 César Marín
César Marín C. Guillermo Bueno
C. Guillermo Bueno Jianqing Wang
Jianqing Wang Vasilis Kokkoris
Vasilis Kokkoris- 1Centro de Investigación e Innovación para el Cambio Climático (CiiCC), Universidad Santo Tomás, Valdivia, Chile
- 2Amsterdam Institute for Life and Environment, Section Ecology & Evolution, Vrije Universiteit Amsterdam, Amsterdam, Netherlands
- 3Instituto Pirenaico de Ecología, Spanish National Research Council (CSIC), Jaca, Spain
- 4Key Laboratory for Humid Subtropical Eco-geographical Processes of the Ministry of Education, Institute of Geography, Fujian Normal University, Fuzhou, China
Editorial on the Research Topic
Biodiversity and ecosystem-level function of the rhizosphere
How soil biodiversity affects ecosystem functioning has long been a central issue in ecology (Darwin, 1881). Models encompassing measurements of microbial processes and biodiversity exhibit superior explanatory capabilities for terrestrial carbon and nitrogen cycling compared to models that only include environmental variables (Graham et al., 2016). However, soil biodiversity and ecosystem functioning have rarely been jointly investigated (Guerra et al., 2020).
Soil and rhizospheric ecologists are only starting to reveal the correlational and causal relationships between macro-organismal/microbial biodiversity and multiple ecosystem functions and services, concept deemed ‘ecosystem multifunctionality’ (Hector and Bagchi, 2007). Fortunately, research on such multifunctionality has increased over the last few years (Figure 1). For example, Delgado-Baquerizo et al. (2020) found that the biodiversity of all soil taxa is positively related to soil ecosystem multifunctionality (11 functions including pathogen control, soil respiration and phosphorous mineralization, among others) at a global scale. Soil ecosystem multifunctionality was also directly related to soil carbon and indirectly related to climate and plant-related variables (Delgado-Baquerizo et al., 2020). These positive associations between soil-rhizospheric biodiversity and ecosystem functioning are not only derived from correlational studies. When experimentally simplifying the complexity of soil microcosms (by increasingly sieving soil into smaller particles), both Wagg et al. (2019) and Romero et al. (2023) have consistently observed a reduction in all (about 9-10) ecosystem functions and multifunctionality.
