 Andrea Perrottelli
Andrea Perrottelli Giulia Maria Giordano
Giulia Maria Giordano Francesco Brando
Francesco Brando Luigi Giuliani
Luigi Giuliani Armida Mucci
Armida Mucci
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
SYSTEMATIC REVIEW article
Front. Psychiatry , 04 May 2021
Sec. Schizophrenia
Volume 12 - 2021 | https://doi.org/10.3389/fpsyt.2021.653642
This article is part of the Research Topic New Developments with Magnetoelectrical Techniques in Schizophrenia View all 5 articles
Introduction: Electrophysiological (EEG) abnormalities in subjects with schizophrenia have been largely reported. In the last decades, research has shifted to the identification of electrophysiological alterations in the prodromal and early phases of the disorder, focusing on the prediction of clinical and functional outcome. The identification of neuronal aberrations in subjects with a first episode of psychosis (FEP) and in those at ultra high-risk (UHR) or clinical high-risk (CHR) to develop a psychosis is crucial to implement adequate interventions, reduce the rate of transition to psychosis, as well as the risk of irreversible functioning impairment. The aim of the review is to provide an up-to-date synthesis of the electrophysiological findings in the at-risk mental state and early stages of schizophrenia.
Methods: A systematic review of English articles using Pubmed, Scopus, and PsychINFO was undertaken in July 2020. Additional studies were identified by hand-search. Electrophysiological studies that included at least one group of FEP or subjects at risk to develop psychosis, compared to healthy controls (HCs), were considered. The heterogeneity of the studies prevented a quantitative synthesis.
Results: Out of 319 records screened, 133 studies were included in a final qualitative synthesis. Included studies were mainly carried out using frequency analysis, microstates and event-related potentials. The most common findings included an increase in delta and gamma power, an impairment in sensory gating assessed through P50 and N100 and a reduction of Mismatch Negativity and P300 amplitude in at-risk mental state and early stages of schizophrenia. Progressive changes in some of these electrophysiological measures were associated with transition to psychosis and disease course. Heterogeneous data have been reported for indices evaluating synchrony, connectivity, and evoked-responses in different frequency bands.
Conclusions: Multiple EEG-indices were altered during at-risk mental state and early stages of schizophrenia, supporting the hypothesis that cerebral network dysfunctions appear already before the onset of the disorder. Some of these alterations demonstrated association with transition to psychosis or poor functional outcome. However, heterogeneity in subjects' inclusion criteria, clinical measures and electrophysiological methods prevents drawing solid conclusions. Large prospective studies are needed to consolidate findings concerning electrophysiological markers of clinical and functional outcome.
Schizophrenia (SCZ) is a severe and complex mental disorder, demonstrating heterogeneity in terms of risk factors, comorbidities, clinical presentations, course, response to treatment, and functional outcome. It approximately affects 26 million people and is currently regarded as one of the leading causes of disability worldwide (1–3). Despite significant advances in the available pharmacological and psychosocial interventions, the impairment in real-life functioning, represents, to date, an unmet need in the care of people suffering from this disorder, with a huge burden on patients, their families, and health-care systems (4–10). Different factors, some related to the illness, others to personal resources, and others to the context, have been demonstrated to contribute to the impairment in functioning (6–8, 10, 11). Among these factors, prolonged periods of untreated psychosis have a role in determining a chronic course of symptoms and a poor functional outcome (12, 13). Therefore, in the last decades much more effort has been invested in the early detection and intervention in schizophrenia, aiming to decrease the risk of deterioration associated to a chronic and relapsing course of the illness.
The first episode of psychosis (FEP) is a crucial stage in the course of schizophrenia, representing the transition from a premorbid to a morbid state. This stage is usually preceded by a “prodromal” period, during which subjects might present gradual and subtle changes in thoughts, perceptions, behaviors, cognition, and functioning (14–16). This period is associated with affective symptoms, social withdrawal, cognitive deficits, attenuated positive psychotic symptoms, and impairment in functioning, which is strongly related to cognitive deficits (17–19). This clinical syndrome has been termed as “at risk mental state” (ARMS) and operationalized criteria were developed to categorize subjects within the clinical high-risk (CHR) or ultra high-risk (UHR) status (14, 20, 21).
It has been demonstrated that within 3 years following the onset of prodromal symptoms, about 18–36% of ARMS subjects make a transition from a premorbid to a morbid state (17). Those ARMS subjects that do not develop a psychosis will present another psychiatric disorder or persistent attenuated symptoms, while only about 14% will have symptomatic remission (22–24).
It seems, therefore, crucial to promptly detect early stages of psychosis, including the state of vulnerability and the onset of psychosis, in order to implement adequate interventions, reduce the rate of transition to psychosis, as well as the risk of further progression to deteriorating stages and impairment in real-life functioning.
In order to characterize subjects that are at risk to develop a psychosis and those with a FEP, research efforts have been directed toward the establishment of the neurobiological underpinnings of these early illness stages, excluding the bias of chronicity, medications, and institutionalization, present in subjects with chronic schizophrenia (18, 25–28). These neurobiological correlates can be investigated effectively with electroencephalography (EEG). Indeed, EEG represents a good and appropriate technique to analyze the neurophysiology of both normal and psychotic experience and behavior, based on an integrative, complex and in-vivo model of the brain (29–33). In addition, this technique is non-invasive, and, in comparison to other imaging techniques, such as functional magnetic resonance imaging (fMRI), EEG has the advantages that it is more flexible in study design, has lower costs and it exhibits a superior temporal resolution. Furthermore, through source analysis methodology, such as Low Resolution Electromagnetic Tomography (LORETA), it is possible to obtain information about brain areas from which the neuronal activity is generated (34, 35).
EEG-based measures can be categorized schematically into three categories. The first one considers the oscillatory nature of neuronal activity. This usually involves dividing the continuous recorded EEG signal into its different frequency bands (delta, theta, alpha, beta, and gamma) (36). These oscillatory rhythms can be recorded while subjects are in state of relaxation and without any external stimulation or, alternatively, during sensory stimulation or while performing a task.
The second category is represented by microstates (MS), defined as brief periods during which global electrical brain activity remains semi-stable. These transient periods of stability last between 80 and 120 ms (37, 38). Each microstate is classified on the basis of its corresponding EEG scalp potential map (39, 40). Microstates are hypothesized to be the most basic instantiations of human neuronal functions and are thus nicknamed as “the atoms of thought.”
Finally, the third category is constituted by event-related potentials (ERPs), which reflect the neuronal response following a specific sensory, cognitive, or motor event (32, 41). These EEG indices can manifest as positive and negative voltage deflections, waves, or components with a precise temporal correlation to the onset of a specific event (42).
A vast EEG literature has documented different abnormalities of neuronal activity in subjects with chronic SCZ, as compared to healthy controls (HCs) (29, 33, 43–49). In particular, several studies have consistently reported alterations in the activity of the whole spectrum of frequency bands (49–53), changes in MSs topography and/or other parameters (46, 47, 54) and a reduction of amplitude in ERPs, such as N100 (55–58), mismatch negativity (MMN) (59, 60), and P300 (33, 57, 61). Furthermore, these alterations have been related to the severity of symptoms, as well as cognitive and functional impairment in schizophrenia. For instance, aberrant frequency bands activity has been associated to cognitive deficits and positive and negative symptoms (50–53); alterations in MS parameters to negative symptoms, hallucinations, and duration of illness (46, 47, 54); reductions in N100 amplitude to auditory hallucinations and attention deficits (55, 57, 58); deficits in MMN to positive symptoms and functioning impairment (57, 59, 60); P300 to neurocognitive impairments and negative symptoms (57, 61).
In the last decades, a large number of studies has highlighted how some of the abnormalities of EEG indices, reported in subjects with chronic SCZ, can also be observed in the ARMS and prodromal or early phases of schizophrenia. Previous reviews on this topic (25, 44, 62–68) have considered either at-risk/prodromal states or early stages of schizophrenia or only one specific electrophysiological index.
In the light of these observations, the aim of the present study is to review the current evidence concerning abnormalities of electrophysiological indices, including all three categories of indices mentioned above, in both CHR/UHR and FEP subjects.
The Preferred Reporting Items for Systematic Reviews (PRISMA) statement has been followed to design and conduct the systematic review (69).
In brief, we performed a comprehensive literature search on abnormalities of EEG indices in the early stages of schizophrenia (FEP and CHR/UHR subjects).
In order to facilitate the comprehension of the reader, we clarify the terms that we used in the manuscript. The term FEP encompasses both affective-spectrum disorders (bipolar disorder and major depressive disorder with psychotic features) and schizophrenia spectrum disorders (schizophrenia, schizoaffective disorder, and schizophreniform disorder). The term first episode of schizophrenia (FES) is used to indicate exclusively those subjects who, although falling also within the FEP categorization, are specifically at the onset of a schizophrenia spectrum disorder. Therefore, in the present manuscript, whenever studies did not specify the characteristics of the FEP sample or if the sample was heterogeneous and included subjects with either a first-episode of a schizophrenia-spectrum disorder or of an affective disorder (i.e., bipolar disorder or major depressive disorder with psychotic features), we indicated the sample as “FEP.” Conversely, we indicated as “FES” the samples for which the authors clearly specified a diagnosis of a schizophrenia-spectrum disorder. Through the whole manuscript with the term “high risk” (HR) we will refer to subjects in the ARMS or prodromal stages of the illness. The original nomenclature of the study samples will be kept within tables reporting the description of the studies included in the present review.
A systematic literature search was conducted in three electronic databases: PubMed, Scopus, and PsychINFO on 13th July 2020 with no time limit and with English language as the only selected filter, in order to ensure that it was as comprehensive as possible (Table 1).

Table 1. Systematic search strategies.
The following combination of search terms was used:
(EEG OR electroencephalography OR “EEG microstate” OR “dipole source localization” OR sLORETA OR LORETA OR eLORETA OR ERP OR “event-related potential” OR “spectral analysis” OR “frequency domain analysis” OR “spectral band” OR “neural oscillations” OR “spectral power” OR N100 OR N1 OR MMN OR “mismatch negativity” OR P300 OR P3a OR P3b OR “event-related” OR “evoked potential” OR “evoked-response”) AND (“ultra-high risk psychosis” OR “clinical high risk psychosis” OR “prodromal psychosis” OR “first episode schizophrenia” OR “first episode psychosis” OR “early onset schizophrenia”).
The search terms were selected to include both general terms related to EEG research and more specific indices (such as specific ERPs) that have been consistently investigated in research papers attaining to schizophrenia. In addition, reference lists were hand-searched to identify additional publications missed by the search strategy.
Firstly, any duplicate from the combination of the three databases was excluded. The remaining articles were included in the systematic review only if they met the following criteria:
Inclusion criteria
1. meta-analysis, reviews, case-control studies concerning the abnormalities of EEG indices in the at-risk, prodromal and early stages of schizophrenia;
2. studies carried out in humans;
3. studies published in English;
4. studies that included at least one group of subjects during their at-risk, prodromal or early stages of illness, compared with a healthy control group;
Exclusion criteria
1. books chapters, comments, editorials, case reports/case series, theses, proceedings, letters, short surveys, notes;
2. studies irrelevant to the topic;
3. unavailable full-text.
If the studies included in the present review reported data concerning differences between HR subjects who made the transition to psychosis (HR-T) from those who did not (HR-NT), as well as data concerning differences between subjects at at-risk/prodromal/early stages of psychosis and subjects with chronic schizophrenia, these data have been also incorporated in the present paper.
Two researchers (AP, FB) independently screened for eligibility all the articles by titles and abstracts and then proceeded to read the full text. Discrepancies in the selection of the eligible articles have been discussed in advance with the whole group and were resolved by discussion and consensus.
We recorded the following variables from each included article: author/s, year of publication, EEG index evaluated, study population, assessment instruments for diagnosis and EEG data results (Supplementary Tables 1–3). Given the heterogeneity of experimental paradigms and considered variables in the eligible studies, we did not plan to carry out a meta-analysis.
The combined outcome of the three databases results yielded a total of 614 records (Figure 1). In addition, 40 studies were included by hand search. Of the total studies, 335 were duplicates, leaving 319 articles. After reading the titles and abstracts, 154 of these were excluded because they were not relevant to the topic of the review or because they were articles other than meta-analysis, reviews and case-control studies. The full text of the remaining 165 studies was examined in more detail. It appeared that 32 studies did not meet the inclusion criteria due to methodological discrepancies (i.e., no control group included, no clear EEG data reported, no clear explanation of diagnostic criteria of sample). Therefore, a total number of 133 studies were finally identified as eligible for inclusion in the current review (Figure 1).
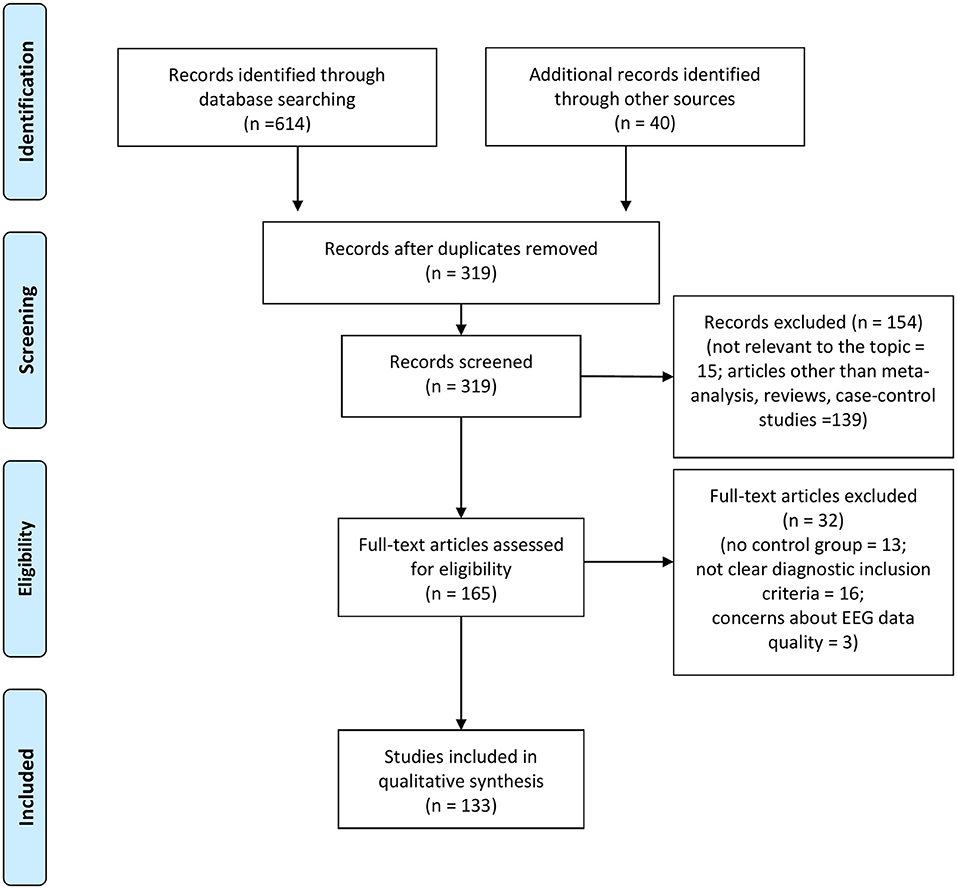
Figure 1. PRISMA flow chart of included studies.
The EEG raw signal can be decomposed into five main oscillatory rhythms or broad frequency bands, namely delta (0.5–4.0 Hz), theta (4–8 Hz), alpha (8–13 Hz), beta (13–30 Hz), and gamma (30–100 Hz) bands (43, 62, 70). These bands can be investigated during both resting-state condition and during sensory stimulation or task performance. In the context of frequency bands analysis, different measures have been considered, such as the power and the source localization of each frequency band activity and indices of coherence, synchronization, and neuronal connectivity. The studies included for this section are reported in Supplementary Table 1.
