 Jun Chen*
Jun Chen* Xin Pang
Xin Pang- Faculty of Horticulture Science & Technology, Suzhou Polytechnic Institute of Agriculture, Suzhou, China
Globally, abiotic stresses are drastically reducing the productivity of vegetable crops. Among abiotic stresses, drought and salinity are more challenging constraints for the sustainable production of vegetables. A great variety of vegetables are facing dry and hot summer spells, poor water availability, and higher salinity mainly due to irrigation with brackish water. Vegetables are considered higher water-dependent crops, requiring water for proper growth and yield. Drought and salinity impair plant metabolism. The disruption in plant metabolism leads to a reduction in growth, developmental processes, and ultimately crop yield. Appropriate management measures are needed to cope with the adverse effects of drought and salinity. Different agronomic and molecular approaches contributed to improving tolerance. Therefore, the present review significantly explores the impact of phytohormones on vegetable crops under drought and salinity stresses. Phytohormones (salicylic acid, melatonin, jasmonates, Brassinosteroids, ascorbic acid, and numerous others) can be sprayed for improvement of plant growth, yield, and photosynthetic pigments by modulation of physiological and biochemical processes. In this manner, these phytohormones should be explored for sustainable production of vegetable crops growing under abiotic stress conditions.
Introduction
Phytohormones are considered plant-protecting hormones under abiotic stress conditions. Different phytohormones are well-known management strategies, which act as stress-relieving bioactive compounds in vegetable crops (Fahad et al., 2015; Altaf et al., 2022a). The exogenous spray of numerous phytohormones can reduce the drought and salinity stresses and also improve the plant defense mechanism focusing on sustainable production. Fascinatingly, phytohormones are more effective for the reduction of challenges that occur from stressful conditions at any growth or developmental phases even from germination to plant senescence. These hormones are contributing to numerous signaling and transduction pathways through hormonal reception and regulatory actions (Hu et al., 2012). The membrane receptors, ionic networks, reactive oxygen species (ROS) indications, and mitogen-activated protein kinase (MAPK) indications are noticeable to numerous fundamental utilities in the synchrony by phytohormones to cope with the negative effects of abiotic stress. Understanding the interactive mechanism of phytohormones and transcriptomics can be effective for the development of tolerant germplasm of vegetables (Diao et al., 2015; Mangal et al., 2022). Modulation of physiological and photosynthetic pigments was found to be helpful for an increase in plant yield growing under water stress and saline conditions. The susceptible germplasm can also become higher yielding by sufficient use of phytohormones based on genetic makeup and climatic conditions of the characterized germplasm, as reported by Forni et al. (2017). However, the impact of phytohormones on the regulation of secondary metabolites and other signaling molecules needs further investigation for better understanding. For the development of tolerant germplasm, traditional breeding ways are time-consuming and not specific. However, the application of phytohormones is more effective for the alleviation of abiotic stress tolerance.
Plants growing under field conditions could be exposed to multiple stresses, which can damage the crops’ yield (Glick, 2012). Severe climatic conditions in summer, irregular nutrition management, and unavailability of irrigation water are causing stunted growth and poor crop yield. Sustainable agricultural crop production is drastically affected by numerous biotic stresses (such as insects, pests, and disease) and abiotic stresses (like drought, salinity, temperature extremes, humidity, light, ultraviolet radiations, mineral nutrition deficiencies, and heavy metals) (Akram et al., 2017; Shakoor et al., 2017). Drought and salinity are considered more destructive conditions, extensively affecting growth, developmental stages, and yield. Plants can change their defense system against stressful conditions to regulate metabolism, growth, and development (Ahmad et al., 2008). Vegetable crops are potentially growing under diverse environmental conditions by natural acclimation, as well as numerous adaptation strategies. However, these approaches may not be sufficient to reduce losses from variations in climate change (Shahid et al., 2021; Zhang et al., 2022a). The severity of abiotic stress is mainly based on the type of species and intensity and duration of stress (Zhang et al., 2022b). Stressful conditions cause variations in plant physiological and biochemical processes, either reversible or irreversible. However, these constraints affect vegetable crops primarily, which are susceptible to abiotic stress (Parveen et al., 2020). Presently, vegetable crop demand is higher; therefore, it is necessary to develop some excellent approaches or tolerant germplasm to tackle the severity of drought and salinity stresses. Drought and salinity stresses are critical global concerns and harm the sustainable production of crops. Irrigation water resources are depleting due to climate change, urbanization, and industrialization (Gruda et al., 2019). Soil salinity is also increasing, mainly due to irrigation with poor-quality and brackish water. The unavailability of quality water in various regions is causing salt accumulation in the soil, which further translocates toward the root zone of vegetable crops. It has been estimated that approximately 20% of global land is negatively affected by salt extremes (Forni et al., 2017).
Vegetables are considered an essential source of the human diet because they are rich in dietary fibers, vitamins, antioxidants, and minerals. Their consumption is also due to good taste, excellent texture, and religious value (Gamalero and Glick, 2022). Global vegetable production in 2020 increased by nearly 66%, from 447 to 1,130 Mt (FAO, 2021). Farmers are investing considerable efforts in improving vegetable production and nutritional aspects under stressful environments (Gruda et al., 2019). The severity of drought and salinity is mainly based on different climatic constraints like the distribution of solar radiation, the need for evapotranspiration, and the retention of soil moisture content (Sabir et al., 2022). Hence, numerous agricultural practices and breeding approaches can be employed for the alleviation of tolerance in vegetable crops against drought and salinity.
Plant researchers urge sustainable management practices to increase vegetable production under drought and salinity stresses (Ahmad et al., 2010; Checker et al., 2018). The exogenous application of phytohormones is a more promising approach to cope with the adverse effects of drought and salinity for sustainable vegetable production. The involvement of phytohormones is attracting much attention from plant researchers due to their multifunctioning behavior against drought and salinity stresses. However, their utilization is still limited in vegetable crops growing under drought and salinity. Therefore, the present study elaborates on the utilization of phytohormones in vegetable crops under drought and salinity stresses. Deep insights into physiological, biochemical, and molecular basis were also explored in the vegetables to cope with the adverse effects of drought and salinity.
Phytohormones are major modulators of plant responses to drought and salinity
Vegetable production is low in different growing areas due to water deficit and salinity. Restricted growth and low yield are due to the unavailability and shortage of water and excessive salt accumulation in the root zone of plants. The higher uptake of Na+ through roots by xylem vessels resulted in restriction in the uptake of nutrients and minerals necessary for sufficient growth and yield (Maksimovic and Ilin, 2012). Salinity, sodicity, and water stress revealed adverse effects on the growth, yield, and quality of vegetable crops. Higher accumulation of salts disturbed the soil structure, texture, porosity, and permeability of water, which ultimately reduces the productivity of vegetable crops (Malhi et al., 2021). Soil provides better anchor and acts as a reservoir of mineral nutrients necessary for better growth, development, and yield. Therefore, the development of mechanistic approaches is needed to minimize the damaging effects of drought and salinity in vegetable crops. Drought and salinity affect vegetable crops, causing restriction in growth with poor yield (Hossain et al., 2022). Drought stress and excessive Na+ accumulation are causing a disturbance in the metabolism of vegetable crops. Moreover, the osmotic potential of plants is also adversely affected due to drought and excessive salt accumulation in different plant cells and compartments (Zaidi et al., 2015). Alterations in metabolism and disturbances in osmotic potential are the leading causes of restricted growth and low yield and sometimes complete or partial death of a plant (Neha et al., 2021).
In addition to supporting signaling pathways, endogenous plant hormones are critical in the response to drought and salinity. Phytohormones play a major role in mediating how plants respond to osmotic adjustment under stress conditions. Small signaling molecules called phytohormones have a significant impact on almost every aspect of the development of plants. The methods of action taken by different hormones for various activities may be very different. Furthermore, it is well recognized that even a single hormone can have an impact on a wide range of cellular and developmental processes or that multiple hormones can regulate a single function concurrently. Phytohormones protect and control plants from biotic and abiotic stresses. As a result, phytohormone application aims to expand crop stress research in the future (Ahmad et al., 2008).
General signs and ion toxicity under drought and salinity stresses
Drought and salinity stress decrease the uptake of Ca2+ and K+ in vegetable crops, which is the primary reason why nutritional imbalances occur in plants. Plant physiology and morphology are also affected by numerous stresses and thus are susceptible to drought and salinity stresses (Rodrıguez et al., 2005). The initial response of vegetable plants under drought and salinity is the dropping of leaves or the initiation of leaf senescence. After that, a reduction in fresh and dry weights may also be considered an early response of plants growing under water shortage and salinity stress conditions (Zhu, 2002). The decline in fresh and dry weights ultimately reduces the plant yield. Yield reduction is evident in vegetable crops growing under drought and salinity stresses (Alian et al., 2000). However, a reduction in yield can also be a responsive mechanism, especially in aerial plant parts (Sharma et al., 2011) (Figure 1).
