
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Psychiatry , 04 August 2022
Sec. Aging Psychiatry
Volume 13 - 2022 | https://doi.org/10.3389/fpsyt.2022.925012
 Lucong Liang1†
Lucong Liang1† Jing Yan2†
Jing Yan2† Xiaohua Huang3†
Xiaohua Huang3† Chun Zou1
Chun Zou1 Liechun Chen1Rongjie Li2,4
Liechun Chen1Rongjie Li2,4 Jieqiong Xie1
Jieqiong Xie1 Mika Pan1
Mika Pan1 Donghua Zou1*Ying Liu2,4*
Donghua Zou1*Ying Liu2,4*Background: Alzheimer’s disease (AD) and sleep disorders are both neurodegenerative conditions characterized by impaired or absent sleep. However, potential common pathogenetic mechanisms of these diseases are not well characterized.
Methods: Differentially expressed genes (DEGs) were identified using publicly available human gene expression profiles GSE5281 for AD and GSE40562 for sleep disorder. DEGs common to the two datasets were used for enrichment analysis, and we performed multi-scale embedded gene co-expression network analysis (MEGENA) for common DEGs. Fast gene set enrichment analysis (fGSEA) was used to obtain common pathways, while gene set variation analysis (GSVA) was applied to quantify those pathways. Subsequently, we extracted the common genes between module genes identified by MEGENA and genes of the common pathways, and we constructed protein-protein interaction (PPI) networks. The top 10 genes with the highest degree of connectivity were classified as hub genes. Common genes were used to perform Metascape enrichment analysis for functional enrichment. Furthermore, we quantified infiltrating immune cells in patients with AD or sleep disorder and in controls.
Results: DEGs common to the two disorders were involved in the citrate cycle and the HIF-1 signaling pathway, and several common DEGs were related to signaling pathways regulating the pluripotency of stem cells, as well as 10 other pathways. Using MEGENA, we identified 29 modules and 1,498 module genes in GSE5281, and 55 modules and 1,791 module genes in GSE40562. Hub genes involved in AD and sleep disorder were ATP5A1, ATP5B, COX5A, GAPDH, NDUFA9, NDUFS3, NDUFV2, SOD1, UQCRC1, and UQCRC2. Plasmacytoid dendritic cells and T helper 17 cells had the most extensive infiltration in both AD and sleep disorder.
Conclusion: AD pathology and pathways of neurodegeneration participate in processes contributing in AD and sleep disorder. Hub genes may be worth exploring as potential candidates for targeted therapy of AD and sleep disorder.
Neurodegenerative disease is a general designation for conditions involving progressive loss of structure or function of the nervous system, leading ultimately to neuronal death, including Alzheimer’s disease (AD) and prion disease (1). These diseases vary in their pathophysiology: some can cause memory and cognitive impairments, while others affect the ability to move, speak, and breathe (2, 3). The symptoms of AD begin to develop with mild memory impairment and evolve into cognitive impairment and dysfunction in complex daily activities (4). AD can be ultimately fatal. Cognitive deficits are ameliorated by reduction in amyloid β accumulation (5). AD is one of the most common factors of dementia and frailty (4), which occurs mostly in people over 65 years (6). The presence of amyloid β and tau proteins is a defining characteristic of AD, characterized by loss of neurons and synapses in the cerebral cortex and certain subcortical regions (7). Nearly half of all AD adults older than 60 years also report sleep disturbances (8). In addition, disturbed sleep or lack of sleep has been identified as one of the risk factors for AD (9).
The pathology of AD may overlap with the pathology of other diseases; for example, some patients with prion mutations are initially diagnosed with AD (10). Prion diseases are neurological disorders characterized by neuronal loss, spongiform degeneration, and activation of astrocytes or microglia cells. Although rare in humans, prion diseases can lead to dementia and ataxia (11). Fatal familial insomnia (FFI) is a prion disease that involves a mutation in codon 178 of the gene encoding the prion protein (PRNP) (12). The disease is characterized by loss of sleep, abnormal autonomic function, and selective atrophy, usually without spongiform changes (13). The thalamus is the main brain region affected in this sleep disorder (6), and FFI has been considered to be dominated by sleep disorder. The disease is currently incurable and has a mean course of 18 months, ultimately leading to death (14).
There is growing evidence that poor sleep accelerates the progression of neurodegenerative disorders and may play a role in their pathogenesis (15). Since sleep disorders are present in AD, we need to understand the pathogenic factors and mechanisms of AD and sleep disorder in order to design effective treatments. Although sleep disorders and AD present different pathogenic factors and clinical features, DNA damage or accumulation of misfolded proteins may be activated as a common mechanism in the pathogenesis of both conditions, ultimately leading to irreversible neuronal loss and cell death (16, 17).
In our previous studies, we also found several biomarkers may contribute to AD. CXCR4, EGFR, MAP4K4, IGF1R (18), and RBM8A (19) may serve as biomarkers for AD diagnosis. A study identified significant marker genes, which could be related to a potential AD molecular type (20). Studies have shown that REPS1 serves as a potential biomarker for AD and Vascular dementia that were both associated with Ras signaling (21). Furthermore, lncRNAs and miRNAs determining the progression of AD, including miR-34a (22) and long ncRNA MALAT1 (23). Chan has reported that high expression of PMP2, KIF5B, and ADD3 as common molecule between AD and sleep disorders that involved in Ras signaling (24). These genes can be used for the diagnosis and treatment of neurodegenerative diseases, and we need to further exploration and verification.
However, relevant mechanisms of AD and sleep disorder are not well characterized and need to be further explored. In the present study, we used bioinformatics analyses to investigate the similarities and differences in gene expression among patients with sleep disorder or AD in order to identify hub genes and intersecting pathways. The resulting insights may help guide experimental studies to elucidate pathways common to both types of disease.
The workflow of the study is shown in Figure 1. Gene expression profiles from publicly available datasets GSE5281 and GSE40562 based on the GPL570 platform (Affymetrix Human Genome U133 Plus 2.0 Array) were downloaded from the Gene Expression Omnibus database (GEO)1 (25). The array data for GSE5281 consisted of 87 AD samples (obtained from six brain regions as follows: 10 entorhinal cortex, 10 hippocampus, 16 medial temporal gyrus, 9 posterior cingulate, 23 superior frontal gyrus, and 19 primary visual cortex) and 74 controls (obtained from the same six brain regions: 13 entorhinal cortex, 13 hippocampus, 12 medial temporal gyrus, 13 posterior cingulate, 11 superior frontal gyrus, and 12 primary visual cortex). The individuals included in this dataset had a mean age of 79.9 ± 6.9 years (26). A total of eight samples of thalamus and parietal lobe were analyzed in the GSE40562 dataset, from three patients with sleep disorder and one normal individual. The three patients included a 48-year-old man and a 26-year-old female from the same family (27), as well as a 55-year-old man.
