 Wenjie Cao
Wenjie Cao Yuqiang Li
Yuqiang Li Yun Chen1,3
Yun Chen1,3 Xuyang Wang
Xuyang Wang
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Plant Sci., 21 December 2023
Sec. Plant Nutrition
Volume 14 - 2023 | https://doi.org/10.3389/fpls.2023.1326244
Introduction: Vegetation restoration is an effective measure to improve the ecosystem service of degraded sandy land ecosystem. However, it is unclear how vegetation restoration on severely desertified land affect soil organic carbon (SOC) sequestration and nutrients balance. Therefore, this study was designed to clarify the response of SOC, total nitrogen (TN), total phosphorus (TP), and the resulting stoichiometric ratios (C:N:P) to afforestation and grazing exclusion, and to quantify their dynamics over time.
Methods: We conducted vegetation community investigation and soil sampling in natural sparse-forest grassland (the climax community stage), afforestation (Pinus sylvestris var. mongolica (40-year, 48-year), Caragana microphylla (20-year, 40-year)), and grazing exclusion (20-year, 40-year) in China’s Horqin Sandy Land. Soil C:N:P stoichiometry and its driving factors under different restoration measures were then studied.
Results: Afforestation and grazing exclusion significantly (p < 0.05) increased SOC, TN, and TP concentrations. Vegetation restoration significantly increased C:N, C:P, and N:P ratios, indicating that nutrient limitations may occur in the later stages of restoration. The C:N, C:P, and N:P ratios after a 40-year grazing exclusion were closest to those of natural sparse-forest grassland. The N:P under grazing exclusion increased from 3.1 to 4.1 with increasing restoration age (from 20 to 40 years), which was close to the national mean values (4.2). Moreover, afforestation may lead to water deficit in the surface soil. Vegetation restoration is the main factor leading to changes in soil C:N:P stoichiometry, and indirectly affects soil C:N:P stoichiometry by altering soil structure and chemical properties.
Conclusion: In terms of ecological stoichiometry, grazing exclusion was more conducive to restore SOC and nutrient balance than afforestation on severely desertified land. Due to the poor soil nutrients, attentions should be paid to the soil nutrients and water conditions in the later stages of vegetation restoration. Those findings can provide valuable information for the restoration of degraded sandy land in semi-arid areas.
Dryland covers 45% of earth’s surface and about 40% of the world’s population lives here (Li et al., 2023a). Soil degradation caused by desertification had a negative impact on soil health (e.g., loss of soil carbon (C) and nutrients, destruction of soil structure, and reduction of land productivity) (Geng et al., 2024). If degraded land continues to be exploited without any protective measures, it is highly likely to cause irreversible damage to the land productivity (Winowiecki et al., 2018). A series of ecological restoration programs around the world have been adopted (e.g., ‘Grain for Green’ project in China; the United Nations Decade on Ecosystem Restoration; the Great Green Wall in Africa) to protect vulnerable ecosystems (Han et al., 2023). However, the effect of restoration measures developed to combat desertification remains controversial.
Ecological stoichiometry studies the balance among energy and chemical elements, especially C, nitrogen (N) and phosphorus (P) in biological systems, to clarify ecological interactions and the underlying processes (Sterner and Elser, 2002). Soil C sequestration is closely related to nutrient cycling, and the fixation of C into soil organic matter (SOM) requires sufficient N and P (Chen et al., 2022a). This coupling relationship not only changes the distribution of photosynthetic products, but also regulates soil microbial metabolism and vegetation community composition (Zhang et al., 2018a; Jia et al., 2022). Stoichiometric ratios (mainly C:N:P) are crucial indicators of element cycling and functioning in ecosystems. Applying the ecological stoichiometric framework to study the biogeochemical cycling during restoration will strengthen our ability to predict the potential of soil C sequestration and manage soil fertility efficiently through additional interventions (e.g., fertilization).
Vegetation restoration is an effective measure to improve the ecosystem service of degraded ecosystem (Nadal-Romero et al., 2023). The biogeochemical cycles (mainly C, N, and P) may change during vegetation restoration, affecting plant growth, microbial community structure, and ecosystem succession processes (Deng et al., 2019; Zhang et al., 2019a). Du and Gao (2021) found that soil C and nutrients gradually increased under short-term (< 9 years) grazing exclusion in degraded alpine grassland. This may be because grazing exclusion increases soil C storage by increasing plant biomass and higher litter C:N (Du and Gao, 2021). There were some similar conclusions that soil C and N increased but P did not change significantly after the natural restoration of agricultural abandonment in the karst area of Southwest China (Lu et al., 2022). Therefore, the C:P and N:P increased, while C:N did not change significantly. The canopy density, solar radiation, soil conditions (e.g., soil moisture, pH, and soil temperature) may change during afforestation periods. This may affect soil C and nutrients by affecting the microbial activity related to organic matter decomposition (Deng et al., 2019; Zhang et al., 2019b). For example, when lands converted from farmland to grassland or plantation, soil moisture content decreased more than 35% (Yang et al., 2012). Tian et al. (2018) found that soil moisture was the primary driver explained variations of soil stoichiometry, as high soil moisture increased C and N by increasing plant biomass. Dijkstra et al. (2012) found that soil moisture is important in controlling P supply from inorganic sources. Nadal-Romero et al. (2023) found that C:N significantly increased after afforestation on abandoned farmland in the Mediterranean due to the litter input and the low decomposition rates. However, previous research mainly focused on the vegetation restoration succession after farmland abandonment and the natural restoration of degraded grasslands. We still know little about how restoration measures on severely desertified land affect soil stoichiometry.
Before 1782, the Horqin Sandy Land, located in northern China’s agro-pastoral ecotone, originally developed native vegetation dominated by grass species along with sparsely scattered woody species (Cao et al., 2023). However, due to the large-scale agricultural development and overgrazing, vegetation degradation has occurred. In addition, this region is dry and windy in the spring (Li et al., 2004). The coupling of human activities with regional climatic characteristics has led to serious aeolian desertification. Grazing exclusion and afforestation have been extensively implemented to combat desertification and restore degraded ecosystems in the Horqin Sandy Land. Effects of afforestation and grazing exclusion on C and N storage in this region has been well described (Li et al., 2012; Li et al., 2013; Li et al., 2017). However, it remained unclear that how different restoration measures affect soil stoichiometry and their dynamics over time in the Horqin Sandy Land. These knowledge gaps limit our ability to timely optimize restoration measures.
In this context, we designed the present study based on the theory of ecological stoichiometry to analyze the responses of soil organic carbon (SOC), total nitrogen (TN), and total phosphorus (TP) and the stoichiometric ratios (C:N:P) in the Horqin Sandy Land to ecological restoration. We examined Pinus sylvestris var. mongolica plantation, Caragana microphylla plantation, and grazing exclusion in severely degraded sandy grassland. We hypothesized that (1) soil C, N, and P would increase after afforestation and grazing exclusion on active dunes due to the improvement of environmental conditions; and (2) nutrient limitations may occur as restoration age increases. Our goals were: (i) to clarify the responses of soil C:N:P stoichiometry to afforestation and grazing exclusion over time; and (ii) to examine the relationships among vegetation factors, soil physicochemical properties, and soil C:N:P stoichiometry to identify the driving factors responsible for variations of soil stoichiometry during restoration.
The study area is in the core area of the Horqin Sandy Land, which is a Chinese national ecological function zone for preventing and mitigating aeolian desertification (MEE and CAS, 2015). The study area (with a temperate semi-arid continental climate) is located east of China’s Inner Mongolia (42.71°N to 44.83°N and 118.86°E to 123.70°E) (Figure 1). The mean annual temperature ranges from 3 to 7°C. The mean annual precipitation ranges from 350 to 500 mm, of which 70% falls from July to September. The annual average potential evaporation ranges from 1500 to 2500 mm. The annual average wind speed ranges from 3.4 to 4.4 m·s−1 (Duan et al., 2014). The annual sandstorm frequency is 10 to 15 days and the storms mainly occur in the spring.
