 Meng Li†
Meng Li† Dongdong Qin
Dongdong Qin
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Psychiatry, 25 January 2022
Sec. Psychopathology
Volume 12 - 2021 | https://doi.org/10.3389/fpsyt.2021.827541
This article is part of the Research TopicNeural Circuits and Neuroendocrine Mechanisms of Major Depressive Disorder and Premenstrual Dysphoric Disorder: Towards Precise Targets for Translational Medicine and Drug DevelopmentView all 10 articles
The incidence rates of depression are increasing year by year. As one of the main clinical manifestations of depression, sleep disorder is often the first complication. This complication may increase the severity of depression and lead to poor prognosis in patients. In the past decades, there have been many methods used to evaluate sleep disorders, such as polysomnography and electroencephalogram, actigraphy, and videography. A large number of rodents and non-human primate models have reproduced the symptoms of depression, which also show sleep disorders. The purpose of this review is to examine and discuss the relationship between sleep disorders and depression. To this end, we evaluated the prevalence, clinical features, phenotypic analysis, and pathophysiological brain mechanisms of depression-related sleep disturbances. We also emphasized the current situation, significance, and insights from animal models of depression, which would provide a better understanding for the pathophysiological mechanisms between sleep disturbance and depression.
Sleep is an essential physiological requirement for human and most animals. A mechanistic link is evident between sleep and depression at the molecular and neurophysiological level. The periodic regulation of awake and sleep requires the participation of many neurotransmitters, including excitatory neurotransmitters (such as acetylcholine) and inhibitory neurotransmitters (such as gamma aminobutyric acid, GABA). Abnormalities of these neurotransmitter systems not only lead to sleep-wake rhythm disorders, but also can contribute to developing depression. Depression and sleep disturbances are common co-morbid conditions (1, 2). More than 90% percent of patients with major depressive disorder will suffer from sleep disorders, which changed the patients' sleep structure. A further demonstration of the link between depression and sleep is that sleep can be improved by most clinically effective antidepressant drugs. Compared with lower mammals, the sleep of non-human primates (NHPs) is better comparable with that of humans. Recently, significant progress has been made in the study of using NHPs to establish depression models. Monitoring the sleep status of animals during modeling will help us further understand the role of sleep in the development of depression, and provide an objective biomarker for the early diagnosis, treatment, and efficacy evaluation.
Sleep is vital for human beings and most animals, and control mechanisms are embodied in all levels of biological organizations, from genes and intracellular mechanisms to cell population networks, and then to all central nervous systems, including systems that control movement, arousal, autonomic function, behavior, and cognition. Mammalian sleep is characterized by the periodic alternation of rapid eye movement sleep (REMs) and non-rapid eye movement sleep (NREMs). NREMs includes two stages: slow-wave sleep (SWS) and light sleep. In humans, SWS and REMs, which are the specific modes of potential electric field oscillations and neuromodulator activities, dominate the first half of the night and the latter half of the night, respectively (3).
The mutual transformation between sleep and wakefulness is caused by the excitation or inhibition of many neurotransmitters in the brain, which are released by sleep-promoting neurons in the anterior hypothalamus or sleep-inhibiting neurons in the lateral and posterior hypothalamus activity. These neurons release excitatory or inhibitory neurotransmitters to promote the brainstem to control the mutual transformation of wakefulness and sleep (2, 4).
The ascend arousal system mainly comes from a group of explicit cells with definite neurotransmitters. The arousal system actually consists of two channels (5). The ascending pathway to the thalamus is the first branch, which activates the thalamus and is essential to relay neurons for transmitting information to the cerebral cortex. The main sources of input from the upper brainstem to the thalamic-relay nucleus, the thalamic reticular nucleus, the pedunculopontine, and laterodorsally tegmental nucleus (PPT/LDT) are a couple of acetylcholine producing cell populations. The neurons in PPT/LDT discharge fastest during awake and REMs, and are often accompanied by cortical activation, loss of body muscle tone and active dreams. During NREMs, the activity of these cells is much lower. They are important for the input of reticular nucleus, because they are located between thalamic relay nucleus and cerebral cortex. It is very important for arousal that they can block the transmission between thalamic and cerebral cortex, thus acting as a gating mechanism. From the reticular structure and PPT/LDT, monoamine nervous system and parabrachial nucleus in the upper part of the brain stem, have more extensive input to the midline of thalamus and tabular nucleus. The laminar nucleus and midline nucleus are also considered to play a role in cortical arousal (5). Bypassing the thalamus and activating the neuronal pathway of the lateral hypothalamic area, basal forebrain (BF), and the whole cerebral cortex is the second branch of the ascending arousal system. This pathway, which covers noradrenergic locus coeruleus, serotonin dorsal nucleus, and median raphe nucleus, dopaminergic midbrain periaqueductal gray matter ventral and histaminergic nodule papillary neurons, is derived from monoamine neurons in the upper brainstem and caudal hypothalamus. Cortical input is increased by hypothalamic lateral peptidergic neurons (containing melanin concentrating hormone or orexin/retinol) and BF neurons (containing acetylcholine or GABA) (5). Lesions along this path, especially in the left hemisphere and the rostral midbrain, produce the most profound and lasting drowsiness and even coma. The neurons in each monoaminergic nucleus involved in this pathway discharge fastest during waking, slow down during NREMs and completely stop during REMs. It should be noted that all these ascending pathways pass through the regions at the junction of forebrain and brainstem. While, the descending pathways responsible for synchronizing phenomena still remain largely unknown at the brain-stem level.
The pathogenesis of sleep disorder is closely related to sleep-wake homeostasis, but the specific mechanism remains still unclear. During NREMs and REMs, different kinds of neurotransmitters are released in the brain. The interaction between aminergic neurons and cholinergic neurons at the meso-pontine junction leads each other to bring about the Ultradian rhythms alternation of REMs and NREMs. During NREMs, aminergic inhibition is decreased and cholinergic excitation is increased. At the onset of REMs, aminergic inhibition is turned off, cholinergic excitability reaches a peak, and other outputs are inhibited (2). When awake, the pontine aminergic system is tensely activated and the pontine cholinergic system is inhibited. In addition to aminergic and cholinergic neurons, other neurotransmitter systems are also involved in modulating REMs/NREMs alternation and may interact with aminergic and cholinergic systems (2, 6, 7). Extrinsically augmented dopaminergic neurotransmission can influence both REMs and NREMs cycles. Moreover, gamma-amino butyric acid and glutamate also affect the REMs/NREMs cycle (2).
In short, the growth and decline of these neurotransmitters promote the mutual transformation between sleep and wake. If these related neurotransmitters are released abnormally, it will cause sleep problems, such as difficulties in falling asleep and maintaining sleep state, changes of REMs latency, abnormal REMs behavior, and disturbed alternating pattern of REMs/NREMs.
Depression is the main cause of the burden of mental health-related diseases in the world, and about 300 million people around the world are affected by depression (8). One aspect of efforts to understand depression focuses on its relationship with sleep. In many cases, the onset of depression is announced through sleep disorders, and sleep deterioration occurs before depression and manic episodes (9). There are many forms of sleep disorders reported in patients with depression. It may be only exhibited by the shortening of sleep time, but it also indicates a reduction in sleep efficiency. The latter is defined as the ratio of total sleep time to total time spent in bed over the night. Lack of sleep increases the risk of depressive episodes and depression relapses. Likewise, depression increases the risk of sleep disorders. However, the self-assessment of sleep quality in patients with depression is unreliable. Similarly, there are differences in the subjective and objective assessment of daytime alertness (10). This leads to bias in the evaluation of sleep efficiency.
