
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
PERSPECTIVE article
Front. Psychiatry , 28 January 2016
Sec. Neuroimaging
Volume 7 - 2016 | https://doi.org/10.3389/fpsyt.2016.00009
This article is part of the Research Topic Third-generation neuroimaging: translating research into clinical utility View all 22 articles
 Frauke Schultze-Lutter1*
Frauke Schultze-Lutter1* Martin Debbané2,3
Martin Debbané2,3 Anastasia Theodoridou4
Anastasia Theodoridou4 Stephen J. Wood5
Stephen J. Wood5 Andrea Raballo6
Andrea Raballo6 Chantal Michel1
Chantal Michel1 Stefanie J. Schmidt1
Stefanie J. Schmidt1 Jochen Kindler1Stephan Ruhrmann7
Jochen Kindler1Stephan Ruhrmann7 Peter J. Uhlhaas8
Peter J. Uhlhaas8
In its initial formulation, the concept of basic symptoms (BSs) integrated findings on the early symptomatic course of schizophrenia and first in vivo evidence of accompanying brain aberrations. It argued that the subtle subclinical disturbances in mental processes described as BSs were the most direct self-experienced expression of the underlying neurobiological aberrations of the disease. Other characteristic symptoms of psychosis (e.g., delusions and hallucinations) were conceptualized as secondary phenomena, resulting from dysfunctional beliefs and suboptimal coping styles with emerging BSs and/or concomitant stressors. While BSs can occur in many mental disorders, in particular affective disorders, a subset of perceptive and cognitive BSs appear to be specific to psychosis and are currently employed in two alternative risk criteria. However, despite their clinical recognition in the early detection of psychosis, neurobiological research on the aetiopathology of psychosis with neuroimaging methods has only just begun to consider the neural correlate of BSs. This perspective paper reviews the emerging evidence of an association between BSs and aberrant brain activation, connectivity patterns, and metabolism, and outlines promising routes for the use of BSs in aetiopathological research on psychosis.
Over the past two decades, preventive research in psychosis has renewed interest in subjective and subclinical psychopathology beyond positive and negative symptoms. One approach to a detailed description of such subtle disturbances, developed since the 1960s, is Huber’s “basic symptoms” (BSs) concept (Figure S1 in Supplementary Material).
Basic symptoms are subtle, subclinical disturbances in stress tolerance, drive, affect, thinking, speech, (body) perception, motor action, and central-vegetative functions that are self-experienced with full insight into their abnormal nature (1, 2). Despite having insight, people find these subjective experiences so new and strange that they remain almost inexplicable, and therefore usually require guided questioning for their assessment. Being different from what is considered to be one’s “normal” mental state, BSs remain predominately private and apparent only to the individual. Thus, rather than BSs themselves, it will be a person’s affective reactions and self-initiated coping strategies in response to their BSs that may be recognized by others. Therefore, BSs differ from (secondary) negative symptoms in their current understanding as dysfunctional mental and behavioral response observable to others (3). Being experienced with full insight, BSs are also distinct from positive symptoms which are experienced by the individual as real, normal thinking, and feeling (2, 4).
Basic symptoms are an integral part of psychosis and appear throughout various stages of the disorder (Figure S1 in Supplementary Material). In combination with selected attenuated psychotic symptoms (APS), a subgroup of BSs was recently conceptualized as “self-disorders” (SDs) and a core schizophrenia vulnerability phenotype [Figures S2 and S3 in Supplementary Material (5–7)].
Huber’s pioneer pneumoencephalographic in vivo studies on chronic schizophrenia patients with persistent negative or deficit symptoms led him to initially assume that a deficit syndrome characterized by BSs was caused in most cases by an atrophy of the basal ganglia and inherent small dysplastic lateral ventricles (8). Later on, he put emphasis on the limbic system by conceptualizing BSs as “substrate-close” or “basic,” i.e., the most immediate symptomatic “expression of pathologically cerebral function in the region of the integrative system, which is responsible for the regulation of the cerebral filter and protection processes” [(9) p. 78]. While structural changes would be irreversible and potentially progressive, Huber (9) hypothesized that abnormal rhythms in EEG related to functional structures of the limbic system would only be seen in certain active (particularly early) stages.
While studies in the 1980s and 1990s indicated that most BSs are indeed not specific to psychosis and may occur in other, especially non-psychotic affective disorders (4), 14 BSs were specific to the development of first-episode schizophrenia within 9.6 years (10) and employed in two clinical high risk (CHR) criteria (4, 11, 12): Cognitive Disturbances, COGDIS, Cognitive-Perceptive BSs, and COPER (Table 1).

Table 1. CHR criteria according to the BSs concept.
A recent meta-analysis (11) revealed pooled conversion rates in COGDIS-defined samples of up to 54.9% within 4 years. Four-year conversion rates of COGDIS samples were significantly higher than those of samples established by ultra-high risk (UHR) criteria (11), mainly by APS. Thus, COGDIS is one of three criteria recommended for CHR assessment by the European Psychiatric Association (11).
Neurocognitive deficits are a common feature of schizophrenia and are also reported in CHR samples (13). To date, few studies separately examined BSs samples and reported rather inconsistent findings. Generally, patients exclusively meeting BSs criteria performed intermediate to UHR patients and controls. While neurocognitive deficits and cognitive BSs did not correlate, there is some evidence for an association between exclusive presence of COPER and executive control/verbal memory dysfunction (Table 2) (14). Thus, BSs samples without APS or brief limited intermittent psychotic symptoms (BLIPS) exhibit fewer and less pronounced neurocognitive impairments compared to samples with APS/BLIPS. This might indicate that BSs generally precede neurocognitive impairments.
