PV and SST subtypes compared to the condition without LHX6 overexpression (14). MicroRNA has also been found to benefit the neuronal conversion (39), and Sun et al., reported that the addition of miR-9/9*-124 into the iPSCs with ASCL1, LHX6, and DLX2 overexpression significantly increased the percentage of MAP2+ cells while maintaining the high GABA+/MAP2+ ratio and enhancing dendritic arborization (12). Overall, the rapid induction of MGE-derived cINs from iPSCs can be achieved by the overexpression of transcription factors that can induce their fate determination or maturation, either on their own or supplemented by other signaling molecules. By utilizing direct genetic manipulation, Ishii et al., successfully generated cINs and glutamatergic neurons from iPSCs derived from patients with bipolar disorder (with copy number variations of PCDH15) and SCZ (with copy number variations of RELN) (40), and observed specific abnormalities in the neuronal routing and synaptic function underlying the psychiatric disorders.
2.3 Considerations in cIN induction method selection
For functional studies of mature cINs in vitro, direct genetic manipulation offer specific advantages. Ishii et al. conducted a study where they differentiated GABAergic and glutamatergic mature neurons from iPSCs obtained from bipolar disorder and SCZ patients with copy number variations of PCDH15 and RELN (40). Notably, the authors compared two methods and found that direct genetic manipulation was more efficient and resulted in higher neuron maturity, enabling them to study neuronal routing and synaptic abnormality phenotypes in psychiatric disorders. The researchers in the first example investigated the developmental status of cINs to examine the correlation between prenatal immune activation and SCZ risk. In the second example, the researchers compared the mature functions of cINs with the findings of postmortem brain studies, aiming to identify shared neuronal defects across psychiatric disorders.
Depending on their specific requirements and applications, researchers can choose optimal methods of cIN generation, considering their advantages and disadvantages. The most significant difference between the two methods is the time consumption. Direct genetic regulation can obtain functional cINs in a relatively short period, which is of great use for fast screening to quickly identify potential therapeutic targets. Furthermore, direct genetic manipulation methods can also transdifferentiate terminally differentiated somatic cells (e.g., fibroblasts) directly into cINs (30), which greatly expands the application of this method, especially providing the possibility of generating neuronal populations with an aging signature (41), which is not easy to achieve in the case of iPSC-derived fetal neurons. However, caution needs to be taken, considering the fact that direct genetic modification methods bypass normal neurodevelopmental processes and may not be fully identical to their in vivo counterparts. In addition, the introduction of exogenous factors by viral vectors’ integration into the genome could generate potential confounders during analysis (42), especially when studying genetic risks spread throughout genome as in the case of SCZ.
In contrast, the signaling molecules induction methods require a longer time but closely recapitulate the native neurodevelopmental process, and thus are more suitable to observe and study developmental pathogenetic mechanisms (43, 44). Signaling molecule induction can be combined with self-assembled 3D neuro-organoids differentiation for the generation of more homogeneous cell population within the organoid. Signaling molecule-induced neuro-organoids exhibit cyto-architectural features of developing brains and provide a model where neuronal migration, projections, and circuit formation can be studied in 3D (17, 22, 26). Furthermore, cIN phenotype-induced neuro-organoids have been optimized for large scale spinner culture systems, allowing industrial-scale culture for the use for high throughput drug screening or cell therapy (21).
When choosing the appropriate cIN differentiation methods, it is also necessary to consider the appropriate developmental stage and maturity of cINs depending on the experimental purpose. One needs to keep in mind the fact that signaling molecule induction methods well recapitulate normal neurodevelopmental timeline, especially developmentally protracted maturation of cINs (15, 45), so the researcher needs to determine length of differentiation based on the developmental timeline of interest where specific disease pathogenesis may occur. Overall, signaling molecule-mediated induction methods can be mostly useful to model fetal neurodevelopmental processes. There has been some methods that have shown to facilitate the maturation of human pluripotent stem cell-derived neurons such as co-culture with astrocytes (12, 13, 15, 30), co-culture with glutamatergic neurons (20), and grafting them into the animal brains (14). Still, these methods have not allowed for generation of fully mature fast-spiking cINs like in adult brains. Alternatively, disease modeling for more mature stages of neurons may be accomplished by utilizing genetic modification along with the use of more mature somatic cells as a starting material (41). In case cINs are studied after transplantation into rodent brains, it would be prudent to use post-mitotic neurons to avoid the potential of uncontrolled proliferation that can damage host cyto-architecture. Several reagents have been proven to accelerate cell cycle exit without affecting cIN phenotype, including CultureOne (Invitrogen), NOTCH pathway inhibitors (DAPT, 2634 Tocris), MEK inhibitors (PD0325901 and 4,192 Tocris), and CDK pathway inhibitors (PD0332991 and S1116 Selleck Chemicals) (21, 46). Use of synchronized cIN populations by induced cell cycle exit will be also useful to obtain reliable disease modeling results by avoiding heterogenous maturity within generated cell populations that can confound the assay results.