Epidemiological investigations confirm that there is a closer relationship between insomnia and the onset of depression. It is reported that most patients often have insomnia and depressive episodes at the same time (11). Approximately 90% of major depressive disorder (MDD) patients have been found to suffer from sleep disorders, including initial insomnia, difficulty in sleep maintenance, non-restorative sleep, and early morning awakenings (12, 13). In reality, the most common subjective sleep complaints reported by depressed patients are insomnia (up to 88%) and hypersomnia (27%) (14). The insomnia and emotional symptoms are bidirectional correlated that poor sleep may precede the onset of depression, and depressive mood may also disrupt sleep patterns. In addition, patients with MDD are three times more likely to suffer from insomnia than those without (15, 16). Furthermore, fatigue, hypersomnia, and sleepiness are closely related to depressive symptoms (14). Many depressed patients complain about non-recovery sleep and excessive daytime sleepiness (16), and about 15% of patients report symptoms of daytime sedation and hypersomnia (17). However, these findings are inconsistent (16). Depression and hypersomnia are two conditions linked in a complex and bidirectional manner. In addition, many patients with depression call their complaints a combination of daytime sleepiness and nighttime anxiety.
Since the 1960s, polysomnography (PSG) sleep studies have repeatedly shown that depression is also associated with disrupted sleep architecture. These abnormalities include increase in RA (REMs activity) and RD (REMs density), as well as a decrease in REMs latency and SWS (18). During REMs, patients with depression often show short latency, prolonged cycle, and increased density (19). Disorders of REMs usually persist throughout the clinical episode, and it is considered to increase the possibility of recurrence, and may reduce the therapeutic effect (19–21). After antidepressant treatment, the number of REMs is decreased and the latency of REMs is increased. Most antidepressants inhibit REMs in patients and healthy volunteers (22).
The increase of serotonin content may be the main reason affecting REMs (23). Antidepressants that increased the contents of serotonin (5-HT) in synapses are effective inhibitors of REMs. 5HT1A agonists can be used as antidepressants and can significantly inhibit REMs (24). However, tryptophan depletion leads to a decrease in serotonin, which has been shown to reverse REMs inhibition caused by antidepressants (25). In addition, trazodone and nefazodone are also used as antidepressants because they have a strong antagonistic effect on serotonergic 5-HT2 receptors, which often promotes sleep and improves sleep continuity (26). The percentage of REMs was most significantly decreased in the early stage of treatment. Additionally, a subsequent study evaluated the changes in sleep structure of 20 patients with unipolar MDD after administration of sustained-release bupropion, and the results showed that 8 weeks of bupropion treatment significantly prolonged REMs latency, increased REMs activity and density in the first REMs period, which led to increased total REM density (27).
Glutamatergic and GABAergic neurons also play a role in the generation of REMs (28). Ketamine is a rapid-acting antidepressant (29), and AMPA-mediated increased neurotransmission is the basis of the antidepressant-like behavioral effects of ketamine (30, 31). The enhancement of AMPA receptor signal is participated in the pathophysiology and the mediation of ketamine-induced rapid antidepressant treatment (32, 33). Importantly, increased levels of ionic AMPA receptor could promote net synaptic strength and induce prolonged waking time in rodents and humans (34).
The REMs density of patients with depression continues to increase, which is regarded as an endophenotype. The reduction of the initial latency and the delta sleep ratio (DSR, the ratio of SWS between the first two NREMs episodes) of the rapid eye movement can be explained by cholinergic-aminergic imbalance (35). The monoaminergic inhibition of PPT/LDT cholinergic cells in patients with depression is weakened and/or the cholinergic-driven effect in pontine reticular formation is enhanced, resulting in an increase in REMs tendency and intensity.
The initiation and maintenance of NREMs also seem to be dependent on the role of monoamine neurotransmitters (26). Sedative antidepressants enhance SWS and prolong sleep duration. For instance, selective serotonin reuptake inhibitors (SSRIs) and non-sedating tricyclic antidepressants (TCA) can result in lighter sleep. In patients with depression, SWS and DSR tends to be low (36, 37). Compared with REMs latency, the measurement of SWS and DSR distribution may be a more reliable predictor of clinical response of antidepressant treatment and recurrence of depressive symptoms. Higher DSR may be more conducive to the treatment of depression (38). Some lines of evidence suggest that ketamine administration significantly increased the intensity of both SWS and DSR in humans and rats (39–41).
In addition, other types of antidepressants can also improve sleep. For example, antidepressants with anti-histaminergic action, such as mirtazapine and ipsapirone, act on their own receptors to support homeostatic maintenance of monoamine levels, block specifically monoamine receptors to enhance serotoninergic neurotransmission. Some patients' sleep can become better even after the first treatment of mirtazapine (42). However, increased levels of noradrenergic and dopaminergic neurotransmission, and raised activation of serotonergic 5-HT2 receptors can worsen the quality of sleep, which are also adverse effects of several antidepressants, such as serotonin and norepinephrine reuptake inhibitors, norepinephrine reuptake inhibitors, monoamine oxidase inhibitors (MAOI), SSRIs, and activated TCA (43). During REMs, monoaminergic neurons reduced significantly their discharge rate or stop their activity, but cholinergic neurons become highly active (44). However, MAOI increases the amounts of monoamine by preventing enzyme degradation and tends to cause the absence of REMs. One possible explanation is the antagonism of three receptors, namely H1 histamine or cholinergic receptor and postsynaptic 5HT2C receptor (26, 45). Therefore, most antidepressants alleviate depressed symptoms by improving sleep quality.
It is necessary to obtain the best animal model for studying disease in biomedical research. Validity of animal models depends on the extent to which how they can mimic human diseases. Researchers have made exogenous and endogenous animal models to simulate the symptoms of depressed patients and elucidate the mechanisms of antidepressant action, involving acute and chronic stress model, secondary depression model, and genetic model (46). Translation validity of animal models is the key to sleep disorders research. As shown in Table 1, zebrafish, mice, rats, cats, dogs, and monkeys are generally useful to develop animal models to study sleep disorders (49, 62, 98–101). Among them, the most used laboratory animals are mice and rats. However, they are quite different from humans as they are nocturnal and adopt a monophasic sleep schedule. While, humans follow a polyphasic sleep pattern and are very flexible in choosing the sleep time (80, 102). Similar to humans, more fragmented and polyphasic sleep patterns are observed in monkeys, and they are generally active during the day and sleep at night (84). In view of this, compared with other animals, the sleep pattern of monkeys is closer to that of humans.
