 Aamir Manzoor
Aamir Manzoor Michaela A. Dippold
Michaela A. Dippold Sebastian Loeppmann
Sebastian Loeppmann Evgenia Blagodatskaya
Evgenia Blagodatskaya- 1Biogeochemistry of Agroecosystems, University of Goettingen, Goettingen, Germany
- 2Geo-Biosphere Interactions, Department of Geosciences, University of Tuebingen, Tuebingen, Germany
- 3Institute of Plant Nutrition and Soil Science, Christian–Albrechts University, Kiel, Germany
- 4Helmholtz Centre for Environmental Research–UFZ, Halle (Saale), Germany
The activity of extracellular phosphatases is a dynamic process controlled by both plant roots and microorganisms, which is responsible for the mineralization of soil phosphorus (P). Plants regulate the availability of soil P through the release of root mucilage and the exudation of low-molecular weight organic acids (LMWOAs). Mucilage increases soil hydraulic conductivity as well as pore connectivity, both of which are associated with increased phosphatase activity. The LMWOAs, in turn, stimulate the mineralization of soil P through their synergistic effects of acidification, chelation, and exchange reactions. This article reviews the catalytic properties of extracellular phosphatases and their interactions with the rhizosphere interfaces. We observed a biphasic effect of root metabolic products on extracellular phosphatases, which notably altered their catalytic mechanism. In accordance with the proposed conceptual framework, soil P is acquired by both plants and microorganisms in a coupled manner that is characterized by the exudation of their metabolic products. Due to inactive or reduced root exudation, plants recycle P through adsorption on the soil matrix, thereby reducing the rhizosphere phosphatase activity. The two-phase conceptual framework might assist in understanding P-acquisition (substrate turnover) and P-restoration (phosphatase adsorption by soil) in various terrestrial ecosystems.
Introduction
The total soil phosphorus (P) pool (e.g., 224–6,725 kg per ha) consists of organic and inorganic forms of P, of which 80% are immobile and not readily available to plants (Menezes–Blackburn et al., 2013). The organic fraction (Po) of soil P consists of dead plant and animal residues, representing 30–65% of the total soil P (Lu et al., 2020). The Po is not bioavailable unless it is mineralized to orthophosphate ( or its protonated forms) (Wang et al., 2021). The remaining 35–70% of soil P pool is inorganic P (Pi) and is found as insoluble forms of primary (e.g., apatite, smectite, and variscite) and secondary phosphate minerals of calcium (Ca), iron (Fe), and aluminum (Al) that cannot be absorbed by plants until mobilized (Richardson et al., 2009; Pizzeghello, 2011). The availability of P in the rhizosphere is greatly affected by several biophysical processes, including the catalysis of soil Po by extracellular phosphatases, which releases Pi (Oehl et al., 2001; Nannipieri et al., 2011). Phosphatase activity can be considered a dynamic process that responds to both root and soil microbial activities, as well as abiotic environmental conditions such as temperature, moisture, and soil adsorption characteristics (Schachtman et al., 1998; Dalling et al., 2016; Tang and Riley, 2021). The root penetration and rhizodeposition processes during nutrient uptake alter both the spatial and temporal distributions of pore spaces in the soil (Crawford et al., 2005; Hill et al., 2015). The altered soil pore space geometry also affects soil porosity and the pathways for transporting nutrients, water, and metabolic products (Hinsinger et al., 2008; Peth et al., 2008; Pettridge and Firestone, 2017). The volumetric water content and structural heterogeneity of the soil influence enzyme activity (Reed et al., 2015; Benard et al., 2019a). As volumetric water content increases in soil, tortuosity and fragmentation of the liquid phase are reduced, facilitating the diffusion of both enzymes and substrates; thus, both molecules can meet, and the substrate is enzymatically catalyzed (Allison et al., 2011; Ali et al., 2015; Ahmed et al., 2018).
On an ecosystem scale, plants acquire soil P through complex interactions with biotic (e.g., soil microorganisms) and abiotic (e.g., soil mineral surfaces) competitors. It has been demonstrated that plant roots and soil microorganisms (e.g., mycorrhizal fungi and their bacterial partners) form a symbiotic relationship that facilitates the hydrolysis of soil Po by soil phosphatases (Haque and Dave, 2005). The rhizosphere's physicochemical conditions are influenced by root exudation of metabolic products [e.g., mucilage and low-molecular weight organic acids (LMWOAs)], affecting the P availability (Araujo et al., 2012). For example, mucilage secretion facilitates root P uptake by modifying soil physical characteristics, such as increasing soil hydraulic conductivity and decreasing Pi adsorption to soil surfaces (Zarebanadkouki et al., 2019). Mucilage also affects phosphatase activity by establishing a moist biofilm-like environment (Ahmed et al., 2018; Bilyera et al., 2022). Similarly, LMWOAs have been shown to enhance the desorption of sparingly soluble phosphate monoesters via acidification, chelation, and exchange reactions, thus acting synergistically with phosphatases in mineralizing soil Po (Lambers et al., 2015; Koester et al., 2021).
Until now, the microbiology and biochemistry of the root–soil interface have not been sufficiently discussed in relation to root architecture and the physicochemical properties of zones adjacent to the roots. This review aimed to analyze the effects of root exudates and soil abiotic environment on phosphatase dynamics and provide answers to the following research questions (RQs).
RQ1: How do plants and microorganisms acquire soil P by releasing their metabolic products, which in addition to their direct role indirectly contribute to the acquisition of soil P by altering soil structure and structure-dependent processes?
RQ2: How do soil physicochemical properties influence phosphatase activity (e.g., by adsorption, immobilization, and inhibition), which lead to considerable changes in phosphatase catalytic properties?
RQ3: How can phosphatase activity be incorporated into a conceptual framework for quantifying P cycling in soil via processes, such as soil Po hydrolysis and phosphatase adsorption to soil matrix?
In order to address these questions, we described the effects of root and microbial exudates on phosphatase activity, examined the interaction of phosphatases with colloidal and mineral soil surfaces, and presented a conceptual two-phase framework that can be used to interpret soil P cycling (Figure 1).
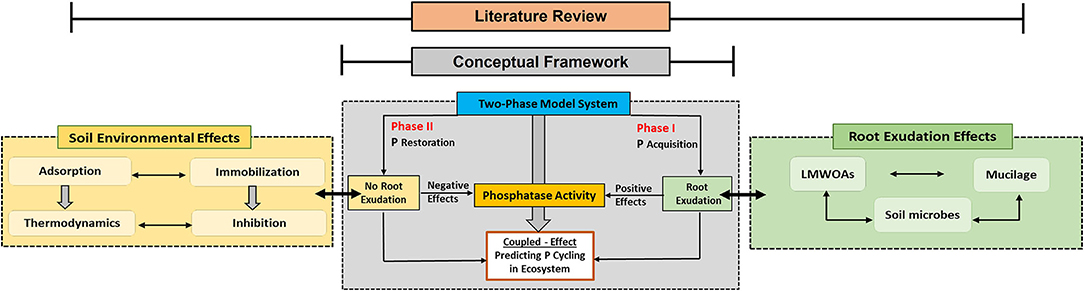
Figure 1. A conceptual two-phase framework that integrates root exudation and soil environment as main drivers of soil phosphatase activity. The dotted lines in each block of main sections indicate their combined effects on phosphatase activity. Based on the conceptual framework block, Phase I shows the soil Po acquisition pathway that is regulated by active root exudation, while Phase II shows soil Po restoration in the absence or with reduced root exudation effects. Both phases are controlled by the P demand of plants with the initiation of P mining processes (green boxes) or by reducing C investment into P mining (yellow boxes).
Effects of Root Exudation on Phosphatase Activity
A plant utilizes 20–40% of its photosynthetically fixed carbon (C) through root exudation to expedite nutrient uptake by supporting the growth of beneficial microorganisms, e.g., symbionts (Badri and Vivanco, 2009). Phloem metabolites (e.g., sugars, amino acids, and LMWOAs) are exuded via (i) passive diffusion, (ii) exudation mediated by transporter channels, and (iii) active transport across plasma membranes (Oburger and Jones, 2018). The exuded metabolic products are used by plants to sense nutrient availability relative to their demand at different physiological stages (Canarini et al., 2019). The efflux of exudate into the soil is carried out by plasmodesmata, which connect the cytoplasm of neighboring cells to establish living bridges between them (Ross–Elliott et al., 2017). The physiological mechanisms regulating exudation efflux are strongly affected by external environmental stimuli and exuded metabolic products (Williams and de Vries, 2019; Korenblum et al., 2020). The concentration of primary metabolites present in the root tip serves as a cue for sensing the soil environment and signaling between roots and shoots in order to modify root growth and nutrient allocation (De Schepper et al., 2013; Hu et al., 2018). The soil microorganisms affect the rate of exudation at the root tips through their consumption and enzymatic transformation of released metabolites (Sasse et al., 2019).
Low Molecular Weight Organic Acids
The root-exuded LMWOAs contain one to three carboxylic groups; among them, malic and citric acids (>2 mmol kg−1 soil) are the most abundant and prevalent organic acids associated with P mobilization (Denton et al., 2007; Aziz et al., 2011; Hunter et al., 2014). A typical concentration of LMWOAs in soil solution ranges between 0–50 μmol for dicarboxylic acids and 0–1 mmol for monocarboxylic acids (Strobel, 2001; Nwoke et al., 2008). Under abiotic stress, root tips of plants regulate malate anion efflux channels (ALMT) that initiate malate exudation in response to P deficiency (Ramesh et al., 2015; Gilliham and Tyerman, 2016; Mora–Macías et al., 2017). A high exudation of citric acid can lower soil pH and enhance P uptake by white lupin (Weisskopf et al., 2006). Citric acid, as a tricarboxylic acid, is more effective than dicarboxylic and monocarboxylic acids at mobilizing soil P (Khademi et al., 2009; Gang et al., 2012). This is because tricarboxylic acids are more capable of (i) forming stable chelation complexes with Ca in alkaline soils, thereby preventing precipitation of P (Kirk et al., 1999), (ii) complexing Al to reduce precipitation of Al–P hydroxy-phosphates, thus promoting weathering of P-bearing rocks (Pearse et al., 2007), (iii) remobilizing adsorbed P from soil surfaces via ion exchange, and (iv) preventing adsorption of P by soil matrix (Oburger et al., 2011). The majority of LMWOAs (57%) is consumed by soil microorganisms as part of their metabolic processes (e.g., respiration), while the rest is absorbed by soil colloids (Andrade et al., 2013).
Plant roots and soil microorganisms actively release phosphatases and LMWOAs as a result of ATP consumption for the acquisition of P in the P-deficient rhizospheres (Kelleher et al., 2004; Nannipieri et al., 2011; Ajmera et al., 2019). LMWOA anions either occupy sorption sites on the soil mineral surfaces to replace Pi or contribute to the hydrolysis of adsorbed Po (Lambers et al., 2015; Wang Y. et al., 2017). Their mutual relationship in mobilizing and mineralizing soil Po is shown to be synergistic in the conceptual model of Clarholm et al. (2015). The bioavailability of P depends on the interaction of pH, phosphatase activity, and the concentration of LMWOAs in the soil solution (Adeleke et al., 2012). For example, a significant positive correlation was found between pH and phosphatase activity in the rhizosphere of drought-tolerant and non-drought-tolerant corn varieties under water stress (Song et al., 2012). The authors observed that osmotic stress increased the concentration of LMWOAs exuded by roots, resulting in increased phosphatase activity during elongation, tasseling, and filling stages, whereas drought-tolerant corn varieties showed higher phosphatase activity. Similarly, glucose, glutamate, and citrate were found to significantly increase phosphatase activity in both clayey and sandy soils at pH 6.9 and 5.1 (Renella et al., 2007). A significant increase in phosphatase activity was observed in the rhizospheres of Crotalaria juncea and Tithonia diversifolia at a distance of 0–1 mm from the root surface (George et al., 2002). The above examples illustrate a close relationship between LMWOA exudation and phosphatase activity in promoting phosphatase-mediated Po mineralization by multiple mechanisms.