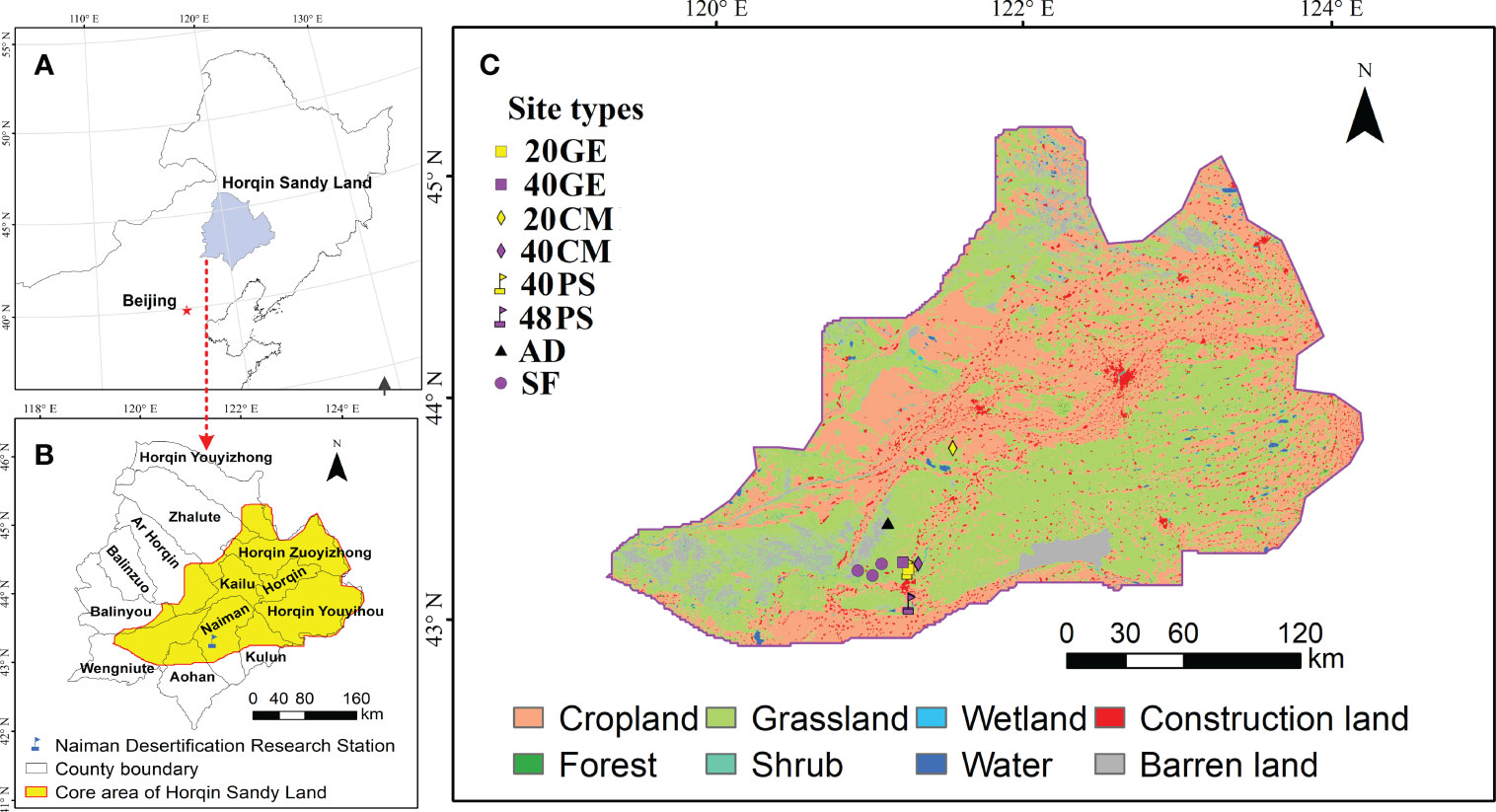
Figure 1 Locations of the core area of the Horqin Sandy Land and the different site types sampled in this study. Site types: 20GE, 20-year grazing exclusion; 40GE, 40-year grazing exclusion; 20CM, 20 years after planting Caragana microphylla; 40CM, 40 years after planting Caragana microphylla; 40PS, 40 years after planting Pinus sylvestris var. mongolica; 48PS, 48 years after planting Pinus sylvestris var. mongolica; AD, active dunes; SF, natural sparse-forest grassland.
The zonal soils are Kastanozems and Chernozems (FAO, 2006) derived from Quaternary alluvial sediments, but the soils have changed to Cambic Arenosols in large areas due to desertification. The native vegetation is dominated by palatable grass species with sparsely scattered woody species, which are dominated by Ulmus macrocarpa, Ulmus pumila, Crataegus pinnatifida, Armeniaca (Prunus) sibirica, and Lespedeza bicolor (Liu and Zhao, 1993). The vegetation has been dominated by xerophytes and psammophytes as a result of excessive grazing and agricultural reclamation, with the dominant species being Salix gordejevii, Periploca sepium, C. microphylla, Artemisia halodendron, and Pennisetum flaccidum (Liu and Zhao, 1993).
We conducted field investigation and sampling from July to August 2022 using the space-for-time substitution approach. Our study was performed near the Naiman Desertification Research Station of the Chinese Academy of Sciences (42.93°N, 120.70°E, 377 m asl.). Based on site investigations and site location information obtained in our previous research (Li et al., 2012; Li et al., 2013; Li et al., 2017), we selected the following restoration site types: (1) 38-year-old and 48-year-old P. sylvestris plantation, (2) 20-year-old and 40-year-old C. microphylla plantation, and (3) 22-year-old and 41-year-old grazing exclusion sites (Figure S1). Before implementing restoration, all of the site types were active dunes, which had similar soil parent material, climate, and topography and not been subjected to any management measures. For the convenience of statistical analysis, we defined 22-year-old and 41-year-old grazing exclusion sites as 20-year-old and 40-year-old grazing exclusion sites, respectively; and defined 38-year-old P. sylvestris site as 40-year-old P. sylvestris site. In addition, we surveyed areas with active dunes (i.e., severely degraded sandy grassland, with an equivalent restoration age of 0 years) and natural sparse-forest grassland (the climax community stage) as references (Figure S1). In total, we examined eight site types (Figure 1, Table S1), and each site type comprised three independent replicates. We randomly established three 20 m×20 m plots at each site for trees and shrubs investigations (Table S2), and established three 1 m×1 m quadrats along the diagonal of each plot at an interval of 10 m for herbs investigation and soil sampling.
The species composition, the density, height, and vegetation cover of each herbaceous species were recorded in each quadrat. The aboveground living herbaceous plants and litter were then collected. We performed soil sampling in the same quadrats where the vegetation had been removed. For the grazing exclusion, natural sparse-forest grassland, and active dune sites, we collected five soil samples to a depth of 20 cm from the quadrats using a soil auger with a diameter of 2.5 cm (Figure S2A) and then combined them to produce a well-mixed composite sample. For the afforestation sites, we tried to avoid the “islands of fertility effect” of shrubs and the possible effect of tree roots on the rhizosphere soil. We then randomly established three sampling locations (corresponding to the three 1 m × 1 m quadrats) along the diagonal of the surrounding plot at an interval of 10 m in each plot. We collected soil samples at 20-cm intervals between woody plants adjacent to the vegetation sampling location (Figure S2B) and then mixed them thoroughly to form a composite sample. The soil samples were sieved through a 2-mm mesh to remove stones, animal debris, and plant residues, then were transported to the laboratory. In total, we collected 216 soil samples (eight site types × three replicate sites × three replicate plots × three replicate quadrats). We chose two additional locations in each quadrat to collect soil cores to a depth of 20 cm using a cylindrical 100-cm3 sampler to determine soil bulk density (BD), field water capacity (FC), and the saturated water content (SAW).
Soil samples were air-dried before analysis of the physicochemical properties. We determined SOC, TN, and TP concentrations (in g·kg−1) using the Walkley-Black dichromate oxidation method, Kjeldahl method, and molybdenum–antimony colorimetric method, respectively (Chen et al., 2022c). Soil pH (1:2.5 w:v) and electrical conductivity (EC, in μS·cm−1; 1:5 w:v) were measured in deionized water using PHS-3C and DDS-307A instruments (INESA Scientific Instrument Co., Ltd., Shanghai, China), respectively. Soil water content (SWC) and BD were measured after oven-drying the soil samples at 105 °C for 48 hours to constant weight. The living plants and litter were dried at 75 °C for 48 hours to constant weight and then weighed to measure the aboveground living biomass (AGB, in g·m−2) and litter biomass (LB, in g·m−2), respectively. We determined FC, SAW, and the particle-size distribution using the methods described by Chen et al. (2022b). The soil samples were separated into five fractions: coarse sand (from 2.00 to 0.50 mm), medium sand (from 0.50 to 0.25 mm), fine sand (from 0.25 to 0.10 mm), very fine sand (from 0.10 to 0.05 mm), and silt and clay (<0.05 mm). We also calculated the Shannon–Wiener diversity index (H) in each quadrat (Ma and Liu, 1994).
Where S represents the total number of species in a quadrat, Pi is the proportion of the total number of species accounted for by a given species “i”.
We conducted normality and homogeneity of variance tests using the Kolmogorov-Smirnov test and the Levene test, respectively. If needed, we used ln-transformation to meet the assumptions of normality and homogeneity. Two-way ANOVA was used to analyze the effects of restoration measures, restoration ages, and their interactions on soil C:N:P stoichiometry. We then used one-way ANOVA followed by LSD test to test for differences in soil C:N:P stoichiometry among different restoration measures and restoration ages at the significant level of α = 0.05. The statistical analyses were conducted in SPSS (version 20.0; https://www.ibm.com/analytics/spss-statistics-software). We used Origin 2018 (https://www.originlab.com/) to perform linear regression to fit the relationships among SOC, TN, and TP. We calculated Pearson’s correlation coefficient (r) using the “cor” function in the R software to quantify the relationships among soil C:N:P stoichiometry and environmental factors. We used version 0.92 of the “corrplot” package for R (https://cran.r-project.org/web/packages/corrplot/index.html) to visualize the correlation matrix. Given the strong correlation between factors, we operated principal component analysis (PCA) (He et al., 2022) using SPSS to achieve dimensionality reduction and calculate scores (Table S3). Structural equation modeling (SEM) was then used to quantify the relative contribution of the factors regulating soil C:N:P stoichiometry using AMOS 24.0 (http://amosdevelopment.com/). The standardized path coefficients were used to indicate correlations between factors in SEM.
