
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
OPINION article
Front. Plant Sci. , 28 April 2023
Sec. Functional Plant Ecology
Volume 14 - 2023 | https://doi.org/10.3389/fpls.2023.1135549
This article is part of the Research Topic Biodiversity and Ecosystem-level Function of the Rhizosphere View all 9 articles
 Jianqing Wang1,2
Jianqing Wang1,2 Xiuzhen Shi1,2*
Xiuzhen Shi1,2*The Intergovernmental Panel on Climate Change (IPCC) has reported that global surface temperature will continue to rise until the middle of the century, and global warming of 1.5°C and 2°C will be exceeded during the century (IPCC, 2021). It is widely accepted that global warming will cause a mass extinction in the coming years (Geisen et al., 2019; Turney et al., 2020; Wang et al., 2021), and biodiversity protection is increasingly important. Forests stand for one of the most crucial ecosystems on Earth and reserve a large proportion of the global biodiversity (Lladó et al., 2017). Unraveling the changes in biodiversity in forest ecosystems under global warming is a challenging field of research in theoretical ecology that remains under-explored in forest ecosystems.
Forest soil is an important living repository of biodiversity, with interactions between microorganisms, fauna, and plants providing multiple functioning and services to human health (Banerjee & van der Heijden, 2023). Soil biodiversity played a vital role in the forest ecosystem, owing to its inherent complexity (the traits, sizes, functions, and types of soil organisms), directly or indirectly underpinning multiple functioning and services, such as nutrient cycling, carbon sequestration, vegetation health, plant growth as well as soil stability (Delgado-Baquerizo et al., 2020b; Shi et al., 2021; Wang et al., 2022a; Wang et al., 2023b). Furthermore, soil biodiversity is increasingly acknowledged as beneficial to human well-being, as a consequence of reducing disease-causing soil microbes and also improving the quality and quantity of water, air, and food (Wall et al., 2015). Yet, forest soil biodiversity is frequently inadvertently altered by human-induced global warming.
Forests distributed in the tropical and subtropical regions are highly concentrated habitats of the Earth’s terrestrial biodiversity (Alroy, 2017). However, subtropical and tropical forests are assumed to be more vulnerable to global warming than temperate forests due to the relatively narrow upper thermal limits and temperature variation in subtropical and tropical regions (Sentinella et al., 2020). To combat global warming, many countries have committed to restoring forest areas (Bastin et al., 2019; Lewis et al., 2019). Almost half of the global forest area is set to become plantations of commercial trees (Lewis et al., 2019). However, several studies have pointed out that the adaptability of soil biodiversity in plantations to global warming might be lower than that of natural forests (Gibson et al., 2011; Lewis et al., 2019). Here, we provide several aspects of consideration that natural forests may exhibit higher soil biodiversity resistance than planted forests with respect to global warming, especially in subtropical forests.
Natural forests play a pivotal role in conserving soil biodiversity and maintaining multiple ecosystem functions and services (Edwards et al., 2014). However, two-thirds of the land area has been set aside for reforestation worldwide (Lewis et al., 2019). Although plantations can contribute to mitigating some of the detrimental impacts of deforestation on soil biodiversity, the resulting soil quality and functions are declining compared to natural forests (Veldkamp et al., 2020). Indeed, our previous study found that long-term Cunninghamia lanceolate plantation greatly increased the abundance of plant parasite nematodes, consequently threatening soil and plant health in subtropical China (Zheng et al., 2022). Delgado-Baquerizo et al. (2020a) pointed out that the relative abundance of soil-borne fungal plant pathogens increased with warmer temperatures based on a global field survey and a nine-year field experiment. And as a consequence, the impacts of natural forests and planted forests on soil biodiversity raise serious concerns under global warming.
First of all, compared to scarce human disturbance, planted forest development is frequently subjected to regular harvesting and clearing of plantations (Poorter et al., 2016). Once trees were harvested, the land is cleared for massive planting of saplings, and chemical fertilizers and pesticides are normally applied to facilitate the rapid accumulation of standing tree biomass. These plantation management practices possibly lead to forest land degradation, further decreasing soil biodiversity and consequent ecosystem multiple functions in subtropical forests (Wang et al., 2022a). By contrast, the natural forest possibly increases soil biodiversity by protecting land from fire and human disturbances and thus improving multiple ecosystem functions and services in the subtropical forest (Shi et al., 2021).