Root Mucilage
Mucilage secreted by the plant roots is composed of polysaccharides or long-chain sugar molecules and proteins that form a gelatinous substance that adheres to the root cap (Bais et al., 2008). As a result of mucilage secretion, a biopolymer layer forms in the soil around the root tips, known as rhizosheath structures (Delhaize et al., 2012). Rhizosheath structures are correlated to the length and density of the root hairs and modify the rhizosphere transport processes (George et al., 2014; Pausch et al., 2016). The gelatinous nature of mucilage facilitates the uptake of water and nutrients by increasing the hydraulic conductivity of soil particles and binding them together and with the roots (Kroener et al., 2014; Carminati et al., 2016). Mucilage, with its water absorption properties, affects soil hydraulic properties due to the presence of surfactants (Zickenrott et al., 2016). Mucilage can absorb water up to 1,000 times its dry weight to maintain a moist rhizosphere where bulk soil dries out more quickly (McCully and Boyer, 1997). The decrease in volumetric water content of the soil reduces surface tension and increases mucilage viscosity, which improves its ability to sustain liquid bridges across soil particles (Carminati et al., 2017; Benard et al., 2019b).
Phosphatase activity is closely related to root mucilage and its ability to increase hydraulic conductivity in soil pores (Jones et al., 2009). Due to mucilage secretion, rhizosphere volumetric water content is higher than bulk soil. These effects enhance enzyme diffusion into the root zone, creating a hotspot of enzymatic activity (Carminati et al., 2010; Manzoni et al., 2014). For example, a reduction in volumetric water content in the rhizosphere of barley caused a 97% decline in phosphatase activity, demonstrating a strong and reversible impact on soil phosphatase activity (Holz et al., 2019). In another study, a low concentration of mucilage secreted by maize roots (40 μg C g−1 soil is equivalent to 10% of microbial biomass C) significantly increased the soil phosphatase activity under drought (30% of soil water holding capacity) (Ahmed et al., 2018). In the presence of a high mucilage concentration (200 μg C g−1 soil is equivalent to 50% of microbial biomass C), the drought effect was overcompensated, leading to a one-third increase in phosphatase activity. Plants increase phosphatase activity directly by producing and exuding phosphatases through roots and indirectly by providing labile C, as found in root mucilage, to promote microbial activity (Rejsek et al., 2012; Spohn and Kuzyakov, 2013). The increase in viscosity of mucilage under drought conditions causes it to transform into a hydrophobic inter-particulate glue that strongly inhibits the diffusion of substrates toward phosphatases, resulting in decreased availability of Pi to plants (Hunter et al., 2014; Brax et al., 2020). Microorganisms in thin water films around soil particles may suffer dehydration and go dormant (as cysts or spores) or even die as a result of denaturation of their cellular components under osmotic stress (Schimel and Balser, 2007; Williams and Rice, 2007; Loeppmann et al., 2018).
Substrate–Product–Enzyme Interactions and Their Kinetic Description
The economic model of extracellular enzyme production treats soil microbial communities as economic units during their resource allocation to produce C-, N-, and P-releasing enzymes (Sinsabaugh and Moorhead, 1994). In spite of contrasting soil properties and nutrient stocks, the C- and P-cycling enzyme network enhanced nutrient acquisition to maintain microbial growth, which indicates a similar trade-off between C- and P-cycling enzymes (Loeppmann et al., 2020). Extracellular phosphomonoesterase activity regulates the P forms in soil and depends on the enzyme production itself, such as the release of the enzyme by plants and microbes into the soil solution, or the availability of labile P substrate (Turner and Haygarth, 2005; Burns et al., 2013). Total composition and quality of soil Po influence enzyme- and substrate-dependent catalysis of soil Po compounds (Quiquampoix and Mousain, 2005; Noll et al., 2019). Both acid and alkaline phosphomonoesterases hydrolyze phosphomonoesters, e.g., inositol phosphates and phytins, which constitute between 20 and 50% of the total soil P (Dalal, 1977; Turner, 2008). Compared with phosphomonoesterases, phosphodiesterases exhibit relatively low activity (e.g., in acidic soils) as a result of the resiliency of phosphodiesters to degradation or sorption, both due to their protected phosphate ester groups (Jarosch et al., 2019). The phosphodiesters found in fresh detritus of plant or microbial origin have a low persistence in the soil and rarely exceed 1% of the total soil P (Paul and Clark, 1989). The activities of phosphomonoesterases and phosphodiesterases cannot be separated completely. The phosphodiesterase produces highly labile substrates for the phosphomonoesterase, which, in turn, releases orthophosphate, which is a potential inhibitor for both enzyme groups (Leake and Miles, 1996).
In nature, enzyme-driven substrate catalysis occurs spontaneously and is primarily regulated by the diffusion of substrate monomers toward enzymes present in solution or adsorbed at the soil surface (Datta et al., 2017). The breakdown of soil Po by phosphatases involves the production of metaphosphate as an intermediate product, which is then converted into orthophosphate in the presence of water (Lassila et al., 2011). As a result of the rapid protonation of phosphoryl groups, phosphate-monoester anions have increased electrophilicity, which allows them to react more quickly with substrates in contrast to phosphate-diester anions (Hengge, 2005). Experimental evidence concerning the co-occurrence of phosphatase activity and the P depletion zone has been obtained up to a distance of 2–4 mm from the root surface (Nuruzzaman et al., 2006; Hummel et al., 2021). Phosphatase activity near the root surface reduces the concentration of Po substrate compared with the surrounding soil (Burns et al., 2013). As a result, a concentration gradient drives substrate diffusion from solution, while catalysis depends largely on phosphatases present in a free state or adsorbed on the soil surfaces. The rate of product formation and/or the rate of substrate catalysis (v) can be determined by the Michaelis–Menten Equation (1).
where Vmax is the maximum reaction rate of product formation and Km is the Michaelis constant, which indicates the affinity of enzymes to specific substrates (e.g., Km is inversely proportional to affinity) and is defined as substrate concentration (S) at half of Vmax.
The kinetic parameters of the substrate-dependent Michaelis–Menten model are merely the weighted means of the characteristics of the enzymatic activity catalyzed by many diverse enzyme systems in soil; hence, they are often defined as apparent Vmax and Km. However, the Michaelis–Menten approach in environmental modeling remains important for understanding the effects of plant–microbial interactions and soil physicochemical conditions on average substrate turnover rates (Eberwein et al., 2017).
Plant Photosynthesis and Microbial P Acquisition: A Coupled Relationship
Root secretion of mucilage and exudation of LMWOAs is carried out by passive and active mechanisms, respectively (Jones et al., 2009). Together with dying root cells, the released C sources serve as microbial substrates that promote microbial activity and growth (Loeppmann et al., 2016a,b). A coupled relationship has been observed between photosynthetic activity and root exudation, which is mediated by soil microorganisms through consumption and biotransformation of released metabolites (Doan et al., 2017; Vidal et al., 2018). For example, the photosynthetic P-use efficiency was found to be exceedingly high for Proteaceae and non-Proteaceae species with different leaf traits (Pereira et al., 2019). On nutrient-poor sites, P was used much more efficient for photosynthesis, indicating a species-independent increase in P-use efficiency in both Proteaceae and non-Proteaceae with decreasing soil P availability (Pereira et al., 2019). The competitive saturation of sorption sites by organic C anions caused by elevated organic C content of soil led to an increase in the lability, solubility, and transport of Pi in soil (Reddy et al., 1980; Ohno and Crannell, 1996; Brucker et al., 2020). The low availability of dissolved Pi in soil solution (<0.01–1 mg L−1) results in starvation conditions for soil microbes, thus they have to use multiple strategies to acquire P, such as improving P assimilation, optimizing intracellular P metabolism, and mobilizing extracellular P (Steinweg et al., 2013; Grafe et al., 2018; Pistocchi et al., 2020). We propose a coupled relationship that entails the provision of labile C to soil microorganisms through root exudates (e.g., mucilage, LMWOAs) and the microbial response to access soil P through phosphatase activity. This coupled relationship (Figure 2) controls both the uptake of Pi by plant roots and soil microorganisms, and hence phosphatase activity, which in turn is determined by phosphatase production and dissolved Pi in soil solution.
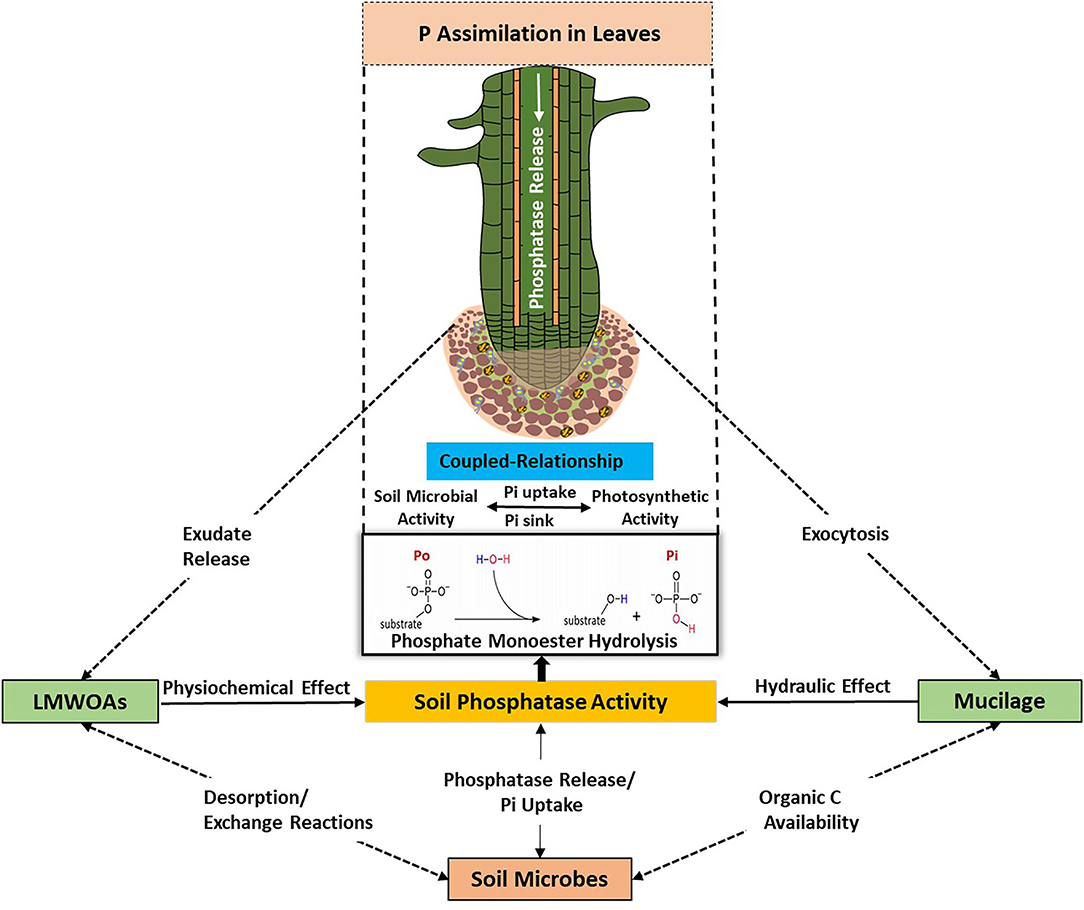
Figure 2. A scheme visualizing the coupled relationship between plants and soil microbes for the acquisition of available soil P resources and/or P mineralization induced via phosphatase activity. Root exudation of mucilage and LMWOAs exert favorable physicochemical and hydraulic changes in the rhizosphere for P accessibility. Soil microbes ensure a mutual relationship with plant roots by the production of phosphatases and release of LMWOAs, which favor P supply from soil P reservoir.
Effects of Abiotic Soil Environment on Phosphatase Activity
The abiotic soil environment strongly regulates substrate degradation rates since it influences the nutrient-foraging strategies of soil microorganisms (Schimel and Weintraub, 2003; Gianfreda, 2015). After being released from plant roots as exudates or microbial secretions, extracellular phosphatases form many types of associations (Figure 3), including (i) enzyme–substrate complexes, (ii) adsorption to clay minerals, (iii) complexing with soil organic matter surfaces through entrapment, absorption, or co-polymerization, and/or (iv) present in freely diffusible forms in soil solution (Wallenstein and Burns, 2011).