The restoration measures and restoration ages significantly affected SOC, TN, and TP and the resulting stoichiometric ratios (Figure 2 and Table 1). Two-way ANOVA indicated that the SOC, TN, TP, C:N, and N:P were significantly affected by both site types and restoration ages and their interaction (Table 1; p < 0.01). The C:P was affected by site types and restoration ages independently (p < 0.01), while C:P was not affected by the interaction between site types and restoration ages (p > 0.05). The SOC, TN, and TP concentrations were the highest in the natural sparse-forest grassland. The SOC, TN, TP concentrations, and C:N, C:P, and N:P ratios of soils under the three restoration measures were significantly (p < 0.05) greater than those in the active dunes, with increases of 7.4 to 13.1, 4.5 to 8.8, 0.6 to 1.2, 1.4 to 2.1, 4.4 to 7.6, and 3.1 to 4.3 times, respectively. The 40-year grazing exclusion had the highest SOC, TN, and TP among the restoration measures. In addition, the SOC, TN, and TP concentrations and three stoichiometric ratios of the 40-year grazing exclusion were closest to the corresponding values of natural sparse-forest grassland. The SOC, TN, TP, C:P, and N:P of the 40-year grazing exclusion increased by 58.2, 77.1, 30.9, 20.5, and 35.1%, respectively, compared with the 20-year grazing exclusion. In contrast, the C:N of the 40-year grazing exclusion decreased by 10.6% compared with the 20-year grazing exclusion. Compared with the 20-year C. microphylla plantations, the SOC, TN, C:P, and N:P of the 40-year C. microphylla plantations increased by 26.6, 19.0, 26.0, and 18.3%, respectively. The TP and C:N of C. microphylla plantations did not change significantly with increasing restoration age (p > 0.05). Compared with the 40-year P. sylvestris plantations, the SOC, C:N, and C:P of the 48-year P. sylvestris plantations increased by 16.3, 14.4, and 15.5%, respectively. The TN, TP, and N:P of P. sylvestris plantations did not change significantly (p > 0.05) with increasing restoration age. In addition, the SWC content of C. microphylla and P. sylvestris plantations decreased with the increase of afforestation years.
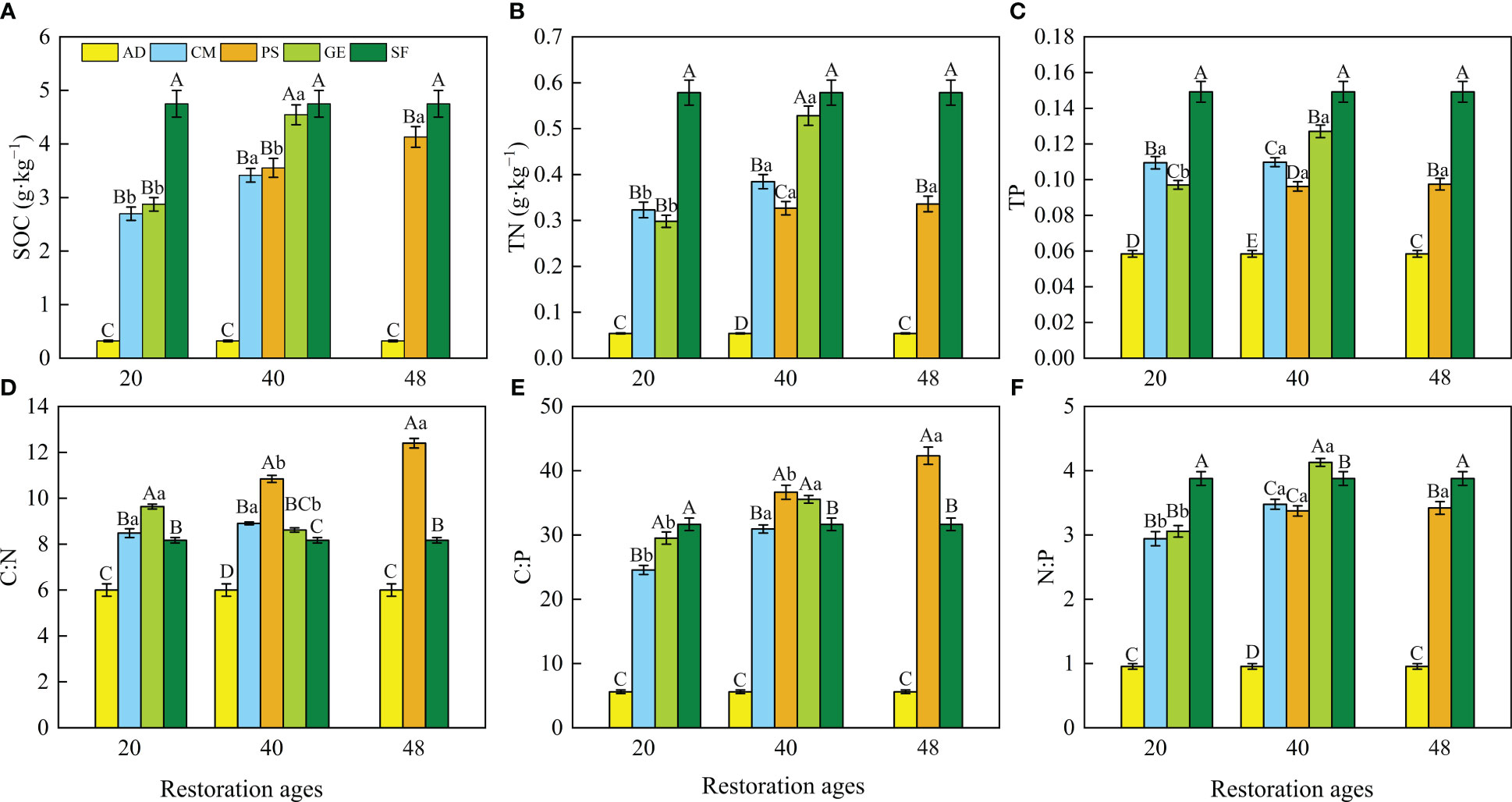
Figure 2 (A) Soil organic carbon (SOC), (B) total nitrogen (TN), and (C) total phosphorus (TP), and the (D) soil organic carbon:total nitrogen (C:N) ratio, (E) soil organic carbon:total phosphorus (C:P) ratio, and (F) total nitrogen:total phosphorus (N:P) ratio for the eight site types. Different lowercase letters indicate significant difference among different restoration ages at the same site types (p < 0.05). Different capital letters indicate the significant difference among different site types at the same restoration age (p < 0.05). Values are means ± SE (n=27 for each site type). AD, active dunes; SF, natural sparse-forest grassland; CM, C. microphylla plantation; GE, grazing exclusion; PS, P. sylvestris plantation.
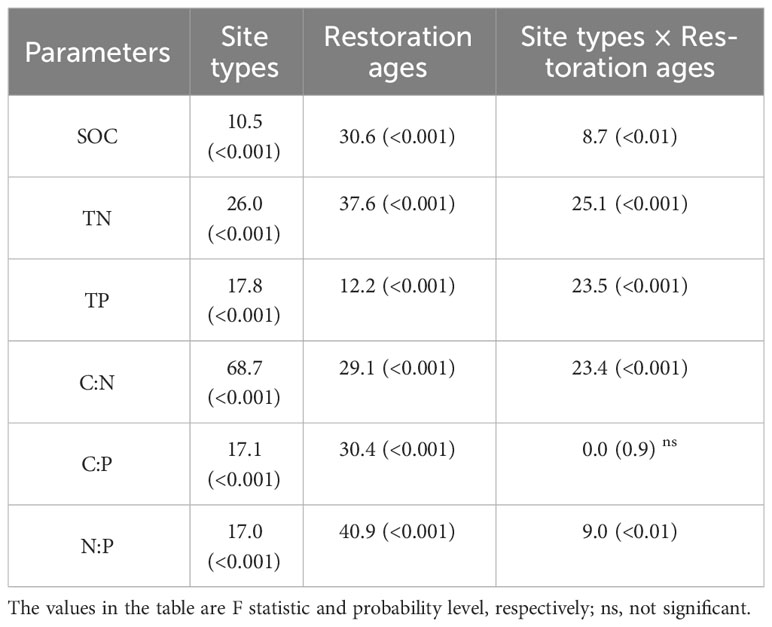
Table 1 The two-way ANOVA on the effects of site types and restoration ages on soil organic carbon (SOC), total nitrogen (TN), and total phosphorus (TP) concentrations and the resulting stoichiometric ratios (C:N:P).
Except for active dunes, we found a significant positive linear relationship between SOC, TN, and TP for all site types (p < 0.001, Table 2, Figure S3). The R2 values for the C–N, C–P, and N–P relationships of the other seven site types were 0.85 to 0.98, 0.39 to 0.86, and 0.47 to 0.91, respectively (Table 2). As restoration age increased, the R2 values for the C–P and N–P relationships increased under grazing exclusion, whereas the R2 for C. microphylla plantations increased for all three relationships. In contrast, R2 of the C–N and C–P relationships for P. sylvestris plantations decreased, and R2 of N–P in P. sylvestris plantations remained relatively constant with increasing restoration age (Table 2).