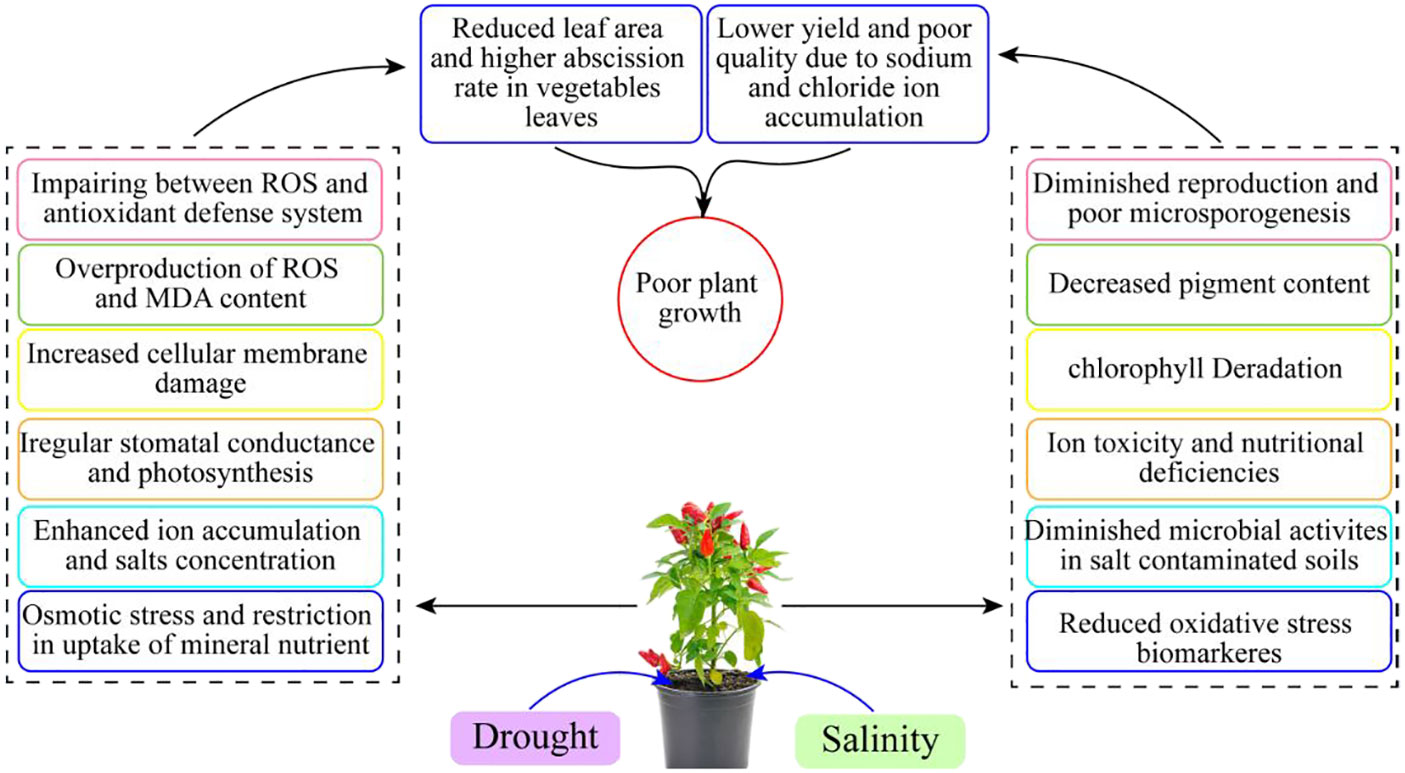
Figure 1 Adverse effects of salinity in vegetable crops.
Plants have been categorized into two main groups, halophytes and glycophytes. It has been reported that halophytes are more tolerant than glycophytes (Zhang et al., 2017). The potential of halophytes was much imperative, and higher survival and reproduction rates were observed as compared to glycophytes due to improved root architecture, regulation in stomatal conductance, balanced nutrition, improved metabolism, and distinctive genetic makeup (Gao et al., 2018). Halophytes can tolerate approximately 200 mM of NaCl because, at this level, glycophytes cannot survive. Furthermore, the halophyte group constitutes a 1% proportion of global flora, and the individuals of this group were grown naturally (Patane et al., 2013). Leaf growth, especially leaf area, is also considered an initial response in stressful conditions within plant cells and compartments. Numerous other signs include leaf scorching from tip and margins, yellowing and bronzing, leaf dropping of leaves, dieback in twigs, necrosis, blackening, and burning (Bernstein et al., 2004).
Different ion movements continue within plant organelles and compartments under normal conditions. Higher regulation of cytosolic K+ and Na+ ratio was recorded in the vegetable crops grown under favorable conditions (Zhu and Gong, 2014). Under salinity and drought conditions, ion balances are disturbed, and abnormal movements of ions continue until the availability of favorable conditions. Water-deficit conditions increased the accumulation of salts in the root zone (Sattar et al., 2021). Excessive Na+ in the root zone and its translocation to other plant parts are also improved. Na+ and K+ channels are also present in the xylem vessels. The discrimination of both ions is necessary, although both are similar in power to hydrated ions, and their discrimination is difficult for plants. However, some transporters of ions with high-affinity potassium transporters (HKTs) are more effective for the discrimination and movement of ions through xylem vessels in all plant parts. Furthermore, some proteins, such as integrated membrane proteins, are also involved in the regulation of solute movements within plant cells and compartments (Ahmad et al., 2008). Moreover, these transporters and proteins are specific for ion regulation; for example, some are specific for the discrimination of Na+ and others for the discrimination of K+. Hence, it has been reported that regulation of Na+ and K+ is necessary for sufficient plant growth, development, and yield of potatoes (Kamran et al., 2021).
Recently, vegetable crops are facing numerous biotic and abiotic stresses; however, a single abiotic stress is also sufficient for the drastic reduction in crop yield. The water shortage and excessive salt concentrations show a direct effect on the reduction in vegetable crop yield (Lin et al., 2006). Any plant parts, even underground or aerial parts, can be damaged due to low soil moisture levels and excessive salts (Li et al., 2022). Under stressful conditions, vegetable plants and their response to drought and salinity are mainly based on the type of species, cultivars, and even landraces. It has been studied that Cl− ions are effective for the catabolism of numerous enzymatic and non-enzymatic activities, and these are also known as co-factors for the regulation of the photosynthesis process (Rodríguez-Delfín et al., 2011). The behavior of sensitive and tolerant germplasm of vegetable crops toward Cl− is more different. The excess of Cl− is toxic; however, Na+ is more toxic than Cl−. Numerous genes are also involved in regulating Cl− produced in plants. Aquaporin has also been involved in the characterization of numerous genes that contributed to the regulation of Cl− efflux, which has significant involvement in the sustainable production of crops.
Avoidance mechanism of vegetable crops against stressful conditions
Salt exclusion and excretion restrict the salt’s access to the xylem vessels of vegetable crops. The exclusion of salts like Na+ and Cl− via roots revealed that the storage of Na+ and Cl− in leaves is not at a toxic level (Andre et al., 2009). However, their increased concentration disturbed physiological mechanisms, further resulting in leaf drop (Sobhanian et al., 2011). Grafting will be successful in numerous vegetable families and species like Solanaceae and Cucurbitaceae. Rootstock and scion combination contributed to the avoidance of salt mechanism in vegetable crops (Colla et al., 2010). Excessive salts in the root zone further translocated toward other plant parts. However, salt translocation can be reduced and not transported toward leaves. The rootstock’s basal portion can absorb the salts (Giordano et al., 2021). Therefore, it has been distinguished that the rootstock and scion combination is most necessary for alleviating salinity in vegetable crops (Figure 2).
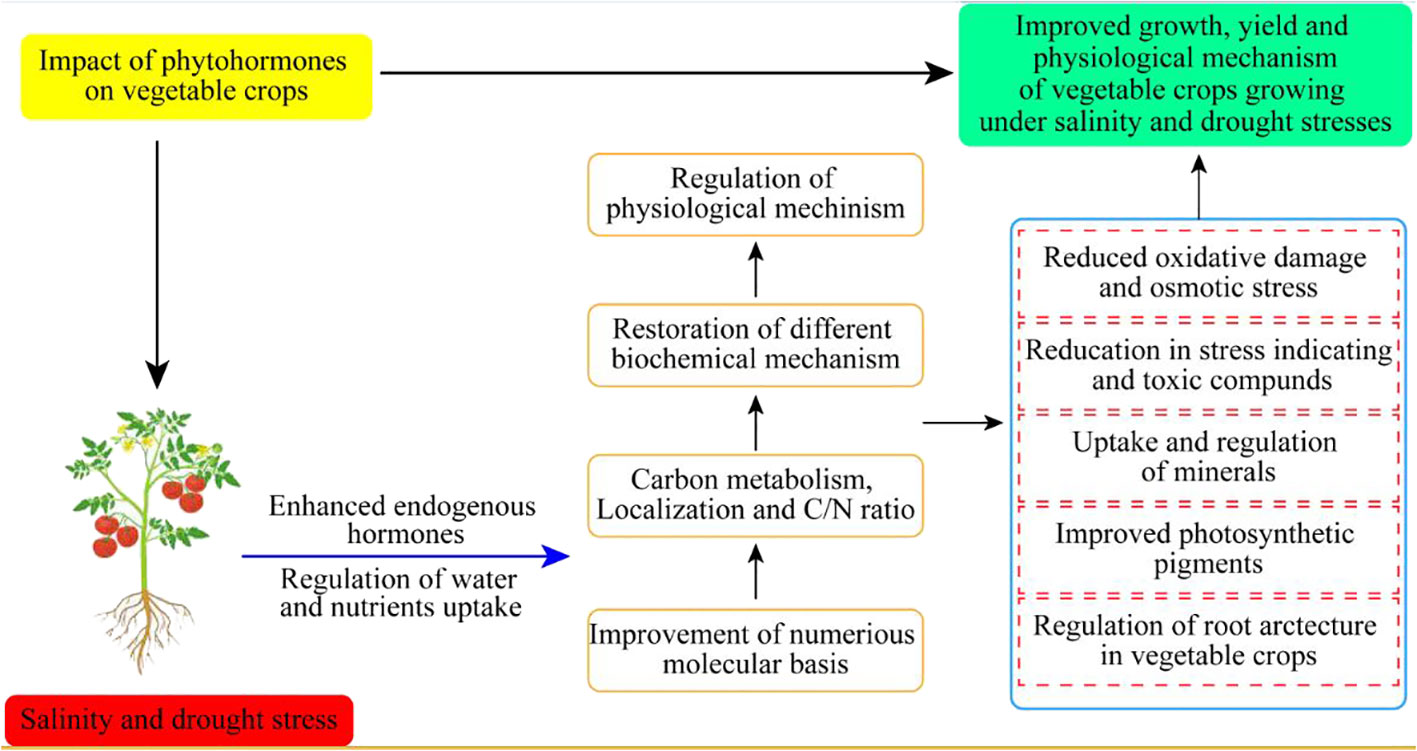
Figure 2 Critical stages of irrigation water at different growth stages of vegetable crops.