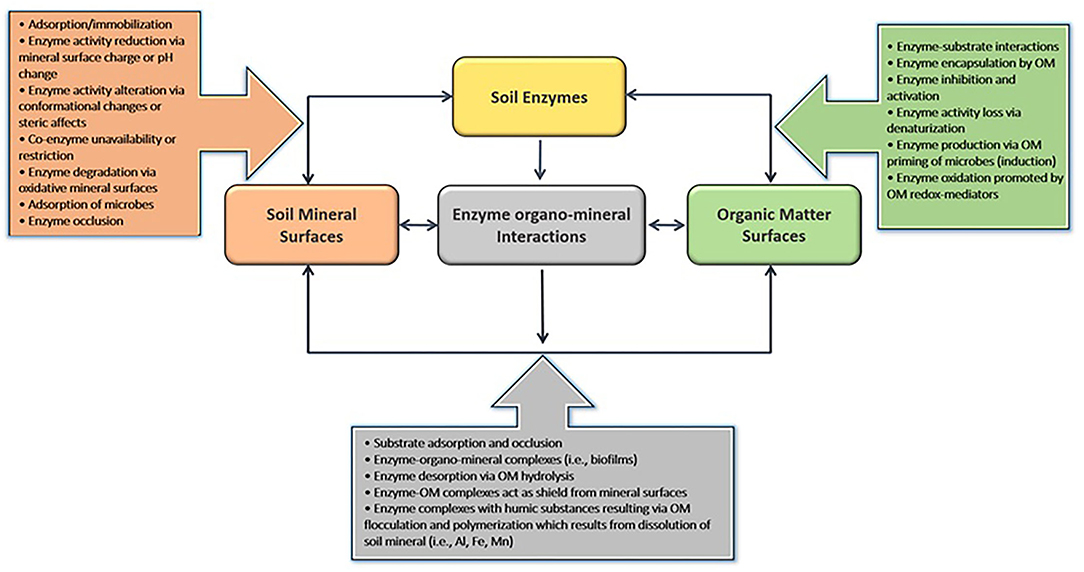
Figure 3. A summary of the interactions between soil colloidal and organic matter surfaces and extracellular enzymes and their effect on enzyme–substrate complex formation.
Phosphatase Adsorption on Colloidal and Mineral Soil Surfaces
In soil, extracellular phosphatases are present in solution or they bind reversibly to soil colloidal and mineral surfaces, which protect them against microbial decomposition and exposure to environmental stresses (Burns, 1982; Kedi et al., 2013; George et al., 2014). Several mechanisms can account for the strong adsorption affinity of phosphatases for soil colloidal components (<2 μm), including electrostatic Van der Waals forces, Lewis's acid–base effects, hydrophobic interactions, and conformational entropy (Ikeda and Kuroda, 2011). In the rhizosphere, organic and inorganic ligands (e.g., LMWOAs and phosphate anions) contribute to phosphatase adsorption through ion exchange, covalent complexation, and hydrogen bonding (Nannipieri et al., 1988). The abundance of silica (SiO2) in soil facilitates the physicochemical interaction of phosphatases with soil surfaces through the negative charge of silanol groups (Si–OH) and siloxane bridges (–Si–O–Si–) (Zhuravlev, 2000). Typically, free or colloid-associated phosphatases are transported to soil surfaces by convection (laminar or turbulent flows); however, their final adsorption to soil surfaces is carried out by diffusion (Datta et al., 2017; Guber et al., 2022). Due to electrostatic attraction, oppositely charged phosphatases adopt a spatial orientation toward soil surfaces during adsorption (Margalef et al., 2017). The phosphatase adsorption by colloidal and mineral surfaces of soil can be explained by Langmuir's Equation (2).
where Xm is the maximum adsorption capacity of enzymes, C is the equilibrium concentration of enzymes in solution, and K is the binding energy of enzyme molecules (e.g., low K values indicate high affinity of enzymes for colloids and minerals) (Huang et al., 2003). Langmuir's equation assumes uniform surface area and is commonly used to estimate phosphate adsorption, but it can underestimate the amount of adsorption since it neglects multiple possible sorption pathways (Hussain et al., 2012; Kruse et al., 2015).
In acidic soil environments, phosphatases adsorb to finer colloidal particles (e.g., goethite and montmorillonite), which can be exacerbated by the presence of iron oxides with large surface areas and high anion exchange capacity (Huang et al., 2005; Zhao et al., 2012). Organic components (e.g., humic acids) facilitate phosphatase adsorption by trapping them within their macromolecular networks (Kelleher et al., 2004). Adsorption of phosphatase by LMWOAs and inorganic anions decreased in the following order: phosphate > tartrate > oxalate > acetate (Huang et al., 2003). Nevertheless, addition of acetate (0–10 mmol) increased phosphatase adsorption onto goethite and kaolinite surfaces by protonating the surfaces of the acetate adsorbates, thereby creating more adsorption sites. As anion adsorption density increased, the promoter effect of acetate diminished at concentrations above 10 mmol as a result of steric and competitive interactions (Kafkafi et al., 1988; Geelhoed et al., 1998). By contrast, higher concentrations of oxalate (0–50 mmol) reduced phosphatase adsorption on goethite surfaces (Zhao et al., 2012). Oxalate affects the adsorption properties of clay minerals by forming mono- (pH 3.5) and di-coordinate (pH 4.5 and 5.5) complexes that reduce phosphatase adsorption through conformational changes (Bhatti et al., 1998). Phosphatases maintain a neutral charge at their isoelectric point (pH~5), which is necessary to preserve their natural molecular configuration (Huang et al., 2003). A reduction in adsorption at the isoelectric point would facilitate the achievement of maximum specific activity by reducing the congestion among phosphatase molecules. When pH decreases from 5, an electrostatic repulsion develops between phosphatases and clay minerals (both being positively charged), resulting in reduced adsorption (Leprince and Quiquampoix, 1996). In addition, the negative charge on the surface of phosphatases and clay minerals above the isoelectric point inhibits adsorption, thereby allowing the phosphatases to diffuse into the water-filled pores of the soil. We therefore speculate that phosphatase activity and adsorption may be affected by the competitive adsorption of enzymes and organic/inorganic anions. Both mechanisms (phosphatase–Po substrate and phosphatase–soil interactions) are controlled by changes in enzyme configuration/coverage on soil colloids and organic anion-induced dissolution of soil colloids and minerals.
Phosphatase Immobilization and Inhibition Kinetics
Adsorption of enzymes generally prevents their degradation since immobilized enzymes are less susceptible to environmental stresses, because their 3D structure is stabilized through the surface–surface interactions, and stressors are prevented from accessing them (Joner and Johansen, 2000). Due to immobilization, phosphatase activity is largely controlled by pH fluctuations, temperature variations, structural orientation effects, and steric hindrances caused by organic or inorganic ligands (Demanèche et al., 2009; Kedi et al., 2013). For example, after immobilization by minerals and sediments, alkaline phosphatases showed a 5% decline in activity (Zhu et al., 2016). However, after immobilization by soil inorganic components, acid phosphatases showed distinct patterns of activity in the following order: allophane > kaolin > Fe oxide > montmorillonite > Al oxide = Mn oxide (Shindo et al., 2002). Acid phosphatases immobilized on Al, Fe, Mn oxides, or montmorillonite retained 13–23% of their activity, whereas acid phosphatases immobilized on kaolin and allophane retained 57 and 77% of their activity, respectively. As a result of adsorption, acid phosphatases showed a decrease in Vmax, an increase in Km, and a decrease in Vmax/Km ratio (Shindo et al., 2002). The specific activity and adsorption strength of acid phosphatases increased with the increasing concentration of organic anions (e.g., oxalate) (Zhao et al., 2012). The general consensus is that organic ligands increase phosphatase activity (a stimulatory effect) compared with inorganic ligands (a competitive effect) (Pascual et al., 2002).
The inhibition of phosphatases by immobilization occurs through interactions with colloidal and mineral soil surfaces via competitive inhibition, non-competitive inhibition, or mixed mechanisms (Deng and Tabatabai, 1995). Phosphatase inhibition causes the formation of enzyme–substrate, enzyme–inhibitor, or enzyme–substrate–inhibitor complexes (Quiquampoix et al., 2002; Tietjen and Wetzel, 2003). The presence of certain inorganic anions (e.g., phosphate) and heavy metals (e.g., arsenate) may inhibit phosphatases, leading to a reduction in their affinity for substrates (Tian et al., 2018). As phosphatases are synthesized by plants under conditions of Pi limitation, their activity is referred to as Pi-repressible activity (Rombola et al., 2014; Čapek et al., 2021). Phosphatase activity is inhibited by dissolved Pi concentrations in solution, suggesting a close relationship between the use of dissolved Pi in the soil and the mobilization of plant's internal P reserves (Gianfreda and Ruggiero, 2006; Maseko and Dakora, 2013). A direct, positive relationship exists between phosphatase activity and product concentration when soil biota gains optimal access to dissolved Pi under conditions of high P demand (Treseder and Vitousek, 2001). The inverse relationship can be observed when high levels of phosphatases are present without significant formation of products (Allison S. D. et al., 2007; Allison V. J. et al., 2007). In addition, the negative correlation is also evident when phosphatases are adsorbing to colloidal and mineral soil surfaces, e.g., lowering their catalytic efficiency (Weintraub and Schimel, 2005).
Phosphatase immobilization on montmorillonite or goethite surfaces results in a reduced inhibitor affinity by reducing the accessibility of inhibitors to the catalytic sites (Bhattacharyya et al., 2008; Wang Z. Q. et al., 2017). For example, the competitive inhibition represents the direct competition between an inhibitor and substrate at the enzyme's active site, resulting in a decrease in Km but an increase in Vmax, indicating that the enzyme's active sites are temporarily bound to the organic matter surfaces (Zimmerman and Ahn, 2010). The ability to characterize the competitive inhibition of phosphatases can be useful in predicting inhibitor toxicity to soil phosphatase activity (Wang et al., 2018). As a result of high inhibitor concentrations, Km increases, resulting in a more difficult-to-break ESI complex, which reduces Vmax (Dick and Tabatabai, 1987; Cornish–Bowden, 2015; Wang Z. Q. et al., 2017). Phosphatases immobilized on mineral soil surfaces exhibit a complete inhibitory effect, whereas free phosphatases in solution show a mixed, linear inhibitory effect (Tian et al., 2018). The reason for this is that inhibitors (e.g., phosphate anions, arsenate) cannot fully compete with the substrate on the active sites of immobilized phosphatases due to their adsorption by mineral soil surfaces. Conversely, inhibitors are better able to compete with the substrate at the active sites of free phosphatases when there are few adsorption sites supplied by the enzyme–substrate complex. A non-competitive inhibition mechanism is characterized by inhibitors binding to the enzyme–substrate complex or enzyme itself, but not the active site, causing a reduction in Vmax and no change in Km (Ahn et al., 2006). In acidic soils, non-competitive inhibition reduces phosphatase activity to a significant extent, but this can be slowly recovered with an increase in organic matter content and cation exchange capacity (Wang et al., 2018). A decline in phosphatase activity in alkaline soils can be attributed to competitive inhibition and non-competitive inhibition effects, with an increase in Km resulting from increasing inhibitor concentration.
Conceptual Two-Phase Framework
In a heterogeneous soil environment, diffusion and mass flow-based transport processes co-regulate Po mineralization through extracellular phosphatases. During water flow toward the plant's xylem, mass flow facilitates substrate transport to phosphatases close to the roots, while diffusion governs Po hydrolysis and is dominated by readily available dissolved Pi in soil. The fast decomposition of the substrate (Po) and the uptake of released products (Pi) in the immediate vicinity of roots enhance the diffusion of P toward root surfaces. Due to the strong adsorption of Pi ions on the soil surfaces, diffusion alone is not sufficient to meet the plant's P requirements. This results in a non-equilibrium shift between high Pi uptake and low replenishment from the soil, resulting in P depletion zones that extend from the root surface toward the bulk soil. The production and subsequent activity of extracellular phosphatases are strongly related to the depletion of Pi from the soil solution, suggesting that plants meet most of their P needs by mineralizing soil Po. In addition, metabolic products that are excreted by the roots (e.g., mucilage and LMWOAs) play an important role in both modifying the soil physicochemical conditions to increase P accessibility and indirectly enhancing extracellular phosphatase activity. In our conceptual two-phase framework, we explain how root-exuded metabolic products, such as mucilage and LMWOAs, regulate phosphatase activity by altering soil physicochemical and hydraulic conditions. It is hypothesized that exudate-imposed conditions can affect soil P bioavailability in response to the P demand of soil biota. An opposite scenario occurs when the active P demand has been met, resulting in reduced exudation of bioactive compounds, which conserves available P resources and phosphatases by adsorption to the soil matrix.