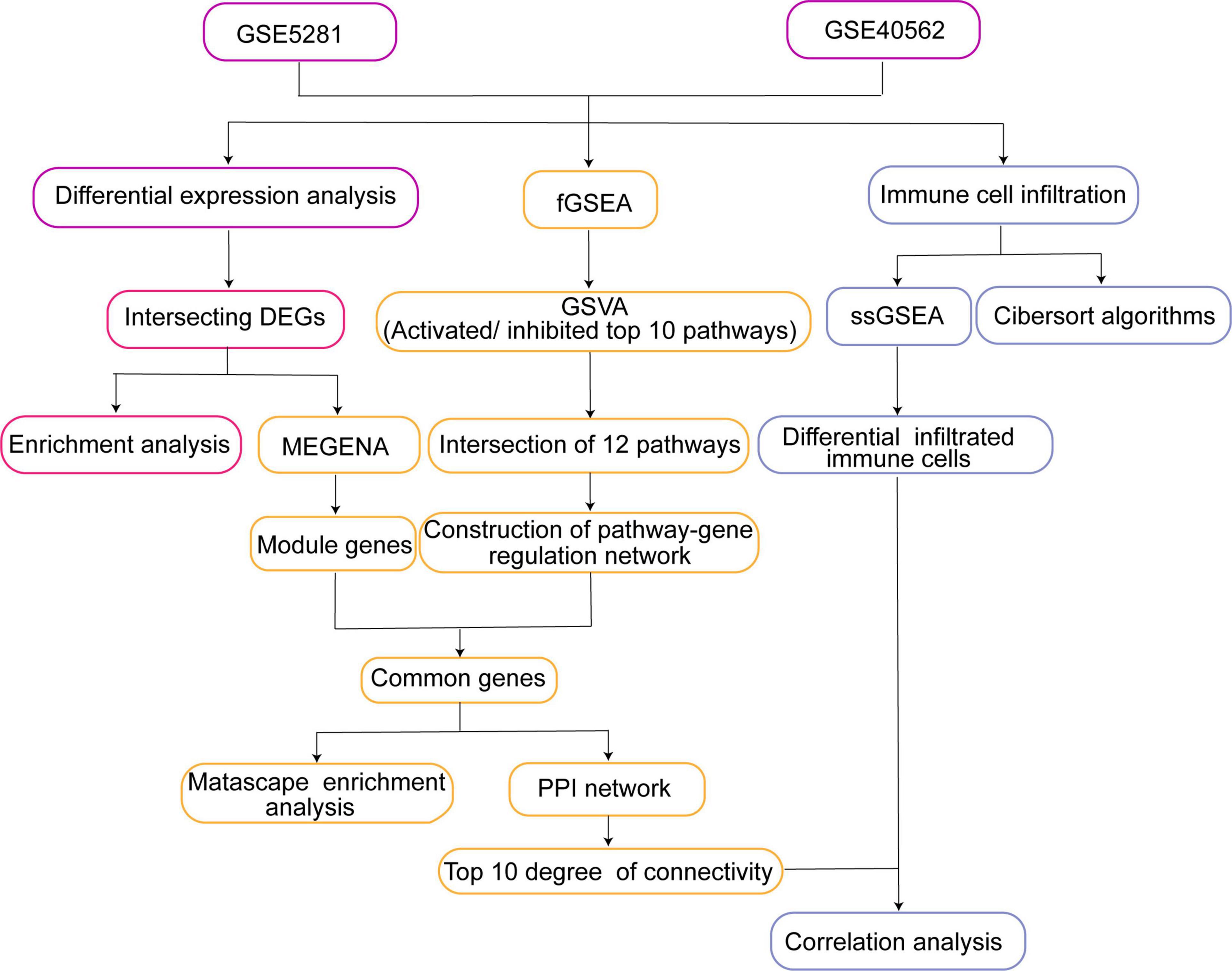
Figure 1. Study workflow. DEG, differentially expressed gene; fGSEA, fast gene set enrichment analysis; MEGENA, multiscale embedded gene co-expression network analysis; GSVA, gene set variation analysis; PPI, protein-protein interaction; ssGSEA, single-sample gene set enrichment analysis.
Differentially expressed genes (DEGs) between patients with AD or sleep disorder and controls were identified in GSE5281 and GSE40562 datasets using the “limma” package in R (28). DEGs were defined as genes showing adjusted P < 0.05. The intersecting DEGs common to the two datasets were identified using the ggVennDiagram package (29), with selection criteria defined as P < 0.05 and log2(fold change) > 0.5. Only intersecting DEGs were analyzed further.
We analyzed the intersecting DEGs for enrichment in Gene Ontology (GO) terms and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways. GO terms, consisting of molecular functions, biological processes, and cell components, were analyzed using the clusterProfiler package (30) in R. P < 0.05 was considered to indicate a statistically significant difference.
Fast gene set enrichment analysis (fGSEA) was performed to obtain common pathways using the expression profiles of GSE5281 and GSE40562. Gene set variation analysis (GSVA) is a non-parametric unsupervised analysis method to evaluate the gene set enrichment of chips and transcriptomes (31). The GSVA algorithm was applied to quantify pathways common to AD and sleep disorder. The Cytoscape tool (32) was used to explore interactions among common pathways and genes.
We examined the intersecting DEGs using multiscale embedded gene co-expression network analysis (MEGENA), which can effectively improve performance over well-established clustering method. We constructed and analyzed large-scale planar filtered co-expression networks using the MEGENA package in R (33). Filtered networks were used to calculate correlations among intersecting DEGs and thereby identify module genes.
PPIs help predict how cells function in health and disease (34). We identified module genes that also participated in pathways involved in both AD and sleep disorder, then we used these genes to construct a PPI network, which was visualized using Cytoscape (32). Among them, the top 10 genes with highest degree of connectivity were identified as hub genes in AD and sleep disorder. Furthermore, common genes to both disorders were used to explore potential functional aspects using Metascape tool (35). The expression of common genes was visualized in a heat map created using “pheatmap” package (23) and compared with the expression of hub genes, which were visualized in AD, sleep disorder, and controls.
The levels of 24 types of immune cells that infiltrated tissues of AD patients, sleep disorder patients and controls were investigated using single-sample gene set enrichment analysis (ssGSEA) and CIBERSORT.2 We also used the top 10 genes with highest degree of connectivity in the PPI network and immune cell infiltration data to perform Pearson correlation analysis using the “ggstatsplot” package.
A total of 2,057 DEGs were identified in AD patients compared to controls in GSE5281, including 509 upregulated and 1,548 downregulated genes (Figure 2A). We identified 2,826 DEGs between sleep disorder patients and control individuals in GSE40562, including 726 upregulated and 2,100 downregulated genes (Figure 2B).
