 Yunhui Wang
Yunhui Wang Tianxiao Lv
Tianxiao Lv Tian Fan
Tian Fan Yuping Zhou
Yuping Zhou Chang-en Tian
Chang-en Tian- Guangdong Provincial Key Laboratory of Plant Adaptation and Molecular Design, School of Life Sciences, Guangzhou University, Guangzhou Higher Education Mega Center, Guangzhou, China
Flowering represents a pivotal phase in the reproductive and survival processes of plants, with the photoperiod serving as a pivotal regulator of plant-flowering timing. An investigation of the mechanism of flowering inhibition in the model plant Arabidopsis thaliana under short-day (SD) conditions will facilitate a comprehensive approach to crop breeding for flowering time, reducing or removing flowering inhibition, for example, can extend the range of adaptation of soybean to high-latitude environments. In A. thaliana, CONSTANS (CO) is the most important component for promoting flowering under long-day (LD) conditions. However, CO inhibited flowering under the SD conditions. Furthermore, the current studies revealed that A. thaliana delayed flowering through multiple pathways that inhibit the transcription and sensitivity of FLOWERING LOCUS T (FT) and suppresses the response to, or synthesis of, gibberellins (GA) at different times, for potential crop breeding resources that can be explored in both aspects. However, the underlying mechanism remains poorly understood. In this review, we summarized the current understanding of delayed flowering under SD conditions and discussed future directions for related topics.
1 Introduction
Plant flowering is regulated by several environmental and endogenous conditions, which directly influence crop yield and quality. Most of the current research focuses on long-day (LD) not short-day (SD) to regulate flowering and it is particularly important to improve the mechanism of SD flowering network in some important crops. For example, the discovery and application of the long juvenile period has improved the status quo of soybean’s extremely low yields at low latitudes, which has led to the expansion of soybean to low latitudes and large-scale cultivation, and the inhibition of flowering has played a decisive role. The problem is that when only different latitude regions are planted to grow crops, especially for photoperiod-sensitive crops, timely flowering germplasm resource is needed (Lai et al., 2023), so mining flowering factors and realizing the precise flowering of crops through modern scientific means is a very important task. A. thaliana is a facultative long-day (LD) plant that exhibits a photoperiodic response that promotes flowering in LD (longer than 12 h of light) and delays flowering in SD (8-h light/16-h dark or 10-h light/14-h dark) (Takagi et al., 2023). A. thaliana regulates flowering by integrating information from multiple pathways, including photoperiodic, temperature, gibberellin (GA), vernalization, age and autonomous pathways (Fornara et al., 2010). Information from multiple pathways is integrated to regulate floral integrators that promote flowering. The photoperiod, or day length, is a stable external condition that plants can perceive and play a crucial role in controlling flowering time (Kinmonth-Schultz et al., 2023). As a florigen in plants, the FLOWERING LOCUS T (FT) protein functions as a systemic signal that induces flowering in the shoot apex (Zhu et al., 2021). The study of the key molecular mechanisms underlying the promotion of flowering in LD compared with the inhibition of flowering in short days (SD) is more in-depth in A. thaliana. The promotion of flowering by LD is primarily achieved through regulation of FT expression. The B-box (BBX) transcription factor CONSTANS (CO) is regarded as the most pivotal regulator of gene expression in A. thaliana (Imaizumi, 2010). The CO protein exhibited stabilization in the afternoon under LD conditions, yet only slight accumulation around ZT8 under SD conditions (Fernandez et al., 2016; Hayama et al., 2017; Valverde et al., 2004). The discrepancy in CO protein stability can account for the discrepancy in FT transcript levels between the LD and SD conditions (Fernandez et al., 2016). Notably, CO, which functions as a flowering promoter under LD conditions, delays flowering under SD conditions (Luccioni et al., 2019). Genetic evidence indicates that the TERMINAL FLOWER 1(TFL1) mutant tfl1 is epistatic to co and ft is epistatic to tfl1. CO may increase the response to FT by shifting TFL1 expression out of the peak of the maximal sensitivity to FT (Luccioni et al., 2019).
The delayed flowering observed under SD conditions necessitates the collective involvement of multiple repressors that exert pronounced inhibitory effects on flowering. The transcription factor SHORT VEGETATIVE PHASE (SVP) plays a pivotal role in the repression of GA biosynthesis and the expression of flowering integration factor genes FT, SUPPRESSOR OF OVEREXPRESSION OF CONSTANS 1(SOC1), and others (Andrés et al., 2014). A. thaliana plants can flower under SD conditions. The GA pathway is believed to play a crucial role in promoting flowering in A. thaliana (Wilson et al., 1992). However, the molecular mechanism by which SD delays flowering in A. thaliana is not yet fully understood, particularly when compared to the well-studied mechanism by which LD promotes flowering. This study presents a review of the literature on the inhibition of flowering in A. thaliana under SD to elucidate the underlying molecular mechanisms. It also serves as a reference for further research and breeding in the field of plant-flowering regulation.
2 CO-dependent regulations
2.1 co mutants and phenotypes
CO proteins are so important for the function of flowering, but there is still controversy about the phenotypes in CO mutants in A. thaliana in SD, so it is necessary to make a detailed list of the current CO mutants and phenotypes here, which will help us to discuss the role of CO under SD condition.
Twelve CO mutants exhibiting diverse backgrounds and mutagenesis methods were identified (Table 1). Most of them displayed an early flowering phenotype under SD conditions.
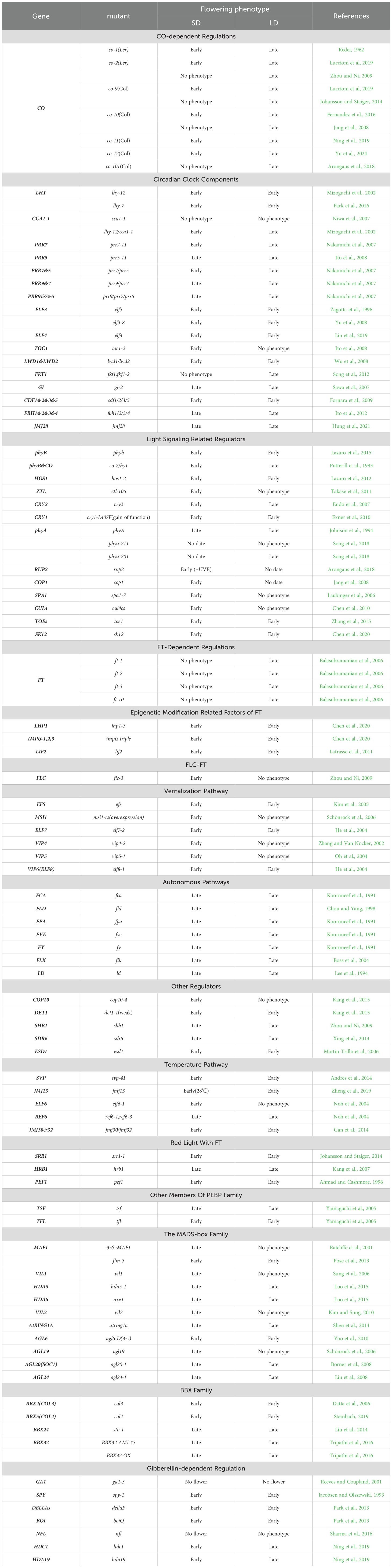
Table 1. Covered in the paper mutant phenotypes and their flowering responses under SD versus LD conditions.
The co-1 mutant was created using X-ray mutagenesis in 1962, and has been documented to flower prematurely in SD (Redei, 1962). The co-12 mutant site was identical to that of co-1, except for the genetic background of the mutant. co-12 is a Columbia (Col) background mutant, whereas co-1 is a Landsberg erecta (Ler) background mutant. Both mutants exhibit early flowering in SD (Redei, 1962; Balasubramanian et al., 2006; Zhang et al., 2015; Yu et al., 2024).
While co-2, which has been mutated at the carboxyl-terminus of the first B-BOX domain, has been observed to convert arginine to histidine in a Ler background (Robson et al., 2001), co-2 has been found to flower earlier than the wild-type (WT) plant under an 8-hour photoperiod and slightly earlier under a 10-hour photoperiod (An et al., 2004; Balasubramanian et al., 2006; Datta et al., 2006; Luccioni et al., 2019).
co-9, co-10, and co-11 are T-DNA insertion mutants isolated from a Col background that flowered early in SD (Balasubramanian et al., 2006; Luccioni et al., 2019; Ning et al, 2019). However, further investigation revealed that co-10 exhibited flowering patterns that differed from those of the wild type at both 21°C and 27°C in SD, suggesting that temperature may play a role in CO-mediated flowering (Fernandez et al., 2016).
The co-1, co-2, co-9, co-10, co-11, and co-12 plants displayed varying degrees of early flowering phenotypes in SD. The flowering phenotype of the co mutants in SD may also be related to the ecotype, with the Col background exhibiting a more pronounced early flowering phenotype and the Ler background displaying a milder early flowering phenotype. Notably, co-2 (Martin-Trillo et al., 2006; Zhou and Ni, 2009), co-9 (Johansson and Staiger, 2014), co-10 (Jang et al., 2008; Laubinger et al., 2006), and co-101 (Arongaus et al., 2018) have been shown to lack early flowering phenotypes in SD, which may be attributed to disparate culture conditions. The early flowering phenotype of the co mutants under SD conditions depended on TFL1 and FT. However, the function of CO under SD conditions is not achieved through a reduction in FT transcript levels (Luccioni et al., 2019). Further investigations are required to elucidate the mechanism of CO function under SD conditions. Further understanding of the role of CO in flowering regulation under SD conditions may be achieved by investigating the relevant regulators that alter CO transcript levels and protein levels/stability.
Therefore, CO has the function of inhibiting flowering under SD, although not strongly. Based on the fact that CO has the function of integrating information from circadian clock and and light signaling, it is therefore important for us to be able to use CO as a cue to mine the factors regulating CO under SD that have the potential to refine the network of SD inhibition of flowering. The transcriptional level of CO is primarily regulated by the circadian clock, whereas its protein level and stability are controlled by light signals (Suarez-Lopez et al., 2001; Valverde et al., 2004). Although the key components of the circadian clock have been demonstrated to regulate CO proteins during LD (Hayama et al., 2017), there is a paucity of evidence regarding the detection of CO protein levels in mutants of the key components of the circadian clock during SD.
2.2 CO transcription regulators: circadian clock components
The CO transcript remains at a low level under light (ZT0-ZT8), begins to increase after entering darkness, reaches its peak after 4 h of darkness (ZT12), and then decreases (Figure 1). The photoperiodic control of flowering time is inextricably linked to the circadian clock, which serves as the timing mechanism for measuring the duration of the day and night. In A. thaliana, complex transcriptional repression mechanisms interlocked with core clock components comprise the circadian clock (Shim et al., 2017).
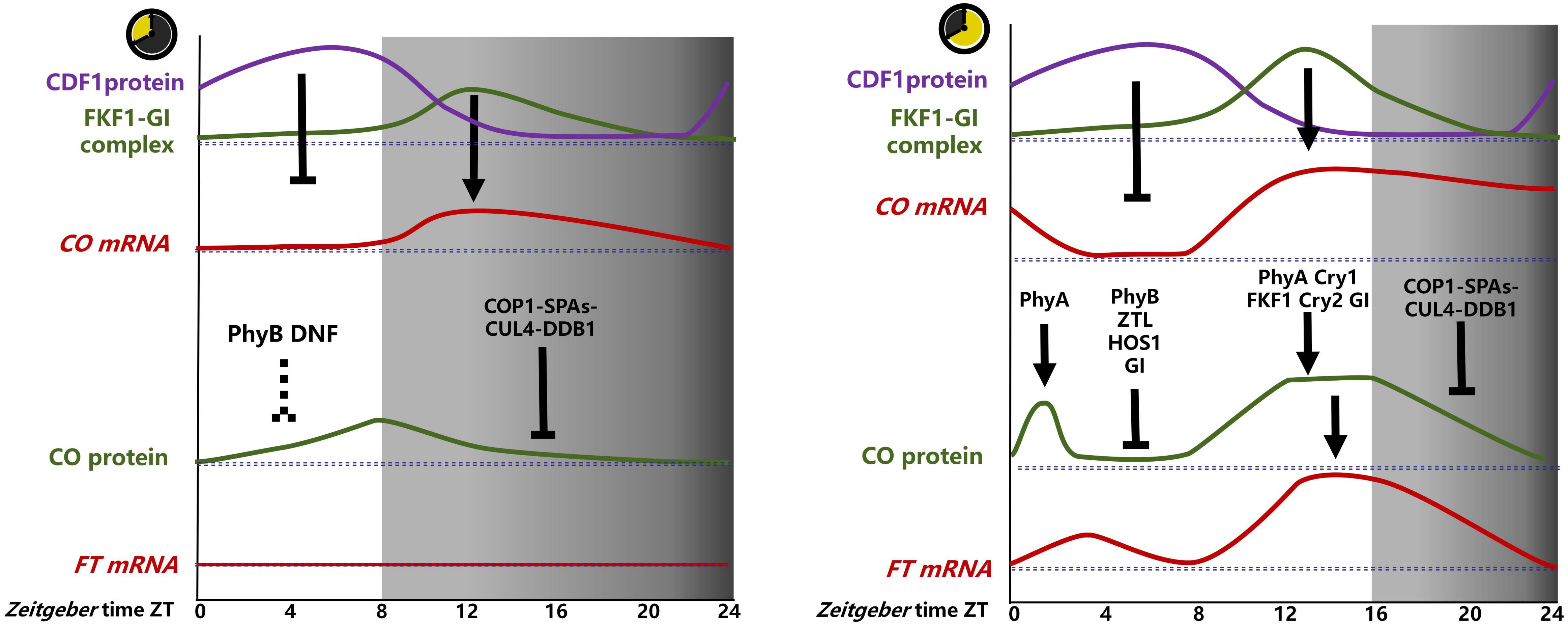
Figure 1. Photoperiodic regulation of CO-FT in A. thaliana. The CO transcript levels are mainly repressed by CDF1, which is mainly repressed by the FKF1-GI complex. The FKF1-GI complex accumulated more in ZT12-16 in LD and less in ZT12-16 in SD, resulting in high levels of CO at ZT12-ZT16 in LD and low levels at ZT12-ZT16 in SD. CO proteins were subjected to complex regulation in LD, stabilized by PHYA at ZT0-ZT4, degraded by GI, PHYB, HOS1, ZTL at ZT4-ZT8, PHYA, CRY1, FKF1, CRY2, and GI at ZT8-ZT16, stabilized, degraded by COP1-SPAs in dark ZT16-ZT24, and FT transcription was induced by CO mainly at ZT12-ZT16. In contrast, the regulation of CO proteins is less well studied in SD and is likely degraded by PHYB and DNF at ZT0-ZT4, degraded by COP1-SPAs-CUL4-DDB1 in the dark at ZT8-ZT24, and FT transcription is not induced by small amounts of accumulated CO proteins at ZT4-ZT12. (→, promote;  , inhabit).