Based on molecular markers and the expression of neuropeptides or calcium-binding proteins, cINs can be divided into many subtypes such as somatostatin (SST)+, parvalbumin (PV)+, calbindin (CB)+, calretinin (CR)+, neuronal nitric oxide synthase (nNOS), and vasoactive intestinal peptide (VIP)+ cINs. The MGE progenitors give rise mostly to SST+ and PV+ interneurons (47), and among these, PV neurons requires an lengthy time of maturation that matches their protracted maturation during in vivo development (15, 20). Among various subtypes, CB+, CR+, and SST+ cINs start to appear at early differentiation stage during MGE-type cIN generation and their populations increase over time, whereas PV+, nNOS+, and VIP+ neurons can be observed at a very low proportion even after several months’ differentiation (15–17, 20, 27). Hence, one needs to be aware of the changing composition of cIN subtypes as the maturation process goes on, and the timeline of assays may need to be adjusted depending on the cINs subtype of interest.
The cIN induction methods reviewed in this review (Table 1) provide relatively homogeneous cell populations, which are useful for genomics studies without the confounders resulting from changing and heterogeneous cell populations. Such homogeneity can provide advantages in the study of cellular and molecular mechanisms of neurodevelopmental disorders (9, 29, 48–50). However, such homogeneous populations lack interactions among diverse cell types as in the brain. Un-induced organoids or organoids assembled after induction can compensate for such a limitation, providing more physiological cell–cell interactions, and cytoarchitectures (16, 26, 51–53), and even circuit-level functionalities in vitro or in vivo (54–57).
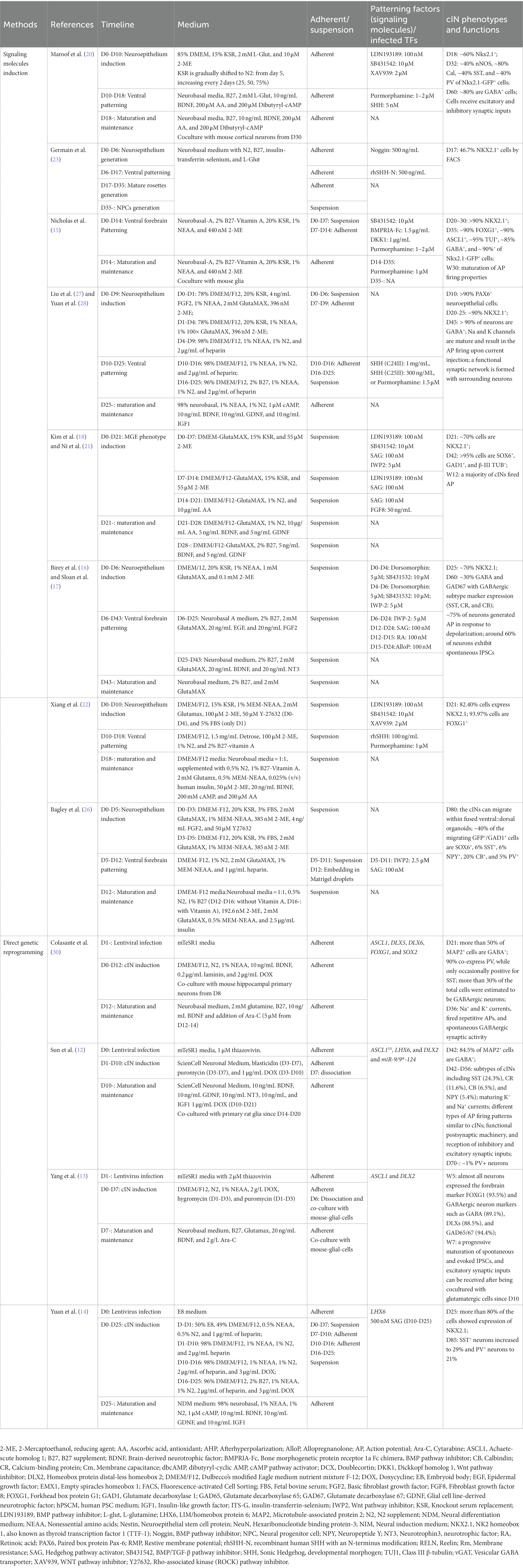
Table 1. The cIN induction methods reviewed in this review.