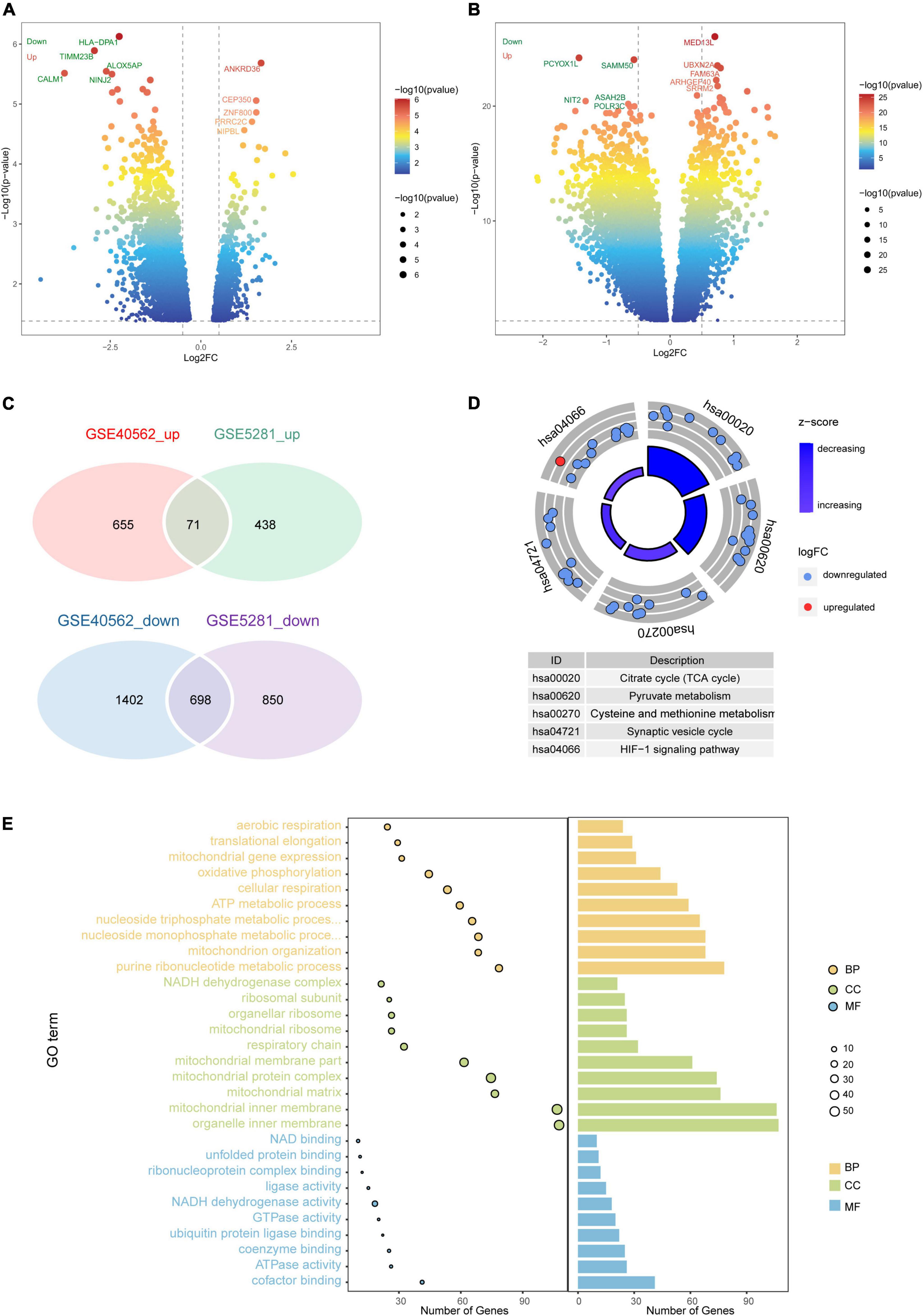
Figure 2. Differential gene expression and enrichment analysis. Volcano plot of differentially expressed genes (DEGs) between (A) patients with Alzheimer’s disease (AD) and controls in GSE40562, or (B) patients with sleep disorder and controls in GSE5281. Red color indicates upregulated genes; green, downregulated genes. (C) Venn diagrams showing common up- or downregulated genes in AD, sleep disorder, and controls. (D) Intersecting DEGs were predicted to be involved in five pathways. (E) Protein functional analysis in terms of biological processes, cellular components, and molecular functions. GO, Gene Ontology; BP, biological process; CC, cellular component; MF, molecular function; log2FC, log2(fold change).
Among the DEGs, 769 were intersecting DEGs found in both GSE5281 and GSE40562 (71 upregulated and 698 downregulated) (Figure 2C). Intersecting DEGs were involved mainly in the hypoxia-inducible family (HIF)-1 signaling pathway and tricarboxylic acid (TCA) cycle (Figure 2D). The intersecting DEGs were enriched in the GO biological processes of cellular respiration and adenosine triphosphate metabolism, the GO cell components of mitochondrial inner membrane and mitochondrial protein complexes, and the GO molecular functions of nicotinamide adenine dinucleotide (NAD) binding and NADH dehydrogenase activity (Figure 2E).
fGSEA showed that some intersecting DEGs were involved in the activation of signaling pathways regulating the pluripotency of stem cells and breast cancer, while others were linked to inhibition of AD and Parkinson’s disease (Figures 3A,B). Our results identified 12 pathways common to AD and sleep disorder: non-alcoholic fatty liver disease, diabetic cardiomyopathy, chemical carcinogenesis-reactive oxygen species, thermogenesis, Parkinson’s disease, prion disease, Huntington’s disease, amyotrophic lateral sclerosis, AD, and pathways related to neurodegeneration in multiple diseases. Twenty pathways were quantified by GSVA, which identified 12 pathways with high scores common to AD and sleep disorder (Figures 3C,D).
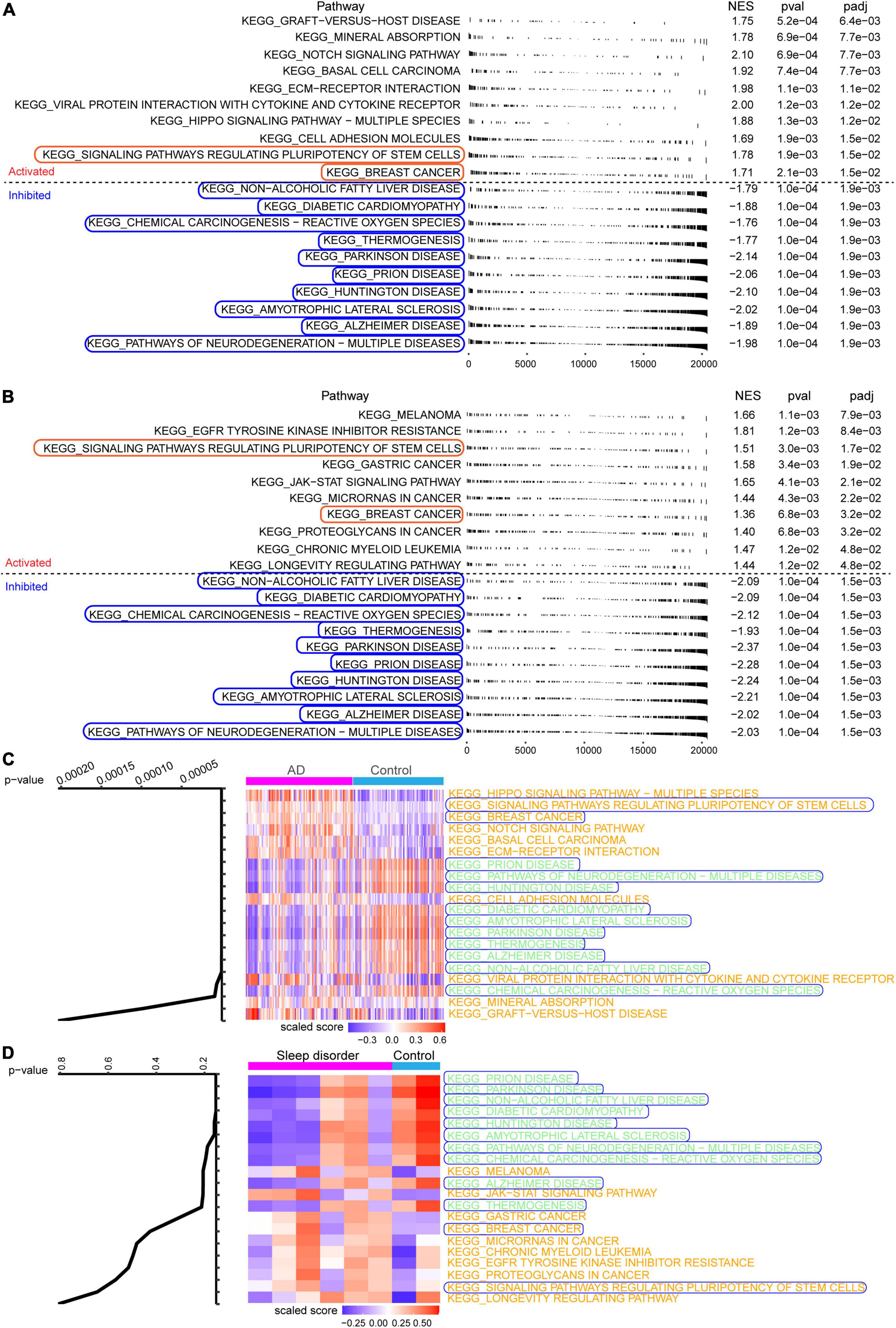
Figure 3. Fast gene set enrichment analysis (fGSEA) and gene set variation analysis. Top 10 pathways significantly activated (red) or inhibited (blue) in the (A) GSE5281 or (B) GSE40562 dataset, based on fGSEA. (C,D) Heat map showing the Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analysis in Alzheimer’s disease (AD), sleep disorder (fatal familial insomnia), and controls.