Salt exclusion is a variety-specific character in vegetable crops, and the higher exclusion of salts is the capability of a specific variety. However, this mechanism does not reveal the tolerance mechanism in vegetable crops. The avoidance mechanism of drought and salinity is also based on the root architecture of vegetable crops. Zhang et al. (2019) reported that grafting improves plant performance under drought and salinity stress in tomatoes. Similarly, phytohormones could improve the Solanaceae vegetable crop performance.
Phytohormones and gene expression under drought and salinity stresses
Expression of genes related to drought and salts is a more imperative utilization for the development of tolerant germplasm. Different genes and their expression in agronomic crops are widely discussed in the literature; however, in vegetable crops, this molecular phenomenon is still in progress. Most functional markers are related to numerous genes involved in the stress tolerance mechanism of vegetable crops (Ahmad et al., 2008). Gene expression potentially contributes to the development of salt-tolerant germplasm. Characterization of drought- and salt-tolerant and susceptible germplasm is a prerequisite for the sustainable production of vegetable crops (Malhi et al., 2021). Wild germplasm had more significant variation in genetic makeup and novel alleles, which can be explored to develop salt-tolerant germplasm. Numerous resistant genes can be identified, and further genome editing and transformation can be helpful for the development of tolerant germplasm in vegetable crops (Malhi et al., 2021). However, the expression of genes can be regulated by exogenous and endogenous improvements of phytohormones for the increase of tolerance against stress drought and salinity. Phytohormones are involved in the upregulation of transcriptomics of ATPase. Moreover, they were also involved in the reduction of the expression level of PpATG for the regulation of numerous morpho-physiological and biochemical activities in cucumbers. Moreover, similar findings were also reported by Parveen et al. (2021) for the expression of genes related to the tolerance mechanism.
Management approaches for mitigation of drought and salinity in vegetables
Different management approaches comprised of proteomics, marker-assisted selection, genome characterization, genome editing, genome mapping, quantitative trait locus (QTL) mapping, genomic editing, and genomic transformation are promising molecular bases for salinity and drought tolerance in vegetable crops (Saidi and Hajibarat, 2020). Furthermore, the molecular bases can be utilized for the backcrossing of genes present in wild species toward offspring or landraces. The first genome map was developed in the 1980s on potatoes in relation to sexual recombination regularities. Plant breeders have successfully characterized disease-tolerant genes in potatoes (Byun et al., 2007). Moreover, numerous economic traits were detected in potatoes. Furthermore, “NL25” is one of the functional markers with excellent capability to identify candidate genes related to tolerance characteristics of potato warts (Saidi and Hajibarat, 2020).
Phytohormones and vegetable crops under drought and salinity stresses
Phytohormones have the potential to enhance vegetables’ growth and development by interacting with numerous processes responsive to stressful conditions (Groppa and Benavides, 2008). Phytohormones have the capability to improve the defense system of vegetable crops growing under drought and salinity stresses. Plants activate their defense system against adverse climatic conditions for their survival (Choudhary et al., 2012). Therefore, supplementation of phytohormones boosts the immune system of plants growing under drought and salinity stresses. Plant defense system comprises the activation of enzymatic compounds (i.e., superoxide dismutase (SOD), peroxidase (POD), and catalases (CATs), non-enzymatic activities (i.e., ascorbic acid (AsA), phenolic content, and different sugars), osmolytes (i.e., glycine betaine (GB), ascorbate peroxidase (APX), and proline), and oxidative stress-indicating activities (ROS, malondialdehyde (MDA), and hydrogen peroxide (H2O2) (Table 1). Therefore, the impact of phytohormones on vegetable crops is imperative and needs more investigation on the molecular level to enhance plant tolerance (Diao et al., 2015).

Table 1 Role of different antioxidant activities in drought and salt tolerance mechanism of vegetable crops.
Brassinosteroids
These are more emerging, eco-friendly, and multifunctional plant hormones involved in regulating physiological mechanisms occurring within the plants (Mumtaz et al., 2022). Plant researchers and physiologists are working on utilizing these plant hormones for sustainable crop production. Alhaithloul et al. (2020) reported that Brassinosteroids (BRs) are more effective for plants growing under drought and salinity stress environments. It has been studied that BRs enhanced seed germination, root growth, seedling development, cell expansion and differentiation, ripening of fruits, leaf senescence, and reproduction of floral parts of vegetable crops (Bhandari and Nailwal, 2020). Moreover, in the findings of Kaya (2021), it has been discovered that BRs can improve growth traits, mineral content, antioxidant activities, and osmolytes and protect from membrane injury. Similarly, Shahid et al. (2011) evaluated that BRs elevated pea productivity against drought and salinity. Thus, it has been confirmed that BRs effectively elevate salinity tolerance in vegetable crops. BRs are effective for amelioration of drought and salinity tolerance in numerous vegetable crops like tomatoes (Jangid and Dwivedi, 2017), cucumber (Jakubowska and Janicka, 2017), and radish (Ramakrishna and Rao, 2015). Furthermore, elevated enzymatic activities like SOD, POD, CAT, and improved metabolites were recorded with exogenous application of these phytohormones. Moreover, improved physiological systems and reduction in oxidative injury were also observed in numerous vegetables, i.e., tomatoes (Jordan et al., 2020), peppers, and cucumbers (Per et al., 2017; Fahad et al., 2019), by application of BRs. Abiotic stress tolerance can be mitigated in the radish by supplemental use of BRs. Reduction in the over-generation of ROS, MDA, H2O2, and electrolyte leakage indicated that BRs are stress-relieving compounds for radish plants growing under stressful conditions as studied by Ramakrishna and Rao (2015). Furthermore, the increase in plant defense indicated that BRS is effective for the improvement of the plant immune system against harsh environments. In another study by Jakubowska and Janicka (2017), it has been indicated that BRs are much more effective for abiotic stress tolerance, as a similar tolerance mechanism was reported in the cucumbers. The exogenous spray of 24-EBRs on cucumbers improved the gaseous exchange processes and all its related traits, chlorophyll fluorescence, starch, soluble sugars, and rubisco activities. Therefore, it is much more effective for higher-yielding vegetable crops growing under normal and abiotic stress conditions. Similarly, in the other research by Choudhary et al. (2012), it has been studied that free radicle-scavenging potential in radishes was improved with enhanced antioxidant potential along with improvements in morphological traits of roots under heavy metal (copper) excess. From previous literature, it has been indicated that BRs are the more effective, eco-friendly, naturally occurring substances that might be extensively utilized for the reduction of drought and salinity stresses in vegetables.
