 Inês Mansinhos
Inês Mansinhos Sandra Gonçalves
Sandra Gonçalves Anabela Romano*
Anabela Romano*- Mediterranean Institute for Agriculture, Environment and Development (MED) and CHANGE – Global Change and Sustainability Institute, Faculdade de Ciências e Tecnologia, Universidade do Algarve, Campus de Gambelas, Faro, Portugal
The interest in medicinal and aromatic plants (MAPs) has increased significantly in recent years, driven by the growing demand for natural products. MAPs are a valuable source of secondary metabolites, which renders them useful to a number of industries, including cosmetics, pharmaceuticals, and food. The Lamiaceae family includes economically important MAPs that produce valuable secondary metabolites such as essential oils (EOs) and phenolic compounds (PCs). The quantity and quality of these secondary metabolites are affected by abiotic stress factors. In a climate change scenario, the Lamiaceae is one of the most affected families, especially due to its wide distribution in the Mediterranean region. In the present study, the most common climate-related environmental stress factors, namely, drought, salinity, temperature, light, and heavy metals, were reviewed and discussed in order to assess their impact on the chemical profiles of EOs and PCs, as well as on the biological properties (antioxidant, antibacterial, antimelanogenic, pest-repellent, and UV-protective) of Lamiaceae species. It can be posited that these stresses typically act as a catalyst for the secondary metabolism of these plants, resulting in increased production of EO compounds (e.g., 1,8-cineole, linalool, camphor, borneol, and limonene) and PCs (e.g., rosmarinic, caffeic, and salvianolic acids) and subsequent enhancement of their biological activities. In view of the industrial applications of these bioactive compounds, it is of interest to explore the changes in secondary metabolism induced by environmental factors as it is possible to increase the accumulation of valuable secondary metabolites.
1 Introduction
Globally, plant growth and development are increasingly affected by climate change-related factors. This is mainly due to the fact that these stresses decrease the uptake and diffusion of CO2 and modify many biochemical reactions. Indeed, these abiotic stresses represent the primary cause of losses in agricultural productivity, with a reduction in yield of over 50% observed in the agriculture sector (Mahajan et al., 2020). In numerous regions, the most significant and detrimental factors are drought and salinity. It is estimated that over 50% of the arable land will be salinized by 2050 (Jan et al., 2018). Another crucial factor that has been stimulated by anthropogenic global warming is temperature. It is anticipated that the frequency and intensity of heat waves and extremely warm summers will increase, which will result in an exacerbation of drought events (Lehner et al., 2018). An essential physical factor involved in plant growth and development is light/radiation, which is particularly influenced by photoperiod (duration of illumination), quality (wavelength, color), and quantity (fluence rate) (Mahajan et al., 2020). In the context of climate change, ultraviolet (UV) radiation, particularly UV-B radiation, is likely to have a significant impact on plants due to its high capacity to cause biological damage (Mansinhos et al., 2024). Furthermore, climate change also affects the pathways of heavy metal pollution, including photo-enhanced toxicity, uptake, metabolism, biodegradation, adsorption, photodegradation, volatilization, and hydrolysis (Mok et al., 2023). The impact of climate change is of particular concern for endemic and rare species, which are at high risk of extinction (Kougioumoutzis et al., 2022). It is of utmost importance to gain a comprehensive understanding of the responses of plants to these constraints if we are to enhance food security and minimize economic losses (Butcher et al., 2016). The effects of climatic changes result in plants being subjected to conditions that exceed their physiological homeostasis, thereby necessitating adaptation. Plants typically respond to stressful conditions by reducing or inhibiting growth, inhibiting photosynthesis, and activating several tolerance mechanisms, including the accumulation of osmolytes, enzymatic and non-enzymatic antioxidant systems, and the expression of specific defense proteins (Szekely-Varga et al., 2020b). Secondary metabolites, such as phenolics, alkaloids, and terpenoids, play an important role in the adaptation and recovery of plants to environmental conditions, mainly due to their ability to scavenge reactive oxygen species (ROS) (Albergaria et al., 2020; Mahajan et al., 2020). It has been reported that environmental stress conditions affect both the composition and concentration of these metabolites, as well as their biological properties (e.g., antioxidant, antitumor, antimicrobial, anti-inflammatory, gastrointestinal, and cardioprotective) (Albergaria et al., 2020) and the consequent health benefits and/or industrial applicability (e.g., food, pharmaceutical, cosmetic, and agrochemical) (Nabi et al., 2021) (Figure 1).
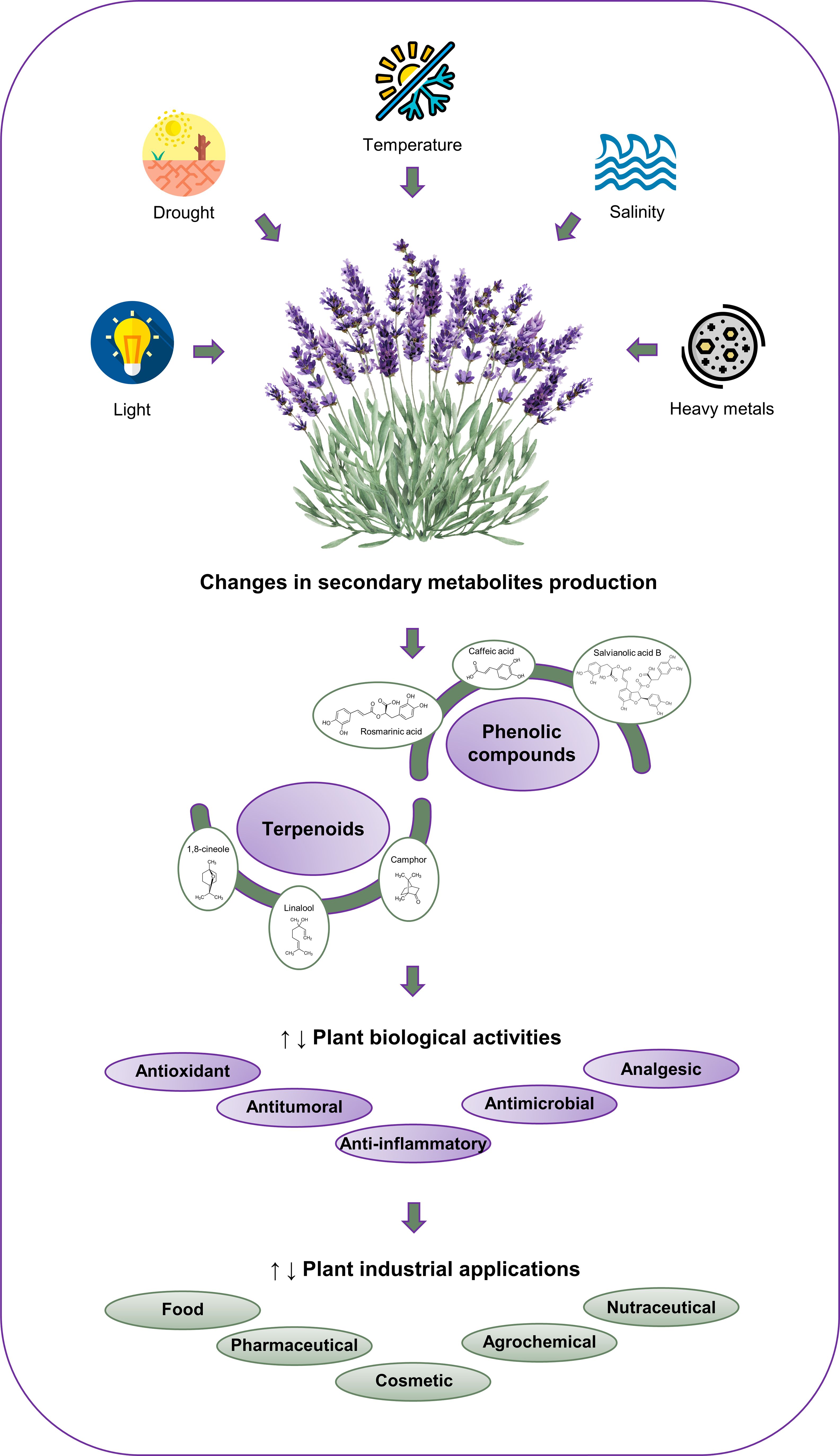
Figure 1 Effect of different abiotic stress factors (drought, salinity, temperature, light and heavy metals) on the production of the main Lamiaceae secondary metabolites and their potential industrial applications.
The demand for medicinal and aromatic plants (MAPs) that produce a broad range of secondary metabolites has increased radically in recent years. Geographically, the global herbal medicine market is segmented into the Americas, Europe, Asia-Pacific, the Middle East, and Africa. In 2021, the global market value of MAP products (prepared in the form of extracts, powders, capsules, tablets, and syrups) was approximately US$145 billion and is expected to reach more than US$356 billion by 2030 (Herbal Medicine Market, 2021). In addition, billions of people worldwide depend (partially or completely) on more than 70,000 MAPs. According to the World Health Organization (WHO), it is estimated that 88% of all countries use traditional medicine such as herbal medicines, yoga, acupuncture, indigenous therapies, and others. In terms of pharmaceutical formulations, more than 40% are based on natural products and breakthrough drugs derived from traditional medicine (e.g., aspirin and artemisinin) (WHO, 2022). Unfortunately, the natural stocks of MAPs are declining due to the increasing demand for them, which is closely linked to changes in the external climatic context (Rahman et al., 2022). The Lamiaceae family is of great significance in the realm of MAPs, as it is responsible for the production of essential oils (EOs) and phenolic compounds (PCs) with industrial applications (Mamadalieva et al., 2017). Although there are some review articles reporting the effects of environmental stress factors related to climate change on MAP secondary metabolite production, a systematic review on Lamiaceae plants is lacking. Consequently, the objective of this study is to provide a comprehensive overview of the impact of environmental stress conditions on the production of EOs and PCs in this significant family of plants which is anticipated to be one of the most affected by climate change due to its extensive distribution in the Mediterranean region.
2 Lamiaceae family: general description, economic importance, and main secondary metabolites
Lamiaceae is a very important angiosperm family consisting of aromatic, medicinal, and ornamental plants. It is the largest family in the order Lamiales, comprising 236 genera with more than 7,000 species, mostly herbs and shrubs (Assaf et al., 2022; Diab et al., 2022). Among the genera, the largest belonging to this family is Salvia (900), followed by Scutellaria (360), Stachys (300), Plectranthus (300), Hyptis (280), Teucrium (250), Vitex (250), Thymus (220), and Nepeta (200) (Tamokou et al., 2017). This family is distributed throughout the world, with a particular incidence in the Mediterranean and subtropical regions. Morphologically, the stems of these plants are usually square in cross section (with some exceptions), and the flowers are bilaterally symmetrical with five united petals and five united sepals. They are usually bisexual and verticillastrate (a flower cluster that appears to be a whorl of flowers but is actually two crowded clusters) (Raja, 2012). The plants are often aromatic in all parts (Raja, 2012), and for this, the Lamiaceae family has significant economic importance due to their EOs or use as culinary herbs/spices (Khojasteh et al., 2020). For culinary use, oregano and marjoram (Origanum), thyme (Thymus), sage (Salvia), mint (Mentha), rosemary (Rosmarinus), basil (Ocimum), and lavender (Lavandula) are the most commonly used species. In addition, several Lamiaceae genera (e.g., Salvia, Lavandula, Teucrium, Phlomis) are also cultivated as ornamental garden/houseplants (Patrignani et al., 2021) due to their aromatic properties and ease of cultivation. In fact, these plants are among the easiest plants to propagate by stem cuttings (Raja, 2012). From an industrial and economic perspective, these MAPs are of significant importance to the development of the cosmetics, agro-alimentary, pharmaceutical, and perfume sectors (Avasiloaiei et al., 2023). Mentha and Lavandula are two important genera that are widely used in cosmetics. Species such as Rosmarinus officinalis L., Origanum majorana L., and Ocimum basilicum L. are commonly used as flavorings in the food industry and Satureja montana L., Mentha x piperita L., Salvia officinalis L., and Sideritis scardica Griseb. as beverages and teas (Patrignani et al., 2021). All these species are known to be rich sources of bioactive compounds, especially PCs and terpenes.
Lamiaceae plants have a long traditional history based on their medicinal use (Patrignani et al., 2021) as anti-inflammatory, sedative, and analgesic agents. In addition, these plants have also shown great applicability as common teas, flavors, and insect repellents. Most of the species belonging to this family are aromatic and contain a mixture of bioactive compounds used in many industries (Mamadalieva et al., 2017), mainly in the form of EOs (Chakrabartty et al., 2022). In addition to EOs, these plants can be used as extracts or isolated bioactive compounds (Mamadalieva et al., 2017; Patrignani et al., 2021). Plants produce different types of bioactive compounds, which are classified into three main groups based on their origin: terpenoids, PCs, and nitrogen-containing compounds (Mahajan et al., 2020). Lamiaceae plants mainly produce terpenes (terpenoids) and PCs (Mamadalieva et al., 2017).
EOs are volatile and highly concentrated hydrophobic liquids isolated by distillation from different parts of plants, specifically flowers, leaves, fruits, stems, bark, peels, roots, seeds, or whole plants from a single botanical source (Gonçalves et al., 2020; Masyita et al., 2022). The chemical constituents of EOs are classified into terpenes, terpenoids, phenylpropanoids, and other components (Masyita et al., 2022). While terpenes are simple hydrocarbons containing isoprene units (five carbons), terpenoids are more complex terpenes with additional functional groups. Terpenoids (oxygenated hydrocarbons) contain oxygen molecules that are formed through biochemical variations by the removal or addition of methyl groups. However, both terms (terpenes and terpenoids) are often used interchangeably (Nagegowda and Gupta, 2020; Masyita et al., 2022). Plants produce two main classes of terpenoids: the primary (essential) terpenoid metabolites (such as carotenoids, sterols, and many hormones) and the secondary (non-essential) or specialized terpenoid metabolites (terpenes). The most abundant group of terpenoids—terpenes—are often specific or unique to individual plant species or groups of species and provide the plant with overall fitness to interact with the environment (Nagegowda and Gupta, 2020). Depending on the number of isoprene units, terpenes are classified into different categories, namely, hemiterpenes formed by one isoprene unit (C5), monoterpenes (C10), sesquiterpenes (C15), diterpenes (C20), triterpenes (C30), and tetraterpenes (C40). Monoterpenes are the major compounds of EOs (90%), followed by sesquiterpenes (Masyita et al., 2022). In plants, terpene biosynthesis occurs via the mevalonate (MVA) pathway (localized in the cytosol) and the 2-methylerythritol 4-phosphate (MEP) pathway (localized in the plastid) and involves different enzymatic reactions (Senji and Mandoulakani, 2018; Gonçalves et al., 2020). Monoterpenes, diterpenes, and tetraterpenes are synthesized in the plastid, whereas sesquiterpenes and triterpenes are synthesized in the cytosol (Mahajan et al., 2020). EOs and their individual compounds have a wide range of biological activities (e.g., antimicrobial, anti-inflammatory, analgesic, antioxidant), and more than 3,000 EOs have shown industrial importance, especially in perfumery and cosmetics (Senji and Mandoulakani, 2018; Gonçalves et al., 2020). The emission of these bioactive compounds depends on several environmental factors (Mahajan et al., 2020).
PCs are an important group of secondary metabolites produced in response to abiotic stress. Chemically, they are defined as compounds containing more than one hydroxyl group that is glycosylated or methylated (Mahajan et al., 2020). These compounds are divided into five major subclasses: phenolic acids, flavonoids, stilbenes, tannins, and lignans. In terms of secondary metabolism pathways, the shikimate and malonate pathways are responsible for the biosynthesis of the precursors of phenolics (Gonçalves et al., 2020). In plants, the shikimate pathway is responsible for most PC biosynthesis, whereas in bacteria and fungi, the malonate pathway is more important for their biosynthesis (Mahajan et al., 2020). As they are derived from phenylalanine, PCs are also known as phenylpropanoids (Sharma et al., 2019). Due to the presence of hydroxyl and carboxyl functional groups, phenolics have the ability to chelate heavy metal ions, such as iron, manganese, and copper, and to scavenge free radicals. In addition to their antioxidant capacity, other pharmacological properties (e.g., antitumor, antimicrobial, anti-inflammatory, gastrointestinal, and cardioprotective) have been widely attributed to different phenolic species (Albergaria et al., 2020).
3 Methodology
In light of the considerable medicinal, industrial, and economic value of Lamiaceae species, a review was conducted of the literature published over the past two decades (2005–2024) on the impact of environmental stress conditions, namely, drought, salinity, temperature, light, and heavy metals, on the production of secondary metabolites (EOs and PCs) and the biological properties of these species. A comprehensive search was conducted across a range of scientific online databases, including Web of Science, PubMed, Scopus, Science Direct, and Google Scholar. The following search terms were used: “Lamiaceae,” “environmental stress,” “abiotic stress,” “drought,” “water stress,” “salinity,” “salt stress,” “temperature,” “heat stress,” “cold stress,” “light stress,” “UV-B stress,” “heavy metals,” “plant secondary metabolites,” “bioactive compounds,” “essential oils,” “phenolic compounds,” and “biological activities.” This review has assembled a total of 180 references, comprising research and review articles, book chapters, a case study, and some clinical trial reports. The present review article presents five tables, separated into EOs and PCs, which indicate mostly the individual compounds that are affected by the abiotic stresses previously mentioned. Furthermore, a figure has been included to illustrate the impact of abiotic stress factors on the production of the main PCs and terpenoids from Lamiaceae MAPs, as well as their biological properties and potential industrial applications.