Mechanistic Interpretation
The framework aimed to address the challenges involved in developing efficient, reliable, and useful P models for ecosystem studies. To provide a mechanistic explanation, a two-phase dynamic system (Figure 4) is constructed to describe the acquisition of soil P by plants via root exudation and the conservation of soil P via adsorption. Both phases are influenced by the presence or absence of stimulating root exudates.
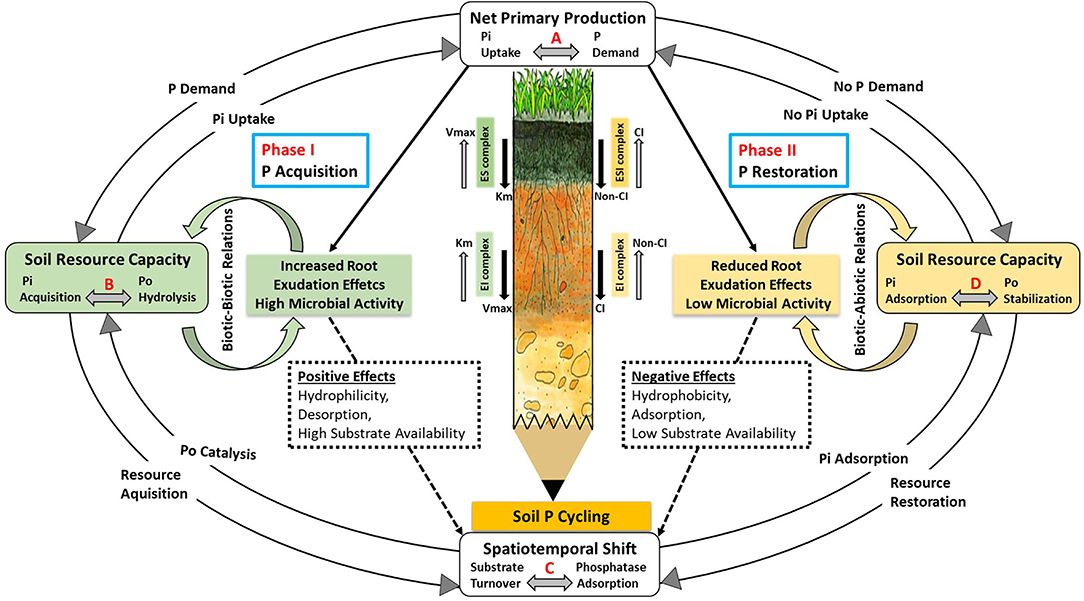
Figure 4. A conceptual two-phase framework for predicting P bioavailability in ecosystems based on phosphatase activity and soil phosphatase interactions. In “Phase I”, plants acquire soil P via phosphatase activity, which is controlled by a coupled relationship (biotic relations) with soil microorganisms in the presence of exuded metabolic products such as mucilage and LMWOAs. The “phase II” is characterized by the conservation of soil P resources under the dominance of biotic and abiotic interaction among microbes, phosphatases, and the different P forms with the soil mineral surfaces in the absence of root exudation. During phase II, extracellular phosphatases are immobilized and adsorbed onto colloidal and mineral soil surfaces, which inhibits their activities through modifications in hydraulic and physicochemical properties. CI, competitive inhibition; non-CI, non–competitive inhibition; ES, enzyme–substrate; EI, enzyme–inhibitor; ESI, enzyme–substrate–inhibitor.
The proposed two-phase model system is composed of several interrelated processes marked with letters A, B, C, and D. The processes designated with the letter “A” are connected with the P demand of plants, which in turn facilitates root uptake of dissolved Pi through diffusion, which is relatively low (0.1–5 × 10−13 m2 s−1) as compared to other nutrients (Bhadoria et al., 1991). Due to a continuous uptake of Pi by plant root systems, a rapid equilibrium between dissolved Pi and labile soil Pi fractions is disturbed, leading to a faster release of Pi into solution (Tinker and Nye, 2000; Mengel et al., 2001). Those processes with the letter “B” involve an increase in the P requirement of plants due to a low level of dissolved Pi, leading to physiological changes in plants. Consequently, plants exude metabolic products (e.g., LMWOAs and phosphatases), which facilitate the mobilization of sparingly soluble soil P forms and/or the mineralization of soil Po (Chen et al., 2002; Merlin et al., 2016). The hydrolysis of soil Po is inhibited when dissolved Pi is no longer the limiting factor for plant uptake, suggesting a negative relationship between Pi availability and phosphatase activity (Moscatelli et al., 2005; Marklein and Houlton, 2012). The letter “C” represents catalytic processes involving Po mineralization and/or phosphatase soil adsorption in the absence of root-exuded metabolic products. The coregulation of labile Pi uptake and Po mineralization is contingent on the plant's P demand, which then drives the system either toward P-mineralization or microbial P-immobilization (Zhang et al., 2018; McConnell et al., 2020). The letter “D” denotes processes that are executed under P restricted conditions that may result in P immobilization (e.g., increasing P storage in microbial cells), which will eventually result in the transition from P acquiring systems to an efficient P recycling system (Lang et al., 2017; Manzoni et al., 2021). The decrease in P demand for immobilization by plants and soil microbes also results in a decrease in the exudation of their metabolic products, including the production and exudation of extracellular phosphatases (Rejsek et al., 2012). Following the cessation of exudation of metabolic products (e.g., mucilage, LMWOAs) from plants' roots, the rhizosphere conditions modify to trigger P immobilization pathways involving adsorption of Pi, Po, and phosphatases to soil surfaces.
Phase I: P Acquisition by Increased Root Exudation Effects
The secretion of mucilage at the root tips has multiple functions, including (i) mobilizing Pi from mineral sorption sites, (ii) transporting mobilized Pi to the roots, and (iii) facilitating the mineralization of Soil Po by extracellular phosphatases. Mucilage stimulates the growth and activity of soil microorganisms by providing them with labile C, thereby indirectly increasing phosphatase activity. The texture of mucilage gel promotes the retention of more water during drought events, so mucilage increases soil hydraulic conductivity. When the soil dries out, water diffuses from the gel and forms liquid bridges with soil surfaces that encapsulate root tips. These liquid bridges act as small water channels connecting the soil particles with each other and with the surface of the roots to facilitate the transport and uptake of Pi. In situations of high P demand, mucilage reduces the diffusion barrier of phosphate monoesters and allows them to reach phosphatases near the roots. The increased secretion of mucilage reduces its viscosity, which increases the surface tension of the solution directly attached to the soil pore surfaces. Ultimately, this increases the hydrophilicity of phosphatase catalytic active sites required for Po catalysis and enhances phosphate desorption from soil surfaces. Mucilage gels are capable of retaining water, creating a hydrodynamic barrier that can restrict the diffusion of phosphatases, thereby preventing their adsorption at soil surfaces. The catalytic efficiency of phosphatases increases with desorption resulting in the formation of an ES complex as indicated by a high Vmax and a low Km. In summary, mucilage secretion can either increase or decrease phosphatase activity by increasing soil hydraulic conductivity and desorbing phosphate anions. Similarly, root-exuded LMWOAs alter soil physicochemical properties in two ways: they (i) increase P bioavailability through the solubilization of crystalline and/or amorphous P-containing minerals and (ii) function synergistically with phosphatases through anion-exchange reactions. This synergistic relationship results in (i) the blocking of phosphatase adsorption sites at the soil surface, which helps to reduce (re-) adsorption and (ii) the introduction of pH changes to the isoelectric point of phosphatases, which increases their activity. A net neutral charge on phosphatases is ensured by the isoelectric point, which maintains their natural molecular configuration during the formation of ES complexes. A maximum level of phosphatase activity is observed between pH values of 3 and 5, which subsequently increases due to root exudation of LMWOAs. At pH < 3, exuded LMWOAs represent an indirect source of competitive inhibition (e.g., desorption of Pi and inhibitor ions from soil surfaces), thereby decreasing the substrate accessibility to phosphatases. Similarly, phosphatase soil adsorption reaches its maximum level below the isoelectric point and decreases at a low pH. Both phosphatases and adsorbing soil surfaces are positively charged at low pH, resulting in a strong electrostatic repulsion, which may account for phosphatase desorption in acidic soils.
Bridging Phase: A Spatiotemporal Shift in Root Activity
Phase I and its underlying processes are continued until Pi availability is no longer a limiting factor for plant uptake. The acquisition of dissolved Pi by plant roots depends on many factors, including (i) the P demand of plants at each stage of their physiological development, (ii) the soil physicochemical properties, and (iii) the environmental factors associated with P accessibility to plants (e.g., water availability). Our hypothesis states that the P demand of plants is not limited by excessive dissolved Pi, but it is shifted toward alternative locations during root development in the soil. Root exudation activity may improve effective utilization of dissolved Pi through reallocation of soil P resources during root growth. The term “effective utilization” refers to acquiring soil Pi in a way that is advantageous over the addition of exuded metabolites. Plant roots secrete mucilage and exude LMWOAs, which contribute to effective soil P acquisition. These processes may also have residual effects due to spatiotemporal shifts in root activity while the roots are still growing in the soil. In our opinion, either the plants' P requirement has been met at the “depleted hotspots” or there is a decline in root exudation activity as a result of environmental factors (e.g., water scarcity). In phase II of the current framework, we refer to these residual rhizosphere effects as “depleted root activity” effects that contribute to P restoration in the soil matrix.
Phase II: P Restoration by Reduced Root Exudation Effects
The spatiotemporal shifts encountered during root growth result in reduced P demand of plants at individual sites, and as a consequence, reduced root exudation activity, thus leading to the “depleted root activity effects” effects. These effects increase during drought, disrupting the coupled relationship between plant roots and soil microorganisms that was established during phase I. When root exudation is reduced, microorganisms struggle to mobilize sufficient P, resulting in a joint C and P deficiency, which results in a reduced level of activity and growth. The resulting effect is the interaction between soil microbes and the mineral surfaces of the soil to maintain the basic requirement of microbial P acquisition. The transition to microbial dormancy when nutrients are scarce starts phase II of our conceptual framework leading to soil P restoration. Drought accelerates P restoration through the adsorption of extracellular phosphatases and Pi anions, as well as labile soil Po as high-quality substrates for future reserves when soil location shifts toward a phase I condition. A decrease in mucilage secretion reduces hydraulic conductivity and soil pore connectivity, which in turn has a negative effect on the transport and uptake of dissolved Pi, as well as its desorption from the soil surfaces. The decrease in hydraulic conductivity as a result of low soil volumetric water content restricts the transport of phosphatases and substrate to thin water films covering the soil pore surfaces. In the absence of fresh mucilage secretion by plant roots, the concentration of mucilage in the rhizosphere increases due to drying, which increases viscosity and decreases surface tension within the biogel. As a result of the lower surface tension, the hydrodynamic barrier between phosphatases and adsorbing soil surfaces is reduced, and phosphatases adsorb to soil surfaces at different concentration gradients. During Phase I, this concentration gradient exists between adsorbed phosphatases and product concentrations (dissolved Pi) in the soil solution and serves as a source of competitive inhibition for phosphatases. During Phase II, this gradient exists between adsorbed phosphatases and their concentration near the soil surfaces. Moreover, a reduction in LMWOAs exudation by plant roots may result in an increase in phosphatase adsorption, since LMWOAs desorption frees up phosphatase adsorption sites. In the event of a pH rise above the isoelectric point (>5), a negative charge is generated, causing electrostatic repulsion between phosphatases and soil colloids. Consequently, the presence of high alkalinity also results in an increase in inhibitor concentration in solution, which leads to a competitive inhibition that reduces substrate accessibility to phosphatases, leading to the formation of the enzyme–inhibitor or enzyme–substrate–inhibitor complexes.