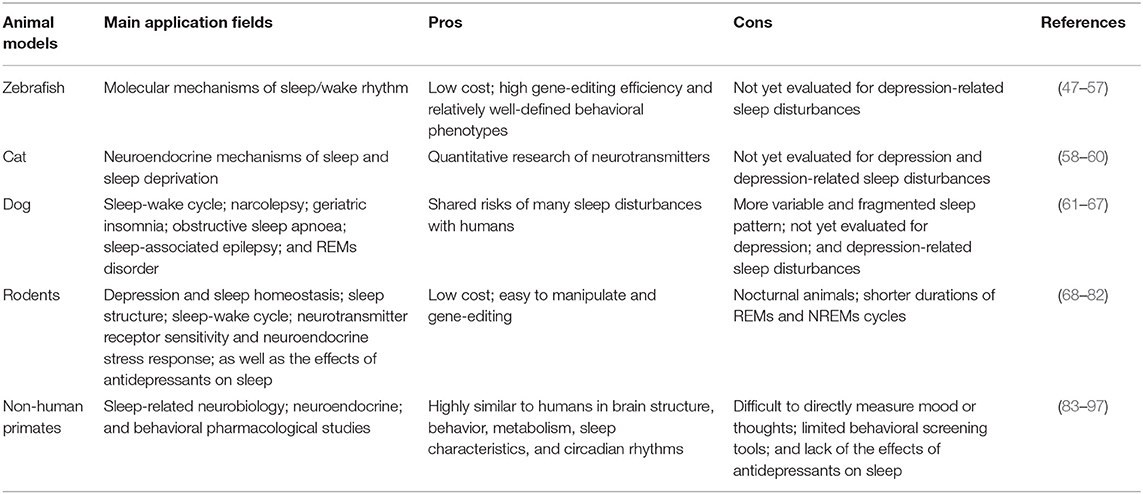
Table 1. Comparison of different animal models used to study sleep disturbances and depression.
Rodents are more usual choice of preclinical models to develop new pharmacological and non-pharmacological strategies. In the study of sleep deprivation, rodents (i.e., rats, mice) and humans have many similarities in sleep electroencephalogram (EEG) and sleep structure (103). External stressors or risk factors of diseases can affect the number or pattern of REMs (22, 104, 105). In humans, REMs latency is negatively correlated with the severity of depression (37). In rodents, changes in the REMs can precede those of other sleep/wake stages. For example, mice that were applied to water immersion for 2 h and restraint stress exhibited an immediate reduction in REMs (106).
As for the effect of stress on rodents' total sleep time, the primary stressors are immobilization and mild electrical shock. Immobilization increased the time spent in SWS and REMs, while electrical shock resulted in a decrease in total sleep time and total REMs time (107). Similarly, fear conditioning paradigms can also induce a decrease in REMs during both the shock training and cue exposure (104). Chronic unpredictable mild stress can lead to changes in the amplitude of both physiological (i.e., locomotion, temperature) and molecular circadian rhythm, which may cause depressive-like behaviors (108).
Continuous light exposure (LL) increases depressive-like behavior in mice, and light exposure at night (LAN) can lead to depressive-like behavior in diurnal rodents, such as grass rats and hamsters (109–111). This may be because LL brings about the interrupted rhythm of locomotion, temperature, and hormonal release, causing the disruption of circadian rhythm, and increases of NREMs during the rest period and REMs in the active period (112, 113).
For social species such as rats and mice, repeated fighting and/or defeat may be a more natural source of stress. Often, the consequences of chronic social defeat stress (CSDS) can persist until the termination of the stressors, which makes it a particularly attractive method to model stress-related psychiatric illnesses (114). Previous studies have found that CSDS has a direct effect on subsequent sleeping. Specifically, it can increase both the total time of REMs and NREMs, as well as the density of NREMs. However, the number of REMs is significantly decreased in the first few hours after conflict (114, 115). Another experiment also reported a brief increase in REMs time following 10 days of social conflict, but no changes in SWS were detected (115, 116). Interestingly, there was no difference in NREMs and slow-wave activity between winner and loser, suggesting it is a consequence induced by the conflict process.
NHPs bridge the gap between rodents and humans (117). Like humans, NHPs have stable sleep at night and some nap during the day. Many kinds of non-human primates, such as baboon, Kenya baboon, South African ape, macaque, cynomolgus monkey, Pada monkey, lemur, and chimpanzee, can be used to study sleep. By comparing the sleep of non-human primates, researchers generally believe that chimpanzee, olive baboon, and rhesus monkey are better model animals. In monkeys, four sleep EEG patterns can be easily identified. Due to its stable and perfect sleep architecture, macaque has become the best model to study the biological characteristics of human sleep (118–121). During the whole night, macaques experienced the alternation of awake, NREMs and REMs, and the total sleep time of rhesus monkey is about 10.5 h per day. It has been found that REMs time accounts for 23%, each of which lasts about 6 min and occurs every 51 min. In the early stage, it is mainly deep sleep, such as SWS. While, it is mainly REMs in the late stage of sleep. These sleep characteristics are very similar to humans that the interval of this cycle is about 90 min. Nevertheless, in rats, the interval is only 13 min. Like humans, obvious theta waves cannot be recorded in the hippocampus during macaques' sleep (122, 123).
EEG is a common method in sleep research, which can provide objective functional indexes for sleep (124). Although EEG can be performed in constrained animals under laboratory conditions, this technique is invasive. Even if it is minimally invasive, it also needs to drill holes in the skull and implant electrodes directly on the brain. PSG plays a cornerstone role in long-term recording of sleep, and has become the gold standard to evaluate sleep disorders. The recorded parameters include the brain activity (EEG), electrooculogram (EOG), expanded EEG montages, and transcutaneous or end-tidal capnography waveform, which are used to comprehensively monitor the normal and abnormal physiological indicators during sleep (125). However, an important limitation of PSG is that it requires electrodes and sensors (126). In addition, expensive and long-term recording intervals may be another limitation. Obviously, these are difficult and impossible to use in freely moving monkeys. A recent study compared videography and actigraphy methods in 10 cynomolgus monkeys during seven nights. It is verified that in the sleep study of NHPs, actigraphy can be regarded as a supplementary technique for routine EEG and/or video analysis to measure the sleep (127).
Researchers have used NHPs to make great efforts in the research of depression. It has been demonstrated for the first time that long-term intracerebroventricular administration of IFN-α (5 days/week for 6 weeks) can induce the monkeys showing considerable depressive-like symptoms with changes in the concentration of monoamine metabolites (128). The relationship between early adversity, chronic stress and depression was also investigated in adolescent monkeys. Eight male rhesus monkeys went through unpredictable chronic stress for 2 months and exhibited significant depression-like behaviors (88). The mechanisms underlying stress-induced depression were also explored in monkeys, and it was found that cortisol hypersecretion interacted with stress to accelerate the development of depressive behaviors (129).
In addition, researchers have employed NHPs animal model to make many beneficial explorations on the association between light deprivation and depression. The results showed that monkeys could develop the main symptoms of seasonal affective disorder under short lighting conditions (130). Analogous to depression in humans, sleep disorders have been also reported in spontaneous depressed monkeys (86). Notably, only the hypersomnia subgroup of spontaneously depressed monkeys shows a specific response to acute ketamine administration, characterized as extended wakefulness and shortening of nocturnal sleep. As a matter of fact, these changes are similar to sleep deprivation in depressed patients, suggesting alternation of nocturnal sleep pattern might help improve depressed mood (86, 119, 131).
There is increasing evidence that sleep plays a causal role in emotional processing and regulation (132). Depression and sleep disturbances are common co-morbid conditions, and almost all depressed patients show some types of sleep disturbances (133, 134). Most antidepressants can change sleep, and the effects appear to be most significant and consistent on REMs (135). Selective REMs deprivation (such as forced awakenings) can produce an antidepressant effect, illustrating the closer association between REMs regulation and mechanisms involved in the development of depression (136). Some neurotransmitter reuptake inhibitors can alleviate depression by suppressing REMs through inhibition of serotonin and norepinephrine reuptake (26). However, many questions remain to be answered in future studies. Firstly, in previous studies, it was found that the effects of antidepressants on sleep initiation and maintenance were inconsistent. Secondly, the mechanism of different effects of antidepressants on sleep continuity is unclear. In rodent experiments, many paradigms of chronic stress have been used to simulate the pathogenesis of human depression, but it is hard to provide a unified description about the impact of chronic stress on sleep patterns. In fact, in addition to the types of stress, the number and persistent time are also important factors for stress responses, which must be carefully considered. NHPs are suitable animal models for experiments related to sleep, however, the study of depression and sleep disorders is far from enough. Although researchers have made continuous efforts and good progress in relevant animal models, it must be recognized that there are deficiencies.