4 Effect of environmental stress factors on secondary metabolites production by Lamiaceae species
Climate change is expected to have a major impact on Lamiaceae species due to their wide distribution in the Mediterranean basin. This region is characterized by a large number of endemic plants and enormous diversity and is even considered the second largest terrestrial biodiversity hotspot in the world (Tomou et al., 2022). According to recent data, the number of days above 37°C is expected to double from 30 to 60 by 2050 in northern Africa, southern Spain, and Turkey. These rising temperatures will increase the risk of droughts, water stress in basins, wildfires, and floods and significantly alter the Mediterranean climate. Spain, Italy, Portugal, and parts of Turkey and Greece are expected to experience at least 6 months of drought each year by 2050. Basin water supplies could decline by approximately 10% by 2030 and up to 25% by 2050 (McKinsey Global Institute, 2020). These environmental changes could affect the distribution of these MAPs, causing them to move to more climatically appropriate and suitable habitats. This could lead to financial problems if economically important plants of local rural communities are lost from locally accessible areas (Kougioumoutzis et al., 2022). In addition, the significant decline in the populations of some MAPs is mainly due to anthropogenic activities (e.g., uncontrolled collection, expansion of the urban/tourist areas, deforestation, fires). Oregano, thyme, and sage are three examples of Lamiaceae plants that have a very limited distribution in some areas and that are threatened with extinction mainly due to uncontrolled collection by free gatherers (Tomou et al., 2022). Thus, given the environmental and human causes, the conservation of these species must be of public interest to protect biodiversity and its sustainable management.
The industrial applicability of Lamiaceae plants is related to their biological properties, which are known to be dependent on their secondary metabolites. However, the production of these compounds is sensitive to small variations in the surrounding environment (Assaf et al., 2022). Drought, salinity, temperature, light and heavy metals are the main stresses that plants face today (Mahajan et al., 2020). The following is an update of studies from the last two decades on the secondary metabolic responses of Lamiaceae species to the five abovementioned abiotic stresses.
4.1 Drought
Water scarcity, one of the most damaging stresses, is responsible for the productivity losses in several crops and MAPs, especially in arid and semiarid regions (Albergaria et al., 2020). This abiotic stress, which is expected to increase in duration, frequency, and intensity as a result of climate change, affects the physiological and biochemical processes of several plants (Moustakas et al., 2022). Under water limitation, roots send a chemical signal through the xylem to the leaves, stimulating a partial stomatal closure to reduce water loss (transpiration) and prevent dehydration. As a result, there is a direct decrease in the CO2 uptake (which is detrimental to photosynthesis) and, since NADPH is not used in the Calvin cycle, NADP+ is not available. These changes lead to increased levels of free radicals and ROS (Albergaria et al., 2020; Karalija et al., 2022; Moustakas et al., 2022), which negatively affect a wide range of plant processes, such as growth and development, loss of turgor leading to cell dehydration, photosynthesis, and many other metabolic parameters (Jan et al., 2018; Khare et al., 2020; Moustakas et al., 2022). Although some researchers suggest a positive effect of elevated CO2 (resulting from fossil fuels burned by humans for energy and exacerbating the greenhouse effect) on crop yield by increasing photosynthesis, this does not appear to be sufficient to counteract the negative effects of severe drought on photosynthesis and yield (Moustakas et al., 2022). Drought tolerance traits are correlated with the accumulation of several compounds. Plants accumulate various organic solutes known as osmoregulators, osmoprotectants, or osmolytes (e.g., proline, soluble sugars, glycine betaine) to help cells maintain turgor pressure in their cytosol. Furthermore, another strategy to cope with drought stress is the enzymatic [e.g., catalase (CAT), superoxide dismutase (SOD), glutathione reductase (GR), ascorbate peroxidase (APX)] and non-enzymatic antioxidant defense systems (e.g., glutathione, ascorbate, α-tocopherol, carotenoids, PC) capable of reducing ROS (Khare et al., 2020; Karalija et al., 2022; Moustakas et al., 2022).
Table 1 summarizes the effect of drought on EO and PC production in several Lamiaceae species. Salvia spp. are among the most important MAPs, and several studies have been conducted to evaluate their ability to respond to drought stress. Ramezani et al. (2020) investigated the effect of water deficit on the EO composition, namely, the five main metabolites (1,8-cineole, camphor, borneol, α- and β-thujone), in three genotypes of Salvia spp. [S. officinalis, S. reuterana (Yasuj), and S. reuterana (Urmia)] and showed that genetic differences could influence the quality and quantity of these compounds. In Salvia dolomitica Codd, drought stimulated the biosynthesis of sesquiterpene hydrocarbons, an important class of terpenoids for the pharmaceutical, food, and cosmetics industries. However, a negative effect was observed for monoterpene hydrocarbons (Caser et al., 2019). Camphor, the main oxygenated monoterpene in Salvia sinaloensis Fern (Caser et al., 2018) and Salvia fruticosa Mill (Chrysargyris et al., 2016) increased after water stress conditions. Similarly, in S. officinalis, the amount of monoterpenes increased tremendously in response to water stress, doubling after only 2 days (Radwan et al., 2017). In S. officinalis, all the EO subclasses were increased after drought exposure (Bettaieb et al., 2009).
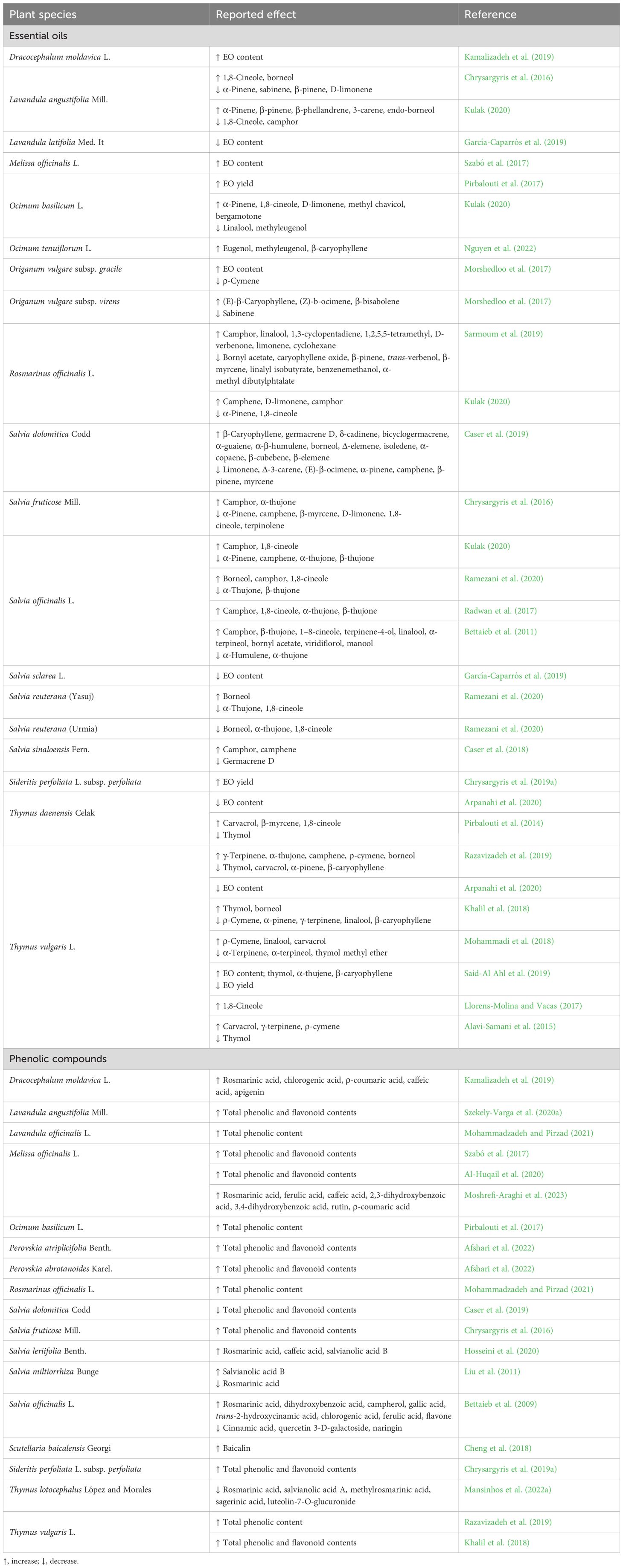
Table 1 Alteration in secondary metabolite (essential oils and phenolic compounds) production in Lamiaceae species under drought stress.
Thymus is another genus that has been well studied in terms of drought tolerance, particularly Thymus vulgaris L., a MAP native to the western Mediterranean region of Europe. In soil-grown plants of T. vulgaris, water shortage led to a decrease in γ-terpinene content and a significant increase in thymol (Khalil et al., 2018). According to the study by Arpanahi et al. (2020), when Thymus daenensis Celak and T. vulgaris were subjected to low, mild, and severe water stress, EO content decreased in both species, although this decrease was more pronounced in T. daenensis. In other studies, the EO yield and composition in plants of this species differed among plant organs, as well as with developmental periods and growth conditions (Alavi-Samani et al., 2015; Llorens-Molina and Vacas, 2017; Mohammadi et al., 2018). With regard to in vitro culture, the major compounds of EOs from T. vulgaris callus were differentially affected by drought, with γ-terpinene increasing its content and thymol and carvacrol decreasing (Razavizadeh et al., 2019). The percentage of some chemical compounds in the EOs extracted from T. daenensis plants under reduced irrigation was higher than from unstressed plants. Nevertheless, the content of its main component (thymol) was significantly reduced under stressed conditions (Pirbalouti et al., 2014).
To investigate the effect of water shortage on the EO content of five Melissa officinalis L. genotypes, plants were irrigated with 40% of the soil water capacity for 3 months, and three of the five cultivars (‘Lorelei,’ ‘Gold Leaf,’ and ‘Quedlinburger Niederliegende’) showed no significant differences between stressed plants and the control. Nevertheless, under drought conditions, the cultivar ‘Soroksar’ increased EO production by 58%, while ‘Lemona’ accumulated only 35% (Szabó et al., 2017). To induce EO biosynthesis, holy basil (Ocimum tenuiflorum L. or O. sanctum L.) at juvenile and preflowering stages was exposed to drought stress for 5 days. In both stages, the content of three major compounds (eugenol, methyl eugenol, and β-caryophyllene) was significantly enhanced compared to the control (Nguyen et al., 2022).
Six species of the Lamiaceae family commonly cultivated in the Mediterranean region, namely, M. piperita (peppermint), Salvia lavandulifolia Vahl. (Spanish sage), Salvia sclarea L. (clary sage or clear eye), Thymus mastichina L. (Spanish marjoram), Thymus capitatus (L.) Hoff. et Link., and Lavandula latifolia Med., were subjected to water stress, and the EO content was evaluated. The results indicated that the only species affected by drought were S. sclarea and L. latifolia, with a corresponding decrease in their EO contents (García-Caparrós et al., 2019). Drought affects the chemical profile of EOs from R. officinalis. Gas chromatography/mass spectrometry (GC/MS) analysis showed that 10 volatile compounds were identified in the EOs of well-irrigated plants, while 13 volatile compounds were detected in non-irrigated plants. Linalool, limonene, 1,3-cyclopentadiene, 1,2,5,5-tetramethyl, and cyclohexane compounds were characteristic of the plants exposed to drought (Sarmoum et al., 2019). Two different subspecies of Origanum vulgare L., namely, ‘gracile’ and ‘virens,’ were exposed to mild and moderate drought, and it was observed that this stress significantly improved the EO content of gracile subsp. but did not affect the EO content of virens subsp (Morshedloo et al., 2017). The content of oxygenated monoterpenes, especially the major compound 1,8-cineole, was increased in drought-stressed Lavandula angustifolia Mill (Chrysargyris et al., 2016), S. officinalis (Kulak, 2020), and O. basilicum (Kulak, 2020). The yield and content of EOs extracted from plants such as O. basilicum (Pirbalouti et al., 2017; Kulak, 2020), Dracocephalum moldavica L (Kamalizadeh et al., 2019), Sideritis perfoliata L. subsp. perfoliate (Chrysargyris et al., 2019a), and S. officinalis (Kulak, 2020) were also higher under stressed than unstressed conditions.
As with EOs, several studies have shown that water stress affects PC production in Lamiaceae plants. The content of the main active constituents of Salvia miltiorrhiza Bunge (salvianolic acid B) (Liu et al., 2011) and S. officinalis (rosmarinic acid) grown in soil (Bettaieb et al., 2011) increased under moderate and severe water deficit. On the other hand, S. dolomitica (Caser et al., 2019) and S. sinaloensis (Caser et al., 2018) were subjected to moderate and severe drought stress, and total phenolic and flavonoid contents significantly decreased in both species compared to the control. Plantlets and callus cultures of Salvia leriifolia Benth. were exposed to polyethylene glycol (PEG) to induce osmotic stress in the culture media, and the levels of rosmarinic acid, caffeic acid, and salvianolic acid B increased significantly in all cultures grown under drought conditions (Hosseini et al., 2020). Scutellaria baicalensis Georgi is a traditional MAP known for its high flavonoid content, of which baicalin is the most important. Cheng et al. (2018) showed a significant improvement in baicalin production after subjecting S. baicalensis to moderate water stress, but this production decreased under severe stress. Drought stress also favored the production of secondary metabolites in Asiatic Moldavian dragonhead (D. moldavica), especially the phenolics rosmarinic, chlorogenic, and ρ-coumaric acids under moderate stress and caffeic acid and apigenin under higher stress conditions (Kamalizadeh et al., 2019). Thymus vulgaris plants irrigated with reduced amounts of water achieved an increase in polyphenol and flavonoid contents (Khalil et al., 2018). Thymus vulgaris in vitro cultures exposed to PEG-induced osmotic stress obtained the highest amount of total phenolics at 6% PEG (Razavizadeh et al., 2019), while in Thymus lotocephalus López and Morales, the total phenolic concentration decreased under 2%, 5%, and 7% PEG (Mansinhos et al., 2022a). Sweet basil (O. basilicum) showed an increase in total phenolic (Pirbalouti et al., 2017; Al-Huqail et al., 2020) and flavonoid contents in response to water deficit and an extremely significant decrease in carotenoid content (Al-Huqail et al., 2020). More recently, drought has been shown to improve the contents of the most abundant compounds in M. longifolia L., namely, rosmarinic, ferulic, caffeic, 2,3-dihydroxybenzoic, and 3,4-dihydroxybenzoic acids, as well as rutin. Furthermore, drought has been demonstrated to trigger the production of ρ-coumaric acid which was not detected in some accessions under control conditions (Moshrefi-Araghi et al., 2023). Other Lamiaceae species such as M. officinalis (Szabó et al., 2017), Perovskia atriplicifolia Benth., Perovskia abrotanoides Karel (Afshari et al., 2022), and L. angustifolia (Szekely-Varga et al., 2020a) improved their phenolic content when subjected to water stress.
4.2 Salinity
Soil salinity is known as the accumulation of excess ions, such as chloride (Cl−), sodium (Na+), sulfate (SO42−), magnesium (Mg2+), and calcium (Ca2+) in the soil (Butcher et al., 2016). This problem has a major impact on soil properties, altering texture and reducing aeration, porosity, and water conductance (Mahajan et al., 2020). According to Butcher et al. (2016), there are two main causes of salinization—primary (natural) and secondary (anthropogenic) salinization. Primary (or dryland), which usually occurs in arid and semiarid regions (with low rainfall and high evapotranspiration), is caused by the capillary rise of saline groundwater and/or by the natural accumulation of salts from a saline parent material. Secondary (or irrigation) salinity has an anthropogenic cause and results from the use of poor-quality irrigation water. The main consequence is a significant inhibition of plant growth, development, and function, as salinity interferes with the uptake of water and nutrients, such as carbon and nitrogen (Cordovilla et al., 2014). This inhibition occurs through two main mechanisms: osmotic/drought stress and specific ionic effects. The first one is related to the change in the osmotic potential of the soil solution surrounding the root, which inhibits the root water uptake. As a result, similar to drought, plants use osmolytes to increase their water uptake. Specific ion effects are physiological effects of individual salt components that accumulate in organic tissues and cause toxicity. A common indicator of most excessive ion levels is membrane damage (Butcher et al., 2016). Another adaptive strategy of plants against stress is the accumulation of secondary metabolites. Several reports have shown that these compounds are significantly affected by salinity and tend to be more abundant in plants exposed to salt stress (Assaf et al., 2022).
Changes in the production of EOs and PCs in Lamiaceae species in response to salinity have been reported by several authors (Table 2). Origanum is an important genus of the Lamiaceae family. The response of O. vulgare plants to soil moisture regimes using fresh and saline water irrigation was evaluated, and it was found that the EO yield decreased significantly with increasing salinity. The amount of carvacrol, the dominant compound in this species (46.44%–77.96%), decreased with salt stress, in contrast to ρ-cymene and γ-terpinene, the second and the third most abundant compounds, respectively (Said-Al Ahl and Wahby, 2010). Origanum majorana (syn. Majorana hortensis Moench), known as marjoram or sweet marjoram, is a perennial herbaceous plant native to Cyprus and the eastern Mediterranean. The salinity effect is highly significant for the EO content of this species but depends on the severity of the treatment. EO yield increased significantly up to 55.5% at moderate salinity and decreased at high salinity (Jelali et al., 2011). In another study, 50–100 mM of NaCl decreased the EO yield and the content of several individual compounds of marjoram, including the most abundant (trans-sabinene hydrate), but increased the percentages of the other predominant compounds, namely, terpinene-4-ol and cis-sabinene hydrate (Baâtour et al., 2009). Two years later, the same authors showed that the influence of this stress on the EO composition of marjoram depends on several factors, such as the plant variety (i.e., Tunisian vs. Canadian) (Baâtour et al., 2011). In 19 accessions of O. basilicum, EO content was higher under salt stress than under control conditions. All accessions showed a high content of methyl chavicol, an active ingredient of various natural pesticides, pharmaceuticals, and fragrances, and the production of anethole and related structural analogs (Akbari et al., 2018). An increase in EO yield and compound content due to salinity has also been reported in O. basilicum plants (Bernstein et al., 2010; Hassanpouraghdam et al., 2011; Tarchoune et al., 2013; Elhindi et al., 2016; Farsaraei et al., 2020). In contrast, different authors showed a negative effect of salt stress on the EO amount of O. basilicum major compounds (Bahcesular et al., 2020; Masoudniaragh et al., 2021). More recently, Es-sbihi et al. (2021) investigated how salt stress affected the chemical profile of S. officinalis and found important changes in the composition of EO. NaCl decreased the EO yield from 1.2% to 0.4%, and while α-thujone, camphor, and 1,8-cineole were the major compounds of EOs under control treatment, these three components decreased in stressed plants, and thujanone emerged as a new major compound. Other reports also showed a strong effect of salt stress on the EO composition of Salvia spp. but with different results, especially depending on the salt concentration (Taarit et al., 2009; Valifard et al., 2014; Kulak et al., 2020). In S. sclarea, the application of 25 mM NaCl significantly increased the oil yield which decreased with increasing salt concentration (Taarit et al., 2011). Years later, the same authors found that the transcript of the cineole synthase 1 gene (SmCin1) accumulated in the leaves (11-fold) during the first hours of salinity treatment compared to the control.