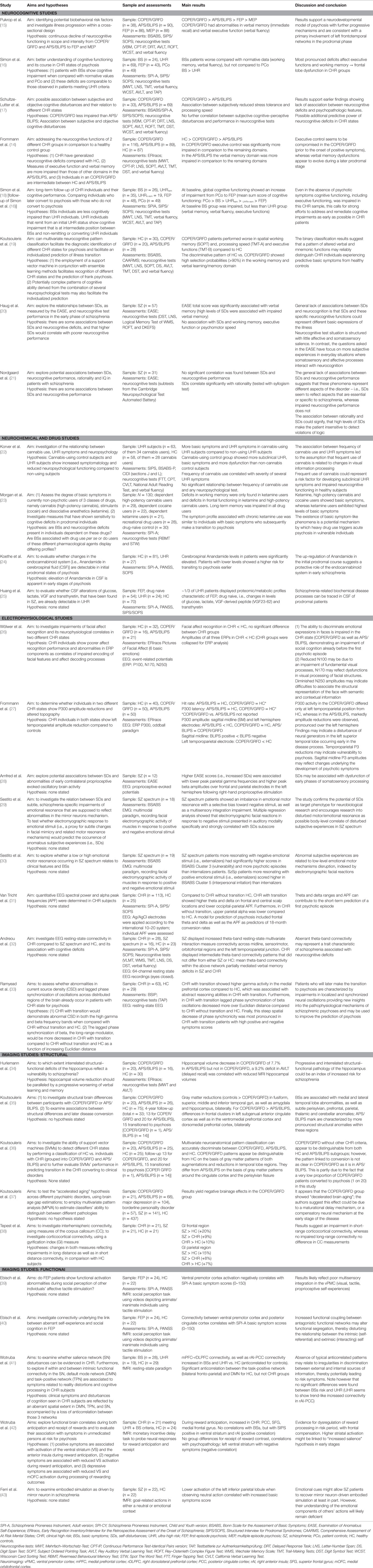
Table 2. Summary of neurobiological studies of basic symptoms.
Neurochemical findings suggest a role for dopaminergic, glutamatergic, serotonergic, and GABAergic systems in schizophrenia (44, 45). In vivo research on CHR states, defined by either UHR or BSs criteria, has focused mainly on dopamine (46, 47), glutamate (48, 49), and GABA (50, 51). Currently, the strongest evidence comes from Positron Emission Tomography studies indicating an increase of presynaptic striatal dopamine synthesis in APS patients compared with controls (46, 47). In addition, an increased dopamine synthesis capacity was also reported for individuals at genetic high risk for schizophrenia (52). Dopamine studies in BSs samples however are still missing.
These neurochemical studies have been complemented by pharmacological models of psychosis, e.g., the ketamine and endocannabinoid models (22, 23). Ketamine is an NMDA receptor antagonist, whereas cannabis or delta-9 tetrahydrocannabinol (THC) is an agonist on the cannabinoid receptor CB1. While commonly the effect of substances has to be ruled out to rate subjective experiences as BSs, recent studies ignoring this rule demonstrated an association of cannabis and ketamine use with cognitive and perceptive BSs (22, 23). Cannabis-using CHR patients had more BSs than non-using patients (22), while non-CHR cannabis users had significantly more positive, disorganization, general symptoms and BSs and also more neurocognitive deficits than non-users (23). Furthermore, the profiles of BSs and neurocognitive deficits of high-potency cannabis and ketamine users resembled COPER patients who subsequently converted to psychosis more closely than the profiles of users of other substances (23). Additional support of an association between the endocannabinoid system and BSs comes from one study investigating anandamide a bioactive lipid binding to cannabinoid receptors, in the cerebrospinal fluid (CSF) of CHR states (24). CHR individuals with higher anandamide levels showed a lower risk for transitioning to psychosis. Finally, one study of the metabolic profile in CSF (25) reported increased levels of glucose and VGF peptide (a polypeptide expressed by neurons and neuroendocrine tissues), and decreased levels of lactate and transthyretin protein in CHR patients (Table 2).
EEG and magnetoencephalographic (MEG) recordings permit the non-invasive assessment of electric currents of large populations of neurons, thus providing an estimate of both spontaneous and task-induced activity with millisecond resolution.
In a study of event-related potentials (ERP: P100, N170, and N250) using a facial recognition paradigm (26), emotion recognition was reduced in CHR groups and accompanied by a decrease in ERP amplitudes. As emotion recognition is already completed within 100 msec (53), these findings may reflect more complex perceptual processes. Further research on their relation to visual BSs may be promising. Moreover, reduced P300 amplitudes during an auditory oddball paradigm were found in a CHR sample (27). The COPER group showed a significant lower amplitude at a left temporoparietal site, whereas the APS/BLIPS group elicited smaller amplitudes at midline and left hemispheric electrode sites. These findings suggest potential differences in ERPs between BSs-defined and APS-defined CHR patients that might be related to different states of disturbed information processing.
In addition, Arnfred et al. (28) examined correlations between proprioceptively evoked event-related responses and changes in SDs in a small sample of schizophrenia patients. EEG data were examined for the spectral power of evoked-activity at beta/gamma-band frequencies (13–80 Hz) in response to a sudden change in muscle contraction. Increased total scores as well as increased ratings in the subscales “cognition and stream of consciousness,” “self-awareness and presence” and “bodily experiences” correlated significantly with lower gamma-band activity over parietal electrodes and higher peak frequencies in beta-activity (Table 2). Beside task-related activity, resting-state neural oscillations have also been recently investigated in CHR samples defined by both APS and BSs (31, 32), indicating increased delta/theta-band activity with reduced alpha-band power (31) and reduced theta-band activity which correlated with neurocognitive impairments (32), respectively. Moreover, there is emerging evidence that spontaneous gamma-band activity may differentiate CHR patients from controls (33).