2.4 Future improvements
To date, iPSC-derived cINs have been increasingly used to study the pathogenesis mechanisms of neurodevelopmental disorders and have shown great potential. However, there are still issues that need to be addressed for the realization of their full potential. Long-term culture processes are needed to obtain more mature cINs, and in general, neuro-organoid cultures are more robust for this purpose than adherent cultures. Still, there are issues with neuro-organoid long-term culture, including insufficient nutrient delivery in large spheres and difficulty with structural maintenance. Possible solutions for these issues are: (1) biocompatible 3D scaffolds that can provide appropriate mechanical support, and whose spatial structure can also provide channels to transport nutrients and oxygen to the cells in the inner layer of the sphere. Such approaches lead to the attenuation of the hypoxic response pathway, lower metabolic dysfunction, and decreased interior cell death (58). (2) vascularization that can create a more physiological microenvironment support for 3D neuro-organoids (59–61). Several methods have been tested for this approach, including co-culturing the neurospheres with endothelial cells (ECs) differentiated from iPSCs (62), or co-culturing iPSCs with human umbilical vein endothelial cells (HUVECs) (63). The vascularized organoids exhibited robust neurogenesis and chemical and electrical synapses in vitro, as well as constructed functional blood vessels inside the grafts and in-between human-mouse interfaces after transplantation in vivo. (3) “trimming” the organoid into slices and exposing the interior of organoids to the culture environment leads to sustained neurogenesis, which also bypasses the diffusion limit to prevent cell death over long-term cultures. This method leads to sustained neurogenesis and the formation of an expanded cortical plate which resembles late-stage cortical development (64).
As mentioned above, the development of fast-spiking PV interneurons and their circuit connectivity is quite challenging to achieve during in vitro differentiation, because of their prolonged maturation process that recapitulates the in vivo developmental process where many of them start to express PV only post-natally and reach adolescence to complete maturation (65, 66). There have been several efforts to facilitate the generation of PV neurons from iPSCs, including the use of a potent adenylate cyclase activator Forskolin (Coleonol) (67) and overexpression of transcription factors of ASCL1 (67) and LHX6 (14). However, it is still a time-consuming process to generate PV interneurons from iPSCs [>2 months to generate ~20% PV+ cINs with the overexpression of LHX6 together with SHH activator (14)], and fully mature fast-spiking-cINs are still difficult to achieve even after months’ culture in vitro or grafting in vivo. To compensate for a low % of PV generation at this gestational period, FACS sorting of PV+ neurons was attempted to enrich them utilizing cell type-specific reporter expression (68). However, achieving their full maturation to model their adult phenotype is still awaiting further technical development.
3 Conclusion
In this review, we discussed diverse methods of generating inhibitory cINs from iPSCs, including the advantages and disadvantages of different methods. Researchers will need to select appropriate cIN generation methods based on the scientific questions and requirements. There are still aspects of current cIN modeling that needs further improvement, such as how to better simulate physiological neurodevelopmental environments and how to generate more mature cell types beyond the fetal neural types generated in many cases. This can be the focus of future research to be optimized. In summary, we systematically reviewed the various methods of cIN generation from human pluripotent stem cells, which will provide valuable tools to study the mechanisms of neurodevelopmental disorders in real human tissues during developmental time points of their vulnerability and to develop novel therapeutics based on a human model system.