, inhabit).
Circadian clock components exhibit distinct temporal expression patterns. For instance, the primary morning-time circadian clock components include two MYB transcription factors, CIRCADIAN CLOCK ASSOCIATED1 (CCA1) and LATE ELONGATED HYPOCOTYL (LHY) (Nagel et al., 2015; Kamioka et al., 2016), as well as PSEUDO RESPONSE REGULATOR 9 (PRR9), PRR7, and PRR5 identified noon-time components (Nakamichi et al., 2010; Nakamichi et al., 2012; Liu et al., 2014; Liu et al., 2016), LUX, ELF3, ELF4, and TOC1/PRR1 as key evening- and night- time components of the circadian clock (Doyle et al., 2002; Helfer et al., 2011; Nusinow et al., 2011; Herrero et al., 2012; Huang et al., 2016). Mutants in most genes exhibit aberrant flowering times under both LD and SD conditions.
The LHY mutant lhy-12 and CCA1 mutant cca1-1 flowered prematurely in SD, and the double mutant lhy-12/cca1-1 flowered especially early. This indicates that these two genes can function redundantly to inhibit flowering in SD (Mizoguchi et al., 2002)Subsequently, the expression peaks of the FLAVIN-BINDING, KELCH REPEAT, F-BOX PROTEIN 1 (FKF1), and GIGANTEA (GI) clock output, which are related to photoperiodic flowering, were advanced by approximately four hours in the lhy-7 mutant. Conversely, when the endogenous circadian cycle of the lhy-7 mutant aligns with the external light/dark cycle, the upregulation of FT is no longer observed (Park et al., 2016). This finding suggests that the early flowering observed in SD in the lhy-7 mutant is due to aberrant expression of photoperiodic flowering genes (Park et al., 2016). The flowering phenotype of lhy-7/co-101 under SD conditions has yet to be documented.
The midday circadian clock components are PPR9, PPR7, and PPR5. Among these, the PPR7 mutant Δ7 flowered prematurely under SD conditions (Nakamichi et al., 2007), the PPR5 mutant prr5-11 flowered slightly later (Ito et al., 2008), and the prr7/prr5 double mutant Δ7 flowered at the same time as Δ7 and earlier than the PPR9/PPR/PPR5 triple mutant Δ7 (Nakamichi et al., 2007). This indicates that PPR7 may play a dominant role in inhibiting flowering in SD. Nevertheless, no observable phenotypes have been documented for these PPR9 single mutants under SD conditions.
The evening circadian clock component ELF3 mutants, elf3 and elf3-8, flowered early during SD (Yu et al., 2008; Zagotta et al., 1996). The evening circadian clock component ELF4 mutant, elf4, also flowered early in SD (Lin et al., 2019). Subsequent studies demonstrated that ELF4 negatively regulates CO expression by forming an ELF4-GI complex with GI, which segregates GI from the CO promoter to specific nucleosomes (Kim et al., 2013). Furthermore, the early flowering phenotype of elf4 under SD conditions can be fully compensated by the overexpression of EFL1, partially compensated by the overexpression of EFL3, and not compensated by the overexpression of EFL2 (Lin et al., 2019), indicating that EFL1 is the primary flowering regulator through the circadian clock pathway under SD conditions.
The evening circadian clock component TIMING OF CAB2 EXPRESSION 1 (TOC1)/PRR1 mutants, toc1 and toc1-2, flowered at an early stage under SD conditions (Niwa et al., 2007). LIGHT-REGULATED WD1 (LWD1) and LWD2 affect flowering onset by regulating the circadian clock. lwd1/lwd2 double mutants exhibit an early flowering response in SD (Wu et al., 2008).
In SD, CO expression commences in the dark due to the accumulation of GI protein during the light period, reaching a peak at the end of the light phase. In contrast, the FKF1 protein does not reach a peak until ZT12 in the dark, resulting in the level of the GI-FKF complex remaining at a low level. This results in the accumulation of CYCLING DOF FACTORs (CDFs) proteins in light, which subsequently inhibits CO transcription. Following the transition to the dark, CDFs undergo degradation, allowing FBHs to bind to the E-box element in the CO promoter, activating CO transcription (Song et al., 2015; Figure 1).
Flowering regulation by these factors under SD conditions differed from that under LD conditions. The fkf1 and fkf1-2 mutants exhibited an unremarkable flowering phenotype (Song et al., 2012), indicating that FKF1 does not directly regulate flowering but that the transcript levels of CO and FT were moderately reduced in the afternoon (Imaizumi et al., 2003). The GI mutant gi-2 exhibits a late-flowering phenotype (Sawa et al., 2007; Sawa and Kay, 2011). However, the precise mechanism by which GI promotes flowering remains unclear. However, it has been demonstrated that GI does not regulate CO (Sawa and Kay, 2011). In addition, FKF1-GI complexes have been observed to form in SD (Sawa et al., 2007). The overexpression line 35S∷HA-FKF1#18 35S∷GI-TAP/fkf1 exhibited an extremely early flowering phenotype (Sawa et al., 2007), This indicates that the SD of 35S∷HA-FKF1#18 35S∷GI-TAP/fkf1 could promote CO transcripts by enhancing the level of the FKF1-GI complex, thereby accelerating flowering (Sawa et al., 2007). Nevertheless, the fkf1/gi mutant does not exhibit a flowering phenotype under SD conditions (Sawa et al., 2007).
The CDF family comprised five members. The quadruple mutant cdf1/2/3/5 exhibited an extremely early flowering phenotype in SD (Fornara et al., 2009). The FBH family comprises four members. The single mutants did not exhibit a flowering phenotype. However, fbh quadruple mutants displayed a slight delay in flowering compared to the wild type (Ito et al., 2012). In contrast, both 35S:FBH1 and 35S:FBH2 plants exhibit an extremely early flowering phenotype (Ito et al., 2012). Demethylase JMJ28 interacts with FBH transcription factors to activate the CO promoter by removing the repressor marker H3K9me2. The JMJ28 mutant jmj28 exhibited a slight delay in flowering under SD conditions (Hung et al., 2021).
In summary, under SD conditions, except for prr5-11 and gi-2, mutants of the circadian clock component and related regulators exhibited early flowering phenotypes, particularly toc1-2, elf3, and lhy-12/cca1-1. This suggests that most circadian clock components are repressed in SD, Some studies have indicated that the key components of the circadian clock during LD regulate CO transcription and directly regulate CO protein stability (Hayama et al., 2017), The mechanism by which these genes regulate flowering under SD needs to be deepened, and the components related to the circadian clock pathway are a very good candidate for breeding targets.Furthermore, these mutants exhibited elevated levels of FT transcripts, suggesting that repression was achieved by the suppression of FT transcription.
2.3 CO protein regulators
CO primarily functions as a transcription factor at the protein level. The exact mechanism of how CO proteins function in SD in contrast to LD,which inhibits not promotes flowering, is not yet clear. The main regulating CO proteins are light signaling factors, so through the phenotype of the mutant and the level of CO protein in the mutant, explore the possible mechanisms by which CO proteins regulate flowering in SD.
The level of CO protein increases with prolonged illumination time, reaches its peak at the end of 8 h (ZT8), and then decreases rapidly after entering darkness (Fernandez et al., 2016; Yu et al., 2024) (Figure 1). In contrast the level of CO transcript, these findings suggest that the regulation of CO protein levels may primarily depend on degradation rather than synthesis, and is not significantly correlated with alterations in CO transcript levels under SD conditions.
2.3.1 Light signaling related regulators
CO protein regulators have been studied more systematically under LD conditions and are mainly regulated by light signalling pathway factors, so how these factors regulate CO proteins under short day conditions and whether changes in CO proteins contribute to flowering will be discussed here.
Under LD conditions, the influence of different light signals on the photoperiodic flowering response varied. In the morning, CO degradation relies mainly on the red-light photoreceptor Phytochrome B (PhyB), PHYA and ZEITLUPE (ZTL) (Putterill et al., 1995; Reed et al., 1994; Hwang et al., 2019). In the afternoon, FKF1,Cryptochrome 1 (Cry2),Cry1,all are blue light photoreceptor, stabilize CO (Takase et al., 2011; Endo et al., 2007; Valverde et al., 2004). In the absence of light signaling at night or in the dark, CO is primarily degraded by CONSTITUTIVE PHOTOMORPHOGENIC 1 (COP1), SUPPRESSOR OF PHYA-105s (SPAs), CULLIN4 (CUL4) and Damaged DNA Binding Protein1 (DDB1) (Jang et al., 2008; Hwang et al., 2019); When the time comes to dawn, PHYA also stabilize CO transiently (Song et al., 2018; Figure 1).
PhyB, which receives red light, plays an important role in reducing the abundance of CO proteins and their activity in LD. phyb mutants exhibited early flowering in response to SD (Reed et al., 1994). The CO and PhyB double mutant co-2/hy1 exhibited a flowering time that was earlier than co-2 and later than hy1, indicating that CO is necessary for the early flowering phenotype of hy1 under SD conditions (Putterill et al., 1995). In SD, CO proteins are degraded via a pathway that requires PhyB (Andrés and Coupland, 2012; Figure 1); however, there is a paucity of relevant biochemical experiments. HOS1 and PhyB have been shown to play redundant roles in the regulation of flowering during SD (Lazaro et al., 2012; Lazaro et al., 2015). Under SD conditions, the hos1-2 mutant flowered early, whereas the hos1-2/co-2 double mutant flowered even earlier. This suggests that the early flowering of hos1 under SD conditions was not achieved through the CO pathway (Lazaro et al., 2012).
PhyA, which receives far-red light, contributes to CO stabilization in the dawn and afternoon under LD conditions. phyA flowered late in SD, which promotes flowering in SD (Johnson et al., 1994).
ztl-105 and ztl-4 mutants of the blue light-responsive ZTL flowered early under SD conditions (Takase et al., 2011; Hwang et al., 2019). A comparable CO protein abundance was observed in 35S:3HA-CO and 35S:3HA-CO/ztl-4 plants (Hwang et al., 2019) This mechanism appears to be distinct from that of ZTL-degrading CO activity in the morning of an LD (Song et al., 2014).
The early flowering observed in ztl mutants under SD conditions depends on FKF1 (Takase et al., 2011). In addition, the flowering phenotypes of the FKF1 mutants, fkf1 and fkf1-2, are not evident (Song et al., 2012), indicating that FKF1 does not independently regulate flowering under SD conditions. Nevertheless, 35S:3HA-CO showed a early flower phenotypes, but no notable discrepancy was observed in CO protein abundance between 35S:3HA-CO and 35S:3HA-CO/fkf1-2 (Song et al., 2012), indicating that early flowering of 35S:3HA-CO is independent of FKF1.
CRY2, which receives blue light in the afternoon on LD, degrades the COP1-SPAs complex and stabilizes CO (Endo et al., 2007). cry2 mutants have been observed to flowered early in SD (Endo et al., 2007)
cry1-L407F,a gain of function allele of CRY1, showed a very early flowering. It can increase the sensitivity of phytochrome signaling cascades (Exner et al., 2010).