We constructed a MEGENA network using the common genes to obtain module genes in AD and sleep disorder. We identified 29 modules and 1,498 module genes in GSE5281 (Figure 4A). The largest modules (C1_3 and C1_4) consisted of 135 genes, followed by module C1_19 with 125 genes. Moreover, we identified 55 modules and 1,791 module genes in GSE40562. Each of the three largest modules (C1_2, C1_4, and C1_5) had 135 genes (Figure 4B).
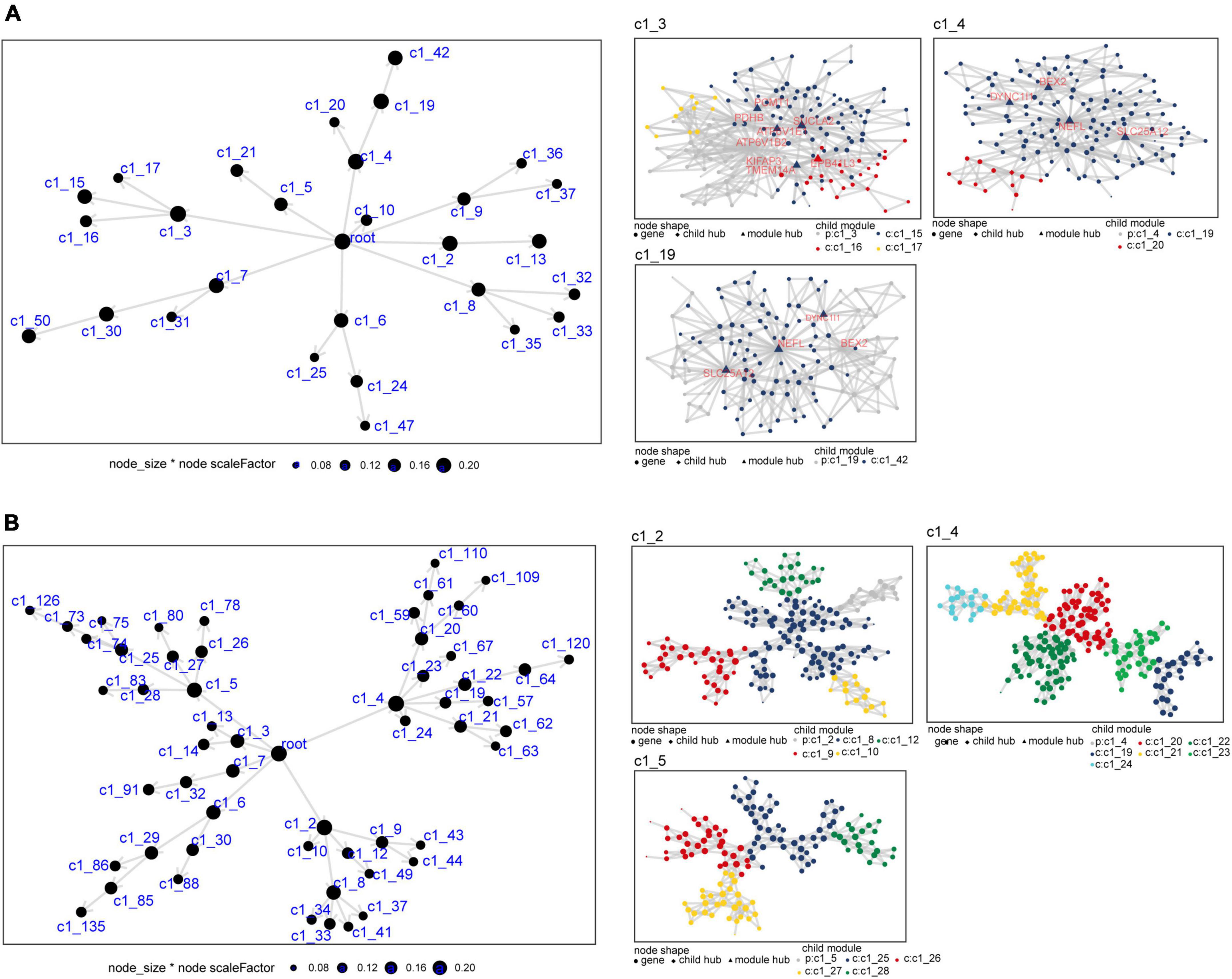
Figure 4. Multiscale embedded gene co-expression network analysis of common genes in Alzheimer’s disease (AD) and sleep disorder. (A) Differentially expressed genes in the co-expression network for the GSE5281 dataset. The three largest gene modules are shown: C1_3, C1_4, and C1_19. (B) Differentially expressed genes in the co-expression network for the GSE40562 dataset. The three largest gene modules are shown: C1_2, C1_4, and C1_5. Each node represents a module, with the larger nodes indicating a higher number of genes. Each color represents one module, and triangles represent key genes in the module.
To identify potential pathways involved in the mechanisms of sleep disorder and AD, genes of the identified 12 pathways that were also module genes were selected as genes that may promote the development of both AD and sleep disorder.
The potential role of gene-regulated pathways was analyzed by Cytoscape (Figure 5A). Similarly, Figure 5B showed that interactions among the genes common to AD and sleep disorder could influence the development of both disorders. Metascape enrichment analysis showed that common genes regulated proton transmembrane transport in sleep disorder and AD (Figure 5C). Heat maps showed the expression of 110 common genes in GSE5281 and GSE40562 datasets, of which four were upregulated and 106 downregulated (Figure 5D and Supplementary Figure 1). Furthermore, the top 10 genes with highest degree of connectivity (ATP5A1, ATP5B, COX5A, GAPDH, NDUFA9, NDUFS3, NDUFV2, SOD1, UQCRC1, and UQCRC2) were identified as hub genes in sleep disorder and AD. Interestingly, 10 hub genes were downregulated in patients with AD or sleep disorder relative to the corresponding controls (Figures 5E,F).