Delta activity, characterized by a spectral bandwidth 0.5–4.0 Hz, is the predominant slow wave in states of unconsciousness, such as sleep and anesthesia. Furthermore, following external stimulation, synchronization of evoked responses in delta band across neuronal regions, plays also a role in motivational, emotional, and cognitive functions (71, 72).
Resting-state EEG data showed consistent increases in delta band power in subjects with chronic SCZ, as compared to healthy controls (33, 43, 72). Therefore, different studies have been conducted aiming to verify whether this abnormality is present since the early phases of the illness.
Findings concerning delta band during resting-state in the early phases of schizophrenia are controversial, with some studies reporting no abnormality in delta power (73) or synchrony (74), while others finding differences between these subjects and HCs in power (75–79) or functional connectivity (80) within this band activity.
Specifically, studies during a resting-state condition that found abnormalities in delta band in FES compared to HCs, have reported a higher power (75–77). Furthermore, additional studies and analyses also showed a lower delta amplitude peak (77), an excess of delta activity in prefrontal areas (78), and a distributed hypo-synchronization of delta activity between cerebral regions, mostly observed in parietal areas as suggested through LORETA analysis (78, 80). One longitudinal study in FEP subjects, with a 1-year follow-up, which also considered electrode location for its analysis, found that higher baseline delta activity at posterior regions predicted improvement in positive symptoms after 1 year, while lower values in the same band at frontal regions were associated with the amelioration of negative symptoms (76).
An increase in delta power in frontal areas, considering scalp electrode location, has been observed also in HR-T compared to HR-NT and to HCs, while no difference between HR-NT and HCs was recorded (79).
Three studies investigated delta activity while subjects performed a task (81–83). In one study, FES subjects, as compared to HCs, had a decrease in the occurrence of anterior–to-posterior propagation of delta waves during auditory and visual tasks (82). A second study, carried out in FEP subjects, demonstrated a workload-dependent increase of the duration of delta oscillations during a working memory task (81) not observed in HCs. This was revealed by longer delta oscillation cycles in FEP during the performance of tasks of increasing difficulty. Another study reported no difference between FES and HCs in terms of delta functional connectivity and its topological properties assessed while subjects performed an executive functioning task (83).
Theta oscillatory activity (4–8 Hz) seems to be involved in the orchestration of several cognitive processes, such as working memory, detection of new sensory stimuli and attentional control (84). It has been reported that subjects with chronic SCZ present different abnormalities in this frequency range, often characterized by excessive frontal theta activity (33, 43, 85, 86).
Inconsistent findings have been reported in studies investigating abnormalities of the theta band during a resting state condition in FES, FEP, and HR subjects, as compared to HCs. In particular, some studies reported no significant difference in the theta band activity between HCs and FEP (87), FES (73, 76, 77), or HR (87) subjects. Other studies reported an increase in theta band activity in FES subjects compared to HCs (75, 88) during resting conditions. The increase in theta activity in FES was associated with the severity of negative symptoms (88) and this alteration was observed also in HR-T if compared to HR-NT and HCs (79). Abnormalities in theta oscillations were also found in studies that performed a source localization analysis through LORETA (78, 89). These studies found a decrease of theta activity specifically in the anterior regions in FEP (89) and FES subjects (78) as compared to HCs. Furthermore, a complex LORETA study investigating theta-gamma amplitude phase coupling (regulation of the gamma band activity depending on the phase of theta band activity) showed an alteration in this index in a FES sample in the posterior cingulate cortex (90). Some studies investigated the neural connectivity during resting-state and found a decrease in the global field synchronization (74), or an increase in the synchronization between posterior cingulate cortex, cuneus, and precuneus (80) or an increased connectivity between brain networks (91–93) in the theta band in FES subjects, as compared to HCs. Connectivity values were also related to clinical symptoms (91), worse verbal memory (92), and processing speed (93). On the other side, a study that included also HR subjects, did not detect any significant difference between these subjects and HCs in theta functional connectivity (92).
Abnormalities in theta activity were reported also during task performance (83, 88, 94–96). In particular, task-related abnormal theta activity was detected during a processing speed (94), an arithmetic (88) or an auditory task (95) in FES (88, 95) and FEP subjects (94). In addition, two studies highlighted brain network dysfunctions in theta band in FES as compared to HCs, as suggested by abnormalities in neuronal information transmission during an executive functioning task (83) and a reduction of the “small-world network” index (96), a parameter assessing cerebral networks topology and the efficiency of neuronal signaling processing. Conversely, the latter index has not been found to be different between HR subjects and HCs in another study (96).
Alpha oscillation is one of the most prominent neuronal rhythm in the adult human brain, both during resting-state condition and task performance, and it is characterized by a frequency spectrum ranging from 8 to 12 Hz (97). Neuronal oscillations within this frequency band play a pivotal role in cognition, consciousness, sensorimotor and emotional processes (98). In subjects with SCZ, a decrease in absolute power during resting-state conditions (43) and disruptions in temporal coherence in evoked oscillations during sensory stimulation and cognitive tasks have been reported (99).
During at-risk and early phases of the disorder, alpha activity shows already alterations in its features. Several studies revealed a reduction in alpha frequency activity (75, 88) in diverse and widespread cerebral areas including frontal (73, 89, 100), parietal (73, 78, 100), temporal (78, 100), and occipital (100) regions in FES and FEP subjects compared to HCs, as assessed through LORETA. Conversely, other studies found no significant difference in the alpha power between HCs and FEP (87, 100), FES (73, 76, 77), and HR (87) subjects. Furthermore, no robust differences were detected in alpha power when HR-T, HR-NT, and HCs were compared (79).
Inconsistent results have been reported also for EEG connectivity indices recorded during rest. One study (74) did not find any difference between FES subjects and HCs in terms of global field synchronization. Conversely, other studies reported abnormalities in connectivity-related indices in alpha band, such as a lower coherence (101), a generalized hypo-synchronization across cerebral regions (80) and lower phase-lag index (PLI) (93) in FES subjects, compared to HCs. Yet, another study reported significantly higher PLI values of alpha in FES and HR subjects compared to HCs (102), contradicting previous results (93).
In studies focusing on stimuli or task-related activity, it was shown that alpha activity, evoked by error commitment, was significantly increased in FES subjects (94) compared to HCs. Task-related connectivity studies focusing on alpha band found either no difference in signal complexity and brain network communication (83), or a decrease in coherence (101) in FES compared to HCs. Finally, a reduction in alpha desynchronization (103) and a higher clustering coefficient of the alpha band (96) were detected in HR as compared to HCs.
Beta oscillation (12–30 Hz) has been studied mostly in relation to sensorimotor behavior and cognitive processes, such as working memory and top-down regulation of attention (104, 105). In schizophrenia, beta-band abnormalities manifest as increased activity in resting-state (106, 107) and in relationship to perceptual integration (108).
Five studies found no differences between FEP or FES subjects, as compared to HCs in the power of the beta frequency band during a resting-state condition (73, 75–77, 109). Conversely, three studies found differences in the beta power between FEP (89) or FES (78, 88) subjects and HCs. In particular, these studies found an increase in beta power in FEP and FES, which was associated with the severity of negative symptoms (88) and localized mainly in the right parietal area (78) and frontal gyrus (89), as revealed by LORETA analysis (78, 89). Focusing on connectivity measures during rest, FES subjects, compared to HCs, showed a lower EEG coherence (109), as well as a generalized hypo-synchronization in the beta band (80). However, a study reported no significant difference between FES subjects and HCs in the global field synchronization of the beta band (74).
In task-related recordings, an increase in the power (88) and a reduction in coherence (101) were reported for the beta band in FES subjects, compared to HCs. The decrease in coherence was also associated to the severity of positive symptoms in one of the studies (101). Finally, no abnormal mean value was recorded through the evaluation of path length of networks activity within the beta band, while subjects performed an executive functioning task (83).
Gamma activity represents the fastest oscillations (30–100 Hz) of the spectrum, and these fast waves are generated through the synchronized activation of pyramidal neurons located in the cerebral cortex (62, 110). Gamma band oscillations have been linked to a vast variety of cognitive and perceptual integration processes (110) and showed several abnormalities in subjects with SCZ, often associated to impairments in neurocognitive functions (111).
A systematic review has already summarized how abnormalities in gamma band have been vastly reported in FEP and HR subjects (62).
During resting state conditions, an increase in gamma power was reported in FES (112), FEP (89), and HR-T subjects (113) vs. HCs, mainly located in the frontal regions (89, 113) when LORETA analysis was implemented. However, no difference in resting-state gamma activity was observed in three other studies (88, 91, 109), between FES subjects and HCs. As regard to connectivity indices during rest, a generalized hypo-synchronization (80), an increase in connectivity (91), a decrease in coherence (101, 109) and a reduced phase lag index (PLI) (93) in the gamma band were found in FES subjects, as compared to HCs.
During sensory stimulation or the performance of a task, different results have been reported. In particular, several studies reported abnormalities in the evoked responses (mainly decrease in evoked gamma), synchronization (reduction or a delay in the synchronization) and connectivity (generally a reduction in functional connectivity) of gamma activity during the performance of memory (81, 114), cognitive control (115), emotion processing (116, 117), or auditory tasks (118–122) both in FES (114, 115, 117–122) and FEP (81, 116) subjects, as compared to HCs. Furthermore, alterations in gamma synchrony were also related to social cognition in one study (117), while another one, employing a longitudinal design, highlighted that the improvement in positive symptoms in FES subjects was related to the increase in gamma synchrony (121). Findings concerning the impairment in synchronization of gamma during the performance of a task in HR subjects are discrepant. Perez et al. (123) reported a decrease in gamma evoked response. A similar result was also reported in a study using an auditory task and multimodal recordings (simultaneous fMRI and EEG), which showed that HR subjects presented a reduction in gamma evoked response, characterized mainly by deficits in activity of the auditory, thalamus, and frontal brain areas (124). Furthermore, although Oribe et al. (118) found a decrease in the coherence of auditory-evoked gamma activity, assessed through phase-locking factor (PLF) in FES, as in the study by Leicht et al. (122), no abnormality was found in HR subjects, as compared to HCs, indicating that this alteration might be linked to more advanced illness phases (118).
Another way to investigate gamma activity is through the analysis of the auditory steady-state response (ASSR), which involves the presentation of auditory stimuli at high frequencies with subsequent entrainment of oscillatory activity at the same frequencies. In a study involving auditory stimulation at 20, 30, and 40 Hz, the FEP group had significantly reduced phase locking and evoked power compared to HCs (125) for gamma ASSR elicited with 30 and 40 Hz stimuli. In this study, higher phase locking was related to more severe positive symptoms (125). Furthermore, studies found a decrease in the evoked gamma power (125), inter-trial phase coherence and spectral perturbation of ASSR (126, 127) in FES subjects compared to HCs. It was also noticed that these alterations in ASSR activity were related to general psychopathology and attentional deficits (126).
In a study with subjects at-risk, no difference was detected in ASSR-evoked gamma power or PLF between the HR subjects and control groups (128), while in another study a decrease of inter-trial phase coherence (ITC) and event-related spectral perturbations in late phases of ASSR was found in HR compared to HCs (126).
The microstates (MS) are EEG-based measures that define the global functional state of the brain by its momentary scalp electric field configuration (39, 129, 130). There is a small set of prototypical MS configurations, which constitutes a basic repertoire of brain functional states: MS-A, MS-B, MS-C, MS-D. In particular, MS-A and MS-B have been found to be associated to BOLD signal within fronto-temporal and occipital regions, areas belonging to the phonological and visual networks, while MS-C and MS-D have been linked to cingulate cortex, right superior and middle frontal gyri, the right superior and inferior parietal lobules, regions involved in the default mode, salience and attention networks (131, 132). Evidence of EEG MS alterations in subjects with SCZ has been widely reported (25, 46, 47, 133). However, few studies investigated these alterations in the early phases of the illness. The studies included in the current review are summarized Supplementary Table 2.
FES subjects, as compared to HCs, showed a reduced duration of MS-B (134) and D (54, 134), an increased occurrence of MS-A (54, 134) and C (134), as well as an increased contribution of MS-A (134) and D (54). The reduced duration of MS-D has been found to correlate with the severity of paranoid symptomatology (54). In addition, the MS syntax A → C → D → A, which predominated in HCs, was reversed in FES (A → D → C → A) (134). Finally, the topography differed between FES and HCs, with FES showing a stronger left and anterior activity of MS-B (134) and central activity of MS-D (54).
Some of the above-mentioned abnormalities have been reported already in HR subjects (135, 136). In particular, HR, as compared to HCs and SCZ subjects, showed an increase in the contribution and occurrence of MS-A; they also showed a reduction in the contribution of MS-B, as compared to SCZ. The aberrant spatial configuration of MS-B, which exhibited a stronger activity in the left posterior cingulate in SCZ subjects, was displayed to a lesser extent also in HR, as compared to HCs (136). In individuals with 22q11.2 deletion syndrome (22q11DS), known to have a 30-fold increased risk to develop schizophrenia, an increased presence of MS-C vs. HCs, was found and was associated with hallucinations (135).
A variety of ERPs, related to sensory-perceptual and cognitive events, have been utilized in schizophrenia research due to their high sensitivity to transient changes in neuronal activity. The following paragraphs report if alterations in P50, N100, mismatch negativity (MMN), P300, and N400 components are detectable already in FEP and HR subjects. These studies are summarized in Supplementary Table 3.
P50 is an early event-related positive potential, which is recorded ~50 ms after the presentation of an auditory click stimulus. In a paired-click paradigm, characterized by two subsequent stimuli, a reduction in P50 response after the second stimulus (S2), compared to P50 recorded after the first one (S1), is expected. This is the outcome of a regulatory mechanism known as sensory gating, which is assessed using the P50 ratio (S2-P50 divided by S1-P50). When subjects do not show a diminished response to the second stimulus, a defect in sensory gating is likely to have occurred. Several articles reported an increase in the P50 ratio and difference in SCZ, suggesting a deficit in sensory gating (137–142).
A recent meta-analysis reported a consistent impairment in P50 sensory gating in FEP subjects, as compared to HCs, and highlighted that this deficit had a similar magnitude to the one reported in subjects with chronic SCZ (44). Different studies found a deficit in sensory gating, as measured with the P50 ratio, in FEP (143–146), FES (147, 148), and HR subjects (143, 148, 149), as compared to HCs. The grade of the impairment seemed to be influenced by the clinical presentation of the illness (no sensory gating deficit has been detected in FES subjects during the post-acute phase, after improvement of positive symptoms) (147). Some other studies did not find an impairment in sensory gating in FEP (150–153), FES (154, 155), and HR (150, 151, 154, 156, 157) subjects. Furthermore, a study involving the innovative implementation of machine learning (ML) to distinguish FES from HCs with P50-related measures (amplitude and ratio), in addition to other neuroimaging and clinical evaluations, highlighted that this EEG-index did not contribute significantly to the discrimination performed by the mathematical model (158).
In a study involving FES and HR subjects, which analyzed P50 through LORETA, it was shown that both groups presented differences in the brain functional networks sustaining this ERP, and that these two groups actually showed similarities, suggesting compromised gating already at at-risk stages (154). In particular, FES subjects showed a greater connectivity in the right superior frontal gyrus and right insula, while HR subjects had a greater connectivity in the paracentral lobule and the middle temporal gyrus, as compared to HCs (154).