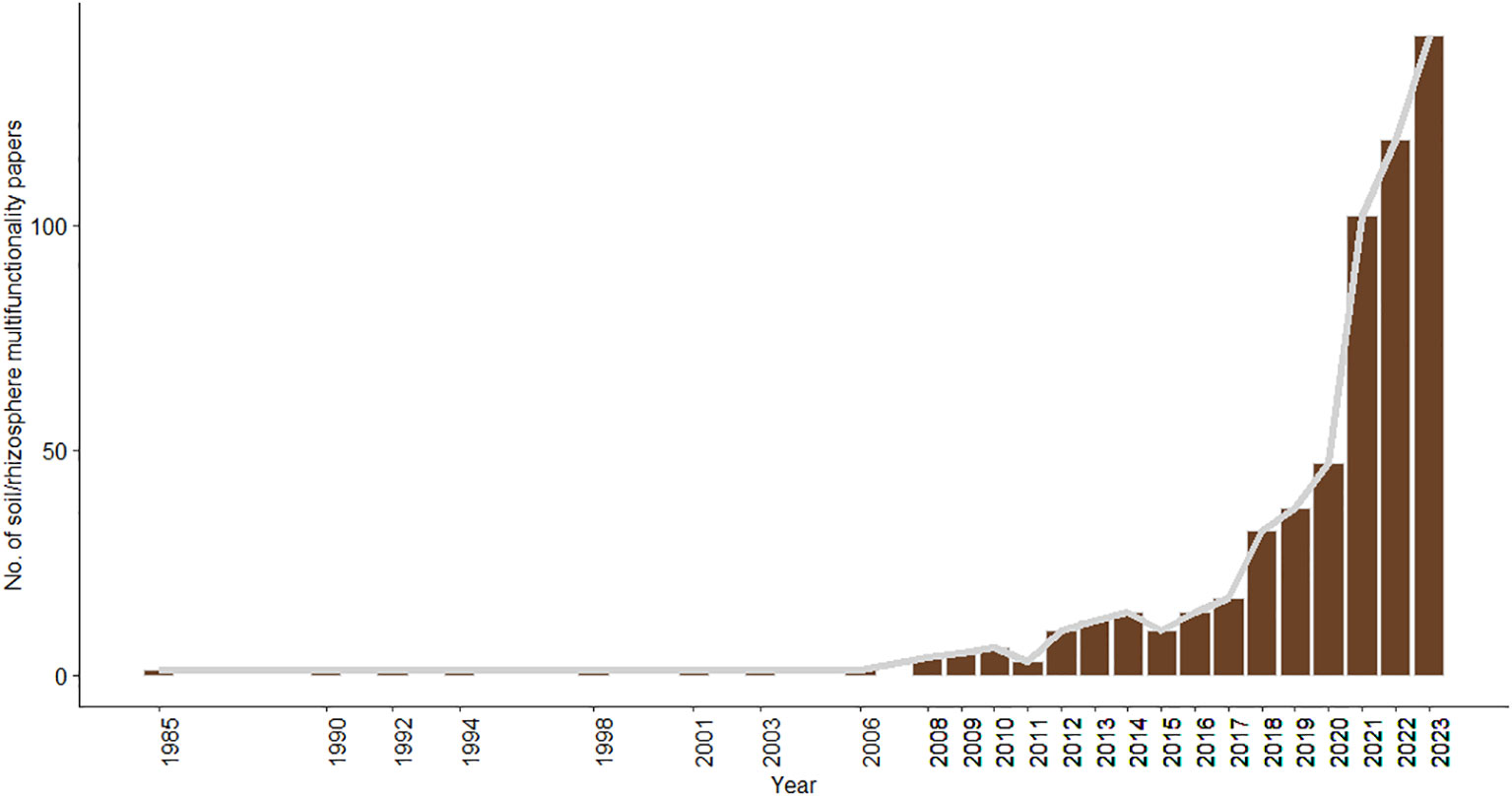
Figure 1 Number of SCOPUS-indexed English (research) articles on soil and rhizospheric ecosystem multifunctionality over the last years. Search terms in SCOPUS (7 August 2023): rhizosph OR soil AND multifunctionality.
Additional research paths on soil/rhizospheric biodiversity and ecosystem functioning relationships include contrasting global biogeographic patterns between flora and soil biota (Cameron et al., 2019; Vasar et al., 2022), understanding functional redundancy and why some ecosystem functions seem to be more related to certain taxa or guilds (Delgado-Baquerizo et al., 2020), and addressing methodological questions on how to accurately assess biodiversity and functions (Guerra et al., 2021). In this Research Topic, we addressed some of these research paths and other aspects related to biodiversity and ecosystem functioning of the rhizosphere. Wang et al. investigated the role of rhizospheric (locally and regionally) rare/abundant/entire bacterial and fungal communities on multiple ecosystem functions in monocultures and intercropped Zea mays croplands in subtropical China. At the local scale the authors found that rare bacteria were associated to ecosystem functions such as N-cycling multifunctionality and aboveground net primary productivity, whereas rare fungi were related to C and P cycling and overall ecosystem multifunctionality. Intercropping increased the network complexity and positive interactions of rare bacteria of the Z. mays rhizosphere. Also in an agronomic setting, Moura et al. found that neither different cover crops (in the Brazilian Cerrado ecosystem) nor N fertilization affected glomalin production and root colonization by arbuscular mycorrhizal fungi (AMF), and AMF spore density was slightly (yet significantly) affected by the different cover crop types evaluated. AMF diversity did not exhibit significant differences across cover crops. These results might be related to the ubiquity, low endemism levels (Davison et al., 2015; Davison et al., 2022), and high resilience of AMF communities, particularly those of the Cerrado (Moura et al., 2022).