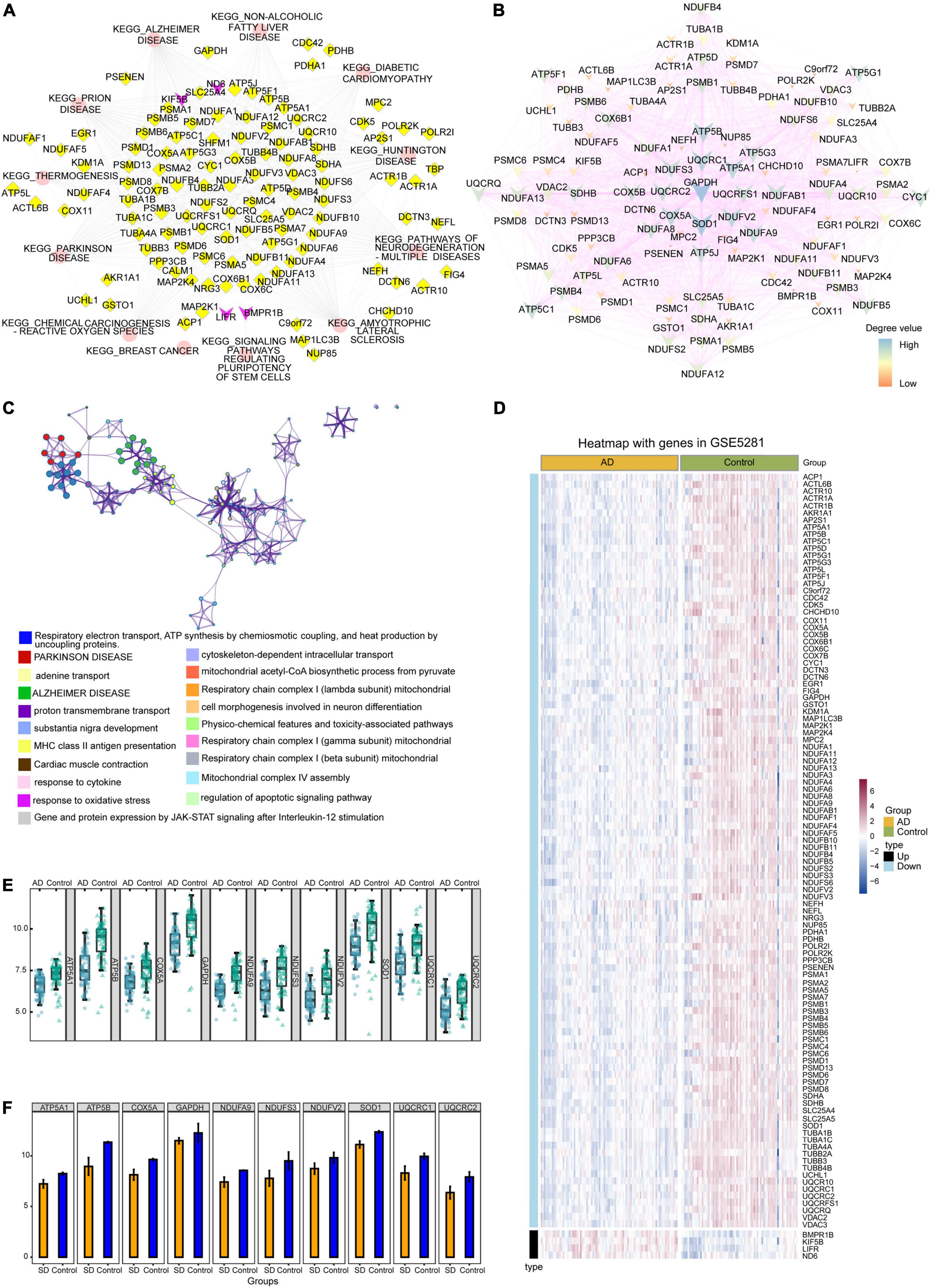
Figure 5. Protein-protein interaction (PPI) network and expression analysis. (A) Interaction of the 12 common pathways and common genes analyzed by Cytoscape. Red ellipses indicate pathways; yellow diamonds, downregulated genes; purple V-symbols, upregulated genes. (B) Interaction of common genes in the PPI network. Low-degree values are indicated by the small nodes and bright color, while high-degree values are indicated by larger nodes and dark color. (C) Functional enrichment analysis showed that common genes were involved mainly in Parkinson’s disease and Alzheimer’s disease (AD). (D) Heat map showing the expression of common genes in the GSE5281 dataset. (E) Box diagram showing degrees of connectivity of the top 10 genes in AD and controls. (F) Bar plot showing the degree values of the top 10 genes in sleep disorder and control group. SD, sleep disorder.
Infiltration of immune cells was compared between AD or sleep disorder patients and controls in both datasets. Plasmacytoid dendritic cells and T helper 17 (Th17) cells had the most extensive infiltration in both datasets (Figures 6A,B), according to ssGSEA. To further verify the correlation between hub genes and infiltrating immune cells, we performed a Pearson correlation analysis. We found a significant correlation between hub genes and infiltrating immune cells in the GSE5281 (Figure 6C) and GSE40562 datasets (Figure 6D). Moreover, in our CIBERSORT analysis, we found that CD8 + T cells constituted the highest proportion of infiltrated immune cells in GSE5281 (Figure 6E) and GSE40562 (Figure 6F), suggesting that these cells may play a role in the development of AD and sleep disorder.