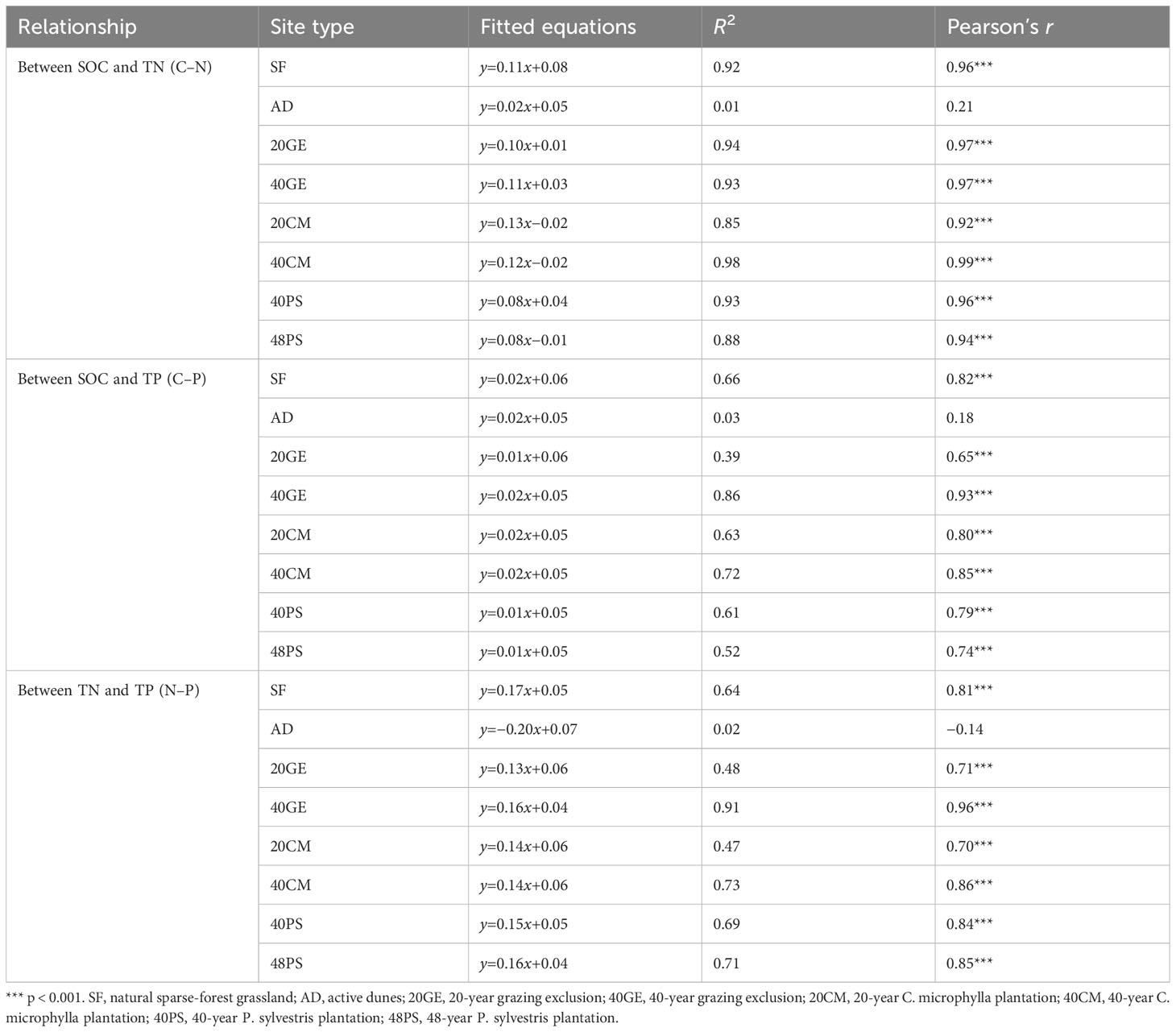
Table 2 The relationships among soil organic carbon (SOC), total nitrogen (TN), and total phosphorus (TP) for the eight site types.
Silt and clay contents, EC, H, and AGB were positively correlated with SOC, TN, TP, C:P and N:P. Elevation and LB were positively correlated with SOC, C:N, C:P and N:P. Soil BD and SAW were negatively correlated with SOC, C:N, C:P and N:P. Soil pH, SWC, and FC were positively correlated with TN and TP, but negatively correlated with C:N. Slope and aspect were negatively correlated with TN and TP, but positively correlated with C:N (p < 0.05, Figure 3). Soil C:N:P stoichiometry was directly affected by restoration measures and soil properties (Figure 4). Restoration measures exhibited the highest direct effect (with the standardized path coefficient of 0.69, p < 0.001) on soil C:N:P stoichiometry (Figure 4). In addition, restoration measures and topography indirectly affected soil C:N:P stoichiometry by regulating vegetation and soil properties. Vegetation indirectly affected soil C:N:P stoichiometry by regulating soil properties (Figure 4). All predictor variables together explained 39.3% of the variation in soil C:N:P stoichiometry.
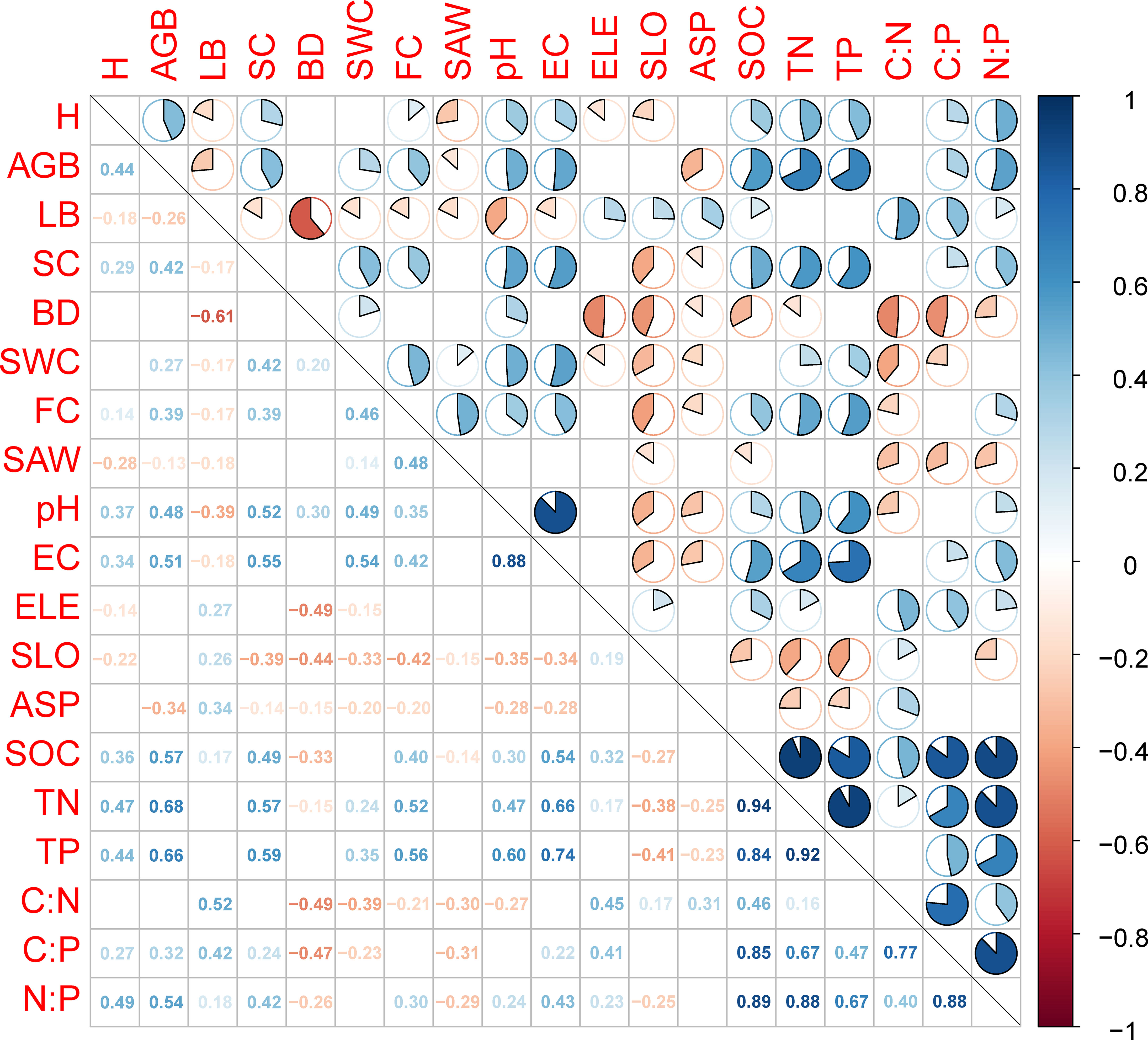
Figure 3 The correlation coefficient matrix (Pearson’s r) for the relationships between pairs of environmental factors and soil organic carbon (SOC), total nitrogen (TN), and total phosphorus (TP) and the resulting stoichiometric ratios (C:N:P). Numbers below the main diagonal are the correlation coefficients (p < 0.05). Above the main diagonal, correlation coefficients are displayed in a pie chart. A positive correlation is represented by blue (p < 0.05), whereas a negative correlation is displayed as red (p < 0.05). Cells with a non-significant correlation coefficient are not shown (p > 0.05). Variables: AGB, aboveground living biomass; ASP, aspect; BD, soil bulk density; C:N, soil organic carbon:total nitrogen; C:P, soil organic carbon:total phosphorus; EC, electrical conductivity; ELE, elevation; FC, field water capacity; H, Shannon–Wiener diversity index; LB, litter biomass; N:P, total nitrogen:total phosphorus; SAW, saturated water content; SC, silt and clay; SLO, slope; SWC, soil water content.