N100 is one of the largest auditory and visually evoked ERP and can be visualized as a negative deflection peaking between 80 and 120 ms after the stimulus onset, with its maximal amplitude recorded over fronto-central leads (159). Subjects with chronic schizophrenia, compared to HCs, show a robust reduction in the amplitude of N100 and in N100-related measures of sensory gating (56, 159, 160). Findings concerning the presence of abnormalities of N100 in the early stages of psychosis are controversial. In particular, some studies demonstrated that FEP subjects, compared to HCs, showed a reduced N100 amplitude both during visual (161) and auditory paradigms (162–164), while other studies did not detect any N100 amplitude impairment in FEP subjects (152, 165–167). In HR subjects one study reported a reduction in N100 amplitude (163), while most of the studies reported similar values of N100 amplitude in HR subjects and HCs (161, 165, 168–171).
When sensory gating was assessed through the presentation of two subsequent stimuli, the N100 amplitude difference (S2-S1) (143, 150) and the N100 gating ratio (S2/S1) (143, 150, 152) showed a significant increase in FEP (143, 150, 152) and HR subjects (143, 150), suggesting an impairment in the processing of redundant stimuli (143, 150). Conversely, some studies did not find any statically significant difference in N100 amplitude (152) or N100 gating ratio (151) in FEP (150–152) and HR subjects (151), as compared to HCs.
N100 latency did not differ in FEP or in HR subjects compared to HCs in any of the studies cited above that analyzed this variable (143, 161–163, 167, 168, 170, 172).
MMN is a negative ERP elicited by the presentation of a “deviant” rare sound in a repetitive sequence of “standard” tones that generally occurs after 150–250 ms upon the presentation of the deviant stimulus (173–176). The deviant stimulus in the auditory modality can differ from the standard one in terms of duration (dMMN) or pitch (pMMN) (177). Currently, a deficit in MMN elicitation is one of the most robust and replicable findings in schizophrenia and it has been related to cognitive dysfunctions, as well as functional impairment in people suffering from this disorder (178–185). Interestingly, the impairment in MMN has been reported already in the early stages of psychotic disorders (63, 65, 186, 187). The reproducibility of dMMN deficit for at-risk and early stages of schizophrenia is greater than that of pMMN, with the latter emerging more robustly only during the chronic stage of schizophrenia. Thus, dMMN can be a more sensitive marker than pMMN, in the context of early psychosis (30, 65).
Discrepant findings have been reported with the regard to the dMMN amplitude. In particular, some studies found a reduction of dMMN amplitude in FEP (66, 151, 188–191) and FES subjects (65, 95, 181, 182, 190, 192–196) as compared to HCs, while other studies did not find any abnormality in dMMN amplitude in FEP and FES subjects (183, 197–200). Furthermore, a study, involving ML to distinguish FES from HCs with dMMN measures in addition to other neuroimaging and clinical evaluations, highlighted that this EEG-index did not contribute significantly to the discriminant ML-model (158).
A reduction in dMMN amplitude has been reported also in HR subjects (66, 151, 191, 192, 194, 196, 201–204). In particular, it seems that abnormalities in dMMN amplitude might predict the onset of psychosis since they were present in HR-T and not in HR-NT subjects (181, 196, 201–203, 205, 206). However, some studies failed to find differences between HR subjects and HCs in the dMMN amplitude (180, 181, 205, 207).
Most of the studies did not find significant differences between HCs, FEP (188, 189), FES (193, 194, 198, 201, 206), and HR subjects (180, 194, 201, 204, 206, 207) in dMMN latency, with the exception of one study that found delayed latency in dMMN peak in FEP subjects (196), as compared to HCs.
Findings concerning pMMN amplitude are also inconsistent. Specifically, several studies reported that FES subjects, as compared to HCs, showed a significant reduction of pMMN amplitude (164, 190, 198, 208); a multimodal-longitudinal study showed that the deficit became evident approximately after 1.5 years from illness onset and was correlated with a reduction in Heschl's gyrus volume (198). Other studies, however, did not confirm the above-mentioned results (65, 164, 183, 192, 194, 197, 199, 200, 209–211). As for the dMMN, also the pMMN amplitude did not contribute to the ML model created to differentiate FES subjects from HCs (158).
Most studies reported no abnormalities in pMMN amplitude in HR subjects (both HR-T and HR-NT) (180, 192, 194, 205, 207, 209), while only two studies reported a pMMN amplitude reduction in HR subjects (203, 204). All of the studies that considered pMMN latency did not find significant differences between HCs and FEP (209) or FES (194) or HR subjects (194, 204, 207, 209).
The P300 is an ERP positive deflection that appears after the onset of rare deviant “target” stimuli embedded in a sequence of frequent “standard” stimuli (212). Previous reviews have showed impairments of this EEG index in at-risk, prodromal, early, and chronic phases of schizophrenia (33, 63, 67).
P300 is not a unitary phenomenon, but it is composed of 2 functionally different subcomponents: an early component, the P3a, which usually peaks within a time window of 130–275 ms after the stimulus onset, that reflects an involuntary shift in attention toward a deviant stimulus; a later component, the P3b, observed within a time window of 275–600 ms after the stimulus, that reflects a conscious and controlled attentive process toward a stimulus and its task-relevance (60). Some studies explicitly differentiated P3a and P3b, analyzing one or both subcomponents in the same sample, while others referred more generally to P300 recordings. In most of studies which referred to a general P300, P300 was considered as the most positive deflection within a time window of 250–600 ms after the stimulus, thus referring mainly to the P3b component.
In order to record and characterize the P300, most of the studies addressing impairments in at-risk and early stages of schizophrenia have employed visual and auditory tasks. In particular, only three studies used a visual paradigm (161, 165, 170) and found a reduced amplitude (161, 165, 170) and prolonged latency (161, 170) of P300 in FES (161, 165) and HR (161, 165, 170), as compared to HCs. The two following subsections will include results of those studies that recorded P3a and P3b components all during auditory paradigms.
In FEP and FES subjects different studies have highlighted a decrease of the P3a amplitude compared to HCs (163, 188, 189, 194, 196, 200, 209, 210, 213). On the other side, some studies failed to detect differences in P3a amplitude between HCs and FEP (190, 195). Interestingly, in one study, deficits in P3a were not present at the baseline, but emerged when FEP subjects were evaluated at 12 and 24 months follow-up visits (166).
The decrease in P3a amplitude has been reported also in several studies including HR subjects, as compared to HCs (68, 128, 163, 194, 196, 209, 214), while two studies did not find any difference in P3a amplitude between HR and control groups (206, 207). One study found P3a amplitude reduction in HR-T compared to HR-NT (196), while two studies did not find any difference between these two groups (68, 214).
Regarding the P3a latency just one study of those mentioned above highlighted a delayed P3a latency in FES subjects (213), while all other studies did not identify any alteration in P3a latency, in FEP, FES and HR subjects (68, 163, 188, 189, 196, 206, 207, 209, 213–215).
In FEP and FES subjects, as compared to HCs, different studies have highlighted a decrease of the P3b amplitude (67, 163, 164, 172, 199, 210, 213, 215–223). One study demonstrated that deficits in P3b, as observed for P3a, were not present at baseline, but emerged when FEP subjects were evaluated at 12- and 24-month follow-up visits (166). Furthermore, using LORETA, it was shown that the reduction of P3b amplitude was mainly driven by dysfunctions in the left temporal regions (222), while a multimodal study, using MRI and EEG, has shown that P3b reduction was specifically associated with left superior temporal gyrus gray matter volume reduction (224). Only one study did not detect any alteration of the P3b amplitude in FEP subjects, compared to HCs (167).
Different studies have reported how the decrease in P3b amplitude is present since the at-risk phases of the illness (68, 163, 168, 169, 171, 214, 225–227). Interestingly, one study highlighted a step-wise decline in P3b amplitude throughout illness course, characterized by a progressive decrease of P3b in subjects at at-risk, early and chronic stages of SCZ (P3b amplitude in HCs>HR>FEP> chronic SCZ) (226). However, other studies found that P3b amplitude was as impaired in HR as in subjects at more advanced stages of illness (163, 214, 227). Several studies showed P3b amplitude decrease in HR-T compared to HR-NT and HCs (68, 171, 214), while just one study did not detect any difference in P3b amplitude between converters and not converters (225). A study divided the HR sample in HR-T and HR-NT and the authors performed a further subdivision for the latter group into remitted and non-remitted. It was observed that HR-T subjects and HR-NT who continued to present attenuated symptoms showed reduced P300 compared to HR-NT who remitted and HCs (228).
With regard to the P3b latency, two studies reported an increase of this EEG measure in FEP (215, 229), while other studies did not detect any abnormalities of this feature in FEP, FES (162, 163, 165, 167, 172, 213, 216–219, 221, 222, 224, 227, 230–232), and HR subjects (68, 163, 165, 168, 169, 227, 228), as compared to HCs.
N400 is a negative-going deflection that peaks around 400 ms post-stimulus onset and it is typically maximal over centro-parietal electrode sites. The N400 is part of the normal brain response to words and other meaningful stimuli, including visual and auditory words, sign-language, pictures, faces, environmental sounds, and smells (233). Several studies have revealed that subjects with chronic schizophrenia presents abnormalities in this ERPs values, with some results suggesting that N400 semantic priming deficits may reflect an underlying neurophysiological mechanism of delusions (234).
However, to date, very few articles have addressed this EEG-index in early stages of schizophrenia. In one study, N400 presented a reduced amplitude and a prolonged latency in FES, as compared to HCs (235). The reduced N400 amplitude has also been found in HR subjects, as compared to HCs (128), and was associated with neurocognitive impairment (236).
EEG recordings provide in-vivo access to neuronal activity and each EEG index can reflect distinct sensory and cognitive processes. This review illustrates how multiple EEG-based indices result already altered during at-risk and early stages of schizophrenia, supporting the hypothesis that cerebral networks dysfunctions appear early in the course of the disorder (25, 237, 238). However, although a large number of studies have highlighted differences between FEP or HR subjects and HCs on EEG variables, only few of these showed homogeneous and consistent results.
Studies on frequency bands have reported several abnormalities across all five bands in at-risk and early stages of schizophrenia.
Within delta band, an increase in resting-state activity in both at-risk and first-episode subjects compared to HCs has often been reported (75–79). Studies have suggested that dysfunctions in this band might arise from changes in dopamine synthesis levels in the fronto-striatal-thalamic loops, which are detectable already at the onset of psychotic disorders and result associated to the severity of prodromal psychotic symptoms (29, 239). Furthermore, this EEG index has also been successfully used to predict the trajectory of negative symptoms and functioning in FES subjects (76).
Studies on theta and alpha bands reported mixed results (73, 75–77, 87, 88, 100). However, the activity of these two bands during a task performance revealed alterations in both FEP and HR samples in almost all of the studies considered. The association of these bands to clinical symptoms (88, 91, 94) and cognitive domains (91, 92, 94, 95) suggests that abnormalities in neuronal oscillations in these frequency bands could contribute to the clinical presentation in early disease stages.
For the beta band, only few studies (78, 88, 89) reported alterations, while the majority did not find significant abnormalities in early and at-risk subjects (73–77, 109). The same is true for studies investigating stimuli-related activity, coherence, and connectivity measures in this band.
Finally, for the gamma band, abnormalities of the resting-state power, evoked power, synchrony, coherence, and connectivity were observed in the early stages of schizophrenia (62). Therefore, considering the role of gamma oscillations in the cognitive processes (110, 111), the widespread cognitive deficits observed already at early stages of schizophrenia (17, 240, 241) might be connected to abnormalities in the gamma activity across cerebral networks.
Considering these results on the whole, it is important to underline that the diversity of the EEG paradigms and analysis methods employed in the studies, do not allow drawing solid conclusions.
Abnormalities in the characteristics of MS, such as their mean duration and the presence of abnormal patterns in the MS syntax, suggest that sustainment of neuronal activity in interconnected cerebral regions is impaired already at early stages of the disorder (25, 54, 134–136).
Furthermore, in a recent study, which included FEP, HR-T, HR-NT, and HCs, the authors discussed how abnormalities of different microstate parameters might be linked to different aspects of the illness (242). In particular, abnormalities of MS-A in FEP and HR subjects could represent an unspecific state biomarker of general psychopathology; abnormalities of MS-B in FEP may represent a state biomarker specific to psychotic illness progression; and finally, abnormalities of MS-D in HR-T (and not HR-NT) might represent a biomarker of future transition of HR subjects (242). However, to date, very few articles have investigated the microstates in HR and FEP subjects, revealing a research gap opportunity for future EEG studies.
The studies included in the current review showed that in FEP and HR subjects compared to HCs, all considered ERPs presented abnormalities, generally manifested as a reduction in their amplitude. These deficits were detected both for ERPs linked to the basic levels of sensory processing, such as P50 and N100, and for indices related to higher levels of cognitive functions, such as MMN and P3b (63). For instance, the results reported for P50, support previous evidence that sensory gating is already impaired in at-risk and early stages of schizophrenia and may reflect a diminished capacity to filter repetitive sensory signals (44). Conversely, impairments in N100 seem to be more consistent in FEP rather than HR subjects, suggesting that this type of deficit might emerge only at a morbid stage of the disorder. The relationship of this ERP abnormalities with primary negative symptoms (160, 243), which might be present only in a subgroup of subjects with schizophrenia, might partially explain the heterogeneity of the results. One general limitation of the studies addressing at-risk and prodromal stages is the poor characterization of negative symptoms and cognitive deficits, which are not included in diagnostic criteria though they might be predictive of poor clinical and functional outcome and conversion to schizophrenia (18). Recent data have demonstrated that early auditory processing deficits, as assessed either by neuropsychological tasks or MMN (244, 245), are present only in a subgroup of subjects with schizophrenia. In at-risk subjects the same deficits are not always found (244), indicating that either cognitive and neurophysiological deficits develop only from the onset of the morbid phase or that individuals with premorbid deficits are not being captured by current criteria which do not consider core aspects of schizophrenia-spectrum disorders, such as negative symptoms.
Amongst the ERPs indices, MMN and P300 showed the highest rate of abnormal values in the studies included in the present review. Articles focusing on MMN have shown that reductions in the amplitude of dMMN and pMMN seem to be different depending on the phase of the disorder. Reductions in dMMN, in fact, precedes the one in pMMN, which is almost never observed in HR subjects and only few times in FEP subjects (64). Thus, dMMN can be a more sensitive marker than pMMN in the context of prodromal and early psychosis (30, 65, 95, 185, 186). Alternatively, pMMN reductions could reflect illness chronicity and could be used to monitor treatment efficacy and disease progression (64, 198). In addition to MMN results, also P300 might be an index used as a robust and effective biomarker for transition to psychosis in HR and for prognosis of the disorder in FEP subjects (67, 68, 171, 214, 226).
Finally, the low number of studies measuring N400 in FEP and HR prevents the formulation of any inference on this component.
The integration of the results reported in the current systematic review is compromised by three major limitations: the use of different diagnostic criteria and assessment scales, the heterogeneity of EEG paradigms and analysis methods used in the included studies and, finally, the intra and inter-subjects' variability of EEG recordings.
Firstly, both for at-risk and early phases of schizophrenia, inclusion criteria were based on different operational definitions. For instance, for at-risk or prodromal phases, ARMS, UHR, and CHR operational criteria have been used in different studies. In addition, the occurrence of a first episode was defined as the first contact at a clinical setting, or the duration of antipsychotic medication use or the duration of psychotic symptoms (26).
Secondly, there was a huge heterogeneity in experimental paradigms, types of EEG indices considered and the definition of their characteristics such as band ranges cut-off points and the employed analysis methods. This was mostly evident in frequency band rather than in ERP studies.
Finally, EEG studies are generally characterized by a high variability within and between subjects, which might not always be indicative of any pathological status, making difficult a generalization of the reports. In fact, EEG data recordings might be influenced by normal temporal fluctuations in subject physiology, as well as caffeine and nicotine intake, skull conductivity, several medical conditions, and medication (246, 247).