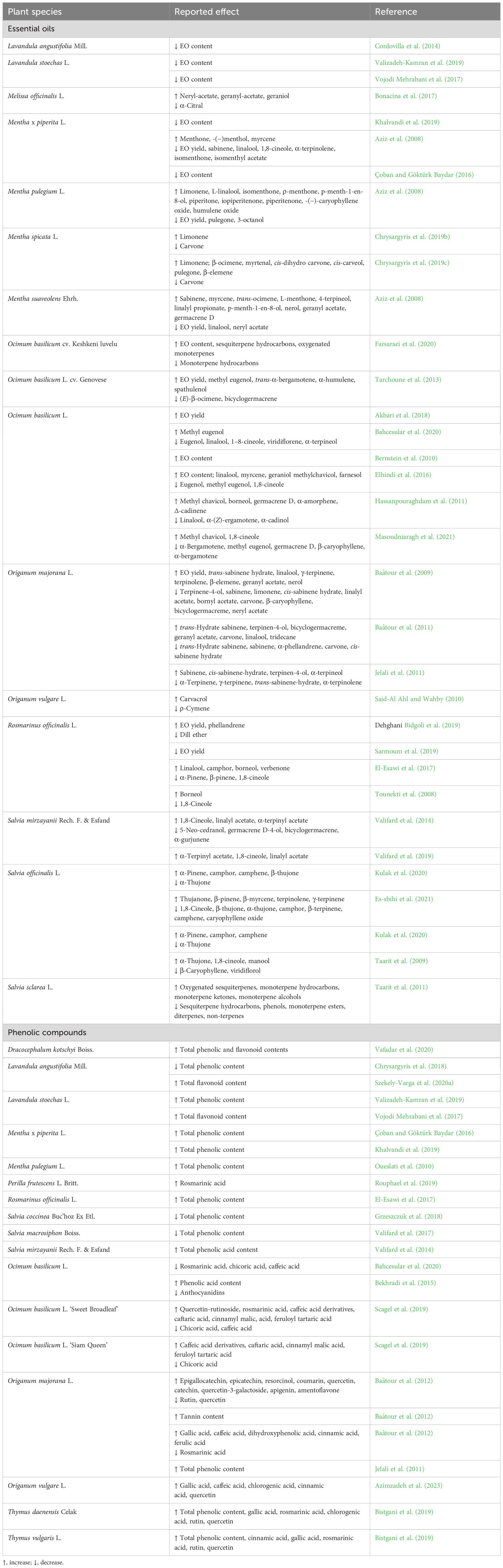
Table 2 Alteration in secondary metabolite (essential oils and phenolic compounds) production in Lamiaceae species under salinity stress.
Mentha sp. is another important Lamiaceae genera with a wide range of culinary and aromatic uses, mainly due to their richness in EOs. Aziz et al. (2008) compared the effect of salt stress on the EO chemical composition of pennyroyal (Mentha pulegium L.), peppermint (Mentha piperita), and apple mint (Mentha suaveolens Ehrh.) and observed that the EO yields of the three species decreased compared to the untreated controls. The same occurred in M. piperita in another study, where the increase in salinity led to a decrease in EO content (Çoban and Göktürk Baydar, 2016). In hydroponically grown Mentha spicata L. plants, the highest amount of NaCl tested by the authors (100 mM) was the only one that improved the EO yield (Chrysargyris et al., 2019b). In hydroponically grown M. spicata plants, 150 mM of NaCl did not affect the total EO content, but there was an increase in total monoterpene hydrocarbons and a decrease in total oxygenated monoterpenes (Chrysargyris et al., 2019c).
The use of rosemary (R. officinalis) in the Mediterranean region is an attractive solution to avoid desertification and rapid soil erosion due to its great high resistance to environmental conditions (Sarmoum et al., 2019). Bidgoli et al. (2019) found a significant relationship between the percentage of rosemary EO and the increasing salinity stress. More specifically, in another study with R. officinalis, salinity altered the major EO compounds, strongly increasing the content of camphor, borneol, linalool, and verbenone, but decreasing β-pinene, α-pinene, and 1,8-cineole (El-Esawi et al., 2017). On the other hand, a 50% decrease in 1,8-cineole content was observed with increasing NaCl concentration (Tounekti et al., 2008). In the case of Thymus maroccanus Ball., which behaved as a moderately salt-tolerant species, the EO content did not change with increasing salt concentration (Belaqziz et al., 2009). An important finding was reported by Cordovilla et al. (2014), who showed that plant–plant interactions play a crucial role in stress tolerance. These authors studied the effects of salinity and the interaction between T. vulgaris and L. angustifolia and found that both plants grown alone were more tolerant to salt stress than when grown together. When exposed to the highest stress intensity (100 mM NaCl), all lavender plants survived, whereas thyme plants died. EO production increased with salinity in T. vulgaris, whereas in lavender, it depended on the presence of thyme. Salinity has an obvious effect on Lavandula stoechas L. (Vojodi Mehrabani et al., 2017; Valizadeh-Kamran et al., 2019), strongly influencing the EO composition. According to some authors, this could be due to the disruption of cytokine transport from roots to shoots and consequently changes in the ratio of cytokines and abscisic acid (Vojodi Mehrabani et al., 2017). A decrease in EO yield from M. officinalis under salt stress was observed although the number of new compounds increased. For example, neryl acetate and geranyl acetate were detected at 100–200 mM of NaCl, suggesting that this species activates the metabolic pathways to produce monoterpenes under salinity (Bonacina et al., 2017).
In addition to EO, salt stress altered the production of PCs by increasing its concentration in the tissues although it limited the biomass production of the plants (Baâtour et al., 2012). The highest levels of PCs were recorded in several Lamiaceae species grown in a medium containing NaCl (Baâtour et al., 2012; Valifard et al., 2014; Vojodi Mehrabani et al., 2017; Valizadeh-Kamran et al., 2019). A recent study investigated the influence of salinity on the production of PCs by two subspecies of O. vulgare, namely ssp. gracile and vulgare. The results indicated an improvement in the contents of quercetin and gallic, caffeic, and chlorogenic acids in ssp. gracile and quercetin and caffeic and cinnamic acids in ssp. vulgare following salt stress (Azimzadeh et al., 2023). Baâtour et al. (2012) investigated the effect of salinity on phenolics production by O. majorana at three developmental stages (early vegetative stage, late or preflowering vegetative stage, and flowering stage). The authors observed that salt stress led to an increase in total and individual phenolic contents (e.g., gallic acid, caffeic acid, and amentoflavone) particularly at the late or preflowering vegetative stage. In the flowering stage, salinity caused a significant stimulation of other phenolic acids (caffeic, syringic, chlorogenic, cinnamic, dihydroxyphenolic, and trans-2-hydrocinnamic acids). Furthermore, the degree of salinity stress (moderate or high) significantly influenced the phenolic content of O. majorana (Jelali et al., 2011). The results of different studies showed that the positive or negative effects of salinity on basil (O. basilicum) depend on the degree of tolerance of the different genotypes. For example, when comparing green Iranian and Genovese basil, salinity stress improved the phenolic acid content only in the former (Bekhradi et al., 2015). In the case of O. basilicum ‘Sweet Broadleaf’ and ‘Siam Queen’ cultivars, salt stress increased the leaf content of the two main phenolics (quercetin-rutinoside and rosmarinic acid) only in the ‘Sweet Broadleaf’ cultivar (Scagel et al., 2019). A salt-induced improvement in PC biosynthesis has also been described in Salvia mirzayanii (Valifard et al., 2014), M. piperita (Çoban and Göktürk Baydar, 2016; Khalvandi et al., 2019), M. pulegium (Oueslati et al., 2010), L. stoechas (Valizadeh-Kamran et al., 2019), Dracocephalum kotschyi Boiss (Vafadar et al., 2020), and Perilla frutescens L. Britt (Rouphael et al., 2019). In the case of L. angustifolia, flavonoids have been shown to play an important role in salinity tolerance (Szekely-Varga et al., 2020a). On the other hand, in other Lamiaceae species, such as Salvia coccinea (Grzeszczuk et al., 2018) and Salvia macrosiphon Boiss (Valifard et al., 2017), salinity-exposed plants showed a reduced accumulation of phenolics. Cinnamic acid was the major constituent in T. daenensis and T. vulgaris, and the concentration of 60 mM NaCl increased its content by more than 30% in both species compared to the control (Bistgani et al., 2019). With regard to rosmarinic acid, the most abundant compound in several Lamiaceae species, the results found in the literature vary, with significant decreases (Baâtour et al., 2012; Bahcesular et al., 2020) or increases (Bistgani et al., 2019; Rouphael et al., 2019; Scagel et al., 2019; Vafadar et al., 2020) in its content observed under salinity stress.
4.3 Temperature
Temperature is also one of the most important abiotic factors limiting plant growth and productivity (Israel et al., 2022). Heat and cold stresses affect various physiological, biochemical, and molecular plant characteristics (Mahajan et al., 2020). High temperature can induce leaf senescence and damage to cell membranes, chlorophylls, and proteins (Mansinhos et al., 2022b). Cold stress is also known to be one of the most damaging abiotic stresses, resulting in morphological changes, such as reduced biomass, reduced leaf surface area, leaf curling, and abscission. Biochemically, low temperature stimulates the biosynthesis of ROS, leading to the oxidation of proteins and lipids (Lianopoulou and Bosabalidis, 2014). The transcriptome and metabolism of plants are also significantly affected by cold, probably due to the direct inhibition of metabolic enzymes at low temperatures (Rastogi et al., 2019). Temperature strongly affects the production of secondary metabolites. However, reports on the effect of temperature on the EO content or phenolic profile of aromatic plants remain limited. The studies available in the literature on the effect of temperature stress on the production of EOs and PCs in Lamiaceae species are presented in Table 3.

Table 3 Alteration in secondary metabolite (essential oils and phenolic compounds) production in Lamiaceae species under temperature stress.
Some studies have shown that EO composition is affected by low temperatures. For example, O. tenuiflorum, an Ocimum species that grows well in tropical, subtropical, and warm temperate regions, was exposed to cold stress (4°C) for 5 days. After this period, the eugenol content increased from 17.7 to 45.2 mg/gFW at the juvenile stage (30 days old). A significant improvement in methyleugenol and β-caryophyllene content was also observed at the juvenile and preflowering stages (Nguyen et al., 2022). On the other hand, other authors reported that eugenol and methyl eugenol concentrations in O. tenuiflorum EO decreased under cold conditions (Rastogi et al., 2019). O. basilicum plants were exposed to different temperatures of 4°C, 10°C, and 22°C (control) for 12 h, 24 h, and 48 h, and significant differences between treatments were found for almost all EO compounds. Linalool was the main compound, but there were no differences between the cold treatments and the control. On the contrary, the highest amount of the sesquiterpene germacrene D (the second most abundant compound) was found at 10°C for 12 h. In the case of geraniol and γ-cadinene, their maximum levels were observed at 4°C for 24 h (Senji and Mandoulakani, 2018). Lianopoulou and Bosabalidis (2014) compared the EO composition of Origanum dictamnus L. in winter and summer and observed that in summer, these plants produced more than twice the EO yield at higher temperatures than at lower temperatures. According to the authors, this higher EO yield in summer is correlated with the greater number of peltate glandular hairs found on summer leaves. This means that peltate hairs (and especially their head cells) are the only cells of all leaf tissues that possess the required enzymatic apparatus required for EO biosynthesis. Regarding the EO profile, there was also a variation in the main compounds, with the most abundant compound being ρ-cymene (59.2%) in winter and carvacrol (42.4%) in summer. Interestingly, the same authors repeated the same study on another Lamiaceae species, Teucrium polium L., and observed the opposite results, a significantly higher EO yield of winter leaves compared to summer leaves. Nevertheless, the authors again correlated the higher EO yield with the higher number of peltate glandular hairs in winter leaves (Lianopoulou et al., 2014). The maximum content and yield of Thymus transcaucasicus Ronn. EO were observed at 25°C compared to 15°C and 20°C (Manukyan, 2019).
There are some studies in the literature related to heat stress. Plant extracts of R. officinalis are routinely used for food preservation and cooking, cosmetics, or in herbal medicine for anti-inflammatory and antimicrobial purposes, as well as for the treatment of diabetes and cardiovascular disorders (Degner et al., 2009). Two accessions of this species were exposed to 12°C/6°C day/night, and both accessions showed an increase in carnosic and caffeic acid levels, while rosmarinic acid, carnosol, naringin, hispidulin, and cirsimaritin showed different responses (Luis et al., 2007). Elevated temperature (2.5°C–3.0°C above ambient) had a significant effect on the EO composition of S. sclarea, especially for the abundant linalyl acetate, almost doubling its concentration (Kumar et al., 2020). Similarly, the pattern of the monoterpene linalool, the most abundant component of the EO of L. angustifolia, was affected by temperature (Hassiotis et al., 2014). Linalool is widely used as an additive in 60%–80% of perfumed hygiene and cleaning products such as detergents, lotions, soaps, and shampoos and as an insecticide against fruit flies, cockroaches, and fleas (Senji and Mandoulakani, 2018). Some reports were found in the literature regarding the influence of temperature on PC biosynthesis. Recently, the effect of different temperature treatments (15°C, 20°C, 25°C, and 30°C) on the phenolic profile of in vitro cultures and micropropagated plants of two Mediterranean aromatic plants, T. lotocephalus and Lavandula viridis L’Hér, was evaluated. For both species, the production of phenolic acids (especially rosmarinic acid) increased with increasing temperature in micropropagated plants, whereas the opposite was observed in in vitro cultures (Mansinhos et al., 2022b). Another study also investigated the effect of constant air temperature (15°C, 20°C, and 25°C) on phenolic production of T. transcaucasicus plants grown in pots, and the temperature of 25°C was the most favorable for the best recovery of rosmarinic acid (Manukyan, 2019). Temperature stress led to a significant increase in total phenolics and flavonoids in basil (O. basilicum) leaves subjected to the highest temperature treatment (55°C) (Al-Huqail et al., 2020). In spearmint (M. spicata), heat significantly reduced the levels of total phenolic acids and especially rosmarinic acid (Fletcher et al., 2005). A high accumulation of rosmarinic acid was also obtained in M. officinalis under heat stress (38°C for 5 h) (Pistelli et al., 2019).
4.4 Light
The three main factors of light that strongly affect plant growth and other physiologic, biochemical, and metabolic processes are quality, quantity, and duration (Israel et al., 2022; Gonçalves et al., 2023). Despite the importance of light in several plant processes, it can also act as a stress factor. Secondary metabolites play an essential role in coping with light stress. The impact of light stress on the biosynthesis of EO and PC in Lamiaceae species is presented in Table 4. The spectral quality of light is a common parameter that is typically accessed through the use of light-emitting diodes (LEDs). For example, O. basilicum, one of the most extensively studied species within the Lamiaceae family, was cultivated under four distinct monochromatic light sources, each of which was irradiated with a specific wavelength: blue (470 nm), blue-green (500 nm), green (525 nm), and red (660 nm). After 70 days, the authors observed that the greatest concentration of EO was observed in the leaves grown under blue light and that 1,8-cineole was the most abundant compound, irrespective of light quality (Amaki et al., 2011). Similarly, P. abrotanoides and P. atriplicifolia were subjected to four distinct artificial light treatments (red, blue, red-blue, and white) generated by LEDs. The results indicated that the highest wavelengths caused a reduction and an increase in monoterpene and sesquiterpene contents, respectively (Ghaffari et al., 2019). The production of EOs from other Lamiaceae species, including M. piperita, M. spicata, and M. longifolia, was enhanced by the application of LED treatments (Sabzalian et al., 2014).