As darkness decreases, plants cease to receive light signals, CONSTITUTIVE PHOTOMORPHOGENIC 1 (COP1) degradation CO protein (Jang et al., 2008). The cop1 mutant flowers prematurely under SD conditions, and substantial accumulation of CO proteins was observed in the dark (Jang et al., 2008). The cop1 co mutant exhibited a flowering time that was later than cop1 and earlier than co, indicating that CO is necessary for the early flowering of cop1 (Mcnellis et al., 1994; Jang et al., 2008). The SPA family comprises four members. The SPA family comprises four members: SPA1, SPA2, SPA3, and SPA4. Of these, SPA1 plays a dominant role in flowering, and SPA1 is sufficient for normal photoperiodic flowering. Furthermore, SPA1 is essential for maintaining flowering in wild-type plants under SD conditions (Laubinger et al., 2006). The spa1 mutant exhibited early flowering under SD conditions, which may be attributed to the presence of a substantial number of FT transcripts (Laubinger et al., 2006). Subsequently, considerable accumulation of CO proteins was observed in spa1-7 mutants under dark conditions (Jang et al., 2008). Consequently, in the absence of light signaling, plants require the COP1-SPAs complex to inhibit flowering by suppressing CO (Yu et al., 2008). CULLIN4 (CUL4)-Damaged DNA Binding Protein1(DDB1) may function with COP1-SPAs complexes to regulate CO protein degradation. Mutant cul4cs (for CUL4 co-suppression) exhibit an early flowering phenotype in SD (Chen et al., 2010). DDB1 has two isoforms: DDB1a,DDB1b.ddb1b mutant is embryo-lethal, whereas the knockout line ddb1a exhibits no obvious phenotype (Schroeder et al., 2002), indicating that DDB1 inhibits flowering via CUL4.
Ultraviolet B (UV-B) radiation is an essential component of light. The Repressor of UV-B Photomorphogenesis 2 (RUP2) has been identified as a flowering repressor under SD conditions containing UV-B. This repressor depends on the UV-B photoreceptor UVR8 (UV RESISTANCE LOCUS 8), and represses FT expression by inhibiting the binding of CO to the FT promoter (Arongaus et al., 2018). The rup2 mutant exhibited an early flowering phenotype under SD conditions (+UV-B) (Arongaus et al., 2018).
2.3.2 Other regulators
Besides the previously discussed CO protein regulators, other mutants of regulators that regulate flowering through CO under LD conditions have been observed to exhibit flowering phenotypes under SD conditions. The observations are presented in the following section. The TARGET OF EAT (TOE) proteins are members of the APETALA2 (AP2)-LIKE family of proteins, which includes TOE1, TOE2, TOE3, SCHLAFMÜTZE (SMZ), and SCHNARCHZAPFEN (SNZ) (Aukerman and Sakai, 2003; Chen, 2004). During the morning of an LD cycle (ZT0-ZT4), holidays inhibit CO activity by directly interacting with it (Zhang et al., 2015). In SD, TOE1 was expressed exclusively during the light period (ZT0-ZT8), with the highest level of expression occurring at ZT4. The toe1 mutant flowered earlier than the wild type, whereas toe1/co flowered later than the wild type and toe1 mutant (Zhang et al., 2015). This suggests that CO plays a role in the early flowering of toe1. Flowering was likely facilitated in the background of toe1 in SD; TOE2 was also involved in flower formation in SD, with toe1/toe2 flowering earlier than toe1 and later than toe1/toe2/co flowering (Zhang et al., 2015). It has been postulated that TOE1 and TOE2 may compete redundantly with CO for the FT promoter, functioning as inhibitors of CO binding to FT under SD conditions. Further experiments are required to substantiate this hypothesis. SHAGGY-like kinase 12 (SK12) is a member of the glycogen synthase kinase-3 family. Under SD conditions, sk12 also flowered early because of the inability of most of its CO proteins to be phosphorylated and their subsequent degradation through ubiquitination. This results in the accumulation of CO proteins and the promotion of flowering through the FT pathway (Chen et al., 2020). DAY NEUTRAL FLOWERING (DNF) encodes a functional membrane-bound E3 ligase, suggesting that DNF targets a repressor of CO for degradation by the proteasome pathway, dnf mutants flowered early (Morris et al., 2010). The GI protein is indirectly involved in the regulation of CO protein stability by forming a complex with the FKF1 and ZTL proteins under conditions of LD, and the gi-2 allele flowered late under SD (Song et al., 2014). The flowering time of 35S:3HA-CO was earlier than 35S:3HA-CO/gi-2 #1 and later than 35S:3HA-CO/fkf1-2 gi-2 #20 in SD (Song et al., 2014), indicating that early flowering of 35S:3HA-CO depends on GI. However, whether GI promotes flowering in nature remains to be determined using CO.
In summary, two insights can be drawn. One is that in the mutants with a large accumulation level of CO proteins, it still led to its early flowering phenotype, so a large number of CO proteins may still have promoting flowering function in SD, such as cop1, spa1, 35S:3HA-CO, sk12. And the regulation of flowering by CO proteins may be a dosage effect, which may also be why A. thaliana needs to maintain low levels of CO proteins in SD, and this mechanism is important for the function of CO proteins in SD. The others is the most of photoreceptor mutants showed an early flowering phenotypes, such as phyb,ztl-105,cry2 etc. suggesting that the photoreceptors mainly inhibit flowering in SD, and this inhibition is firstly due to unable to stabilize a large number of CO proteins. However, how the photoreceptors regulate to these low levels of CO proteins in SD may be the direction of the future research.
3 FT-dependent regulations
A. thaliana exhibits delayed flowering and a notable reduction in FT transcription under SD conditions. Besides the CO-FT pathway, A. thaliana represses FT transcription through epigenetic modifications of FT, FLC-FT pathway, and various related FT-regulated genes.
3.1 FT mutants and phenotypes
FT is a mobile protein synthesized in the companion cells of leaves and transported to the SAM through the phloem, where it promotes flowering (Maple et al., 2024). Under SD conditions, flowering is delayed in the wild type, which does not express or express low levels of FT transcripts (Luccioni et al., 2019). Furthermore, the FT mutants ft-1, ft-2, ft-3, and ft-10 do not exhibit obvious flowering phenotypes (Balasubramanian et al., 2006). In addition, natural variation in FT-creating promoter length does not alter flowering time in SD (Liu et al., 2014), suggesting that FT and FT under natural conditions in SD are not directly involved in regulating flowering.
3.2 Epigenetic modification related factors of FT
Photoperiods can directly regulate FT through cis-regulatory changes at its gene locus. Furthermore, under SD conditions, a minimal distance between the regulatory regions is required to fully suppress FT expression (Liu et al., 2014).
LIKE HETEROCHROMATIN PROTEIN 1 (LHP1) is a transcriptional repressor of flowering-related genes. It represses FT expression by directly associating with FT chromatin. The lhp1-3 mutant flowered under SD conditions have been shown to have elevated FT transcripts (Chen et al., 2020). Under SD conditions, LHP1 interacts with IMPα-1, 2, 3 to regulate flowering by modulating epigenetic modifications of FT. Impα-1, 2, 3 triple mutants flowered early, exhibited severely impaired nuclear targeting of LHP1, and displayed a substantial elevation of FT transcripts under SD conditions (Chen et al., 2020). Moreover, LHP1 interacts with LHP1-Interacting Factor 2 (LIF2) in the nucleus. LIF2 belongs to the hnRNP family of proteins and is involved in RNA processing, and lif2 flowered early under SD conditions (Latrasse et al., 2011).
3.3 FLC-FT dependent pathways
FLC plays a pivotal role in regulating flowering under LD conditions by engaging numerous genetic regulatory pathways associated with flowering (Michaels et al., 2003; Whittaker and Dean, 2017). In addition, FLC inhibits flowering under SD conditions and flc-3 exhibits early flowering (Zhou and Ni, 2009). Notably, the transcript levels of FLC in the wild type are comparable under LD and SD conditions (Zhou and Ni, 2009).
3.3.1 Vernalization pathway
Vernalization is the process by which plants undergo prolonged low-temperature treatments to promote flowering. Vernalization primarily disengages FLC inhibition of flowering by repressing FLC expression, optimizing the timing of flowering to align with the cessation of winter and onset of spring, which allows for maximal reproductive acclimation (Whittaker and Dean, 2017). The expression levels of FLC in the vernalization pathway are primarily regulated by upstream FRIGIDA (FRI) (Shindo et al., 2005; Zhang and Jiménez-Gómez, 2020). Allelic variation at the FRI locus in A. thaliana is a significant factor influencing the natural variation in flowering time (Johanson et al., 2000; Kim and Michaels, 2006). FRI can methylate FLC chromatin in complex with the histone methyltransferase EARLY FLOWERING IN SDS (EFS), which promotes FLC expression and efs mutant flowered early in SD (Kim et al., 2005).
In the vernalization pathway, the Polycomb Repressive Complex (PRC2) can cause FLC silencing through its specific component, PLANT HOMEODOMAIN (PHD) (Whittaker and Dean, 2017). PRC2 is a polycomb group (PcG) protein complex that is a cellular memory module that maintains the repression of gene transcription (Brock and Fisher, 2005). In the PcG protein MSI1 overexpression line msi1-cs, there was a marked reduction in MSI1 levels. These plants also exhibit early flowering in SD (Schönrock et al., 2006).
The RNA polymerase II-associated factor 1 (PAF1) complex also exerts a negative regulatory effect on FLC and FLC-like proteins via the vernalization pathway. The PAF1 complex comprises four subunits, ELF7, ERNALIZATION INDEPENDENCE 4 (VIP4), VIP5, and VIP6/ELF8. The elf7-3, vip4-2, vip5-1, and elf8-1 mutants exhibit an early flowering phenotype (He et al., 2004; Zhang and Van Nocker, 2002). Of these, elf7-3 and elf8-1 exhibited an early flowering phenotype (He et al., 2004). SKIP interacts with ELF7 to regulate flowering by activating FLC transcription and skip flowered early in SD (Cao et al., 2015). Therefore, the vernalization pathway also inhibited flowering in SD.
3.3.2 Autonomous pathways
The effect of FRI on FLC expression is antagonized by a group of proteins that have been termed the autonomous pathway, because their activity appears to be largely independent of the environment (Koornneef et al., 1998). The main components of the autonomous pathways involved in FLC are FCA, FLOWERING LOCUS D (FLD), FPA, FVE, FY, FLOWERING LOCUS K (FLK), and LD (LUMINI DEPENDENS). Loss-of-function of these genes delays flowering under any photoperiod (Chou and Yang, 2002; Koornneef et al., 1991; Lee et al., 1994).
3.3.3 Other regulators
COP10 epigenetically induces FLC expression by interacting with MULTICOPY SUPPRESSOR OF IRA14 (MSI4)/FVE (MSI4/FVE) (Kang et al., 2015). cop10-4 flowered early under SD conditions (Kang et al., 2015). COP10, DE-ETIOLATED1 (DET1), and Damaged DNA Binding Protein1 (DDB1) interacts to form the CDD complex and inhibit photomorphogenesis in the dark (Kang et al., 2015); det1-1 weak mutants flowered early in SD (Kang et al., 2015), suggesting that the CDD complex inhibits SD flowering. There are also genes that regulate flowering by modulating autonomous pathways. SHORT HYPOCOTYL UNDER BLUE1 (SHB1) encodes a yeast SYG1-like protein that represses the FLC pathway under SD conditions, resulting in FT transcription activation (Zhou and Ni, 2009). The shb1 mutant flowered late, whereas the functionally acquired mutant shb1-D flowered early (Zhou and Ni, 2009). SDR6 encodes a short-chain dehydrogenase/reductase containing an NAD(P) domain that regulates flowering through autonomous pathways. The SDR6 mutant, sdr6, flowered late (Xing et al., 2014).HIGH PLOIDY2 (HPY2) as an E3 SUMO ligase for FLC, regulates FLC function and stability at both the transcriptional and post-translational levels through its E3 SUMO ligase activity (Kwak et al., 2016). hpy2-2 mutants flowered early than wild-type plants (Kwak et al., 2016). EARLY IN SDS 1 (ESD1) is required for the expression of FLC repressors at levels that inhibit flowering. The ESD1 mutant esd1 flowered early (Martin-Trillo et al., 2006), REF6 encodes an H3K27 demethylation transferase that represses FLC expression by demethylating FLC, and the REF6 mutants ref6-1 and ref6-2 flowered late (Noh et al., 2004).
3.4 Temperature pathway
A. thaliana can overcome delayed flowering by increasing suitable temperature under SD conditions. The 28°C FT transcript level in WT plants under SD conditions was over 10-fold higher than that at 23°C, and diurnal oscillations were not affected (Balasubramanian et al., 2006).
The SVP mutant svp-41 flowered early in SD (Fernandez et al., 2016) and requires only a small amount of FT to flower at 27°C. This increased sensitivity to FT during flowering may be due to a reduction in SVP activity at the apex at 27°C. In addition, svp-41 ft-10 tsf -1 plants flowered simultaneously under 21°C-SD and 27°C-SD (Fernandez et al., 2016), indicating that the flowering response to temperature was not affected by the ft-10/tsf -1 mutation. These findings indicate that SVP, FT, and TSF are indispensable for the thermosensory induction of flowering under SD conditions. Furthermore, SVP selectively binds to FLOWERING LOCUS M (FLM)- and MADS AFFECTING FLOWERING2(MAF2)-specific transcripts and regulates flowering through temperature (Airoldi et al., 2015; Pose et al., 2013). The SVP plays a pivotal role in delaying flowering during SD. The JUMONJI (JMJ) family members JMJ13, JMJ30, and JMJ32 regulate A. thaliana flowers at different temperatures through epistatic regulation of FLC in SD.JMJ13 is an H3K27me3 demethylase that may negatively regulate temperature-driven flowering by suppressing temperature-photoperiod compensation, jmj13 mutant flowered early at 29°C rather than at 22°C (Zheng et al., 2019). JMJ13, along with ELF6 and REF6, influences the genome-wide distribution of H3K27me3, regulates the activation of tissue-specific genes (Pajoro et al., 2017), and plays a role in the regulation of a flowering pathway that also affects flowering in SD. elf6-1 showed an early flowering, whereas ref6-1 showed a late flowering (Noh et al., 2004). The jmj30/jmj32 double mutant exhibited an early flowering phenotype when cultivated under SD conditions at 29°C. JMJ30 has been demonstrated to directly binds to FLC, removing the inhibitory histone modification H3 lysine 27 trimethylation (H3K27me3) (Gan et al., 2014).