The application of EEG indices into clinical settings to predict the likelihood of conversion to psychosis from a high-risk state or the onset and the progression of the illness is still under scrutiny. Due to the high variance of the results considered in the present review, none of these EEG indices can be regarded as a flawless marker of at-risk or early stages of schizophrenia, and no clinical translation has yet been envisaged for any of them (62, 64, 68). For instance, alterations in EEG-based measures such as N100, MMN or P300 are indicators of deficits in early sensory processing. However, the literature reported that these impairments are dichotomously distributed among schizophrenia subjects during their early and chronic stages of illness, so they are not found in all affected patients. For this reason, also in subjects at-risk of schizophrenia, deficits in early sensory processes are not always reported, leading to discrepant results. Furthermore, it should be considered that the presence of sensory processing deficits is not included among ARMS criteria, which might imply that some subjects can be missed using current diagnostic systems (244, 245).
Another consideration that should be made, is that most of the studies included in the review rely on comparisons of HR and FEP subjects to HCs on only one EEG index, while only few analyzed multiple EEG indices simultaneously (76, 79, 163, 248, 249). The latter approach could be more effective in predicting clinical and functional outcome both in early and prodromal phases of schizophrenia or to characterize the neurobiological alterations in initial phases. For instance, Renaldi et al. (76) used a multiple regression analysis with delta, theta, alpha, and beta spectral power with the aim to predict symptomatic and functional improvement in FES subjects. Furthermore, a study investigated the utilization of N100, P3a, and P3b amplitudes to discriminate HR and FES subjects from HCs, showing that only the first two EEG-indices were significant predictors of the diagnosis (163). Some authors analyzed changes in multiple frequency band indices, using a longitudinal design, to identify which could predict conversion to psychosis in HR subjects (79, 248, 249). In one of these studies, delta, theta and alpha activity contributed to a predictive model of conversion (79). Furthermore, another study (249) showed that a regression model using delta, theta, and beta activity, combined with clinical data, was able to predict with an accuracy of over 80% the transition to psychosis in HR subjects. Conversely, one study reported that absolute power values of frontal alpha, beta, and delta activity were not associated to transition to psychosis (248).
In the last decades, machine learning technique became popular to overcome the limits of univariate analyses, which require a preselection of variables to be used in prediction. This promising approach is based on the use of multiple variables (e.g., electrophysiological, genetic, or clinical data) and the algorithm enables general hypotheses and previsions (e.g., discrimination between patients and controls, prediction of the response to treatment or clinical course). This approach has been used in different imaging studies, with the aim to predict clinical information at individual level (250–252). However, in studies with subjects at early and prodromal stages of schizophrenia, only few research groups have employed this promising method with EEG data. Among these, one study utilized the machine learning algorithm, which incorporated measures of current-source density (CSD) and LORETA synchronization indices of beta and gamma oscillations, in order to predict HR subjects who would develop psychosis in a 3-years follow-up (253). Furthermore, another study found that EEG-measures, such as P50 and MMN, could identify the presence of two distinct subgroups within a FES sample, which could potentially assist clinicians in treatment design based on the individual neurobiological differences (254). In a recent longitudinal study (255), machine learning was used to discriminate two subgroups of FEP subjects, according to changes in dMMN amplitude, revealing that subjects with improvement of dMMN had better clinical, cognitive, and functional follow-up outcomes than those with worsening of dMMN (255). However, another study did not find a significant contribution of electrophysiological indices, such as P50 and MMN, for the discrimination between FES and HCs (158).
Besides the multivariate approach, it could be very useful to use multimodal analyses in which, for instance, simultaneous EEG-MRI is recorded in order to achieve a good spatial-temporal resolution. However, few of the included studies have been conducted using a multimodal approach, analyzing and correlating MRI and EEG abnormalities in these groups of subjects (124, 198, 224, 256).
Finally, given that progressive changes in EEG measures are associated with transition to psychosis and disease course, more studies with a longitudinal design (76, 79, 121, 255, 256) are needed.
The current systematic review advocates the conduction of further studies on EEG indices that could support clinicians in their decision-making process in the early stage of the disorder. In order to draw reliable conclusions from the combination of the various studies, standardized subjects' inclusion criteria, electrophysiological protocols, and analysis methods should be adopted.
Studies should include multiple EEG indices, integrate them with other clinical variables and apply multivariate approaches, such as machine learning algorithms, in order to provide a reliable tool in the diagnosis and prognosis of early and prodromal stages of schizophrenia.
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
AP, GG, and AM contributed to the conceptualization and supervision of the manuscript. AP, GG, and FB contributed to the establishment of the methodology and the literature research. All authors contributed to writing, critically revising, and editing the content of the manuscript and approved the final manuscript for submission to Frontiers in Psychiatry.
General sources of potential conflict of interest, considered unrelated to this work include the following: AM received honoraria, advisory board or consulting fees from the following companies: Amgen Dompé, Angelini-Acraf, Astra Zeneca, Bristol-Myers Squibb, Gedeon Richter Bulgaria, Innova-Pharma, Janssen Pharmaceuticals, Lundbeck, Otsuka, Pfizer and Pierre Fabre.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpsyt.2021.653642/full#supplementary-material
1. Charlson FJ, Ferrari AJ, Santomauro DF, Diminic S, Stockings E, Scott JG, et al. Global epidemiology and burden of schizophrenia: findings from the global burden of disease study 2016. Schizophrenia Bull. (2018) 44:1195–203. doi: 10.1093/schbul/sby058
2. Vos T, Abajobir AA, Abate KH, Abbafati C, Abbas KM, Abd-Allah F, et al. Global, regional, and national incidence, prevalence, and years lived with disability for 328 diseases and injuries for 195 countries, 1990-2016: a systematic analysis for the Global Burden of Disease Study 2016. Lancet. (2017) 390:1211–59. doi: 10.1016/S0140-6736(17)32154-2
3. Leucht S, Cipriani A, Spineli L, Mavridis D, Orey D, Richter F, et al. Comparative efficacy and tolerability of 15 antipsychotic drugs in schizophrenia: a multiple-treatments meta-analysis. Lancet. (2013) 382:951–62. doi: 10.1016/S0140-6736(13)60733-3
4. Harvey PD, Strassnig M. Predicting the severity of everyday functional disability in people with schizophrenia: cognitive deficits, functional capacity, symptoms, and health status. World Psychiatry. (2012) 11:73–9. doi: 10.1016/j.wpsyc.2012.05.004
5. Fleischhacker WW, Arango C, Arteel P, Barnes TR, Carpenter W, Duckworth K, et al. Schizophrenia–time to commit to policy change. Schizophrenia Bull. (2014) 40(Suppl. 3):S165–94. doi: 10.1093/schbul/sbu006
6. Galderisi S, Rucci P, Kirkpatrick B, Mucci A, Gibertoni D, Rocca P, et al. Interplay among psychopathologic variables, personal resources, context-related factors, and real-life functioning in individuals with schizophrenia: a network analysis. JAMA Psychiatry. (2018) 75:396–404. doi: 10.1001/jamapsychiatry.2017.4607
7. Galderisi S, Mucci A, Buchanan RW, Arango C. Negative symptoms of schizophrenia: new developments and unanswered research questions. Lancet Psychiatry. (2018) 5:664–77. doi: 10.1016/S2215-0366(18)30050-6
8. Galderisi S, Rossi A, Rocca P, Bertolino A, Mucci A, Bucci P, et al. The influence of illness-related variables, personal resources and context-related factors on real-life functioning of people with schizophrenia. World Psychiatry. (2014) 13:275–87. doi: 10.1002/wps.20167
9. Harvey PD, Strassnig MT, Silberstein J. Prediction of disability in schizophrenia: symptoms, cognition, and self-assessment. J Exp Psychopathol. (2019) 10:2043808719865693. doi: 10.1177/2043808719865693
10. Galderisi S, Rucci P, Mucci A, Rossi A, Rocca P, Bertolino A, et al. The interplay among psychopathology, personal resources, context-related factors and real-life functioning in schizophrenia: stability in relationships after 4 years and differences in network structure between recovered and non-recovered patients. World Psychiatry. (2020) 19:81–91. doi: 10.1002/wps.20700
11. Mucci A, Galderisi S, Gibertoni D, Rossi A, Rocca P, Bertolino A, et al. Factors associated with real-life functioning in persons with schizophrenia in a 4-year follow-up study of the Italian Network for Research on Psychoses. JAMA Psychiatry. (2021) e204614. doi: 10.1001/jamapsychiatry.2020.4614. [Epub ahead of print].
12. Galderisi S, Mucci A, Bitter I, Libiger J, Bucci P, Fleischhacker WW, et al. Persistent negative symptoms in first episode patients with schizophrenia: results from the European First Episode Schizophrenia Trial. Euro Neuropsychopharmacol. (2013) 23:196–204. doi: 10.1016/j.euroneuro.2012.04.019
13. Lieberman JA, Small SA, Girgis RR. Early detection and preventive intervention in schizophrenia: from fantasy to reality. Am J Psychiatry. (2019) 176:794–810. doi: 10.1176/appi.ajp.2019.19080865
14. Yung AR, McGorry PD. The prodromal phase of first-episode psychosis: past and current conceptualizations. Schizophrenia Bull. (1996) 22:353–70. doi: 10.1093/schbul/22.2.353
15. Kuharic DB, Kekin I, Hew J, Kuzman MR, Puljak L. Preventive treatments in patients at high risk of psychosis. Lancet Psychiatry. (2020) 7:384–5. doi: 10.1016/S2215-0366(20)30100-0
16. Bosnjak Kuharic D, Kekin I, Hew J, Rojnic Kuzman M, Puljak L. Interventions for prodromal stage of psychosis. Cochrane Database Syst Rev. (2019) 2019:CD012236. doi: 10.1002/14651858.CD012236.pub2
17. Fusar-Poli P, Deste G, Smieskova R, Barlati S, Yung AR, Howes O, et al. Cognitive functioning in prodromal psychosis: a meta-analysis. Arch General Psychiatry. (2012) 69:562–71. doi: 10.1001/archgenpsychiatry.2011.1592
18. Fusar-Poli P, Borgwardt S, Bechdolf A, Addington J, Riecher-Rössler A, Schultze-Lutter F, et al. The psychosis high-risk state: a comprehensive state-of-the-art review. JAMA Psychiatry. (2013) 70:107–20. doi: 10.1001/jamapsychiatry.2013.269
19. Dondé C, Laprévote V, Lavallé L, Haesebaert F, Fakra E, Brunelin J. Cognitive insight in individuals with an at-risk mental state for psychosis: a meta-analysis. Early Intervent Psychiatry. (2020) 1–8. doi: 10.1111/eip.12993
20. Yung AR, Stanford C, Cosgrave E, Killackey E, Phillips L, Nelson B, et al. Testing the Ultra High Risk (prodromal) criteria for the prediction of psychosis in a clinical sample of young people. Schizophrenia Res. (2006) 84:57–66. doi: 10.1016/j.schres.2006.03.014
21. Yung AR, Phillips LJ, McGorry PD, McFarlane CA, Francey S, Harrigan S, et al. Prediction of psychosis. A step towards indicated prevention of schizophrenia. Br J Psychiatry Supplement. (1998) 172:14–20. doi: 10.1192/S0007125000297602
22. Nelson B, Yuen HP, Wood SJ, Lin A, Spiliotacopoulos D, Bruxner A, et al. Long-term follow-up of a group at ultra high risk (“prodromal”) for psychosis: the PACE 400 study. JAMA Psychiatry. (2013) 70:793–802. doi: 10.1001/jamapsychiatry.2013.1270
23. Lin A, Yung AR, Nelson B, Brewer WJ, Riley R, Simmons M, et al. Neurocognitive predictors of transition to psychosis: medium- to long-term findings from a sample at ultra-high risk for psychosis. Psychol Med. (2013) 43:2349–60. doi: 10.1017/S0033291713000123
24. Kempton MJ, Bonoldi I, Valmaggia L, McGuire P, Fusar-Poli P. Speed of psychosis progression in people at ultra-high clinical risk: a complementary meta-analysis. JAMA Psychiatry. (2015) 72:622–3. doi: 10.1001/jamapsychiatry.2015.0094
25. Begré S, Koenig T. Cerebral disconnectivity: an early event in schizophrenia. Neuroscientist. (2008) 14:19–45. doi: 10.1177/1073858406298391
26. Breitborde NJK, Srihari VH, Woods SW. Review of the operational definition for first-episode psychosis. Early Intervent Psychiatry. (2009) 3:259–65. doi: 10.1111/j.1751-7893.2009.00148.x
27. Radua J, Borgwardt S, Crescini A, Mataix-Cols D, Meyer-Lindenberg A, McGuire PK, et al. Multimodal meta-analysis of structural and functional brain changes in first episode psychosis and the effects of antipsychotic medication. Neurosci Biobehav Rev. (2012) 36:2325–33. doi: 10.1016/j.neubiorev.2012.07.012
28. Smieskova R, Fusar-Poli P, Allen P, Bendfeldt K, Stieglitz RD, Drewe J, et al. Neuroimaging predictors of transition to psychosis–a systematic review and meta-analysis. Neurosci Biobehav Rev. (2010) 34:1207–22. doi: 10.1016/j.neubiorev.2010.01.016
29. Boutros NN, Arfken C, Galderisi S, Warrick J, Pratt G, Iacono W. The status of spectral EEG abnormality as a diagnostic test for schizophrenia. Schizophrenia Res. (2008) 99:225–37. doi: 10.1016/j.schres.2007.11.020
30. Koukkou M, Koenig T, Bänninger A, Rieger K, Diaz Hernandez L, Higuchi Y, et al. Neurobiology of schizophrenia: electrophysiological indices. In: Javed A, Fountoulakis KN, editors. Advances in Psychiatry. Cham: Springer International Publishing (2019). p. 433–59.
31. Başar E, Schmiedt-Fehr C, Mathes B, Femir B, Emek-Savaş DD, Tülay E, et al. What does the broken brain say to the neuroscientist? Oscillations and connectivity in schizophrenia, Alzheimer's disease, and bipolar disorder. Int J Psychophysiol. (2016) 103:135–48. doi: 10.1016/j.ijpsycho.2015.02.004
32. Helfrich RF, Knight RT. Cognitive neurophysiology: event-related potentials. Handb Clin Neurol. (2019) 160:543–58. doi: 10.1016/B978-0-444-64032-1.00036-9
33. Galderisi S, Mucci A, Volpe U, Boutros N. Evidence-based medicine and electrophysiology in schizophrenia. Clin EEG Neurosci. (2009) 40:62–77. doi: 10.1177/155005940904000206
34. Michel CM, Brunet D. EEG source imaging: a practical review of the analysis steps. Front Neurol. (2019) 10:325. doi: 10.3389/fneur.2019.00325
35. Pascual-Marqui RD. Standardized low-resolution brain electromagnetic tomography (sLORETA): technical details. Methods Findings Exp Clin Pharmacol. (2002) 24(Suppl. D):5–12.
36. Schomer DL, Lopes da Silva FH. Niedermeyer's Electroencephalography Basic Principles, Clinical Applications, and Related Fields: Basic Principles, Clinical Applications, and Related Fields. Oxford: Oxford University Press (2017).