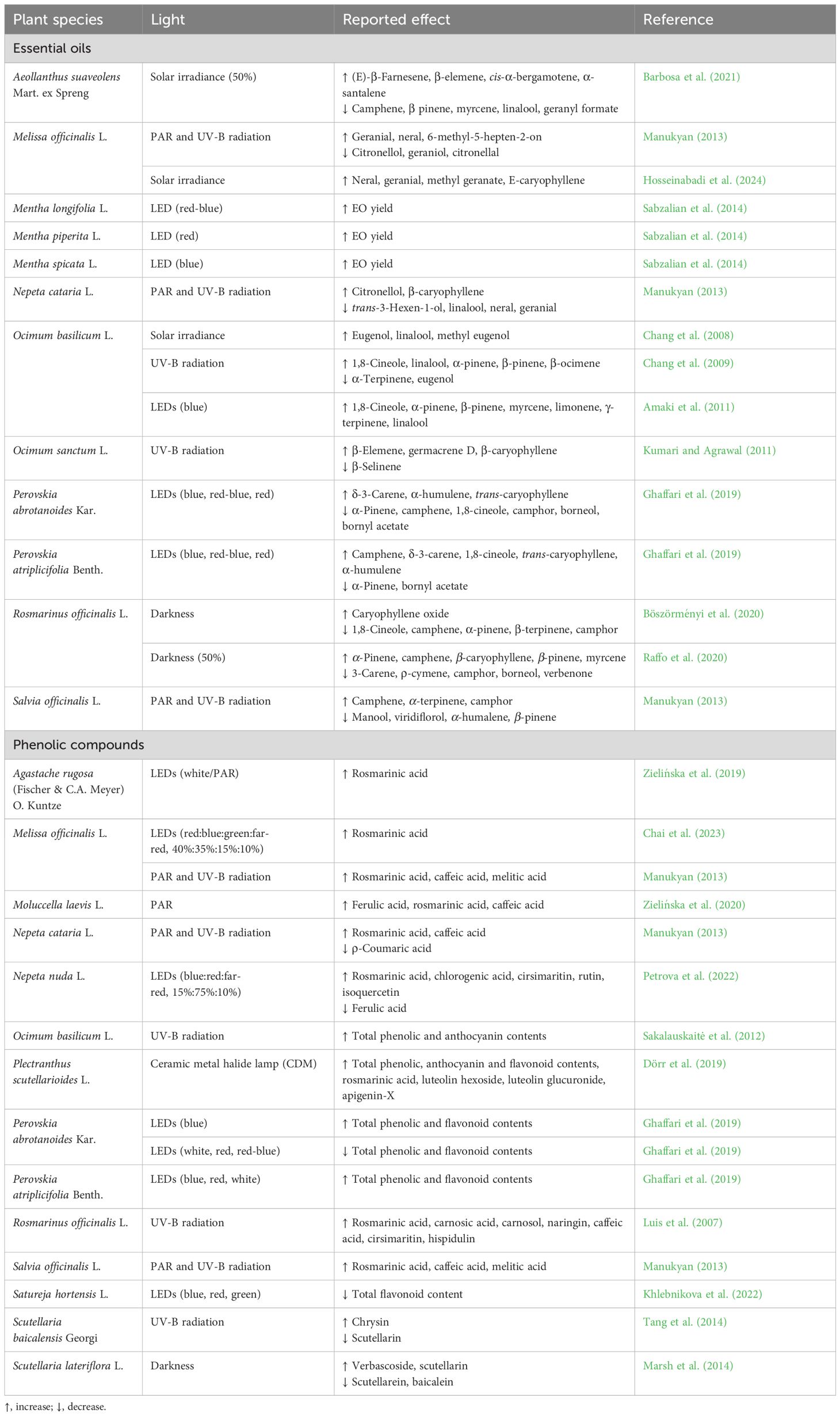
Table 4 Alteration in secondary metabolite (essential oils and phenolic compounds) production in Lamiaceae species under light stress.
The impact of UV radiation on plants is a topic that has been extensively researched. Although the Earth’s surface only receives 20% of UV-B radiation, this energy provides sufficient energy to cause several biological injuries to plants, including damage to DNA. For this reason, UV-B is the most extensively studied type of UV radiation (Mansinhos et al., 2024). The plant photoreceptors of plants respond to UV-B radiation, regulating plant growth and metabolism. This increases the plant’s capability to protect itself from the harmful consequences of UV radiation (Hosseinabadi et al., 2024). Chang et al. (2008) investigated the impact of solar irradiance and UV-B radiation (Chang et al., 2009) on O. basilicum, demonstrating a positive effect of both treatments on linalool, a key taste compound in basil. Similarly, O. sanctum was subjected to UV-B radiation. While the content of the EO major compound (eugenol) was not affected by the treatment, the EO yield increased by 42% (Kumari and Agrawal, 2011). The analysis of volatile oils in fresh leaf samples of basil demonstrated that UV-B irradiation also stimulated the synthesis of 1,8-cineole and linalool. In contrast, no effect was observed on the volatile composition of the EO. The (partial) absence of light can also serve as a stimulus for the production of bioactive compounds. Aeollanthus suaveolens, popularly known as Catinga-de-mulata, is employed by the Amazonian population for the treatment of pain. The chemical profile of this medicinal plant was studied in response to two light intensities, namely, 50% and 100% of sunlight. It was observed that the concentration of the most abundant compound (E)-β-farnesene was increased by half shade treatment (Barbosa et al., 2021). Additionally, intermediate shading (50% of sunlight) demonstrated a beneficial impact on the yield of R. officinalis EO, resulting in an increase of 29% compared to full solar irradiance (Raffo et al., 2020). Conversely, the impact of prolonged darkness on the composition of EO from this species was examined, and no discernible differences in monoterpenes and sesquiterpenes were observed between the light- and dark-grown leaves (Böszörményi et al., 2020). In a recent study, Hosseinabadi et al. (2024) examined the impact of varying harvesting times on the yield of EO from M. officinalis. Their findings indicated a positive correlation between the increase in UV-B levels during the summer months and the EO yield. The influence of photosynthetically active radiation (PAR) and UV-B radiation on the EO of sage (S. officinalis), lemon balm (M. officinalis), and lemon catmint (Nepeta cataria L.) was investigated by Manukyan (2013). The highest yield and content of EO were observed in N. cataria, with both parameters significantly enhanced by both radiations. In the case of S. officinalis, no significant differences were observed. In contrast, the radiation displayed a negative effect on the EO content of M. officinalis.
The utilization of spectrally tunable LEDs for elicitation has also demonstrated promising potential for augmenting PC production in several species, including M. officinalis (Chai et al., 2023), Plectranthus scutellarioides L (Dörr et al., 2019), Agastache rugosa (Fischer & C.A. Meyer) O. Kuntze (Zielińska et al., 2019), and Nepeta nuda L. (Petrova et al., 2022). In a recent study, Chai et al. (2023) evaluated the impact of three distinct light treatments on rosmarinic acid production. The treatments included a) three different spectrally tuned light treatments, b) white LED, and c) sunlight. The optimal result was achieved with the blue-enriched light treatment (red:blue:green:far-red, 40%:35%:15%:10%). Additionally, the leaves of P. scutellarioides grown with a ceramic metal halide lamp (CDM) and LED light exhibited the highest concentration of rosmarinic acid (Dörr et al., 2019).
In addition to the type of light, the intensity of light also influences the production of PC. In the case of N. nuda, the high light intensity of LEDs (blue:red:far-red, 15%:75%:10%) stimulated the production of PC-like compounds, including rosmarinic acid, isoquercetin, rutin, cirsimaritin, and naringenin. Conversely, a lower intensity significantly upregulated the production of chlorogenic acid (Petrova et al., 2022). With regard to ultraviolet radiation, there is substantial evidence indicating the considerable impact of UV-B radiation on the production of PC (Luis et al., 2007; Sakalauskaitė et al., 2012; Tang et al., 2014). O. basilicum was subjected to two different intensities of UV-B radiation (2 kJ m−2 day-1 and 4 kJ m−2 day-1) for 7 days. A comparison with the control revealed that the total PC levels were significantly higher in both UV treatments (Sakalauskaitė et al., 2012). The application of UV-B radiation resulted in an increase in the concentrations of rosmarinic and carnosic acids in rosemary plants, as well as naringin and carnosol (Luis et al., 2007). Similarly, the use of different UV-B light intensities was found to activate chrysin synthesis in S. baicalensis L., which was not detected in the control plants (Tang et al., 2014). The impact of UV-B radiation and PAR on PC production was also investigated in S. officinalis, M. officinalis, and N. cataria. The application of PAR treatment was found to be conducive to PC accumulation in M. officinalis, while intensive UV-B radiation was observed to stimulate the highest PC production in S. officinalis (Manukyan, 2013). Furthermore, PAR enhanced the biosynthesis of the phenolic acids rosmarinic, caffeic, and ferulic in Moluccella laevis L (Zielińska et al., 2020).
4.5 Heavy metals
The accumulation of heavy metals (HMs) in the soil, air, and water is influenced by a number of factors, including metal industries, mining, and intensive agriculture. These activities can lead to the accumulation of essential HMs, such as copper (Cu), iron (Fe), zinc (Zn), and magnesium (Mg), which are required for plant growth and development. However, they can also result in the accumulation of non-essential HMs, such as lead (Pb), nickel (Ni), arsenic (As), mercury (Hg), chromium (Cr), and cadmium (Cd), in excess of acceptable limits (Mishra and Chandra, 2022; Gonçalves et al., 2023; Das et al., 2024). Soil contamination with HMs represents a significant environmental concern, with significant implications for human health. HMs can disrupt the normal functioning of the organs, leading to a range of illnesses, including abdominal pain, skin eruptions, ulcers, and cancer (Jezler et al., 2015). Furthermore, HMs are a significant threat to plant health, impeding photosynthesis, respiration, and nutrient uptake. The literature indicates that essential HMs are harmful to plants when present in high amounts, whereas non-essential metals are particularly injurious even when present in small concentrations (Mishra and Chandra, 2022; Das et al., 2024). In order to mitigate the effects of free radicals associated with HM stress, plants typically accumulate elevated levels of secondary metabolites that function as antioxidant agents (Gonçalves et al., 2023). Table 5 presents a summary of the effects of HM stress on the production of EOs and PCs of MAPs from the Lamiaceae family. A number of studies have been conducted in the literature which report the impact of HMs on the EOs of Ocimum spp. (Rai et al., 2004; Zheljazkov et al., 2006; Prasad et al., 2011; Siddiqui et al., 2013; Fattahi et al., 2019). The composition of the EOs of O. basilicum was found to be altered by the presence of Ni, Cr, Cd, and Pb, with the type and concentration of the HMs influencing the extent of the changes (Prasad et al., 2011). Fattahi et al. (2019) reported that Cd and Pb stimulated the EO yield and induced a positive influence on EO composition, with a notable impact on the production of its most abundant compound estragole. Similarly, Cd and Pb (individually and in combination) enhanced the limonene and 1,8-cineole contents in O. basilicum EOs, yet had no impact on the content of their major compounds menthol and menthone (Zheljazkov et al., 2006). O. tenuiflorum L. was subjected to a range of Cr concentrations (10, 20, 50, and 100 mM), which resulted in the production of eugenol, its major compound, particularly at 20 mM (Rai et al., 2004). Lower doses of As induced the accumulation of the major compounds (eugenol, methyl chavicol, and linalool) of O. tenuiflorum, O. basilicum, and O. gratissimum L. EOs. It is encouraging to note that no evidence of As was found in the EOs of any species, which makes these EOs suitable for industrial applications (Siddiqui et al., 2013).
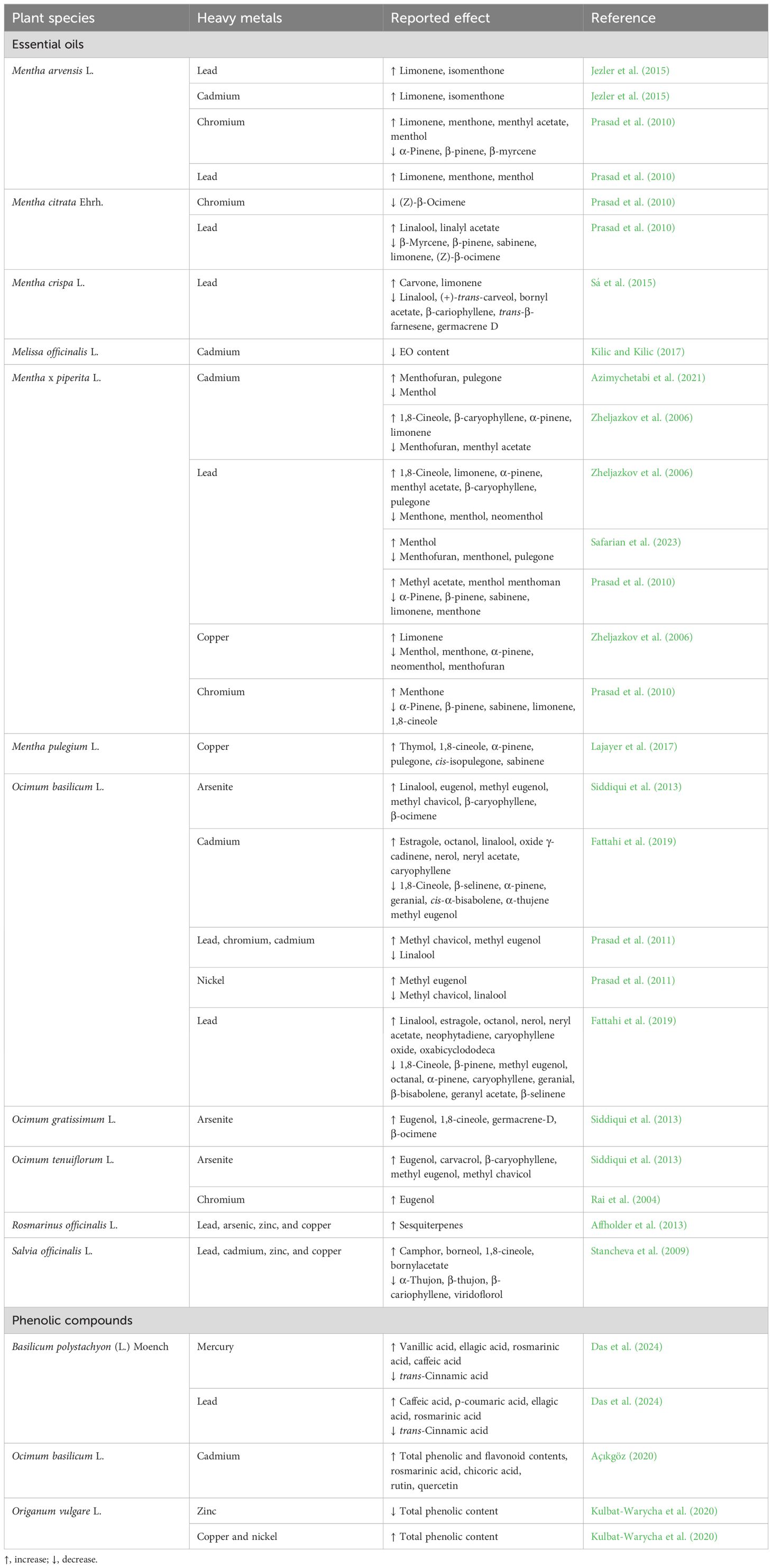
Table 5 Alteration in secondary metabolite (essential oils and phenolic compounds) production in Lamiaceae species under heavy metal stress.
Mentha is another Lamiaceae genus extensively studied in terms of EO production in response to HM stress (Zheljazkov et al., 2006; Prasad et al., 2010; Affholder et al., 2013; Jezler et al., 2015; Sá et al., 2015; Kilic and Kilic, 2017; Lajayer et al., 2017; Azimychetabi et al., 2021). The application of Cu (5 mg kg−1) and Zn (10 mg kg−1) (Lajayer et al., 2017) resulted in an increase in the yield and content of EO in M. pulegium. The presence of Cd (10–40 ppm) was found to decrease the major compound, menthol, while simultaneously increasing the menthofuran and pulegone contents in M. piperita EO (Azimychetabi et al., 2021). In a recent study, Safarian et al. (2023) demonstrated that the relative abundance of the enzymes menthol reductase and pulegone reductase were upregulated in M. piperita, in contrast to menthofuran synthase, which was downregulated during Pb stress. Furthermore, the authors demonstrated the crucial role of menthol in mitigating the effects of this environmental stress. In Mentha arvensis L., the cellular alterations induced by HM application (Pb and Cd, 8–128 mg kg−1) were insufficient to produce phytotoxicity in the plant or modify the EO yield (Jezler et al., 2015). A pot culture experiment was conducted to investigate the impact of two concentrations (30 and 60 mg kg−1) of Cr and Pb on the yield and chemical composition of EOs from three plant species: marjoram (M. arvensis), bergamot mint (M. citrata Ehrh.), and peppermint (M. piperita). The EO content of the three species decreased with both concentrations of both metals but increased EO yield in M. piperita. The effect of increasing concentrations of Pb (900, 1,800, 3,600, 7,200, and 9,000 mg kg−1) on the yield and chemical composition of EO of garden mint (Mentha crispa L.) was investigated. The EO yield was approximately 10-fold higher under extreme contamination conditions compared to that produced by the control. Moreover, although the majority of compounds were not detected under the Pb context, the content of its major component (carvone) increased from 39.3% (control) to 85.32%–95.04% after growth in Pb-contaminated soils (Sá et al., 2015). The presence of Cd in the soil negatively influenced the EO content in M. officinalis L. subsp. officinalis. The highest Cd concentration (30 mg kg−1) resulted in a 97% decrease in EO content (Kilic and Kilic, 2017). Conversely, an increased biosynthesis of sesquiterpenes in EO was observed in rosemary plants growing in contaminated regions with Pb, As, Zn, and Cu (Affholder et al., 2013). The presence of heavy metals, including Cd, Zn, Cu, and Pb, in the environment resulted in a reduction in the concentrations of α-thujon, β-thujon, viridoflorol, and β-cariophyllene in the EOs of S. officinalis. Conversely, the levels of 1,8-cineole, camphor, bornylacetate, and borneol were significantly enhanced (Stancheva et al., 2009).
A limited number of studies were identified in the literature that examined the influence of HMs on the biosynthesis of PCs in Lamiaceae species. Basilicum polystachyon (L.) Moench, a MAP found in Asia, Africa, and India, was cultivated in vitro with mercury (II) chloride and lead (II) nitrate. In cultures exposed to Hg and Pb, the esterified forms of vanillic acid and caffeic acid were respectively the major PCs, which were not detected in the control (Das et al., 2024). In a further in vitro study, using cell suspension cultures, O. basilicum was exposed to different concentrations (5–200 µM) of cadmium chloride (CdCl2) (Açıkgöz, 2020). The highest contents of rosmarinic and chicoric acids were achieved at 5 µM, while the highest accumulation of the flavonoids rutin and quercetin was observed at 10 µM. Finally, the effect of three HMs (Ni, Cu, Zn), at distinct concentrations, was studied in O. vulgare. It was observed that increasing levels of Zn and Cu caused a decrease in the PC concentration (Kulbat-Warycha et al., 2020).
5 Effect of environmental stress factors on biological properties and applications
Plants from the Lamiaceae family are known for their strong antioxidant activity, mainly due to their high PC content, which is widely used in health supplements and food preservation (Baâtour et al., 2012). In addition to antioxidant activity, plants belonging to this family have other important biological properties, such as anti-inflammatory, antitumoral, antibacterial, and antiviral, among others, which made them very interesting for researchers, food producers, and consumers (Grzeszczuk et al., 2018). Nevertheless, as concluded from the above data, the production of EOs and PCs from many Lamiaceae species is strongly influenced by environmental conditions, and the accelerating context of climate change may affect the future production and use of these species in various industries. However, most studies only focus on the effect on the percentage/amount of constituents, and concrete details on the effect on the biological properties are lacking. For example, there are few studies in the literature on the effects of drought, salinity, temperature, light, and heavy metals on the biological properties (mostly antioxidant activity) of Lamiaceae species.