Jasmonates
This group is comprised of methyl jasmonate (MeJA) and jasmonic acid (JA), which have been explored for their impacts on vegetable crops (Dar et al., 2015). Deprivation of photosynthetic pigments and tuber formation can be regulated under the exogenous application of JAs, as studied by Viswanath et al. (2020). The exogenous spray of this plant hormone improved sugar beet growth and defense system under drought (Ghaffari et al., 2019). Importantly, the exogenous JA application improved the endogenous production of JAs, and consequently, it can be used for hormonal regulation (Shahzad et al., 2015). MJ improved the drought resistance in cauliflower by improving oxidative bioactive compounds (Wu et al., 2012). Therefore, it has been exhibited that vegetable production can be increased with JA supplementation. JAs strengthen the defense system against environmental stresses in horticultural crops (Dar et al., 2015). These are significant for horticultural crops growing in areas with drought (Ge et al., 2010) and salinity (Pedranzani et al., 2003). Environmental threats can be regulated by the application of JAs. Similarly, Zou et al. (2017) revealed that the defense mechanism of plants was improved under environmental stresses like waterlogged conditions in peppers. JAs have good potential as a regulatory mechanism of vegetable crops against drought and salinity stresses. Abouelsaad and Renault (2018) reported that ROS mediation can be improved with JA because ROS is an indication of stress occurrence in tomato plants. Manan et al. (2016) reported that MeJA had the good capability to enhance the yield-related traits of tomato cultivars growing under elevated salinity as evaluated by Manan et al. (2016). The exogenous spray of MeJA on peas growing under stressful situations results in the improvement of indigenous hormonal levels of JA (Shahzad et al., 2015). Cauliflower grows under water-deficit conditions, facing challenges in growth at the seedling stage and poor yield at the reproductive stage. Wu et al. (2012) examined whether MeJA potentially triggered both oxidative and non-oxidative activities. Absorption and uptake of heavy metals were decreased in eggplant through the exogenous application of MeJA as supplementation (Yan et al., 2015). Seed priming is also an effective way to reduce challenges due to stress conditions. Therefore, it has been recorded that JA contributed to the increase in the germination of okra seeds, increase of seedlings, improved level of osmoprotectants, defense activities, photopigments, ROS reduction, lessening of H2O2, and low MDA level against salinity as studied by Iqbal et al. (2022). Exogenous application of MeJA on peppers improved osmolyte generation, oxidative and non-oxidative bioactive molecules, and metabolism and also improved the uptake of minerals via roots. Furthermore, decreases in MDA, H2O2, electrolyte leakage, and ROS were also reported by supplemental application of MeJA in the peppers. Therefore, it has been considered that JAs are suitable phytohormones for the mitigation of adverse effects of salinity and water-deficit conditions in horticultural crops.
Salicylic acid
This is a phenolic-based hormone that contributes to the elevated growth and yield of vegetable crops grown under drought and salinity environments, mainly by improving the plant defense system (Khan et al., 2015). Similarly, in another vegetable crop (pea), different concentrations of salicylic acid were applied exogenously (nearly 1–4 mM) under salinity conditions (50, 100, and 150 mM of NaCl) (Saidi and Hajibarat, 2020). In this study, it has been noted that salicylic acid improved pea growth, yield, enzymatic and non-enzymatic activities, and osmolytes. Salicylic acid (SA) (300 ppm) improved the mineral content in garlic and decreased Na+ uptake and translocation to other plant parts. Therefore, it has been considered that SA is helpful for vegetable crops growing under drought and salinity stresses (Shama et al., 2016). Similarly, in another study, nearly 0.11 mM of SA improved the tolerance of potatoes against abiotic stress (chilling). Priming seeds with salicylic acid at 100 mg/L is an effective strategy for the mitigation of adverse effects of salinity in cucumber (Rehman et al., 2011). The use of salicylic acid is an effective strategy for tolerance of abiotic stresses in vegetable crops, i.e., potatoes (Li et al., 2019), bell pepper (Zhang et al., 2020), spinach (Gilani et al., 2020), and peppermint (Ahmad et al., 2018). Spraying 1 mM of SA on tomatoes growing under heat stress resulted in an improved process of gaseous exchange, good water use potential, enzymatic activity generation, non-oxidative activation, and reduced oxidative stress conditions as studied by Zulfiqar et al. (2021). Moreover, biomass reduction on a fresh or dry basis was decreased, ultimately reducing the yield because of salinity and drought stresses. Furthermore, disruption in photopigments, photosynthesis disturbances, and irregularities in the functions of stomata are causes of osmotic stress. Therefore, it has been explored that regularities in the process of photosynthesis and stomatal function are important by application of different levels of SA. Similarly, in other findings, nearly 0.1 mM of SA enhanced the fresh and dried biomass, regulated photosynthesis, generation of oxidative and non-oxidative compounds, regulation in electrolyte leakage, protection from membrane injury, efficient water use potential, and excellent anatomical responses (Galviz et al., 2021). Moreover, it has been recorded that SA had the capability to mitigate challenges that occur from drought and salinity stresses by reduction of oxidative and osmotic injuries (Kaya, 2021).
Polyamines
The polyamine (PA) group from phytohormones primarily comprised spermidine, putrescine, and spermine having a lower molecular weight (Ahmad et al., 2012). Several physiological and biochemical processes were administered through polyamines by improving root, leaf differentiation, pollen viability, flower development, fruit growth, gene transcription, morphogenesis, embryo-genesis, leaf senesce, organogenesis, embryogenesis, and fruit maturation of the respective vegetable crop (Chen et al., 2019). Multiple abiotic stresses can be regulated by the application of varying concentrations of polyamines in horticultural crops, especially vegetable crops. Abiotic stresses can be regulated by the alteration of numerous processes of plants with a spray of polyamines available in the markets globally, as reported by Kamran et al. (2019). Moreover, the exogenous application of spermidine revealed good outcomes for tomato seedlings grown under stressful conditions. Moreover, the application of spermidine also enhanced the concentration of polyamine compounds within cells and compartments, especially in the root zone of tomato seedlings. The higher concentration of spermidine can be effective for tomato plants growing under saline conditions. The differentiation of ions and their translocation to other plant parts can be improved by supplementing polyamines (Hu et al., 2012). Exogenous application of spermidine is found to be effective for the improvement of plant growth, chlorophyll content, proline level, and different sugars, as reported by Zapata et al. (2004). Furthermore, it has also been reported that the reduction in ROS, MDA, and H2O2 was also measured in tomato plants. Pepper seeds were treated with different polyamines (spermine, putrescine, and spermidine), and it has been studied that the improved rate of germination, higher germination index, and early germination were recorded in treated seeds as compared to non-treated seeds. Similarly, in another study by Wu et al. (2018), it was revealed that the application of polyamines in cucumber seedlings improved crop performance under stressful conditions. Ormrod and Beckerson (1986) reported that polyamines are stress-relieving molecules as in the tomato for higher yield. The reduction and balance in the generation of oxidative stress markers, i.e., ROS, H2O2, MDA, free radicles, and movement of electrons, indicate the reduced stress in plants. Therefore, it has been studied that PA is an appropriate hormone for the improvement of endogenous hormones and also improved the activation of scavengers of toxic compounds.
Ascorbic acid
This contributed to the regulation of biosynthesis of ascorbates within the plant body. It is involved in the detoxification and compartmentation of H2O2 and MDA activities. Ascorbic acid is an important phytohormone necessary for sustainable vegetable production globally, grown under drought and salt stress conditions. The increased concentration of ascorbic acid on lettuce revealed that ascorbic acid is also effective for increasing the fresh and dry weights of lettuce and the number of leaves, which are considered yield-contributing factors against salinity. Seed germination is disturbed due to stressful conditions. Therefore, the exogenous application of AsA significantly enhanced seed germination with the endogenous improvement of ascorbates, which further activates the scavengers of toxic bioactive molecules within the plant cells. Hence, the initiation of seed germination is regulated with supplemental AsA as studied by Akram et al. (2017). Moreover, its application had a good role in the balance and neutralization of free radicals and toxic ROS generated within the plant cells. The exogenous application of ascorbic acid effectively improves the endogenous ascorbic acid content.
Abscisic acid
Its production is enhanced due to low moisture availability in the root zone of plants (Seiler et al., 2011; Lim et al., 2015). The enhanced production of abscisic acid (ABA) adversely affected plant growth and yield by producing nutritional imbalances (Sreenivasulu et al., 2012). The optimum production of ABA regulates the osmotic stress conditions (Ali et al., 2021a; Ali et al., 2021b). Induced levels of Na+ and ABA are the main causes of nutrient uptake restriction and nutrient translocation from roots toward leaves for food synthesis (Soma et al., 2021). Vegetables faced a reduction in stem and leaf, cell membrane injury, chlorophyll instability, lipid peroxidation, low water potential, degradation of photosynthetic pigments, poor gas exchange, higher Na+, Cl−, ABA, reduced K+, turgidity in leaf, osmolyte generation, and ROS scavenger production under stress (Malhi et al., 2021).
Melatonin
Melatonin (MLE) is a stress-reducing molecule by exogenous supplementation. It is involved in the improvement of seed germination, the proliferation of roots, better flowering, fruit set and enlargement, fruit ripening, shelf life, and quality as studied by Wang et al. (2021). Drought and salinity stresses can be mitigated by the supplemental spray of MLE on many vegetable crops because of MLE’s multifaceted functions (Wang et al., 2013; Qi et al., 2018; Altaf et al., 2022b). MLE has the good capability to scavenge toxic ROS, MDA, H2O2, and electrolyte leakage as studied by Wang et al. (2012). The enhanced level of endogenous MLE has the capability to mitigate challenges that occur from drought and salinity in agricultural crops as reported by Nawaz et al. (2018). Restricted translocation of minerals (macronutrients and micronutrients) is upregulated by supplemental application of MLE. Moreover, the uptake and absorption of minerals by roots are regulated due to the application of appropriate melatonin levels. Oxidative and osmotic stresses are relieved by MLE due to the regulation of endogenous hormones and activation of scavengers of oxidative stress markers (Neha et al., 2021). The morphology of roots is improved regarding uptake, absorption, and further translocation toward other plant parts by supplemental MLE. Drought and salinity are involved in the disruption of plant metabolism. Therefore, disturbance in the plant metabolism is an indication of a stress situation. The activation of oxidative stress markers like ROS, H2O2, and MDA is reduced, and their scavengers (enzymatic, non-enzymatic, and osmolytes) are activated. Therefore, the availability of nutrients to plants is imperative especially when growing under stressful conditions. Hence, MLE can be a suitable method for the alleviation of drought and salinity in vegetable crops.