37. Lehmann D, Skrandies W. Reference-free identification of components of checkerboard-evoked multichannel potential fields. Electroencephalogr Clin Neurophysiol. (1980) 48:609–21. doi: 10.1016/0013-4694(80)90419-8
38. Lehmann D, Strik WK, Henggeler B, Koenig T, Koukkou M. Brain electric microstates and momentary conscious mind states as building blocks of spontaneous thinking: I. Visual imagery and abstract thoughts. Int J Psychophysiol. (1998) 29:1–11. doi: 10.1016/S0167-8760(97)00098-6
39. Pascual-Marqui RD, Michel CM, Lehmann D. Segmentation of brain electrical activity into microstates: model estimation and validation. IEEE Trans Bio Med Eng. (1995) 42:658–65. doi: 10.1109/10.391164
40. Wackermann J, Lehmann D, Michel CM, Strik WK. Adaptive segmentation of spontaneous EEG map series into spatially defined microstates. Int J Psychophysiol. (1993) 14:269–83. doi: 10.1016/0167-8760(93)90041-M
41. Hajcak G, Klawohn J, Meyer A. The utility of event-related potentials in clinical psychology. Annu Rev Clin Psychol. (2019) 15:71–95. doi: 10.1146/annurev-clinpsy-050718-095457
42. van der Stelt O, Belger A. Application of electroencephalography to the study of cognitive and brain functions in schizophrenia. Schizophrenia Bull. (2007) 33:955–70. doi: 10.1093/schbul/sbm016
43. Newson JJ, Thiagarajan TC. EEG frequency bands in psychiatric disorders: a review of resting state studies. Front Hum Neurosci. (2018) 12:521. doi: 10.3389/fnhum.2018.00521
44. Atagun MI, Drukker M, Hall MH, Altun IK, Tatli SZ, Guloksuz S, et al. Meta-analysis of auditory P50 sensory gating in schizophrenia and bipolar disorder. Psychiatry Res Neuroimaging. (2020) 300:111078. doi: 10.1016/j.pscychresns.2020.111078
45. Kambeitz J, Kambeitz-Ilankovic L, Leucht S, Wood S, Davatzikos C, Malchow B, et al. Detecting neuroimaging biomarkers for schizophrenia: a meta-analysis of multivariate pattern recognition studies. Neuropsychopharmacology. (2015) 40:1742–51. doi: 10.1038/npp.2015.22
46. Rieger K, Diaz Hernandez L, Baenninger A, Koenig T. 15 years of microstate research in schizophrenia - where are we? A meta-analysis. Front Psychiatry. (2016) 7:22. doi: 10.3389/fpsyt.2016.00022
47. Giordano GM, Koenig T, Mucci A, Vignapiano A, Amodio A, Di Lorenzo G, et al. Neurophysiological correlates of Avolition-apathy in schizophrenia: a resting-EEG microstates study. NeuroImage Clin. (2018) 20:627–36. doi: 10.1016/j.nicl.2018.08.031
48. Vignapiano A, Koenig T, Mucci A, Giordano GM, Amodio A, Altamura M, et al. Disorganization and cognitive impairment in schizophrenia: new insights from electrophysiological findings. Int J Psychophysiol. (2019) 145:99–108. doi: 10.1016/j.ijpsycho.2019.03.008
49. Javitt DC, Siegel SJ, Spencer KM, Mathalon DH, Hong LE, Martinez A, et al. A roadmap for development of neuro-oscillations as translational biomarkers for treatment development in neuropsychopharmacology. Neuropsychopharmacology. (2020) 45:1411–22. doi: 10.1038/s41386-020-0697-9
50. Mitra S, Nizamie SH, Goyal N, Tikka SK. Electroencephalogram alpha-to-theta ratio over left fronto-temporal region correlates with negative symptoms in schizophrenia. Asian J Psychiatry. (2017) 26:70–6. doi: 10.1016/j.ajp.2017.01.013
51. Angelopoulos E, Koutsoukos E, Maillis A, Papadimitriou GN, Stefanis C. Cortical interactions during the experience of auditory verbal hallucinations. J Neuropsychiatry Clin Neurosci. (2011) 23:287–93. doi: 10.1176/jnp.23.3.jnp287
52. Martínez A, Gaspar PA, Hillyard SA, Bickel S, Lakatos P, Dias EC, et al. Neural oscillatory deficits in schizophrenia predict behavioral and neurocognitive impairments. Front Hum Neurosci. (2015) 9:371. doi: 10.3389/fnhum.2015.00371
53. Hasey GM, Kiang M. A review of recent literature employing electroencephalographic techniques to study the pathophysiology, phenomenology, and treatment response of schizophrenia. Curr Psychiatry Rep. (2013) 15:388. doi: 10.1007/s11920-013-0388-x
54. Koenig T, Lehmann D, Merlo MC, Kochi K, Hell D, Koukkou M. A deviant EEG brain microstate in acute, neuroleptic-naive schizophrenics at rest. Euro Arch Psychiatry Clin Neurosci. (1999) 249:205–11. doi: 10.1007/s004060050088
55. Thoma RJ, Meier A, Houck J, Clark VP, Lewine JD, Turner J, et al. Diminished auditory sensory gating during active auditory verbal hallucinations. Schizophrenia Res. (2017) 188:125–31. doi: 10.1016/j.schres.2017.01.023
56. Boutros NN, Brockhaus-Dumke A, Gjini K, Vedeniapin A, Elfakhani M, Burroughs S, et al. Sensory-gating deficit of the N100 mid-latency auditory evoked potential in medicated schizophrenia patients. Schizophrenia Res. (2009) 113:339–46. doi: 10.1016/j.schres.2009.05.019
57. Galderisi S, Vignapiano A, Mucci A, Boutros NN. Physiological correlates of positive symptoms in schizophrenia. Curr Topics Behav Neurosci. (2014) 21:103–28. doi: 10.1007/7854_2014_322
58. Smith AK, Edgar JC, Huang M, Lu BY, Thoma RJ, Hanlon FM, et al. Cognitive abilities and 50- and 100-msec paired-click processes in schizophrenia. Am J Psychiatry. (2010) 167:1264–75. doi: 10.1176/appi.ajp.2010.09071059
59. Perrin MA, Kantrowitz JT, Silipo G, Dias E, Jabado O, Javitt DC. Mismatch negativity (MMN) to spatial deviants and behavioral spatial discrimination ability in the etiology of auditory verbal hallucinations and thought disorder in schizophrenia. Schizophrenia Res. (2018) 191:140–7. doi: 10.1016/j.schres.2017.05.012
60. Kim JS, Kwon YJ, Lee HY, Lee HS, Kim S, Shim SH. Mismatch negativity indices as a prognostic factor for remission in schizophrenia. Clin Psychopharmacol Neurosci. (2020) 18:127–35. doi: 10.9758/cpn.2020.18.1.127
61. Nieman DH, Koelman JH, Linszen DH, Bour LJ, Dingemans PM, Ongerboer de Visser BW. Clinical and neuropsychological correlates of the P300 in schizophrenia. Schizophrenia Res. (2002) 55:105–13. doi: 10.1016/S0920-9964(01)00184-0
62. Reilly TJ, Nottage JF, Studerus E, Rutigliano G, Micheli AI, Fusar-Poli P, et al. Gamma band oscillations in the early phase of psychosis: a systematic review. Neurosci Biobehav Rev. (2018) 90:381–99. doi: 10.1016/j.neubiorev.2018.04.006
63. Lepock JR, Mizrahi R, Korostil M, Bagby RM, Pang EW, Kiang M. Event-related potentials in the clinical high-risk (CHR) state for psychosis: a systematic review. Clin EEG Neurosci. (2018) 49:215–25. doi: 10.1177/1550059418755212
64. Tada M, Kirihara K, Mizutani S, Uka T, Kunii N, Koshiyama D, et al. Mismatch negativity (MMN) as a tool for translational investigations into early psychosis: a review. Int J Psychophysiol. (2019) 145:5–14. doi: 10.1016/j.ijpsycho.2019.02.009
65. Haigh SM, Coffman BA, Salisbury DF. Mismatch negativity in first-episode schizophrenia: a meta-analysis. Clin EEG Neurosci. (2017) 48:3–10. doi: 10.1177/1550059416645980
66. Nagai T, Kirihara K, Tada M, Koshiyama D, Koike S, Suga M, et al. Reduced mismatch negativity is associated with increased plasma level of glutamate in first-episode psychosis. Sci Rep. (2017) 7:2258. doi: 10.1038/s41598-017-02267-1
67. Qiu YQ, Tang YX, Chan RC, Sun XY, He J. P300 aberration in first-episode schizophrenia patients: a meta-analysis. PLoS ONE. (2014) 9:e97794. doi: 10.1371/journal.pone.0097794
68. Hamilton HK, Boos AK, Mathalon DH. Electroencephalography and event-related potential biomarkers in individuals at clinical high risk for psychosis. Biol Psychiatry. (2020) 88:294–303. doi: 10.1016/j.biopsych.2020.04.002
69. Moher D, Liberati A, Tetzlaff J, Altman DG, The PG. Preferred reporting items for systematic reviews and meta-analyses: the PRISMA statement. PLoS Med. (2009) 6:e1000097. doi: 10.1371/journal.pmed.1000097
70. Babiloni C, Barry RJ, Başar E, Blinowska KJ, Cichocki A, Drinkenburg WHIM, et al. International Federation of Clinical Neurophysiology (IFCN) – EEG research workgroup: recommendations on frequency and topographic analysis of resting state EEG rhythms. Part 1: Applications in clinical research studies. Clin Neurophysiol. (2020) 131:285–307. doi: 10.1016/j.clinph.2019.06.234
71. Harmony T. The functional significance of delta oscillations in cognitive processing. Front Integr Neurosci. (2013) 7:83. doi: 10.3389/fnint.2013.00083
72. Güntekin B, Başar E. Review of evoked and event-related delta responses in the human brain. Int J Psychophysiol. (2016) 103:43–52. doi: 10.1016/j.ijpsycho.2015.02.001
73. Begré S, Federspiel A, Kiefer C, Schroth G, Dierks T, Strik WK. Reduced hippocampal anisotropy related to anteriorization of alpha EEG in schizophrenia. NeuroReport. (2003) 14:739–42. doi: 10.1097/00001756-200304150-00016
74. Koenig T, Lehmann D, Saito N, Kuginuki T, Kinoshita T, Koukkou M. Decreased functional connectivity of EEG theta-frequency activity in first-episode, neuroleptic-naïve patients with schizophrenia: preliminary results. Schizophrenia Res. (2001) 50:55–60. doi: 10.1016/S0920-9964(00)00154-7
75. Clementz BA, Sponheim SR, Iacono WG, Beiser M. Resting EEG in first-episode schizophrenia patients, bipolar psychosis patients, and their first-degree relatives. Psychophysiology. (1994) 31:486–94. doi: 10.1111/j.1469-8986.1994.tb01052.x
76. Renaldi R, Kim M, Lee TH, Kwak YB, Tanra AJ, Kwon JS. Predicting symptomatic and functional improvements over 1 year in patients with first-episode psychosis using resting-state electroencephalography. Psychiatry Investig. (2019) 16:695–703. doi: 10.30773/pi.2019.06.20.1
77. Harris A, Melkonian D, Williams L, Gordon E. Dynamic spectral analysis findings in first episode and chronic schizophrenia. Int J Neurosci. (2006) 116:223–46. doi: 10.1080/00207450500402977
78. Pascual-Marqui RD, Lehmann D, Koenig T, Kochi K, Merlo MC, Hell D, et al. Low resolution brain electromagnetic tomography (LORETA) functional imaging in acute, neuroleptic-naive, first-episode, productive schizophrenia. Psychiatry Res. (1999) 90:169–79. doi: 10.1016/S0925-4927(99)00013-X
79. van Tricht MJ, Ruhrmann S, Arns M, Müller R, Bodatsch M, Velthorst E, et al. Can quantitative EEG measures predict clinical outcome in subjects at Clinical High Risk for psychosis? A prospective multicenter study. Schizophrenia Res. (2014) 153:42–7. doi: 10.1016/j.schres.2014.01.019
80. Krukow P, Jonak K, Grochowski C, Plechawska-Wójcik M, Karakuła-Juchnowicz H. Resting-state hyperconnectivity within the default mode network impedes the ability to initiate cognitive performance in first-episode schizophrenia patients. Progress Neuro Psychopharmacol Biol Psychiatry. (2020) 102:109959. doi: 10.1016/j.pnpbp.2020.109959
81. Missonnier P, Prévot A, Herrmann FR, Ventura J, Padée A, Merlo MCG. Disruption of gamma-delta relationship related to working memory deficits in first-episode psychosis. J Neural Transmission. (2020) 127:103–15. doi: 10.1007/s00702-019-02126-5
82. Alexander DM, Flynn GJ, Wong W, Whitford TJ, Harris AW, Galletly CA, et al. Spatio-temporal EEG waves in first episode schizophrenia. Clin Neurophysiol. (2009) 120:1667–82. doi: 10.1016/j.clinph.2009.06.020
83. Zhao Z, Cheng Y, Li Z, Yu Y. Altered small-world networks in first-episode schizophrenia patients during cool executive function task. Behav Neurol. (2018) 2018:2191208. doi: 10.1155/2018/2191208
84. Cavanagh JF, Frank MJ. Frontal theta as a mechanism for cognitive control. Trends Cogn Sci. (2014) 18:414–21. doi: 10.1016/j.tics.2014.04.012
85. Hunt MJ, Kopell NJ, Traub RD, Whittington MA. Aberrant network activity in schizophrenia. Trends Neurosci. (2017) 40:371–82. doi: 10.1016/j.tins.2017.04.003
86. Adams RA, Bush D, Zheng F, Meyer SS, Kaplan R, Orfanos S, et al. Impaired theta phase coupling underlies frontotemporal dysconnectivity in schizophrenia. Brain. (2020) 143:1261–77. doi: 10.1093/brain/awaa035
87. Ranlund S, Nottage J, Shaikh M, Dutt A, Constante M, Walshe M, et al. Resting EEG in psychosis and at-risk populations–a possible endophenotype? Schizophrenia Res. (2014) 153:96–102. doi: 10.1016/j.schres.2013.12.017
88. Garakh Z, Zaytseva Y, Kapranova A, Fiala O, Horacek J, Shmukler A, et al. EEG correlates of a mental arithmetic task in patients with first episode schizophrenia and schizoaffective disorder. Clin Neurophysiol. (2015) 126:2090–8. doi: 10.1016/j.clinph.2014.12.031
89. Ramyead A, Studerus E, Kometer M, Heitz U, Gschwandtner U, Fuhr P, et al. Neural oscillations in antipsychotic-naïve patients with a first psychotic episode. World J Biol Psychiatry. (2016) 17:296–307. doi: 10.3109/15622975.2016.1156742
90. Lee TH, Kim M, Hwang WJ, Kim T, Kwak YB, Kwon JS. Relationship between resting-state theta phase-gamma amplitude coupling and neurocognitive functioning in patients with first-episode psychosis. Schizophrenia Res. (2020) 216:154–60. doi: 10.1016/j.schres.2019.12.010
91. Andreou C, Nolte G, Leicht G, Polomac N, Hanganu-Opatz IL, Lambert M, et al. Increased resting-state gamma-band connectivity in first-episode schizophrenia. Schizophrenia Bull. (2015) 41:930–9. doi: 10.1093/schbul/sbu121
92. Andreou C, Leicht G, Nolte G, Polomac N, Moritz S, Karow A, et al. Resting-state theta-band connectivity and verbal memory in schizophrenia and in the high-risk state. Schizophrenia Res. (2015) 161:299–307. doi: 10.1016/j.schres.2014.12.018
93. Krukow P, Jonak K, Karakuła-Juchnowicz H, Podkowiński A, Jonak K, Borys M, et al. Disturbed functional connectivity within the left prefrontal cortex and sensorimotor areas predicts impaired cognitive speed in patients with first-episode schizophrenia. Psychiatry Res Neuroimaging. (2018) 275:28–35. doi: 10.1016/j.pscychresns.2018.03.001
94. Boudewyn MA, Carter CS. Electrophysiological correlates of adaptive control and attentional engagement in patients with first episode schizophrenia and healthy young adults. Psychophysiology. (2018) 55:e12820. doi: 10.1111/psyp.12820
95. Xiong YB, Bo QJ, Wang CM, Tian Q, Liu Y, Wang CY. Differential of frequency and duration mismatch negativity and theta power deficits in first-episode and chronic schizophrenia. Front Behav Neurosci. (2019) 13:37. doi: 10.3389/fnbeh.2019.00037
96. Jhung K, Cho SH, Jang JH, Park JY, Shin D, Kim KR, et al. Small-world networks in individuals at ultra-high risk for psychosis and first-episode schizophrenia during a working memory task. Neurosci Lett. (2013) 535:35–9. doi: 10.1016/j.neulet.2012.11.051
97. Klimesch W. Alpha-band oscillations, attention, and controlled access to stored information. Trends Cogn Sci. (2012) 16:606–17. doi: 10.1016/j.tics.2012.10.007
98. Bazanova OM, Vernon D. Interpreting EEG alpha activity. Neurosci Biobehav Rev. (2014) 44:94–110. doi: 10.1016/j.neubiorev.2013.05.007
99. Başar E, Güntekin B. Chapter 19 - review of delta, theta, alpha, beta, and gamma response oscillations in neuropsychiatric disorders. In: Başar E, Başar-Eroglu C, Özerdem A, Rossini PM, Yener GG, editors. Supplements to Clinical Neurophysiology, Vol. 62. Amsterdam: Elsevier (2013). p. 303–41.