A. thaliana can release the inhibition of FT through the SVP-FLM and JMJ-FLC pathways, flowering earlier under the high-temperature conditions of SD.
3.5 Red light with FT
SENSITIVITY TO RED LIGHT REDUCED 1 (SRR1) is a protein with reduced sensitivity to red light, which was previously involved in the regulation of the circadian clock and PhyB signaling pathway in A. thaliana (Staiger et al., 2003). Mutant srr1-1 flowered early in SD (Johansson and Staiger, 2014). SRR1 suppresses FT expression and thus inhibits flowering under SD conditions by activating the expression of FT-binding repressors CDF1, TEM1, TEM2, and FLC (Johansson and Staiger, 2014).
Hypersensitivity to Red and Blue 1 (HRB1) mutant hrb1 flowered late, whereas HRB1 overexpressing line flowered early in SD (Kang et al., 2007), indicating that HRB1 promotes flowering in SD. The SD flowering phenotype of hrb1/phyB-9 was the same as that of phyB-9, and hrb1/cry2 showed a phenotype similar to that of hrb1 (Kang et al., 2007), suggesting that HRB1 mediates the regulation of flowering via red, but not blue, light signaling. hrb1/ft-2 flowered later than ft-2 (Kang et al., 2007), indicating that HRB1 promotes flowering in SD. The Phytochrome-signaling Early Flowering 1 (PEF1) mutant pef1, screened earlier in the SD early flowering mutant, showed an early flowering phenotype (Ahmad and Cashmore, 1996).
3.6 Other FT-dependent regulations
3.6.1 Other members of the phosphatidylethanolamine-binding protein family
FT is a member of the Phosphatidylethanolamine-binding Protein (PEBP) family, which comprises six members that can be categorized into three branches: FT-like, and TERMINAL FLOWER1 (TFL1)-like, MOTHER OF FT (MFT)-like (Chardon and Damerval, 2005). Another floral integrator, the TWIN SISTER OF FT (TSF) mutant tsf and its overexpression, was found in late and early flowering under SD conditions (Yamaguchi et al., 2005), although it is thought to function redundantly with FT in LD to promote flowering, and its specific mechanism under SD conditions is not clear. Mutant tfl flowered early under SD conditions and TFL negatively regulates the transcription of FD-dependent target genes, participating in the transcriptional repression of FT-activated genes (Hanano and Goto, 2011). To our knowledge, no study has reported the involvement of MFT in the regulation of flowering by SD.
3.6.2 The MADS-box family
Besides the MADS-box family members above mentioned, MADS AFFECTING FLOWERING1 (MAF1)/FLM/AGL27, MAF4, MAF5, AGL6, AGL19, AGL20, and AGL24family members play a pivotal role in the regulation of SD flowering.
The MAF1/FLM/AGL27 is responsible for the natural variation in the SD flowering time observed in certain ecotypes (Werner et al., 2005). The 35S::MAF1 construct was shown to flower approximately one month later (Ratcliffe et al., 2001). The VERNALIZATION INSENSITIVE 3-LIKE 1 (VIL1) mutant, vil1, flowered only late in SD, and VIL reduces the transcript levels of MAF1/FLM in SD (Sung et al., 2006). Furthermore, transcriptome changes induced by warm ambient temperatures in A.thaliana require VIL1, and its loss-of-function resulted in insensitivity to ambient temperatures (Sung et al., 2006; Kim et al., 2023).Histone deacetylation/acetylation plays a crucial role in maintaining genomic stability, regulating transcription, and influencing plant development. Histone acetylation is controlled by histone histone acetyltransferases and histone deacetylases (HDACs or HDAs) (Liu et al., 2014b).The HDACs family member, HDA5 mutant hda5-1, exhibited elevated FLC and MAF1 transcript levels under SD conditions, resulting in a late-flowering phenotype (Luo et al., 2015). Conversely, the HDA6 mutant axe1 displays late flowering under SD conditions and elevated MAF4 transcript levels (Luo et al., 2015). MAF5 repressed the VIN3-LIKE 2 (VIL2) gene to accelerate flowering under SD conditions, the vil2 mutant exclusively exhibits a late-flowering phenotype under SD conditions, but the maf5 mutant showed no obvious phenotype (Kim and Sung, 2010). Furthermore, the PRC1 RING-finger protein AtRING1A promotes flowering by suppressing MAF4 and MAF5 expression, which downregulates two floral integrators, FT and SUPPRESSOR OF OVEREXPRESSION OF CONSTANS 1 (SOC1). The atring1a mutant exhibited late flowering in SD (Shen et al., 2014). AGL6 has been demonstrated to negatively regulate FLC, MAF4, and MAF5 expression and positively regulate FT expression at the transcriptional level, promoting flowering in A.thaliana agl6-D (in which AGL6 is activated by the 35S enhancer). This results in early flowering in SD (Yoo et al., 2010). AGL19 has been identified as a negative regulator of msi1-cs. The AGL19 mutant agl19 flowered late in SD (Schönrock et al., 2006). SIN3-like proteins (SNLs) and their homologous protein MSI delay the flowering time of SNLs by repressing AGL19-regulated HDA9, snl2/3/4, and hda9 during early flowering in SD plants (Ning et al., 2019; Kim et al., 2013). Subsequent studies have demonstrated that HDA9 inhibits premature flowering under SD conditions by modifying the local chromatin environment and suppressing hyperactivation of the FT upstream activator AGL19 (Kang et al., 2015).The AGL20/SOC1 mutant exhibited delayed flowering time, whereas the SOC1 overexpression line 35S::AGL20-13 displayed an exceptionally early flowering time. SOC1 is a major floral integrator that integrates both developmental and environmental cues into floral genetic networks, but under SD conditions, SOC1 is only a minor target of GA signaling at the shoot meristem (Borner et al., 2008; Galvão et al., 2012). Another MADS-box gene, agl24, has been demonstrated to delays flowering under SD conditions by regulating SOC1 (Liu et al., 2008).
Therefore, in SD, the FT repressors MAF1, MAF4, and MAF5 are associated with delayed flowering, the FT activators AGL19 and ALG20 are associated with the promotion of flowering, and HDAs proteins are also actively involved in flowering through epigenetic modifications.
3.6.3 The BBX family
Besides CO, several members mutants of the BBX family also exhibited flowering phenotypes. Their function in regulating flowering is related to the FT in SD. BBX4/COL3 and BBX5/COL4 inhibit flowering, and upregulation of FT expression has been detected in both col3 and col4 (Datta et al., 2006; Steinbach, 2019). In contrast, an overexpression line of COL5 caused early flowering, and upregulation of FT levels was also detected; however, low levels of COL5 expression did not affect flowering in SD (Hassidim et al., 2009). BBX24/STO mutant sto-1 flowered late in SD, and the STO overexpression line STO-OE reduces the expression level of FLC, and at the same time, due to competition with FLC, the regulated downstream genes are not affected (Liu et al., 2014). Competition is a regulated downstream gene that activates FT and SOC1 expression (Liu et al., 2014). COL3 targets FT in the presence of BBX32 to regulate the flowering pathway, but both BBX32 overexpression lines BBX32-OX #5 and BBX32 artificial microRNA lines BX32-AMI #3 resulted in late flowering under SD conditions (Tripathi et al., 2016). Further studies are required to elucidate these underlying mechanisms.
4 Gibberellin-dependent regulation
In SD, rapid GA synthesis begins only when plants have been growing for a reproductive phase, and promotes flowering by facilitating floral meristem identity LEAFY (LFY) transcription via an independent FT pathway (Wilson et al., 1992; Eriksson et al., 2006). At the juvenile stage, plants inhibit flowering by suppressing the gibberellin response; at the adult phase, plants inhibit flowering by suppressing the gibberellin content; and at the critical stage of floral transition, flowering is promoted by rapidly increasing GA synthesis and response (Eriksson et al., 2006).
The exogenous application of GAs accelerates flowering in wild-type A. thaliana, particularly in SD (Langridge, 1957). Genetic analysis has suggested that GA has the most important function in flowering under SD conditions (Porri et al., 2012).
The GA1 gene encodes the first enzyme involved in GA biosynthesis and regulates GA biosynthesis at an early stage; ga1-3 did not flower in SD unless given exogenous GA, and weakly flowers late in LD (Wilson et al., 1992). ga1-3 had a significantly stronger effect on flowering under SD than under LD, possibly because the photoperiodic pathway masked the effects of gibberellins in ga1-3 under LD conditions (Reeves and Coupland, 2001). It has been shown that overexpression of SOC1 or simultaneous inactivation of two GA-responsive GATA transcription factors, GATA NITRATE-INDUCIBLE CARBONMETABOLISM INVOLVED (GNC) and GNC-LIKE/CYTOKININRESPONSIVE GATA FACTOR1 (GNL) could rescue the flowering phenotype of the ga1-3 plants in SD conditions (Moon et al., 2003; Richter et al., 2010). In contrast, SPINDLY (SPY) negatively regulates GA signaling, and spy-1 is flowered early in SD (Jacobsen and Olszewski, 1993). The enzyme GIBBERELLIN 2 OXIDASE 7 (GA2ox7) catabolizes active GAs, and transgenic plants SUCROSE TRANSPORTER 2 (SUC2):GA2ox7 or KNAT1:GA2ox7 were constructed to specifically express GA2ox7 in vascular or shoot apical meristems. flowered later than the wild type, and KNAT1:GA2ox7 flowered later, indicating that the role of GA in flowering under SD conditions is tissue specific, and that GA from shoot apical meristem tissues contributes more to flowering (Porri et al., 2012).
Three key GA signaling pathway components have been identified in A. thaliana: GA receptor GA INSENSITIVE DWARF1 (GID1), GA-response inhibitory protein factors DELLAs, and SLEEPY1 (SLY), of which GID1 and DELLAs have been reported to be associated with SD flowering; DELLAs inhibit all GA responses, whereas GID1 activates the GA response by binding and ubiquitinating DELLAs (Park et al., 2013).
DELLAs proteins play important roles as central regulatory nodes in the GA signaling pathway and are repressors that block GA signaling (Fleet and Sun, 2005). DELLAs proteins contain five members of the GRAS family of transcription factors: REPRESSOR OF GA1-3 (RGA), GA INSENSITIVE (GAI), RGA-LIKE 1 (RGL1), RGL2, and RGL3 (Sun and Gubler, 2004). gaiΔ17, rgaΔ17, rgl1Δ17, rgl2Δ17, and rgl3Δ17 are GA-insensitive lines, collectively referred to as dellaΔ17. Different dellaΔ17 proteins were identified by constructing pSUC2:dellaΔ17, pFD:dellaΔ17, and pCLV3:dellaΔ17, each of which has its own specific expression in the phloem, meristem, and shoot stem cell niches (Galvão et al., 2012). Under SD conditions, dellaΔ17 expression in the meristem delayed flowering or did not flower, whereas dellaΔ17 expression in the phloem and shoot stem had little effect on flowering time (Galvão et al., 2012). This differs from the finding that the expression of rgl3Δ17 in the shoot stem does not affect flowering in LD (Galvão et al., 2012). It is hypothesized that DELLAs proteins regulate flowering tissues differently in LD and SD, and that they function mainly at the meristem in SD, which is consistent with the tissues in which GA2ox7 exerts its function (Porri et al., 2012; Galvão et al., 2012; Yu et al., 2012).
DELLA proteins and BOTRYTIS SUSCEPTIBLE1 INTERACTOR (BOI), BOI-RELATED GENE1 (BRG1), BRG2, and BRG3 (collectively referred to as BOIs) repress the GA response by interacting with and binding to the promoters of responsive GA genes (Park et al., 2013). The BOIs quadruple mutant boiQ and the DELLA pentuple mutant dellaP both flowered early in SD, consistent with their function in repressing the GA response (Park et al., 2013). The bHLH transcription factor MYC3 stabilized by DELLAs in SD inhibits flowering by inhibiting CO binding to FT and myc3 early flowering in SD (Bao et al., 2019).
In addition, NO FLOWERING IN SD (NFL) is an obligate factor for the induction of flowering (Sharma et al., 2016) and promotes flowering by responding to gibberellins, which are active upstream of the GA signaling pathway; NFL belongs to the basic helix-loop-helix transcription factors, and its mutant, nfl, fails to flower in SD, but can flower by externally applying GA4 or by transferring nfl into the DELLA quadruple mutant rga/gai/rgl1/rgl2, and is therefore hypothesized to be a key transcription factor necessary for A. thaliana evolve into a parthenogenetic LD plant (Sharma et al., 2016).