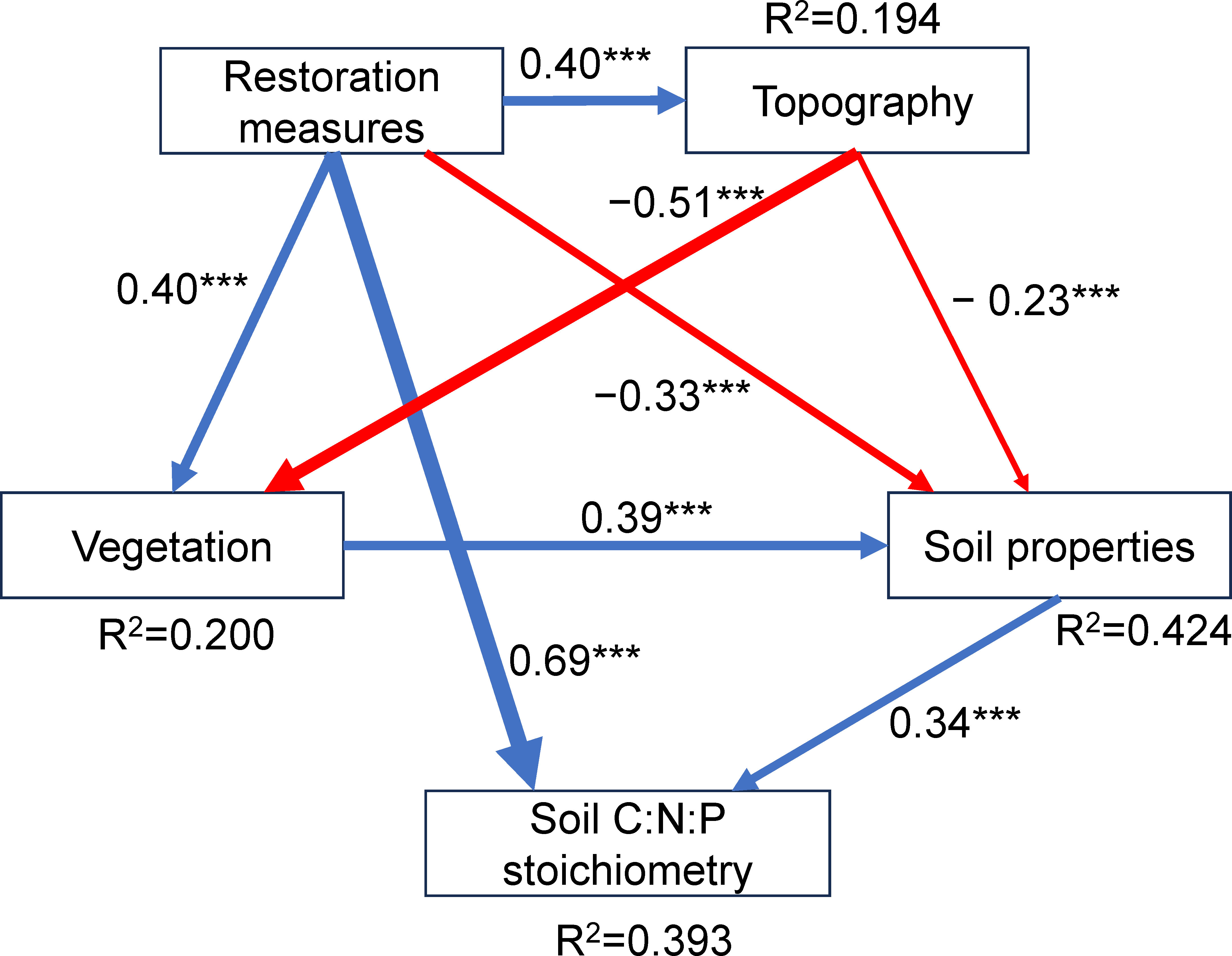
Figure 4 Structural equation models (SEM) revealing the direct and indirect effects of environmental factors and restoration measures on soil C:N:P stoichiometry. The numbers are the standardized path coefficients and R2. Blue and red solid arrows indicate positive and negative associations, respectively. Non-significant paths are not shown. ***p < 0.001. The model fit summary: p = 0.59, χ2/df = 0.53, GFI = 1.00, AGFI = 0.99, CFI = 1.00, RMSEA = 0.00.
Afforestation and grazing exclusion in areas with active dunes can help to reverse desertification by enhancing soil fertility. Overall, soil C:N:P stoichiometry of the 40-year grazing exclusion were closest to those of the natural sparse-forest grassland, indicating that the restoration effect of grazing exclusion was better than afforestation in the severely degraded sandy grassland under the perspective of soil C:N:P stoichiometry. The result was consistent with previous study which found that the soil health index increased after 21-year-old grazing exclusion in Brazilian drylands and was close to the level of native vegetation (Lima et al., 2024). However, the SOC, TN, and TP concentrations in the eight site types were significantly lower than previously reported values in Chinese (24.6, 1.9, and 0.8 g·kg−1, respectively; Tian et al., 2010) and in global terrestrial ecosystems (57.2, 4.1, and 0.5 g·kg−1, respectively; Xu et al., 2013). In addition, the SOC, TN, and TP concentrations of grazing exclusion and P. sylvestris sites in our research were lower than those in the fenced and afforested soils of China’s Loess Plateau, respectively (Zeng et al., 2016; Bai et al., 2019), which had a shorter restoration time. These results indicated that SOC and nutrients in the Horqin Sandy Land were very low. Although vegetation restoration increases the input of plant sources, it has a lower decomposition rate in desertification areas (Zechmeister-Boltenstern et al., 2015). Moreover, compared to vegetation restoration, the restoration of soil fertility is more difficult, and usually takes decades or even hundreds of years in the severely degraded sandy grasslands (Li et al., 2007).
Except for the 48-year P. sylvestris, whose C:N (12.4) was slightly higher than that in China as a whole, the C:N values of the remaining seven site types and the C:P and N:P values of all site types in the Horqin Sandy Land were lower than those in China as a whole, with values of 12.4, 52.7, and 4.2, respectively (Tian et al., 2010) and in global terrestrial ecosystems, with values of 14.1, 111.1, and 7.9, respectively (Xu et al., 2013). The C:N of the 20-year grazing exclusion in our research was higher and C:P and N:P were lower than those in the 17 years of grazing exclusion in degraded semi-arid region in Brazil (Filho et al., 2019). Moreover, the C:N of the 48-year P. sylvestris plantations in our research was higher than that (10.3) in the 50-year afforested soils of Mediterranean mountain agroecosystems (Nadal-Romero et al., 2023). These results indicated that soil in our study region lacks C and N. In other words, the decomposition rate of plant litter is relatively slow, resulting in insufficient soil C and N accumulation (Zechmeister-Boltenstern et al., 2015). Wang et al. (2014) suggested that the C:N ratio was relatively consistent under different land-use and cover types in a subtropical wetland, including sites that received nitrogenous fertilizer. There is a strong positive correlation between C and N (r = 0.94) in our study. Because N is distributed in several types of organic polymers and humus, and organic N is both a N source and a C source. Therefore, when microorganisms decompose organic N, they also affect the mineralization flux of C (Zechmeister-Boltenstern et al., 2015). As a result, the synchronous changes of C and N lead to relatively stable ratios. The tight coupling of C and N for most site types supports this conclusion.
With increasing restoration age, the N:P under grazing exclusion increased from 3.1 to 4.1, which was close to the national mean values (4.2) (Tian et al., 2010) and much higher than that in active dunes (1.0). In addition, the enhanced N–P relationship under grazing exclusion supports the conclusion that soil N:P converged after long-term grazing exclusion in Inner Mongolia grasslands (Yang et al., 2017). The C:P and N:P of 20-year C. microphylla plantations were higher than those of a 20-year-old Caragana korshinskii plantation on the Loess Plateau (Chen et al., 2022a). The N:P of C. microphylla plantations increased with increasing age, suggesting that N limitation was mitigated but P limitation was exacerbated with increasing time since restoration, which partially supports our second hypothesis. Phosphate fertilizers should therefore be applied at C. microphylla sites in the later stage of the restoration. There may be two reasons for these changes: First, the silt and clay content at C. microphylla sites increased significantly compared with that in active dunes because C. microphylla has pinnate leaves that can capture blowing soil fine particles (which are rich in C and N) and plant debris (Zhao et al., 2007). Second, C. microphylla is a leguminous plant species that can fix N (Li et al., 2023b).