100. Murphy M, Öngür D. Decreased peak alpha frequency and impaired visual evoked potentials in first episode psychosis. NeuroImage Clin. (2019) 22:101693. doi: 10.1016/j.nicl.2019.101693
101. Zaytseva Y, Garakh Z, Novototsky-Vlasov V, Gurovich IY, Shmukler A, Papaefstathiou A, et al. EEG coherence in a mental arithmetic task performance in first episode schizophrenia and schizoaffective disorder. Clin Neurophysiol. (2018) 129:2315–24. doi: 10.1016/j.clinph.2018.08.020
102. Liu T, Zhang J, Dong X, Li Z, Shi X, Tong Y, et al. Occipital alpha connectivity during resting-state electroencephalography in patients with ultra-high risk for psychosis and schizophrenia. Front Psychiatry. (2019) 10:553. doi: 10.3389/fpsyt.2019.00553
103. Kayser J, Tenke CE, Kroppmann CJ, Alschuler DM, Fekri S, Ben-David S, et al. Auditory event-related potentials and α oscillations in the psychosis prodrome: neuronal generator patterns during a novelty oddball task. Int J Psychophysiol. (2014) 91:104–20. doi: 10.1016/j.ijpsycho.2013.12.003
104. Spitzer B, Haegens S. Beyond the status quo: a role for beta oscillations in endogenous content (re)activation. eNeuro. (2017) 4:ENEURO.0170-17.2017. doi: 10.1523/ENEURO.0170-17.2017
105. Brinkman L, Stolk A, Dijkerman HC, de Lange FP, Toni I. Distinct roles for alpha- and beta-band oscillations during mental simulation of goal-directed actions. J Neurosci. (2014) 34:14783–92. doi: 10.1523/JNEUROSCI.2039-14.2014
106. Venables NC, Bernat EM, Sponheim SR. Genetic and disorder-specific aspects of resting state EEG abnormalities in schizophrenia. Schizophrenia Bull. (2009) 35:826–39. doi: 10.1093/schbul/sbn021
107. Narayanan B, O'Neil K, Berwise C, Stevens MC, Calhoun VD, Clementz BA, et al. Resting state electroencephalogram oscillatory abnormalities in schizophrenia and psychotic bipolar patients and their relatives from the bipolar and schizophrenia network on intermediate phenotypes study. Biol Psychiatry. (2014) 76:456–65. doi: 10.1016/j.biopsych.2013.12.008
108. Liddle EB, Price D, Palaniyappan L, Brookes MJ, Robson SE, Hall EL, et al. Abnormal salience signaling in schizophrenia: the role of integrative beta oscillations. Hum Brain Mapp. (2016) 37:1361–74. doi: 10.1002/hbm.23107
109. Yeragani VK, Cashmere D, Miewald J, Tancer M, Keshavan MS. Decreased coherence in higher frequency ranges (beta and gamma) between central and frontal EEG in patients with schizophrenia: a preliminary report. Psychiatry Res. (2006) 141:53–60. doi: 10.1016/j.psychres.2005.07.016
110. Bosman CA, Lansink CS, Pennartz CMA. Functions of gamma-band synchronization in cognition: from single circuits to functional diversity across cortical and subcortical systems. Euro J Neurosci. (2014) 39:1982–99. doi: 10.1111/ejn.12606
111. McNally JM, McCarley RW. Gamma band oscillations: a key to understanding schizophrenia symptoms and neural circuit abnormalities. Curr Opin Psychiatry. (2016) 29:202–10. doi: 10.1097/YCO.0000000000000244
112. Tikka SK, Yadav S, Nizamie SH, Das B, Tikka DL, Goyal N. Schneiderian first rank symptoms and gamma oscillatory activity in neuroleptic naïve first episode schizophrenia: a 192 channel EEG study. Psychiatry Investig. (2014) 11:467–75. doi: 10.4306/pi.2014.11.4.467
113. Ramyead A, Kometer M, Studerus E, Koranyi S, Ittig S, Gschwandtner U, et al. Aberrant current source-density and lagged phase synchronization of neural oscillations as markers for emerging psychosis. Schizophrenia Bull. (2015) 41:919–29. doi: 10.1093/schbul/sbu134
114. Missonnier P, Curtis L, Ventura J, Herrmann FR, Merlo MCG. Differences of temporal dynamics and signal complexity of gamma band oscillations in first-episode psychosis during a working memory task. J Neural Transmission. (2017) 124:853–62. doi: 10.1007/s00702-017-1728-5
115. Minzenberg MJ, Firl AJ, Yoon JH, Gomes GC, Reinking C, Carter CS. Gamma oscillatory power is impaired during cognitive control independent of medication status in first-episode schizophrenia. Neuropsychopharmacology. (2010) 35:2590–9. doi: 10.1038/npp.2010.150
116. Flynn G, Alexander D, Harris A, Whitford T, Wong W, Galletly C, et al. Increased absolute magnitude of gamma synchrony in first-episode psychosis. Schizophrenia Res. (2008) 105:262–71. doi: 10.1016/j.schres.2008.05.029
117. Williams LM, Whitford TJ, Nagy M, Flynn G, Harris AW, Silverstein SM, et al. Emotion-elicited gamma synchrony in patients with first-episode schizophrenia: a neural correlate of social cognition outcomes. J Psychiatry Neurosci. (2009) 34:303–13.
118. Oribe N, Hirano Y, Del Re E, Seidman LJ, Mesholam-Gately RI, Woodberry KA, et al. Progressive reduction of auditory evoked gamma in first episode schizophrenia but not clinical high risk individuals. Schizophrenia Res. (2019) 208:145–52. doi: 10.1016/j.schres.2019.03.025
119. Slewa-Younan S, Gordon E, Harris AW, Haig AR, Brown KJ, Flor-Henry P, et al. Sex differences in functional connectivity in first-episode and chronic schizophrenia patients. Am J Psychiatry. (2004) 161:1595–602. doi: 10.1176/appi.ajp.161.9.1595
120. Symond MP, Harris AW, Gordon E, Williams LM. “Gamma synchrony” in first-episode schizophrenia: a disorder of temporal connectivity? Am J Psychiatry. (2005) 162:459–65. doi: 10.1176/appi.ajp.162.3.459
121. Williams LM, Whitford TJ, Gordon E, Gomes L, Brown KJ, Harris AWF. Neural synchrony in patients with a first episode of schizophrenia: tracking relations with grey matter and symptom profile. J Psychiatry Neurosci. (2009) 34:21–9.
122. Leicht G, Andreou C, Polomac N, Lanig C, Schöttle D, Lambert M, et al. Reduced auditory evoked gamma band response and cognitive processing deficits in first episode schizophrenia. World J Biol Psychiatry. (2015) 16:387–97. doi: 10.3109/15622975.2015.1017605
123. Perez VB, Roach BJ, Woods SW, Srihari VH, McGlashan TH, Ford JM, et al. Early auditory gamma-band responses in patients at clinical high risk for schizophrenia. Supplement Clin Neurophysiol. (2013) 62:147–62. doi: 10.1016/B978-0-7020-5307-8.00010-7
124. Leicht G, Vauth S, Polomac N, Andreou C, Rauh J, Mußmann M, et al. EEG-informed fMRI reveals a disturbed gamma-band-specific network in subjects at high risk for psychosis. Schizophrenia Bull. (2016) 42:239–49. doi: 10.1093/schbul/sbv092
125. Spencer KM, Salisbury DF, Shenton ME, McCarley RW. γ-band auditory steady-state responses are impaired in first episode psychosis. Biol Psychiatry. (2008) 64:369–75. doi: 10.1016/j.biopsych.2008.02.021
126. Tada M, Nagai T, Kirihara K, Koike S, Suga M, Araki T, et al. Differential alterations of auditory gamma oscillatory responses between pre-onset high-risk individuals and first-episode schizophrenia. Cerebral Cortex. (2016) 26:1027–35. doi: 10.1093/cercor/bhu278
127. Wang J, Tang Y, Curtin A, Chan RCK, Wang Y, Li H, et al. Abnormal auditory-evoked gamma band oscillations in first-episode schizophrenia during both eye open and eye close states. Progress Neuro Psychopharmacol Biol Psychiatry. (2018) 86:279–86. doi: 10.1016/j.pnpbp.2018.04.016
128. Lepock JR, Ahmed S, Mizrahi R, Gerritsen CJ, Maheandiran M, Drvaric L, et al. Relationships between cognitive event-related brain potential measures in patients at clinical high risk for psychosis. Schizophrenia Res. (2019) 226:84–94. doi: 10.1016/j.schres.2019.01.014
129. Koenig T, Prichep L, Lehmann D, Sosa PV, Braeker E, Kleinlogel H, et al. Millisecond by millisecond, year by year: normative EEG microstates and developmental stages. NeuroImage. (2002) 16:41–8. doi: 10.1006/nimg.2002.1070
130. Khanna A, Pascual-Leone A, Michel CM, Farzan F. Microstates in resting-state EEG: current status and future directions. Neurosci Biobehav Rev. (2015) 49:105–13. doi: 10.1016/j.neubiorev.2014.12.010
131. Britz J, Van De Ville D, Michel CM. BOLD correlates of EEG topography reveal rapid resting-state network dynamics. NeuroImage. (2010) 52:1162–70. doi: 10.1016/j.neuroimage.2010.02.052
132. Custo A, Van De Ville D, Wells WM, Tomescu MI, Brunet D, Michel CM. Electroencephalographic resting-state networks: source localization of microstates. Brain Connect. (2017) 7:671–82. doi: 10.1089/brain.2016.0476
133. da Cruz JR, Favrod O, Roinishvili M, Chkonia E, Brand A, Mohr C, et al. EEG microstates are a candidate endophenotype for schizophrenia. Nat Commun. (2020) 11:3089. doi: 10.1038/s41467-020-16914-1
134. Lehmann D, Faber PL, Galderisi S, Herrmann WM, Kinoshita T, Koukkou M, et al. EEG microstate duration and syntax in acute, medication-naive, first-episode schizophrenia: a multi-center study. Psychiatry Res. (2005) 138:141–56. doi: 10.1016/j.pscychresns.2004.05.007
135. Tomescu MI, Rihs TA, Becker R, Britz J, Custo A, Grouiller F, et al. Deviant dynamics of EEG resting state pattern in 22q11.2 deletion syndrome adolescents: a vulnerability marker of schizophrenia? Schizophrenia Res. (2014) 157:175–81. doi: 10.1016/j.schres.2014.05.036
136. Andreou C, Faber PL, Leicht G, Schoettle D, Polomac N, Hanganu-Opatz IL, et al. Resting-state connectivity in the prodromal phase of schizophrenia: insights from EEG microstates. Schizophrenia Res. (2014) 152:513–20. doi: 10.1016/j.schres.2013.12.008
137. Adler LE, Pachtman E, Franks RD, Pecevich M, Waldo MC, Freedman R. Neurophysiological evidence for a defect in neuronal mechanisms involved in sensory gating in schizophrenia. Biol Psychiatry. (1982) 17:639–54.
138. Boutros NN, Zouridakis G, Overall J. Replication and extension of P50 findings in schizophrenia. Clin EEG. (1991) 22:40–5. doi: 10.1177/155005949102200109
139. Clementz BA, Geyer MA, Braff DL. Multiple site evaluation of P50 suppression among schizophrenia and normal comparison subjects. Schizophrenia Res. (1998) 30:71–80. doi: 10.1016/S0920-9964(97)00122-9
140. Light GA, Geyer MA, Clementz BA, Cadenhead KS, Braff DL. Normal P50 suppression in schizophrenia patients treated with atypical antipsychotic medications. Am J Psychiatry. (2000) 157:767–71. doi: 10.1176/appi.ajp.157.5.767
141. Bramon E, Rabe-Hesketh S, Sham P, Murray RM, Frangou S. Meta-analysis of the P300 and P50 waveforms in schizophrenia. Schizophrenia Res. (2004) 70:315–29. doi: 10.1016/j.schres.2004.01.004