Besides investigating abiotic and biotic drivers of AMF communities, Albracht et al. evaluated the effects of recurrent summer droughts (for 9 years) in experimental grasslands differing in plant richness and functional group richness. Plant functional composition, rather than mere plant species richness, shaped fungal communities during recurrent summer droughts. The aboveground components of grasslands also showed distinct patterns. As an illustration, Mao et al. discovered that six years of artificial afforestation led to the gradual development of grassland plant communities. These communities optimized composition, coverage, and aboveground biomass, with post-afforestation grassland diversity nearing that of a naturally recovering 10-year abandoned community.
As previously mentioned, understanding the global biogeography of soil biota remains a central concern in soil ecology. In line with this, Sepp et al. collected 327 soil samples around the globe. Their study employed metabarcoding sequencing of the nifH gene and the general prokaryotic 16S SSU region to delve into the global distribution of nitrogen-fixing bacteria in soils. Globally, the distribution of N-fixing bacteria was primarily influenced by climatic conditions, and soil pH and N content, with variations in these responses observed among specific taxa, such as Cyanobacteria. Similarly, using global-scale metabarcoding to elucidate global plant biodiversity, Vasar et al. found that large-scale plant diversity patterns and community composition elucidated through sequencing of 325 soil samples broadly coincided with the patterns revealed by traditional sources. Approximately half of local GBIF records were represented in this environmental DNA sequencing at the species level.
Changes in soil properties caused by plants, which in turn affect plant performance, termed ‘plant–soil feedback’ (PSF) (Van der Putten et al., 2013), are fundamental to understand relationships between soil biodiversity and functioning. Using a tree productivity gradient from long-term consistent reforestation experiments, Stefani et al. assessed the relationships between microbial communities and soil and tree nutrient stock, as affected by a positive PSF induced by wood mulch amendment in boreal forests. They found that adding such amendment resulted in a microbially-mediated positive PSF that increases mineral weathering and non-symbiotic N fixation. This shift turned unproductive plots into productive ones, contributing to rapid reforestation in a harsh environment. Similarly to Wang et al. regarding intercropping, Stefani et al. found that mulching increased the complexity and connectivity of soil microbial networks. Regarding the current effect of global warming, Wang and Shi calls out to the restoration-involved community, which encompasses forestry experts, legislators, and others, urging them to prioritize natural forest regeneration over different tree-planting techniques. They argue that such tree-planting techniques might have adverse impacts on soil and litter diversity, as well on certain crucial soil parameters such as organic matter, pH, nutrient/water retention, and overall structure.
In summary, the relationship between soil biodiversity and ecosystem functioning is gaining attention. Recent studies reveal correlations between biodiversity and multifunctionality, as well as the impact of factors like rare microbial communities and recurring droughts. These findings underscore the importance of PSF and pollution effects. This Research Topic contributes to understanding soil-rhizospheric interactions, highlighting their significance in ecosystem processes. As we advance, considering global biogeographic patterns, functional redundancies, and assessment methods will further enrich our comprehension.
Author contributions
CM: Conceptualization, Formal Analysis, Writing – original draft, Writing – review & editing. CB: Conceptualization, Writing – review & editing. JW: Conceptualization, Writing – review & editing. VK: Conceptualization, Formal Analysis, Writing – review & editing.
Funding
CM thanks ANID + Convocatoria Nacional Subvención a Instalación en la Academia Convocatoria Año 2021 + Folio No. SA77210019. CB was supported by a Ramon y Cajal fellowship (RYC2021-032533-I) from the Spanish Minister of Science and Innovation.