In any way whatever, the research on animal models of sleep disorders provides a good clue and basis for clinical diagnosis and treatment of depression. NHPs are considered as a further valuable and translational animal model, which is necessary for sleep and related diseases (137, 138). It is also an important entry point for increased efforts dedicated to collaborative translational endeavors.
DQ, LX, XL, and ML designed the structure. ML, JC, BX, YW, CF, and DQ wrote the first draft of the manuscript. DQ, LX, and XL supervised and revised the final version of the manuscript. All authors contributed substantially to the scientific process, writing of the manuscript, and have approved the final version of the manuscript being submitted.
This work was supported by the National Natural Science Foundation of China (31960178, 82074421, and 82160923), the Applied Basic Research Programs of Science and Technology Commission Foundation of Yunnan Province (2019FA007), the Joint Project of Applied Basic Research of Yunnan University of Chinese Medicine & Yunnan Provincial Science and Technology Department [2019FF002(-001)], Yunnan Provincial Department of Education Science Research Fund Project (2020Y0203), the Key Realm R&D Program of Guangdong Province (2019B030335001), the China Postdoctoral Science Foundation (2018M631105), and the Yunnan Provincial Academician and Expert Workstation (202005AF150017 and 2019IC051).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Noguti J, Andersen ML, Cirelli C, Ribeiro DA. Oxidative stress, cancer, and sleep deprivation: is there a logical link in this association? Sleep Breath. (2013) 17:905–10. doi: 10.1007/s11325-012-0797-9
2. Pace-Schott EF, Hobson JA. The neurobiology of sleep: genetics, cellular physiology and subcortical networks. Nat Rev Neurosci. (2002) 3:591–605. doi: 10.1038/nrn895
3. Diekelmann S, Born J. The memory function of sleep. Nat Rev Neurosci. (2010) 11:114–26. doi: 10.1038/nrn2762
4. Jouvet M. Neurophysiology of the states of sleep. Physiol Rev. (1967) 47:117–77. doi: 10.1152/physrev.1967.47.2.117
5. Saper CB, Scammell TE, Lu J. Hypothalamic regulation of sleep and circadian rhythms. Nature. (2005) 437:1257–63. doi: 10.1038/nature04284
6. Lydic R, Baghdoyan HA. Handjournal of Behavioral State Control: Cellular and Molecular Mechanisms. Boca Raton, Florida: CRC Press (2019). doi: 10.1201/9780429114373
7. Sakai K, Crochet S, Onoe H. Pontine structures and mechanisms involved in the generation of paradoxical (REM) sleep. Arch Ital Biol. (2001) 139:93–107. doi: 10.4449/aib.v139i1.207
8. Herrman H, Kieling C, McGorry P, Horton R, Sargent J, Patel V. Reducing the global burden of depression: a Lancet-World Psychiatric Association Commission. Lancet. (2019) 393:e42–3. doi: 10.1016/S0140-6736(18)32408-5
9. Van Moffaert MM. Sleep disorders and depression: the 'chicken and egg' situation. J Psychosom Res. (1994) 38(Suppl 1):9–13; discussion 12–3. doi: 10.1016/0022-3999(94)90131-7
10. Matousek M, Cervena K, Zavesicka L, Brunovsky M. Subjective and objective evaluation of alertness and sleep quality in depressed patients. BMC Psychiatry. (2004) 4:14. doi: 10.1186/1471-244X-4-14
11. Ohayon MM, Caulet M, Lemoine P. Comorbidity of mental and insomnia disorders in the general population. Comprehensive Psychiatry. (1998) 39:185–97. doi: 10.1016/S0010-440X(98)90059-1
12. Buysse DJ, Reynolds CF 3rd, Kupfer DJ, Thorpy MJ, Bixler E, Manfredi R, et al. Clinical diagnoses in 216 insomnia patients using the International Classification of Sleep Disorders (ICSD), DSM-IV and ICD-10 categories: a report from the APA/NIMH DSM-IV field trial. Sleep. (1994) 17:630–7. doi: 10.1093/sleep/17.7.630
13. Mellinger GD, Balter MB, Uhlenhuth EH. Insomnia and its treatment. Prevalence and correlates. Arch Gen Psychiatry. (1985) 42:225–32. doi: 10.1001/archpsyc.1985.01790260019002
14. Yates WR, Mitchell J, Rush AJ, Trivedi MH, Wisniewski SR, Warden D, et al. Clinical features of depressed outpatients with and without co-occurring general medical conditions in STAR*D. Gen Hosp Psychiatry. (2004) 26:421–9. doi: 10.1016/j.genhosppsych.2004.06.008
15. Johnson EO, Roth T, Breslau N. The association of insomnia with anxiety disorders and depression: exploration of the direction of risk. J Psychiatr Res. (2006) 40:700–8. doi: 10.1016/j.jpsychires.2006.07.008
16. Peterson MJ, Benca RM. Sleep in mood disorders. Psychiatr Clin North Am. (2006) 29:1009–32. doi: 10.1016/j.psc.2006.09.003
17. Reynolds CF 3rd, Kupfer DJ. Sleep research in affective illness: state of the art circa (1987) Sleep. (1987) 10:199–215. doi: 10.1093/sleep/10.3.199
18. Palagini L, Baglioni C, Ciapparelli A, Gemignani A, Riemann DREM. sleep dysregulation in depression: state of the art. Sleep Med Rev. (2013) 17:377–90. doi: 10.1016/j.smrv.2012.11.001
19. Clark C, Dupont R, Golshan S, Gillin JC, Rapaport MH, Kelsoe JR. Preliminary evidence of an association between increased REM density and poor antidepressant response to partial sleep deprivation. J Affect Disord. (2000) 59:77–83. doi: 10.1016/S0165-0327(99)00135-4
20. Giles DE, Jarrett RB, Roffwarg HP, Rush AJ. Reduced rapid eye movement latency. A predictor of recurrence in depression Neuropsychopharmacology : official publication of the American College of Neuropsychopharmacology. (1987) 1:33–9. doi: 10.1016/0893-133X(87)90007-8
21. Mendlewicz J. Sleep disturbances: core symptoms of major depressive disorder rather than associated or comorbid disorders. World J Biol Psychiatry. (2009) 10:269–75. doi: 10.3109/15622970802503086
22. Steiger A, Pawlowski M. Depression and sleep. Int J Mol Sci. (2019) 20:607. doi: 10.3390/ijms20030607
23. Monaca C, Boutrel B, Hen R, Hamon M, Adrien J. 5-HT 1A/1B receptor-mediated effects of the selective serotonin reuptake inhibitor, citalopram, on sleep: studies in 5-HT 1A and 5-HT 1B knockout mice. Neuropsychopharmacology. (2003) 28:850–6. doi: 10.1038/sj.npp.1300109
24. Gillin JC, Jernajczyk W, Valladares-Neto DC, Golshan S, Lardon M, Stahl SM. Inhibition of REM sleep by ipsapirone, a 5HT1A agonist, in normal volunteers. Psychopharmacology. (1994) 116:433–6. doi: 10.1007/BF02247474
25. Landolt HP, Kelsoe JR, Rapaport MH, Gillin JC. Rapid tryptophan depletion reverses phenelzine-induced suppression of REM sleep. J Sleep Res. (2003) 12:13–8. doi: 10.1046/j.1365-2869.2003.00336.x