These findings provide preliminary evidence for a potential link between BSs and abnormalities in EEG parameters in patients with schizophrenia and CHR groups. However, it is essential for these findings to be replicated and expanded in large samples. Overall, data on aberrant neural oscillations and ERP-parameters are consistent with data from APS-only (54) and schizophrenia samples (55), suggesting a continuum between psychosis-risk and progression to ScZ. Thus, neural oscillations and their synchronization could constitute a candidate mechanism for BSs. During normal brain functioning, rhythmic activity, especially at gamma-frequency ranges, are important for ensuring effective communication between and within neuronal assembles and correlate with a range of cognitive processes, including attention, perception, and working memory (55). Moreover, 30–80 Hz activity is generated by the interplay between GABAergic-interneurons and excitatory drive mediated through NMDA/AMPA-receptors (56, 57). These cellular mechanisms have been shown to be disrupted in schizophrenia (58).
To date, a handful of magnetic resonance imaging (MRI) studies have included the assessment of BSs by specialized instruments (Table 2). Their findings were similar to those reported for UHR and schizophrenia patients (59–61).
Five studies have investigated structural characteristics in relation to BSs. These studies distinguished “early risk” for psychosis, characterized by either COPER or the UHR genetic risk criterion (GRFD) in the absence of symptomatic UHR states, from “late risk,” which encompasses individuals meeting APS or BLIPS criteria, irrespective of the presence of BSs (34–37). Hurlemann et al. (34) reported bilaterally reduced hippocampi in COPER/GRFD and in APS/BLIPS subjects, correlating in the latter group with delayed recall in a verbal memory test. Koutsouleris et al. (35) employed voxel-based morphometry analyses to examine morphological differences between early- and late-risk samples. In comparison to controls, the COPER/GRFD group presented gray matter reductions involving the fusiform, superior, middle, and inferior temporal gyri, as well as amygdala and hippocampus, bilaterally. While they were associated with medial and lateral temporal lobe abnormalities, as well as subtle perisylvian, prefrontal, parietal, thalamic, and cerebellar anomalies, these alterations were continuous with late-risk participants (35). In a parallel study using multivariate neuroanatomical pattern classification, morphological patterns of COPER/GRFD were distinguishable from controls on the basis of gray matter patterns of both augmentations and reductions in temporal lobe regions; they differed from late risk on the basis of gray matter patterns covering the anteroposterior cingulate cortex and the perisylvian fissure (36). Using brain age gap estimations, the same group of participants reporting COPER/GRFD presented a “decelerated brain aging,” suggesting differential maturational dynamics at different stages of risk. Such deceleration could be due to a maturational delay mechanism, or a compensatory neural mechanism at the early stage of the disease (37). A further study indicated increased gyrification in frontal and parietal regions in BSs individuals (identified using COGDIS) compared to controls, implicating impaired short-range corticocortical connectivity (38).
To date, only five fMRI studies have examined BSs (39–43). Studies examining cerebral activation in patients with schizophrenia consistently reported significant associations between BSs and localized decreased activations in the ventral premotor cortex (40) and in the left inferior parietal lobule during passive viewing of neutral actions (43). Notably, increased connectivity between ventral premotor cortex and posterior cingulate cortex was associated with the severity of BSs in first-episode schizophrenia patients (39). This is consistent with another study examining the relationship between task positive and default-mode connectivity in CHR subjects, which reported a lack of anticorrelation between task positive and task negative networks (41).
The preliminary nature of these studies does not enable any definite conclusion as to the links between altered functional connectivity and BSs. Differences in methodology as well as heterogeneity of samples, which included both CHR and frankly psychotic patients, limit the interpretation of the available results and likely contributed to their inconsistencies (62). Furthermore, the variety of BSs involved, which touch upon motivation, cognitive and perceptual domains are likely to involve a variety cortical regions and networks. Overall, these pioneering studies suggest atypical patterns of neural activation in relation to BSs in terms of both reduced activity in discrete regions associated to self-other boundary distinction and atypical cross-talk between networks, which is similar to what is observed in UHR populations (59).
Although, in their original formulation, BSs were thought to reflect core abnormalities in brain functioning, investigations have only recently begun to look for their neurobiological origins, especially in CHR groups. Given the importance of BSs criteria in CHR research (11), further studies are needed to provide more detailed insights into the underlying neurobiological correlates that give rise to self-experienced disturbances in perception and cognition. Such candidate mechanisms could be of crucial importance for understanding the etiology of psychosis-risk as well as provide potential biomarkers for early detection and diagnosis. Moreover, such insights could point toward useful targets for novel pharmacological and psychological interventions that might ultimately reduce conversion rates.
Current studies provide only preliminary evidence for neurobiological mechanisms underlying BSs. Available data highlights that diverse anatomical, pharmacological and functional correlates may be involved in the manifestation of BSs in psychotic and CHR individuals. These include structural alterations, changes in ERPs and neural oscillations, neurotransmitter systems as well as evidence for changes in large-scale networks as assessed with fMRI. For more detailed and possibly mechanistic insights into neurobiology of BSs, different strategies need to be employed.