The overproduction of ROS leading to oxidative stress is known to be induced by abiotic stresses. In this context, plants are provided with an antioxidant defense system, including enzymatic and non-enzymatic ones, to counteract oxidative stress. According to the literature, the radical scavenging properties of the plant extracts have been shown to be involved in abiotic stress tolerance, and most of the studies are related to osmotic stress, namely, drought and salinity. Salvia is the largest genus in the Lamiaceae family with over 900 species. Many have been used in traditional medicine for centuries as antiseptics, antispasmodics, antirheumatics, astringents, antihydrotics, tonics, antimicrobials, carminatives, digestives, anti-inflammatories, and chronic painkillers. Many reports have identified Salvia as one of the most important sources of natural antioxidants that help to maintain health and protect against various diseases, such as diabetes, stroke, cancer, atherosclerosis, and neurodegenerative diseases (Grzeszczuk et al., 2018). In response to different abiotic stresses, namely, salinity in the case of S. mirzayanii (Valifard et al., 2014) and S. macrosiphon (Valifard et al., 2017) and drought in S. fruticosa (Chrysargyris et al., 2016), the extracts of these plants exhibited increased antioxidant activity. Similarly, Chrysargyris et al. (2019a) studied the effects of drought in another Lamiaceae plant (S. perfoliata) and observed that deficit irrigation enhanced the antioxidant properties of plant extracts. Conversely, water scarcity was found to significantly reduce the antioxidant activity of S. dolomitica (Caser et al., 2019) and S. sinaloensis (Caser et al., 2018) extracts accessed by the ferric reducing antioxidant power (FRAP) method. O. basilicum is a highly valuable annual herb that is commercially cultivated in numerous countries worldwide. It has been reported that this species has several biological properties, including antimicrobial, antioxidant, anti-inflammatory, analgesic, anaesthetic, anti-ulcerogenic, anti-tuberculosis, and cardiac and immune-stimulant effects (Tamokou et al., 2017; Açıkgöz, 2021). O. basilicum plants were subjected to reduced irrigation, and a positive effect on the antioxidant capacity of their extract was observed (Pirbalouti et al., 2017). In contrast, salt stress was found to reduce the antioxidant activity of O. basilicum extracts (Bahcesular et al., 2020). In Mentha species, the impact of osmotic stress is also variable. The DPPH˙ scavenging potential of M. pulegium was found to be significantly elevated in the leaves under salinity conditions, whereas the scavenging potential of the root extracts remained unaltered (Oueslati et al., 2010). Chrysargyris et al. (2019c) observed that exposure of M. spicata plants to 150 mM of NaCl did not affect the antioxidant capacity, as assessed by the FRAP, DPPH˙, and ABTS˙ methods. Conversely, in M. piperita (Khalvandi et al., 2019), R. officinalis (El-Esawi et al., 2017; Mohammadzadeh and Pirzad, 2021), T. daenensis (Bistgani et al., 2019), T. vulgaris (Khalil et al., 2018; Bistgani et al., 2019; Razavizadeh et al., 2019; Mohammadzadeh and Pirzad, 2021), and D. kotschyi (Vafadar et al., 2020), the capacity of plant extracts to scavenge free radicals has been demonstrated to improve with increasing osmotic stress induced by salinity or drought. In several cases and as demonstrated above, osmotic stress induced by climate change factors stimulates the antioxidant activity of plant extracts, but sometimes the opposite effect is observed. Lavender is an example of a cultivated plant, either as an ornamental or for EO production and pharmaceutical purposes, and is widely used in medicines, perfumes, balms, lotions, and other cosmetics. L. angustifolia was exposed to 25, 50, and 100 mM of NaCl, and the antioxidant capacity (FRAP, DPPH˙, and ABTS˙) of its extracts significantly decreased with the increasing stress intensity (Chrysargyris et al., 2018). The results from other Lamiaceae spp. such as O. majorana (Baâtour et al., 2012), S. coccinea (Grzeszczuk et al., 2018), P. atriplicifolia, and P. abrotanoides (Afshari et al., 2022) demonstrated a negative effect on the antioxidant activity of plant extracts when exposed to salt stress or drought conditions in T. lotocephalus (Mansinhos et al., 2022a). Temperature also has a major effect on the antioxidant activity of the plant extracts. T. transcaucasicus plants were exposed to 15°C, 20°C, and 25°C, and the strongest antioxidant activity was observed at 20°C (Manukyan, 2019). Similarly, T. lotocephalus and L. viridis were grown at 15°C, 20°C, 25°C, and 30°C for 2 weeks, but the highest antioxidant activities were achieved at the extreme temperatures (15°C and 30°C) (Mansinhos et al., 2022b). On the other hand, after exposure of M. spicata to 30°C, the plant lost 21%–60% of its total antioxidant capacity after week 1 and up to 95% by week 4 (Pistelli et al., 2019).
A study by Sakalauskaitė et al. (2012) found that plant extracts from basil plants exposed to UV-B radiation exhibited a significantly higher activity in quenching DPPH radicals in comparison to non-treated plants. Furthermore, in a more recent study, the authors evaluated the impact of different spectrally tuned light treatments on the antioxidant activity of M. officinalis leaf extracts. The leaves harvested from the 40% red, 35% blue, 15% green, and 10% far-red treatments evidenced the strongest activity (Chai et al., 2023). In the context of HM pollution, increasing concentrations of Cu and Zn were found to result in a reduction in the antioxidant capacity of O. vulgare extracts, as assessed by the FRAP assay (Kulbat-Warycha et al., 2020). Conversely, the greatest enhancements in the radical scavenging capacity of O. basilicum were observed in the presence of 5 μM of CdCl2.
Although less studied, the antioxidant capacity of EO is also affected by climate change conditions. This is potentially important given the advantages of EOs over synthetic antioxidants, which often have negative health effects, to be used as alternative additives for the food industry and medicine (Tit and Bungau, 2023). Pirbalouti et al. (2014) demonstrated that the antioxidant activity of the EOs from T. daenensis plants cultivated under normal irrigation conditions was significantly higher than that observed under limited irrigation. Although Cd (10–40 ppm) significantly decreased the major compound of M. officinalis EO (menthol), it did not compromise its antioxidant properties, which increased with the increasing concentration of Cd stress (Azimychetabi et al., 2021). Similarly, there was a dose-dependent increase in the antioxidant capacity of peppermint, with the highest value obtained at 600 ppm of Pb (Safarian et al., 2023).
Few studies were found in the literature on the effect of abiotic stress on other biological properties of Lamiaceae species (beyond antioxidant activity). The antibacterial activity of R. officinalis EO against Listeria monocytogenes, Bacillus cereus, Staphylococcus aureus, Micrococcus flavus, Escherichia coli, and Pseudomonas aeruginosa was improved with increasing salt concentration. According to the authors, these activities were much higher than the antibiotics tested (El-Esawi et al., 2017). Similarly, the EO of O. majorana exposed to saline conditions (75 mM of NaCl) showed higher antibacterial activity (especially against Gram-negative bacteria) compared to plants grown under normal conditions (Olfa et al., 2016). The extracts from M. officinalis plants subjected to LEDs (red:blue:green:far-red, 40%:35%:15%:10%) were assessed against three Gram-positive bacteria (S. aureus, methicillin-resistant S. aureus, Enterococcus faecalis) and two Gram-negative bacteria (E. coli and K. pneumoniae). They demonstrated inhibitory activity against all pathogens, with the exception of E. faecalis, K. pneumoniae, and E. coli at a concentration of 5 g/mL. Nevertheless, the outcomes were more favorable than those observed in the control (Chai et al., 2023). Chrysargyris et al. (2016) investigated the toxicity of L. angustifolia and S. fruticosa EOs from plants grown under different deficit irrigation treatments to a crop pest (two-spotted spider mite, Tetranychus urticae) and found that EO from lavender under moderate water deficit caused significantly higher mortality to T. urticae than the EO from plants irrigated with adequate water. In contrast, EO from sage showed no significant differences between drought treatments. T. daenensis was irrigated at 50% field capacity, and its ability to inhibit two gram-positive (B. cereus and L. monocytogenes) and two gram-negative (Proteus vulgaris and Salmonella typhimurium) bacterial strains was evaluated. In general, EO extracted from control plants showed superior antibacterial activity against B. cereus and P. vulgaris than under drought conditions. On the other hand, thyme EO showed higher activity against S. typhimurium and L. monocytogenes in stressed than unstressed plants (Pirbalouti et al., 2014). The effect of drought on other biological properties of plant extracts was also investigated. T. lotocephalus in vitro cultures grown in culture media supplemented with PEG to simulate drought stress and the antimelanogenic and ultraviolet (UV) protective capacities of the extracts were analyzed. According to the results, all cultures exposed to PEG showed the worst biological activities (Mansinhos et al., 2022a). In addition to these biological properties, MAPs have been increasingly reported as an environmentally friendly strategy for phytoremediation. This is due to the fact that HMs are incorporated into the plant system by chelation or sequestration, neutralizing them. Furthermore, the Lamiaceae is identified as one of the most promising families of MAPs for sustainable phytoremediation. Nevertheless, it is important to note that HMs may infiltrate the food chain and cause dangerous effects. Therefore, it is recommended that non-edible MAPs need to be used in phytoremediation (Mishra and Chandra, 2022).
Although there is some ambiguous information available in the literature regarding the effect of abiotic stress on the biological properties of plants, especially because there are several factors that influence the production of bioactive compounds (e.g., duration/concentration of stress or plant genotype), in most cases, climate-related stress seems to have a positive effect. According to the wide range of drought, salinity, temperature, light, and heavy metal experiments described so far, approximately 70% of the studies showed that these factors positively affected the biological properties of Lamiaceae species, while 30% of these activities were reduced. This evidence could be very favorable to increase the industrial applications of these species. Moreover, this consideration highlights the importance of the production of secondary metabolites as one of the main adaptive mechanisms adopted by plants under stress conditions.
A review of the available data in Tables 1–5 indicates that 61% of the studies relate to the investigation of EOs. Furthermore, 1,8-cineole (also known as eucalyptol, one of the main compounds of Lamiaceae EOs) was the most frequently reported compound in Lamiaceae EOs, occurring in 56% of the studies. It was also observed that 1,8-cineole production was stimulated by abiotic stress. This monoterpene, commonly added to cosmetic or perfumery products (e.g., bath additives, mouthwashes, insect repellents), has been shown to have a wide range of pharmacological properties over the last two decades, mainly anti-inflammatory and antioxidant via NF-kB and Nrf2 signaling pathways, respectively. 1,8-Cineole can be used to treat cardiovascular, digestive, and respiratory disorders, as well as Alzheimer’s disease. In clinical trials, this compound has been administered by capsule but has high volatility and poor water solubility, which limits the absorption of 1,8-cineole by oral administration. Thus, the development of suitable formulations for further in vivo application is needed (Cai et al., 2021). Linalool is another monoterpene widely used in cosmetics, food additives, and household cleaners (An et al., 2021), which in 59% of the cases increased under abiotic stress (Tables 1–5). According to An et al. (2021), this terpene can be used as an adjuvant to antibiotics or anticancer treatments due to its low toxicity and protective effects. Linanool has the ability to induce apoptosis in cancer cells while protecting normal cells, and it has also been reported to protect the kidney, liver, and lungs due to its anti-inflammatory activity. Borneol and camphor were the EO components found in the Lamiaceae studies to be more stimulated by environmental factors, particularly drought. In 91% (for borneol) and 85% (for camphor) of the drought and salinity studies reported (Tables 1, 2), the production of this monoterpene increased significantly after exposure to these two environmental stresses. Borneol is widely used in traditional Chinese medicine for its analgesic, antipyretic, and resuscitative effects, as well as in perfumery, medicine, and the chemical industry (Ma et al., 2021). In terms of therapeutic uses, this compound helps to relieve the discomfort associated with hemorrhoids. Furthermore, in 2017, borneol completed the phase III clinical trial of capsules for the prevention and treatment of stable angina (Tasly Pharmaceuticals, Inc, 2017) and more recently, in 2022, the phase III clinical trial of tablets for the treatment of acute ischemic stroke (Peking University Third Hospital, 2022). Camphor is another terpene with multiple applications such as alternative medicine, topical medication, respiratory aerosol, flavoring additive, household cleaner, and perfumery (Chen et al., 2013; McAllister and Burnett, 2015).
With regard to PCs, rosmarinic acid is the most relevant compound in the Lamiaceae plants. According to Tables 1–5, this phenolic acid was positively affected by drought, salinity, temperature, light, and heavy metals in 81% of the reported studies. Rosmarinic acid has a wide range of applications, from food preservatives to cosmetics, and is also an interesting molecule for pharmaceuticals (Marchev et al., 2021). In terms of its wide range of health-promoting properties, this compound has demonstrated therapeutic benefits in neurodegenerative and liver disorders, cancer, diabetes, and inflammatory diseases (Noor et al., 2022). However, rosmarinic acid has a low lipophilicity, which is not good for its transmembrane penetration and oral availability. In addition, the presence of excipients in cosmetic and pharmaceutical products, as well as the acidic atmosphere of the human digestive tract or many foods, affects the stability of rosmarinic acid, resulting in low bioavailability and a significant decrease in its biological properties (Marchev et al., 2021). Nevertheless, some clinical trials with rosmarinic acid have been conducted in Japan, namely, for cognitive function (JPRN Search Portal, 2016a), dementia (JPRN Search Portal, 2016b), and Alzheimer’s disease (JPRN Search Portal, 2012). Caffeic acid was the second most abundant phenolic compound in Lamiaceae species and was also highly stimulated by abiotic stress (89% of the studies) (Tables 1–5). Caffeic acid is one of the FDA National Drugs approved and is an active ingredient in at least five products (PubChem, 2023). In 2016, this compound completed phase III clinical trials for tablets as a second-line therapy for immune thrombocytopenia (ITP) (Hou, 2016) and, in 2020, completed phase IV for caffeic acid in combination with high-dose dexamethasone in the treatment of ITP (Hou, 2020). In 2019, it entered phase III as a GASC1 inhibitor for advanced esophageal squamous cell carcinoma (The First Affiliated Hospital of Henan University of Science and Technology, 2020). According to the National Cancer Institute, caffeic acid plays a key role in the development of tumor cells. This phenolic acid is orally bioavailable and has great anti-inflammatory, antioxidant, and antineoplastic properties. When administered, this hydroxycinnamic acid acts as an antioxidant molecule, preventing oxidative stress and consequent DNA damage induced by free radicals (NCI Thesaurus, 2023).
6 Conclusions and future prospects
The Lamiaceae MAPs have been used in traditional medicine and perfumery for millennia, and their diverse array of biological properties has led to an increasing interest from various industries. Regrettably, these plants face several environmental stresses, which have a detrimental effect on the quality and quantity of their secondary metabolites. The data presented in this review indicate that the impact of climate-related stresses varies according to several factors, including plant genus, species, genetic background (cultivar), type, level, and duration of stress, as well as experimental conditions. This makes it challenging to generalize about the effect of abiotic stresses on the secondary metabolism of these plants. Nevertheless, the results of the reviewed studies demonstrated that in the majority of cases, these stresses induce the production of secondary metabolites, particularly compounds such as 1,8-cineole, borneol, camphor, linalool, limonene, rosmarinic acid, and caffeic acid. This may potentially enhance the industrial value of the plants. Furthermore, the results indicate the crucial role of these compounds in mitigating the effects of abiotic stress. In the context of climate change scenario, particularly in the Mediterranean basin, the reviewed results are of significant relevance in predicting the response of Lamiaceae plants to environmental changes and assisting the implementation of alternative agricultural and cultivation practices, such as suitable water management, which would allow for the sustainable cultivation of these plants for industrial or other applications. Moreover, several studies have demonstrated that certain Lamiaceae MAPs can accumulate heavy metals without contaminating the EOs. For these reasons, these plants may be considered viable alternatives for phytoremediation while simultaneously serving as a valuable source of bioactive compounds for numerous applications.