Author contributions
SC: Supervision, Validation, Writing – original draft, Writing – review & editing. PN: Supervision, Validation, Writing – original draft, Writing – review & editing. LF: Writing – original draft, Writing – review & editing. YJ: Conceptualization, Writing – review & editing. CZ: Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was supported by NS129630 (SC), MH133205 (SC), MH131610 (SC), and the National Natural Science Foundation of China (82071502 [PN], 81871054 [PN]).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Chattopadhyaya, B, and Cristo, GD. Gabaergic circuit dysfunctions in neurodevelopmental disorders. Front Psych. (2012) 3:51. doi: 10.3389/fpsyt.2012.00051
2. Gonzalez-Burgos, G, Fish, KN, and Lewis, DA. Gaba neuron alterations, cortical circuit dysfunction and cognitive deficits in schizophrenia. Neural Plast. (2011) 2011:723184. doi: 10.1155/2011/723184
3. Yang, J, Yang, X, and Tang, K. Interneuron development and dysfunction. FEBS J. (2021) 289:2318–36. doi: 10.1111/febs.15872
4. Sequeira, A, Martin, MV, Rollins, B, Moon, EA, Bunney, WE, Macciardi, F, et al. Mitochondrial mutations and polymorphisms in psychiatric disorders. Front Genet. (2012) 3:103. doi: 10.3389/fgene.2012.00103
5. Ramaker, RC, Bowling, KM, Lasseigne, BN, Hagenauer, MH, Hardigan, AA, Davis, NS, et al. Post-mortem molecular profiling of three psychiatric disorders. Genome Med. (2017) 9:72. doi: 10.1186/s13073-017-0458-5
6. Kaiser, T, and Feng, G. Modeling psychiatric disorders for developing effective treatments. Nat Med. (2015) 21:979–88. doi: 10.1038/nm.3935
7. Salgado, JV, and Sandner, G. A critical overview of animal models of psychiatric disorders: challenges and perspectives. Rev Bras Psiquiatr. (2013) 35:S77–81. doi: 10.1590/1516-4446-2013-1156
8. Hodge, RD, Bakken, TE, Miller, JA, Smith, KA, Barkan, ER, Graybuck, LT, et al. Conserved cell types with divergent features in human versus mouse cortex. Nature. (2019) 573:61–8. doi: 10.1038/s41586-019-1506-7
9. Shao, Z, Noh, H, Bin Kim, W, Ni, P, Nguyen, C, Cote, SE, et al. Dysregulated protocadherin-pathway activity as an intrinsic defect in induced pluripotent stem cell-derived cortical interneurons from subjects with schizophrenia. Nat Neurosci. (2019) 22:229–42. doi: 10.1038/s41593-018-0313-z
10. Zhao, X, and Bhattacharyya, A. Human models are needed for studying human neurodevelopmental disorders. Am J Hum Genet. (2018) 103:829–57. doi: 10.1016/j.ajhg.2018.10.009
11. Simão, D, Silva, MM, Terrasso, AP, Arez, F, Sousa, MFQ, Mehrjardi, NZ, et al. Recapitulation of human neural microenvironment signatures in IPSC-derived NPC 3D differentiation. Stem Cell Rep. (2018) 11:552–64. doi: 10.1016/j.stemcr.2018.06.020
12. Sun, AX, Yuan, Q, Tan, S, Xiao, Y, Wang, D, Khoo, AT, et al. Direct induction and functional maturation of forebrain gabaergic neurons from human pluripotent stem cells. Cell Rep. (2016) 16:1942–53. doi: 10.1016/j.celrep.2016.07.035
13. Yang, N, Chanda, S, Marro, S, Ng, Y-H, Janas, JA, Haag, D, et al. Generation of pure Gabaergic neurons by transcription factor programming. Nat Methods. (2017) 14:621–8. doi: 10.1038/nmeth.4291
14. Yuan, F, Chen, X, Fang, KH, Wang, Y, Lin, M, Xu, SB, et al. Induction of human somatostatin and parvalbumin neurons by expressing a single transcription factor Lim homeobox 6. elife. (2018) 7:e37382. doi: 10.7554/eLife.37382