Secondly, planted forests intrinsically comprise substantially lower tree diversity than native forests. However, to alleviate global warming, ongoing reforestation efforts accelerate the current loss of biodiversity (Carnus et al., 2006). We previously found that soil nematode abundance significantly increased with forest restoration and improved soil health status in natural forests rather than in planted forests in subtropical regions (Wang et al., 2022b; Zheng et al., 2022). This is mostly attributed to the fact that species-rich ecosystems can enhance soil microbial growth and biomass by providing a higher amount of plant-derived resources (e.g., litter inputs and root exudates) (Figure 1) (Prommer et al., 2020). It is commonly established that species-rich natural forests better support biodiversity than planted forests in the tropical zone (Gibson et al., 2011).
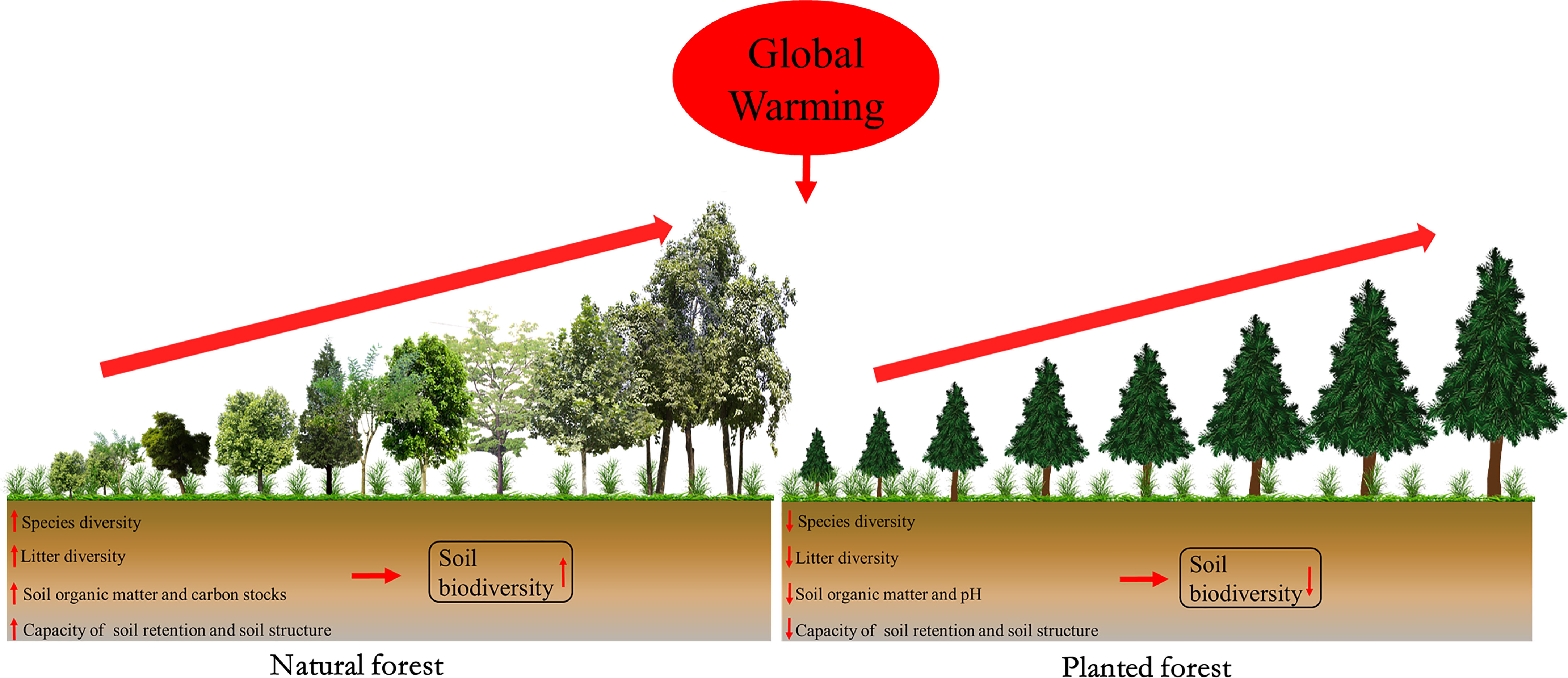
Figure 1 Conceptual graph of the changes in ecosystem processes and variables with forest restoration between natural and planted forests in the subtropical zone under global warming. The red upward arrow represents a positive effect; the red downward arrow indicates a negative effect.