Pinus sylvestris var. mongolica afforestation greatly improved soil fertility, because afforestation both stabilizes dunes by reducing wind speed and contributes a large amount of high-quality litter to the soil (Zhang et al., 2019b; Wu et al., 2020). However, P. sylvestris is a tree species with a shallow root system, resulting in a vulnerability to fluctuations in soil moisture and severe drought stress in the shallow rooting zone (Song et al., 2017). Previous studies have shown that vegetation restoration decreased SWC in arid and semi-arid areas (Deng et al., 2019), and the low moisture content may have decreased microbial biomass and decomposition rate of litter (Borken et al., 2006). In addition, soil N mineralization was affected by changes in soil moisture (Beier et al., 2008). In this study, soil moisture decreases with increasing age of P. sylvestris afforestation may result in insignificant changes in N and P nutrients and increasing C:N with increasing age (Beier et al., 2008; Tao et al., 2020). Soil in these plantations may become N-limited over time. We therefore recommend measures such as irrigation and application of nitrogenous fertilizer in the later stages of P. sylvestris afforestation to avoid the development of dried soil layers and to improve soil fertility.
Previous studies have shown that soil stoichiometry in restored ecosystems was most strongly affected by the vegetation type (Su et al., 2019; Wang et al., 2022), restoration age (Su and Shangguan, 2021), soil properties (Yang and Liu, 2019), and management measures (Tao et al., 2021). In the present study, vegetation restoration is the main reason for improving SOC sequestration and soil fertility in severely desertified land. Those improvements enhanced markedly over time after vegetation restoration, which is consistent with the previous study of shrub afforestation in the Tengger Desert (Yang et al., 2014). These results supported our first hypothesis. Firstly, afforestation and grazing exclusion have formed actual protected areas in the areas with active dunes, increasing surface roughness to reduce wind speed and reducing human activities interference (e.g., grazing), which protect soil from wind erosion (Han et al., 2023). Secondly, afforestation and grazing exclusion have increased the productivity, species richness, and diversity, resulting in the increase of soil C and nutrient sources by increasing the quantity and quality of litter and root exudates (Lange et al., 2015; Yang et al., 2019; Zhang et al., 2019b). In addition, the diversity of soil root exudates and litter inputs after restoration not only stimulates microbial metabolism, but also provides abundant energy and nutrients for decomposers, which is beneficial for the accumulation of C, N, and P (Zhang et al., 2018b). The previous study found that roots had a greater impact on soil nutrients than other plant components, which was related to the rhizosphere microbial community and root exudates (Zeng et al., 2016). Although we did not attempt to quantify the contributions of roots to SOM and nutrients, the sampling depth was only 20 cm in the present study, which was the main rooting zone of herbaceous plants (Yang et al., 2017). The increase in herbaceous plant richness promotes the improvement of fertility in the topsoil due to the short life history, the large amount of root residues, and high litter turnover rate of herbaceous plant (Wei et al., 2012; Wang et al., 2021). This may partially explain why the effectiveness of the grazing exclusion is better than afforestation.
Soil properties are another important influencing factor for soil C:N:P stoichiometry, which is consistent with previous study (Tian et al., 2018). Vegetation restoration and changes in topography caused by grassland degradation–restoration process affects soil C:N:P stoichiometry by altering soil structure and chemical properties. Soil texture and BD are important parameters for evaluating soil compaction strength and soil structure, and play an important role in regulating the litter decomposition, root growth, species composition, and community structure (Dodd et al., 2002; Don et al., 2011). Long term vegetation restoration reduces BD and increases soil fine particulate matter, promoting the formation of soil aggregates. In this context, it is beneficial for SOC sequestration and nutrient accumulation (Zhou et al., 2008; Chen et al., 2022b; Wang et al., 2022). Soil BD is greatly affected by porosity, and soils with low BD were porous. Therefore, soil texture and structure typically alter soil moisture and have an impact on soil C:N:P stoichiometry. Because SWC can change litter decomposition and nutrient release mediated by decomposers (Cui et al., 2020). Borken et al. (2006) showed that low SWC adversely affected microbial metabolism. Water is the main limiting factor for plant primary productivity in arid and semi-arid regions (Wang et al., 2018), and this may explain the negative correlation between SWC and the C:N ratios and the positive correlation between SWC and nutrients. Therefore, soil water deficit caused by the increase in afforestation years may have adverse effects on SOC sequestration and soil fertility.
Soil pH and EC are parameters of soil quality, and have an impact on soil C:N:P stoichiometry by affecting soil enzyme and microbial activity (Zhang et al., 2019c; Qiao et al., 2020; Zhang et al., 2021). Soil pH and EC also indirectly change the microbial community by changing the soil structure and the availability of nutrients (Chen et al., 2022b). For example, bacterial and fungal communities differed significantly between soils with different pH (Puissant et al., 2019). Fungi have lower nutrient requirements and slower metabolic activity than bacteria, so they affect soil stoichiometry at different rates (Zechmeister-Boltenstern et al., 2015; Zhou et al., 2023). An experiment with manipulation of desert grasslands in northern China showed that the availability of nitrate N increased after soil acidification, whereas soil alkalization reduced total C and TN (Liu et al., 2022). In the present study, we found a positive correlation between pH and TP, which was inconsistent with the conclusions drawn by Wang et al. (2014) in subtropical wetlands. This may be due to the large climatic differences between the two regions.
Our results indicated that afforestation and grazing exclusion in active dunes contributed to increasing SOC, TN, and TP. The SOC, TN, and TP and the resulting stoichiometric ratios in the 40-year grazing exclusion were close to those of natural sparse-forest grassland (which represents the undamaged natural ecological condition), indicating that grazing exclusion was more beneficial for restoring SOC and nutrient balance than afforestation. With increasing duration (from 20 to 40 years), the N:P increased under grazing exclusion, which was close to the national mean values. In addition, the R2 of the N–P relationship under grazing exclusion increased, indicating that N:P converged with increasing time since restoration. The N:P of C. microphylla sites increased with increasing restoration age (from 20 to 40 years), suggesting that N limitation was mitigated whereas P limitation was exacerbated. Therefore, phosphate fertilizers should be applied later during the restoration process. With increasing afforestation age (from 40 to 48 years), the SOC and C:N in the P. sylvestris plantation increased. Therefore, the soil may be subject to an increasing N limitation over time. We recommend applying nitrogenous fertilizers in the later stages of P. sylvestris afforestation. Moreover, soil drought stress may be a challenge that plantations will face in the future. Vegetation restoration is the main factor directly leading to changes in soil C:N:P stoichiometry, and indirectly affects soil C:N:P stoichiometry by altering soil structure and chemical properties. Our findings provide a framework for linking ecological restoration measures to soil stoichiometry in semi-arid regions.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
WC: Investigation, Software, Visualization, Writing – original draft. YL: Conceptualization, Funding acquisition, Project administration, Supervision, Writing – review & editing. YC: Funding acquisition, Investigation, Software, Writing – review & editing. XW: Funding acquisition, Methodology, Writing – review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was supported by the Strategic Priority Research Program of the Chinese Academy of Sciences (grant number XDA26020104), the National Natural Science Foundation of China (grant numbers 42301080, 31971466, 32001214), and Grassland Elite Projects of Inner Mongolia Autonomous Region (grant number E339050701).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2023.1326244/full#supplementary-material
C, carbon; N, nitrogen; P, phosphorus; SOC, soil organic carbon; TN, total nitrogen; TP, total phosphorus; C:N, soil organic carbon:total nitrogen; C:P, soil organic carbon:total phosphorus; N:P, total nitrogen:total phosphorus; C:N:P, soil organic carbon:total nitrogen:total phosphorus; SOM, soil organic matter; BD, bulk density; EC, electrical conductivity; SWC, soil water content; FC, field water capacity; SAW, saturated water content; AGB, aboveground living biomass; LB, litter biomass; H, Shannon–Wiener diversity index.