First, BSs in their original formulation are a heterogeneous set of symptoms comprising disturbances in perception, affect, drive, and cognition. Dimensional analyses of BSs indicated six BSs dimensions in adult psychosis patients that appear stable across various states of the illness [i.e., from the prodromal phase via the first episode to chronic states/relapse (4, 63)]. However, these six dimensions could not be replicated in a sample of early-onset schizophrenia patients, where four slightly different dimensions emerged [Figures S4–S6 and Table S1 in Supplementary Material (4)]. Thus, while past studies have either considered BSs in total (e.g., SPI-A sum score) or only considered COPER or COGDIS, BSs dimensions or even more differentiated BSs cluster (e.g., distinguishing even further between cognitive, speech, visual, and acoustic disturbances) might offer a more appropriate route to detecting neurobiological mechanisms underlying (if related to specific BSs) or further promoting (if related to unspecific BSs) development of psychosis. Furthermore, as recently indicated for single attenuated and manifest psychotic “Ich-Störungen (64), even investigating the neurobiology of single BSs may provide some significant insight. For example, when considering subjective disturbances of receptive and expressive speech as subtle, functional and only transient variants of receptive and amnesic aphasia, respectively (65), on a phenomenological level, it seems promising to investigate the role of brain regions which are highly correlated with theses neurologic syndromes also in patients exhibiting these BS.
Further insights into the origins of BSs may be derived from considering their developmental pattern and relationship to brain development. Psychoses, in particular of the schizophrenia-spectrum, are considered a fundamentally neurodevelopmental disorder involving two critical time windows [early (perinatal) brain development and adolescence] that together produce the symptomatic manifestations of the disorder. In this framework, early developmental insults may lead to dysfunction of specific neural networks that would account for early and (in some cases) trait-like signs and symptoms, which may have little or no clinical significance. The development of a CHR state (e.g., by additional occurrence or an increase in frequency of COGDIS symptoms) might index an ongoing imbalance of excessive synaptogenesis/pruning in critical networks, and ultimately the emergence of diagnostically relevant psychotic symptoms. This is supported by a recent analysis of two general population samples spanning the age of 8–40 years (66). Age seemed to affect the 14 perceptive and cognitive BSs included in COPER and COGDIS differently, indicating an age threshold of perceptive BS in late adolescents, around age 18, and of cognitive BSs in young adulthood, in the early twenties – with higher prevalence but reduced association to functional deficits and presence of mental disorder in the below-threshold groups. Thereby, differential age effects seem to follow normal back-to-front brain maturation processes, during which BSs might occur as temporary, in most cases infrequently occurring non-pathological disturbances. Their persistence or onset after conclusion of main brain maturation processes, however, might signify aberrant brain processes. Furthermore, an alternative or complementary explanation of this age-related pattern might be given against the background of decreasing brain plasticity after myelination and pruning processes reach maturity in the early 20s (67): BSs developing in childhood and adolescence, or rather their underlying neurobiological aberrations, might be much better compensated for by the still developing brain and thus, despite the larger number of affected youths, might only be reported by them as infrequent, momentary phenomena. These assumptions might be examined in future studies (i) by cross-sectionally comparing brain development in adolescents reporting BSs with and without clinical significance, (ii) by comparing subjects with an onset of BSs before and after age 18, and (iii) by comparing non-converters and converters to psychosis with regard to their onset and course of BSs.
In addition, if BSs reflect brain processes related to psychosis, then manifestation of BSs should closely be linked to genetic risk. Indeed, studies on first-degree relatives (7, 68–70) and schizotypal samples (71–73) indicated increased rates of BSs compared to HCs and non-schizophrenia-spectrum patients, respectively. In first-degree relatives, this involved more specific cognitive, perceptive and stress-tolerance-related BSs in particular (68). A recent genome-wide association study (74) identified 108 genetic variants associated with biological pathways central to the pathophysiology of schizophrenia. Thus, another route of future studies could be to explore links between certain BSs and possibly related risk-variants, e.g., between certain cognitive BSs and SNPs involved in cognition related neurotransmission.
Additional insights into the neurobiological basis of BSs might also be gained by using pharmacological perturbations in healthy samples. Preliminary evidence indicates that ketamine as well as THC may be associated with the expression of BSs (22, 23). Further studies in healthy populations using a range of pharmacological models which target specific cannabinoid, glutamatergic, dopaminergic, and GABAergic systems could provide important information on the contribution of dysfunctions in neurotransmitter systems and manifestation of BSs.
Finally, studying CHR patients at different stages may reveal the neurobiological correlates of BSs and evolution of schizophrenia. Theoretical and empirical evidence suggests that BSs may constitute the earliest signposts, preceding the development of UHR and psychotic symptoms. Recent studies point toward anatomical and electrophysiological differences between BSs and UHR samples; but these findings require replication and extension with functional imaging approaches. Longitudinal investigations of individuals identified on the basis of BSs alone could provide crucial information on their neurobiological correlates and potential progressive pathophysiological processes that might lead eventually to psychosis.
In conclusion, despite the fact that BSs were in their original (and pioneering) formulation considered to be a direct manifestation of brain abnormalities in patients with schizophrenia, the nature of such abnormalities appear subtle and heterogeneous, requiring sophisticated methods of analyses to be detected. Our review suggests that the CHR paradigm may constitute a fruitful paradigm to investigate the relationship between phenomenologically grounded perceptual and cognitive alterations and underlying abnormalities in the functionality of anatomical and functional brain networks. These insights may not only be useful for an improved general understanding of BSs but may ultimately give critical insights into the development of psychosis, which could be crucial for early diagnosis and intervention. Furthermore, as BSs are not only present in the initial prodromal phase, but throughout the course of the illness, they may also enable new insights into the neurobiological determinants of unfavorable outcomes including functional deterioration.