Thirdly, natural forests exhibit higher water use efficiency and soil organic matter storage compared to planted forests (Yu et al., 2019). The conversion of natural forests to planted forests caused substantial declines in soil organic carbon retention by around 60% (Zarafshar et al., 2020). Furthermore, carbon capture capacity in planted forests was less stable than in natural forests, particularly in the face of increasing global warming in the tropical zone (Figure 1) (Osuri et al., 2020). Ghosh et al. (2020) also highlighted that the quality and temperature sensitivity (Q10) of soil organic matter were higher in the natural ecosystem than in the managed ecosystems in North Eastern India, indicating protecting natural ecosystems is very important to mitigate climate warming. Amoo et al. (2021) revealed that planted forests consistently decreased multiple soil functions linked to soil carbon cycling and nutrient dynamics, and altered the functional profile and activity of soil microbial communities in South Africa. The planted forest can therefore have lasting detrimental impacts on soil biodiversity and health, in addition to posing a significant threat to terrestrial ecosystem functions in subtropical forests (Carnus et al., 2006; Wang et al., 2022a; Zheng et al., 2022). Therefore, we expect greater soil biodiversity in the species-rich natural forests than that in the species-poor plantations, particularly in subtropical forests.
Therefore, it is generally believed that natural forests can assist to relieve the negative impact of global warming by conserving biodiversity, as well as supporting multiple ecosystem functions and services (Alroy, 2017). In the future, forest natural restoration rather than planted forests would continue to be one of the most effective ways to combat global warming in subtropical forests.
Comparatively little is known about the responses of soil biodiversity, with most climate warming experiments concentrating on the aboveground ecosystems (Fierer et al., 2009). Soil organisms are an extremely diversified assemblage of organisms, which involves soil microorganisms (i.e., archaea, bacteria, and fungi), and soil fauna (i.e., protozoa, nematode) (Orgiazzi et al., 2016; Van Den Hoogen et al., 2019; Aslani et al., 2022). As important components of forest ecosystems, soil organisms undertake a wide range of ecosystem functions and services (Shi et al., 2021; Wang et al., 2023a), such as mediating biogeochemical cycling and ecosystem health maintenance, their responses to global warming are potentially important in subtropical forest ecosystems. However, the impacts of global warming on soil organisms and biodiversity are rather equivocal to date. Therefore, the authors encourage filling this knowledge gap from the perspective of the soil food web to thoroughly understand soil biodiversity, including vegetation resources, soil microorganisms, and soil fauna.
This is a research area receiving little attention in forest ecosystems but could potentially have a tremendous impact on combating global warming in the future. However, disentangling soil food webs and biodiversity in the forest soil may be a daunting task, because of the enormous phylogenetic variety in the soil (Kardol et al., 2016). Traditionally, soil web food studies have used identification procedures based strictly on morphological traits. This approach requires a high degree of taxonomic competence, which usually restricts the number of species that can be investigated concurrently (Oliverio et al., 2018). Furthermore, the procedures of morphological identification are time-consuming, and intensive efforts are required from taxonomists aiming to recognize biodiversity at lower taxonomic levels (Madden et al., 2016). In recent years, molecular analysis tools for soil food web and biodiversity are now widely used in soil ecology, and molecular methods have greatly improved our knowledge of soil biodiversity (Thomsen & Willerslev, 2015). Molecular ecological approaches, such as metagenome, 13C isotope identification, and network analysis, have been increasingly applied for the characterization of the soil food web and biodiversity (Shi et al., 2018; Chomel et al., 2022).
Overall, to mitigate the adverse impacts of global warming on soil biodiversity, we urge the restoration community, forestry specialists, and legislators to emphasize natural forest regeneration over various tree-planting approaches, thus maintaining soil biodiversity and improving multiple ecosystem functions. Additionally, we call for new theories and technologies to preserve soil biodiversity based on the soil food web to combat global warming in the future. This perspective will emphasize the necessity of strengthening natural regeneration rather than planted forests for maintaining soil biodiversity and ecosystem functioning.