15. Nicholas, CR, Chen, J, Tang, Y, Southwell, DG, Chalmers, N, Vogt, D, et al. Functional maturation of HPSC-derived forebrain interneurons requires an extended timeline and mimics human neural development. Cell Stem Cell. (2013) 12:573–86. doi: 10.1016/j.stem.2013.04.005
16. Birey, F, Andersen, J, Makinson, CD, Islam, S, Wei, W, Huber, N, et al. Assembly of functionally integrated human forebrain spheroids. Nature. (2017) 545:54–9. doi: 10.1038/nature22330
17. Sloan, SA, Andersen, J, PaȘca, AM, Birey, F, and PaȘca, SP. Generation and assembly of human brain region-specific three-dimensional cultures. Nat Protoc. (2018) 13:2062–85. doi: 10.1038/s41596-018-0032-7
18. Kim, T-G, Yao, R, Monnell, T, Cho, J-H, Vasudevan, A, Koh, A, et al. Efficient specification of interneurons from human pluripotent stem cells by dorsoventral and rostrocaudal modulation. Stem Cells. (2014) 32:1789–804. doi: 10.1002/stem.1704
19. Kriks, S, Shim, J-W, Piao, J, Ganat, YM, Wakeman, DR, Xie, Z, et al. Dopamine neurons derived from human Es cells efficiently engraft in animal models of Parkinson’s disease. Nature. (2011) 480:547–51. doi: 10.1038/nature10648
20. Maroof, AM, Keros, S, Tyson, JA, Ying, SW, Ganat, YM, Merkle, FT, et al. Directed differentiation and functional maturation of cortical interneurons from human embryonic stem cells. Cell Stem Cell. (2013) 12:559–72. doi: 10.1016/j.stem.2013.04.008
21. Ni, P, Noh, H, Shao, Z, Zhu, Q, Guan, Y, Park, JJ, et al. Large-scale generation and characterization of homogeneous populations of migratory cortical interneurons from human pluripotent stem cells. Mol Ther Methods Clin Dev. (2019) 13:414–30. doi: 10.1016/j.omtm.2019.04.002
22. Xiang, Y, Tanaka, Y, Patterson, B, Kang, YJ, Govindaiah, G, Roselaar, N, et al. Fusion of regionally specified HPSC-derived organoids models human brain development and interneuron migration. Cell Stem Cell. (2017) 21:383–398.e7. doi: 10.1016/j.stem.2017.07.007
23. Germain, ND, Banda, EC, Becker, S, Naegele, JR, and Grabel, LB. Derivation and isolation of Nkx2.1-positive basal forebrain progenitors from human embryonic stem cells. Stem Cells Dev. (2013) 22:1477–89. doi: 10.1089/scd.2012.0264
24. Chambers, SM, Fasano, CA, Papapetrou, EP, Tomishima, M, Sadelain, M, and Studer, L. Highly efficient neural conversion of human Es and ips cells by dual inhibition of Smad signaling. Nat Biotechnol. (2009) 27:275–80. doi: 10.1038/nbt.1529
25. Watanabe, K, Kamiya, D, Nishiyama, A, Katayama, T, Nozaki, S, Kawasaki, H, et al. Directed differentiation of telencephalic precursors from embryonic stem cells. Nat Neurosci. (2005) 8:288–96. doi: 10.1038/nn1402
26. Bagley, JA, Reumann, D, Bian, S, Lévi-Strauss, J, and Knoblich, JA. Fused cerebral organoids model interactions between brain regions. Nat Methods. (2017) 14:743–51. doi: 10.1038/nmeth.4304
27. Liu, Y, Liu, H, Sauvey, C, Yao, L, Zarnowska, ED, and Zhang, S-C. Directed differentiation of forebrain Gaba interneurons from human pluripotent stem cells. Nat Protoc. (2013) 8:1670–9. doi: 10.1038/nprot.2013.106
28. Yuan, F, Fang, K-H, Cao, S-Y, Qu, Z-Y, Li, Q, Krencik, R, et al. Efficient generation of region-specific forebrain neurons from human pluripotent stem cells under highly defined condition. Sci Rep. (2015) 5:18550. doi: 10.1038/srep18550