26. Argyropoulos SV, Wilson SJ. Sleep disturbances in depression and the effects of antidepressants. Int Rev Psychiatry. (2005) 17:237–45. doi: 10.1080/09540260500104458
27. Ott GE, Rao U, Lin KM, Gertsik L, Poland RE. Effect of treatment with bupropion on EEG sleep: relationship to antidepressant response. Int J Neuropsychopharmacol. (2004) 7:275–81. doi: 10.1017/S1461145704004298
28. Weber F, Dan Y. Circuit-based interrogation of sleep control. Nature. (2016) 538:51–9. doi: 10.1038/nature19773
29. Abdallah CG, Sanacora G, Duman RS, Krystal JH. Ketamine and rapid-acting antidepressants: a window into a new neurobiology for mood disorder therapeutics. Annu Rev Med. (2015) 66:509–23. doi: 10.1146/annurev-med-053013-062946
30. Autry AE, Adachi M, Nosyreva E, Na ES, Los MF, Cheng PF, et al. receptor blockade at rest triggers rapid behavioural antidepressant responses. Nature. (2011) 475:91–5. doi: 10.1038/nature10130
31. Zanos P, Moaddel R, Morris PJ, Georgiou P, Fischell J, Elmer GI, et al. NMDAR inhibition-independent antidepressant actions of ketamine metabolites. Nature. (2016) 533:481–6. doi: 10.1038/nature17998
32. Aleksandrova LR, Phillips AG, Wang YT. Antidepressant effects of ketamine and the roles of AMPA glutamate receptors and other mechanisms beyond NMDA receptor antagonism. J Psychiatry Neurosci. (2017) 42:222–9. doi: 10.1503/jpn.160175
33. Maeng S, Zarate CA Jr, Du J, Schloesser RJ, McCammon J, Chen G, et al. Cellular mechanisms underlying the antidepressant effects of ketamine: role of alpha-amino-3-hydroxy-5-methylisoxazole-4-propionic acid receptors. Biol Psychiatry. (2008) 63:349–52. doi: 10.1016/j.biopsych.2007.05.028
34. Tononi G, Cirelli C. Sleep and the price of plasticity: from synaptic and cellular homeostasis to memory consolidation and integration. Neuron. (2014) 81:12–34. doi: 10.1016/j.neuron.2013.12.025
35. Pandi-Perumal SR, Kramer M. Sleep and Mental Illness. Cambridge: Cambridge University Press (2010). doi: 10.1017/CBO9781139042734
36. Ehlers CL, Havstad JW, Kupfer DJ. Estimation of the time course of slow-wave sleep over the night in depressed patients: effects of clomipramine and clinical response. Biol Psychiatry. (1996) 39:171–81. doi: 10.1016/0006-3223(95)00139-5
37. Kupfer DJ, Foster FG. Interval between onset of sleep and rapid-eye-movement sleep as an indicator of depression. Lancet. (1972) 2:684–6. doi: 10.1016/S0140-6736(72)92090-9
38. Zhang MQ Li R, Wang YQ, Huang ZL. Neural plasticity is involved in physiological sleep, depressive sleep disturbances, and antidepressant treatments. Neural Plast. (2017) 2017:5870735. doi: 10.1155/2017/5870735
39. Duncan WC, Sarasso S, Ferrarelli F, Selter J, Riedner BA, Hejazi NS, et al. Concomitant BDNF and sleep slow wave changes indicate ketamine-induced plasticity in major depressive disorder. Int J Neuropsychopharmacol. (2013) 16:301–11. doi: 10.1017/S1461145712000545
40. Duncan WC Jr, Selter J, Brutsche N, Sarasso S, Zarate CA Jr. Baseline delta sleep ratio predicts acute ketamine mood response in major depressive disorder. J Affect Disord. (2013) 145:115–9. doi: 10.1016/j.jad.2012.05.042
41. Feinberg I, Campbell IG. Ketamine administration during waking increases delta EEG intensity in rat sleep. Neuropsychopharmacology. (1993) 9:41–8. doi: 10.1038/npp.1993.41
42. Schmid DA, Wichniak A, Uhr M, Ising M, Brunner H, Held K, et al. Changes of sleep architecture, spectral composition of sleep EEG, the nocturnal secretion of cortisol, ACTH, GH, prolactin, melatonin, ghrelin, and leptin, and the DEX-CRH test in depressed patients during treatment with mirtazapine. Neuropsychopharmacology. (2006) 31:832–44. doi: 10.1038/sj.npp.1300923
43. Wichniak A, Wierzbicka A, Walecka M, Jernajczyk W. Effects of antidepressants on sleep. Curr Psychiatry Rep. (2017) 19:63. doi: 10.1007/s11920-017-0816-4
44. Mallick BN, Singh A, Khanday MA. Activation of inactivation process initiates rapid eye movement sleep. Prog Neurobiol. (2012) 97:259–76. doi: 10.1016/j.pneurobio.2012.04.001
45. Haas H, Panula P. The role of histamine and the tuberomamillary nucleus in the nervous system. Nat Rev Neurosci. (2003) 4:121–30. doi: 10.1038/nrn1034
46. Krishnan V, Nestler EJ. Animal models of depression: molecular perspectives. Curr Top Behav Neurosci. (2011) 7:121–47. doi: 10.1007/7854_2010_108
47. Yokogawa T, Marin W, Faraco J, Pézeron G, Appelbaum L, Zhang J, et al. Characterization of sleep in zebrafish and insomnia in hypocretin receptor mutants. PLoS Biol. (2007) 5:e277. doi: 10.1371/journal.pbio.0050277
48. Barlow IL, Rihel J. Zebrafish sleep: from geneZZZ to neuronZZZ. Curr Opin Neurobiol. (2017) 44:65–71. doi: 10.1016/j.conb.2017.02.009
49. Rihel J, Prober DA, Arvanites A, Lam K, Zimmerman S, Jang S, et al. Zebrafish behavioral profiling links drugs to biological targets and rest/wake regulation. Science. (2010) 327:348–51. doi: 10.1126/science.1183090
50. Sigurgeirsson B, Thornorsteinsson H, Sigmundsdóttir S, Lieder R, Sveinsdóttir HS, Sigurjónsson ÓE, et al. Sleep-wake dynamics under extended light and extended dark conditions in adult zebrafish. Behav Brain Res. (2013) 256:377–90. doi: 10.1016/j.bbr.2013.08.032
51. Sorribes A, Thornorsteinsson H, Arnardóttir H, Jóhannesdóttir I, Sigurgeirsson B, de Polavieja GG, et al. The ontogeny of sleep-wake cycles in zebrafish: a comparison to humans. Front Neural Circuits. (2013) 7:178. doi: 10.3389/fncir.2013.00178
53. Panula P, Chen YC, Priyadarshini M, Kudo H, Semenova S, Sundvik M, et al. The comparative neuroanatomy and neurochemistry of zebrafish CNS systems of relevance to human neuropsychiatric diseases. Neurobiol Dis. (2010) 40:46–57. doi: 10.1016/j.nbd.2010.05.010
54. de Abreu MS, Friend AJ, Demin KA, Amstislavskaya TG, Bao W, Kalueff AV. Zebrafish models: do we have valid paradigms for depression? J Pharmacol Toxicol Methods. (2018) 94(Pt 2):16–22. doi: 10.1016/j.vascn.2018.07.002
55. Egan RJ, Bergner CL, Hart PC, Cachat JM, Canavello PR, Elegante MF, et al. Understanding behavioral and physiological phenotypes of stress and anxiety in zebrafish. Behav Brain Res. (2009) 205:38–44. doi: 10.1016/j.bbr.2009.06.022