Author contributions
IM: Conceptualization, Writing – original draft. SG: Conceptualization, Funding acquisition, Project administration, Supervision, Writing – review & editing. AR: Conceptualization, Funding acquisition, Project administration, Supervision, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This research was funded by National Funds through FCT-Foundation for Science and Technology under the Projects UIDB/05183/2020 (https://doi.org/10.54499/UIDB/05183/2020) and LA/P/0121/2020 (https://doi.org/10.54499/LA/P/0121/2020). IM (https://doi.org/10.54499/SFRH/BD/145243/2019) and SG (CEECINST/00052/2021) acknowledge the financial support from FCT.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Açıkgöz, M. A. (2020). Establishment of cell suspension cultures of Ocimum basilicum L. and enhanced production of pharmaceutical active ingredients. Ind. Crops Prod. 148, 112278. doi: 10.1016/j.indcrop.2020.112278
Açıkgöz, M. A. (2021). Effects of sorbitol on the production of phenolic compounds and terpenoids in the cell suspension cultures of Ocimum basilicum L. Biologia 76, 395–409. doi: 10.2478/s11756-020-00581-0
Affholder, M.-C., Prudent, P., Masotti, V., Coulomb, B., Rabier, J., Nguyen-The, B., et al. (2013). Transfer of metals and metalloids from soil to shoots in wild rosemary (Rosmarinus officinalis L.) growing on a former lead smelter site: Human exposure risk. Sci. Total Environ. 454–455, 219–229. doi: 10.1016/j.scitotenv.2013.02.086
Afshari, M., Rahimmalek, M., Sabzalian, M. R., Bielecka, M., Matkowski, A., Talebi, M. (2022). Changes in physiological, phytochemical traits and gene expression of two Perovskia species in response to water deficit. Scientia Hortic. 293, 110747. doi: 10.1016/j.scienta.2021.110747
Akbari, G. A., Soltani, E., Binesh, S., Amini, F. (2018). Cold tolerance, productivity and phytochemical diversity in sweet basil (Ocimum basilicum L.) accessions. Ind. Crops Prod. 124, 677–684. doi: 10.1016/j.indcrop.2018.08.048
Alavi-Samani, S. M., Kachouei, M. A., Pirbalouti, A. G. (2015). Growth, yield, chemical composition, and antioxidant activity of essential oils from two thyme species under foliar application of jasmonic acid and water deficit conditions. Hortic. Environ. Biotechnol. 56, 411–420. doi: 10.1007/s13580-015-0117-y
Albergaria, E. T., Oliveira, A. F. M., Albuquerque, U. P. (2020). The effect of water deficit stress on the composition of phenolic compounds in medicinal plants. South Afr. J. Bot. 131, 12–17. doi: 10.1016/j.sajb.2020.02.002
Al-Huqail, A., El-Dakak, R. M., Sanad, M. N., Badr, R. H., Ibrahim, M. M., Soliman, D., et al. (2020). Effects of climate temperature and water stress on plant growth and accumulation of antioxidant compounds in sweet basil (Ocimum basilicum L.) leafy vegetable. Scientifica 2020, e3808909. doi: 10.1155/2020/3808909
Amaki, W., Yamazaki, N., Ichimura, M., Watanabe, H. (2011). Effects of light quality on the growth and essential oil content in Sweet basil. Acta Hortic. 907, 91–94. doi: 10.17660/ActaHortic.2011.907.9
An, Q., Ren, J.-N., Li, X., Fan, G., Qu, S.-S., Song, Y., et al. (2021). Recent updates on bioactive properties of linalool. Food Funct. 12, 10370–10389. doi: 10.1039/D1FO02120F
Arpanahi, A. A., Feizian, M., Mehdipourian, G., Khojasteh, D. N. (2020). Arbuscular mycorrhizal fungi inoculation improve essential oil and physiological parameters and nutritional values of Thymus daenensis Celak and Thymus vulgaris L. under normal and drought stress conditions. Eur. J. Soil Biol. 100, 103217. doi: 10.1016/j.ejsobi.2020.103217
Assaf, M., Korkmaz, A., Karaman, Ş., Kulak, M. (2022). Effect of plant growth regulators and salt stress on secondary metabolite composition in Lamiaceae species. South Afr. J. Bot. 144, 480–493. doi: 10.1016/j.sajb.2021.10.030
Avasiloaiei, D. I., Calara, M., Brezeanu, P. M., Murariu, O. C., Brezeanu, C. (2023). On the Future Perspectives of Some Medicinal Plants within Lamiaceae Botanic Family Regarding Their Comprehensive Properties and Resistance against Biotic and Abiotic Stresses. Genes 14, 955. doi: 10.3390/genes14050955
Azimychetabi, Z., Sabokdast Nodehi, M., Karami Moghadam, T., Motesharezadeh, B. (2021). Cadmium stress alters the essential oil composition and the expression of genes involved in their synthesis in peppermint (Mentha piperita L.). Ind. Crops Prod. 168, 113602. doi: 10.1016/j.indcrop.2021.113602
Azimzadeh, Z., Hassani, A., Mandoulakani, B. A., Sepehr, E. (2023). Physiological, biochemical, and antioxidant responses of oregano subspecies (Origanum vulgare L. ssp. gracile and ssp. vulgare) to NaCl stress. Acta Physiol. Plant 46, 14. doi: 10.1007/s11738–023-03636–2
Aziz, E. E., Al-Amier, H., Craker, L. E. (2008). Influence of salt stress on growth and essential oil production in peppermint, pennyroyal, and apple mint. J. Herbs Spices Med. Plants 14, 77–87. doi: 10.1080/10496470802341375
Baâtour, O., Kaddour, R., Aidi Wannes, W., Lachaâl, M., Marzouk, B. (2009). Salt effects on the growth, mineral nutrition, essential oil yield and composition of marjoram (Origanum majorana). Acta Physiol. Plant 32, 45. doi: 10.1007/s11738–009-0374–4
Baâtour, O., Kaddour, R., Mahmoudi, H., Tarchoun, I., Bettaieb, I., Nasri, N., et al. (2011). Salt effects on Origanum majorana fatty acid and essential oil composition. J. Sci. Food Agric. 91, 2613–2620. doi: 10.1002/jsfa.4495
Baâtour, O., Tarchoun, I., Nasri, N., Kaddour, R., Harrathi, J., Drawi, E., et al. (2012). Effect of growth stages on phenolics content and antioxidant activities of shoots in sweet marjoram (Origanum majorana L.) varieties under salt stress. AJB 11, 16486–16493. doi: 10.5897/AJB12.1722
Bahcesular, B., Yildirim, E. D., Karaçocuk, M., Kulak, M., Karaman, S. (2020). Seed priming with melatonin effects on growth, essential oil compounds and antioxidant activity of basil (Ocimum basilicum L.) under salinity stress. Ind. Crops Prod. 146, 112165. doi: 10.1016/j.indcrop.2020.112165
Barbosa, S. M., Abreu, N., do, C., de Oliveira, M. S., Cruz, J. N., Andrade, E.H.d. A., et al. (2021). Effects of light intensity on the anatomical structure, secretory structures, histochemistry and essential oil composition of Aeollanthus suaveolens Mart. ex Spreng. (Lamiaceae). Biochem. System. Ecol. 95, 104224. doi: 10.1016/j.bse.2021.104224
Bekhradi, F., Delshad, M., Marín, A., Luna, M. C., Garrido, Y., Kashi, A., et al. (2015). Effects of salt stress on physiological and postharvest quality characteristics of different Iranian genotypes of basil. Hortic. Environ. Biotechnol. 56, 777–785. doi: 10.1007/s13580-015-1095-9
Belaqziz, R., Romane, A., Abbad, A. (2009). Salt stress effects on germination, growth and essential oil content of an endemic thyme species in Morocco (Thymus maroccanus Ball.). J. Appl. Sci. Res. 5, 858–863.
Bernstein, N., Kravchik, M., Dudai, N. (2010). Salinity-induced changes in essential oil, pigments and salts accumulation in sweet basil (Ocimum basilicum) in relation to alterations of morphological development. Ann. Appl. Biol. 156, 167–177. doi: 10.1111/j.1744-7348.2009.00376.x
Bettaieb, I., Hamrouni-Sellami, I., Bourgou, S., Limam, F., Marzouk, B. (2011). Drought effects on polyphenol composition and antioxidant activities in aerial parts of Salvia officinalis L. Acta Physiol. Plant 33, 1103–1111. doi: 10.1007/s11738-010-0638-z
Bettaieb, I., Zakhama, N., Wannes, W. A., Kchouk, M. E., Marzouk, B. (2009). Water deficit effects on Salvia officinalis fatty acids and essential oils composition. Scientia Hortic. 120, 271–275. doi: 10.1016/j.scienta.2008.10.016
Bidgoli, D. R., Azarnezhad, N., Akhbari, M., Ghorbani, M. (2019). Salinity stress and PGPR effects on essential oil changes in Rosmarinus officinalis L. Agric. Food Secur. 8, 2. doi: 10.1186/s40066–018-0246–5
Bistgani, Z. E., Hashemi, M., DaCosta, M., Craker, L., Maggi, F., Morshedloo, M. R. (2019). Effect of salinity stress on the physiological characteristics, phenolic compounds and antioxidant activity of Thymus vulgaris L. and Thymus daenensis Celak. Ind. Crops Prod. 135, 311–320. doi: 10.1016/j.indcrop.2019.04.055
Bonacina, C., Trevizan, C. B., Stracieri, J., Santos, T., Gonçalves, J. E., Gazim, Z. C., et al. (2017). Changes in growth, oxidative metabolism and essential oil composition of lemon balm (Melissa officinalis L.) subjected to salt stress. Aust. J. Crop Sci. 11, 1665–1674. doi: 10.21475/ajcs.17.11.12.pne921
Böszörményi, A., Dobi, A., Skribanek, A., Pávai, M., Solymosi, K. (2020). The effect of light on plastid differentiation, chlorophyll biosynthesis, and essential oil composition in rosemary (Rosmarinus officinalis) leaves and cotyledons. Front. Plant Sci. 0. doi: 10.3389/fpls.2020.00196
Butcher, K., Wick, A. F., DeSutter, T., Chatterjee, A., Harmon, J. (2016). Soil salinity: A threat to global food security. Agron. J. 108, 2189–2200. doi: 10.2134/agronj2016.06.0368
Cai, Z.-M., Peng, J.-Q., Chen, Y., Tao, L., Zhang, Y.-Y., Fu, L.-Y., et al. (2021). 1,8-Cineole: a review of source, biological activities, and application. J. Asian Nat. Prod. Res. 23, 938–954. doi: 10.1080/10286020.2020.1839432
Caser, M., Chitarra, W., D’Angiolillo, F., Perrone, I., Demasi, S., Lovisolo, C., et al. (2019). Drought stress adaptation modulates plant secondary metabolite production in Salvia dolomitica Codd. Ind. Crops Prod. 129, 85–96. doi: 10.1016/j.indcrop.2018.11.068
Caser, M., D’Angiolillo, F., Chitarra, W., Lovisolo, C., Ruffoni, B., Pistelli, L., et al. (2018). Ecophysiological and phytochemical responses of Salvia sinaloensis Fern. to drought stress. Plant Growth Regul. 84, 383–394. doi: 10.1007/s10725-017-0349-1
Chai, W. Y., Goh, J. K., Kalavally, V., Rahman, S., Lim, Y. Y., Choo, W. S. (2023). Enhancing rosmarinic acid production and regulating enzyme activity in Melissa officinalis L. using spectrally tunable light-emitting diodes. Ind. Crops Prod. 204, 117332. doi: 10.1016/j.indcrop.2023.117332
Chakrabartty, I., Mohanta, Y. K., Nongbet, A., Mohanta, T. K., Mahanta, S., Das, N., et al. (2022). Exploration of lamiaceae in cardio vascular diseases and functional foods: medicine as food and food as medicine. Front. Pharmacol. 13. doi: 10.3389/fphar.2022.894814
Chang, X., Alderson, P. G., Wright, C. J. (2008). Solar irradiance level alters the growth of basil (Ocimum basilicum L.) and its content of volatile oils. Environ. Exp. Bot. 63, 216–223. doi: 10.1016/j.envexpbot.2007.10.017
Chang, X., Alderson, P. G., Wright, C. J. (2009). Enhanced UV-B radiation alters basil (Ocimum basilicum L.) growth and stimulates the synthesis of volatile oils. JHF 1, 27–31. doi: 10.5897/JHF.9000049
Chen, W., Vermaak, I., Viljoen, A. (2013). Camphor—A fumigant during the black death and a coveted fragrant wood in ancient Egypt and Babylon—A review. Molecules 18, 5434–5454. doi: 10.3390/molecules18055434
Cheng, L., Han, M., Yang, L., Li, Y., Sun, Z., Zhang, T. (2018). Changes in the physiological characteristics and baicalin biosynthesis metabolism of Scutellaria baicalensis Georgi under drought stress. Ind. Crops Prod. 122, 473–482. doi: 10.1016/j.indcrop.2018.06.030
Chrysargyris, A., Kloukina, C., Vassiliou, R., Tomou, E.-M., Skaltsa, H., Tzortzakis, N. (2019a). Cultivation strategy to improve chemical profile and anti-oxidant activity of Sideritis perfoliata L. subsp. perfoliata. Ind. Crops Prod. 140, 111694. doi: 10.1016/j.indcrop.2019.111694
Chrysargyris, A., Laoutari, S., Litskas, V. D., Stavrinides, M. C., Tzortzakis, N. (2016). Effects of water stress on lavender and sage biomass production, essential oil composition and biocidal properties against Tetranychus urticae (Koch). Scientia Hortic. 213, 96–103. doi: 10.1016/j.scienta.2016.10.024
Chrysargyris, A., Loupasaki, S., Petropoulos, S. A., Tzortzakis, N. (2019b). Salinity and cation foliar application: Implications on essential oil yield and composition of hydroponically grown spearmint plants. Scientia Hortic. 256, 108581. doi: 10.1016/j.scienta.2019.108581
Chrysargyris, A., Michailidi, E., Tzortzakis, N. (2018). Physiological and biochemical responses of lavandula angustifolia to salinity under mineral foliar application. Front. Plant Sci. 9. doi: 10.3389/fpls.2018.00489
Chrysargyris, A., Papakyriakou, E., Petropoulos, S. A., Tzortzakis, N. (2019c). The combined and single effect of salinity and copper stress on growth and quality of Mentha spicata plants. J. Hazard. Mater. 368, 584–593. doi: 10.1016/j.jhazmat.2019.01.058
Çoban, Ö., Göktürk Baydar, N. (2016). Brassinosteroid effects on some physical and biochemical properties and secondary metabolite accumulation in peppermint (Mentha piperita L.) under salt stress. Ind. Crops Prod. 86, 251–258. doi: 10.1016/j.indcrop.2016.03.049
Cordovilla, M. P., Bueno, M., Aparicio, C., Urrestarazu, M. (2014). Effects of salinity and the interaction between thymus vulgaris and lavandula angustifolia on growth, ethylene production and essential oil contents. J. Plant Nutr. 37, 875–888. doi: 10.1080/01904167.2013.873462
Das, S., Sultana, K. W., Mondal, M., Chandra, I., Ndhlala, A. R. (2024). Unveiling the dual nature of heavy metals: stressors and promoters of phenolic compound biosynthesis in Basilicum polystachyon (L.) moench in vitro. Plants 13, 98. doi: 10.3390/plants13010098
Degner, S. C., Papoutsis, A. J., Romagnolo, D. F. (2009). “Chapter 26 - health benefits of traditional culinary and medicinal mediterranean plants,” in Complementary and Alternative Therapies and the Aging Population. Ed. Watson, R. R. (Academic Press, San Diego), 541–562. doi: 10.1016/B978–0-12–374228–5.00026–3
Diab, F., Zbeeb, H., Baldini, F., Portincasa, P., Khalil, M., Vergani, L. (2022). The potential of lamiaceae herbs for mitigation of overweight, obesity, and fatty liver: studies and perspectives. Molecules 27, 5043. doi: 10.3390/molecules27155043
Dörr, O. S., Zimmermann, B. F., Kögler, S., Mibus, H. (2019). Influence of leaf temperature and blue light on the accumulation of rosmarinic acid and other phenolic compounds in Plectranthus scutellarioides (L.). Environ. Exp. Bot. 167, 103830. doi: 10.1016/j.envexpbot.2019.103830
El-Esawi, M. A., Elansary, H. O., El-Shanhorey, N. A., Abdel-Hamid, A. M. E., Ali, H. M., Elshikh, M. S. (2017). Salicylic acid-regulated antioxidant mechanisms and gene expression enhance rosemary performance under saline conditions. Front. Physiol. 8. doi: 10.3389/fphys.2017.00716
Elhindi, K. M., Al-Suhaibani, N. A., El-Din, A. F. S., Yakout, S. M., Al-Amri, S. M. (2016). Effect of foliar-applied iron and zinc on growth rate and essential oil in sweet basil (Ocimum basilicum L.) under saline conditions. Progr Nutr. 18, 288–298.
Es-sbihi, F. Z., Hazzoumi, Z., Aasfar, A., Amrani Joutei, K. (2021). Improving salinity tolerance in Salvia officinalis L. by foliar application of salicylic acid. Chem. Biol. Technol. Agric. 8, 25. doi: 10.1186/s40538–021-00221-y
Farsaraei, S., Moghaddam, M., Pirbalouti, A. G. (2020). Changes in growth and essential oil composition of sweet basil in response of salinity stress and superabsorbents application. Scientia Hortic. 271, 109465. doi: 10.1016/j.scienta.2020.109465
Fattahi, B., Arzani, K., Souri, M. K., Barzegar, M. (2019). Effects of cadmium and lead on seed germination, morphological traits, and essential oil composition of sweet basil (Ocimum basilicum L.). Ind. Crops Prod. 138, 111584. doi: 10.1016/j.indcrop.2019.111584
Fletcher, R. S., Slimmon, T., McAuley, C. Y., Kott, L. S. (2005). Heat stress reduces the accumulation of rosmarinic acid and the total antioxidant capacity in spearmint (Mentha spicata L). J. Sci. Food Agric. 85, 2429–2436. doi: 10.1002/jsfa.2270
García-Caparrós, P., Romero, M. J., Llanderal, A., Cermeño, P., Lao, M. T., Segura, M. L. (2019). Effects of drought stress on biomass, essential oil content, nutritional parameters, and costs of production in six lamiaceae species. Water 11, 573. doi: 10.3390/w11030573
Ghaffari, Z., Rahimmalek, M., Sabzalian, M. R. (2019). Variation in the primary and secondary metabolites derived from the isoprenoid pathway in the Perovskia species in response to different wavelengths generated by light emitting diodes (LEDs). Ind. Crops Prod. 140, 111592. doi: 10.1016/j.indcrop.2019.111592
Gonçalves, S., Mansinhos, I., Romano, A. (2020). “Chapter 11 - Aromatic plants: A source of compounds with antioxidant and neuroprotective effects,” in Oxidative Stress and Dietary Antioxidants in Neurological Diseases. Eds. Martin, C. R., Preedy, V. R. (London, United Kingdom: Academic Press), 155–173. doi: 10.1016/B978–0-12–817780–8.00011–6
Gonçalves, S., Mansinhos, I., Romano, A. (2023). “Influence of biotic and abiotic factors on the production and composition of essential oils,” in Essential Oils (Beverly, MA, United States: John Wiley & Sons, Ltd), 69–97. doi: 10.1002/9781119829614.ch4
Grzeszczuk, M., Salachna, P., Meller, E. (2018). Changes in photosynthetic pigments, total phenolic content, and antioxidant activity of salvia coccinea buc’hoz ex etl. Induced by exogenous salicylic acid and soil salinity. Molecules 23, 1296. doi: 10.3390/molecules23061296
Hassanpouraghdam, M., Gohari, G., Tabatabaei, S., Dadpour, M., Shirdel, M. (2011). NaCl salinity and Zn foliar application influence essential oil composition of basil (Ocimum basilicum L.). Acta agriculturae Slovenica 97, 93–98. doi: 10.14720/aas.2011.97.2
Hassiotis, C. N., Ntana, F., Lazari, D. M., Poulios, S., Vlachonasios, K. E. (2014). Environmental and developmental factors affect essential oil production and quality of Lavandula angustifolia during flowering period. Ind. Crops Prod. 62, 359–366. doi: 10.1016/j.indcrop.2014.08.048
Herbal Medicine Market, M. R. F (2021) Herbal Medicine Market. Available online at: https://www.marketresearchfuture.com/reports/herbal-medicine-market-3250 (Accessed February 24, 2023).