29. Park, GH, Noh, H, Shao, Z, Ni, P, Qin, Y, Liu, D, et al. Activated microglia cause metabolic disruptions in developmental cortical interneurons that persist in interneurons from individuals with schizophrenia. Nat Neurosci. (2020) 23:1352–64. doi: 10.1038/s41593-020-00724-1
30. Colasante, G, Lignani, G, Rubio, A, Medrihan, L, Yekhlef, L, Sessa, A, et al. Rapid conversion of fibroblasts into functional forebrain gabaergic interneurons by direct genetic reprogramming. Cell Stem Cell. (2015) 17:719–34. doi: 10.1016/j.stem.2015.09.002
31. Fode, C, Ma, Q, Casarosa, S, Ang, SL, Anderson, DJ, and Guillemot, F. A role for neural determination genes in specifying the dorsoventral identity of telencephalic neurons. Genes Dev. (2000) 14:67–80. doi: 10.1101/gad.14.1.67
32. Long, JE, Cobos, I, Potter, GB, and Rubenstein, JL. Dlx1&2 and Mash1 transcription factors control Mge and Cge patterning and differentiation through parallel and overlapping pathways. Cereb Cortex. (2009) 19:i96–i106. doi: 10.1093/cercor/bhp045
33. Schuurmans, C, and Guillemot, F. Molecular mechanisms underlying cell fate specification in the developing telencephalon. Curr Opin Neurobiol. (2002) 12:26–34. doi: 10.1016/S0959-4388(02)00286-6
34. Ali, FR, Cheng, K, Kirwan, P, Metcalfe, S, Livesey, FJ, Barker, RA, et al. The phosphorylation status of Ascl1 is a key determinant of neuronal differentiation and maturation in vivo and in vitro. Development. (2014) 141:2216–24. doi: 10.1242/dev.106377
35. Anastasiades, PG, and Butt, SJ. Decoding the transcriptional basis for Gabaergic interneuron diversity in the mouse neocortex. Eur J Neurosci. (2011) 34:1542–52. doi: 10.1111/j.1460-9568.2011.07904.x
36. Wonders, C, and Anderson, SA. Cortical interneurons and their origins. Neuroscientist. (2005) 11:199–205. doi: 10.1177/1073858404270968
37. Du, T, Xu, Q, Ocbina, PJ, and Anderson, SA. Nkx2.1 specifies cortical interneuron fate by activating Lhx6. Development. (2008) 135:1559–67. doi: 10.1242/dev.015123
38. Neves, G, Shah, MM, Liodis, P, Achimastou, A, Denaxa, M, Roalfe, G, et al. The Lim homeodomain protein Lhx6 regulates maturation of interneurons and network excitability in the mammalian cortex. Cereb Cortex. (2013) 23:1811–23. doi: 10.1093/cercor/bhs159
39. Yoo, AS, Sun, AX, Li, L, Shcheglovitov, A, Portmann, T, Li, Y, et al. Microrna-mediated conversion of human fibroblasts to neurons. Nature. (2011) 476:228–31. doi: 10.1038/nature10323
40. Ishii, T, Ishikawa, M, Fujimori, K, Maeda, T, Kushima, I, Arioka, Y, et al. In vitro modeling of the bipolar disorder and schizophrenia using patient-derived induced pluripotent stem cells with copy number variations of PCDH15 and RELN. eNeuro. (2019) 6:ENEURO.0403-18.2019. doi: 10.1523/ENEURO.0403-18.2019
41. Mertens, J, Paquola, ACM, Ku, M, Hatch, E, Böhnke, L, Ladjevardi, S, et al. Directly reprogrammed human neurons retain aging-associated transcriptomic signatures and reveal age-related nucleocytoplasmic defects. Cell Stem Cell. (2015) 17:705–18. doi: 10.1016/j.stem.2015.09.001
42. Fernandopulle, MS, Prestil, R, Grunseich, C, Wang, C, Gan, L, and Ward, ME. Transcription factor-mediated differentiation of human IPSCS into neurons. Curr Protoc Cell Biol. (2018) 79:e51. doi: 10.1002/cpcb.51