FS-L and PU wrote the outline of the article. All authors managed the literature searches to draft their respective chapters. FS-L drafted the introduction and parts 1 and 3 of the Supplementary Material; CM drafted the chapter on Neurocognition and BSs; AT and JK drafted the chapter on Neurochemistry and BSs; PU drafted the chapter on Electrophysiology and BSs and the discussion; MD drafted the chapter on Neuroimaging and BSs; AR drafted part 2 of the Supplementary Material. All authors contributed to and have approved the final manuscript.
Stephan Ruhrmann served as a consultant for Boehringer Ingelheim, received Speaker’s honoraria from Otsuka and Johnson & Johnson, and travel support from Servier. All other authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The Supplementary Material for this article can be found online at http://journal.frontiersin.org//article/10.3389/fpsyt.2016.00009
1. Huber G, Gross G. The concept of basic symptoms in schizophrenic and schizoaffective psychoses. Recent Prog Med (1989) 80:646–52.
2. Schultze-Lutter F. Subjective symptoms of schizophrenia in research and the clinic: the basic symptom concept. Schizophr Bull (2009) 35(1):5–8. doi: 10.1093/schbul/sbn139
3. Koehler K, Sauer H. Huber’s basic symptoms: another approach to negative psychopathology in schizophrenia. Compr Psychiatry (1984) 25(2):174–82. doi:10.1016/0010-440X(84)90006-3
4. Schultze-Lutter F, Ruhrmann S, Fusar-Poli P, Bechdolf A, Schimmelmann BG, Klosterkötter J. Basic symptoms and the prediction of first-episode psychosis. Curr Pharm Des (2012) 18(4):351–7. doi:10.2174/138161212799316064
5. Parnas J, Handest P, Jansson L, Sæbye D. Anomalous subjective experience among first-admitted schizophrenia spectrum patients: empirical investigation. Psychopathology (2005) 38:259–67. doi:10.1159/000088442
6. Parnas J, Møller P, Kircher T, Thalbitzer J, Jansson L, Handest P, et al. Examination of anomalous self-experience. Psychopathology (2005) 38(5):236–58. doi:10.1016/S0013-7006(12)70090-8
7. Raballo A, Parnas J. The silent side of the spectrum: schizotypy and the schizotaxic self. Schizophr Bull (2011) 37(5):1017–26. doi:10.1093/schbul/sbq008
8. Huber G. Reine Defektsyndrome und Basisstadien endogener Psychosen. Fortschr Neurol Psychiatr (1966) 34:409–26.
9. Huber G. Indizien für die Somatosehypothese bei den Schizophrenien. Fortschr Neurol Psychiatr (1976) 44:77–94.
10. Klosterkötter J, Hellmich M, Steinmeyer EM, Schultze-Lutter F. Diagnosing schizophrenia in the initial prodromal phase. Arch Gen Psychiatry (2001) 58(2):158–64. doi:10.1001/archpsyc.58.2.158
11. Schultze-Lutter F, Michel C, Schmidt SJ, Schimmelmann BG, Maric NP, Salokangas RK, et al. EPA guidance on the early detection of clinical high risk states of psychoses. Eur Psychiatry (2015) 30(3):405–16. doi:10.1016/j.eurpsy.2015.01.010
12. Schultze-Lutter F, Addington J, Ruhrmann S, Klosterkötter J. Schizophrenia Proneness Instrument, Adult version (SPI-A). Rom: Giovanni Fioriti Editore s.r.l. (2007). 97 p.
13. de Paula AL, Hallak JE, Maia-de-Oliveira JP, Bressan RA, Machado-de-Sousa JP. Cognition in at-risk mental states for psychosis. Neurosci Biobehav Rev (2015) 57:199–208. doi:10.1016/j.neubiorev.2015.09.006
14. Frommann I, Pukrop R, Brinkmeyer J, Bechdolf A, Ruhrmann S, Berning J, et al. Neuropsychological profiles in different at-risk states of psychosis: executive control impairment in the early – and additional memory dysfunction in the late – prodromal state. Schizophr Bull (2011) 37(4):861–73. doi:10.1093/schbul/sbp155
15. Pukrop R, Schultze-Lutter F, Ruhrmann S, Brockhaus-Dumke A, Tendolkar I, Bechdolf A, et al. Neurocognitive functioning in subjects at risk for a first episode of psychosis compared with first- and multiple-episode schizophrenia. J Clin Exp Neuropsychol (2006) 28(8):1388–407. doi:10.1080/13803390500434425
16. Simon AE, Cattapan-Ludewig K, Zmilacher S, Arbach D, Gruber K, Dvorsky DN, et al. Cognitive functioning in the schizophrenia prodrome. Schizophr Bull (2007) 33(3):761–71. doi:10.1093/schbul/sbm018
17. Schultze-Lutter F, Ruhrmann S, Picker H, von Reventlow HG, Daumann B, Brockhaus-Dumke A, et al. Relationship between subjective and objective cognitive function in the early and late prodrome. Br J Psychiatry Suppl (2007) 51:s43–51. doi:10.1192/bjp.191.51.s43
18. Simon AE, Grädel M, Cattapan-Ludewig K, Gruber K, Ballinari P, Roth B, et al. Cognitive functioning in at-risk mental states for psychosis and 2-year clinical outcome. Schizophr Res (2012) 142(1–3):108–15. doi:10.1016/j.schres.2012.09.004