The P restoration phase is in effect until the “depleted root activity” effects are mitigated by increasing the P demand of plants, thereby transitioning from phase II to phase I. The dynamic spatiotemporal shifts between both phases are considered essential to our cyclic two-phase framework.
Conclusion
Extracellular phosphatase activity is considered to be the mechanism by which soil Po is made available to plants and microorganisms through enzymatic catalysis. The production of extracellular phosphatases and their subsequent activity are governed by the P demand of the plant, as well as the availability of Po substrate in the rhizosphere. Upon the plant's demand for P, non-equilibrium conditions develop between readily available dissolved Pi and the soil's labile P pool, resulting in an increase in P mineralization through root and microbial phosphatase activity. In addition to the catalytic mechanism, several factors determine phosphatase activity, such as the accessibility of Po substrates, the pH-sensitive charge of phosphatases, and the adsorption of phosphatases and their substrates to colloidal and mineral soil surfaces. The catalysis of soil Po is strongly influenced by physicochemical properties of soil, including the presence of organic and phosphate anions. It has been observed that the amount of dissolved Pi in soil exerts a bimodal effect on extracellular phosphatase activity (i) directly, when Pi anions are excessive, they act as competitive inhibitors of phosphatases, preventing them from binding to substrates, and (ii) indirectly, by reducing their adsorption on soil surfaces. In addition, the availability of water influences the interactions between phosphatases and soil organic matter and mineral surfaces, hence controlling the catalysis and adsorption processes. The lack of water availability leads to diffusion limitations, which, in turn, affect the hydrolysis of soil Po by phosphatases, and thereby facilitate their adsorption to the soil matrix.
The exudation of metabolic products from plant roots (e.g., mucilage, LMWOAs) increases soil P availability through alteration of soil physicochemical conditions. The hydraulic and physicochemical effects of mucilage and LMWOAs influence the process of soil P acquisition by phosphatase activity, and the mobilization of insoluble P compounds. Based on our conceptual framework, root exudation serves as a controlling factor that regulates the acquisition and restoration of P within the soil matrix and is responsible for driving the two phases of Po-phosphatase interactions. This conceptual framework provides both theoretical- and process-based insights into the dynamics of soil P. Phosphatases are secreted by plant roots to meet their requirement for P, particularly when dissolved Pi is insufficient in the soil. The spatial and temporal shift between P acquisition and restoration affects the dynamics of P, implying a demand-driven strategy for resource acquisition by plants. As root exudation is largely controlled by plants, it is often poorly understood in its regulation, which makes it difficult to predict the transition from an acquiring phase I to a restoring phase II. To develop a detailed quantitative description and a reliable model of soil phosphatase activity, and thus Po and Pi dynamics, future studies are needed that investigate the spatiotemporal heterogeneity of both phases of soil P cycling.
Author Contributions
AM contributed to the conceptualization and drafting of this review manuscript. MD, SL, and EB provided critical feedback, improved and further developed a concept, suggested revisions, and approved the manuscript. All authors contributed to the article and approved the submitted version.
Funding
AM acknowledges support from the Open Access Publication Funds of the Göttingen University. EB acknowledges support from the priority program 2089 Rhizosphere Spatiotemporal Organization–A Key to Rhizosphere Functions funded by the Deutsche Forschungsgemeinschaft (DFG, German Research Foundation) (Project No. 403664478). The Robert Bosch Foundation provided funding for MD and part of the publication fee via the Junior Professorship 2017 grant.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Adeleke, R., Cloete, T. E., and Khasa, D. P. (2012). Culturable microorganisms associated with Sishen iron ore and their potential roles in biobeneficiation. World J. Microbiol. Biotechnol. 28, 1057–1070. doi: 10.1007/s11274-011-0904-2
Ahmed, M. A., Sanaullah, M., Blagodatskaya, E., Mason-Jones, K., Jawad, H., Kuzyakov, Y., et al. (2018). Soil microorganisms exhibit enzymatic and priming response to root mucilage under drought. Soil Biol. Biochem. 116, 410–418. doi: 10.1016/j.soilbio.2017.10.041
Ahn, M. Y., Martínez, C. E., Archibald, D. D., Zimmerman, A. R., Bollag, J.–M., and Dec, J. (2006). Transformation of catechol in the presence of a laccase and birnessite. Soil Biol. Biochem. 38, 1015–1020. doi: 10.1016/j.soilbio.2005.08.016
Ajmera, I., Hodgman, T. C., and Lu, C. (2019). An integrative systems perspective on plant phosphate research. Genes (Basel). 13, 139. doi: 10.3390/genes10020139
Ali, R. S., Ingwersen, J., Demyan, M. S., Funkuin, Y. N., Wizemann, H., Kandeler, E., et al. (2015). Modelling in situ activities of enzymes as a tool to explain seasonal variation of soil respiration from agro–ecosystems. Soil Biol. Biochem. 81, 291–303. doi: 10.1016/j.soilbio.2014.12.001
Allison, S. D., Gartner, T., Holland, K., Weintraub, M., and Sinsabaugh, R. (2007). “Soil enzymes: linking proteomics and ecological processes,” in: Man. Environ. Microbiol. Hurst, C., Crawford, R., Garland, J., Lipson, D., Mills, A., Stetzenbach, L. (Eds.) p. 704e711. doi: 10.1128/9781555815882.ch58
Allison, S. D., Weintraub, M. N., Gartner, T. B., and Waldrop, M. P. (2011). “Evolutionary–economic principles as regulators of soil enzyme production and ecosystem function”, in Soil Enzymology. Soil Biology Shukla G., Varma A. (eds). Berlin, Heidelberg: Springer. p. 229–243. doi: 10.1007/978-3-642-14225-3_12
Allison, V. J., Condron, L. M., Peltzer, D. A., Richardson, S. J., and Turner, B. L. (2007). Changes in enzyme activities and soil microbial community composition along carbon and nutrient gradients at the Franz Josef chronosequence, New Zealand. Soil Biol. Biochem. 39, 1770e1781. doi: 10.1016/j.soilbio.2007.02.006
Andrade, F. V., Mendonca, E. E., and Silva, I. R. (2013). Organic acid adsorption and mineralization in oxisols with different textures. Rev. Bras. Cienc. Solo 37, 976–985. doi: 10.1590/S0100-06832013000400015
Araujo, A., Leite, L., De–Iwata, B., De–Lira, M., Xavier, G., and Do–Figueiredo, M. (2012). Microbiological process in agroforestry systems. A review. Agron. Sustainable Dev. 32, 215–226. doi: 10.1007/s13593-011-0026-0
Aziz, T., Steffens, D., and Rahmatullah, Schubert, S. (2011). Variation in phosphorus efficiency among brassica cultivars II: Changes in root morphology and carboxylate exudation. J. Plant Nutr. 34, 2127–2138. doi: 10.1080/01904167.2011.618573
Badri, D. V., and Vivanco, J. M. (2009). Regulation and function of root exudates. Plant Cell Environ. 32, 666–681. doi: 10.1111/j.1365-3040.2009.01926.x
Bais, H. P., Broeckling, C. D., and Vivanco, J. M. (2008). “Root exudates modulate plant–microbe interactions in the rhizosphere,” in Secondary Metabolites in Soil Ecology. Soil Biol, Karlovsky P. (eds). doi: 10.1007/978-3-540-74543-3_11
Benard, P., Zarebanadkouki, M., Brax, M., Kaltenbach, R., Jerjen, I., Marone, F., et al. (2019b). Microhydrological niches in soils: how mucilage and EPS alter the biophysical properties of the rhizosphere and other biological hotspots. Vadose Zone. J. 18, 1–10. doi: 10.2136/vzj2018.12.0211
Benard, P., Zarebanadkouki, M., and Carminati, A. (2019a). Physics and hydraulics of the rhizosphere network. J. Plant Nutr. Soil Sci. 182, 5–8. doi: 10.1002/jpln.201800042
Bhadoria, P. B. S., Kaselowsky, J., Claassen, N., and Jungk, A. (1991). Phosphate diffusion coefficients in soil as affected by bulk density and water content. Zeitschrift für Pflanzenernährung und Bodenkunde. 154, 53–57. doi: 10.1002/jpln.19911540111
Bhattacharyya, P., Tripathy, S., Kim, K., and Kim, S.-H. (2008). Arsenic fractions and enzyme activities in arsenic-contaminated soils by groundwater irrigation in West Bengal. Ecotoxicol. Environ. Saf. 71, 149–156. doi: 10.1016/j.ecoenv.2007.08.015
Bhatti, J. S., Comerford, N. B., and Johnston, C. T. (1998). Influence of soil organic matter removal and pH on oxalate sorption onto a spodic horizon. Soil Sci. Soc. Am. J. 62, 152–158. doi: 10.2136/sssaj1998.03615995006200010020x
Bilyera, N., Hummel, C., Daudin, G., Santangeli, M., Zhang, X., Santner, J., et al. (2022). Co–localised phosphorus mobilization processes in the rhizosphere of field-grown maize jointly contribute to plant nutrition. Soil Biol. Biochem. 165, 108497. doi: 10.1016/j.soilbio.2021.108497
Brax, M., Buchmann, C., Kenngott, K., Schaumann, G. E., and Diehl, D. (2020). Influence of the physico-chemical properties of root mucilage and model substances on the microstructural stability of sand. Biogeochemistry 147, 35–52. doi: 10.1007/s10533-019-00626-w
Brucker, E., Kernchen, S., and Spohn, M. (2020). Release of phosphorus and silicon from minerals by soil microorganisms depends on the availability of organic carbon. Soil Biol. Biochem. 143, 107737. doi: 10.1016/j.soilbio.2020.107737
Burns, R. G. (1982). Enzyme activity in soil: location and a possible role in microbial ecology. Soil Biol. Biochem. 14, 423–427. doi: 10.1016/0038-0717(82)90099-2
Burns, R. G., DeForest, J. L., Marxsen, J., Sinsabaugh, R. L., Stromberger, M. E., Wallenstein, M. D., et al. (2013). Soil enzymes in a changing environment: current knowledge and future directions. Soil Biol. Biochem. 58, 216–234. doi: 10.1016/j.soilbio.2012.11.009
Canarini, A., Kaiser, C., Merchant, A., Richter, A., and Wanek, W. (2019). Root exudation of primary metabolites: mechanisms and their roles in plant responses to environmental stimuli. Front. Plant Sci. 10, 157. doi: 10.3389/fpls.2019.00157
Čapek, P., Kasanke, C. P., and Starke, R. (2021). Biochemical inhibition of acid phosphatase activity in two mountain spruce forest soils. Biol Fertil Soils 57, 991–1005. doi: 10.1007/s00374-021-01587-9
Carminati, A., Benard, P., Ahmed, M. A., and Zarebanadkouki, M. (2017). Liquid bridges at the root–soil interface. Plant Soil. 417, 1–15. doi: 10.1007/s11104-017-3227-8
Carminati, A., Moradi, A. B., Vetterlein, D., Vontobel, P., Lehmann, E., Weller, U., et al. (2010). Dynamics of soil water content in the rhizosphere. Plant Soil 332, 163–176. doi: 10.1007/s11104-010-0283-8
Carminati, A., Zarebanadkouki, M., Kroener, E., Ahmed, M. A., and Holz, M. (2016). Biophysical rhizosphere processes affecting root water uptake. Ann. Bot. (Oxford, U. K.). 118, 561–571. doi: 10.1093/aob/mcw113
Chen, C. R., Condron, L. M., Davis, M. R., and Sherlock, R. R. (2002). Phosphorus dynamics in the rhizosphere of perennial ryegrass (Lolium perenne L.) and radiata pine (Pinus Radiata D. Don.). Soil Biol. Biochem. 34, 487–499. doi: 10.1016/S0038-0717(01)00207-3
Clarholm, M., Skyllberg, U., and Rosling, A. (2015). Organic acid induced release of nutrients from metal–stabilized soil organic matter–The unbutton model. Soil Biol. Biochem. 84, 168–176. doi: 10.1016/j.soilbio.2015.02.019
Cornish–Bowden, A. (2015): One hundred years of michaelis-menten kinetics. Perspectives in Sci. 4, 3–9. doi: 10.1016/j.pisc.2014.12.002.