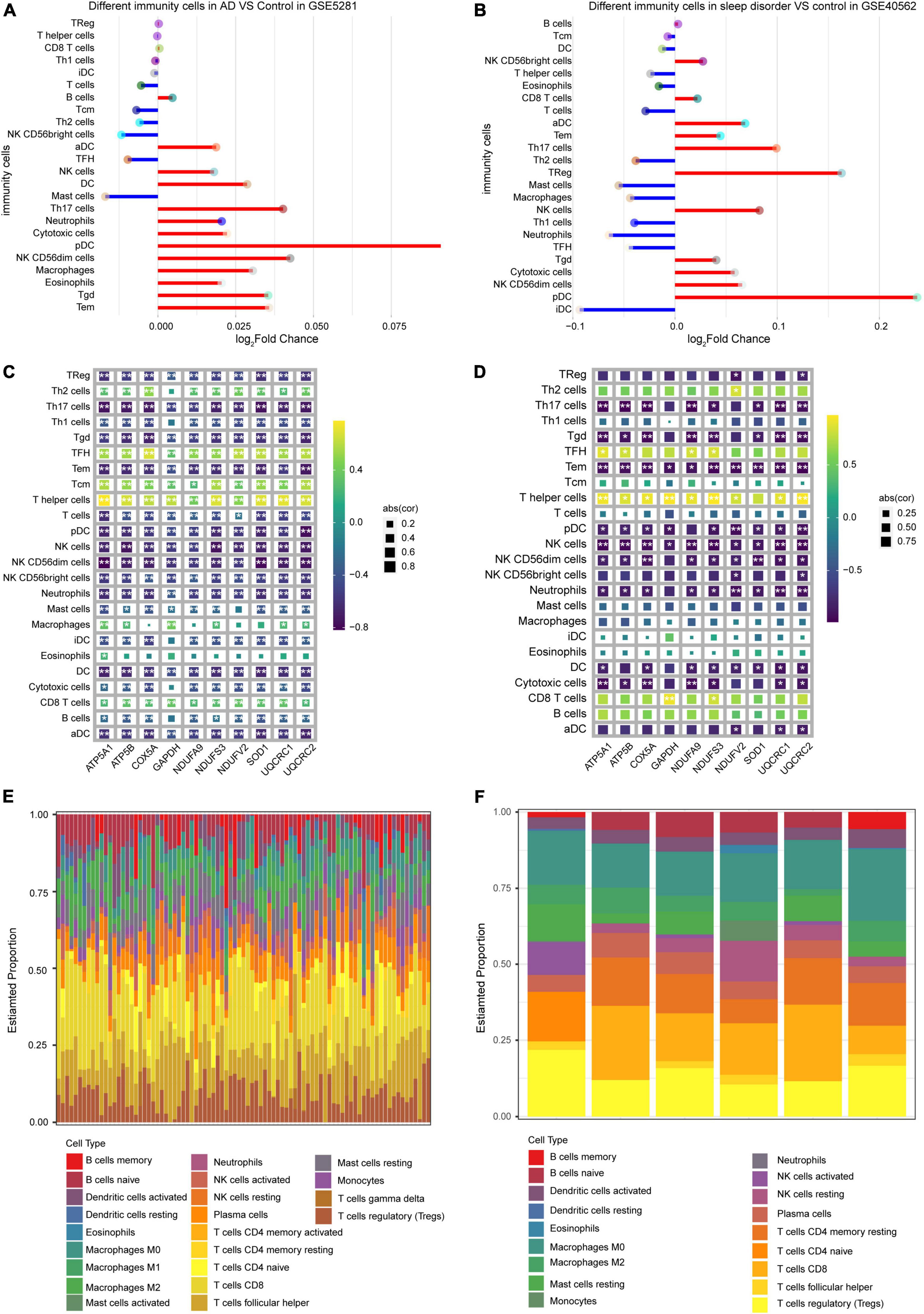
Figure 6. Level of immune cell infiltration in Alzheimer’s disease (AD) and sleep disorder. Comparison of immune cell infiltration (A) between AD patients and controls, and (B) between sleep disorder patients and controls, based on ssGSEA. Plots of correlations between hub genes and immune cell types in (C) AD and (D) sleep disorder. Proportions of 22 types of infiltrating immune cells in (E) AD and (F) sleep disorder, based on the CIBERSORT algorithm.
AD is frequently characterized by disturbed or absent sleep, similarly to sleep disorders. In addition, sleep disturbance is a risk factor for neurodegenerative diseases that are ultimately fatal. However, few studies have looked in detail at molecular pathways that may be common to both disorders. In our study, we integrated transcriptomic data from patients with either condition and controls and performed bioinformatics analyses to identify common pathogenic mechanisms.
Enrichment analysis showed that intersecting DEGs in GSE5281 and GSE40562 were involved in the HIF-1 signaling pathway and the TCA cycle, which produces energy for the cell. HIF appears to play a role in brain response to hypoxia and has been associated with the development of AD (36) and Parkinson’s disease (37). In neurodegenerative diseases, HIF signaling may be associated with impaired brain response to hypoxia. Furthermore, during the development and occurrence of AD and other neurodegenerative disorders, the TCA cycle in mitochondria may be altered. Enzymes and metabolites involved in these reactions suffer various changes during neurodegeneration (38). Patients with sleep disorder show abnormalities in metabolic pathways involving glucose and amino acids, and these pathways interact with the TCA cycle (39, 40).
The results of our GSEA showed that DEGs common to AD and sleep disorder were associated with activation of signaling pathways regulating pluripotency of stem cells and breast cancer, as well as associated with inhibition of Alzheimer’s disease, Parkinson’s disease, and other pathways. Pluripotent stem cells enable the acquisition of a large number of neural cells to improve cell recovery after neurodegenerative disorders, such as AD or Parkinson’s disease (41). Furthermore, women with breast cancer can also have cognitive problems (42). Interestingly, there is a possible connection between sleep disorder and breast cancer (43).
We identified several hub genes common to sleep disorder and AD: ATP5A1, ATP5B, COX5A, GAPDH, NDUFA9, NDUFS3, NDUFV2, SOD1, UQCRC1, and UQCRC2. ATP5A1 has been identified as the target of some components of Lavandula angustifolia extract, suggesting that some extract components may have therapeutic effects on AD and other neurodegenerative diseases (44). Downregulated ATP5B was also previously identified as a hub gene involved in AD (45). COX5A plays a vital role in aging-related memory impairment (46). Overexpression of glyceraldehyde-3-phosphate dehydrogenase in AD animal models increases apoptosis of hippocampal cells, neural degeneration, and cognitive dysfunction (47). NDUFA9, NDUFV2, and NDUFS3 are considered critical genes in oxidative phosphorylation and Parkinson’s disease, Huntington’s disease, and AD pathways (48). SOD1 (49) and UQCRC2 (50) have been linked to neurodegeneration. ATP5B (51), GAPDH (52), and SOD1 (53) were reported as sleep deprivation and sleep disorders related genes. One study showed an association among DNA methylation, UQCRC1 expression and risk of AD in a Chinese population (54). Notably, these genes may be potential markers or key genes in AD or other neurological diseases, but they have rarely been investigated in sleep disorders.
In our ssGSEA analysis, plasmacytoid dendritic cells and Th17 cells showed the most extensive infiltration in both AD and sleep disorder patients. Plasmacytoid dendritic cells in the central nervous system have been identified as the primary infiltrating cells in experimental autoimmune encephalomyelitis (55). Additionally, Th17 cells have been associated with cognitive impairment in multiple sclerosis and AD (56). Th17 cells strongly contribute to chronic neuroinflammation, thus perpetuating neurodegenerative processes (57). However, Th17 cells also have a role in barrier protection in many inflammatory diseases (58). Notably, when we applied the CIBERSORT algorithm, the abundance of CD8 + T cell infiltration was the highest in both AD and sleep disorder patients. A previous study found increased numbers of CD8 + T cells in the cerebrospinal fluid in AD patients (59). Differences in the results of the two immune infiltration analyses may be due to differences among individual patients.
This study presents several limitations. The main limitation is that our results are based purely on bioinformatics of small samples. Our analyses should be validated and extended in experimental studies. Furthermore, the infiltration of different immune cells may vary among patients, so we need to validate our results in large samples.
In the present study, ATP5A1, ATP5B, COX5A, GAPDH, NDUFA9, NDUFS3, NDUFV2, SOD1, UQCRC1, and UQCRC2 were identified as hub genes differentially expressed in AD and sleep disorder relative to healthy individuals. These genes may be involved in AD and sleep disorder through pathways related to neurodegeneration and multiple diseases, and they may be candidate biomarkers to facilitate diagnosis and therapy. We also identified some immune cell populations that strongly infiltrate tissues in both disorders and which therefore may contribute to pathogenesis.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary material.
The studies involving human participants were reviewed and approved by the First People’s Hospital of Nanning. Written informed consent for participation was not required for this study in accordance with the national legislation and the institutional requirements. Written informed consent was obtained from the individual(s) for the publication of any potentially identifiable images or data included in this article.