HDC1 and HDA19 are directly responsible for HDAC and transcriptional repression of two flowering repressor genes in the gibberellin signaling pathway, GASA5 and GA2OX6, which together form a multi-subunit complex that regulates flowering; hdc1 and hda19 flowered early under SD conditions (Ning et al., 2019). The early flowering of svp-41 is associated with an increase in GA20ox2 mRNA, and it is possible that GAs progressively induce the expression of SOC1 under SD conditions, which represses SVP transcription and promotes flowering (Andrés et al., 2014). Therefore, gibberellin plays an important role in promoting flowering under SD conditions, and the meristem is the main tissue in which it exerts its promoting function.
5 Prospects
In this paper, we reviewed the mechanisms underlying delayed flowering under SD conditions (Figures 2A, B). First, in contrast to its function in promoting flowering in LD, CO inhibits flowering dependent on FT in SD, at least through the TFL. Second, A. thaliana inhibits flowering by repressing FT transcription via multiple pathways. Finally, plants inhibit flowering by suppressing the response to or synthesis of gibberellins at different times before floral transition. Nevertheless, our understanding of the molecular mechanisms underlying SD-mediated delay in flowering in A. thaliana remains incomplete. Therefore, further in-depth investigation should be conducted.
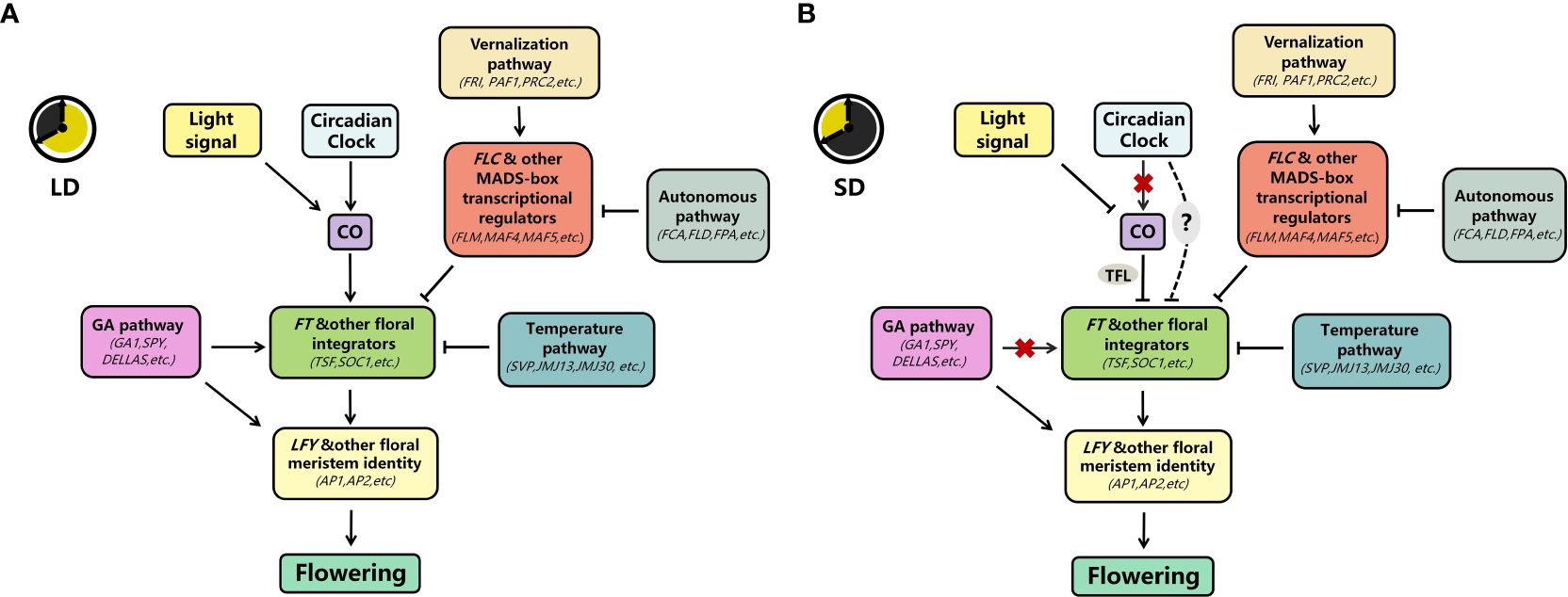
Figure 2. Key pathways for the photoperiodic regulation of flowering at 22°C (A) In LD, CO is mainly stabilized by light signals (PHYA, CRY1, etc.; see Figure 1 for details) and circadian clock signals to promote FT transcription and thus flowering under LD; the gibberellin pathway promotes flowering either through FT or directly through LFY; the vernalization and autonomous pathways mainly inhibit flowering by inhibiting FT transcription through FLC and other MADS-box transcriptional regulators; and the temperature pathway inhibits FT transcription/sensitivity at 22°C by SVP and others. (B) In SD, CO is mainly degraded by light signals (PHYB, see Figure 1 for details). The circadian clock does not promote CO transcription but represses FT transcription; the gibberellin pathway also does not promote FT transcription but directly promotes LFY transcription; the temperature pathway, vernalization pathway, and the autonomous pathway maintain a similar repression of FT transcription as LD under these conditions (→, promote; ⟞, inhabit; ‐‐‐, unclear;  , inactive).
, inactive).
5.1 Pathways and mechanisms involving CO
Typically, CO proteins in A. thaliana repress flowering in SD, as evidenced by the varying degrees of early flowering observed in several co-mutants (An et al., 2004; Datta et al., 2006; Redei, 1962; Balasubramanian et al., 2006; Fernandez et al., 2016; Luccioni et al., 2019; Ning et al., 2019; Yu et al., 2024; Zhang et al., 2015). The capacity of CO to bind to the FT promoter under SD conditions is constrained by competing factors (e.g., MYC3 and potentially others), and the activities of CO proteins that cannot bind to the FT promoter may be closely associated with the mechanism through which CO inhibits flowering. Recent studies have demonstrated that CO inhibits flowering via the TFL-FT pathway. However, it is noteworthy that the flowering phenotype of co-9/tfl was earlier than co-9 but later than tfl, indicating a competitive inhibition of flowering between CO and TFL. It has been postulated that additional factors may operate in a co-background to promote flowering. The early flowering observed in the PHYB mutants hy1 and hy3 depends on CO (Putterill et al., 1995). In addition, PHYB degrades CO proteins through ubiquitination. However, there is a lack of biochemical evidence confirming whether early flowering in PHYB under SD conditions is caused by CO accumulation. It is also noteworthy that mutants containing elevated levels of CO proteins, such as 35S:3HA-CO, cop1, and spa1-7, also exhibit early flowering phenotypes under SD conditions (Song et al., 2015; Laubinger et al., 2006; Jang et al., 2008). This differs from the function of CO in suppressing flowering under natural conditions, and further studies on the related mechanisms are required.
5.2 Mechanisms involving the FT pathway
The study of FT sensitivity is crucial for the inhibition of flowering by SD and the establishment of a precise quantification technique for plant sensitivity to FT is of paramount importance, cause FT levels cannot explain the similarity in flowering between LD and SD in some accessions (Kinmonth-Schultz et al., 2021). Studies have demonstrated that alterations in FT transcription within a specific range under SD conditions have a limited effect on flowering. Conversely, inducing FT expression during a period of heightened sensitivity to FT (ZT12-ZT20) is more likely to promote flowering in A. thaliana (Krzymuski et al., 2015). The mechanism underlying this period of heightened sensitivity depends on the circadian clock and remains to be elucidated. The sensitivity of the FT promoter can be regulated by its distance to its key elements, but the relevant trans-acting factor(s) and associated cis-element(s) remain unknown (Bao et al., 2019).
5.2.1 Role of circadian clock
Several mutants that affect key components of the circadian clock have been identified, including lhy-7, cca1-1, elf4, toc1-2, lwd1/lwd2, and cdf1/2/3/5 (Lin et al., 2019; Niwa et al., 2007; Park et al., 2016). Fornara et al. (2009) observed early flowering phenotypes and upregulation of FT transcription, indicating that the circadian clock pathway generally inhibits flowering under SD conditions by repressing FT transcription (Figure 1). Nevertheless, the precise mechanisms by which other components inhibit FT transcription and flowering remain unclear, except for LHY, which has been the subject of extensive research (Park et al., 2016).
5.2.2 Role of red light
Several mutant studies have indicated that red light plays a role in the regulation of SD light suppression during flowering. Early flowering of the red-light receptor PHYB mutant phyB-5 is not associated with FT transcription and is independent of the GA pathway (Blázquez and Weigel, 1999). However, the early flowering phenotype of srr1 and hrb1 mutants of the red-light pathway has been observed to suppress flowering by repressing FT transcription (Johansson and Staiger, 2014; Kang et al., 2007). This suggests that the mechanism by which red light inhibits flowering is complex and requires further investigation.
5.2.3 Mechanisms of FT inhibition of plant response to GA
Under SD conditions, GA promotes flowering independently of the FT pathway by binding to the GA response element in the LFY promoter and promoting its transcription (Blázquez et al., 1998; Eriksson et al., 2006). However, when GA4 was externally applied under SD conditions, ft-1 flowered earlier than Col (Wang et al., 2009) and the ft/tsf double mutant flowered later than Col (Porri et al., 2012), suggesting that deletion of FT increased GA sensitivity of the plants, whereas deletion of both FT and TSF decreased GA sensitivity of the plants; however, the mechanism remains to be investigated.
5.3 Mechanisms involved in the GA pathway
5.3.1 Unstudied key components of the GA pathway with flowering
The GA receptor GID1 has three homologous genes in A. thaliana: GID1a, GID1b, and GID1c, which are functionally redundant in regulating the GA signaling pathway (Griffiths et al., 2006). The gida-1/gidb-1/gidc-1 triple mutant does not exhibit flowering under LD conditions, continuous light, or gibberellic acid (GA3) (Griffiths et al., 2006). Although there is a paucity of data regarding the flowering of this mutant under SD conditions, it probably does not flower. Further studies are needed to verify this hypothesis. Moreover, the flowering function of SLY1, a pivotal component of the GA pathway, under SD conditions, remains to be elucidated.
5.3.2 Mechanism of co-regulation of flowering by GA and sucrose
Although GA4 plays a significant role in the flowering process, there was not a simple linear relationship between GA4 and LFY transcripts during the vegetative phase. Until the transition to the reproductive phase, when GA4 synthesis commences in substantial quantities and LFY transcripts increase, which may be associated with the varying levels of sucrose at different times (Eriksson et al., 2006), the mechanism by which GA and sucrose interact to stimulate flowering in SD remains uninvestigated.
5.4 Suggestions for crop breeding
Flowering is an important trait for improving crop yields (Carrera et al., 2024). A. thaliana, as a model plant, still has the ability to mine potential reference gene resources. In this article, it is summarized that A. thaliana achieves the suppression of FT under SD through multiple pathways, and the suppression of GA by the plant at the juvenile stage. So in the future, we can mine breeding resources from these two aspects in SD. Firstly, we can explore the genes that are the main regulators of FT in SD, such as mutants with extreme early flowering phenotypes, such as elf3, elf8,cry1-L407F(gain of function), etc, and mutants with flowering phenotypes only in SD, such as vil1, vip4-2, prr7-11, vip5-1, etc. Secondly, the research on the juvenile stage regulation of gibberellin in A. thaliana under SD condition is in-depth,exploring more primary repressors in this period is the most important task. Modulating them to access different seed resources through biotechnology (e.g., CRISPR-Cas) is more efficient.