56. Marcon M, Herrmann AP, Mocelin R, Rambo CL, Koakoski G, Abreu MS, et al. Prevention of unpredictable chronic stress-related phenomena in zebrafish exposed to bromazepam, fluoxetine and nortriptyline. Psychopharmacology. (2016) 233:3815–24. doi: 10.1007/s00213-016-4408-5
57. Song C, Liu BP, Zhang YP, Peng Z, Wang J, Collier AD, et al. Modeling consequences of prolonged strong unpredictable stress in zebrafish: complex effects on behavior and physiology. Prog Neuropsychopharmacol Biol Psychiatry. (2018) 81:384–94. doi: 10.1016/j.pnpbp.2017.08.021
58. Vanini G, Wathen BL, Lydic R, Baghdoyan HA. Endogenous GABA levels in the pontine reticular formation are greater during wakefulness than during rapid eye movement sleep. J Neurosci. (2011) 31:2649–56. doi: 10.1523/JNEUROSCI.5674-10.2011
59. Gardner JP, Fornal CA, Jacobs BL. Effects of sleep deprivation on serotonergic neuronal activity in the dorsal raphe nucleus of the freely moving cat. Neuropsychopharmacology. (1997) 17:72–81. doi: 10.1016/S0893-133X(97)00025-0
60. Frank MG. Sleep and developmental plasticity not just for kids. Prog Brain Res. (2011) 193:221–32. doi: 10.1016/B978-0-444-53839-0.00014-4
61. Reicher V, Kis A, Simor P, Bódizs R, Gácsi M. Interhemispheric asymmetry during NREM sleep in the dog. Sci Rep. (2021) 11:18817. doi: 10.1038/s41598-021-98178-3
62. Schubert TA, Chidester RM, Chrisman CL. Clinical characteristics, management and long-term outcome of suspected rapid eye movement sleep behaviour disorder in 14 dogs. J Small Anim Pract. (2011) 52:93–100. doi: 10.1111/j.1748-5827.2010.01026.x
63. Kovács E, Kosztolányi A, Kis A. Rapid eye movement density during REM sleep in dogs (Canis familiaris). Learn Behav. (2018) 46:554–60. doi: 10.3758/s13420-018-0355-9
64. Takahashi Y, Ebihara S, Nakamura Y, Takahashi K. Temporal distributions of delta wave sleep and rem sleep during recovery sleep after 12-h forced wakefulness in dogs; similarity to human sleep. Neurosci Lett. (1978) 10:329–34. doi: 10.1016/0304-3940(78)90248-3
65. Kis A, Szakadát S, Gácsi M, Kovács E, Simor P, Török C, et al. The interrelated effect of sleep and learning in dogs (Canis familiaris); an EEG and behavioural study. Sci Rep. (2017) 7:41873. doi: 10.1038/srep41873
66. Takeuchi T, Harada E. Age-related changes in sleep-wake rhythm in dog. Behav Brain Res. (2002) 136:193–9. doi: 10.1016/S0166-4328(02)00123-7
67. Aserinsky E. Relationship of rapid eye movement density to the prior accumulation of sleep and wakefulness. Psychophysiology. (1973) 10:545–58. doi: 10.1111/j.1469-8986.1973.tb00804.x
68. Sánchez C, Brennum LT, Stórustovu S, Kreilgård M, Mørk A. Depression and poor sleep: the effect of monoaminergic antidepressants in a pre-clinical model in rats. Pharmacol Biochem Behav. (2007) 86:468–76. doi: 10.1016/j.pbb.2007.01.006
69. Andersen SL. Exposure to early adversity: Points of cross-species translation that can lead to improved understanding of depression. Dev Psychopathol. (2015) 27:477–91. doi: 10.1017/S0954579415000103
70. Yamada Y, Yoshikawa T, Naganuma F, Kikkawa T, Osumi N, Yanai K. Chronic brain histamine depletion in adult mice induced depression-like behaviours and impaired sleep-wake cycle. Neuropharmacology. (2020) 175:108179. doi: 10.1016/j.neuropharm.2020.108179
71. Cheeta S, Ruigt G, van Proosdij J, Willner P. Changes in sleep architecture following chronic mild stress. Biol Psychiatry. (1997) 41:419–27. doi: 10.1016/S0006-3223(96)00058-3
72. Novati A, Roman V, Cetin T, Hagewoud R, den Boer JA, Luiten PG, et al. Chronically restricted sleep leads to depression-like changes in neurotransmitter receptor sensitivity and neuroendocrine stress reactivity in rats. Sleep. (2008) 31:1579–85. doi: 10.1093/sleep/31.11.1579
73. Dugovic C, Solberg LC, Redei E, Van Reeth O, Turek FW. Sleep in the Wistar-Kyoto rat, a putative genetic animal model for depression. Neuroreport. (2000) 11:627–31. doi: 10.1097/00001756-200002280-00038
74. Lagus M, Gass N, Saharinen J, Savelyev S, Porkka-Heiskanen T, Paunio T. Inter-tissue networks between the basal forebrain, hippocampus, and prefrontal cortex in a model for depression caused by disturbed sleep. J Neurogenet. (2012) 26:397–412. doi: 10.3109/01677063.2012.694932
75. Le Dantec Y, Hache G, Guilloux JP, Guiard BP, David DJ, Adrien J, et al. sleep hypersomnia and reduced sleep/wake continuity in a neuroendocrine mouse model of anxiety/depression based on chronic corticosterone administration. Neuroscience. (2014) 274:357–68. doi: 10.1016/j.neuroscience.2014.05.050
76. Mendoza J. Circadian insights into the biology of depression: symptoms, treatments and animal models. Behav Brain Res. (2019) 376:112186. doi: 10.1016/j.bbr.2019.112186
77. Vogel G, Neill D, Kors D, Hagler M. REM sleep abnormalities in a new animal model of endogenous depression. Neurosci Biobehav Rev. (1990) 14:77–83. doi: 10.1016/S0149-7634(05)80163-0
78. Popa D, El Yacoubi M, Vaugeois JM, Hamon M, Adrien J. Homeostatic regulation of sleep in a genetic model of depression in the mouse: effects of muscarinic and 5-HT1A receptor activation. Neuropsychopharmacology. (2006) 31:1637–46. doi: 10.1038/sj.npp.1300948
79. Wang ZJ, Yu B, Zhang XQ, Sheng ZF, Li SJ, Huang YL, et al. Correlations between depression behaviors and sleep parameters after repeated corticosterone injections in rats. Acta Pharmacol Sin. (2014) 35:879–88. doi: 10.1038/aps.2014.44
80. Yasenkov R, Deboer T. Circadian modulation of sleep in rodents. Prog Brain Res. (2012) 199:203–18. doi: 10.1016/B978-0-444-59427-3.00012-5
81. Ivarsson M, Paterson LM, Hutson PH. Antidepressants and REM sleep in Wistar-Kyoto and Sprague-Dawley rats. Eur J Pharmacol. (2005) 522:63–71. doi: 10.1016/j.ejphar.2005.08.050
82. Kennaway DJ. Clock genes at the heart of depression. J Psychopharmacol. (2010) 24 (2 Suppl):5–14. doi: 10.1177/1359786810372980
83. Worlein JM. Nonhuman primate models of depression: effects of early experience and stress. ILAR J. (2014) 55:259–73. doi: 10.1093/ilar/ilu030
84. Ishikawa A, Sakai K, Maki T, Mizuno Y, Niimi K, Oda Y, et al. Investigation of sleep-wake rhythm in non-human primates without restraint during data collection. Exp Anim. (2017) 66:51–60. doi: 10.1538/expanim.16-0073
85. Suárez FR. Neurobiology of neuronal circuits involved in depression. Anal Real Academia Nacional Medicina. (2012) 129:75–97; discussion 97–101.