Crawford, J. W., Harris, J. A., Ritz, K., and Young, I. M. (2005). Towards an evolutionary ecology of life in soil. Trends Ecol Evol. 20, 81–87. doi: 10.1016/j.tree.2004.11.014
Dalal, R. C. (1977). Soil organic phosphorus. Adv. Agron. 29, 83–117. doi: 10.1016/S0065-2113(08)60216-3
Dalling, J. W., Heineman, K., Lopez, O. R., Wright, S. J., and Turner, B. L. (2016). “Nutrient availability in tropical rain forests: the paradigm of phosphorus limitation,” in Tropical Tree Physiology, Goldstein, G., and Santiago, L. S. (eds). Cham: Springer. p. 261–273. doi: 10.1007/978-3-319-27422-5_12
Datta, R., Anand, S., Moulick, A., Baraniya, D., Pathan, S. I., Rejsek, K., et al. (2017). How enzymes are adsorbed on soil solid phase and factors limiting its activity: A review. Int. Agrophys. 31, 287–302. doi: 10.1515/intag-2016-0049
De Schepper, V., De Swaef, T., Bauweraerts, I., and Steppe, K. (2013). Phloem transport: a review of mechanisms and controls. J. Exp. Bot. 64, 4839–4850. doi: 10.1093/jxb/ert302
Delhaize, E., James, R. A., and Ryan, P. R. (2012). Aluminium tolerance of root hairs underlies genotypic differences in rhizosheath size of wheat (Triticum aestivum) grown on acid soil. New Phytol. 195, 609–619. doi: 10.1111/j.1469-8137.2012.04183.x
Demanèche, S., Chapel, J.-P., Monrozier, L. J., and Quiquampoix, H. (2009). Dissimilar pH-dependent adsorption features of bovine serum albumin and α-chymotrypsin on mica probed by AFM. Colloids Surf. B. 70, 226–231. doi: 10.1016/j.colsurfb.2008.12.036
Deng, S. P., and Tabatabai, M. A. (1995). Cellulase activity of soils: effect of trace elements. Soil Biol. Biochem. 27, 977–979. doi: 10.1016/0038-0717(95)00005-Y
Denton, M. D., Veneklaas, E. J., Freimoser, F. M., and Lambers, H. (2007). Banksia species (Proteaceae) from severely phosphorus impoverished soils exhibit extreme efficiency in the use and re-mobilization of phosphorus. Plant, Cell Environ. 30, 1557–1565. doi: 10.1111/j.1365-3040.2007.01733.x
Dick, W. A., and Tabatabai, M. A. (1987). Kinetics and activities of phosphatase–clay complexes1. Soil Sci. 5–15. doi: 10.1097/00010694-198701000-00002
Doan, T. H., Doan, T. A., Kangas, M. J., Ernest, A. E., Tran, D., Wilson, C. L., et al. (2017). A low–cost imaging method for the temporal and spatial colorimetric detection of free amines on maize root surfaces. Front. Plant Sci. 8, 1513. doi: 10.3389/fpls.2017.01513
Eberwein, J., Shen, W., and Jenerette, G. D. (2017). Michaelis–Menten kinetics of soil respiration feedbacks to nitrogen deposition and climate change in subtropical forests. Sci. Rep. 7, 1752. doi: 10.1038/s41598-017-01941-8
Gang, X., Hongbo, S., Rongfu, X., Nie, Y., Pei, Y., Sun, Z., et al. (2012). The role of root-released organic acids and anions in phosphorus transformations in a sandy loam soil from Yantai, China. Afr. J. Microbiol. Res. 6, 674–679. doi: 10.5897/AJMR11.1296
Geelhoed, J. S., Hiemstra, T., and Van Riemsdijk, W. H. (1998). Competition interaction between phosphate and citrate on goethite. Environ. Sci. Technol. 32, 2119–2123. doi: 10.1021/es970908y
George, T. S., Brown, L. K., Ramsay, L., White, P. H., Newton, A. C., Bengough, A. G., et al. (2014). Understanding the genetic control and physiological traits associated with rhizosheath production by barley (Hordeum vulgare). New Phytol. 203, 195–205. doi: 10.1111/nph.12786
George, T. S., Gregory, P. J., Wood, M., Read, D., and Buresh, R. J. (2002). Phosphatase activity and organic acids in the rhizosphere of potential agroforestry species and maize. Soil Biol. Biochem. 34, 1487–1494. doi: 10.1016/S0038-0717(02)00093-7
Gianfreda, L. (2015). Enzymes of importance to rhizosphere processes. J. Soil Sci. Plant Nutr. 15, 283–306. doi: 10.4067/S0718-95162015005000022
Gianfreda, L., and Ruggiero, P. (2006). “Enzyme activities in soil,” in Nucleic acids and proteins in soil. Series Soil Biology Nannipieri, P., and Smalla, K. (eds). Berlin, Germany: Springer-Verlag. p. 257–311. doi: 10.1007/3-540-29449-X_12
Gilliham, M., and Tyerman, S. D. (2016). Linking metabolism to membrane signaling: the GABA-Malate connection. Trends Plant Sci. 21, 295–301. doi: 10.1016/j.tplants.2015.11.011
Grafe, M., Goers, M., von Tucher, S., Baum, C., Zimmer, D., Leinweber, P., et al. (2018). Bacterial potentials for uptake, solubilization and mineralization of extracellular phosphorus in agricultural soils are highly stable under different fertilization regimes. Environ. Microbiol. Rep. 10, 320–327. doi: 10.1111/1758-2229.12651
Guber, A., Blagodatskaya, E., and Kravchenko, A. (2022). Are enzymes transported in soils by water fluxes? Soil Biol. Biochem. 168, 108633. doi: 10.1016/j.soilbio.2022.108633
Haque, N. A., and Dave, S. R. (2005). Ecology of phosphate solubilizers in semi-arid agricultural soils. Indian J. Microbiol. 45, 27–32
Hengge, A. C. (2005). Mechanistic studies on enzyme–catalyzed phosphoryl transfer. Adv. Phys. Org. Chem. 40, 49–108. doi: 10.1016/S0065-3160(05)40002-7
Hill, P. W., Garnett, M. H., Farrar, J., Iqbal, Z., Khalid, M., Soleman, N., et al. (2015). Living roots magnify the response of soil organic carbon decomposition to temperature in temperate grassland. GCB Bioenergy. 21, 1368–1375. doi: 10.1111/gcb.12784
Hinsinger, P., Bravin, M. N., Devau, N., Gerard, F., Le Cadre, E., and Jaillard, B. (2008). Soil–root–microbe interactions in the rhizosphere – a key to understanding and predicting nutrient bioavailability to plants. J. Soil Sci. Plant Nutr. 8, 39–47. doi: 10.4067/S0718-27912008000400008
Holz, M., Zarebanadkouki, M., Carminati, A., Hovind, J., Kaestner, A., and Spohn, M. (2019). Increased water retention in the rhizosphere allows for high phosphatase activity in drying soil. Plant Soil. 443, 259–271. doi: 10.1007/s11104-019-04234-3
Hu, L., Bi, A., Hu, Z., Amombo, E., Li, H., and Fu, J. (2018). Antioxidant metabolism, photosystem II, and fatty acid composition of two tall fescue genotypes with different heat tolerance under high temperature stress. Front. Plant Sci. 9, 1242. doi: 10.3389/fpls.2018.01242
Huang, Q., Liang, W., and Cai, P. (2005). Adsorption, desorption and activities of acid phosphatase on various colloidal particles from an Ultisol. Colloids Surf., B. 45, 209–214. doi: 10.1016/j.colsurfb.2005.08.011
Huang, Q., Zhao, Z., and Chen, W. (2003). Effects of several low-molecular weight organic acids and phosphate on the adsorption of acid phosphatase by soil colloids and minerals. Chemosphere. 52, 571–579. doi: 10.1016/S0045-6535(03)00238-8
Hummel, C., Boitt, G., Santner, J., Lehto, N. J., Condron, L., and Wenzel, W. W. (2021). Co–occurring increased phosphatase activity and labile P depletion in the rhizosphere of Lupinus angustifolius assessed with a novel, combined 2D–imaging approach. Soil Biol. Biochem. 153, 107963. doi: 10.1016/j.soilbio.2020.107963
Hunter, P. J., Teakle, G. R., and Bending, G. D. (2014). Root traits and microbial community interactions in relation to phosphorus availability and acquisition, with particular reference to Brassica. Front. Plant Sci. 5, 1. doi: 10.3389/fpls.2014.00027
Hussain, A., Murtaza, G., Ghafoor, A., and Mehdi, S. M. (2012). Use of two–surface Langmuir-type equations for assessment of phosphorus requirements of lentil on differently textured alluvial soils. Comm. Soil Sci. Plant Anal. 43, 2575–2589. doi: 10.1080/00103624.2012.716121
Ikeda, T., and Kuroda, A. (2011). Why does the silica-binding protein “Si-tag” bind strongly to silica surfaces? Implications of conformational adaptation of the intrinsically disordered polypeptide to solid surfaces. Colloids Surf., B. 86, 359–363. doi: 10.1016/j.colsurfb.2011.04.020
Jarosch, K. A., Kandeler, C., Frossard, E., and Bünemann, E. K. (2019). Is the enzymatic hydrolysis of soil organic phosphorus compounds limited by enzyme or substrate availability? Soil Biol. Biochem. 139, 107628. doi: 10.1016/j.soilbio.2019.107628
Joner, E. J., and Johansen, A. (2000). Phosphatase activity of external hyphae of two arbuscular mycorrhizal fungi. Mycol. Res. 104, 81–86. doi: 10.1017/S0953756299001240
Jones, D. L., Nguyen, C., and Finlay, R. D. (2009). Carbon flow in the rhizosphere: carbon trading at the soil–root interface. Plant Soil 321, 5–33. doi: 10.1007/s11104-009-9925-0
Kafkafi, U., Bar–Yosef, B., Rosenberg, R., and Sposito, G. (1988). Phosphorus adsorption by kaolinite and montmorillonite: II. organic anion competition. Soil Sci. Soc. Am. J. 52, 1585–1589. doi: 10.2136/sssaj1988.03615995005200060012x
Kedi, B., Abadie, J., Sei, J., Quiquampoix, H., and Staunton, S. (2013). Diversity of adsorption affinity and catalytic activity of fungal phosphatases adsorbed on some tropical soils. Soil Biol. Biochem. Vol. 56, 13–20. doi: 10.1016/j.soilbio.2012.02.006
Kelleher, B. P., Willeford, K. O., Simpson, A. J., Simpson, M. J., Stout, R., Rafferty, R., et al. (2004). Acid phosphatase interactions with organo-mineral complexes: influence on catalytic activity. Biogeochemistry. 71, 285–297. doi: 10.1023/B:BIOG.0000049348.53070.6f
Khademi, Z., Jones, D. L., Malakouti, M. J., Asadi, F., and Ardebili, M. (2009). Organic acid mediated nutrient extraction efficiency in three calcareous soils. Aust. J. Soil Res. 47, 213–220. doi: 10.1071/SR07179
Kirk, G. J. D., Santos, E. E., and Santos, M. B. (1999). Phosphate solubilization by organic anion excretion from rice growing in aerobic soil: rates of excretion and decomposition, effects on rhizosphere pH and effects on phosphate solubility and uptake. New Phytol. 142, 185–200. doi: 10.1046/j.1469-8137.1999.00400.x
Koester, M., Stock, S. C., Nájera, F., Abdallah, K., Gorbushina, A., Prietzel, J., et al. (2021). From rock eating to vegetarian ecosystems — disentangling processes of phosphorus acquisition across biomes. Geoderma. 388, 114827. doi: 10.1016/j.geoderma.2020.114827
Korenblum, E., Dong, Y., Szymanski, J., Panda, S., Jozwiak, A., Massalha, H., et al. (2020). Rhizosphere microbiome mediates systemic root metabolite exudation by root-to-root signaling. PNAS. 117, 3874–3883. doi: 10.1073/pnas.1912130117
Kroener, E., Zarebanadkouki, M., Kaestner, A., and Carminati, A. (2014). Nonequilibrium water dynamics in the rhizosphere: How mucilage affects water flow in soils. Water Resour. Res. 50, 6479–6495. doi: 10.1002/2013WR014756