Author contributions
YW: Writing – original draft, Writing – review & editing. TL: Conceptualization, Writing – review & editing, Supervision, Validation. YZ: Supervision, Writing – review & editing. TF: Supervision, Validation, Writing – review & editing. CT: Funding acquisition, Investigation, Supervision, Validation, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was supported by Natural Science Foundation of Guangdong Province(2020A1515011423, 2023A1515011645, 2024A1515013246) and the National Natural Science Foundation of China (31770342).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Ahmad, M., Cashmore, A. R. (1996). The pef mutants of arabidopsis thaliana define lesions early in the phytochrome signaling pathway. Plant Journal: For Cell And Mol. Biol. 10, 1103–1110. doi: 10.1046/j.1365-313X.1996.10061103.x
Airoldi, C. A., Mckay, M., Davies, B. (2015). Maf2 is regulated by temperaturedependent splicing and represses flowering at low temperatures in parallel with flm. PloS One 10. doi: 10.1371/journal.pone.0126516
An, H., Roussot, C., Suaarez-Lopez, P., Corbesier, L., Vincent, C., Piñeiro, M., et al. (2004). Constans acts in the phloem to regulate A systemic signal that induces photoperiodic flowering of arabidopsis. Development 131, 3615–3626. doi: 10.1242/dev.01231
Andrés, F., Coupland, G. (2012). The genetic basis of flowering responses to seasonal cues. Nat. Rev. Genet. 13, 627–639. doi: 10.1038/nrg3291
Andrés, F., Porri, A., Torti, S., Mateos, J., Romera-Branchat, M., García-Martínez, J. L., et al. (2014). Short vegetative phase reduces gibberellin biosynthesis at the arabidopsis shoot apex to regulate the floral transition. Proc. Of Natl. Acad. Of Sci. 111, E2760–E2769. doi: 10.1073/pnas.1409567111
Arongaus, A. B., Chen, S., Pireyre, M., Glöckner, N., Galvão, V. C., Albert, A., et al. (2018). Arabidopsis rup2 represses uvr8-mediated flowering in noninductive photoperiods. Genes Dev. 32, 1332–1343. doi: 10.1101/gad.318592.118
Aukerman, M. J., Sakai, H. (2003). Regulation of flowering time and floral organ identity by A microrna and its apetala2-like target genes. Plant Cell 15, 2730–2741. doi: 10.1105/tpc.016238
Balasubramanian, S., Sureshkumar, S., Lempe, J., Weigel, D. (2006). Potent induction of arabidopsis thaliana flowering by elevated growth temperature. PloS Genet. 2, 980–989. doi: 10.1371/journal.pgen.0020106
Bao, S., Hua, C., Huang, G., Cheng, P., Gong, X., Shen, L., et al. (2019). Molecular basis of natural variation in photoperiodic flowering responses. Dev. Cell 50, 90–101. doi: 10.1016/j.devcel.2019.05.018
Blázquez, M. A., Weigel, D. (1999). Independent regulation of flowering by phytochrome B and gibberellins in Arabidopsis. Plant Physiology 120, 1025–1032. doi: 10.1104/pp.120.4.1025
Blázquez, M. A., Green, R., Nilsson, O., Sussman, M. R., Weigel, D. (1998). Gibberellins promote flowering of Arabidopsis by activating the LEAFY promoter. The Plant cell 10, 791–800. doi: 10.1105/tpc.10.5.791
Borner, R., Kampmann, G., Chandler, J., Gleißner, R., Wisman, E., Apel, K., et al. (2008). A mads domain gene involved in the transition to flowering in arabidopsis. Plant J. 24, 591–599. doi: 10.1046/j.1365-313x.2000.00906.x
Boss, P. K., Bastow, R. M., Mylne, J. S., Dean, C. (2004). Multiple pathways in the decision to flower: Enabling, promoting, and resetting. Plant Cell 16, S18–S31. doi: 10.1105/tpc.015958
Brock, H. W., Fisher, C. L. (2005). Maintenance of gene expression patterns. Dev. Dynamics 232, 633–655. doi: 10.1002/dvdy.v232:3
Cao, Y., Wen, L., Wang, Z., Ma, L. (2015). Skip interacts with the paf1 complex to regulate flowering via the activation of flc transcription in arabidopsis. Mol. Plant 8, 1816–1819. doi: 10.1016/j.molp.2015.09.004
Carrera, C. S., Savin, R., Slafer, G. A. (2024). Critical period for yield determination across grain crops. Trends In Plant Sci. 29, 329–342. doi: 10.1016/j.tplants.2023.08.012
Chardon, F., Damerval, C. (2005). Phylogenomic analysis of the pebp gene family in cereals. J. Of Mol. Evol. 61, 579–590. doi: 10.1007/s00239-004-0179-4
Chen, X. (2004). A microrna as A translational repressor of apetala2 in arabidopsis flower development. Science 303, 2022–2025. doi: 10.1126/science.1088060
Chen, H., Huang, X., Gusmaroli, G., Terzaghi, W., Lau, O. S., Yanagawa, Y., et al. (2010). Arabidopsis cullin4-damaged dna binding protein 1 interacts with constitutively photomorphogenic1-suppressor of phya complexes to regulate photomorphogenesis and flowering time. Plant Cell 22, 108–123. doi: 10.1105/tpc.109.065490
Chen, C., Kim, D., Yun, H. R., Lee, Y. M., Yogendra, B., Bo, Z., et al. (2020). Nuclear import of like heterochromatin protein1 is redundantly mediated by importins α-1, α-2 and α-3. Plant J. 103, 1205–1214. doi: 10.1111/tpj.v103.3
Chen, Y., Song, S. Y., Gan, Y. B., Jiang, L. X., Yu, H., Shen, L. S. (2020). Shaggy-like kinase 12 regulates flowering through mediating constans stability in arabidopsis. Sci. Adv. 6, eaaw0413. doi: 10.1126/sciadv.aaw0413
Chou, M. L., Yang, C. H. (1998). FLD interacts with genes that affect different developmental phase transitions to regulate arabidopsis shoot development. Plant J. 15, 231–242. doi: 10.1046/j.1365-313X.1998.00204.x
Datta, S., Hettiarachchi, G. H. C. M., Deng, X. W., Holm, M. (2006). Arabidopsis constans-like3 is A positive regulator of red light signaling and root growth. Plant Cell 18, 70–84. doi: 10.1105/tpc.105.038182
Doyle, M. R., Davis, S. J., Bastow, R. M., Mcwatters, H. G., Kozma-Bognár, L., Nagy, F., et al. (2002). The elf4 gene controls circadian rhythms and flowering time in arabidopsis thaliana. Nature 419, 74–77. doi: 10.1038/nature00954
Endo, M., Mochizuki, N., Suzuki, T., Nagatani, A. (2007). Cryptochrome2 in vascular bundles regulates flowering in arabidopsis. Plant Cell 19, 84–93. doi: 10.1105/tpc.106.048157
Eriksson, S., Moritz, T., Nilsson, O. (2006). Ga4 is the active gibberellin in the regulation of leafy transcription and arabidopsis floral initiation. Plant Cell 18, 2172–2181. doi: 10.1105/tpc.106.042317
Exner, V., Alexandre, C., Rosenfeldt, G., Alfarano, P., Nater, M., Caflisch, A., et al. (2010). A gain-of-function mutation of arabidopsis cryptochrome1 promotes flowering. Plant Physiol. 154, 1633–1645. doi: 10.1104/pp.110.160895
Fernandez, V., Takahashi, Y., Le Gourrierec, J., Coupland, G. (2016). Photoperiodic and thermosensory pathways interact through constans to promote flowering at high temperature under short days. Plant J. 86, 426–440. doi: 10.1111/tpj.13183
Fleet, C. M., Sun, T. P. (2005). A dellacate balance: the role of gibberellin in plant morphogenesis. Curr. Opin. In Plant Biol. 8, 77–85. doi: 10.1016/j.pbi.2004.11.015
Fornara, F., De Montaigu, A., Coupland, G. (2010). Snapshot: control of flowering in arabidopsis. Cell 141, 550–550.E2. doi: 10.1016/j.cell.2010.04.024
Fornara, F., Panigrahi, K. C. S., Gissot, L., Sauerbrunn, N., Rühl, M., Jarillo, J. A., et al. (2009). Arabidopsis dof transcription factors act redundantly to reduce constans expression and are essential for A photoperiodic flowering response. Dev. Cell 17, 75–86. doi: 10.1016/j.devcel.2009.06.015
Galvão, V. C., Horrer, D., Küttner, F., Schmid, M. (2012). Spatial control of flowering by della proteins in arabidopsis thaliana. Development 139, 4072–4082. doi: 10.1242/dev.080879
Gan, E. S., Xu, Y., Wong, J. Y., Geraldine Goh, J., Sun, B., Wee, W. Y., et al. (2014). Jumonji demethylases moderate precocious flowering at elevated temperature via regulation of flc in arabidopsis. Nat. Commun. 5, 5098. doi: 10.1038/ncomms6098
Griffiths, J., Murase, K., Rieu, I., Zentella, R., Zhang, Z. L., Powers, S. J., et al. (2006). Genetic characterization and functional analysis of the GID1 gibberellin receptors in Arabidopsis. Plant Cell 19, 726–726. doi: 10.1105/tpc.107.190261
Hanano, S., Goto, K. (2011). Arabidopsis terminal flower1 is involved in the regulation of flowering time and inflorescence development through transcriptional repression. Plant Cell 23, 3172–3184. doi: 10.1105/tpc.111.088641
Hassidim, M., Harir, Y., Yakir, E., Kron, I., Green, R. M. (2009). Over-expression of constans-like 5 can induce flowering in short-day grown arabidopsis. Planta 230, 481–491. doi: 10.1007/s00425-009-0958-7
Hayama, R., Sarid-Krebs, L., Richter, R., Fernández, V., Jang, S., Coupland, G. (2017). Pseudo response regulators stabilize constans protein to promote flowering in response to day length. EMBO J. 36, 904–918. doi: 10.15252/embj.201693907
He, Y., Doyle, M. R., Amasino, R. M. (2004). Paf1-complex-mediated histone methylation of flowering locus C chromatin is required for the vernalization-responsive, winter-annual habit in arabidopsis. Genes Dev. 18, 2774–2784. doi: 10.1101/gad.1244504
Helfer, A., Nusinow, D. A., Chow, B. Y., Gehrke, A. R., Bulyk, M. L., Kay, S. A. (2011). Lux arrhythmo encodes A nighttime repressor of circadian gene expression in the arabidopsis core clock. Curr. Biol. 21, 126–133. doi: 10.1016/j.cub.2010.12.021
Herrero, E., Kolmos, E., Bujdoso, N., Yuan, Y., Wang, M., Berns, M. C., et al. (2012). Early flowering4 recruitment of early flowering3 in the nucleus sustains the arabidopsis circadian clock. Plant Cell 24, 428–443. doi: 10.1105/tpc.111.093807
Huang, H., Yoo, C. Y., Bindbeutel, R., Goldsworthy, J., Tielking, A., Alvarez, S., et al. (2016). Pch1 integrates circadian and light-signaling pathways to control photoperiod-responsive growth in arabidopsis. Elife 5, e13292. doi: 10.7554/eLife.13292.033
Hung, F. Y., Lai, Y. C., Wang, J., Feng, Y. R., Shih, Y. H., Chen, J. H., et al. (2021). The arabidopsis histone demethylase jmj28 regulates constans by interacting with fbh transcription factors. Plant Cell 33, 1196–1211. doi: 10.1093/plcell/koab014
Hwang, D. Y., Park, S., Lee, S., Lee, S. S., Imaizumi, T., Song, Y. H. (2019). Gigantea regulates the timing stabilization of constans by altering the interaction between fkf1 and zeitlupe. Molecules And Cells 42, 693–701. doi: 10.14348/molcells.2019.0199
Imaizumi, T. (2010). Arabidopsis circadian clock and photoperiodism: time to think about location. Curr. Opin. In Plant Biol. 13, 83–89. doi: 10.1016/j.pbi.2009.09.007
Imaizumi, T., Tran, H. G., Swartz, T. E., Briggs, W. R., Kay, S. A. (2003). Fkf1 is essential for photoperiodic-specific light signalling in. Nature 426, 302–306. doi: 10.1038/nature02090
Ito, S., Niwa, Y., Nakamichi, N., Kawamura, H., Yamashino, T., Mizuno, T. (2008). Insight into missing genetic links between two evening-expressed pseudo-response regulator genes toc1 and prr5 in the circadian clock-controlled circuitry in arabidopsis thaliana. Plant And Cell Physiol. 49, 201–213. doi: 10.1093/pcp/pcm178
Ito, S., Song, Y. H., Josephson-Day, A. R., Miller, R. J., Breton, G., Olmstead, R. G., et al. (2012). Flowering bhlh transcriptional activators control expression of the photoperiodic flowering regulator constans in arabidopsis. Proc. Of Natl. Acad. Of Sci. 109, 3582–3587. doi: 10.1073/pnas.1118876109
Jacobsen, S. E., Olszewski, N. E. (1993). Mutations at the spindly locus of arabidopsis alter gibberellin signal transduction. Plant Cell 5, 887–896. doi: 10.1105/tpc.5.8.887
Jang, S., Marchal, V., Panigrahi, K. C. S., Wenkel, S., Soppe, W., Deng, X. W., et al. (2008). Arabidopsis cop1 shapes the temporal pattern of co accumulation conferring A photoperiodic flowering response. EMBO J. 27, 1277–1288. doi: 10.1038/emboj.2008.68
Johanson, U., West, J., Lister, C., Michaels, S., Amasino, R., Dean, C. (2000). Molecular analysis of frigida, A major determinant of natural variation in arabidopsis flowering time. Science 290, 344–347. doi: 10.1126/science.290.5490.344