142. Ritsner MS. The Handbook of Neuropsychiatric Biomarkers, Endophenotypes, and Genes. Dordrecht: Springer (2009).
143. Brockhaus-Dumke A, Schultze-Lutter F, Mueller R, Tendolkar I, Bechdolf A, Pukrop R, et al. Sensory gating in schizophrenia: P50 and N100 gating in antipsychotic-free subjects at risk, first-episode, and chronic patients. Biol Psychiatry. (2008) 64:376–84. doi: 10.1016/j.biopsych.2008.02.006
144. Hong X, Chan RCK, Zhuang X, Jiang T, Wan X, Wang J, et al. Neuroleptic effects on P50 sensory gating in patients with first-episode never-medicated schizophrenia. Schizophrenia Res. (2009) 108:151–7. doi: 10.1016/j.schres.2008.11.016
145. Oranje B, Aggernaes B, Rasmussen H, Ebdrup BH, Glenthøj BY. P50 suppression and its neural generators in antipsychotic-naive first-episode schizophrenia before and after 6 months of quetiapine treatment. Schizophrenia Bull. (2013) 39:472–80. doi: 10.1093/schbul/sbr183
146. Song L, Chen X, Chen M, Tang Y, Wang J, Zhang M, et al. Differences in P50 and prepulse inhibition of the startle reflex between male smokers and non-smokers with first episode schizophrenia without medical treatment. Chin Med J. (2014) 127:1651–5. doi: 10.3760/cma.j.issn.0366-6999.20133306
147. Devrim-Uçok M, Keskin-Ergen HY, Uçok A. P50 gating at acute and post-acute phases of first-episode schizophrenia. Progress Neuro Psychopharmacol Biol Psychiatry. (2008) 32:1952–6. doi: 10.1016/j.pnpbp.2008.09.018
148. Luo Y, Zhang J, Wang C, Zhao X, Chang Q, Wang H, et al. Discriminating schizophrenia disease progression using a P50 sensory gating task with dense-array EEG, clinical assessments, and cognitive tests. Expert Rev Neurother. (2019) 19:459–70. doi: 10.1080/14737175.2019.1601558
149. Myles-Worsley M, Ord L, Blailes F, Ngiralmau H, Freedman R. P50 sensory gating in adolescents from a pacific island isolate with elevated risk for schizophrenia. Biol Psychiatry. (2004) 55:663–7. doi: 10.1016/j.biopsych.2003.12.006
150. Hsieh MH, Lin YT, Chien YL, Hwang TJ, Hwu HG, Liu CM, et al. Auditory event-related potentials in antipsychotic-free subjects with ultra-high-risk state and first-episode psychosis. Front Psychiatry. (2019) 10:223. doi: 10.3389/fpsyt.2019.00223
151. Hsieh MH, Shan JC, Huang WL, Cheng WC, Chiu MJ, Jaw FS, et al. Auditory event-related potential of subjects with suspected pre-psychotic state and first-episode psychosis. Schizophrenia Res. (2012) 140:243–9. doi: 10.1016/j.schres.2012.06.021
152. Morales-Muñoz I, Jurado-Barba R, Fernández-Guinea S, Rodríguez-Jiménez R, Jiménez-Arriero M, Criado JR, et al. Sensory gating deficits in first-episode psychosis: evidence from neurophysiology, psychophysiology, and neuropsychology. J Nervous Mental Dis. (2016) 204:877–84. doi: 10.1097/NMD.0000000000000572
153. Yee CM, Williams TJ, White PM, Nuechterlein KH, Ames D, Subotnik KL. Attentional modulation of the P50 suppression deficit in recent-onset and chronic schizophrenia. J Abnormal Psychol. (2010) 119:31–9. doi: 10.1037/a0018265
154. Chang Q, Liu M, Tian Q, Wang H, Luo Y, Zhang J, et al. EEG-based brain functional connectivity in first-episode schizophrenia patients, ultra-high-risk individuals, and healthy controls during p50 suppression. Front Hum Neurosci. (2019) 13:379. doi: 10.3389/fnhum.2019.00379
155. Düring S, Glenthøj BY, Andersen GS, Oranje B. Effects of dopamine D2/D3 blockade on human sensory and sensorimotor gating in initially antipsychotic-naive, first-episode schizophrenia patients. Neuropsychopharmacology. (2014) 39:3000–8. doi: 10.1038/npp.2014.152
156. van Tricht MJ, Nieman DH, Koelman JT, Mensink AJ, Bour LJ, van der Meer JN, et al. Sensory gating in subjects at ultra high risk for developing a psychosis before and after a first psychotic episode. World J Biol Psychiatry. (2015) 16:12–21. doi: 10.3109/15622975.2012.680911
157. Ziermans TB, Schothorst PF, Sprong M, Magnee MJ, van Engeland H, Kemner C. Reduced prepulse inhibition as an early vulnerability marker of the psychosis prodrome in adolescence. Schizophrenia Res. (2012) 134:10–5. doi: 10.1016/j.schres.2011.10.009
158. Ebdrup BH, Axelsen MC, Bak N, Fagerlund B, Oranje B, Raghava JM, et al. Accuracy of diagnostic classification algorithms using cognitive-, electrophysiological-, and neuroanatomical data in antipsychotic-naïve schizophrenia patients. Psychol Med. (2019) 49:2754–63. doi: 10.1017/S0033291718003781
159. Rosburg T, Boutros NN, Ford JM. Reduced auditory evoked potential component N100 in schizophrenia — a critical review. Psychiatry Res. (2008) 161:259–74. doi: 10.1016/j.psychres.2008.03.017
160. Mucci A, Galderisi S, Kirkpatrick B, Bucci P, Volpe U, Merlotti E, et al. Double dissociation of N1 and P3 abnormalities in deficit and nondeficit schizophrenia. Schizophrenia Res. (2007) 92:252–61. doi: 10.1016/j.schres.2007.01.026
161. Oribe N, Hirano Y, Kanba S, del Re EC, Seidman LJ, Mesholam-Gately R, et al. Early and late stages of visual processing in individuals in prodromal state and first episode schizophrenia: an ERP study. Schizophrenia Res. (2013) 146:95–102. doi: 10.1016/j.schres.2013.01.015
162. Brown KJ, Gonsalvez CJ, Harris AW, Williams LM, Gordon E. Target and non-target ERP disturbances in first episode vs. chronic schizophrenia. Clin Neurophysiol. (2002) 113:1754–63. doi: 10.1016/S1388-2457(02)00290-0
163. del Re EC, Spencer KM, Oribe N, Mesholam-Gately RI, Goldstein J, Shenton ME, et al. Clinical high risk and first episode schizophrenia: auditory event-related potentials. Psychiatry Res. (2015) 231:126–33. doi: 10.1016/j.pscychresns.2014.11.012
164. Salisbury DF, Kohler J, Shenton ME, McCarley RW. Deficit effect sizes and correlations of auditory event-related potentials at first hospitalization in the schizophrenia spectrum. Clin EEG Neurosci. (2020) 51:198–206. doi: 10.1177/1550059419868115
165. Lee SY, Namkoong K, Cho HH, Song DH, An SK. Reduced visual P300 amplitudes in individuals at ultra-high risk for psychosis and first-episode schizophrenia. Neurosci Lett. (2010) 486:156–60. doi: 10.1016/j.neulet.2010.09.035
166. Monaghan CK, Brickman S, Huynh P, Öngür D, Hall MH. A longitudinal study of event related potentials and correlations with psychosocial functioning and clinical features in first episode psychosis patients. Int J Psychophysiol. (2019) 145:48–56. doi: 10.1016/j.ijpsycho.2019.05.007
167. Valkonen-Korhonen M, Tarkka IM, Pääkkönen A, Purhonen M. Auditory event-related potentials in patients with psychotic illness: a 5-year follow-up. Neurocase. (2012) 18:392–9. doi: 10.1080/13554794.2011.627337
168. Bramon E, Shaikh M, Broome M, Lappin J, Berge D, Day F, et al. Abnormal P300 in people with high risk of developing psychosis. NeuroImage. (2008) 41:553–60. doi: 10.1016/j.neuroimage.2007.12.038
169. Frommann I, Brinkmeyer J, Ruhrmann S, Hack E, Brockhaus-Dumke A, Bechdolf A, et al. Auditory P300 in individuals clinically at risk for psychosis. Int J Psychophysiol. (2008) 70:192–205. doi: 10.1016/j.ijpsycho.2008.07.003
170. Oribe N, Hirano Y, Del Re E, Mesholam-Gately RI, Woodberry KA, Ueno T, et al. Longitudinal evaluation of visual P300 amplitude in clinical high risk subjects: an ERP study. Psychiatry Clin Neurosci. (2020) 74:527–34. doi: 10.1111/pcn.13083
171. van Tricht MJ, Nieman DH, Koelman JH, van der Meer JN, Bour LJ, de Haan L, et al. Reduced parietal P300 amplitude is associated with an increased risk for a first psychotic episode. Biol Psychiatry. (2010) 68:642–8. doi: 10.1016/j.biopsych.2010.04.022
172. Xu Y, Chai H, Zhang B, Gao Q, Fan H, Zheng L, et al. Event-related potentials elicited by the Deutsch “high-low” word illusion in the patients with first-episode schizophrenia with auditory hallucinations. BMC Psychiatry. (2016) 16:33. doi: 10.1186/s12888-016-0747-3
173. Oknina LB, Wild-Wall N, Oades RD, Juran SA, Ropcke B, Pfueller U, et al. Frontal and temporal sources of mismatch negativity in healthy controls, patients at onset of schizophrenia in adolescence and others at 15 years after onset. Schizophrenia Res. (2005) 76:25–41. doi: 10.1016/j.schres.2004.10.003
174. Naatanen R, Paavilainen P, Rinne T, Alho K. The mismatch negativity (MMN) in basic research of central auditory processing: a review. Clin Neurophysiol. (2007) 118:2544–90. doi: 10.1016/j.clinph.2007.04.026
175. Garrido MI, Kilner JM, Stephan KE, Friston KJ. The mismatch negativity: a review of underlying mechanisms. Clin Neurophysiol. (2009) 120:453–63. doi: 10.1016/j.clinph.2008.11.029
176. Earls HA, Curran T, Mittal V. A Meta-analytic review of auditory event-related potential components as endophenotypes for schizophrenia: perspectives from first-degree relatives. Schizophrenia Bull. (2016) 42:1504–16. doi: 10.1093/schbul/sbw047
177. Naatanen R. Mismatch negativity (MMN): perspectives for application. Int J Psychophysiol. (2000) 37:3–10. doi: 10.1016/S0167-8760(00)00091-X
178. Umbricht D, Krljes S. Mismatch negativity in schizophrenia: a meta-analysis. Schizophrenia Res. (2005) 76:1–23. doi: 10.1016/j.schres.2004.12.002
179. Naatanen R, Shiga T, Asano S, Yabe H. Mismatch negativity (MMN) deficiency: a break-through biomarker in predicting psychosis onset. Int J Psychophysiol. (2015) 95:338–44. doi: 10.1016/j.ijpsycho.2014.12.012
180. Brockhaus-Dumke A, Tendolkar I, Pukrop R, Schultze-Lutter F, Klosterkötter J, Ruhrmann S. Impaired mismatch negativity generation in prodromal subjects and patients with schizophrenia. Schizophrenia Res. (2005) 73:297–310. doi: 10.1016/j.schres.2004.05.016
181. Higuchi Y, Sumiyoshi T, Seo T, Miyanishi T, Kawasaki Y, Suzuki M. Mismatch negativity and cognitive performance for the prediction of psychosis in subjects with at-risk mental state. PLoS ONE. (2013) 8:e54080. doi: 10.1371/journal.pone.0054080
182. Magno E, Yeap S, Thakore JH, Garavan H, De Sanctis P, Foxe JJ. Are auditory-evoked frequency and duration mismatch negativity deficits endophenotypic for schizophrenia? high-density electrical mapping in clinically unaffected first-degree relatives and first-episode and chronic schizophrenia. Biol Psychiatry. (2008) 64:385–91. doi: 10.1016/j.biopsych.2008.03.019
183. Murphy TK, Haigh SM, Coffman BA, Salisbury DF. Mismatch negativity and impaired social functioning in long-term and in first episode schizophrenia spectrum psychosis. Front Psychiatry. (2020) 11:544. doi: 10.3389/fpsyt.2020.00544
184. Kiang M, Braff DL, Sprock J, Light GA. The relationship between preattentive sensory processing deficits and age in schizophrenia patients. Clin Neurophysiol. (2009) 120:1949–57. doi: 10.1016/j.clinph.2009.08.019
185. Erickson MA, Ruffle A, Gold JM. a meta-analysis of mismatch negativity in schizophrenia: from clinical risk to disease specificity and progression. Biol Psychiatry. (2016) 79:980–7. doi: 10.1016/j.biopsych.2015.08.025
186. Nagai T, Tada M, Kirihara K, Araki T, Jinde S, Kasai K. Mismatch negativity as a “translatable” brain marker toward early intervention for psychosis: a review. Front Psychiatry. (2013) 4:115. doi: 10.3389/fpsyt.2013.00115
187. Salisbury DF, McCathern AG, Coffman BA, Murphy TK, Haigh SM. Complex mismatch negativity to tone pair deviants in long-term schizophrenia and in the first-episode schizophrenia spectrum. Schizophrenia Res. (2018) 191:18–24. doi: 10.1016/j.schres.2017.04.044
188. Hermens DF, Ward PB, Hodge MA, Kaur M, Naismith SL, Hickie IB. Impaired MMN/P3a complex in first-episode psychosis: cognitive and psychosocial associations. Progress Neuro Psychopharmacol Biol Psychiatry. (2010) 34:822–9. doi: 10.1016/j.pnpbp.2010.03.019
189. Kaur M, Battisti RA, Ward PB, Ahmed A, Hickie IB, Hermens DF. MMN/P3a deficits in first episode psychosis: comparing schizophrenia-spectrum and affective-spectrum subgroups. Schizophrenia Res. (2011) 130:203–9. doi: 10.1016/j.schres.2011.03.025
190. Rydkjær J, Møllegaard Jepsen JR, Pagsberg AK, Fagerlund B, Glenthøj BY, Oranje B. Mismatch negativity and P3a amplitude in young adolescents with first-episode psychosis: a comparison with ADHD. Psychol Med. (2017) 47:377–88. doi: 10.1017/S0033291716002518
191. Solís-Vivanco R, Mondragón-Maya A, León-Ortiz P, Rodríguez-Agudelo Y, Cadenhead KS, de la Fuente-Sandoval C. Mismatch negativity reduction in the left cortical regions in first-episode psychosis and in individuals at ultra high-risk for psychosis. Schizophrenia Res. (2014) 158:58–63. doi: 10.1016/j.schres.2014.07.009
192. Koshiyama D, Kirihara K, Tada M, Nagai T, Koike S, Suga M, et al. Duration and frequency mismatch negativity shows no progressive reduction in early stages of psychosis. Schizophrenia Res. (2017) 190:32–8. doi: 10.1016/j.schres.2017.03.015
193. Lho SK, Kim M, Park J, Hwang WJ, Moon SY, Oh S, et al. Progressive impairment of mismatch negativity is reflective of underlying pathophysiological changes in patients with first-episode psychosis. Front Psychiatry. (2020) 11:587. doi: 10.3389/fpsyt.2020.00587
194. Nagai T, Tada M, Kirihara K, Yahata N, Hashimoto R, Araki T, et al. Auditory mismatch negativity and P3a in response to duration and frequency changes in the early stages of psychosis. Schizophrenia Res. (2013) 150:547–54. doi: 10.1016/j.schres.2013.08.005
195. Randau M, Oranje B, Miyakoshi M, Makeig S, Fagerlund B, Glenthøj B, et al. Attenuated mismatch negativity in patients with first-episode antipsychotic-naive schizophrenia using a source-resolved method. NeuroImage Clin. (2019) 22:101760. doi: 10.1016/j.nicl.2019.101760
196. Atkinson RJ, Michie PT, Schall U. Duration mismatch negativity and P3a in first-episode psychosis and individuals at ultra-high risk of psychosis. Biol Psychiatry. (2012) 71:98–104. doi: 10.1016/j.biopsych.2011.08.023
197. Salisbury DF, Coffman BA, Haigh SM. Reductions in complex mismatch negativity to extra tone gestalt pattern deviance in first-episode schizophrenia. Front Psychiatry. (2020) 11:505. doi: 10.3389/fpsyt.2020.00505
198. Salisbury DF, Shafer AR, Murphy TK, Haigh SM, Coffman BA. Pitch and duration mismatch negativity and Heschl's gyrus volume in first-episode schizophrenia-spectrum individuals. Clin EEG Neurosci. (2020) 51:359–64. doi: 10.1177/1550059420914214
199. Umbricht DSG, Bates JA, Lieberman JA, Kane JM, Javitt DC. Electrophysiological indices of automatic and controlled auditory information processing in first-episode, recent-onset and chronic schizophrenia. Biol Psychiatry. (2006) 59:762–72. doi: 10.1016/j.biopsych.2005.08.030
200. Düring S, Glenthøj BY, Oranje B. Effects of blocking D2/D3 receptors on mismatch negativity and P3a amplitude of initially antipsychotic naïve, first episode schizophrenia patients. Int J Neuropsychopharmacol. (2016) 19:1–7. doi: 10.1093/ijnp/pyv109