JW contributed ideas to the study and drafted the manuscript. XS contributed ideas to the study and improved the draft. All authors contributed to the article and approved the submitted version.
JW was financially supported by the National Natural Science Foundation of China (32271679) and the Natural Science Foundation of Fujian Province (2020J01186). XS was financially supported by the National Natural Science Foundation of China (32071631) and the Natural Science Foundation of Fujian Province (2020J01138).
We are grateful to editor Prof. Avijit Ghosh for the time spent handling our manuscript. We would like to thank you for the constructive comments and suggestions from the editor and independent reviewers that have helped us greatly improve the quality of this manuscript.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Alroy, J. (2017). Effects of habitat disturbance on tropical forest biodiversity. Proc. Natl. Acad. Sci. 114, 6056–6061. doi: 10.1073/pnas.1611855114
Amoo, A. E., Delgado-Baquerizo, M., Babalola, O. O. (2021). Forest plantations reduce soil functioning in terrestrial ecosystems from South Africa. Pedobiologia 89, 150757. doi: 10.1016/j.pedobi.2021.150757
Aslani, F., Geisen, S., Ning, D., Tedersoo, L., Bahram, M. (2022). Towards revealing the global diversity and community assembly of soil eukaryotes. Ecol. Lett. 25, 65–76. doi: 10.1111/ele.13904
Banerjee, S., van der Heijden, M. G. (2023). Soil microbiomes and one health. Nat. Rev. Microbiol. 21 (1), 6–20. doi: 10.1038/s41579-022-00779-w
Bastin, J.-F., Finegold, Y., Garcia, C., Mollicone, D., Rezende, M., Routh, D., et al. (2019). The global tree restoration potential. Science 365, 76–79. doi: 10.1126/science.aax0848
Carnus, J.-M., Parrotta, J., Brockerhoff, E., Arbez, M., Jactel, H., Kremer, A., et al. (2006). Planted forests and biodiversity. J. Forestry 104, 65–77. doi: 10.1093/jof/104.2.65
Chomel, M., Lavallee, J. M., Alvarez-Segura, N., Baggs, E. M., Caruso, T., De Castro, F., et al. (2022). Intensive grassland management disrupts below-ground multi-trophic resource transfer in response to drought. Nat. Commun. 13, 6991.
Delgado-Baquerizo, M., Guerra, C. A., Cano-Díaz, C., Egidi, E., Wang, J.-T., Eisenhauer, N., et al. (2020a). The proportion of soil-borne pathogens increases with warming at the global scale. Nat. Climate Change 10, 550–554. doi: 10.1038/s41558-020-0759-3
Delgado-Baquerizo, M., Reich, P. B., Trivedi, C., Eldridge, D. J., Abades, S., Alfaro, F. D., et al. (2020b). Multiple elements of soil biodiversity drive ecosystem functions across biomes. Nat. Ecol. Evol. 4, 210–220. doi: 10.1038/s41559-019-1084-y
Edwards, D. P., Tobias, J. A., Sheil, D., Meijaard, E., Laurance, W. F. (2014). Maintaining ecosystem function and services in logged tropical forests. Trends Ecol. Evol. 29, 511–520. doi: 10.1016/j.tree.2014.07.003
Fierer, N., Strickland, M. S., Liptzin, D., Bradford, M. A., Cleveland, C. C. (2009). Global patterns in belowground communities. Ecol. Lett. 12, 1238–1249. doi: 10.1111/j.1461-0248.2009.01360.x
Geisen, S., Wall, D. H., van der Putten, W. H. (2019). Challenges and opportunities for soil biodiversity in the anthropocene. Curr. Biol. 29, R1036–R1044. doi: 10.1016/j.cub.2019.08.007
Ghosh, A., Das, A., Das, D., Ray, P., Bhattacharyya, R., Biswas, D. R., et al. (2020). Contrasting land use systems and soil organic matter quality and temperature sensitivity in north Eastern India. Soil Tillage Res. 199, 104573. doi: 10.1016/j.still.2020.104573
Gibson, L., Lee, T. M., Koh, L. P., Brook, B. W., Gardner, T. A., Barlow, J., et al. (2011). Primary forests are irreplaceable for sustaining tropical biodiversity. Nature 478, 378–381. doi: 10.1038/nature10425
IPCC (2021). “Climate change 2021: The physical science basis,” in Contribution of working group I to the sixth assessment report of the intergovernmental panel on climate change. Eds. Masson-Delmotte, V., Zhai, P., Pirani, A., Connors, S. L., Péan, C., Berger, S., Caud, N., Chen, Y., Goldfarb, L., Gomis, M. I., Huang, M., Leitzell, K., Lonnoy, E., Matthews, J. B. R., Maycock, T. K., Waterfield, T., Yelekçi, O., Yu, R., Zhou, X. X. X. B. (Cambridge University Press. In Press).