19. Koutsouleris N, Davatzikos C, Bottlender R, Patschurek-Kliche K, Scheuerecker J, Decker P, et al. Early recognition and disease prediction in the at-risk mental states for psychosis using neurocognitive pattern classification. Schizophr Bull (2012) 38(6):1200–15. doi:10.1093/schbul/sbr037
20. Haug E, Øie M, Melle I, Andreassen OA, Raballo A, Bratlien U, et al. The association between self-disorders and neurocognitive dysfunction in schizophrenia. Schizophr Res (2012) 135(1–3):79–83. doi:10.1016/j.schres.2011.11.015
21. Nordgaard J, Revsbech R, Henriksen MG. Self-disorders, neurocognition and rationality in schizophrenia: a preliminary study. Psychopathology (2015) 48(5):310–6. doi:10.1159/000435892
22. Korver N, Nieman DH, Becker HE, van de Fliert JR, Dingemans PH, de Haan L, et al. Symptomatology and neuropsychological functioning in cannabis using subjects at ultra-high risk for developing psychosis and healthy controls. Aust N Z J Psychiatry (2010) 44(3):230–6. doi:10.3109/00048670903487118
23. Morgan CJ, Duffin S, Hunt S, Monaghan L, Mason O, Curran HV. Neurocognitive function and schizophrenia-proneness in individuals dependent on ketamine, on high potency cannabis (‘skunk’) or on cocaine. Pharmacopsychiatry (2012) 45(7):269–74. doi:10.1055/s-0032-1306310
24. Koethe D, Giuffrida A, Schreiber D, Hellmich M, Schultze-Lutter F, Ruhrmann S, et al. Anandamide elevation in cerebrospinal fluid in initial prodromal states of psychosis. Br J Psychiatry (2009) 194(4):371–2. doi:10.1192/bjp.bp.108.053843
25. Huang JT, Leweke FM, Tsang TM, Koethe D, Kranaster L, Gerth CW, et al. CSF metabolic and proteomic profiles in patients prodromal for psychosis. PLoS One (2007) 2(8):e756. doi:10.1371/journal.pone.0000756
26. Wölwer W, Brinkmeyer J, Stroth S, Streit M, Bechdolf A, Ruhrmann S, et al. Neurophysiological correlates of impaired facial affect recognition in individuals at risk for schizophrenia. Schizophr Bull (2012) 38(5):1021–9. doi:10.1093/schbul/sbr013
27. Frommann I, Brinkmeyer J, Ruhrmann S, Hack E, Brockhaus-Dumke A, Bechdolf A, et al. Auditory P300 in individuals clinically at risk for psychosis. Int J Psychophysiol (2008) 70(3):192–205. doi:10.1016/j.ijpsycho.2008.07.003
28. Arnfred SM, Raballo A, Morup M, Parnas J. Self-disorder and brain processing of proprioception in schizophrenia spectrum patients: a re-analysis. Psychopathology (2015) 48(1):60–4. doi:10.1159/000366081
29. Sestito M, Raballo A, Umiltà MA, Leuci E, Tonna M, Fortunati R, et al. Mirroring the self: testing neurophysiological correlates of disturbed self-experience in schizophrenia spectrum. Psychopathology (2015) 48(3):184–91. doi:10.1159/000380884
30. Sestito M, Raballo A, Umiltà MA, Amore M, Maggini C, Gallese V. Anomalous echo: exploring abnormal experience correlates of emotional motor resonance in Schizophrenia Spectrum. Psychiatry Res (2015) 229(1–2):559–64. doi:10.1016/j.psychres.2015.05.038
31. van Tricht MJ, Ruhrmann S, Arns M, Müller R, Bodatsch M, Velthorst E, et al. Can quantitative EEG measures predict clinical outcome in subjects at clinical high risk for psychosis? A prospective multicenter study. Schizophr Res (2014) 153(1–3):42–7. doi:10.1016/j.schres.2014.01.019
32. Andreou C, Leicht G, Nolte G, Polomac N, Moritz S, Karow A, et al. Resting-state theta-band connectivity and verbal memory in schizophrenia and in the high-risk state. Schizophr Res (2015) 161(2–3):299–307. doi:10.1016/j.schres.2014.12.018
33. Ramyead A, Kometer M, Studerus E, Koranyi S, Ittig S, Gschwandtner U, et al. Aberrant current source-density and lagged phase synchronization of neural oscillations as markers for emerging psychosis. Schizophr Bull (2015) 41(4):919–29. doi:10.1093/schbul/sbu134
34. Hurlemann R, Jessen F, Wagner M, Frommann I, Ruhrmann S, Brockhaus A, et al. Interrelated neuropsychological and anatomical evidence of hippocampal pathology in the at-risk mental state. Psychol Med (2008) 38(6):843–51. doi:10.1017/S0033291708003279
35. Koutsouleris N, Schmitt GJ, Gaser C, Bottlender R, Scheuerecker J, McGuire P, et al. Neuroanatomical correlates of different vulnerability states for psychosis and their clinical outcomes. Br J Psychiatry (2009) 195(3):218–26. doi:10.1192/bjp.bp.108.052068
36. Koutsouleris N, Meisenzahl EM, Davatzikos C, Bottlender R, Frodl T, Scheuerecker J, et al. Use of neuroanatomical pattern classification to identify subjects in at-risk mental states of psychosis and predict disease transition. Arch Gen Psychiatry (2009) 66(7):700–12. doi:10.1001/archgenpsychiatry.2009.62