Hormonal regulations are necessary to enhance vegetable crop tolerance against drought and salinity stress conditions (Figure 3). Different concentrations of antioxidant sprays improved crop performance by modulation of physiological and biochemical mechanisms in sweet potatoes (Lin et al., 2006) (Table 2).

Figure 3 Impact of phytohormones on different vegetable crops growing under drought and salinity stresses.
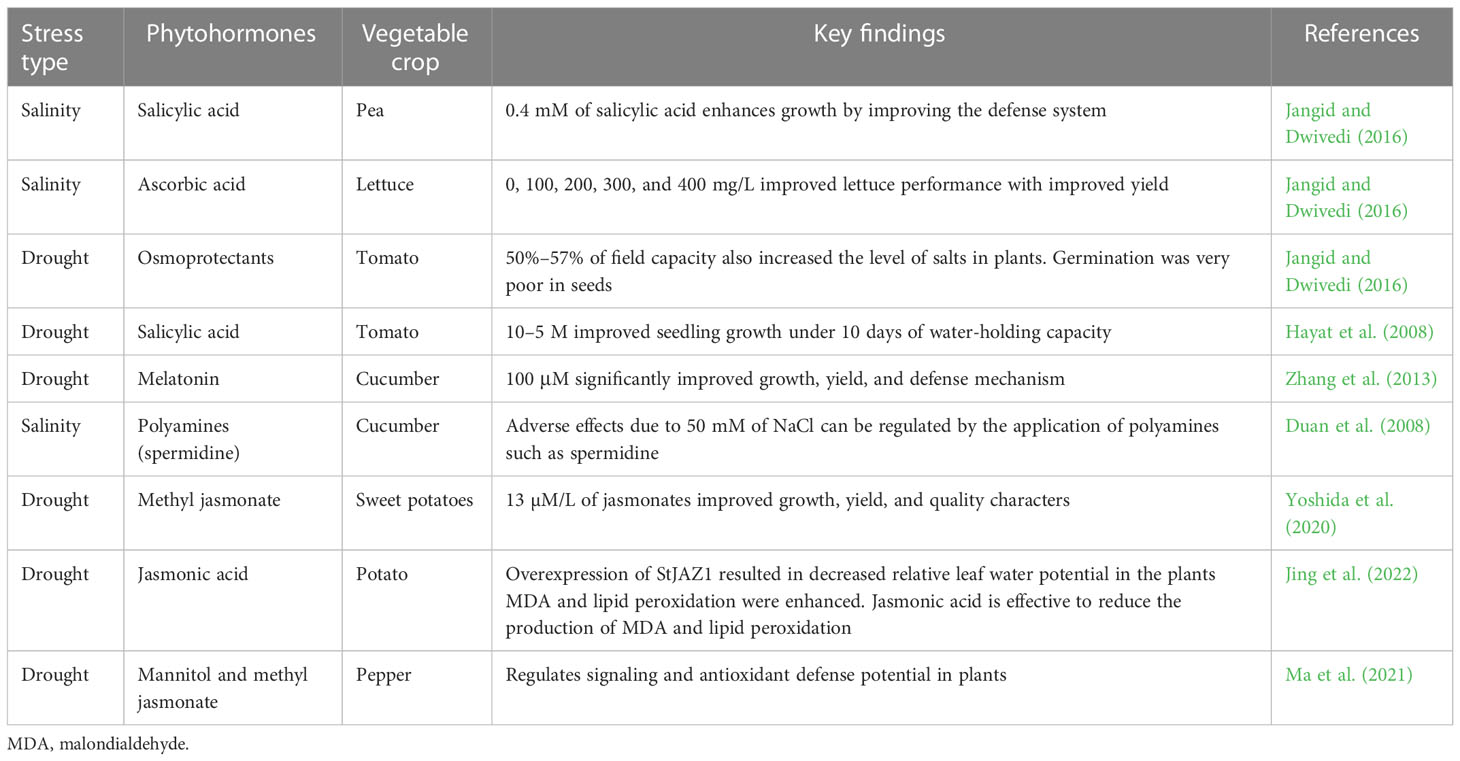
Table 2 The role of exogenous phytohormones against drought and salinity stresses in vegetable production.
Conclusion and prospects
In the present study, it has been explored that modulation of physiological and biochemical mechanisms is necessary for sustainable production of vegetable crops growing under drought and salinity stresses. Exogenous application of phytohormones is necessary for the improvement of vegetable growth, yield, photosynthetic pigments, minerals nutrient content, and defense-related characteristics. It has been concluded that phytohormones are necessary for the sustainable production of vegetable crops.
* Climate change, urbanization, and industrial zones are depleting and polluting water resources. Water shortage is going to worsen. To feed a huge population, it is necessary to develop management approaches to obtain higher vegetable production with limited water resources.
* Elevated drought and salinity conditions severely affect the productivity of vegetable crops. In this situation, phytohormones are considered a supportive strategy for the sustainable production of vegetable crops in the current scenario.
* To achieve zero hunger, it is necessary to elevate drought and salinity tolerance in vegetables. Moreover, the development of tolerant landraces is also a present need.
* Exploration of molecular basis, i.e., genome characterization, QTL mapping, marker-assisted selection (MAS), genome editing, genetic transformation, and genome sequencing are also imperative for the development of tolerant germplasm of vegetable crops.
Author contributions
JC: conceptualization, literature survey, writing major original draft, and review structure. XP: literature survey, writing—review and editing, and figure designing. All authors contributed to the article and approved the submitted version.
Funding
This work was sponsored in part by SZAI Science and Technology Incubation Project (PY2104).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Abouelsaad, I., Renault, S. (2018). Enhanced oxidative stress in the jasmonic acid-deficient tomato mutant def-1 exposed to NaCl stress. J. Plant Physiol. 226, 136–144. doi: 10.1016/j.jplph.2018.04.009
Ahmad, B., Jaleel, H., Sadiq, Y., A Khan, M. M., Shabbir, A. (2018). Response of exogenous salicylic acid on cadmium induced photosynthetic damage, antioxidant metabolism and essential oil production in peppermint. Plant Growth Regul. 86, 273–286. doi: 10.1007/s10725-018-0427-z
Ahmad, P., Kumar, A., Gupta, A., Hu, X., Azooz, M. M., Sharma, S. (2012). “Polyamines: role in plants under abiotic stress,” in Crop production for agricultural improvement (Dordrecht: Springer), 491–512. doi: 10.1007/978-94-007-4116-4_19
Ahmad, P., Umar, S., Sharma, S. (2010). Mechanism of free radical scavenging and role of phytohormones in plants under abiotic stresses. Plant Adapt. Phytoremed. Springer, Dordrecht.: Berline, Germany, 99–118. doi: 10.1007/978-90-481-9370-7_5
Ahmad, R., Kim, M. D., Back, K. H., Kim, H. S., Lee, H. S., Kwon, S. Y., et al. (2008). Stress-induced expression of choline oxidase in potato plant chloroplasts confers enhanced tolerance to oxidative, salt, and drought stresses. Plant Cell Rep. 27, 687–698. doi: 10.1007/s00299-007-0479-4
Akram, N. A., Shafiq, F., Ashraf, M. (2017). Ascorbic acid-a potential oxidant scavenger and its role in plant development and abiotic stress tolerance. Front. Plant Sci. 8. doi: 10.3389/fpls.2017.00613
Alhaithloul, H. A. S., Abu-Elsaoud, A. M., Soliman, M. H. (2020). “Abiotic stress tolerance in crop plants: role of phytohormones,” in Abiotic stress in plants (London: Intech Open), 1–22.