Kardol, P., Throop, H. L., Adkins, J., de Graaff, M.-A. (2016). A hierarchical framework for studying the role of biodiversity in soil food web processes and ecosystem services. Soil Biol. Biochem. 102, 33–36. doi: 10.1016/j.soilbio.2016.05.002
Lewis, S. L., Wheeler, C. E., Mitchard, E. T., Koch, A. (2019). Regenerate natural forests to store carbon. Nature 568, 25–28. doi: 10.1038/d41586-019-01026-8
Lladó, S., López-Mondéjar, R., Baldrian, P. (2017). Forest soil bacteria: diversity, involvement in ecosystem processes, and response to global change. Microbiol. Mol. Biol. Rev. 81, e00063–e00016. doi: 10.1128/MMBR.00063-16
Madden, A. A., Barberán, A., Bertone, M. A., Menninger, H. L., Dunn, R. R., Fierer, N. (2016). The diversity of arthropods in homes across the united states as determined by environmental DNA analyses. Mol. Ecol. 25, 6214–6224. doi: 10.1111/mec.13900
Oliverio, A. M., Gan, H., Wickings, K., Fierer, N. (2018). A DNA metabarcoding approach to characterize soil arthropod communities. Soil Biol. Biochem. 125, 37–43. doi: 10.1016/j.soilbio.2018.06.026
Orgiazzi, A., Bardgett R, D., Barrios, E., Behan-Pelletier, V., Briones, M., Chotte, J.-L., et al. (2016). Global soil biodiversity atlas (European Commission). Available at: https://esdac.jrc.ec.europa.eu/content/global-soil-biodiversity-atlas.
Osuri, A. M., Gopal, A., Raman, T. R. S., DeFries, R., Cook-Patton, S. C., Naeem, S. (2020). Greater stability of carbon capture in species-rich natural forests compared to species-poor plantations. Environ. Res. Lett. 15 (3), 034011. doi: 10.1088/1748-9326/ab5f75
Poorter, L., Bongers, F., Aide, T. M., Almeyda Zambrano, A. M., Balvanera, P., Becknell, J. M., et al. (2016). Biomass resilience of Neotropical secondary forests. Nature 530, 211–214. doi: 10.1038/nature16512
Prommer, J., Walker, T. W., Wanek, W., Braun, J., Zezula, D., Hu, Y., et al. (2020). Increased microbial growth, biomass, and turnover drive soil organic carbon accumulation at higher plant diversity. Global Change Biol. 26, 669–681. doi: 10.1111/gcb.14777
Sentinella, A. T., Warton, D. I., Sherwin, W. B., Offord, C. A., Moles, A. T. (2020). Tropical plants do not have narrower temperature tolerances, but are more at risk from warming because they are close to their upper thermal limits. Global Ecol. Biogeography 29 (8), 1387–1398. doi: 10.1111/geb.13117
Shi, X., Hu, H.-W., Wang, J., He, J.-Z., Zheng, C., Wan, X., et al. (2018). Niche separation of comammox nitrospira and canonical ammonia oxidizers in an acidic subtropical forest soil under long-term nitrogen deposition. Soil Biol. Biochem. 126, 114–122. doi: 10.1016/j.soilbio.2018.09.004
Shi, X., Wang, J., Lucas-Borja, M. E., Wang, Z., Li, X., Huang, Z. (2021). Microbial diversity regulates ecosystem multifunctionality during natural secondary succession. J. Appl. Ecol. 58 (12), 2833–2842. doi: 10.1111/1365-2664.14015
Thomsen, P. F., Willerslev, E. (2015). Environmental DNA–an emerging tool in conservation for monitoring past and present biodiversity. Biol. Conserv. 183, 4–18. doi: 10.1016/j.biocon.2014.11.019