37. Koutsouleris N, Davatzikos C, Borgwardt S, Gaser C, Bottlender R, Frodl T, et al. Accelerated brain aging in schizophrenia and beyond: a neuroanatomical marker of psychiatric disorders. Schizophr Bull (2014) 40(5):1140–53. doi:10.1093/schbul/sbt142
38. Tepest R, Schwarzbach CJ, Krug B, Klosterkötter J, Ruhrmann S, Vogeley K. Morphometry of structural disconnectivity indicators in subjects at risk and in age-matched patients with schizophrenia. Eur Arch Psychiatry Clin Neurosci (2013) 263(1):15–24. doi:10.1007/s00406-012-0343-6
39. Ebisch SJ, Mantini D, Northoff G, Salone A, De Berardis D, Ferri F, et al. Altered brain long-range functional interactions underlying the link between aberrant self-experience and self-other relationship in first-episode schizophrenia. Schizophr Bull (2014) 40(5):1072–82. doi:10.1093/schbul/sbt153
40. Ebisch SJ, Salone A, Ferri F, De Berardis D, Romani GL, Ferro FM, et al. Out of touch with reality? Social perception in first-episode schizophrenia. Soc Cogn Affect Neurosci (2013) 8(4):394–403. doi:10.1093/scan/nss012
41. Wotruba D, Michels L, Buechler R, Metzler S, Theodoridou A, Gerstenberg M, et al. Aberrant coupling within and across the default mode, task-positive, and salience network in subjects at risk for psychosis. Schizophr Bull (2014) 40(5):1095–104. doi:10.1093/schbul/sbt161
42. Wotruba D, Heekeren K, Michels L, Buechler R, Simon JJ, Theodoridou A, et al. Symptom dimensions are associated with reward processing in unmedicated persons at risk for psychosis. Front Behav Neurosci (2014) 8:382. doi:10.3389/fnbeh.2014.00382
43. Ferri F, Costantini M, Salone A, Ebisch S, De Berardis D, Mazzola V, et al. Binding action and emotion in first-episode schizophrenia. Psychopathology (2014) 47(6):394–407. doi:10.1159/000366133
44. Steeds H, Carhart-Harris RL, Stone JM. Drug models of schizophrenia. Ther Adv Psychopharmacol (2015) 5(1):43–58. doi:10.1177/2045125314557797
45. Lebedev AV, Lövdén M, Rosenthal G, Feilding A, Nutt DJ, Carhart-Harris RL. Finding the self by losing the self: neural correlates of ego-dissolution under psilocybin. Hum Brain Mapp (2015) 36(8):3137–53. doi:10.1002/hbm.22833
46. Howes OD, Bose SK, Turkheimer F, Valli I, Egerton A, Valmaggia LR, et al. Dopamine synthesis capacity before onset of psychosis: a prospective [18F]-DOPA PET imaging study. Am J Psychiatry (2011) 168(12):1311–7. doi:10.1176/appi.ajp.2011.11010160
47. Egerton A, Chaddock CA, Winton-Brown TT, Bloomfield MA, Bhattacharyya S, Allen P, et al. Presynaptic striatal dopamine dysfunction in people at ultra-high risk for psychosis: findings in a second cohort. Biol Psychiatry (2013) 74(2):106–12. doi:10.1016/j.biopsych.2012.11.017
48. Allen P, Chaddock CA, Egerton A, Howes OD, Barker G, Bonoldi I, et al. Functional outcome in people at high risk for psychosis predicted by thalamic glutamate levels and prefronto-striatal activation. Schizophr Bull (2015) 41(2):429–39. doi:10.1093/schbul/sbu115
49. de la Fuente-Sandoval C, León-Ortiz P, Favila R, Stephano S, Mamo D, Ramírez-Bermúdez J, et al. Higher levels of glutamate in the associative-striatum of subjects with prodromal symptoms of schizophrenia and patients with first-episode psychosis. Neuropsychopharmacology (2011) 36(9):1781–91. doi:10.1038/npp.2011.65
50. de la Fuente-Sandoval C, Reyes-Madrigal F, Mao X, León-Ortiz P, Rodríguez-Mayoral O, Solís-Vivanco R, et al. Cortico-striatal GABAergic and glutamatergic dysregulations in subjects at ultra-high risk for psychosis investigated with proton magnetic resonance spectroscopy. Int J Neuropsychopharmacol (2015). doi:10.1093/ijnp/pyv105
51. Kang JI, Park HJ, Kim SJ, Kim KR, Lee SY, Lee E, et al. Reduced binding potential of GABA-A/benzodiazepine receptors in individuals at ultra-high risk for psychosis: an [18F]-fluoroflumazenil positron emission tomography study. Schizophr Bull (2014) 40(3):548–57. doi:10.1093/schbul/sbt052
52. Huttunen J, Heinimaa M, Svirskis T, Nyman M, Kajander J, Forsback S, et al. Striatal dopamine synthesis in first-degree relatives of patients with schizophrenia. Biol Psychiatry (2008) 63(1):114–7. doi:10.1016/j.biopsych.2007.04.017
53. Liu L, Ioannides AA. Emotion separation is completed early and it depends on visual field presentation. PLoS One (2010) 5(3):e9790. doi:10.1371/journal.pone.0009790
54. Nagai T, Tada M, Kirihara K, Araki T, Jinde S, Kasai K. Mismatch negativity as a “translatable” brain marker toward early intervention for psychosis: a review. Front Psychiatry (2013) 4:115. doi:10.3389/fpsyt.2013.00115
55. Uhlhaas PJ, Singer W. Oscillations and neuronal dynamics in schizophrenia: the search for basic symptoms and translational opportunities. Biol Psychiatry (2015) 77(12):1001–9. doi:10.1016/j.biopsych.2014.11.019