Kruse, J., Abraham, M., Amelung, W., Baum, C., Bol, R., Kühn, O., et al. (2015). Innovative methods in soil phosphorus research: a review. J. Plant Nutr. Soil Sci. 178, 43–88. doi: 10.1002/jpln.201400327
Lambers, H., Hayes, P. E., Laliberté, E., Oliveira, R. S., and Turner, B. L. (2015). Leaf manganese accumulation and phosphorus-acquisition efficiency. Tren. Plant Sci. 20, 83–90. doi: 10.1016/j.tplants.2014.10.007
Lang, F., Krüger, J., Amelung, W., Willbold, S., Frossard, E., Bünemann, E. K., et al. (2017). Soil phosphorus supply controls P nutrition strategies of beech forest ecosystems in Central Europe. Biogeochemistry. 136, 5–29. doi: 10.1007/s10533-017-0375-0
Lassila, J. K., Zalatan, J. G., and Herschlag, D. (2011). Biological phosphoryl-transfer reactions: understanding mechanism and catalysis. Annu Rev Biochem. 80, 669–702. doi: 10.1146/annurev-biochem-060409-092741
Leake, J. R., and Miles, W. (1996). Phosphodiesters as mycorrhizal P sources I. phosphodiesterase production and the utilization of DNA as a phosphorus source by the ericoid mycorrhizal fungus Hymenoscyphus ericae. New Phytol. 132, 435–443. doi: 10.1111/j.1469-8137.1996.tb01863.x
Leprince, F., and Quiquampoix, H. (1996). Extracellular enzyme activity in soil: effect of pH and ionic strength on the interaction with montmorillonite of two acid phosphatases secreted by the ectomycorrhizal fungus Hebeloma cylindrosporum. Eur. J. Soil Sci. 47, 511–522. doi: 10.1111/j.1365-2389.1996.tb01851.x
Loeppmann, S., Blagodatskaya, E., Pausch, J., and Kuzyakov, Y. (2016b). Substrate quality affects kinetics and catalytic efficiency of exo–enzymes in rhizosphere and detritusphere. Soil Biol. Biochem. Vol. 92, 111–118. doi: 10.1016/j.soilbio.2015.09.020
Loeppmann, S., Breidenbach, A., Spielvogel, S., Dippold, M. A., and Blagodatskaya, E. (2020). Organic nutrients induced coupled C– and P–cycling enzyme activities during microbial growth in forest soils. Front. For. Glob. Change 3, 100. doi: 10.3389/ffgc.2020.00100
Loeppmann, S., Semenov, M., Blagodatskaya, E., and Kuzyakov, Y. (2016a). Substrate quality affects microbial– and enzyme activities in rooted soil. J. Plant Nut. Soil Sci. 179, 39–47. doi: 10.1002/jpln.201400518
Loeppmann, S., Semenov, S., Kuzyakov, Y., and Blagodatskaya, E. (2018), Shift from dormancy to microbial growth revealed by RNA:DNA ratio. Ecol. Indic. 85, 603–612. doi: 10.1016/j.ecolind.2017.11.020
Lu, X. C., Mahdi, A. K., Han, X. Z., Chen, X., Yan, J., Biswas, A., et al. (2020). Long–term application of fertilizer and manures affect P fractions in Mollisol. Sci. Rep. 10. doi: 10.1038/s41598-020-71448-2
Manzoni, S., Ding, Y., Warren, C. R., Banfield, C. C., Dippold, M. A., and Mason–Jones, K. (2021). Intracellular storage reduces stoichiometric imbalances in soil microbial biomass – a theoretical exploration. Front. Ecol. Evol. 663. doi: 10.3389/fevo.2021.714134
Manzoni, S., Schaeffer, S. M., Katul, G., Porporato, A., and Schimel, J. P. (2014). A theoretical analysis of microbial eco–physiological and diffusion limitations to carbon cycling in drying soils. Soil Biol. Biochem. 73, 69–83. doi: 10.1016/j.soilbio.2014.02.008
Margalef, O., Sardans, J., Fernández–Martínez, M., Molowny–Horas, R., Janssens, I. A., Ciais, P., et al. (2017). Global patterns of phosphatase activity in natural soils. Sci. Rep. 7, 1337. doi: 10.1038/s41598-017-01418-8
Marklein, A. R., and Houlton, B. Z. (2012). Nitrogen inputs accelerate phosphorus cycling rates across a wide variety of terrestrial ecosystems. New Phytol. 193, 696–704. doi: 10.1111/j.1469-8137.2011.03967.x
Maseko, S. T., and Dakora, F. D. (2013). Rhizosphere acid and alkaline phosphatase activity as a marker of Pb nutrition in nodulated Cyclopia and Aspalathus species in the Cape fynbos of South Africa. South African J. Bot. 89, 289–295. doi: 10.1016/j.sajb.2013.06.023
McConnell, C. A., Kaye, J. P., and Kemanian, A. R. (2020). Reviews and syntheses: Ironing out wrinkles in the soil phosphorus cycling paradigm. Biogeosciences. 5309–5333. doi: 10.5194/bg-17-5309-2020
McCully, M. E., and Boyer, J. S. (1997). The expansion of maize root-cap mucilage during hydration. 3. Changes in water potential and water content. Physiol. Plant. 99, 169–177. doi: 10.1111/j.1399-3054.1997.tb03445.x
Menezes–Blackburn, D., Jorquera, M. A., Greiner, R., Gianfreda, L., and Dela, L. M. M. (2013). Phytases and phytase-labile organic phosphorus in manures and soils. Crit. Rev. Environ. Sci. Technol. 43, 916–954. doi: 10.1080/10643389.2011.627019
Mengel, K., Kirkby, E. A., Kosegarten, H., and Appel, T. (2001). “Phosphorus,” in Principles of Plant Nutrition, Mengel K., Kirkby E.A., Kosegarten H., Appel T. (eds). Dordrecht: Springer. doi: 10.1007/978-94-010-1009-2
Merlin, A., Rosolem, C. A., and He, Z. (2016). Non-labile phosphorus acquisition by Brachiaria. J. Plant Nutr. 39, 1319–1327. doi: 10.1080/01904167.2015.1109117
Mora–Macías, J., Ojeda–Rivera, J. O., Gutiérrez–Alanís, D., Yong–Villalobos, L., Oropeza–Aburto, A., Raya–González, J., et al. (2017). Malate–dependent Fe accumulation is a critical checkpoint in the root developmental response to low phosphate. Proc. Natl. Acad. Sci. 114, E3563–E3572. doi: 10.1073/pnas.1701952114
Moscatelli, M. C., Lagomarsino, A., De Angelis, P., and Grego, S. (2005). Seasonality of soil biological properties in a poplar plantation growing under elevated atmospheric CO2. Appl. Soil Ecol. 30, 162–173. doi: 10.1016/j.apsoil.2005.02.008
Nannipieri, P., Ceccanti, B., and Bianchi, D. (1988). Characterization of humus–phosphatase complexes extracted from soil. Soil Biol. Biochem. 20, 683–691. doi: 10.1016/0038-0717(88)90153-8
Nannipieri, P., Giagnoni, L., Landi, L., and Renella, G. (2011). “Role of phosphatase enzymes in soil,” in Phosphorus in Action. Springer, Berlin 215–243. doi: 10.1007/978-3-642-15271-9_9
Noll, L., Zhang, S., Zheng, Q., Hu, Y., and Wanek, W. (2019). Wide–spread limitation of soil organic nitrogen transformations by substrate availability and not by extracellular enzyme content. Soil Biol. Biochem. 133, 37–49. doi: 10.1016/j.soilbio.2019.02.016
Nuruzzaman, M., Lambers, H., and Bolland, M. D. A. (2006). Distribution of carboxylates and acid phosphatase and depletion of different phosphorus fractions in the rhizosphere of a cereal and three grain legumes. Plant Soil 281, 109–120. doi: 10.1007/s11104-005-3936-2
Nwoke, O. C., Diels, J., Abaidoo, R., Nziguheba, G., and Merckx, R. (2008). Organic acids in the rhizosphere and root characteristics of soybean (Glycine max) and cowpea (Vigna unguiculata) in relation to phosphorus uptake in poor savannah soils. Afr. J. Biotechnol. 7, 3620–3627.
Oburger, E., and Jones, D. L. (2018). Sampling root exudates – mission impossible? Rhizosphere 6, 116–133. doi: 10.1016/j.rhisph.2018.06.004
Oburger, E., Leitner, D., Jones, D. L., Zygalakis, K. C., Schnepf, A., and Roose, T. (2011). Adsorption and desorption dynamics of citric acid anions in soil. Eur. J. Soil Sci. 62, 733–742. doi: 10.1111/j.1365-2389.2011.01384.x
Oehl, F., Oberson, A., Probst, M., Fliessbach, A., Roth, H. R., and Frossard, E. (2001). Kinetics of microbial phosphorus uptake in cultivated soils. Biol. Fertil. Soils 34, 31–41. doi: 10.1007/s003740100362
Ohno, T., and Crannell, B. S. (1996). Green and animal manure–derived dissolved organic matter effects on phosphorus sorption. J. Environ. Qual. 25, 1137–1143. doi: 10.2134/jeq1996.00472425002500050029x
Pascual, J. A., Moreno, J. L., Hernández, T., and García, C. (2002). Persistence of immobilized and total urease and phosphatase activities in a soil amended with organic wastes. Biores. Tech. 82, 73–78. doi: 10.1016/S0960-8524(01)00127-4
Paul, E. A., and Clark, F. E. (1989). Soil Microbiology and Biochemistry. San Diego: Academic Press. p. 272. doi: 10.1016/B978-0-12-546805-3.50004-7
Pausch, J., Loeppmann, S., Kühnel, A., Forbush, K., Kuzyakov, Y., and Cheng, W. (2016). Rhizosphere priming of barley with and without root hairs. Soil Biol. Biochem. 100, 74–82. doi: 10.1016/j.soilbio.2016.05.009
Pearse, S. J., Veneklaas, E. J., Cawthray, G., Bolland, M. D. A., and Lambers, H. (2007). Carboxylate composition of root exudates does not relate consistently to a crop species' ability to use phosphorus from aluminium, iron or calcium phosphate sources. New Phytol. 173, 181–190. doi: 10.1111/j.1469-8137.2006.01897.x
Pereira, G. C., Hayes, P. E., O'Sullivan, O. S., Weerasinghe, L. K., Clode, P. L., Atkin, O. K., et al. (2019). Trait convergence in photosynthetic nutrient-use efficiency along a 2–million year dune chronosequence in a global biodiversity hotspot. J. Ecol. 107, 2006–2023. doi: 10.1111/1365-2745.13158
Peth, S., Horn, R., Beckmann, F., Donath, T., Fischer, J., and Smucker, A. J. M. (2008). Three–dimensional quantification of intra-aggregate pore-space features using synchrotron–radiation–based microtomography. Soil Sci. Soc. Am. J. 72, 897–907. doi: 10.2136/sssaj2007.0130
Pettridge, J., and Firestone, M. K. (2017). Using stable isotopes to explore root–microbe–mineral interactions in soil. Rhizosphere. 1–10. doi: 10.1016/j.rhisph.2017.04.016
Pistocchi, C., Mészáros, E., Frossard, E., Bünemann, E. K., and Tamburini, F. (2020). In or out of equilibrium? How microbial activity controls the oxygen isotopic composition of phosphate in forest organic horizons with low and high phosphorus availability. Front. Environ. Sci. 8, 1–15. doi: 10.3389/fenvs.2020.564778
Pizzeghello, D., Berti, A., Nardi, S., and Morari, F. (2011). Phosphorus forms and P-sorption properties in three alkaline soils after long-term mineral and manure applications in north-eastern Italy. Agric. Ecosyst. Environ. 141, 58–66. doi: 10.1016/j.agee.2011.02.011
Quiquampoix, H., and Mousain, D. (2005). Enzymatic hydrolysis of organic phosphorus. Organic Phosphorus in the Environment, Turner, B. L. Frossard, E. Baldwin, D. S. (eds). Wallingford, UK: CAB International. p. 89–112. doi: 10.1079/9780851998220.0089
Quiquampoix, H., Servagent-Noinville, S., and Baron, M. -H. (2002). “Enzyme adsorption on soil mineral surfaces and consequences for the catalytic activity,” in Enzymes in the Environment: Activity, Ecology, and Application, eds R. G. Burns and R. P. Dick (New York, NY: Marcel Dekker Inc), 285–306.