Acknowledgments
The authors are thankful to the contributors to this Research Topic as well as the Editorial support of the Journal.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Cameron, E. K., Martins, I. S., Lavelle, P., Mathieu, J., Tedersoo, L., Bahram, M., et al. (2019). Global mismatches in aboveground and belowground biodiversity. Conserv. Biol. 33, 1187–1192. doi: 10.1111/cobi.13311
Darwin, C. (1881). The formation of vegetable mould, through the action of worms, with observations on their habits. London.: J. Murray. doi: 10.5962/bhl.title.107559
Davison, J., Moora, M., Öpik, M., Adholeya, A., Ainsaar, L., Bâ, A., et al. (2015). Global assessment of arbuscular mycorrhizal fungus diversity reveals very low endemism. Science 349, 970–973. doi: 10.1126/science.aab1161
Davison, J., Vasar, M., Sepp, S. K., Oja, J., Al-Quraishy, S., Bueno, C. G., et al. (2022). Dominance, diversity, and niche breadth in arbuscular mycorrhizal fungal communities. Ecology 103, e3761. doi: 10.1002/ecy.3761
Delgado-Baquerizo, M., Reich, P. B., Trivedi, C., Eldridge, D. J., Abades, S., Alfaro, F. D., et al. (2020). Multiple elements of soil biodiversity drive ecosystem functions across biomes. Nat. Ecol. Evol. 4, 210–220. doi: 10.1038/s41559-019-1084-y
Graham, E. B., Knelman, J. E., Schindlbacher, A., Siciliano, S., Breulmann, M., Yannarell, A., et al. (2016). Microbes as engines of ecosystem function: when does community structure enhance predictions of ecosystem processes? Front. Microbiol. 7. doi: 10.3389/fmicb.2016.00214
Guerra, C. A., Bardgett, R. D., Caon, L., Crowther, T. W., Delgado-Baquerizo, M., Montanarella, L., et al. (2021). Tracking, targeting, and conserving soil biodiversity. Science 371, 239–241. doi: 10.1126/science.abd7926
Guerra, C. A., Heintz-Buschart, A., Sikorski, J., Chatzinotas, A., Guerrero-Ramírez, N., Cesarz, S., et al. (2020). Blind spots in global soil biodiversity and ecosystem function research. Nat. Commun. 11, 3870. doi: 10.1038/s41467-020-17688-2
Hector, A., Bagchi, R. (2007). Biodiversity and ecosystem multifunctionality. Nature 448, 188–190. doi: 10.1038/nature05947
Moura, J. B., Souza, R. F., Vieira Júnior, W. G., Lucas, L. S., Santos, J. M., Dutra e Silva, S., et al. (2022). Effects of a megafire in the arbuscular mycorrhizal fungal community and parameters in the Brazilian Cerrado ecosystem. For. Syst. 31, e001. doi: 10.5424/fs/2022311-18557
Romero, F., Hilfiker, S., Edlinger, A., Held, A., Hartman, K., Labouyrie, M., et al. (2023). Soil microbial biodiversity promotes crop productivity and agro-ecosystem functioning in experimental microcosms. Sci. Total. Environ. 885, 163683. doi: 10.1016/j.scitotenv.2023.163683
Van der Putten, W. H., Bardgett, R. D., Bever, J. D., Bezemer, T. M., Casper, B. B., Fukami, T., et al. (2013). Plant–soil feedbacks: the past, the present and future challenges. J. Ecol. 101, 265–276. doi: 10.1111/1365-2745.12054
Vasar, M., Davison, J., Sepp, S. K., Mucina, L., Oja, J., Al-Quraishy, S., et al. (2022). Global soil microbiomes: A new frontline of biome-ecology research. Glob. Ecol. Biogeogr. 31, 1120–1132. doi: 10.1111/geb.13487
Keywords: rhizosphere, soil ecology and microbiome, multifunctionality, mycorrhiza, ecosystem function, ecosystem services, biodiversity
Citation: Marín C, Bueno CG, Wang J and Kokkoris V (2023) Editorial: Biodiversity and ecosystem-level function of the rhizosphere. Front. Plant Sci. 14:1278662. doi: 10.3389/fpls.2023.1278662
Received: 16 August 2023; Accepted: 01 September 2023;
Published: 08 September 2023.
Edited and Reviewed by:
Shrivardhan Dheeman, MVN University, IndiaCopyright © 2023 Marín, Bueno, Wang and Kokkoris. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: César Marín, Y21hcmluZEBzYW50b3RvbWFzLmNs