Johansson, M., Staiger, D. (2014). Srr1 is essential to repress flowering in non-inductive conditions in arabidopsis thaliana. J. Of Exp. Bot. 65, 5811–5822. doi: 10.1093/jxb/eru317
Johnson, E., Bradley, M., Np, H., Gc, W. (1994). Photoresponses of light-grown phya mutants of arabidopsis (Phytochrome A is required for the perception of daylength extensions). Plant Physiol. 105, 141–149. doi: 10.1104/pp.105.1.141
Kamioka, M., Takao, S., Suzuki, T., Taki, K., Higashiyama, T., Kinoshita, T., et al. (2016). Direct repression of evening genes by circadian clock-associated1 in the arabidopsis circadian clock. Plant Cell 28, 696–711. doi: 10.1105/tpc.15.00737
Kang, M. Y., Kwon, H. Y., Kim, N. Y., Sakuraba, Y., Paek, N. C. (2015). Constitutive photomorphogenic 10 (Cop10) contributes to floral repression under non-inductive short days in arabidopsis. Int. J. Of Mol. Sci. 16, 26493–26505. doi: 10.3390/ijms161125969
Kang, X., Zhou, Y., Sun, X., Ni, M. (2007). Hypersensitive to red and blue 1 and its C-terminal regulatory function control flowering locus T expression. Plant J. 52, 937–948. doi: 10.1111/j.1365-313X.2007.03295.x
Kim, J., Bordiya, Y., Xi, Y., Zhao, B., Kim, D.-H., Pyo, Y., et al. (2023). Warm temperature-triggered developmental reprogramming requires vil1-mediated, genome-wide H3k27me3 accumulation in arabidopsis. Development 150. doi: 10.1242/dev.201343
Kim, S. Y., He, Y., Jacob, Y., Noh, Y. S., Michaels, S., Amasino, R. (2005). Establishment of the vernalization-responsive, winter-annual habit in arabidopsis requires A putative histone H3 methyl transferase. Plant Cell 17, 3301–3310. doi: 10.1105/tpc.105.034645
Kim, Y., Lim, J., Yeom, M., Kim, H., Kim, J., Wang, L., et al. (2013). Elf4 regulates gigantea chromatin access through subnuclear sequestration. Cell Rep. 3, 671–677. doi: 10.1016/j.celrep.2013.02.021
Kim, S. Y., Michaels, S. D. (2006). Suppressor of fri 4 encodes A nuclear-localized protein that is required for delayed flowering in winter-annual arabidopsis. Development 133, 4699–4707. doi: 10.1242/dev.02684
Kim, D. H., Sung, S. (2010). The plant homeo domain finger protein, vin3-like 2, is necessary for photoperiod-mediated epigenetic regulation of the floral repressor, maf5. Proc. Of Natl. Acad. Of Sci. 107, 17029–17034. doi: 10.1073/pnas.1010834107
Kinmonth-Schultz, H., Lewandowska-Sabat, A., Imaizumi, T., Ward, J. K., Rognli, O. A., Fjellheim, S. (2021). Flowering times of wild arabidopsis accessions from across Norway correlate with expression levels of ft, co, and flc genes. Front. In Plant Sci. 12. doi: 10.3389/fpls.2021.747740
Kinmonth-Schultz, H., Snsteb, J. H., Croneberger, A. J., Johnsen, S. S., Leder, E., Lewandowska-Sabat, A., et al. (2023). Responsiveness to long days for flowering is reduced in arabidopsis by yearly variation in growing season temperatures. Plant Cell Environ. 46, 3337–3352. doi: 10.1111/pce.v46.11
Koornneef, M., Alonso-Blanco, C., Peeters, A. J. M., Soppe, W. (1998). Genetic control of flowering time in arabidopsis. Annu. Rev. Of Plant Physiol. And Plant Mol. Biol. 49, 345–370. doi: 10.1146/annurev.arplant.49.1.345
Koornneef, M., Hanhart, C. J., Veen, J. H. V. D. (1991). A genetic and physiological analysis of late flowering mutants in arabidopsis thaliana. Mol. Gen. Genet. 229, 57–66. doi: 10.1007/BF00264213
Krzymuski, M., Andrés, F., Cagnola, J. I., Jang, S., Yanovsky, M. J., Coupland, G., et al. (2015). The dynamics of FLOWERING LOCUS T expression encodes long-day information. Plant Journal 83, 952–961. doi: 10.1111/tpj.12938
Kwak, J. S., Son, G. H., Kim, S. I., Song, J. T., Seo, H. S. (2016). Arabidopsis high ploidy2 sumoylates and stabilizes flowering locus C through its E3 ligase activity. Front. In Plant Sci. 7. doi: 10.3389/fpls.2016.00530
Lai, B.-W., Chen, L., Lu, S.-J. (2023). The current status of photoperiod adaptability in soybean. Hereditas 45, 793–800. doi: 10.16288/j.yczz.23-200
Langridge, J. (1957). Effect of day-length and gibberellic acid on the flowering of arabidopsis. Nature 180, 36–37. doi: 10.1038/180036a0
Latrasse, D., Germann, S., Houba-Hérin, N., Dubois, E., Bui-Prodhomme, D., Hourcade, D., et al. (2011). Control of flowering and cell fate by lif2, an rna binding partner of the polycomb complex component lhp1. PloS One 6, e16592. doi: 10.1371/journal.pone.0016592
Laubinger, S., Marchal, V., Gentilhomme, J., Wenkel, S., Adrian, J., Jang, S., et al. (2006). Arabidopsis spa proteins regulate photoperiodic flowering and interact with the floral inducer constans to regulate its stability. Development 133, 4608–4608. doi: 10.1242/dev.02481
Lazaro, A., Mouriz, A., Piñeiro, M., Jarillo, J. A. (2015). Red light-mediated degradation of constans by the E3 ubiquitin ligase hos1 regulates photoperiodic flowering in arabidopsis. Plant Cell 27, 2437–2454. doi: 10.1105/tpc.15.00529
Lazaro, A., Valverde, F., Piñeiro, M., Jarillo, J. A. (2012). The E3 Ubiquitin Ligase HOS1 Negatively Regulates CONSTANS Abundance in the Photoperiodic Control of Flowering. Plant Cell 24, 982–999. doi: 10.1105/tpc.110.081885
Lee, I., Aukerman, M. J., Gore, S. L., Lohman, K. N., Michaels, S. D., Weaver, L. M., et al. (1994). Lsolation of luminidependens: A gene lnvolved in the control of flowering time in arabidopsis. Plant Cell 6, 75–83. doi: 10.1105/tpc.6.1.75
Lin, K., Zhao, H., Gan, S., Li, G. (2019). Arabidopsis elf4-like proteins efl1 and efl3 influence flowering time. Gene 700, 131–138. doi: 10.1016/j.gene.2019.03.047
Liu, L., Adrian, J., Pankin, A., Hu, J., Dong, X., Von Korff, M., et al. (2014). Induced and natural variation of promoter length modulates the photoperiodic response of flowering locus T. Nat. Commun. 5, 4558. doi: 10.1038/ncomms5558
Liu, C., Chen, H., Er, H. L., Soo, H. M., Kumar, P. P., Han, J. H., et al. (2008). Direct interaction of agl24 and soc1 integrates flowering signals in arabidopsis. Development 135, 1481–1491. doi: 10.1242/dev.020255
Liu, T. L., Newton, L., Liu, M. J., Shiu, S. H., Farré, E. M. (2016). A G-box-like motif is necessary for transcriptional regulation by circadian pseudo-response regulators in arabidopsis. Plant Physiol. 170, 528–539. doi: 10.1104/pp.15.01562
Liu, X., Yang, S., Zhao, M., Luo, M., Yu, C. W., Chen, C. Y., et al. (2014). Transcriptional repression by histone deacetylases in plants. Mol. Plant 7, 764–772. doi: 10.1093/mp/ssu033
Luccioni, L., Krzymuski, M., Sánchez-Lamas, M., Karayekov, E., Cerdán, P. D., Casal, J. J. (2019). Constans delays arabidopsis flowering under short days. Plant J. 97, 923–932. doi: 10.1111/tpj.2019.97.issue-5
Luo, M., Tai, R., Yu, C. W., Yang, S., Chen, C. Y., Lin, W. D., et al. (2015). Regulation of flowering time by the histone deacetylase hda5 in arabidopsis. Plant J. 82, 925–936. doi: 10.1111/tpj.2015.82.issue-6
Maple, R., Zhu, P., Hepworth, J., Wang, J. W., Dean, C. (2024). Flowering time: from physiology, through genetics to mechanism. Plant Physiol. 195, 190–212. doi: 10.1093/plphys/kiae109
Martin-Trillo, M., Lázaro, A., Poethig, R. S., Gómez-Mena, C. N., Piñeiro, M. A., Martinez-Zapater, J. M., et al. (2006). Early in short days 1(Esd1) encodes actin-related protein 6 (Atarp6), A putative component of chromatin remodelling complexes that positively regulates flc accumulation in arabidopsis. Development 133, 1241–1252. doi: 10.1242/dev.02301
Mcnellis, T. W., Von Arnim, A. G., Araki, T., Komeda, Y., Misera, S., Deng, X. W. (1994). Genetic and molecular analysis of an allelic series of cop1 mutants suggests functional roles for the multiple protein domains. Plant Cell 6, 487–500. doi: 10.1105/tpc.6.4.487
Michaels, S. D., He, Y. H., Scortecci, K. C., Amasino, R. M. (2003). Attenuation of flowering locus C activity as A mechanism for the evolution of summer-annual flowering behavior in arabidopsis. Proc. Of Natl. Acad. Of Sci. Of United States Of America 100, 10102–10107. doi: 10.1073/pnas.1531467100
Mizoguchi, T., Wheatley, K., Hanzawa, Y., Wright, L., Mizoguchi, M., Song, H. R., et al. (2002). Lhy and cca1 are partially redundant genes required to maintain circadian rhythms in arabidopsis. Dev. Cell 2, 629–641. doi: 10.1016/S1534-5807(02)00170-3
Moon, J., Suh, S. S., Lee, H., Choi, K. R., Hong, C. B., Paek, N. C., et al. (2003). The soc1 mads-box gene integrates vernalization and gibberellin signals for flowering in arabidopsis. Plant J. 35, 613–623. doi: 10.1046/j.1365-313X.2003.01833.x
Morris, K., Thornber, S., Codrai, L., Richardson, C., Craig, A., Sadanandom, A., et al. (2010). Day neutral flowering represses constans to prevent arabidopsis flowering early in short days. Plant Cell 22, 1118–1128. doi: 10.1105/tpc.109.066605
Nagel, D. H., Doherty, C. J., Pruneda-Paz, J. L., Schmitz, R. J., Ecker, J. R., Kay, S. A. (2015). Genome-wide identification of cca1 targets uncovers an expanded clock network in arabidopsis. Proc. Of Natl. Acad. Of Sci. 112, E4802–E4810. doi: 10.1073/pnas.1513609112
Nakamichi, N., Kiba, T., Henriques, R., Mizuno, T., Chua, N. H., Sakakibara, H. (2010). Pseudo-response regulators 9, 7, and 5 are transcriptional repressors in the arabidopsis circadian clock. Plant Cell 22, 594–605. doi: 10.1105/tpc.109.072892
Nakamichi, N., Kiba, T., Kamioka, M., Suzuki, T., Yamashino, T., Higashiyama, T., et al. (2012). Transcriptional repressor prr5 directly regulates clock-output pathways. Proc. Of Natl. Acad. Of Sci. 109, 17123–17128. doi: 10.1073/pnas.1205156109
Nakamichi, N., Kita, M., Niinuma, K., Ito, S., Yamashino, T., Mizoguchi, T., et al. (2007). Arabidopsis clock-associated pseudo-response regulators prr9, prr7 and prr5 coordinately and positively regulate flowering time through the canonical constans-dependent photoperiodic pathway. Plant And Cell Physiol. 48, 822–832. doi: 10.1093/pcp/pcm056
Ning, Y. Q., Chen, Q., Lin, R. N., Li, Y. Q., Li, L., Chen, S., et al. (2019). The hda19 histone deacetylase complex is involved in the regulation of flowering time in A photoperiod-dependent manner. Plant J. 98, 448–464. doi: 10.1111/tpj.2019.98.issue-3
Niwa, Y., Ito, S., Nakamichi, N., Mizoguchi, T., Niinuma, K., Yamashino, T., et al. (2007). Genetic linkages of the circadian clock-associated genes, toc1, cca1 and lhy, in the photoperiodic control of flowering time in arabidopsis thaliana. Plant And Cell Physiol. 48, 925–937. doi: 10.1093/pcp/pcm067
Noh, B., Lee, S. H., Kim, H. J., Yi, G., Shin, E. A., Lee, M., et al. (2004). Divergent roles of A pair of homologous jumonji/zinc-finger–class transcription factor proteins in the regulation of arabidopsis flowering time. Plant Cell 16, 2601–2613. doi: 10.1105/tpc.104.025353
Nusinow, D. A., Helfer, A., Hamilton, E. E., King, J. J., Imaizumi, T., Schultz, T. F., et al. (2011). The elf4–elf3–lux complex links the circadian clock to diurnal control of hypocotyl growth. Nature 475, 398–402. doi: 10.1038/nature10182
Oh, S., Zhang, H., Ludwig, P., van Nocker, S. (2004). A mechanism related to the yeast transcriptional regulator Paf1c is required for expression of the Arabidopsis FLC/MAF MADS box gene family. Plant Cell 16, 2940–2953. doi: 10.1105/tpc.110.081885
Pajoro, A., Severing, E., Angenent, G. C., Immink, R. G. H. (2017). Histone H3 lysine 36 methylation affects temperature-induced alternative splicing and flowering in plants. Genome Biol. 18, 102. doi: 10.1186/s13059-017-1235-x
Park, M. J., Kwon, Y. J., Gil, K. E., Park, C. M. (2016). Late elongated hypocotyl regulates photoperiodic flowering via the circadian clock in arabidopsis. BMC Plant Biol. 16, 114. doi: 10.1186/s12870-016-0810-8
Park, J., Nguyen, K. T., Park, E., Jeon, J. S., Choi, G. (2013). Della proteins and their interacting ring finger proteins repress gibberellin responses by binding to the promoters of A subset of gibberellin-responsive genes in. Arabidopsis Plant Cell 25, 927–943. doi: 10.1105/tpc.112.108951