Bai, X., Wang, B., An, S., Zeng, Q., Zhang, H. (2019). Response of forest species to C:N:P in the plant-litter-soil system and stoichiometric homeostasis of plant tissues during afforestation on the Loess Plateau, China. Catena 183, 104186. doi: 10.1016/j.catena.2019.104186
Beier, C., Emmett, B., Peñuelas, J., Schmidt, I., Tietema, A., Estiarte, M., et al. (2008). Carbon and nitrogen cycles in European ecosystems respond differently to global warming. Sci. Total Environ. 407, 692–697. doi: 10.1016/j.scitotenv.2008.10.001
Borken, W., Savage, K., Davidson, E., Trumbore, S. (2006). Effects of experimental drought on soil respiration and radiocarbon efflux from a temperate forest soil. Global Change Biol. 12, 177–193. doi: 10.1111/j.1365-2486.2005.001058.x
Cao, W., Li, Y., Chen, Y., Chen, Y., Wang, X. (2023). Spatial patterns of soil stoichiometry and their responses to land use in a desertified area: A case study of China's Horqin Sandy Land. Land Degrad. Dev. doi: 10.1002/ldr.4920
Chen, Y., Li, Y., Cao, W., Wang, X., Duan, Y., Liu, X., et al. (2022b). Response of the plant–soil system to desertification in the Hulun Buir Sandy Land, China. Land Degrad. Dev. 34, 2024–2037. doi: 10.1002/ldr.4586
Chen, Y., Li, Y., Duan, Y., Wang, L., Wang, X., Yao, C., et al. (2022c). Patterns and driving factors of soil ecological stoichiometry in typical ecologically fragile areas of China. Catena 219, 106628. doi: 10.1016/j.catena.2022.106628
Chen, M., Yang, X., Shao, M., Wei, X., Li, T. (2022a). Changes in soil C–N–P stoichiometry after 20 years of typical artificial vegetation restoration in semiarid continental climate zones. Sci. Total Environ. 852, 158380. doi: 10.1016/j.scitotenv.2022.158380
Cui, Y., Wang, X., Zhang, X., Ju, W., Duan, C., Guo, X., et al. (2020). Soil moisture mediates microbial carbon and phosphorus metabolism during vegetation succession in a semiarid region. Soil Biol. Biochem. 147, 107814. doi: 10.1016/j.soilbio.2020.107814
Deng, L., Peng, C., Huang, C., Wang, K., Liu, Q., Liu, Y., et al. (2019). Drivers of soil microbial metabolic limitation changes along a vegetation restoration gradient on the Loess Plateau, China. Geoderma 353, 188–200. doi: 10.1016/j.geoderma.2019.06.037
Dijkstra, F., Pendall, E., Morgan, J., Blumenthal, D., Carrillo, Y., LeCain, D., et al. (2012). Climate change alters stoichiometry of phosphorus and nitrogen in a semiarid grassland. New Phytol. 196, 807–815. doi: 10.1111/j.1469-8137.2012.04349.x
Dodd, M., Lauenroth, W., Burke, I., Chapman, P. (2002). Associations between vegetation patterns and soil texture in the shortgrass steppe. Plant Ecol. 158, 127–137. doi: 10.1023/A:1015525303754
Don, A., Schumacher, J., Freibauer, A. (2011). Impact of tropical land-use change on soil organic carbon stocks – a meta-analysis. Global Change Biol. 14, 1658–1670. doi: 10.1111/j.1365-2486.2010.02336.x
Du, C., Gao, Y. (2021). Grazing exclusion alters ecological stoichiometry of plant and soil in degraded alpine grassland. Agr. Ecosyst. Environ. 308, 107256. doi: 10.1016/j.agee.2020.107256
Duan, H., Wang, T., Xue, X., Liu, S., Guo, J. (2014). Dynamics of aeolian desertification and its driving forces in the Horqin Sandy Land, Northern China. Environ. Monit. Assess. 186, 6083–6096. doi: 10.1007/s10661-014-3841-3
FAO (2006). “FAO/IUSS Working Group WRB, World reference base for soil resources 2006,” in World soil resources reports, vol. 103. (Rome: Food and Agriculture Organization).
Filho, J., Vieira, J., Silva, E., Oliveira, J., Pereira, M., Brasileiro, F. (2019). Assessing the effects of 17 years of grazing exclusion in degraded semi-arid soils: Evaluation of soil fertility, nutrients pools and stoichiometry. J. Arid Environ. 166, 1–10. doi: 10.1016/j.jaridenv.2019.03.006
Geng, J., Tan, Q., Lv, J., Fang, H. (2024). Assessing spatial variations in soil organic carbon and C:N ratio in Northeast China's black soil region: Insights from Landsat-9 satellite and crop growth information. Soil Till. Res. 235, 105897. doi: 10.1016/j.still.2023.105897
Han, Y., Zhao, W., Zhou, A., Pereira, P. (2023). Water and wind erosion response to ecological restoration measures in China's drylands. Geoderma 435, 116514. doi: 10.1016/j.geoderma.2023.116514
He, M., Fang, K., Chen, L., Feng, X., Qin, S., Kou, D., et al. (2022). Depth-dependent drivers of soil microbial necromass carbon across Tibetan alpine grasslands. Glob. Change Biol. 28, 936–949. doi: 10.1111/gcb.15969
Jia, B., Jia, L., Zhang, Y., Mou, X., Li, X. (2022). Leguminous Caragana korshinskii evidently enhances microbial necromass carbon accumulation in dryland soils. Catena 215, 106342. doi: 10.1016/j.catena.2022.106342
Lange, M., Eisenhauer, N., Sierra, C., Bessler, H., Engels, C., Griffiths, R., et al. (2015). Plant diversity increases soil microbial activity and soil carbon storage. Nat. Commun. 6, 6707. doi: 10.1038/ncomms7707
Li, Y., Brandle, J., Awada, T., Chen, Y., Han, J., Zhang, F., et al. (2013). Accumulation of carbon and nitrogen in the plant–soil system after afforestation of active sand dunes in China's Horqin Sandy Land. Agr. Ecosyst. Environ. 177, 75–84. doi: 10.1016/j.agee.2013.06.007
Li, Y., Chen, Y., Wang, X., Niu, Y., Lian, J. (2017). Improvements in soil carbon and nitrogen capacities after shrub planting to stabilize sand dunes in China’s Horqin Sandy Land. Sustainability 9, 662. doi: 10.3390/su9040662
Li, C., Fu, B., Wang, S., Stringer, L., Zhou, W., Ren, Z., et al. (2023a). Climate-driven ecological thresholds in China’s drylands modulated by grazing. Nat. Sustain. 6, 1363–1372. doi: 10.1038/s41893-023-01187-5
Li, X., He, M., Duan, Z., Xiao, H., Jia, X. (2007). Recovery of topsoil physicochemical properties in revegetated sites in the sand-burial ecosystems of the Tengger Desert, northern China. Geomorphology 88, 254–265. doi: 10.1016/j.geomorph.2006.11.009
Li, D., Liu, Y., Yang, X., Zhang, X., Shi, Z. (2023b). Shrub encroachment alters plant trait response to nitrogen addition in a semi-arid grassland. Front. Plant Sci. 14. doi: 10.3389/fpls.2023.1103371
Li, F., Zhao, L., Zhang, H., Zhang, T., Shirato, Y. (2004). Wind erosion and airborne dust deposition in farmland during spring in the Horqin Sandy Land of eastern Inner Mongolia, China. Soil Till. Res. 75, 121–130. doi: 10.1016/j.still.2003.08.001
Li, Y., Zhou, X., Brandle, J., Zhang, T., Chen, Y., Han, J. (2012). Temporal progress in improving carbon and nitrogen storage by grazing exclosure practice in a degraded land area of China's Horqin Sandy Grassland. Agr. Ecosyst. Environ. 159, 55–61. doi: 10.1016/j.agee.2012.06.024
Lima, A., Cherubin, M., Silva, D., Mota, J., Silva, F., Araujo, A., et al. (2024). Grazing exclusion restores soil health in Brazilian drylands under desertification process. Appl. Soil Ecol. 193, 105107. doi: 10.1016/j.apsoil.2023.105107
Liu, K., Liu, Z., Zhou, N., Shi, X., Lock, T., Kallenbach, R., et al. (2022). Diversity-stability relationships in temperate grasslands as a function of soil pH. Land Degrad. Dev. 33, 1704–1717. doi: 10.1002/ldr.4259
Liu, X., Zhao, H. (1993). Study on the comprehensive Management of ecological Environment in the Horqin Sandy Land (LanZhou: Gansu Science and Technology Press).
Lu, M., Liu, K., Zhang, L., Zeng, F., Song, T., Peng, W., et al. (2022). Stoichiometric variation in soil carbon, nitrogen, and phosphorus following cropland conversion to forest in southwest China. Forests 13 (8), 1155. doi: 10.3390/f13081155
Ma, K., Liu, Y. (1994). Measurement of biotic community diversity I a diversity (Part 2). Biodivers. Sci. 2, 231–239. doi: 10.17520/biods.1994038
MEE, CAS. (2015). National ecological function regionalization. Available at: https://www.mee.gov.cn/gkml/hbb/bgg/201511/t20151126_317777.htm (Accessed May 29, 2023).
Nadal-Romero, E., Khorchani, M., Gaspar, L., Arnáez, J., Cammeraat, E., Navas, A., et al. (2023). How do land use and land cover changes after farmland abandonment affect soil properties and soil nutrients in Mediterranean mountain agroecosystems? Catena 226, 107062. doi: 10.1016/j.catena.2023.107062
Puissant, J., Jones, B., Goodall, T., Mang, D., Blaud, A., Gweon, H., et al. (2019). The pH optimum of soil exoenzymes adapt to long term changes in soil pH. Soil Biol. Biochem. 138, 107601. doi: 10.1016/j.soilbio.2019.107601
Qiao, Y., Wang, J., Liu, H., Huang, K., Yang, Q., Lu, R., et al. (2020). Depth-dependent soil C-N-P stoichiometry in a mature subtropical broadleaf forest. Geoderma 370, 114357. doi: 10.1016/j.geoderma.2020.114357
Song, L., Zhu, J., Zhang, X. (2017). Forestation and management scheme of Pinus sylvestris var. mongolica plantations in sandy lands based on their decline mechanisms. Chin. J. Ecol. 36, 3249–3256. doi: 10.13292/j.1000-4890.201711.002
Sterner, R. W., Elser, J. J. (2002). Ecological stoichiometry: the biology of elements from molecules to the biosphere (Princeton: Princeton University Press).