201. Jahshan C, Cadenhead KS, Rissling AJ, Kirihara K, Braff DL, Light GA. Automatic sensory information processing abnormalities across the illness course of schizophrenia. Psychol Med. (2012) 42:85–97. doi: 10.1017/S0033291711001061
202. Shaikh M, Valmaggia L, Broome MR, Dutt A, Lappin J, Day F, et al. Reduced mismatch negativity predates the onset of psychosis. Schizophrenia Res. (2012) 134:42–8. doi: 10.1016/j.schres.2011.09.022
203. Perez VB, Woods SW, Roach BJ, Ford JM, McGlashan TH, Srihari VH, et al. Automatic auditory processing deficits in schizophrenia and clinical high-risk patients: forecasting psychosis risk with mismatch negativity. Biol Psychiatry. (2014) 75:459–69. doi: 10.1016/j.biopsych.2013.07.038
204. Carrion RE, Cornblatt BA, McLaughlin D, Chang J, Auther AM, Olsen RH, et al. Contributions of early cortical processing and reading ability to functional status in individuals at clinical high risk for psychosis. Schizophrenia Res. (2015) 164:1–7. doi: 10.1016/j.schres.2015.01.030
205. Bodatsch M, Ruhrmann S, Wagner M, Müller R, Schultze-Lutter F, Frommann I, et al. Prediction of psychosis by mismatch negativity. Biol Psychiatry. (2011) 69:959–66. doi: 10.1016/j.biopsych.2010.09.057
206. Higuchi Y, Seo T, Miyanishi T, Kawasaki Y, Suzuki M, Sumiyoshi T. Mismatch negativity and p3a/reorienting complex in subjects with schizophrenia or at-risk mental state. Front Behav Neurosci. (2014) 8:172. doi: 10.3389/fnbeh.2014.00172
207. Atkinson RJ, Fulham WR, Michie PT, Ward PB, Todd J, Stain H, et al. Electrophysiological, cognitive and clinical profiles of at-risk mental state: the longitudinal Minds in Transition (MinT) study. PLoS ONE. (2017) 12:e0171657. doi: 10.1371/journal.pone.0171657
208. Devrim-Uçok M, Keskin-Ergen HY, Uçok A. Mismatch negativity at acute and post-acute phases of first-episode schizophrenia. Euro Arch Psychiatry Clin Neurosci. (2008) 258:179–85. doi: 10.1007/s00406-007-0772-9
209. Mondragón-Maya A, Solís-Vivanco R, León-Ortiz P, Rodríguez-Agudelo Y, Yáñez-Téllez G, Bernal-Hernández J, et al. Reduced P3a amplitudes in antipsychotic naïve first-episode psychosis patients and individuals at clinical high-risk for psychosis. J Psychiatric Res. (2013) 47:755–61. doi: 10.1016/j.jpsychires.2012.12.017
210. Morales-Muñoz I, Jurado-Barba R, Fernández-Guinea S, Álvarez-Alonso MJ, Rodríguez-Jiménez R, Jiménez-Arriero MA, et al. Cognitive impairments in patients with first episode psychosis: the relationship between neurophysiological and neuropsychological assessments. J Clin Neurosci. (2017) 36:80–7. doi: 10.1016/j.jocn.2016.10.023
211. Salisbury DF, Polizzotto NR, Nestor PG, Haigh SM, Koehler J, McCarley RW. Pitch and duration mismatch negativity and premorbid intellect in the first hospitalized schizophrenia spectrum. Schizophrenia Bull. (2017) 43:407–16. doi: 10.1093/schbul/sbw074
212. Turetsky BI, Calkins ME, Light GA, Olincy A, Radant AD, Swerdlow NR. Neurophysiological endophenotypes of schizophrenia: the viability of selected candidate measures. Schizophrenia Bull. (2007) 33:69–94. doi: 10.1093/schbul/sbl060
213. Kruiper C, Fagerlund B, Nielsen M, Düring S, Jensen MH, Ebdrup BH, et al. Associations between P3a and P3b amplitudes and cognition in antipsychotic-naïve first-episode schizophrenia patients. Psychol Med. (2019) 49:868–75. doi: 10.1017/S0033291718001575
214. Hamilton HK, Perez VB, Ford JM, Roach BJ, Jaeger J, Mathalon DH. Mismatch negativity but not P300 is associated with functional disability in schizophrenia. Schizophrenia Bull. (2018) 44:492–504. doi: 10.1093/schbul/sbx104
215. Hassan WA, Darweesh AEM, Abdel-Rahman AA, Ahmad HEK, Hassaan SH, Noaman MM, et al. P300 cognitive assessment in patients with first-episode psychosis: a prospective case-control study. Middle East Curr Psychiatry. (2020) 27:1–8. doi: 10.1186/s43045-020-00031-2
216. Devrim-Üçok M, Keskin-Ergen Y, Üçok A. Lack of progressive reduction in P3 amplitude after the first-episode of schizophrenia: a 6-year follow-up study. Psychiatry Res. (2016) 243:303–11. doi: 10.1016/j.psychres.2016.02.065
217. Guo Q, Tang Y, Li H, Zhang T, Li J, Sheng J, et al. Both volumetry and functional connectivity of Heschl's gyrus are associated with auditory P300 in first episode schizophrenia. Schizophrenia Res. (2014) 160:57–66. doi: 10.1016/j.schres.2014.10.006
218. Lho SK, Kim M, Lee TH, Kwak YB, Kwon JS. Predicting prognosis in patients with first-episode psychosis using auditory P300: a 1-year follow-up study. Clin Neurophysiol. (2019) 130:46–54. doi: 10.1016/j.clinph.2018.10.011
219. Renoult L, Prévost M, Brodeur M, Lionnet C, Joober R, Malla A, et al. P300 asymmetry and positive symptom severity: a study in the early stage of a first episode of psychosis. Schizophrenia Res. (2007) 93:366–73. doi: 10.1016/j.schres.2007.03.024
220. Salisbury DF, Shenton ME, Sherwood AR, Fischer IA, Yurgelun-Todd DA, Tohen M, et al. First-episode schizophrenic psychosis differs from first-episode affective psychosis and controls in P300 amplitude over left temporal lobe. Arch General Psychiatry. (1998) 55:173–80. doi: 10.1001/archpsyc.55.2.173
221. Xiong P, Zeng Y, Zhu Z, Tan D, Xu F, Lu J, et al. Reduced NGF serum levels and abnormal P300 event-related potential in first episode schizophrenia. Schizophrenia Res. (2010) 119:34–9. doi: 10.1016/j.schres.2010.02.1063
222. Wang J, Tang Y, Li C, Mecklinger A, Xiao Z, Zhang M, et al. Decreased P300 current source density in drug-naive first episode schizophrenics revealed by high density recording. Int J Psychophysiol. (2010) 75:249–57. doi: 10.1016/j.ijpsycho.2009.12.005
223. Wang CH, Li Y, Yang J, Su LY, Geng YG, Li H, et al. A randomized controlled trial of olanzapine improving memory deficits in Han Chinese patients with first-episode schizophrenia. Schizophrenia Res. (2013) 144:129–35. doi: 10.1016/j.schres.2012.12.021
224. McCarley RW, Salisbury DF, Hirayasu Y, Yurgelun-Todd DA, Tohen M, Zarate C, et al. Association between smaller left posterior superior temporal gyrus volume on magnetic resonance imaging and smaller left temporal P300 amplitude in first-episode schizophrenia. Arch General Psychiatry. (2002) 59:321–31. doi: 10.1001/archpsyc.59.4.321
225. Fusar-Poli P, Crossley N, Woolley J, Carletti F, Perez-Iglesias R, Broome M, et al. White matter alterations related to P300 abnormalities in individuals at high risk for psychosis: an MRI-EEG study. J Psychiatry Neurosci. (2011) 36:239–48. doi: 10.1503/jpn.100083
226. Ozgurdal S, Gudlowski Y, Witthaus H, Kawohl W, Uhl I, Hauser M, et al. Reduction of auditory event-related P300 amplitude in subjects with at-risk mental state for schizophrenia. Schizophrenia Res. (2008) 105:272–8. doi: 10.1016/j.schres.2008.05.017
227. van der Stelt O, Lieberman JA, Belger A. Auditory P300 in high-risk, recent-onset and chronic schizophrenia. Schizophrenia Res. (2005) 77:309–20. doi: 10.1016/j.schres.2005.04.024
228. Tang Y, Wang J, Zhang T, Xu L, Qian Z, Cui H, et al. P300 as an index of transition to psychosis and of remission: Data from a clinical high risk for psychosis study and review of literature. Schizophrenia Res. (2019). doi: 10.1016/j.schres.2019.02.014
229. Demiralp T, Üçok A, Devrim M, Isoglu-Alkaç U, Tecer A, Polich J. N2 and P3 components of event-related potential in first-episode schizophrenic patients: scalp topography, medication, and latency effects. Psychiatry Res. (2002) 111:167–79. doi: 10.1016/S0165-1781(02)00133-6
230. Devrim-Uçok M, Keskin-Ergen HY, Uçok A. Novelty P3 and P3b in first-episode schizophrenia and chronic schizophrenia. Progress Neuro Psychopharmacol Biol Psychiatry. (2006) 30:1426–34. doi: 10.1016/j.pnpbp.2006.05.019
231. Hirayasu Y, Asato N, Ohta H, Hokama H, Arakaki H, Ogura C. Abnormalities of auditory event-related potentials in schizophrenia prior to treatment. Biol Psychiatry. (1998) 43:244–53. doi: 10.1016/S0006-3223(97)00275-8
232. Wang J, Hirayasu Y, Hiramatsu K, Hokama H, Miyazato H, Ogura C. Increased rate of P300 latency prolongation with age in drug-naive and first episode schizophrenia. Clin Neurophysiol. (2003) 114:2029–35. doi: 10.1016/S1388-2457(03)00207-4
233. Kutas M, Federmeier KD. Thirty years and counting: finding meaning in the N400 component of the event-related brain potential (ERP). Annu Rev Psychol. (2011) 62:621–47. doi: 10.1146/annurev.psych.093008.131123
234. Kiang M, Gerritsen CJ. The N400 event-related brain potential response: a window on deficits in predicting meaning in schizophrenia. Int J Psychophysiol. (2019) 145:65–9. doi: 10.1016/j.ijpsycho.2019.04.005
235. Du XD, Zhang GY, Yang Y, Li Z, Pan W, Yin GZ, et al. Follow-up of N400 in the rehabilitation of first-episode schizophrenia. Chin Med J. (2015) 128:2215–9. doi: 10.4103/0366-6999.162499
236. Lepock JR, Ahmed S, Mizrahi R, Gerritsen CJ, Maheandiran M, Bagby RM, et al. N400 event-related brain potential as an index of real-world and neurocognitive function in patients at clinical high risk for schizophrenia. Early Intervent Psychiatry. (2019) 15:68–75. doi: 10.1111/eip.12911
237. Fusar-Poli P, McGuire P, Borgwardt S. Mapping prodromal psychosis: a critical review of neuroimaging studies. Euro Psychiatry. (2012) 27:181–91. doi: 10.1016/j.eurpsy.2011.06.006
238. Kahn RS, Sommer IE. The neurobiology and treatment of first-episode schizophrenia. Mol Psychiatry. (2015) 20:84–97. doi: 10.1038/mp.2014.66
239. Dandash O, Pantelis C, Fornito A. Dopamine, fronto-striato-thalamic circuits and risk for psychosis. Schizophrenia Res. (2017) 180:48–57. doi: 10.1016/j.schres.2016.08.020
240. Chu AOK, Chang WC, Chan SKW, Lee EHM, Hui CLM, Chen EYH. Comparison of cognitive functions between first-episode schizophrenia patients, their unaffected siblings and individuals at clinical high-risk for psychosis. Psychol Med. (2019) 49:1929–36. doi: 10.1017/S0033291718002726
241. Zhang H, Wang Y, Hu Y, Zhu Y, Zhang T, Wang J, et al. Meta-analysis of cognitive function in Chinese first-episode schizophrenia: MATRICS consensus cognitive battery (MCCB) profile of impairment. General Psychiatry. (2019) 32:e100043. doi: 10.1136/gpsych-2018-100043
242. de Bock R, Mackintosh AJ, Maier F, Borgwardt S, Riecher-Rössler A, Andreou C. EEG microstates as biomarker for psychosis in ultra-high-risk patients. Transl Psychiatry. (2020) 10:300. doi: 10.1038/s41398-020-00963-7
243. Boutros NN, Mucci A, Vignapiano A, Galderisi S. Electrophysiological aberrations associated with negative symptoms in schizophrenia. Curr Topics Behav Neurosci. (2014) 21:129–56. doi: 10.1007/7854_2014_303
244. Dondé C, Martínez A, Kantrowitz JT, Silipo G, Dias EC, Patel GH, et al. Bimodal distribution of tone-matching deficits indicates discrete pathophysiological entities within the syndrome of schizophrenia. Transl Psychiatry. (2019) 9:221. doi: 10.1038/s41398-019-0557-8
245. Lee M, Sehatpour P, Dias EC, Silipo GS, Kantrowitz JT, Martinez AM, et al. A tale of two sites: differential impairment of frequency and duration mismatch negativity across a primarily inpatient versus a primarily outpatient site in schizophrenia. Schizophrenia Res. (2018) 191:10–7. doi: 10.1016/j.schres.2017.07.032
246. Huiskamp G. Interindividual variability of skull conductivity: an EEG-MEG analysis. Int J Bioelectromag. (2008) 10:25–30.
247. Dager SR, Friedman SD. Brain imaging and the effects of caffeine and nicotine. Ann Med. (2000) 32:592–9. doi: 10.3109/07853890009002029
248. Clark SR, Baune BT, Schubert KO, Lavoie S, Smesny S, Rice SM, et al. Prediction of transition from ultra-high risk to first-episode psychosis using a probabilistic model combining history, clinical assessment and fatty-acid biomarkers. Transl Psychiatry. (2016) 6:e897. doi: 10.1038/tp.2016.170
249. Zimmermann R, Gschwandtner U, Wilhelm FH, Pflueger MO, Riecher-Rössler A, Fuhr P. EEG spectral power and negative symptoms in at-risk individuals predict transition to psychosis. Schizophr Res. (2010) 123:208–16. doi: 10.1016/j.schres.2010.08.031
250. Baradits M, Bitter I, Czobor P. Multivariate patterns of EEG microstate parameters and their role in the discrimination of patients with schizophrenia from healthy controls. Psychiatry Res. (2020) 288:112938. doi: 10.1016/j.psychres.2020.112938
251. Koutsouleris N, Borgwardt S, Meisenzahl EM, Bottlender R, Möller HJ, Riecher-Rössler A. Disease prediction in the at-risk mental state for psychosis using neuroanatomical biomarkers: results from the FePsy study. Schizophrenia Bull. (2012) 38:1234–46. doi: 10.1093/schbul/sbr145
252. Tognin S, Pettersson-Yeo W, Valli I, Hutton C, Woolley J, Allen P, et al. Using structural neuroimaging to make quantitative predictions of symptom progression in individuals at ultra-high risk for psychosis. Front Psychiatry. (2013) 4:187. doi: 10.3389/fpsyt.2013.00187
253. Ramyead A, Studerus E, Kometer M, Uttinger M, Gschwandtner U, Fuhr P, et al. Prediction of psychosis using neural oscillations and machine learning in neuroleptic-naïve at-risk patients. World J Biol Psychiatry. (2016) 17:285–95. doi: 10.3109/15622975.2015.1083614
254. Bak N, Ebdrup BH, Oranje B, Fagerlund B, Jensen MH, Düring SW, et al. Two subgroups of antipsychotic-naive, first-episode schizophrenia patients identified with a Gaussian mixture model on cognition and electrophysiology. Transl Psychiatry. (2017) 7:e1087. doi: 10.1038/tp.2017.59
255. Qu X, Liukasemsarn S, Tu J, Higgins A, Hickey TJ, Hall M-H. Identifying clinically and functionally distinct groups among healthy controls and first episode psychosis patients by clustering on EEG patterns. Front Psychiatry. (2020) 11:541659. doi: 10.3389/fpsyt.2020.541659
Keywords: electroencephalogram, first-episode psychosis, first-episode schizophrenia, clinical high-risk, ultra high-risk, frequency bands, microstates, ERPs
Citation: Perrottelli A, Giordano GM, Brando F, Giuliani L and Mucci A (2021) EEG-Based Measures in At-Risk Mental State and Early Stages of Schizophrenia: A Systematic Review. Front. Psychiatry 12:653642. doi: 10.3389/fpsyt.2021.653642
Received: 14 January 2021; Accepted: 06 April 2021;
Published: 04 May 2021.
Edited by:
Vicente Molina, University of Valladolid, SpainReviewed by:
Clément Dondé, Université Grenoble Alpes, FranceCopyright © 2021 Perrottelli, Giordano, Brando, Giuliani and Mucci. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Giulia Maria Giordano, Z2l1bGlhbWdpb3JkYW5vQGdtYWlsLmNvbQ==
†These authors have contributed equally to this work and share first authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.