Porri, A., Torti, S., Romera-Branchat, M., Coupland, G. (2012). Spatially distinct regulatory roles for gibberellins in the promotion of flowering of arabidopsis under long photoperiods. Development 139, 2198–2209. doi: 10.1242/dev.077164
Pose, D., Verhage, L., Ott, F., Yant, L., Mathieu, J., Angenent, G. C., et al. (2013). Temperature-dependent regulation of flowering by antagonistic flm variants. Nature 503, 414–417. doi: 10.1038/nature12633
Putterill, J., Robson, F., Lee, K., Coupland, G. (1993). Chromosome walking with yac clones in arabidopsis: isolation of 1700 kb of contiguous dna on chromosome 5, including A 300 kb region containing the flowering-time gene co. Mol. Gen. Genet. 239, 145–157. doi: 10.1007/BF00281613
Putterill, J., Robson, F., Lee, K., Simon, R., Coupland, G. (1995). The constans gene of arabidopsis promotes flowering and encodes A protein showing similarities to zinc finger transcription factors. Cell 80, 847–857. doi: 10.1016/0092-8674(95)90288-0
Ratcliffe, O. J., Nadzan, G. C., Reuber, T. L., Riechmann, J. L. (2001). Regulation of flowering in Arabidopsis by an FLC homologue. Plant Physiology 126, 122–132. doi: 10.1104/pp.126.1.122
Redei, G. P. (1962). Supervital mutants of arabidopsis. Genetics 47, 443–460. doi: 10.1093/genetics/47.4.443
Reed, J. W., Nagatani, A., Elich, T. D., Fagan, M., Chory, J. (1994). Phytochrome A and phytochrome B have overlapping but distinct functions in arabidopsis development. Plant Physiol. 104, 1139–1149. doi: 10.1104/pp.104.4.1139
Reeves, P. H., Coupland, G. (2001). Analysis of flowering time control in arabidopsis by comparison of double and triple mutants. Plant Physiol. 126, 1085–1091. doi: 10.1104/pp.126.3.1085
Richter, R., Behringer, C., Müller, I. K., Schwechheimer, C. (2010). The gata-type transcription factors gnc and gnl/cga1 repress gibberellin signaling downstream from della proteins and phytochrome-interacting factors. Genes Dev. 24, 2093–2104. doi: 10.1101/gad.594910
Robson, F., Costa, M. M. R., Hepworth, S. R., Vizir, I., Piñeiro, M., Reeves, P. H., et al. (2001). Functional importance of conserved domains in the flowering-time gene constans demonstrated by analysis of mutant alleles and transgenic plants. Plant J. 28, 619–631. doi: 10.1046/j.1365-313x.2001.01163.x
Sawa, M., Kay, S. A. (2011). Gigantea directly activates flowering locus T in arabidopsis thaliana. Proc. Of Natl. Acad. Of Sci. 108, 11698–11703. doi: 10.1073/pnas.1106771108
Sawa, M., Nusinow, D. A., Kay, S. A., Imaizumi, T. (2007). Fkf1 and gigantea complex formation is required for day-length measurement in arabidopsis. Science 318, 261–265. doi: 10.1126/science.1146994
Schönrock, N., Bouveret, R., Leroy, O., Borghi, L., Köhler, C., Gruissem, W., et al. (2006). Polycomb-group proteins repressthe floral activator agl19 in the flc-independent vernalization pathway. Genes Dev. 20, 1667–1678. doi: 10.1101/gad.377206
Schroeder, D. F., Gahrtz, M., Maxwell, B. B., Cook, R. K., Kan, J. M., Alonso, J. M., et al. (2002). De-etiolated 1 and damaged dna binding protein 1 interact to regulate arabidopsis photomorphogenesis. Curr. Biol. 12, 1462–1472. doi: 10.1016/S0960-9822(02)01106-5
Sharma, N., Xin, R., Kim, D.-H., Sung, S., Lange, T., Huq, E. (2016). No flowering in short day (Nfl) is A bhlh transcription factor that promotes flowering specifically under short-day in arabidopsis. Development., 682–690. doi: 10.1242/dev.128595
Shen, L., Thong, Z., Gong, X., Shen, Q., Gan, Y., Yu, H. (2014). The putative prc1 ring-finger protein atring1a regulates flowering through repressing mads affecting flowering genes in arabidopsis. Development 141, 1303–1312. doi: 10.1242/dev.104513
Shim, J. S., Kubota, A., Imaizumi, T. (2017). Circadian clock and photoperiodic flowering in arabidopsis: constans is A hub for signal integration. Plant Physiol. 173, 5–15. doi: 10.1104/pp.16.01327
Shindo, C., Aranzana, M. J., Lister, C., Baxter, C., Nicholls, C., Nordborg, M., et al. (2005). Role of frigida and flowering locus C in determining variation in flowering time of arabidopsis. Plant Physiol. 138, 1163–1173. doi: 10.1104/pp.105.061309
Song, Y. H., Shim, J. S., Kinmonth-Schultz, H. A., Imaizumi, T. (2015). Photoperiodic flowering: time measurement mechanisms in leaves. Annu. Rev. Of Plant Biol. 66, 441–464. doi: 10.1146/annurev-arplant-043014-115555
Song, Y. H., Smith, R. W., To, B. J., Millar, A. J., Imaizumi, T. (2012). Fkf1 conveys timing information for constans stabilization in photoperiodic flowering. Science 336, 1045–1049. doi: 10.1126/science.1219644
Song, Y. H.Kubota, A.Kwon, M. S.Covington, M. F.Lee, N.Taagen, E. R.. (2018). Molecular basis of flowering under natural long-day conditions in Arabidopsis. Nature Plants 4, 824–835. doi: 10.1038/s41477-018-0253-3
Song, Y. H., Estrada, D. A., Johnson, R. S., Kim, S. K., Lee, S. Y., Maccoss, M. J., et al (2014). Distinct roles of FKF1, GIGANTEA, and ZEITLUPE proteins in the regulation of CONSTANS stability in Arabidopsis photoperiodic flowering. Proceedings of the National Academy of Sciences of the United States of America 111, 17672–17677. doi: 10.1073/pnas.1415375111
Staiger, D., Allenbach, L., Salathia, N., Fiechter, V., Davis, S. J., Millar, A. J., et al. (2003). The arabidopsis srr1 gene mediates phyb signaling and is required for normal circadian clock function. Genes Dev. 17, 256–268. doi: 10.1101/gad.244103
Steinbach, Y. (2019). The arabidopsis thalianaconstans-like 4 (Col4) – A modulator of flowering time. Front. In Plant Sci. 10. doi: 10.3389/fpls.2019.00651
Suarez-Lopez, P., Wheatley, K., Robson, F., Onouchi, H., Valverde, F., Coupland, G. (2001). Constans mediates between the circadian clock and the control of flowering in arabidopsis. Nature 410, 1116–1120. doi: 10.1038/35074138
Sun, T. P., Gubler, F. (2004). Molecular mechanism of gibberellin signaling in plants. Annu. Rev. Of Plant Biol. 55, 197–223. doi: 10.1146/annurev.arplant.55.031903.141753
Sung, S., Schmitz, R. J., Amasino, R. M. (2006). A phd finger protein involved in both the vernalization and photoperiod pathways in arabidopsis. Genes Dev. 20, 3244–3248. doi: 10.1101/gad.1493306
Takagi, H., Hempton, A. K., Imaizumi, T. (2023). Photoperiodic flowering in arabidopsis: multilayered regulatory mechanisms of constans and the florigen flowering locus T. Plant Commun. 4, 100552. doi: 10.1016/j.xplc.2023.100552
Takase, T., Nishiyama, Y., Tanihigashi, H., Ogura, Y., Miyazaki, Y., Yamada, Y., et al. (2011). Lov kelch protein2 and zeitlupe repress arabidopsis photoperiodic flowering under non-inductive conditions, dependent on flavin-binding kelch repeat F-box1. Plant J. 67, 608–621. doi: 10.1111/j.1365-313X.2011.04618.x
Tripathi, P., Carvallo, M., Hamilton, E. E., Preuss, S., Kay, S. A. (2016). Arabidopsis B-box32 interacts with constans-like3 to regulate flowering. Proc. Of Natl. Acad. Of Sci. 114, 172–177. doi: 10.1073/pnas.1616459114
Valverde, F., Mouradov, A., Soppe, W., Ravenscroft, D., Samach, A., Coupland, G. (2004). Photoreceptor regulation of constans protein in photoperiodic flowering. Science 303, 1003–1006. doi: 10.1126/science.1091761
Werner, J. D., Borevitz, J. O., Warthmann, N., Trainer, G. T., Ecker, J. R., Chory, J., et al. (2005). Quantitative trait locus mapping and dna array hybridization identify an flm deletion as A cause for natural flowering-time variation. Proc. Of Natl. Acad. Of Sci. Of United States Of America 102, 2460–2465. doi: 10.1073/pnas.0409474102
Wang, J. W., Czech, B., Weigel, D. (2009). miR156-Regulated SPL transcription factors define an endogenous flowering pathway in Arabidopsis thaliana. Cell 138, 738–749. doi: 10.1016/j.cell.2009.06.014
Whittaker, C., Dean, C. (2017). The flc locus: A platform for discoveries in epigenetics and adaptation. Annu. Rev. Of Cell And Dev. Biol. 33, 555–575. doi: 10.1146/annurev-cellbio-100616-060546
Wilson, R. N., Heckman, J. W., Somerville, C. R. (1992). Gibberellin is required for flowering in arabidopsis thaliana under short days. Plant Physiol. 100, 403–408. doi: 10.1104/pp.100.1.403
Wu, J. F., Wang, Y., Wu, S. H. (2008). Two new clock proteins, lwd1 and lwd2, regulate arabidopsis photoperiodic flowering. Plant Physiol. 148, 948–959. doi: 10.1104/pp.108.124917
Xing, J., Zhang, J., Yang, P., Jiang, C., Fan, J., Han, J., et al. (2014). Sdr6 is involved in regulation of flowering time in arabidopsis thaliana. Plant Biotechnol. 31, 133–139. doi: 10.5511/plantbiotechnology.14.0125b
Yamaguchi, A., Kobayashi, Y., Goto, K., Abe, M., Araki, T. (2005). Twin sister of ft (Tsf) acts as A floral pathway integrator redundantly with ft. Plant And Cell Physiol. 46, 1175–1189. doi: 10.1093/pcp/pci151
Yoo, S. K., Wu, X., Lee, J. S., Ahn, J. H. (2010). Agamous-like 6 is A floral promoter that negatively regulates the flc/maf clade genes and positively regulates ft in arabidopsis. Plant J. 65, 62–76. doi: 10.1111/j.1365-313X.2010.04402.x
Yu, B., Hu, Y., Hou, X. (2024). More than flowering: constans plays multifaceted roles in plant development and stress responses. J. Of Integr. Plant Biol. 00, 1–15. doi: 10.1111/jipb.13798
Yu, J. W., Rubio, V., Lee, N. Y., Bai, S., Lee, S. Y., Kim, S. S., et al. (2008). Cop1 and elf3 control circadian function and photoperiodic flowering by regulating gi stability. Mol. Cell 32, 617–630. doi: 10.1016/j.molcel.2008.09.026
Yu, S., Galvao, V. C., Zhang, Y. C., Horrer, D., Zhang, T. Q., Hao, Y. H., et al. (2012). Gibberellin Regulates the Arabidopsis Floral Transition through miR156-Targeted SQUAMOSA PROMOTER BINDING-LIKE Transcription Factors. Plant Cell 24, 3320–3332. doi: 10.1105/tpc.112.101014
Zagotta, M. T., Hicks, K. A., Jacobs, C. I., Young, J. C., Hangarter, R. P., Meeks-Wagner, D. R. (1996). The arabidopsis elf3 gene regulates vegetative photomorphogenesis and the photoperiodic induction of flowering. Plant J. 10, 691–702. doi: 10.1046/j.1365-313X.1996.10040691.x
Zhang, L., Jiménez-Gómez, J. M. (2020). Functional analysis of frigida using naturally occurring variation in arabidopsis thaliana. Plant J. 103, 154–165. doi: 10.1111/tpj.v103.1
Zhang, H., Van Nocker, S. (2002). The vernalization independence 4 gene encodes A novel regulator of flowering locus C. Plant J. 31, 663–673. doi: 10.1046/j.1365-313X.2002.01380.x
Zhang, B., Wang, L., Zeng, L., Zhang, C., Ma, H. (2015). Arabidopsis toe proteins convey A photoperiodic signal to antagonize constans and regulate flowering time. Genes Dev. 29, 975–987. doi: 10.1101/gad.251520.114
Zheng, S., Hu, H., Ren, H., Yang, Z., Qiu, Q., Qi, W., et al. (2019). The arabidopsis H3k27me3 demethylase jumonji 13 is A temperature and photoperiod dependent flowering repressor. Nat. Commun. 10, 1303. doi: 10.1038/s41467-019-09310-x
Zhou, Y., Ni, M. (2009). Shb1 plays dual roles in photoperiodic and autonomous flowering. Dev. Biol. 331, 50–57. doi: 10.1016/j.ydbio.2009.04.023
Keywords: flowering, short-day, CONSTANS, flowering locus T, gibberellin
Citation: Wang Y, Lv T, Fan T, Zhou Y and Tian C-e (2025) Research progress on delayed flowering under short-day condition in Arabidopsis thaliana. Front. Plant Sci. 16:1523788. doi: 10.3389/fpls.2025.1523788
Received: 06 November 2024; Accepted: 03 February 2025;
Published: 07 March 2025.
Edited by:
Lee Jeong Hwan, Jeonbuk National University, Republic of KoreaReviewed by:
Islam Frahat Hassan, National Research Centre, EgyptKyounghee Lee, Seoul National University, Republic of Korea
Copyright © 2025 Wang, Lv, Fan, Zhou and Tian. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Chang-en Tian, YmlvY2V0aWFuQGd6aHUuZWR1LmNu