Su, L., Du, H., Zeng, F., Peng, W., Rizwan, M., Núñez-Delgado, A., et al. (2019). Soil and fine roots ecological stoichiometry in different vegetation restoration stages in a karst area, southwest China. J. Environ. Manage. 252, 109694. doi: 10.1016/j.jenvman.2019.109694
Su, B., Shangguan, Z. (2021). Response of water use efficiency and plant-soil C:N:P stoichiometry to stand quality in Robinia pseudoacacia on the Loess Plateau of China. Catena 206, 105571. doi: 10.1016/j.catena.2021.105571
Tao, T., Liu, L., Small, G., Chen, J., Wang, Y., Sun, X. (2021). The effects of land management patterns on soil carbon sequestration and C:N:P stoichiometry in sloping croplands in southern China. Agr. Ecosyst. Environ. 320, 107584. doi: 10.1016/j.agee.2021.107584
Tao, Y., Zhou, X., Zhang, S., Lu, H., Shao, H. (2020). Soil nutrient stoichiometry on linear sand dunes from a temperate desert in Central Asia. Catena 195, 104847. doi: 10.1016/j.catena.2020.104847
Tian, H., Chen, G., Zhang, C., Melillo, J. M., Hall, C. A. S. (2010). Pattern and variation of C:N:P ratios in China’s soils: a synthesis of observational data. Biogeochemistry 98, 139–151. doi: 10.1007/s10533-009-9382-0
Tian, L., Zhao, L., Wu, X., Fang, H., Zhao, Y., Hu, G. (2018). Soil moisture and texture primarily control the soil nutrient stoichiometry across the Tibetan grassland. Sci. Total Environ. 622-623, 192–202. doi: 10.1016/j.scitotenv.2017.11.331
Wang, M., Gong, Y., Lafleur, P., Wu, Y. (2021). Patterns and drivers of carbon, nitrogen and phosphorus stoichiometry in Southern China's grasslands. Sci. Total Environ. 785, 147201. doi: 10.1016/j.scitotenv.2021.147201
Wang, W., Sardans, J., Zeng, C., Zhong, C., Li, Y., Peñuelas, J. (2014). Responses of soil nutrient concentrations and stoichiometry to different human land uses in a subtropical tidal wetland. Geoderma 232-234, 459–470. doi: 10.1016/j.geoderma.2014.06.004
Wang, S., Wang, X., Han, X., Deng, Y. (2018). Higher precipitation strengthens the microbial interactions in semi-arid grassland soils. Global Ecol. Biogeogr. 27, 570–580. doi: 10.1111/geb.12718
Wang, L., Zhang, G., Zhu, P., Xing, S., Wang, C. (2022). Soil C, N and P contents and their stoichiometry as affected by typical plant communities on steep gully slopes of the Loess Plateau, China. Catena 208, 105740. doi: 10.1016/j.catena.2021.105740
Wei, J., Cheng, J., Li, W., Liu, W. (2012). Comparing the effect of naturally restored forest and grassland on carbon sequestration and its vertical distribution in the Chinese Loess Plateau. PloS One 7, e40123. doi: 10.1371/journal.pone.0040123
Winowiecki, L., Vågen, T., Kinnaird, M., O'Brien, T. (2018). Application of systematic monitoring and mapping techniques: Assessing land restoration potential in semi-arid lands of Kenya. Geoderma 327, 107–118. doi: 10.1016/j.geoderma.2018.04.017
Wu, W., Zhang, D., Tian, L., Zhang, H. (2020). Aeolian activities and protective effects of artificial plants in re-vegetated sandy land of Qinghai Lake, China. Chin. Geogr. Sci. 30, 1129–1142. doi: 10.1007/s11769-020-1168-2
Xu, X., Thornton, P., Post, W. (2013). A global analysis of soil microbial biomass carbon, nitrogen and phosphorus in terrestrial ecosystems. Global Ecol. Biogeogr. 22, 737–749. doi: 10.1111/geb.12029
Yang, Z., Baoyin, T., Minggagud, H., Sun, H., Li, F. (2017). Recovery succession drives the convergence, and grazing versus fencing drives the divergence of plant and soil N/P stoichiometry in a semiarid steppe of Inner Mongolia. Plant Soil 420, 303–314. doi: 10.1007/s11104-017-3404-9
Yang, H., Li, X., Wang, Z., Jia, R., Liu, L., Chen, Y., et al. (2014). Carbon sequestration capacity of shifting sand dune after establishing new vegetation in the Tengger Desert, northern China. Sci. Total Environ. 478, 1–11. doi: 10.1016/j.scitotenv.2014.01.063
Yang, Y., Liu, B. (2019). Effects of planting Caragana shrubs on soil nutrients and stoichiometries in desert steppe of Northwest China. Catena 183, 104213. doi: 10.1016/j.catena.2019.104213
Yang, Y., Tilman, D., Furey, G., Lehman, C. (2019). Soil carbon sequestration accelerated by restoration of grassland biodiversity. Nat. Commun. 10, 718. doi: 10.1038/s41467-019-08636-w
Yang, L., Wei, W., Chen, L., Mo, B. (2012). Response of deep soil moisture to land use and afforestation in the semi-arid Loess Plateau, China. J. Hydrol. 475, 111–122. doi: 10.1016/j.jhydrol.2012.09.041
Zechmeister-Boltenstern, S., Keiblinger, K., Mooshammer, M., Penuelas, J., Richter, A., Sardans, J., et al. (2015). The application of ecological stoichiometry to plant-microbial-soil organic matter transformations. Ecol. Monogr. 85, 133–155. doi: 10.1890/14-0777.1
Zeng, Q., Li, X., Dong, Y., An, S., Darboux, F. (2016). Soil and plant components ecological stoichiometry in four steppe communities in the Loess Plateau of China. Catena 147, 481–488. doi: 10.1016/j.catena.2016.07.047
Zhang, Y., Li, P., Liu, X., Xiao, L., Shi, P., Zhao, B. (2019a). Effects of farmland conversion on the stoichiometry of carbon, nitrogen, and phosphorus in soil aggregates on the Loess Plateau of China. Geoderma 351, 188–196. doi: 10.1016/j.geoderma.2019.05.037
Zhang, A., Li, X., Wu, S., Li, L., Jiang, Y., Wang, R., et al. (2021). Spatial pattern of C:N:P stoichiometry characteristics of alpine grassland in the Altunshan Nature Reserve at North Qinghai-Tibet Plateau. Catena 207, 105691. doi: 10.1016/j.catena.2021.105691
Zhang, C., Liu, G., Song, Z., Wang, J., Guo, L. (2018b). Interactions of soil bacteria and fungi with plants during long-term grazing exclusion in semiarid grasslands. Soil Biol. Biochem. 124, 47–58. doi: 10.1016/j.soilbio.2018.05.026
Zhang, W., Liu, W., Xu, M., Deng, J., Han, X., Yang, G., et al. (2019b). Response of forest growth to C:N:P stoichiometry in plants and soils during Robinia pseudoacacia afforestation on the Loess Plateau, China. Geoderma 337, 280–289. doi: 10.1016/j.geoderma.2018.09.042
Zhang, W., Xu, Y., Gao, D., Wang, X., Liu, W., Deng, J., et al. (2019c). Ecoenzymatic stoichiometry and nutrient dynamics along a revegetation chronosequence in the soils of abandoned land and Robinia pseudoacacia plantation on the Loess Plateau, China. Soil Biol. Biochem. 134, 1–14. doi: 10.1016/j.soilblo.2019.03.017
Zhang, J., Yan, X., Su, F., Li, Z., Wang, Y., Wei, Y., et al. (2018a). Long-term N and P additions alter the scaling of plant nitrogen to phosphorus in a Tibetan alpine meadow. Sci. Total Environ. 625, 440–448. doi: 10.1016/j.scitotenv.2017.12.292
Zhao, H., Zhou, R., Su, Y., Zhang, H., Zhao, L., Drake, S. (2007). Shrub facilitation of desert land restoration in the Horqin Sand Land of Inner Mongolia. Ecol. Eng. 31, 1–8. doi: 10.1016/j.ecoleng.2007.04.010
Zhou, R., Li, Y., Zhao, H., Drake, S. (2008). Desertification effects on C and N content of sandy soils under grassland in Horqin, northern China. Geoderma 145, 370–375. doi: 10.1016/j.geoderma.2008.04.003
Keywords: ecological stoichiometry, ecological restoration, restoration age, nutrient limitation, soil physicochemical property
Citation: Cao W, Li Y, Chen Y and Wang X (2023) Grazing exclusion is more beneficial for restoring soil organic carbon and nutrient balance than afforestation on degraded sandy land. Front. Plant Sci. 14:1326244. doi: 10.3389/fpls.2023.1326244
Received: 23 October 2023; Accepted: 11 December 2023;
Published: 21 December 2023.
Edited by:
Bo Zhang, Xinjiang Institute of Ecology and Geography (CAS), ChinaReviewed by:
Ling Ling Chen, Inner Mongolia University, ChinaCopyright © 2023 Cao, Li, Chen and Wang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yuqiang Li, bGl5cUBsemIuYWMuY24=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.