56. Wang XJ. Neurophysiological and computational principles of cortical rhythms in cognition. Physiol Rev (2010) 90(3):1195–268. doi:10.1152/physrev.00035.2008
57. Kantrowitz JT, Javitt DC. N-methyl-d-aspartate (NMDA) receptor dysfunction or dysregulation: the final common pathway on the road to schizophrenia? Brain Res Bull (2010) 83(3–4):108–21. doi:10.1016/j.brainresbull.2010.04.006
58. Lewis DA, Hashimoto T, Volk DW. Cortical inhibitory neurons and schizophrenia. Nat Rev Neurosci (2005) 6(4):312–24. doi:10.1038/nrn1648
59. Chung Y, Cannon TD. Brain imaging during the transition from psychosis prodrome to schizophrenia. J Nerv Ment Dis (2015) 203(5):336–41. doi:10.1097/NMD.0000000000000286
60. Fornito A, Yücel M, Patti J, Wood SJ, Pantelis C. Mapping grey matter reductions in schizophrenia: an anatomical likelihood estimation analysis of voxel-based morphometry studies. Schizophr Res (2009) 108(1–3):104–13. doi:10.1016/j.schres.2008.12.011
61. Schultz CC, Fusar-Poli P, Wagner G, Koch K, Schachtzabel C, Gruber O, et al. Multimodal functional and structural imaging investigations in psychosis research. Eur Arch Psychiatry Clin Neurosci (2012) 262(Suppl 2):S97–106. doi:10.1007/s00406-012-0360-5
62. Borgwardt SJ, Stoyanov DS. Emerging psychiatric neuroimaging findings: translating the research into clinical application. In: Stoyanov DS, editor. Towards New Philosophy and Mental Health, Perspectives from Neuroscience and Humanities. Newcastle upon Tyne: Cambridge Scholars Publishing (2015). p. 35–41.
63. Schultze-Lutter F, Steinmeyer EM, Ruhrmann S, Klosterkötter J. The dimensional structure of self-reported ‘prodromal’ disturbances in schizophrenia. Clin Neuropsychiatry (2008) 5(3):140–50.
64. Mishara A, Bonoldi I, Allen P, Rutigliano G, Perez J, Fusar-Poli P, et al. Neurobiological models of self-disorders in early schizophrenia. Schizophr Bull (2015). doi:10.1093/schbul/sbv123
65. Kreisler A, Godefroy O, Delmaire C, Debachy B, Leclercq M, Pruvo JP, et al. The anatomy of aphasia revisited. Neurology (2000) 54(5):1117–23. doi:10.1212/WNL.54.5.1117
66. Schultze-Lutter F, Michel C, Schimmelmann BG. Impact of age on the prevalence and clinical significance of risk-symptoms in 8- to 40-year-olds of the general population. Eur Arch Psychiatry Clin Neurosci (2015) 265(Suppl 1):S11.
67. Paus T, Keshavan M, Giedd JN. Why do many psychiatric disorders emerge during adolescence? Nat Rev Neurosci (2008) 9(12):947–57. doi:10.1038/nrn2513
68. Klosterkötter J, Gross G, Huber G, Wieneke A, Steinmeyer EM, Schultze-Lutter F. Evaluation of the ‘Bonn Scale for the Assessment of Basic Symptoms – BSABS’ as an instrument for the assessment of schizophrenia proneness: a review of recent findings. Neurol Psychiatry Brain Res (1997) 5:137–50.
69. Raballo A, Sæbye D, Parnas J. Looking at the schizophrenia spectrum through the prism of self-disorders: an empirical study. Schizophr Bull (2011) 37(2):344–51. doi:10.1093/schbul/sbp056
70. Maggini C, Raballo A. Subjective experience of schizotropic vulnerability in siblings of schizophrenics. Psychopathology (2004) 37(1):23–8. doi:10.1159/000077016
71. Maggini C, Raballo A. Subjective experience of personality dimensions in 1st degree relatives of schizophrenics. Acta Biomed (2003) 74(3):131–6.
72. Maggini C, Raballo A. Subjective experience of schizotypal dimensions in 1st degree relatives of schizophrenics. Neurol Psychiatry Brain Res (2004) 11:71–6.
73. Handest P, Parnas J. Clinical characteristics of first-admitted patients with ICD-10 schizotypal disorder. Br J Psychiatry Suppl (2005) 48:s49–54. doi:10.1192/bjp.187.48.s49
Keywords: basic symptoms, neurobiology, psychosis, clinical high risk, aetiopathology
Citation: Schultze-Lutter F, Debbané M, Theodoridou A, Wood SJ, Raballo A, Michel C, Schmidt SJ, Kindler J, Ruhrmann S and Uhlhaas PJ (2016) Revisiting the Basic Symptom Concept: Toward Translating Risk Symptoms for Psychosis into Neurobiological Targets. Front. Psychiatry 7:9. doi: 10.3389/fpsyt.2016.00009
Received: 03 December 2015; Accepted: 14 January 2016;
Published: 28 January 2016
Edited by:
Stefan Borgwardt, University of Basel, SwitzerlandReviewed by:
Drozdstoy Stoyanov Stoyanov, Medical University of Plovdiv, BulgariaCopyright: © 2016 Schultze-Lutter, Debbané, Theodoridou, Wood, Raballo, Michel, Schmidt, Kindler, Ruhrmann and Uhlhaas. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Frauke Schultze-Lutter, ZnJhdWtlLnNjaHVsdHplLWx1dHRlckBranAudW5pYmUuY2g=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.