43. Mariani, J, Coppola, G, Zhang, P, Abyzov, A, Provini, L, Tomasini, L, et al. Foxg1-dependent dysregulation of gaba/glutamate neuron differentiation in autism spectrum disorders. Cells. (2015) 162:375–90. doi: 10.1016/j.cell.2015.06.034
44. Wang, P, Mokhtari, R, Pedrosa, E, Kirschenbaum, M, Bayrak, C, Zheng, D, et al. Crispr/Cas9-mediated heterozygous knockout of the autism gene Chd8 and characterization of its transcriptional networks in cerebral organoids derived from IPS cells. Mol Autism. (2017) 8:11. doi: 10.1186/s13229-017-0124-1
45. Anderson, S, and Vanderhaeghen, P. Cortical neurogenesis from pluripotent stem cells: complexity emerging from simplicity. Curr Opin Neurobiol. (2014) 27:151–7. doi: 10.1016/j.conb.2014.03.012
46. Bershteyn, M, Broer, S, Parekh, M, Maury, Y, Havlicek, S, Kriks, S, et al. Human pallial Mge-type Gabaergic interneuron cell therapy for chronic focal epilepsy. Cell Stem Cell. (2023) 30:1331–1350.e11. doi: 10.1016/j.stem.2023.08.013
47. Le Magueresse, C, and Monyer, H. Gabaergic interneurons shape the functional maturation of the cortex. Neuron. (2013) 77:388–405. doi: 10.1016/j.neuron.2013.01.011
48. Lu, K, Hong, Y, Tao, M, Shen, L, Zheng, Z, Fang, K, et al. Depressive patient-derived Gaba interneurons reveal abnormal neural activity associated with Htr2C. EMBO Mol Med. (2023) 15:e16364. doi: 10.15252/emmm.202216364
49. Xu, L, Huo, HQ, Lu, KQ, Tang, XY, Hong, Y, Han, X, et al. Abnormal mitochondria in Down syndrome ipsc-derived Gabaergic interneurons and organoids. Biochim Biophys Acta Mol basis Dis. (2022) 1868:166388. doi: 10.1016/j.bbadis.2022.166388
50. Xu, R, Brawner, AT, Li, S, Liu, JJ, Kim, H, Xue, H, et al. Olig2 drives abnormal neurodevelopmental phenotypes in human IPSC-based organoid and chimeric mouse models of down syndrome. Cell Stem Cell. (2019) 24:908–926.e8. doi: 10.1016/j.stem.2019.04.014
51. Kathuria, A, Lopez-Lengowski, K, Jagtap, SS, Mcphie, D, Perlis, RH, Cohen, BM, et al. Transcriptomic landscape and functional characterization of induced pluripotent stem cell-derived cerebral organoids in schizophrenia. JAMA Psychiatry. (2020) 77:745–54. doi: 10.1001/jamapsychiatry.2020.0196
52. Li, C, Fleck, JS, Martins-Costa, C, Burkard, TR, Themann, J, Stuempflen, M, et al. Single-cell brain organoid screening identifies developmental defects in autism. Nature. (2023) 621:373–80. doi: 10.1038/s41586-023-06473-y
53. Notaras, M, Lodhi, A, Dündar, F, Collier, P, Sayles, NM, Tilgner, H, et al. Schizophrenia is defined by cell-specific neuropathology and multiple neurodevelopmental mechanisms in patient-derived cerebral organoids. Mol Psychiatry. (2022) 27:1416–34. doi: 10.1038/s41380-021-01316-6
54. Dong, X, Xu, SB, Chen, X, Tao, M, Tang, XY, Fang, KH, et al. Human cerebral organoids establish subcortical projections in the mouse brain after transplantation. Mol Psychiatry. (2021) 26:2964–76. doi: 10.1038/s41380-020-00910-4
55. Revah, O, Gore, F, Kelley, KW, Andersen, J, Sakai, N, Chen, X, et al. Maturation and circuit integration of transplanted human cortical organoids. Nature. (2022) 610:319–26. doi: 10.1038/s41586-022-05277-w
56. Sharf, T, Van Der Molen, T, Glasauer, SMK, Guzman, E, Buccino, AP, Luna, G, et al. Functional neuronal circuitry and oscillatory dynamics in human brain organoids. Nat Commun. (2022) 13:4403. doi: 10.1038/s41467-022-32115-4