Ali, M., Afzal, S., Parveen, A., Kamran, M., Javed, M. R., Abbasi, G. H., et al. (2021a). Silicon mediated improvement in the growth and ion homeostasis by decreasing na+ uptake in maize (Zea mays l.) cultivars exposed to salinity stress. Plant Physiol. Biochem. 158, 208–218. doi: 10.1016/j.plaphy.2020.10.040
Ali, M., Kamran, M., Abbasi, G. H., Saleem, M. H., Ahmad, S., Parveen, A., et al. (2021b). Melatonin-induced salinity tolerance by ameliorating osmotic and oxidative stress in the seedlings of two tomato (Solanum lycopersicum l.) cultivars. J. Plant Growth Regul. 40, 2236–2248. doi: 10.1007/s00344-020-10273-3
Alian, A., Altman, A., Heuer, B. (2000). Genotypic difference in salinity and water stress tolerance of fresh market tomato cultivars. Plant Sci. 1, 59–65. doi: 10.1016/S0168-9452(99)00220-4
Altaf, M. A., Shahid, R., Altaf, M. M., Kumar, R., Naz, S., Kumar, A., et al. (2022b). Melatonin: First-line soldier in tomato under abiotic stress current and future perspective. Plant Physiol. Biochem. 185, 188–197. doi: 10.1016/j.plaphy.2022.06.004
Altaf, M. A., Shahid, R., Kumar, R., Altaf, M. M., Kumar, A., Khan, L. U., et al. (2022a). Phytohormones mediated modulation of abiotic stress tolerance and potential crosstalk in horticultural crops. J. Plant Growth Regul., 1–27. doi: 10.1007/s00344-022-10812-0
Andre, C. M., Schafleitner, R., Guignard, C., Oufir, M., Aliaga, C. A. A., Nomberto, G., et al. (2009). Modification of the health-promoting value of potato tubers field grown under drought stress: emphasis on dietary antioxidant and glycoalkaloid contents in five native andean cultivars (Solanum tuberosum l.). J. Agric. Food Chem. 57, 599–609. doi: 10.1021/jf8025452
Bernstein, N., Meiri, A., Zilberstaine, M. (2004). Root growth of avocado is more sensitive to salinity than shoot growth. J. Am. Soc Hortic. Sci. 129, 188–192. doi: 10.21273/JASHS.129.2.0188
Bhandari, S., Nailwal, T. K. (2020). Role of brassinosteroids in mitigating abiotic stresses in plants. Biol. 75, 2203–2230. doi: 10.2478/s11756-020-00587-8
Byun, M. O., Kwon, H. B., Park, S. C. (2007). “Recent advances in genetic engineering of potato crops for drought and saline stress tolerance,” in Advances in molecular breeding toward drought and salt tolerant crops Springer International Publishing Switzerland, 713–737. doi: 10.1007/978-1-4020-5578-2_29
Checker, V. G., Kushwaha, H. R., Kumari, P., Yadav, S. (2018). “Role of phytohormones in plant defense: signaling and cross talk,” in Molecular aspects of plant-pathogen interaction (Singapore: Springer), 159–184.
Chen, D., Shao, Q., Yin, L., Younis, A., Zheng, B. (2019). Polyamine function in plants: metabolism, regulation on development, and roles in abiotic stress responses. Front. Plant Sci. 9. doi: 10.3389/fpls.2018.01945
Choudhary, S. P., Kanwar, M., Bhardwaj, R., Yu, J. Q., Tran, L. S. P. (2012). Chromium stress mitigation by polyamine-brassinosteroid application involves phytohormonal and physiological strategies in Raphanus sativus l. PloS One 7, e33210. doi: 10.1371/journal.pone.0033210
Colla, G., Rouphael, Y., Leonardi, C., Bie, Z. (2010). Role of grafting in vegetable crops grown under saline conditions. Sci. Hortic. 127, 147–155. doi: 10.1016/j.scienta.2010.08.004
Dar, T. A., Uddin, M., Khan, M. M. A., Hakeem, K. R., Jaleel, H. (2015). Jasmonates counter plant stress: a review. Environ. Exp. Bot. 115, 49–57. doi: 10.1016/j.envexpbot.2015.02.010
Diao, Q., Song, Y., Qi, H. (2015). Exogenous spermidine enhances chilling tolerance of tomato (Solanum lycopersicum l.) seedlings via involvement in polyamines metabolism and physiological parameter levels. Acta Physiol. Plant 37, 1–15. doi: 10.1007/s11738-015-1980-y
Duan, J., Li, J., Guo, S., Kang, Y. (2008). Exogenous spermidine affects polyamine metabolism in salinity-stressed Cucumis sativus roots and enhances short-term salinity tolerance. J. Plant Physiol. 165, 1620–1635. doi: 10.1016/j.jplph.2007.11.006
Fahad, S., Hussain, S., Matloob, A., Khan, F. A., Khaliq, A., Saud, S., et al. (2015). Phytohormones and plant responses to salinity stress: a review. Plant Growth Regul. 75, 391–404. doi: 10.1007/s10725-014-0013-y
Fahad, S., Ullah, A., Ali, U., Ali, E., Saud, S., Hakeem, K. R., et al. (2019). “Drought tolerance in plants role of phytohormones and scavenging system of ROS,” in Plant tolerance to environmental stress (CRC Press) Taylor and Francis Group, England and Whales, 103–114.
Forni, C., Duca, D., Glick, B. R. (2017). Mechanisms of plant response to salt and drought stress and their alteration by rhizobacteria. Plant Soil. 410, 335–356. doi: 10.1007/s11104-016-3007-x
Galviz, Y. C., Bortolin, G. S., Guidorizi, K. A., Deuner, S., Reolon, F., de Moraes, D. M. (2021). Effectiveness of seed priming and soil drench with salicylic acid on tomato growth, physiological and biochemical responses to severe water deficit. J. Soil Sci. Plant Nutr. 21, 2364–2377. doi: 10.1007/s42729-021-00528-7
Gamalero, E., Glick, B. R. (2022). Recent advances in bacterial amelioration of plant drought and salt stress. Biol. 11, 437. doi: 10.3390/biology11030437
Gao, W., He, M., Liu, J., Ma, X., Zhang, Y., Dai, S., et al. (2018). Overexpression of Chrysanthemum lavandulifolium ClCBF1 in Chrysanthemum morifolium ‘White snow’ improves the level of salinity and drought tolerance. Plant Physiol. Biochem. 124, 50–58. doi: 10.1016/j.plaphy.2018.01.004
Ge, Y. X., Zhang, L. J., Li, F. H., Chen, Z. B., Wang, C., Yao, Y. C., et al. (2010). Relationship between jasmonic acid accumulation and senescence in drought-stress. Afr. J. Agr. Res. 5, 1978–1983.
Ghaffari, H., Tadayon, M. R., Nadeem, M., Razmjoo, J., Cheema, M. (2019). Foliage applications of jasmonic acid modulate the antioxidant defense under water deficit growth in sugar beet. Spanish. J. Agric. Res. 17, e0805–e0805.
Gilani, M., Danish, S., Ahmed, N., Rahi, A. A., Akrem, A., Younis, U., et al. (2020). Mitigation of drought stress in spinach using individual and combined applications of salicylic acid and potassium. Pak. J. Bot. 52, 1505–1513.
Giordano, M., Petropoulos, S. A., Rouphael, Y. (2021). Response and defense mechanisms of vegetable crops against drought, heat and salinity stress. Agric. 11, 463. doi: 10.3390/agriculture11050463
Glick, B. R. (2012). Plant growth-promoting bacteria: Mechanisms and applications. Scientific., 963401. doi: 10.6064/2012/963401
Groppa, M. D., Benavides, M. P. (2008). Polyamines and abiotic stress: recent advances. Amino Acids 34, 35–45. doi: 10.1007/s00726-007-0501-8
Gruda, N., Bisbis, M., Tanny, J. (2019). Impacts of protected vegetable cultivation on climate change and adaptation strategies for cleaner production–a review. J. Clean. Product. 225, 324–339. doi: 10.1016/j.jclepro.2019.03.295
Hayat, S., Hasan, S. A., Fariduddin, Q., Ahmad, A. (2008). Growth of tomato (Lycopersicon esculentum) in response to salicylic acid under water stress. J. Plant Interact. 3, 297–304. doi: 10.1080/17429140802320797
Hossain, M. N., Sarker, U., Raihan, M. S., Al-Huqail, A. A., Siddiqui, M. H., Oba, S. (2022). Influence of salinity stress on color parameters, leaf pigmentation, polyphenol and flavonoid contents, and antioxidant activity of Amaranthus lividus leafy vegetables. Molecules. 27, 1821. doi: 10.3390/molecules27061821
Hu, X., Zhang, Y., Shi, Y., Zhang, Z., Zou, Z., Zhang, H., et al. (2012). Effect of exogenous spermidine on polyamine content and metabolism in tomato exposed to salinity-alkalinity mixed stress. Plant Physiol. Biochem. 57, 200–209. doi: 10.1016/j.plaphy.2012.05.015
Iqbal, S., Wang, X., Mubeen, I., Kamran, M., Kanwal, I., Díaz, G. A., et al. (2022). Phytohormones trigger drought tolerance in crop plants: Outlook and future perspectives. Front. Plant Sci. 12. doi: 10.3389/fpls.2021.799318
Jakubowska, D., Janicka, M. (2017). The role of brassinosteroids in the regulation of the plasma membrane h+-ATPase and NADPH oxidase under cadmium stress. Plant Sci. 264, 37–47. doi: 10.1016/j.plantsci.2017.08.007
Jangid, K. K., Dwivedi, P. (2016). Physiological responses of drought stress in tomato: a review. Int. J. Agric. Environ. Biotechnol. 9, 53. doi: 10.5958/2230-732X.2016.00009.7
Jangid, K. K., Dwivedi, P. (2017). Physiological and biochemical changes by nitric oxide and brassinosteroid in tomato (Lycopersicon esculentum mill.) under drought stress. Acta Physiol. Plant 39, 1–10. doi: 10.1007/s11738-017-2373-1
Jing, S., Begum, S., Yu, L., Kawochar, M. A., Wang, E., Chen, Y., et al. (2022). StJAZ1-like mediated root architecture plays critical roles in drought susceptibility in potato. Environ. Exp. Bot. 202, 105008. doi: 10.1016/j.envexpbot.2022.105008
Jordan, J. T., Oates, R. P., Subbiah, S., Payton, P. R., Singh, K. P., Shah, S. A., Green, M. J., Klein, D. M., Cañas-Carrell, J. E.. (2020). Carbon nanotubes affect early growth, flowering time and phytohormones in tomato. Chemosphere 256, 127042.