LL, JY, XH, DZ, and YL conceived and designed the study. All authors analyzed the data, prepared the figures and tables, wrote the manuscript, reviewed, the manuscript and approved its submission.
This study was supported by the National Natural Science Foundation of China (82060210), the Nanning Excellent Young Scientist Program and the Guangxi Beibu Gulf Economic Zone Major Talent Program (RC20190103), and the Guangxi Medical and Health Key Discipline Construction Project (Department of Geriatrics, The First People’s Hospital of Nanning).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpsyt.2022.925012/full#supplementary-material
Supplementary Figure 1 | Heat map showing the expression of common genes in the GSE40562 dataset.
2. Canter RG, Penney J, Tsai LH. The road to restoring neural circuits for the treatment of Alzheimer’s disease. Nature. (2016) 539:187–96.
5. Jian C, Zou D, Luo C, Liu X, Meng L, Huang J, et al. Cognitive deficits are ameliorated by reduction in amyloid beta accumulation in Tg2576/p75(NTR+/-) mice. Life Sci. (2016) 155:167–73. doi: 10.1016/j.lfs.2016.05.011
6. Brookmeyer R, Gray S, Kawas C. Projections of Alzheimer’s disease in the United States and the public health impact of delaying disease onset. Am J Public Health. (1998) 88:1337–42.
7. Scheltens P, De Strooper B, Kivipelto M, Holstege H, Chetelat G, Teunissen CE, et al. Alzheimer’s disease. Lancet. (2021) 397:1577–90.
8. Irwin MR, Vitiello MV. Implications of sleep disturbance and inflammation for Alzheimer’s disease dementia. Lancet Neurol. (2019) 18:296–306.
9. Bubu OM, Brannick M, Mortimer J, Umasabor-Bubu O, Sebastiao YV, Wen Y, et al. Sleep, cognitive impairment, and Alzheimer’s disease: a systematic review and meta-analysis. Sleep (2017) 40:zsw032. doi: 10.1093/sleep/zsw032
10. Bagyinszky E, Giau VV, Youn YC, An SSA, Kim S. Characterization of mutations in PRNP (prion) gene and their possible roles in neurodegenerative diseases. Neuropsychiatr Dis Treat. (2018) 14:2067–85.
11. Burchell JT, Panegyres PK. Prion diseases: immunotargets and therapy. Immunotargets Ther. (2016) 5:57–68. doi: 10.2147/ITT.S64795
12. Medori R, Montagna P, Tritschler HJ, LeBlanc A, Cortelli P, Tinuper P, et al. Fatal familial insomnia: a second kindred with mutation of prion protein gene at codon 178. Neurology. (1992) 42(3 Pt 1):669–70.
13. Monari L, Chen SG, Brown P, Parchi P, Petersen RB, Mikol J, et al. Fatal familial insomnia and familial creutzfeldt-jakob disease: different prion proteins determined by a DNA polymorphism. Proc Natl Acad Sci USA. (1994) 91:2839–42. doi: 10.1073/pnas.91.7.2839
16. Rubinsztein DC. The roles of intracellular protein-degradation pathways in neurodegeneration. Nature. (2006) 443:780–6.
18. Jian C, Wei L, Mo R, Li R, Liang L, Chen L, et al. Microglia mediate the occurrence and development of Alzheimer’s disease through ligand-receptor axis communication. Front Aging Neurosci. (2021) 13:731180. doi: 10.3389/fnagi.2021.731180
19. Zou D, Li R, Huang X, Chen G, Liu Y, Meng Y, et al. Identification of molecular correlations of RBM8A with autophagy in Alzheimer’s disease. Aging (Albany NY). (2019) 11:11673–85. doi: 10.18632/aging.102571
20. Ma M, Liao Y, Huang X, Zou C, Chen L, Liang L, et al. Identification of Alzheimer’s disease molecular subtypes based on parallel large-scale sequencing. Front Aging Neurosci. (2022) 14:770136. doi: 10.3389/fnagi.2022.770136
21. Luo J, Chen L, Huang X, Xie J, Zou C, Pan M, et al. REPS1 as a potential biomarker in Alzheimer’s disease and vascular dementia. Front Aging Neurosci. (2022) 14:894824. doi: 10.3389/fnagi.2022.894824
22. Jian C, Lu M, Zhang Z, Liu L, Li X, Huang F, et al. miR-34a knockout attenuates cognitive deficits in APP/PS1 mice through inhibition of the amyloidogenic processing of APP. Life Sci. (2017) 182:104–11. doi: 10.1016/j.lfs.2017.05.023
23. Zou C, Wang J, Huang X, Jian C, Zou D, Li X. Analysis of transcription factor- and ncRNA-mediated potential pathogenic gene modules in Alzheimer’s disease. Aging (Albany NY). (2019) 11:6109–19. doi: 10.18632/aging.102169
24. Tian C, Liu D, Xiang W, Kretzschmar HA, Sun QL, Gao C, et al. Analyses of the similarity and difference of global gene expression profiles in cortex regions of three neurodegenerative diseases: sporadic Creutzfeldt-Jakob disease (sCJD), fatal familial insomnia (FFI), and Alzheimer’s disease (AD). Mol Neurobiol. (2014) 50:473–81. doi: 10.1007/s12035-014-8758-x
25. Barrett T, Wilhite SE, Ledoux P, Evangelista C, Kim IF, Tomashevsky M, et al. NCBI GEO: archive for functional genomics data sets–update. Nucleic Acids Res. (2013) 41:D991–5. doi: 10.1093/nar/gks1193
26. Liang WS, Dunckley T, Beach TG, Grover A, Mastroeni D, Walker DG, et al. Gene expression profiles in anatomically and functionally distinct regions of the normal aged human brain. Physiol Genomics. (2007) 28:311–22.
27. Shi XH, Han J, Zhang J, Shi Q, Chen JM, Xia SL, et al. Clinical, histopathological and genetic studies in a family with fatal familial insomnia. Infect Genet Evol. (2010) 10:292–7. doi: 10.1016/j.meegid.2010.01.007
28. Ritchie ME, Phipson B, Wu D, Hu Y, Law CW, Shi W, et al. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. (2015) 43:e47. doi: 10.1093/nar/gkv007
29. Gao CH, Yu G, Cai P. ggVennDiagram: an intuitive, easy-to-use, and highly customizable R package to generate venn diagram. Front Genet. (2021) 12:706907. doi: 10.3389/fgene.2021.706907
30. Yu G, Wang LG, Han Y, He QY. ClusterProfiler: an R package for comparing biological themes among gene clusters. OMICS. (2012) 16:284–7. doi: 10.1089/omi.2011.0118
31. Sun W, Jia C, Zhang X, Wang Z, Li Y, Fang X. Identification of key genes related with aspartic acid metabolism and corresponding protein expression in human colon cancer with postoperative prognosis and the underlying molecular pathways prediction. Front Cell Dev Biol. (2022) 10:812271. doi: 10.3389/fcell.2022.812271
32. Shannon P, Markiel A, Ozier O, Baliga NS, Wang JT, Ramage D, et al. Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res. (2003) 13:2498–504. doi: 10.1101/gr.1239303
33. Song WM, Zhang B. Multiscale embedded gene co-expression network analysis. PLoS Comput Biol. (2015) 11:e1004574. doi: 10.1371/journal.pcbi.1004574
34. Tomkins JE, Manzoni C. Advances in protein-protein interaction network analysis for Parkinson’s disease. Neurobiol Dis. (2021) 155:105395.