57. Trujillo, CA, Gao, R, Negraes, PD, Gu, J, Buchanan, J, Preissl, S, et al. Complex oscillatory waves emerging from cortical organoids model early human brain network development. Cell Stem Cell. (2019) 25:558–569.e7. doi: 10.1016/j.stem.2019.08.002
58. Fiorenzano, A, Sozzi, E, Birtele, M, Kajtez, J, Giacomoni, J, Nilsson, F, et al. Single-cell transcriptomics captures features of human midbrain development and dopamine neuron diversity in brain organoids. Nat Commun. (2021) 12:7302. doi: 10.1038/s41467-021-27464-5
59. Cakir, B, Xiang, Y, Tanaka, Y, Kural, MH, Parent, M, Kang, YJ, et al. Engineering of human brain organoids with a functional vascular-like system. Nat Methods. (2019) 16:1169–75. doi: 10.1038/s41592-019-0586-5
60. Song, L, Yuan, X, Jones, Z, Griffin, K, Zhou, Y, Ma, T, et al. Assembly of human stem cell-derived cortical spheroids and vascular spheroids to model 3-d brain-like tissues. Sci Rep. (2019) 9:5977. doi: 10.1038/s41598-019-42439-9
61. Sun, XY, Ju, XC, Li, Y, Zeng, PM, Wu, J, Zhou, YY, et al. Generation of vascularized brain organoids to study neurovascular interactions. elife. (2022) 11:e76707. doi: 10.7554/eLife.76707
62. Pham, MT, Pollock, KM, Rose, MD, Cary, WA, Stewart, HR, Zhou, P, et al. Generation of human vascularized brain organoids. Neuroreport. (2018) 29:588–93. doi: 10.1097/WNR.0000000000001014
63. Shi, Y, Sun, L, Wang, M, Liu, J, Zhong, S, Li, R, et al. Vascularized human cortical organoids (vOrganoids) model cortical development in vivo. PLoS Biol. (2020) 18:e3000705. doi: 10.1371/journal.pbio.3000705
64. Qian, X, Su, Y, Adam, CD, Deutschmann, AU, Pather, SR, Goldberg, EM, et al. Sliced human cortical organoids for modeling distinct cortical layer formation. Cell Stem Cell. (2020) 26:766–781.e9. doi: 10.1016/j.stem.2020.02.002
65. Baho, E, Chattopadhyaya, B, Lavertu-Jolin, M, Mazziotti, R, Awad, PN, Chehrazi, P, et al. p75 neurotrophin receptor activation regulates the timing of the maturation of cortical parvalbumin interneuron connectivity and promotes juvenile-like plasticity in adult visual cortex. J Neurosci. (2019) 39:4489–510. doi: 10.1523/JNEUROSCI.2881-18.2019
66. Chattopadhyaya, B, Di Cristo, G, Higashiyama, H, Knott, GW, Kuhlman, SJ, Welker, E, et al. Experience and activity-dependent maturation of perisomatic Gabaergic innervation in primary visual cortex during a postnatal critical period. J Neurosci. (2004) 24:9598–611. doi: 10.1523/JNEUROSCI.1851-04.2004
67. Shi, Z, Zhang, J, Chen, S, Li, Y, Lei, X, Qiao, H, et al. Conversion of fibroblasts to parvalbumin neurons by one transcription factor, ascl1, and the chemical compound forskolin. J Biol Chem. (2016) 291:13560–70. doi: 10.1074/jbc.M115.709808
Keywords: interneurons, induced pluripotent stem cells, signal transduction, gene editing, GABAergic neurons, neurodevelopmental disorders, mental disorders, in vitro techniques
Citation: Ni P, Fan L, Jiang Y, Zhou C and Chung S (2023) From cells to insights: the power of human pluripotent stem cell-derived cortical interneurons in psychiatric disorder modeling. Front. Psychiatry. 14:1336085. doi: 10.3389/fpsyt.2023.1336085
Edited by:
Sehyoun Yoon, Northwestern Medicine, United StatesReviewed by:
Masaru Tanaka, University of Szeged (ELKH-SZTE), HungaryCopyright © 2023 Ni, Fan, Jiang, Zhou and Chung. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Peiyan Ni, cGVpeWFubmlAemp1LmVkdS5jbg==; Sangmi Chung, c2NodW5nOEBueW1jLmVkdQ==