Kamran, M., Danish, M., Saleem, M. H., Malik, Z., Parveen, A., Abbasi, G. H., et al. (2021). Application of abscisic acid and 6-benzylaminopurine modulated morpho-physiological and antioxidative defense responses of tomato (Solanum lycopersicum l.) by minimizing cobalt uptake. Chemos. 263, 128169. doi: 10.1016/j.chemosphere.2020.128169
Kamran, M., Parveen, A., Ahmar, S., Malik, Z., Hussain, S., Chattha, M. S., et al. (2019). An overview of hazardous impacts of soil salinity in crops, tolerance mechanisms, and amelioration through selenium supplementation. Int. J. Mol. Sci. 21, 148. doi: 10.3390/ijms21010148
Kaya, C. (2021). Nitrate reductase is required for salicylic acid-induced water stress tolerance of pepper by upraising the AsA-GSH pathway and glyoxalase system. Physiol. Plant 172, 351–370. doi: 10.1111/ppl.13153
Khan, M. I. R., Fatma, M., Per, T. S., Anjum, N. A., Khan, N. A. (2015). Salicylic acid-induced abiotic stress tolerance and underlying mechanisms in plants. Front. Plant Sci. 6. doi: 10.3389/fpls.2015.00462
Li, Q., Qin, Y., Hu, X., Jin, L., Li, G., Gong, Z., et al. (2022). Physiology and gene expression analysis of potato (Solanum tuberosum l.) in salt stress. Plant. 11, 1565. doi: 10.3390/plants11121565
Li, Q., Wang, G., Wang, Y., Yang, D., Guan, C., Ji, J. (2019). Foliar application of salicylic acid alleviate the cadmium toxicity by modulation the reactive oxygen species in potato. Ecotoxicol. Environ. Saf. 172, 317–325. doi: 10.1016/j.ecoenv.2019.01.078
Lim, C. W., Baek, W., Jung, J., Kim, J. H., Lee, S. C. (2015). Function of ABA in stomatal defense against biotic and drought stresses. Int. Jo. Mol. Sci. 16, 15251–15270. doi: 10.3390/ijms160715251
Ma, X., Li, Y., Gai, W. X., Li, C., Gong, Z. H. (2021). The CaCIPK3 gene positively regulates drought tolerance in pepper. Hortic. Res. 8, 216. doi: 10.1038/s41438-021-00651-7
Maksimovic, I., Ilin, Z. (2012). Effects of salinity on vegetable growth and nutrients uptake. Irrig. System. Pract. Challen. Environ. 9, 384.
Malhi, G. S., Kaur, M., Kaushik, P., Alyemeni, M. N., Alsahli, A. A., Ahmad, P. (2021). Arbuscular mycorrhiza in combating abiotic stresses in vegetables: An eco-friendly approach. Saud. J. Biol. Sci. 28, 1465–1476. doi: 10.1016/j.sjbs.2020.12.001
Manan, A., Ayyub, C. M., Pervez, M. A., Ahmad, R. (2016). Methyl jasmonate brings about resistance against salinity stressed tomato plants by altering biochemical and physiological processes. Pak. J. Agric. Sci. 53, 35–41. doi: 10.21162/PAKJAS/16.4441
Mangal, V., Lal, M. K., Tiwari, R. K., Altaf, M. A., Sood, S., Kumar, D., et al. (2022). Molecular insights into the role of reactive oxygen, nitrogen and sulphur species in conferring salinity stress tolerance in plants. J. Plant Growth Regul., 1–21. doi: 10.1007/s00344-022-10591-8
Mumtaz, M. A., Hao, Y., Mehmood, S., Shu, H., Zhou, Y., Jin, W., et al. (2022). Physiological and transcriptomic analysis provide molecular insight into 24-epibrassinolide mediated cr (VI)-toxicity tolerance in pepper plants. Environ. Poll. 306, 119375. doi: 10.1016/j.envpol.2022.119375
Nawaz, M. A., Jiao, Y., Chen, C., Shireen, F., Zheng, Z., Imtiaz, M., et al. (2018). Melatonin pretreatment improves vanadium stress tolerance of watermelon seedlings by reducing vanadium concentration in the leaves and regulating melatonin biosynthesis and antioxidant-related gene expression. J. Plant Physiol. 220, 115–127. doi: 10.1016/j.jplph.2017.11.003
Neha, P., Kumar, M., Solankey, S. S. (2021). “Impact of drought and salinity on vegetable crops and mitigation strategies,” in Advances in research on vegetable production under a changing climate (Cham: Springer), 235–253. doi: 10.1007/978-3-030-63497-1_13
Ormrod, D. P., Beckerson, D. W. (1986). Polyamines as antiozonants for tomato. HortSci. 21, 1070–1071. doi: 10.21273/HORTSCI.21.4.1070
Parveen, A., Ahmar, S., Kamran, M., Malik, Z., Ali, A., Riaz, M., et al. (2021). Abscisic acid signaling reduced transpiration flow, regulated na+ ion homeostasis and antioxidant enzyme activities to induce salinity tolerance in wheat (Triticum aestivum l.) seedlings. Environ. Technol. Innovat. 24, 101808. doi: 10.1016/j.eti.2021.101808
Parveen, A., Saleem, M. H., Kamran, M., Haider, M. Z., Chen, J. T., Malik, Z., et al. (2020). Effect of citric acid on growth, ecophysiology, chloroplast ultrastructure, and phytoremediation potential of jute (Corchorus capsularis l.) seedlings exposed to copper stress. Biomol. 10, 592. doi: 10.3390/biom10040592
Patane, C., Saita, A., Sortino, O. (2013). Comparative effects of salt and water stress on seed germination and early embryo growth in two cultivars of sweet sorghum. J. Agron. Crop Sci. 199, 30–37. doi: 10.1111/j.1439-037X.2012.00531.x
Pedranzani, H., Racagni, G., Alemano, S., Miersch, O., Ramírez, I., Peña-Cortés, H., et al. (2003). Salt tolerant tomato plants show increased levels of jasmonic acid. Plant Growth Regul. 41, 149–158. doi: 10.1023/A:1027311319940
Per, T. S., Khan, N. A., Reddy, P. S., Masood, A., Hasanuzzaman, M., Khan, M. I. R., et al. (2017). Approaches in modulating proline metabolism in plants for salt and drought stress tolerance: Phytohormones, mineral nutrients and transgenics. Plant Physiol. Biochem. 115, 126–140. doi: 10.1016/j.plaphy.2017.03.018
Qi, Z. Y., Wang, K. X., Yan, M. Y., Kanwar, M. K., Li, D. Y., Wijaya, L., et al. (2018). Melatonin alleviates high temperature-induced pollen abortion in Solanum lycopersicum. Molecules. 23, 386. doi: 10.3390/molecules23020386
Ramakrishna, B., Rao, S. (2015). Foliar application of brassinosteroids alleviates adverse effects of zinc toxicity in radish (Raphanus sativus l.) plants. Prot. 252, 665–677. doi: 10.1007/s00709-014-0714-0
Rehman, H., Farooq, M., Basra, S. M. A., Afzal, I. (2011). Hormonal priming with salicylic acid improves the emergence and early seedling growth in cucumber. J. Agric. Soc Sci. 7, 109–113.
Rodríguez-Delfín, A., Posadas, A., León-Velarde, C., Mares, V., Quiroz, R. (2011). “Effect of salt and water stress on the proline and total chlorophyll content and nutrients uptake on two sweet potato cultivars grown on soilless culture,” in II international symposium soilless culture hydroponics,Acta Horticulturae vol. 947. , 55–62. doi: 10.17660/ActaHortic.2012.947.4
Rodrıguez, P., Torrecillas, A., Morales, M. A., Ortuno, M. F., Sánchez-Blanco, M. J. (2005). Effects of NaCl salinity and water stress on growth and leaf water relations of Asteriscus maritimus plants. Environ. Experiment. Bot. 53, 113–123. doi: 10.1016/j.envexpbot.2004.03.005
Sabir, F., Noreen, S., Malik, Z., Kamran, M., Riaz, M., Dawood, M., et al. (2022). “Silicon improves salinity tolerance in crop plants: Insights into photosynthesis, defense system, and production of phytohormones,” in Silicon and nano-silicon in environmental stress management and crop quality improvement (Elesvier, Amsterdam, Netherlands), 91–103. doi: 10.1016/B978-0-323-91225-9.00022-4
Saidi, A., Hajibarat, Z. (2020). Application of next generation sequencing, GWAS, RNA seq, WGRS, for genetic improvement of potato (Solanum tuberosum l.) under drought stress. Biocat. Agric. Biotechnol. 29, 101801. doi: 10.1016/j.bcab.2020.101801
Sattar, F. A., Hamooh, B. T., Wellman, G., Ali, M. A., Shah, S. H., Anwar, Y., Mousa, M. A. A., et al. (2021). Growth and biochemical responses of potato cultivars under In vitro lithium chloride and mannitol simulated salinity and drought stress. Plants 10, 924. doi: 10.3390/plants10050924
Seiler, C., Harshavardhan, V. T., Rajesh, K., Reddy, P. S., Strickert, M., Rolletschek, H., et al. (2011). ABA biosynthesis and degradation contributing to ABA homeostasis during barley seed development under control and terminal drought-stress conditions. J. Exp. Bot. 62, 2615–2632. doi: 10.1093/jxb/erq446
Shahid, M. A., Pervez, M. A., Balal, R. M. (2011). Brassinosteroid (24-epibrassinolide) enhances growth and alleviates the deleterious effects induced by salt stress in pea (Pisum sativum l.). Aust. J. Crop Sci. 5, 500–510.