86. Chu X. Preliminary validation of natural depression in macaques with acute treatments of the fast-acting antidepressant ketamine. Behav Brain Res. (2019) 360:60–8. doi: 10.1016/j.bbr.2018.11.044
87. Qiu P, Jiang J, Liu Z, Cai Y, Huang T, Wang Y, et al. BMAL1 knockout macaque monkeys display reduced sleep and psychiatric disorders. Natl Sci Rev. (2019) 6:87–100. doi: 10.1093/nsr/nwz002
88. Zhang ZY, Mao Y, Feng XL, Zheng N, Lü LB, Ma YY, et al. Early adversity contributes to chronic stress induced depression-like behavior in adolescent male rhesus monkeys. Behav Brain Res. (2016) 306:154–9. doi: 10.1016/j.bbr.2016.03.040
89. Barr CS, Newman TK, Schwandt M, Shannon C, Dvoskin RL, Lindell SG, et al. Sexual dichotomy of an interaction between early adversity and the serotonin transporter gene promoter variant in rhesus macaques. Proc Natl Acad Sci USA. (2004) 101:12358–63. doi: 10.1073/pnas.0403763101
90. Hrdina PD, von Kulmiz P, Stretch R. Pharmacological modification of experimental depression in infant macaques. Psychopharmacology. (1979) 64:89–93. doi: 10.1007/BF00427351
91. Willard SL, Friedman DP, Henkel CK, Shively CA. Anterior hippocampal volume is reduced in behaviorally depressed female cynomolgus macaques. Psychoneuroendocrinology. (2009) 34:1469–75. doi: 10.1016/j.psyneuen.2009.04.022
92. Shively CA, Register TC, Friedman DP, Morgan TM, Thompson J, Lanier T. Social stress-associated depression in adult female cynomolgus monkeys (Macaca fascicularis). Biol Psychol. (2005) 69:67–84. doi: 10.1016/j.biopsycho.2004.11.006
93. Shively CA, Willard SL. Behavioral and neurobiological characteristics of social stress versus depression in nonhuman primates. Exp Neurol. (2012) 233:87–94. doi: 10.1016/j.expneurol.2011.09.026
94. Harding JD. Nonhuman primates and translational research: progress, opportunities, and challenges. ILAR J. (2017) 58:141–50. doi: 10.1093/ilar/ilx033
95. Bliss-Moreau E, Rudebeck PH. Animal models of human mood. Neurosci Biobehav Rev. (2021) 120:574–82. doi: 10.1016/j.neubiorev.2020.06.024
96. Goncharova ND, Marenin VY, Oganyan TE. Aging of the hypothalamic-pituitary-adrenal axis in nonhuman primates with depression-like and aggressive behavior. Aging. (2010) 2:854–66. doi: 10.18632/aging.100227
97. Xu F, Wu Q, Xie L, Gong W, Zhang J, Zheng P, et al. Macaques exhibit a naturally-occurring depression similar to humans. Sci Rep. (2015) 5:9220. doi: 10.1038/srep09220
98. Lancel M, van Riezen H, Glatt A. Effects of circadian phase and duration of sleep deprivation on sleep and EEG power spectra in the cat. Brain Res. (1991) 548:206–14. doi: 10.1016/0006-8993(91)91123-I
99. Trachsel L, Tobler I, Achermann P, Borbély AA. Sleep continuity and the REM-nonREM cycle in the rat under baseline conditions and after sleep deprivation. Physiol Behav. (1991) 49:575–80. doi: 10.1016/0031-9384(91)90283-T
100. Valentino RJ, Volkow ND. Drugs, sleep, and the addicted brain. Neuropsychopharmacology. (2020) 45:3–5. doi: 10.1038/s41386-019-0465-x
101. Zammit GK. Ramelteon: a novel hypnotic indicated for the treatment of insomnia. Psychiatry. (2007) 4:36–42.
102. Reinhardt KD, Vyazovskiy VV, Hernandez-Aguilar RA, Imron MA, Nekaris KA. Environment shapes sleep patterns in a wild nocturnal primate. Sci Rep. (2019) 9:9939. doi: 10.1038/s41598-019-45852-2
103. Borbély AA, Tobler I. Manifestations and functional implications of sleep homeostasis. Handb Clin Neurol. (2011) 98:205–13. doi: 10.1016/B978-0-444-52006-7.00013-7
104. Sanford LD, Fang J, Tang X. Sleep after differing amounts of conditioned fear training in BALB/cJ mice. Behav Brain Res. (2003) 147:193–202. doi: 10.1016/S0166-4328(03)00180-3
105. Suchecki D, Tiba PA, Machado RB. REM sleep rebound as an adaptive response to stressful situations. Front Neurol. (2012) 3:41. doi: 10.3389/fneur.2012.00041
106. Yasugaki S, Liu CY, Kashiwagi M, Kanuka M, Honda T, Miyata S, et al. Effects of 3 weeks of water immersion and restraint stress on sleep in mice. Front Neurosci. (2019) 13:1072. doi: 10.3389/fnins.2019.01072
107. Pawlyk AC, Morrison AR, Ross RJ, Brennan FX. Stress-induced changes in sleep in rodents: models and mechanisms. Neurosci Biobehav Rev. (2008) 32:99–117. doi: 10.1016/j.neubiorev.2007.06.001
108. Logan RW, Edgar N, Gillman AG, Hoffman D, Zhu X, McClung CA. Chronic stress induces brain region-specific alterations of molecular rhythms that correlate with depression-like behavior in mice. Biol Psychiatry. (2015) 78:249–58. doi: 10.1016/j.biopsych.2015.01.011
109. Bedrosian TA, Fonken LK, Walton JC, Haim A, Nelson RJ. Dim light at night provokes depression-like behaviors and reduces CA1 dendritic spine density in female hamsters. Psychoneuroendocrinology. (2011) 36:1062–9. doi: 10.1016/j.psyneuen.2011.01.004
110. Fonken LK, Finy MS, Walton JC, Weil ZM, Workman JL, Ross J, et al. Influence of light at night on murine anxiety- and depressive-like responses. Behav Brain Res. (2009) 205:349–54. doi: 10.1016/j.bbr.2009.07.001
111. Fonken LK, Kitsmiller E, Smale L, Nelson RJ. Dim nighttime light impairs cognition and provokes depressive-like responses in a diurnal rodent. J Biol Rhythms. (2012) 27:319–27. doi: 10.1177/0748730412448324
112. Eastman C, Rechtschaffen A. Circadian temperature and wake rhythms of rats exposed to prolonged continuous illumination. Physiol Behav. (1983) 31:417–27. doi: 10.1016/0031-9384(83)90061-6
113. Ikeda M, Sagara M, Inoué S. Continuous exposure to dim illumination uncouples temporal patterns of sleep, body temperature, locomotion and drinking behavior in the rat. Neurosci Lett. (2000) 279:185–9. doi: 10.1016/S0304-3940(99)00943-X
114. Wells AM, Ridener E, Bourbonais CA, Kim W, Pantazopoulos H, Carroll FI, et al. Effects of chronic social defeat stress on sleep and circadian rhythms are mitigated by kappa-opioid receptor antagonism. J Neurosci. (2017) 37:7656–68. doi: 10.1523/JNEUROSCI.0885-17.2017
115. Kamphuis J, Lancel M, Koolhaas JM, Meerlo P. Deep sleep after social stress: NREM sleep slow-wave activity is enhanced in both winners and losers of a conflict. Brain Behav Immun. (2015) 47:149–54. doi: 10.1016/j.bbi.2014.12.022
116. Olini N, Rothfuchs I, Azzinnari D, Pryce CR, Kurth S, Huber R. Chronic social stress leads to altered sleep homeostasis in mice. Behav Brain Res. (2017) 327:167–73. doi: 10.1016/j.bbr.2017.03.022
117. Barr CS, Newman TK, Becker ML, Parker CC, Champoux M, Lesch KP, et al. The utility of the non-human primate; model for studying gene by environment interactions in behavioral research. Genes Brain Behav. (2003) 2:336–40. doi: 10.1046/j.1601-1848.2003.00051.x
118. Balzamo E, Santucci V, Seri B, Vuillon-Cacciuttolo G, Bert J. Nonhuman primates: laboratory animals of choice for neurophysiologic studies of sleep. Lab Anim Sci. (1977) 27:879–86.