Turney, C., Ausseil, A. G., Broadhurst, L. (2020). Urgent need for an integrated policy framework for biodiversity loss and climate change. Nat. Ecol. Evol. l4, 996. doi: 10.1038/s41559-020-1242-2
Van Den Hoogen, J., Geisen, S., Routh, D., Ferris, H., Traunspurger, W., Wardle, D. A., et al. (2019). Soil nematode abundance and functional group composition at a global scale. Nature 572, 194–198. doi: 10.1038/s41586-019-1418-6
Veldkamp, E., Schmidt, M., Powers, J. S., Corre, M. D. (2020). Deforestation and reforestation impacts on soils in the tropics. Nat. Rev. Earth Environ. 1, 590–605. doi: 10.1038/s43017-020-0091-5
Wall, D. H., Nielsen, U. N., Six, J. (2015). Soil biodiversity and human health. Nature 528, 69–76. doi: 10.1038/nature15744
Wang, J., Shi, X., Li, L., Zhang, X. (2021). Changes in soil nematodes in rhizosphere and non-rhizosphere soils following combined elevated [CO2] and canopy warming in a winter wheat field. Geoderma 386, 114907. doi: 10.1016/j.geoderma.2020.114907
Wang, J., Shi, X., Lucas-Borja, M. E., Guo, Q., Mao, J., Tan, Y., et al. (2023a). Soil nematode abundances drive agroecosystem multifunctionality under short-term elevated CO2 and O3. Global Change Biol. 29 (6), 1618–1627. doi: 10.1111/gcb.16546
Wang, J., Shi, X., Lucas-Borja, M. E., Lam, S. K., Wang, Z., Huang, Z. (2022a). Plants, soil properties and microbes directly and positively drive ecosystem multifunctionality in a plantation chronosequence. Land Degradation Dev. 33 (16), 3049–3057. doi: 10.1002/ldr.4371
Wang, J., Zheng, C., Lucas-Borja, M. E., Shi, X. (2023b). Soil protist functional composition shifts with atmospheric nitrogen deposition in subtropical forests. J. Appl. Ecol., 1–9. doi: 10.1111/1365-2664.14392
Wang, J., Zheng, Y., Shi, X., Lam, S. K., Lucas-Borja, M. E., Huang, Z. (2022b). Nature restoration shifts the abundance and structure of soil nematode communities in subtropical forests. Plant Soil 471, 315–327. doi: 10.1007/s11104-021-05229-9
Yu, Z., Liu, S., Wang, J., Wei, X., Schuler, J., Sun, P., et al. (2019). Natural forests exhibit higher carbon sequestration and lower water consumption than planted forests in China. Global Change Biol. 25, 68–77. doi: 10.1111/gcb.14484
Zarafshar, M., Bazot, S., Matinizadeh, M., Bordbar, S. K., Rousta, M. J., Kooch, Y., et al. (2020). Do tree plantations or cultivated fields have the same ability to maintain soil quality as natural forests? Appl. Soil Ecol. 151, 103536. doi: 10.1016/j.apsoil.2020.103536
Keywords: climate change, forest management, global warming, soil biodiversity, soil microbes, subtropical forest, forest conservation
Citation: Wang J and Shi X (2023) Soil biodiversity in natural forests potentially exhibits higher resistance than planted forests under global warming. Front. Plant Sci. 14:1135549. doi: 10.3389/fpls.2023.1135549
Received: 01 January 2023; Accepted: 28 March 2023;
Published: 28 April 2023.
Edited by:
Avijit Ghosh, Indian Grassland and Fodder Research Institute (ICAR), IndiaReviewed by:
Javid Ahmad Dar, SRM University, IndiaCopyright © 2023 Wang and Shi. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Xiuzhen Shi, c2h4emg4N0Bob3RtYWlsLmNvbQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.