Shahid, R., Shijie, L., Shahid, S., Altaf, M. A., Shahid, H. (2021). Determinants of reactive adaptations to climate change in semi-arid region of Pakistan. J. Arid. Environ. 193, 104580. doi: 10.1016/j.jaridenv.2021.104580
Shahzad, R., Waqas, M., Khan, A. L., Hamayun, M., Kang, S. M., Lee, I. J. (2015). Foliar application of methyl jasmonate induced physio-hormonal changes in Pisum sativum under diverse temperature regimes. Plant Physiol. Biochem. 96, 406–416. doi: 10.1016/j.plaphy.2015.08.020
Shakoor, A., Abdullah, M., Sarfraz, R., Altaf, M. A., Batool, S. (2017). A comprehensive review on phytoremediation of cadmium (Cd) by mustard (Brassica juncea l.) and sunflower (Helianthus annuus l.). J. Biodivers. Environ. Sci. 10, 88–98.
Shama, A. M., Moussa, S., Abo El Fadel, N. (2016). Salicylic acid efficacy on resistance of garlic plants (Allium sativum l.) to water salinity stress on growth, yield and its quality. Alexandr. Sci. Exch. J. 37, 165–174.
Sharma, J., Upadhyay, A. K., Bande, D., Patil, S. D. (2011). Susceptibility of Thompson seedless grapevines raised on different rootstocks to leaf blackening and necrosis under saline irrigation. J. Plant Nutr. 34, 1711–1722. doi: 10.1080/01904167.2011.592566
Sobhanian, H., Aghaei, K., Komatsu, S. (2011). Changes in the plant proteome resulting from salt stress: toward the creation of salt-tolerant crops? J. Proteom. 74, 1323–1337. doi: 10.1016/j.jprot.2011.03.018
Soma, F., Takahashi, F., Yamaguchi-Shinozaki, K., Shinozaki, K. (2021). Cellular phosphorylation signaling and gene expression in drought stress responses: ABA-dependent and ABA-independent regulatory systems. Plants. 10, 756. doi: 10.3390/plants10040756
Sreenivasulu, N., Harshavardhan, V. T., Govind, G., Seiler, C., Kohli, A. (2012). Contrapuntal role of ABA: does it mediate stress tolerance or plant growth retardation under long-term drought stress? Gene 506, 265–273. doi: 10.1016/j.gene.2012.06.076
Viswanath, K. K., Varakumar, P., Pamuru, R. R., Basha, S. J., Mehta, S., Rao, A. D. (2020). Plant lipoxygenases and their role in plant physiology. J. Plant Biol. 63, 83–95. doi: 10.1007/s12374-020-09241-x
Wang, L., Luo, Z., Ban, Z., Jiang, N., Yang, M., Li, L. (2021). Role of exogenous melatonin involved in phenolic metabolism of Zizyphus jujuba fruit. Food Chem. 341, 128268. doi: 10.1016/j.foodchem.2020.128268
Wang, P., Sun, X., Li, C., Wei, Z., Liang, D., Ma, F. (2013). Long-term exogenous application of melatonin delays drought-induced leaf senescence in apple. J. Pineal. Res. 54, 292–302. doi: 10.1111/jpi.12017
Wang, P., Yin, L., Liang, D., Li, C., Ma, F., Yue, Z. (2012). Delayed senescence of apple leaves by exogenous melatonin treatment: toward regulating the ascorbate–glutathione cycle. J. Pineal. Res. 53, 11–20. doi: 10.1111/j.1600-079X.2011.00966.x
Wu, H., Wu, X., Li, Z., Duan, L., Zhang, M. (2012). Physiological evaluation of drought stress tolerance and recovery in caulifower (Brassica oleracea l.) seedlings treated with methyl jasmonate and coronatine. J. Plant Growth Regul. 31, 113–123. doi: 10.1007/s00344-011-9224-x
Wu, J., Shu, S., Li, C., Sun, J., Guo, S. (2018). Spermidine-mediated hydrogen peroxide signaling enhances the antioxidant capacity of salt-stressed cucumber roots. Plant Physiol. Biochem. 128, 152–162. doi: 10.1016/j.plaphy.2018.05.002
Yan, Z., Zhang, W., Chen, J., Li, X. (2015). Methyl jasmonate alleviates cadmium toxicity in Solanum nigrum by regulating metal uptake and antioxidative capacity. Biol. Plant 59, 373–381. doi: 10.1007/s10535-015-0491-4
Yoshida, C. H. P., Pacheco, A. C., Lapaz, A. D. M., Gorni, P. H., Vítolo, H. F., Bertoli, S. C. (2020). Methyl jasmonate modulation reduces photosynthesis and induces synthesis of phenolic compounds in sweet potatoes subjected to drought. Bragan. 79, 319–334. doi: 10.1590/1678-4499.20200203
Zaidi, A., Ahmad, E., Khan, M. S., Saif, S., Rizvi, A. (2015). Role of plant growth promoting rhizobacteria in sustainable production of vegetables: current perspective. Sci. Hortic. 193, 231–239. doi: 10.1016/j.scienta.2015.07.020
Zapata, P. J., Serrano, M., Pretel, M. T., Amorós, A., Botella, M.Á. (2004). Polyamines and ethylene changes during germination of different plant species under salinity. Plant Sci. 167, 781–788. doi: 10.1016/j.plantsci.2004.05.014
Zhang, D. Y., Kumar, M., Xu, L., Wan, Q., Huang, Y. H., Xu, Z. L., et al. (2017). Genome-wide identification of major intrinsic proteins in Glycine soja and characterization of GmTIP2; 1 function under salt and water stress. Scie. Rep. 7, 1–12. doi: 10.1038/s41598-017-04253-z
Zhang, H., Zhu, J., Gong, Z., Zhu, J. K. (2022a). Abiotic stress responses in plants. Nat. Rev. Genet. 23, 104 119.
Zhang, N., Zhao, B., Zhang, H. J., Weeda, S., Yang, C., Yang, Z. C., et al. (2013). Melatonin promotes water-stress tolerance, lateral root formation, and seed germination in cucumber (Cucumis sativus l.). J. Pineal Res. 54, 15–23. doi: 10.1111/j.1600-079X.2012.01015.x
Zhang, X., Zhang, L., Ma, C., Su, M., Wang, J., Zheng, S., et al. (2022b). Exogenous strigolactones alleviate the photosynthetic inhibition and oxidative damage of cucumber seedlings under salt stress. Sci. Hortic. 297, 110962. doi: 10.1016/j.scienta.2022.110962
Zhang, Z., Cao, B., Gao, S., Xu, K. (2019). Grafting improves tomato drought tolerance through enhancing photosynthetic capacity and reducing ROS accumulation. Protop. 256, 1013–1024. doi: 10.1007/s00709-019-01357-3
Zhang, Z., Lan, M., Han, X., Wu, J., Wang-Pruski, G. (2020). Response of ornamental pepper to high-temperature stress and role of exogenous salicylic acid in mitigating high temperature. J. Plant Growth Regul. 39, 133–146. doi: 10.1007/s00344-019-09969-y
Zhu, J. K. (2002). Salt and drought stress signal transduction in plants. Annu. Rev. Plant Biol. 53, 247. doi: 10.1146/annurev.arplant.53.091401.143329
Zhu, Y., Gong, H. (2014). Beneficial effects of silicon on salt and drought tolerance in plants. Agron. Sust. Dev. 34, 455–472. doi: 10.1007/s13593-013-0194-1
Zou, C., Liu, Z., Wei, G., Yang, B., Zou, X. (2017). Mitigation of water logging-induced damages to pepper by exogenous MeJA. Pak. J. Bot. 49, 127–1135.
Zulfiqar, H., Shahbaz, M., Ahsan, M., Nafees, M., Nadeem, H., Akram, M., et al. (2021). Strigolactone (GR24) induced salinity tolerance in sunflower (Helianthus annuus l.) by ameliorating morpho-physiological and biochemical attributes under in vitro conditions. J. Plant Growth Regul. 40, 2079–2091. doi: 10.1007/s00344-020-10256-4
Keywords: metabolic mechanisms, irrigation practices, sustainable yield, water needs, crops
Citation: Chen J and Pang X (2023) Phytohormones unlocking their potential role in tolerance of vegetable crops under drought and salinity stresses. Front. Plant Sci. 14:1121780. doi: 10.3389/fpls.2023.1121780
Received: 12 December 2022; Accepted: 26 January 2023;
Published: 28 February 2023.
Edited by:
Muhammad Kamran, University of Adelaide, AustraliaReviewed by:
Dr. Safina Naz, Bahauddin Zakariya University, PakistanAhmed H. El-Sappah, Zagazig University, Egypt
Deepesh Bhatt, Sarvajanik University, India
Copyright © 2023 Chen and Pang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jun Chen, Y2hlbmp1bkBzemFpLmVkdS5jbg==