119. Chen YM, Qin DD, Jiang HH, Hu XT, Ma YY. Sleep disorder, a potential early diagnostic marker for psychiatric and neurodegenerative diseases. Zool Res. (2011) 32:81–90. doi: 10.3724/SP.J.1141.2011.01081
120. Deadwyler SA, Porrino L, Siegel JM, Hampson RE. Systemic and nasal delivery of orexin-A (Hypocretin-1) reduces the effects of sleep deprivation on cognitive performance in nonhuman primates. J Neurosci. (2007) 27:14239–47. doi: 10.1523/JNEUROSCI.3878-07.2007
121. Weitzman ED, Kripke DF, Pollak C, Dominguez J. Cyclic activity in sleep of Macaca mulatta. Arch Neurol. (1965) 12:463–7. doi: 10.1001/archneur.1965.00460290019003
122. Hsieh KC, Robinson EL, Fuller CA. Sleep architecture in unrestrained rhesus monkeys (Macaca mulatta) synchronized to 24-hour light-dark cycles. Sleep. (2008) 31:1239–50.
123. Kripke DF, Reite ML, Pegram GV, Stephens LM, Lewis OF. Clinical and laboratory notes. Nocturnal sleep in rhesus monkeys. Electroencephal Clin Neurophysiol. (1968) 24:582–6. doi: 10.1016/0013-4694(68)90047-3
124. Feinberg I, Koresko RL, Heller NEEG. sleep patterns as a function of normal and pathological aging in man. J Psychiatr Res. (1967) 5:107–44. doi: 10.1016/0022-3956(67)90027-1
125. Markun LC, Sampat A. Clinician-focused overview and developments in polysomnography. Curr Sleep Med Rep. (2020) 23:1–13. doi: 10.1007/s40675-020-00197-5
126. Lucey BP, McLeland JS, Toedebusch CD, Boyd J, Morris JC, Landsness EC, et al. Comparison of a single-channel EEG sleep study to polysomnography. J Sleep Res. (2016) 25:625–35. doi: 10.1111/jsr.12417
127. Qin DD, Feng SF, Zhang FY, Wang N, Sun WJ, Zhou Y, et al. Potential use of actigraphy to measure sleep in monkeys: comparison with behavioral analysis from videography. Zool Res. (2020) 41:437–43. doi: 10.24272/j.issn.2095-8137.2020.056
128. Li Z, Li Z, Lv X, Li Z, Xiong L, Hu X, et al. Intracerebroventricular administration of interferon-alpha induced depressive-like behaviors and neurotransmitter changes in rhesus monkeys. Front Neurosci. (2020) 14:585604. doi: 10.3389/fnins.2020.585604
129. Qin DD, Rizak J, Feng XL, Yang SC, Lü LB, Pan L, et al. Prolonged secretion of cortisol as a possible mechanism underlying stress and depressive behaviour. Sci Rep. (2016) 6:30187. doi: 10.1038/srep30187
130. Qin D, Chu X, Feng X, Li Z, Yang S, Lü L, et al. The first observation of seasonal affective disorder symptoms in Rhesus macaque. Behav Brain Res. (2015) 292:463–9. doi: 10.1016/j.bbr.2015.07.005
131. Hemmeter UM, Hemmeter-Spernal J, Krieg JC. Sleep deprivation in depression. Expert Rev Neurother. (2010) 10:1101–15. doi: 10.1586/ern.10.83
132. Walker MP, van der Helm E. Overnight therapy? The role of sleep in emotional brain processing. Psychol Bull. (2009) 135:731–48. doi: 10.1037/a0016570
133. Benca RM, Peterson MJ. Insomnia and depression. Sleep Med. (2008) 9(Suppl 1):S3–9. doi: 10.1016/S1389-9457(08)70010-8
134. Staner L. Comorbidity of insomnia and depression. Sleep Med Rev. (2010) 14:35–46. doi: 10.1016/j.smrv.2009.09.003
135. Wilson S, Argyropoulos S. Antidepressants and sleep: a qualitative review of the literature. Drugs. (2005) 65:927–47. doi: 10.2165/00003495-200565070-00003
136. Berger M, Riemann D. Symposium: Normal and abnormal REM sleep regulation: REM sleep in depression-an overview. J Sleep Res. (1993) 2:211–23. doi: 10.1111/j.1365-2869.1993.tb00092.x
137. Fruth B, Tagg N, Stewart F. Sleep and nesting behavior in primates: a review. Am J Phys Anthropol. (2018) 166:499–509. doi: 10.1002/ajpa.23373
Keywords: depression, sleep, non-human primate, brain development, animal model
Citation: Li M, Cui J, Xu B, Wei Y, Fu C, Lv X, Xiong L and Qin D (2022) Sleep Disturbances and Depression Are Co-morbid Conditions: Insights From Animal Models, Especially Non-human Primate Model. Front. Psychiatry 12:827541. doi: 10.3389/fpsyt.2021.827541
Received: 02 December 2021; Accepted: 31 December 2021;
Published: 25 January 2022.
Edited by:
Sheng Wei, Shandong University of Traditional Chinese Medicine, ChinaReviewed by:
Jiaojian Wang, University of Electronic Science and Technology of China, ChinaCopyright © 2022 Li, Cui, Xu, Wei, Fu, Lv, Xiong and Qin. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Dongdong Qin, cWluZG9uZzEwOEAxNjMuY29t; Lei Xiong, eGxsdWNrQHNpbmEuY29t; Xiaoman Lv, bHhtLmNjQGZveG1haWwuY29t
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.