Hosseinabadi, S., Yavari, A., Abdollahi, F. (2024). Essential oil variation in Melissa officinalis L. cultivated under industrial field conditions: effects of different harvesting times and plant materials. J. Essent. Oil Bear. Plants. 27, 285–299. doi: 10.1080/0972060X.2024.2304581
Hosseini, N. S., Ghasimi Hagh, Z., Khoshghalb, H. (2020). Morphological, antioxidant enzyme activity and secondary metabolites accumulation in response of polyethylene glycol-induced osmotic stress in embryo-derived plantlets and callus cultures of Salvia leriifolia. Plant Cell Tiss Organ Cult 140, 143–155. doi: 10.1007/s11240-019-01718-z
Hou, M. (2016) A Multicentre Randomized Study of Caffeic Acid Tablets as a Second-line Therapy for the Treatment of Immune Thrombocytopenia (ITP) (clinicaltrials.gov). Available online at: https://clinicaltrials.gov/ct2/show/NCT02351622 (Accessed March 13, 2023).
Hou, M. (2020) A Prospective, Multicenter, Placebo Controlled Study on Caffeic Acid Tablet Combining High-dose Dexamethasone in Management of Adult Primary Immune Thrombocytopenia (ITP) (clinicaltrials.gov). Available online at: https://clinicaltrials.gov/ct2/show/NCT02556814 (Accessed March 13, 2023).
Israel, A., Langrand, J., Fontaine, J., Lounès-Hadj Sahraoui, A. (2022). Significance of arbuscular mycorrhizal fungi in mitigating abiotic environmental stress in medicinal and aromatic plants: A review. Foods 11, 2591. doi: 10.3390/foods11172591
Jan, N., Qazi, H. A., Ramzan, S., John, R. (2018). “Developing stress-tolerant plants through in vitro tissue culture: family brassicaceae,” in Biotechnologies of Crop Improvement, Volume 1: Cellular Approaches. Eds. Gosal, S. S., Wani, S. H. (Springer International Publishing, Cham), 327–372. doi: 10.1007/978–3-319–78283-6_10
Jelali, N., Dhifi, W., Chahed, T., Marzouk, B. (2011). Salinity effects on growth, essential oil yield and composition and phenolic compounds content of marjoram (origanum Majorana L.) leaves. J. Food Biochem. 35, 1443–1450. doi: 10.1111/jfbc.2011.35.issue-5
Jezler, C. N., Mangabeira, P. A. O., Almeida, A.-A. F.d., Jesus, R., Oliveira, R., Silva, D., et al. (2015). Pb and Cd on growth, leaf ultrastructure and essential oil yield mint (Mentha arvensis L.). Cienc. Rural 45, 392–398. doi: 10.1590/0103-8478cr20130966
JPRN Search Portal (2012) NIPH Clinical Trials Search - Safety and efficacy of rosmarinic acid in patients with Alzheimer’s disease: Double blind placebo-controlled study. Available online at: https://rctportal.niph.go.jp/en/detail?trial_id=UMIN000007734 (Accessed March 13, 2023).
JPRN Search Portal (2016a) NIPH Clinical Trials Search - Community-based study about the efficacy of Melissa officinalis extract which contained rosmarinic acid on cognitive function in older adults with subjective cognitive impairment and mild cognitive impairment: A double blind, placebo-controlled, parallel-design, randomized control trial. Available online at: https://rctportal.niph.go.jp/en/detail?trial_id=UMIN000021596 (Accessed March 13, 2023).
JPRN Search Portal (2016b) NIPH Clinical Trials Search - Rosmarinic Acid Project for Prevention of Dementia. Available online at: https://rctportal.niph.go.jp/en/detail?trial_id=jRCTs041180064 (Accessed March 13, 2023).
Kamalizadeh, M., Bihamta, M., Zarei, A. (2019). Drought stress and TiO2 nanoparticles affect the composition of different active compounds in the Moldavian dragonhead plant. Acta Physiol. Plant 41, 21. doi: 10.1007/s11738-019-2814-0
Karalija, E., Dahija, S., Tarkowski, P., Zeljković, S.Ć. (2022). Influence of climate-related environmental stresses on economically important essential oils of mediterranean salvia sp. Front. Plant Sci. 13. doi: 10.3389/fpls.2022.864807
Khalil, N., Fekry, M., Bishr, M., El-Zalabani, S., Salama, O. (2018). Foliar spraying of salicylic acid induced accumulation of phenolics, increased radical scavenging activity and modified the composition of the essential oil of water stressed Thymus vulgaris L. Plant Physiol. Biochem. 123, 65–74. doi: 10.1016/j.plaphy.2017.12.007
Khalvandi, M., Amerian, M., Pirdashti, H., Keramati, S., Hosseini, J. (2019). Essential oil of peppermint in symbiotic relationship with Piriformospora indica and methyl jasmonate application under saline condition. Ind. Crops Prod. 127, 195–202. doi: 10.1016/j.indcrop.2018.10.072
Khare, S., Singh, N. B., Singh, A., Hussain, I., Niharika, K., Yadav, V., et al. (2020). Plant secondary metabolites synthesis and their regulations under biotic and abiotic constraints. J. Plant Biol. 63, 203–216. doi: 10.1007/s12374-020-09245-7
Khlebnikova, D. A., Efanova, E. M., Danilova, N. A., Shcherbakova, Y. V., Rivera Sidorova, I. (2022). Flavonoid accumulation in an aseptic culture of summer savory (Satureja hortensis L.). Plants 11, 533. doi: 10.3390/plants11040533
Khojasteh, A., Mirjalili, M. H., Alcalde, M. A., Cusido, R. M., Eibl, R., Palazon, J. (2020). Powerful plant antioxidants: A new biosustainable approach to the production of rosmarinic acid. Antioxidants 9, 1273. doi: 10.3390/antiox9121273
Kilic, S., Kilic, M. (2017). Effects of cadmium-induced stress on essential oil production, morphology and physiology of lemon balm (Melissa officinalis L., Lamiaceae). Appl. Ecol. Env. Res. 15, 1653–1669. doi: 10.15666/aeer/1503_16531669
Kougioumoutzis, K., Papanikolaou, A., Kokkoris, I. P., Strid, A., Dimopoulos, P., Panitsa, M. (2022). Climate change impacts and extinction risk assessment of nepeta representatives (Lamiaceae) in Greece. Sustainability 14, 4269. doi: 10.3390/su14074269
Kulak, M. (2020). Recurrent drought stress effects on essential oil profile of Lamiaceae plants: An approach regarding stress memory. Ind. Crops Prod. 154, 112695. doi: 10.1016/j.indcrop.2020.112695
Kulak, M., Gul, F., Sekeroglu, N. (2020). Changes in growth parameter and essential oil composition of sage (Salvia officinalis L.) leaves in response to various salt stresses. Ind. Crops Prod. 145, 112078. doi: 10.1016/j.indcrop.2019.112078
Kulbat-Warycha, K., Georgiadou, E. C., Mańkowska, D., Smolińska, B., Fotopoulos, V., Leszczyńska, J. (2020). Response to stress and allergen production caused by metal ions (Ni, Cu and Zn) in oregano (Origanum vulgare L.) plants. J. Biotechnol. 324, 171–182. doi: 10.1016/j.jbiotec.2020.10.025
Kumar, R., Joshi, R., Kumari, M., Thakur, R., Kumar, D., Kumar, S. (2020). Elevated CO2 and temperature influence key proteins and metabolites associated with photosynthesis, antioxidant and carbon metabolism in Picrorhiza kurroa. J. Proteomics 219, 103755. doi: 10.1016/j.jprot.2020.103755
Kumar, R., Kaundal, M., Sharma, S., Thakur, M., Kumar, N., Kaur, T., et al. (2017). Effect of elevated [CO2] and temperature on growth, physiology and essential oil composition of Salvia sclarea L. in the western Himalayas. J. Appl. Res. Med. Aromat. Plants 6, 22–30. doi: 10.1016/j.jarmap.2017.01.001
Kumari, R., Agrawal, S. B. (2011). Comparative analysis of essential oil composition and oil containing glands in Ocimum sanctum L. (Holy basil) under ambient and supplemental level of UV-B through gas chromatography–mass spectrometry and scanning electron microscopy. Acta Physiol. Plant 33, 1093–1101. doi: 10.1007/s11738-010-0637-0
Lajayer, H. A., Savaghebi, G., Hadian, J., Hatami, M., Pezhmanmehr, M. (2017). Comparison of copper and zinc effects on growth, micro- and macronutrients status and essential oil constituents in pennyroyal (Mentha pulegium L.). Braz. J. Bot. 40, 379–388. doi: 10.1007/s40415-016-0353-0
Lehner, F., Deser, C., Sanderson, B. M. (2018). Future risk of record-breaking summer temperatures and its mitigation. Climatic Change 146, 363–375. doi: 10.1007/s10584-016-1616-2
Lianopoulou, V., Bosabalidis, A. M. (2014). Traits of seasonal dimorphism associated with adaptation to cold stress in Origanum dictamnus L. (Lamiaceae). J. Biol. Research-Thessaloniki 21, 17. doi: 10.1186/2241-5793-21-17
Lianopoulou, V., Bosabalidis, A. M., Patakas, A., Lazari, D., Panteris, E. (2014). Effects of chilling stress on leaf morphology, anatomy, ultrastructure, gas exchange, and essential oils in the seasonally dimorphic plant Teucrium polium (Lamiaceae). Acta Physiol. Plant 36, 2271–2281. doi: 10.1007/s11738-014-1605-x
Liu, H., Wang, X., Wang, D., Zou, Z., Liang, Z. (2011). Effect of drought stress on growth and accumulation of active constituents in Salvia miltiorrhiza Bunge. Ind. Crops Prod. 33, 84–88. doi: 10.1016/j.indcrop.2010.09.006
Llorens-Molina, J. A., Vacas, S. (2017). Effect of drought stress on essential oil composition of Thymus vulgaris L. (Chemotype 1, 8-cineole) from wild populations of Eastern Iberian Peninsula. J. Essential Oil Res. 29, 145–155. doi: 10.1080/10412905.2016.1211561
Luis, J. C., Martín, R., Frías, I., Valdés, F. (2007). Enhanced carnosic acid levels in two rosemary accessions exposed to cold stress conditions. J. Agric. Food Chem. 55, 8062–8066. doi: 10.1021/jf0712393
Ma, Q., Ma, R., Jin, B.-L., Cui, G.-H. (2021). [Research progress of natural borneol resources]. Zhongguo Zhong Yao Za Zhi 46, 57–61. doi: 10.19540/j.cnki.cjcmm.20200915.609
Mahajan, M., Kuiry, R., Pal, P. K. (2020). Understanding the consequence of environmental stress for accumulation of secondary metabolites in medicinal and aromatic plants. J. Appl. Res. Med. Aromat. Plants 18, 100255. doi: 10.1016/j.jarmap.2020.100255
Mamadalieva, N. Z., Akramov, D., Ovidi, E., Tiezzi, A., Nahar, L., Azimova, S. S., et al. (2017). Aromatic medicinal plants of the lamiaceae family from Uzbekistan: ethnopharmacology, essential oils composition, and biological activities. Medicines (Basel) 4, 8. doi: 10.3390/medicines4010008
Mansinhos, I., Gonçalves, S., Rodríguez-Solana, R., Duarte, H., Ordóñez-Díaz, J. L., Moreno-Rojas, J. M., et al. (2022a). Response of thymus lotocephalus in vitro cultures to drought stress and role of green extracts in cosmetics. Antioxidants 11, 1475. doi: 10.3390/antiox11081475
Mansinhos, I., Gonçalves, S., Rodríguez-Solana, R., Ordóñez-Díaz, J. L., Moreno-Rojas, J. M., Romano, A. (2022b). Impact of Temperature on Phenolic and Osmolyte Contents in In Vitro Cultures and Micropropagated Plants of Two Mediterranean Plant Species, Lavandula viridis and Thymus lotocephalus. Plants 11, 3516. doi: 10.3390/plants11243516
Mansinhos, I., Gonçalves, S., Romano, A. (2024)Impact of abiotic stresses on in vitro production of secondary metabolites. In: In Vitro Propagation and Secondary Metabolite Production from Medicinal Plants: Current Trends (Part 1) (Bentham Science Publishers). Available online at: https://www.eurekaselect.com/chapter/21655 (Accessed May 9, 2024).
Manukyan, A. (2013). Effects of PAR and UV-B radiation on herbal yield, bioactive compounds and their antioxidant capacity of some medicinal plants under controlled environmental conditions. Photochem. Photobiol. 89, 406–414. doi: 10.1111/j.1751-1097.2012.01242.x
Manukyan, A. (2019). Secondary metabolites and their antioxidant capacity of Caucasian endemic thyme (Thymus transcaucasicus Ronn.) as affected by environmental stress. J. Appl. Res. Med. Aromat. Plants 13, 100209. doi: 10.1016/j.jarmap.2019.100209
Marchev, A. S., Vasileva, L. V., Amirova, K. M., Savova, M. S., Koycheva, I. K., Balcheva-Sivenova, Z. P., et al. (2021). Rosmarinic acid - From bench to valuable applications in food industry. Trends Food Sci. Technol. 117, 182–193. doi: 10.1016/j.tifs.2021.03.015
Marsh, Z., Yang, T., Nopo-Olazabal, L., Wu, S., Ingle, T., Joshee, N., et al. (2014). Effect of light, methyl jasmonate and cyclodextrin on production of phenolic compounds in hairy root cultures of Scutellaria lateriflora. Phytochemistry 107, 50–60. doi: 10.1016/j.phytochem.2014.08.020
Masoudniaragh, A., Oraei, M., Gohari, G., Akbari, A., Faramarzi, A. (2021). Using halloysite nanotubes as carrier for proline to alleviate salt stress effects in sweet basil (Ocimum basilicum L.). Scientia Hortic. 285, 110202. doi: 10.1016/j.scienta.2021.110202
Masyita, A., Mustika Sari, R., Dwi Astuti, A., Yasir, B., Rahma Rumata, N., Emran, T. B., et al. (2022). Terpenes and terpenoids as main bioactive compounds of essential oils, their roles in human health and potential application as natural food preservatives. Food Chem. X 13, 100217. doi: 10.1016/j.fochx.2022.100217
McAllister, R. K., Burnett, C. J. (2015). “Topical analgesic medications,” in Pain Management and Palliative Care. Ed. Sackheim, K. A. (Springer New York, New York, NY), 99–102. doi: 10.1007/978–1-4939–2462-2_15
McKinsey Global Institute (2020) A Mediterranean basin without a Mediterranean climate? Available online at: https://www.mckinsey.com/~/media/mckinsey/business%20functions/sustainability/our%20insights/
a%20mediterranean%20basin%20without%20a%20mediterranean%20climate/a-mediterranean-basin-without-a-mediterranean-climate.pdf.
Mishra, B., Chandra, M. (2022). Evaluation of phytoremediation potential of aromatic plants: A systematic review. J. Appl. Res. Med. Aromat. Plants 31, 100405. doi: 10.1016/j.jarmap.2022.100405
Mohammadi, H., Ghorbanpour, M., Brestic, M. (2018). Exogenous putrescine changes redox regulations and essential oil constituents in field-grown Thymus vulgaris L. under well-watered and drought stress conditions. Ind. Crops Prod. 122, 119–132. doi: 10.1016/j.indcrop.2018.05.064
Mohammadzadeh, S., Pirzad, A. (2021). Biochemical responses of mycorrhizal-inoculated Lamiaceae (Lavender, Rosemary and Thyme) plants to drought: a field study. Soil Sci. Plant Nutr. 67, 41–49. doi: 10.1080/00380768.2020.1851144
Mok, W. J., Ghaffar, M. A., Noor, M. I. M., Lananan, F., Azra, M. N. (2023). Understanding climate change and heavy metals in coastal areas: A macroanalysis assessment. Water 15, 891. doi: 10.3390/w15050891
Morshedloo, M. R., Craker, L. E., Salami, A., Nazeri, V., Sang, H., Maggi, F. (2017). Effect of prolonged water stress on essential oil content, compositions and gene expression patterns of mono- and sesquiterpene synthesis in two oregano (Origanum vulgare L.) subspecies. Plant Physiol. Biochem. 111, 119–128. doi: 10.1016/j.plaphy.2016.11.023
Moshrefi-Araghi, A., Nemati, H., Azizi, M., Hadian, J., Moshtaghi, N., Shoor, M. (2023). Screening of distributed horsemint (Mentha longifolia L.) accessions for agricultural drought tolerance based on biological responses. Acta Physiol. Plant 45, 130. doi: 10.1007/s11738–023-03605–9
Moustakas, M., Sperdouli, I., Moustaka, J. (2022). Early drought stress warning in plants: color pictures of photosystem II photochemistry. Climate 10, 179. doi: 10.3390/cli10110179
Nabi, N., Singh, S., Saffeullah, P. (2021). Responses of in vitro cell cultures to elicitation: regulatory role of jasmonic acid and methyl jasmonate: a review. In Vitro Cell.Dev.Biol.-Plant. 57, 341–355. doi: 10.1007/s11627-020-10140-6
Nagegowda, D. A., Gupta, P. (2020). Advances in biosynthesis, regulation, and metabolic engineering of plant specialized terpenoids. Plant Sci. 294, 110457. doi: 10.1016/j.plantsci.2020.110457
NCI Thesaurus (2023) NCI Thesaurus - Caffeic acid. Available online at: https://ncithesaurus.nci.nih.gov/ncitbrowser/ConceptReport.jsp?dictionary=NCI_Thesaurus&ns=ncit&code=C68540 (Accessed March 14, 2023).