35. Zhou Y, Zhou B, Pache L, Chang M, Khodabakhshi AH, Tanaseichuk O, et al. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat Commun. (2019) 10:1523. doi: 10.1038/s41467-019-09234-6
36. Zhou F, Chen D, Chen G, Liao P, Li R, Nong Q, et al. Gene set index based on different modules may help differentiate the mechanisms of Alzheimer’s disease and vascular dementia. Clin Interv Aging. (2021) 16:451–63. doi: 10.2147/CIA.S297483
37. Correia SC, Carvalho C, Cardoso S, Santos RX, Placido AI, Candeias E, et al. Defective HIF signaling pathway and brain response to hypoxia in neurodegenerative diseases: not an “iffy” question! Curr Pharm Des. (2013) 19:6809–22. doi: 10.2174/1381612811319380013
38. Yan X, Hu Y, Wang B, Wang S, Zhang X. Metabolic dysregulation contributes to the progression of Alzheimer’s disease. Front Neurosci. (2020) 14:530219. doi: 10.3389/fnins.2020.530219
39. Benedini S, Cortelli P, Caumo A, Terruzzi I, Beelke M, Perseghin G, et al. Insulin resistance to both glucose and aminoacid metabolism in a patient with fatal familial insomnia. Nutr Metab Cardiovasc Dis. (2008) 18:e47–8. doi: 10.1016/j.numecd.2008.08.003
40. Hui S, Ghergurovich JM, Morscher RJ, Jang C, Teng X, Lu W, et al. Glucose feeds the TCA cycle via circulating lactate. Nature. (2017) 551:115–8.
41. Kolagar TA, Farzaneh M, Nikkar N, Khoshnam SE. Human pluripotent stem cells in neurodegenerative diseases: potentials, advances and limitations. Curr Stem Cell Res Ther. (2020) 15:102–10.
42. Lehrer S, Rheinstein PH. Breast cancer, Alzheimer’s disease, and APOE4 Allele in the UK biobank cohort. J Alzheimers Dis Rep. (2021) 5:49–53. doi: 10.3233/ADR-200266
43. Shorofi SA, Nozari-Mirarkolaei F, Arbon P, Bagheri-Nesamie M. Depression and sleep quality among iranian women with breast cancer. Asian Pac J Cancer Prev. (2021) 22:3433–40. doi: 10.31557/APJCP.2021.22.11.3433
44. Zali H, Zamanian-Azodi M, Rezaei Tavirani M, Akbar-Zadeh Baghban A. Protein Drug Targets of Lavandula angustifolia on treatment of rat Alzheimer’s disease. Iran J Pharm Res. (2015) 14:291–302.
45. Liu L, Wu Q, Zhong W, Chen Y, Zhang W, Ren H, et al. Microarray analysis of differential gene expression in Alzheimer’s disease identifies potential biomarkers with diagnostic value. Med Sci Monit. (2020) 26:e919249.
46. Xiyang YB, Liu R, Wang XY, Li S, Zhao Y, Lu BT, et al. COX5A plays a vital role in memory impairment associated with brain Aging via the BDNF/ERK1/2 signaling pathway. Front Aging Neurosci. (2020) 12:215. doi: 10.3389/fnagi.2020.00215
47. Lazarev VF, Tsolaki M, Mikhaylova ER, Benken KA, Shevtsov MA, Nikotina AD, et al. Extracellular GAPDH promotes Alzheimer disease progression by enhancing amyloid-beta aggregation and cytotoxicity. Aging Dis. (2021) 12:1223–37. doi: 10.14336/AD.2020.1230
48. Zhang L, Guo XQ, Chu JF, Zhang X, Yan ZR, Li YZ. Potential hippocampal genes and pathways involved in Alzheimer’s disease: a bioinformatic analysis. Genet Mol Res. (2015) 14:7218–32.
49. Bader JM, Geyer PE, Muller JB, Strauss MT, Koch M, Leypoldt F, et al. Proteome profiling in cerebrospinal fluid reveals novel biomarkers of Alzheimer’s disease. Mol Syst Biol. (2020) 16:e9356.
50. Nakamura T, Watanabe A, Fujino T, Hosono T, Michikawa M. Apolipoprotein E4 (1-272) fragment is associated with mitochondrial proteins and affects mitochondrial function in neuronal cells. Mol Neurodegener. (2009) 4:35. doi: 10.1186/1750-1326-4-35
51. Nikonova EV, Naidoo N, Zhang L, Romer M, Cater JR, Scharf MT, et al. Changes in components of energy regulation in mouse cortex with increases in wakefulness. Sleep. (2010) 33:889–900.
52. Lee KS, Alvarenga TA, Guindalini C, Andersen ML, Castro RM, Tufik S. Validation of commonly used reference genes for sleep-related gene expression studies. BMC Mol Biol. (2009) 10:45. doi: 10.1186/1471-2199-10-45
53. Chen J, Gu H, Wurster RD, Cheng ZJ. The protective role of SOD1 overexpression in central mediation of bradycardia following chronic intermittent hypoxia in mice. Am J Physiol Regul Integr Comp Physiol. (2021) 320:R317–30. doi: 10.1152/ajpregu.00147.2020
54. Ma SL, Tang NL, Lam LC. Association of gene expression and methylation of UQCRC1 to the predisposition of Alzheimer’s disease in a Chinese population. J Psychiatr Res. (2016) 76:143–7. doi: 10.1016/j.jpsychires.2016.02.010
55. Bailey-Bucktrout SL, Caulkins SC, Goings G, Fischer JA, Dzionek A, Miller SD. Cutting edge: central nervous system plasmacytoid dendritic cells regulate the severity of relapsing experimental autoimmune encephalomyelitis. J Immunol. (2008) 180:6457–61. doi: 10.4049/jimmunol.180.10.6457
56. Cipollini V, Anrather J, Orzi F, Iadecola C. Th17 and cognitive impairment: possible mechanisms of action. Front Neuroanat. (2019) 13:95. doi: 10.3389/fnana.2019.00095
57. Gonzalez H, Pacheco R. T-cell-mediated regulation of neuroinflammation involved in neurodegenerative diseases. J Neuroinflammation. (2014) 11:201.
58. Stockinger B, Omenetti S. The dichotomous nature of T helper 17 cells. Nat Rev Immunol. (2017) 17:535–44.
Keywords: Alzheimer’s disease, sleep disorder, gene expression, bioinformatics analysis, differentially expressed gene (DEG)
Citation: Liang L, Yan J, Huang X, Zou C, Chen L, Li R, Xie J, Pan M, Zou D and Liu Y (2022) Identification of molecular signatures associated with sleep disorder and Alzheimer’s disease. Front. Psychiatry 13:925012. doi: 10.3389/fpsyt.2022.925012
Received: 21 April 2022; Accepted: 20 July 2022;
Published: 04 August 2022.
Edited by:
Francesco Panza, University of Bari Aldo Moro, ItalyReviewed by:
Xuan Ye, Novartis Institutes for BioMedical Research, United StatesCopyright © 2022 Liang, Yan, Huang, Zou, Chen, Li, Xie, Pan, Zou and Liu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Donghua Zou, em91ZG9uZ2h1YUBneG11LmVkdS5jbg==; Ying Liu, bHlseTAwMDAwMDFAMTYzLmNvbQ==
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.