Nguyen, C. T. T., Nguyen, N. H., Choi, W. S., Lee, J. H., Cheong, J.-J. (2022). Biosynthesis of essential oil compounds in Ocimum tenuiflorum is induced by abiotic stresses. Plant Biosyst. - Int. J. Dealing all Aspects Plant Biol. 156, 353–357. doi: 10.1080/11263504.2020.1857870
Noor, S., Mohammad, T., Rub, M. A., Raza, A., Azum, N., Yadav, D. K., et al. (2022). Biomedical features and therapeutic potential of rosmarinic acid. Arch. Pharm. Res. 45, 205–228. doi: 10.1007/s12272-022-01378-2
Olfa, B., Mariem, A., Salah, A. M., Mouhiba, B. A. (2016). Chemical content, antibacterial and antioxidant properties of essential oil extract from Tunisian Origanum majorana L. cultivated under saline condition. Pak J. Pharm. Sci. 29, 1951–1958.
Oueslati, S., Karray-Bouraoui, N., Attia, H., Rabhi, M., Ksouri, R., Lachaal, M. (2010). Physiological and antioxidant responses of Mentha pulegium (Pennyroyal) to salt stress. Acta Physiologiae Plantarum 32, 289–296. doi: 10.1007/s11738-009-0406-0
Patrignani, F., Prasad, S., Novakovic, M., Marin, P. D., Bukvicki, D. (2021). Lamiaceae in the treatment of cardiovascular diseases. Front. Biosci. (Landmark Ed) 26, 612–643. doi: 10.2741/4909
Peking University Third Hospital (2022) Phase III Clinical Trial of Y-2 Sublingual Tablet in the Treatment of Acute Ischemic Stroke - a Multicenter, Randomized, Double-blind, Parallel, Placebo-controlled Phase III Clinical Trial (clinicaltrials.gov). Available online at: https://clinicaltrials.gov/ct2/show/NCT04950920 (Accessed March 9, 2023).
Petrova, D., Gašić, U., Yocheva, L., Hinkov, A., Yordanova, Z., Chaneva, G., et al. (2022). Catmint (Nepeta nuda L.) phylogenetics and metabolic responses in variable growth conditions. Front. Plant Sci. 13. doi: 10.3389/fpls.2022.866777
Pirbalouti, A. G., Malekpoor, F., Salimi, A., Golparvar, A., Hamedi, B. (2017). Effects of foliar of the application chitosan and reduced irrigation on essential oil yield, total phenol content and antioxidant activity of extracts from green and purple basil. Acta Scientiarum Polonorum - Hortorum Cultus 16, 177–186. doi: 10.24326/asphc
Pirbalouti, A. G., Samani, M. R., Hashemi, M., Zeinali, H. (2014). Salicylic acid affects growth, essential oil and chemical compositions of thyme (Thymus daenensis Celak.) under reduced irrigation. Plant Growth Regul. 72, 289–301. doi: 10.1007/s10725-013-9860-1
Pistelli, L., Tonelli, M., Pellegrini, E., Cotrozzi, L., Pucciariello, C., Trivellini, A., et al. (2019). Accumulation of rosmarinic acid and behaviour of ROS processing systems in Melissa officinalis L. under heat stress. Ind. Crops Prod. 138, 111469. doi: 10.1016/j.indcrop.2019.111469
Prasad, A., Kumar, S., Khaliq, A., Pandey, A. (2011). Heavy metals and arbuscular mycorrhizal (AM) fungi can alter the yield and chemical composition of volatile oil of sweet basil (Ocimum basilicum L.). Biol. Fertil Soils 47, 853–861. doi: 10.1007/s00374-011-0590-0
Prasad, A., Singh, A. K., Chand, S., Chanotiya, C. S., Patra, D. D. (2010). Effect of chromium and lead on yield, chemical composition of essential oil, and accumulation of heavy metals of mint species. Commun. Soil Sci. Plant Anal. 41, 2170–2186. doi: 10.1080/00103624.2010.504798
PubChem (2023) Caffeic Acid. Available online at: https://pubchem.ncbi.nlm.nih.gov/compound/689043 (Accessed March 14, 2023).
Radwan, A., Kleinwächter, M., Selmar, D. (2017). Impact of drought stress on specialised metabolism: Biosynthesis and the expression of monoterpene synthases in sage (Salvia officinalis). Phytochemistry 141, 20–26. doi: 10.1016/j.phytochem.2017.05.005
Raffo, A., Mozzanini, E., Ferrari Nicoli, S., Lupotto, E., Cervelli, C. (2020). Effect of light intensity and water availability on plant growth, essential oil production and composition in Rosmarinus officinalis L. Eur. Food Res. Technol. 246, 167–177. doi: 10.1007/s00217-019-03396-9
Rahman, M., Roy, B., Chowdhury, G. M., Hasan, A., Md., S. R. (2022). Medicinal plant sources and traditional healthcare practices of forest-dependent communities in and around Chunati Wildlife Sanctuary in southeastern Bangladesh. Environ. Sustainability 5, 207–241. doi: 10.1007/s42398-022-00230-z
Rai, V., Vajpayee, P., Singh, S. N., Mehrotra, S. (2004). Effect of chromium accumulation on photosynthetic pigments, oxidative stress defense system, nitrate reduction, proline level and eugenol content of Ocimum tenuiflorum L. Plant Sci. 167, 1159–1169. doi: 10.1016/j.plantsci.2004.06.016
Raja, R. R. (2012). Medicinally potential plants of labiatae (Lamiaceae) family: an overview. Res. J. Med. Plants 6, 203–213. doi: 10.3923/rjmp.2012.203.213
Ramezani, S., Abbasi, A., Sobhanverdi, S., Shojaeiyan, A., Ahmadi, N. (2020). The effects of water deficit on the expression of monoterpene synthases and essential oils composition in Salvia ecotypes. Physiol. Mol. Biol. Plants 26, 2199–2207. doi: 10.1007/s12298-020-00892-1
Rastogi, S., Shah, S., Kumar, R., Vashisth, D., Akhtar, M. Q., Kumar, A., et al. (2019). Ocimum metabolomics in response to abiotic stresses: Cold, flood, drought and salinity. PloS One 14, e0210903. doi: 10.1371/journal.pone.0210903
Razavizadeh, R., Farahzadianpoor, F., Adabavazeh, F., Komatsu, S. (2019). Physiological and morphological analyses of Thymus vulgaris L. in vitro cultures under polyethylene glycol (PEG)-induced osmotic stress. In Vitro Cell.Dev.Biol.-Plant 55, 342–357. doi: 10.1007/s11627-019-09979-1
Rouphael, Y., Kyriacou, M. C., Carillo, P., Pizzolongo, F., Romano, R., Sifola, M. I. (2019). Chemical eustress elicits tailored responses and enhances the functional quality of novel food perilla frutescens. Molecules 24, 185. doi: 10.3390/molecules24010185
Sá, R. A., Sá, R. A., Alberton, O., Gazim, Z. C., Laverde, A., Jr., Caetano, J., et al. (2015). Phytoaccumulation and effect of lead on yield and chemical composition of Mentha crispa essential oil. Desalination Water Treat 53, 3007–3017. doi: 10.1080/19443994.2013.874716
Sabzalian, M. R., Heydarizadeh, P., Zahedi, M., Boroomand, A., Agharokh, M., Sahba, M. R., et al. (2014). High performance of vegetables, flowers, and medicinal plants in a red-blue LED incubator for indoor plant production. Agron. Sustain. Dev. 34, 879–886. doi: 10.1007/s13593-014-0209-6
Safarian, M., Nodehi, M. S., Bihamta, M. R., Sobhanverdi, S. (2023). Mentha piperita L. demonstrates a potential tolerance at molecular and biochemical level to Pb stress: New perspectives. South Afr. J. Bot. 159, 635–641. doi: 10.1016/j.sajb.2023.06.035
Said-Al Ahl, H. A. H., Sabra, A. S., Alataway, A., Astatkie, T., Mahmoud, A. A., Bloem, E. (2019). Biomass production and essential oil composition of Thymus vulgaris in response to water stress and harvest time. J. Essential Oil Res. 31, 63–68. doi: 10.1080/10412905.2018.1518794
Said-Al Ahl, H., Wahby, M. S. (2010). Effect of water stress and potassium humate on the productivity of organo plant using saline and fresh water irrigation. Ozean J. Appl. Sci. 3, 125–141.
Sakalauskaitė, J., Viskelis, P., Duchovskis, P., Dambrauskienė, E., Sakalauskienė, S., Samuoliene, G., et al. (2012). Supplementary UV-B irradiation effects on basil (Ocimum basilicum L.) growth and phytochemical properties. J. Food Agric. Environ. 10, 342–346.
Sarmoum, R., Haid, S., Biche, M., Djazouli, Z., Zebib, B., Merah, O. (2019). Effect of salinity and water stress on the essential oil components of rosemary (Rosmarinus officinalis L.). Agronomy 9, 214. doi: 10.3390/agronomy9050214
Scagel, C. F., Lee, J., Mitchell, J. N. (2019). Salinity from NaCl changes the nutrient and polyphenolic composition of basil leaves. Ind. Crops Prod. 127, 119–128. doi: 10.1016/j.indcrop.2018.10.048
Senji, B. M., Mandoulakani, B. A. (2018). The impact of cold stress on genes expression pattern of mono- and sesquiterpene biosynthesis and their contents in Ocimum basilicum L. Phytochemistry 156, 250–256. doi: 10.1016/j.phytochem.2018.09.006
Sharma, A., Shahzad, B., Rehman, A., Bhardwaj, R., Landi, M., Zheng, B. (2019). Response of phenylpropanoid pathway and the role of polyphenols in plants under abiotic stress. Molecules 24, 2452. doi: 10.3390/molecules24132452
Siddiqui, F., Krishna, S. K., Tandon, P. K., Srivastava, S. (2013). Arsenic accumulation in Ocimum spp. and its effect on growth and oil constituents. Acta Physiol. Plant 35, 1071–1079. doi: 10.1007/s11738-012-1145-1
Stancheva, I., Geneva, M., Hristozkova, M., Boychinova, M., Markovska, Y. (2009). Essential oil variation of salvia officinalis (L.), grown on heavy metals polluted soil. Biotechnol. Biotechnol. Equip. 23, 373–376. doi: 10.1080/13102818.2009.10818442
Szabó, K., Radácsi, P., Rajhárt, P., Ladányi, M., Németh, É. (2017). Stress-induced changes of growth, yield and bioactive compounds in lemon balm cultivars. Plant Physiol. Biochem. 119, 170–177. doi: 10.1016/j.plaphy.2017.07.019
Szekely-Varga, Z., González-Orenga, S., Cantor, M., Boscaiu, M., Vicente, O. (2020a). Antioxidant responses to drought and salinity in Lavandula angustifolia Mill. Not Bot. Horti Agrobo 48, 1980–1992. doi: 10.15835/nbha48412150
Szekely-Varga, Z., González-Orenga, S., Cantor, M., Jucan, D., Boscaiu, M., Vicente, O. (2020b). Effects of drought and salinity on two commercial varieties of lavandula angustifolia mill. Plants 9, 637. doi: 10.3390/plants9050637
Taarit, B. M., Msaada, K., Hosni, K., Hammami, M., Kchouk, M. E., Marzouk, B. (2009). Plant growth, essential oil yield and composition of sage (Salvia officinalis L.) fruits cultivated under salt stress conditions. Ind. Crops Prod. 30, 333–337. doi: 10.1016/j.indcrop.2009.06.001
Taarit, B. M., Msaada, K., Hosni, K., Marzouk, B. (2011). Physiological changes and essential oil composition of clary sage (Salvia sclarea L.) rosette leaves as affected by salinity. Acta Physiol. Plant 33, 153–162. doi: 10.1007/s11738-010-0532-8
Tamokou, J. D. D., Mbaveng, A. T., Kuete, V. (2017). “Chapter 8 - antimicrobial activities of african medicinal spices and vegetables,” in Medicinal Spices and Vegetables from Africa. Ed. Kuete, V. (Cambridge, MA, United States: Academic Press), 207–237. doi: 10.1016/B978–0-12–809286–6.00008-X
Tang, W.-T., Fang, M.-F., Liu, X., Yue, M. (2014). Simultaneous quantitative and qualitative analysis of flavonoids from ultraviolet-B radiation in leaves and roots of scutellaria baicalensis georgi using LC-UV-ESI-Q/TOF/MS. J. Anal. Methods Chem. 2014, e643879. doi: 10.1155/2014/643879
Tarchoune, I., Baâtour, O., Harrathi, J., Cioni, P. L., Lachaâl, M., Flamini, G., et al. (2013). Essential oil and volatile emissions of basil (Ocimum basilicum) leaves exposed to NaCl or Na2SO4 salinity. J. Plant Nutr. Soil Sci. 176, 748–755. doi: 10.1002/jpln.201200278
Tasly Pharmaceuticals, Inc (2017) Phase III Confirmatory Trial to Confirm the Anti-anginal Effect of Dantonic® (T89) in Patients With Chronic Stable Angina (clinicaltrials.gov). Available online at: https://clinicaltrials.gov/ct2/show/NCT01659580 (Accessed March 9, 2023).
The First Affiliated Hospital of Henan University of Science and Technology (2020) GASC1 Inhibitor Caffeic Acid for Advanced Squamous Esophageal Cell Cancer (ESCC): a Multicenter, Phase II Trial (GiCAEC) (clinicaltrials.gov). Available online at: https://clinicaltrials.gov/ct2/show/NCT04648917 (Accessed March 13, 2023).
Tit, D. M., Bungau, S. G. (2023). Antioxidant activity of essential oils. Antioxidants 12, 383. doi: 10.3390/antiox12020383
Tomou, E.-M., Skaltsa, H., Economou, G., Trichopoulou, A. (2022). Sustainable diets & medicinal aromatic plants in Greece: Perspectives towards climate change. Food Chem. 374, 131767. doi: 10.1016/j.foodchem.2021.131767
Tounekti, T., Vadel, A. M., Bedoui, A., Khemira, H. (2008). NaCl stress affects growth and essential oil composition in rosemary (Rosmarinus officinalis L.). J. Hortic. Sci. Biotechnol. 83, 267–273. doi: 10.1080/14620316.2008.11512379
Vafadar, F., Amooaghaie, R., Ehsanzadeh, P., Ghanadian, M., Talebi, M., Ghanati, F. (2020). Melatonin and calcium modulate the production of rosmarinic acid, luteolin, and apigenin in Dracocephalum kotschyi under salinity stress. Phytochemistry 177, 112422. doi: 10.1016/j.phytochem.2020.112422
Valifard, M., Mohsenzadeh, S., Kholdebarin, B. (2017). Salinity effects on phenolic content and antioxidant activity of salvia macrosiphon. Iran J. Sci. Technol. Trans. Sci. 41, 295–300. doi: 10.1007/s40995-016-0022-y
Valifard, M., Mohsenzadeh, S., Kholdebarin, B., Rowshan, V. (2014). Effects of salt stress on volatile compounds, total phenolic content and antioxidant activities of Salvia mirzayanii. South Afr. J. Bot. 93, 92–97. doi: 10.1016/j.sajb.2014.04.002
Valifard, M., Mohsenzadeh, S., Kholdebarin, B., Rowshan, V., Niazi, A., Moghadam, A. (2019). Effect of salt stress on terpenoid biosynthesis in Salvia mirzayanii: from gene to metabolite. J. Hortic. Sci. Biotechnol. 94, 389–399. doi: 10.1080/14620316.2018.1505443
Valizadeh-Kamran, R., Vojodi Mehrabani, L., Pessarakli, M. (2019). Effects of foliar application of methanol on some physiological characteristics of Lavandula stoechas L. under NaCl salinity conditions. J. Plant Nutr. 42, 261–268. doi: 10.1080/01904167.2018.1554677
Vojodi Mehrabani, L., Kamran, R. V., Hassanpouraghdam, M. B., Pessarakli, M. (2017). Zinc sulfate foliar application effects on some physiological characteristics and phenolic and essential oil contents of lavandula stoechas L. Under sodium chloride (NaCl) salinity conditions. Commun. Soil Sci. Plant Anal. 48, 1860–1867. doi: 10.1080/00103624.2017.1406105
WHO (2022) WHO Global Centre for Traditional Medicine. Available online at: https://www.who.int/initiatives/who-global-centre-for-traditional-medicine (Accessed February 24, 2023).
Zheljazkov, V. D., Craker, L. E., Xing, B. (2006). Effects of Cd, Pb, and Cu on growth and essential oil contents in dill, peppermint, and basil. Environ. Exp. Bot. 58, 9–16. doi: 10.1016/j.envexpbot.2005.06.008
Zielińska, S., Dryś, A., Piątczak, E., Kolniak-Ostek, J., Podgórska, M., Oszmiański, J., et al. (2019). Effect of LED illumination and amino acid supplementation on phenolic compounds profile in Agastache rugosa in vitro cultures. Phytochem. Lett. 31, 12–19. doi: 10.1016/j.phytol.2019.02.029
Zielińska, S., Piątczak, E., Kozłowska, W., Bohater, A., Jezierska-Domaradzka, A., Kolniak-Ostek, J., et al. (2020). LED illumination and plant growth regulators’ effects on growth and phenolic acids accumulation in Moluccella laevis L. in vitro cultures. Acta Physiol. Plant 42, 72. doi: 10.1007/s11738–020-03060-w
Keywords: bioactivity, climate change, environmental stress, essential oils, phenolic compounds
Citation: Mansinhos I, Gonçalves S and Romano A (2024) How climate change-related abiotic factors affect the production of industrial valuable compounds in Lamiaceae plant species: a review. Front. Plant Sci. 15:1370810. doi: 10.3389/fpls.2024.1370810
Received: 15 January 2024; Accepted: 06 June 2024;
Published: 10 July 2024.
Edited by:
Khalid Rehman Hakeem, King Abdulaziz University, Saudi ArabiaReviewed by:
Saeid Hazrati, Azarbaijan Shahid Madani University, IranMozaniel Oliveira, Emílio Goeldi Paraense Museum, Brazil
Copyright © 2024 Mansinhos, Gonçalves and Romano. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sandra Gonçalves, c21nb25jYWx2ZXNAdWFsZy5wdA==; Anabela Romano, YXJvbWFub0B1YWxnLnB0