Ramesh, S. A., Tyerman, S. D., Xu, B., Bose, J., Kaur, S., Conn, V., et al. (2015). GABA signalling modulates plant growth by directly regulating the activity of plant-specific anion transporters. Nat. Commun. 6, 7879. doi: 10.1038/ncomms8879
Reddy, K. R., Overcash, M. R., Khaleel, R., and Westerman, P. W. (1980). Phosphorus adsorption-desorption characteristics of two soils utilized for disposal of animal wastes, J. Environ. Qual. 9, 86–92. doi: 10.2134/jeq1980.00472425000900010020x
Reed, S. C., Yang, X., and Thornton, P. E. (2015). Incorporating phosphorus cycling into global modeling efforts: a worthwhile, tractable endeavor. New Phytol. 208, 324–329. doi: 10.1111/nph.13521
Rejsek, K., Vranova, V., Pavelka, M., and Formanek, P. (2012). Acid phosphomonoesterase (E.C. 3.1.3.2) location in soil. J Plant Nutr. Soil Sci. 175, 196–211. doi: 10.1002/jpln.201000139
Renella, G., Landi, L., Valori, F., and Nannipieri, P. (2007). Microbial and hydrolase activity after release of low molecular weight organic compounds by a model root surface in a clayey and a sandy soil. Appl. Soil Ecol. 36, 124–129. doi: 10.1016/j.apsoil.2007.01.001
Richardson, A. E., Hocking, P. J., Simpson, R. J., and George, T. S. (2009). Plant mechanisms to optimise access to soil phosphorus. Crop Pasture Sci. 60, 124. doi: 10.1071/CP07125
Rombola, T. H., Pedrinho, E. A. N., de Macedo Lemos, E. G., Gonçalves, A. M., José dos Santos, L. F., and Pizauro, J. J. M. (2014). Identification and enzymatic characterization of acid phosphatase from Burkholderia gladioli. BMC Res. Notes. 7, 221. doi: 10.1186/1756-0500-7-221
Ross–Elliott, T. J., Jensen, K. H., Haaning, K. S., Wager, B. M., Knoblauch, J., Howell, A. H., et al. (2017). Phloem unloading in Arabidopsis roots is convective and regulated by the phloem–pole pericycle. eLife. 6, e24125. doi: 10.7554/eLife.24125.022
Sasse, J., Kant, J., Cole, B. J., Klein, A. P., Arsova, B., Schlaepfer, P., et al. (2019). Multilab EcoFAB study shows highly reproducible physiology and depletion of soil metabolites by a model grass. New Phytol. 222, 1149–1160. doi: 10.1111/nph.15662
Schachtman, D. P., Reid, R. J., and Ayling, S. M. (1998). Phosphorus uptake by plants: from soil to cell. J. Plant Physiol. 116, 447–453. doi: 10.1104/pp.116.2.447
Schimel, J., Balser, T. C. and Wallenstein, M. (2007). Microbial stress-response physiology and its implications for ecosystem function. Ecol. 88, 1386–1394. doi: 10.1890/06-0219
Schimel, J. P., and Weintraub, M. N. (2003). The implications of exoenzyme activity on microbial carbon and nitrogen limitation in soil: a theoretical model. Soil Biol. Biochem. 35, 549–563. doi: 10.1016/S0038-0717(03)00015-4
Shindo, H., Watanabe, D., Onaga, T., Urakawa, M., Nakahara, O., and Huang, Q. (2002). Adsorption, activity, and kinetics of acid phosphatase as influenced by selected oxides and clay minerals. Soil Sci. Plant Nutr. 48, 763–767. doi: 10.1080/00380768.2002.10409268
Sinsabaugh, R. L., and Moorhead, D. L. (1994). Resource allocation to extracellular enzyme production: a model for nitrogen and phosphorus control of litter decomposition. Soil Biol. Biochem. 26, 1305–1311. doi: 10.1016/0038-0717(94)90211-9
Song, F., Han, X., Zhu, X., and Herbert, S. J. (2012). Response to water stress of soil enzymes and root exudates from drought and non-drought tolerant corn hybrids at different growth stages. Can. J. Soil Sci. 92, 501–507. doi: 10.4141/cjss2010-057
Spohn, M., and Kuzyakov, Y. (2013). Phosphorus mineralization can be driven by microbial need for carbon. Soil Biol. Biochem. 61, 69–75. doi: 10.1016/j.soilbio.2013.02.013
Steinweg, J., Dukes, J., Paul, E., and Wallenstein, M. D. (2013). Microbial responses to multi-factor climate change: effects on soil enzymes. Front. Microbiol. 4, 146. doi: 10.3389/fmicb.2013.00146
Strobel, B. W. (2001). Influence of vegetation on low-molecular-weight carboxylic acids in soil solution—a review. Geoderma 99, 169–198. doi: 10.1016/S0016-7061(00)00102-6
Tang, J., and Riley, W. J. (2021). On the modeling paradigm of plant root nutrient acquisition. Plant Soil 459, 441–451. doi: 10.1007/s11104-020-04798-5
Tian, H., Zhao, Y., Megharaj, M., and He, W. (2018). Arsenate inhibition on kinetic characteristics of alkaline phosphatase as influenced by pH. Ecol. Indic. 85, 1101–1106. doi: 10.1016/j.ecolind.2017.11.041
Tietjen, T., and Wetzel, R. G. (2003). Extracellular enzyme–clay mineral complexes: Enzyme adsorption, alteration of enzyme activity, and protection from photodegradation. Aquat. Ecol. 37, 331–339. doi: 10.1023/B:AECO.0000007044.52801.6b
Tinker, P. B., and Nye, P. H. (2000): Solute movement in the rhizosphere. 2nd ed. Oxford UK: Oxford University Press. doi: 10.1093/oso/9780195124927.001.0001
Treseder, K. K., and Vitousek, P. M. (2001). Effects of soil nutrient availability on investment in acquisition of N and P in Hawaiian Rain Forests. Spec. Publ. - Ecol. Soc. Am. 82:946e954. doi: 10.1890/0012-9658(2001)082[0946:EOSNAO]2.0.CO;2
Turner, B. L. (2008). Resource partitioning for soil phosphorus: a hypothesis. J. Ecol. 96, 698–702. doi: 10.1111/j.1365-2745.2008.01384.x
Turner, B. L., and Haygarth, P. M. (2005). Phosphatase activity in temperate pasture soils: Potential regulation of labile organic phosphorus turnover by phosphodiesterase activity. Sci Total Environ. 344, 27–36. doi: 10.1016/j.scitotenv.2005.02.003
Vidal, A., Hirte, J., Bender, S. F., Mayer, J., Gattinger, A., Höschen, C., et al. (2018). Linking 3D soil structure and plant-microbe-soil carbon transfer in the rhizosphere. Front. Environ. Sci. 6, 9 doi: 10.3389/fenvs.2018.00009
Wallenstein, M. D., and Burns, R. G. (2011). “Ecology of extracellular enzyme activities and organic matter degradation in soil: a complex community-driven process,” in Methods of Soil Enzymology, Vol. 9, ed R. P. Dick. doi: 10.2136/sssabookser9.c2
Wang, Y., Krogstad, T., Clarke, N., Øgaard, A. F., and Clarke, J. L. (2017). Impact of phosphorus on rhizosphere organic anions of wheat at different growth stages under field conditions. AoB Plants. 9, plx008. doi: 10.1093/aobpla/plx008
Wang, Y., Zhang, T., Zhao, Y., Ciborowski, J. J. H., Zhao, Y., O'Halloran, I. P., et al. (2021). Characterization of sedimentary phosphorus in lake erie and on-site quantification of internal phosphorus loading. Water Res. 188, 116525. doi: 10.1016/j.watres.2020.116525
Wang, Z., Tian, H., Lu, G., Zhao, Y., Yang, R., Megharaj, M., et al. (2018). Catalytic efficiency is a better predictor of arsenic toxicity to soil alkaline phosphatase. Ecotoxicol. Environ. Saf. 148, 721–728. doi: 10.1016/j.ecoenv.2017.11.040
Wang, Z. Q., Li, Y. B., Tan, X. P., He, W. X., Xie, W., Megharaj, M., et al. (2017). Effect of arsenate contamination on free, immobilized and soil alkaline phosphatases: activity, kinetics and thermodynamics. Eur J Soil Sci. 68, 126–135. doi: 10.1111/ejss.12397
Weintraub, M. N., and Schimel, J. P. (2005). The seasonal dynamics of amino acids and other nutrients in Alaskan Arctic tundra soils. Biogeochemistry 73, 359e380. doi: 10.1007/s10533-004-0363-z
Weisskopf, L., Abou-Mansour, E., Fromin, N., Tomasi, N., Santelia, D., Edelkott, I., et al. (2006). White lupin has developed a complex strategy to limit microbial degradation of secreted citrate required for phosphate acquisition. Plant, Cell Environ. 29, 919–927. doi: 10.1111/j.1365-3040.2005.01473.x
Williams, A., and de Vries, F. T. (2019). Plant root exudation under drought: implications for ecosystem functioning. New Phytol. 5, 1899–1905. doi: 10.1111/nph.16223
Williams, M. A., and Rice, C. W. (2007). Seven years of enhanced water availability influences the physiological, structural, and functional attributes of a soil microbial community. Appl. Soil Ecol. 35, 535–545. doi: 10.1016/j.apsoil.2006.09.014
Zarebanadkouki, M., Trtik, P., Hayat, F., Carminati, A., and Kaestner, A. (2019). Root water uptake and its pathways across the root: quantification at the cellular scale. Sci. Rep. 9, 12979. doi: 10.1038/s41598-019-49528-9
Zhang, L., Ding, X., Peng, Y., George, T. S., and Feng, G. (2018). Closing the Loop on Phosphorus Loss from Intensive Agricultural Soil: A Microbial Immobilization Solution? Front. Microbiol. 104. doi: 10.3389/fmicb.2018.00104
Zhao, Z., Jiang, Y., Mi, T., Ling, W., Huang, Q., and Xia, L. (2012). Effects of Addition Sequence and pH of Oxalate on the Adsorption and Activity of Acid Phosphatase on Soil Colloids and Minerals. Commun. Soil Sci. Plant Anal. 43, 1436–1448. doi: 10.1080/00103624.2012.670343
Zhu, Y., Wu, F., Feng, W., Liu, S., and Giesy, J. P. (2016). Interaction of alkaline phosphatase with minerals and sediments: Activities, kinetics and hydrolysis of organic phosphorus. Colloid Surf. Physicochem. Eng. Asp. 495, 46–53. doi: 10.1016/j.colsurfa.2016.01.056
Zhuravlev, L. (2000). The surface chemistry of Amorphous silica. Zhuravlev model. Colloid Surf. Physicochem. Eng. Asp 173, 1–38. doi: 10.1016/S0927-7757(00)00556-2
Zickenrott, I.–M., Woche, S. K., Bachmann, J., Ahmed, M. A., and Vetterlein, D. (2016). An efficient method for the collection of root mucilage from different plant species-a case study on the effect of mucilage on soil water repellency. J. Plant Nutr. Soil Sci. 179, 294–302. doi: 10.1002/jpln.201500511
Keywords: phosphatase-soil interactions, substrate catalysis, root exudation, mucilage, LMWOAs, phosphatase adsorption
Citation: Manzoor A, Dippold MA, Loeppmann S and Blagodatskaya E (2022) Two-Phase Conceptual Framework of Phosphatase Activity and Phosphorus Bioavailability. Front. Plant Sci. 13:935829. doi: 10.3389/fpls.2022.935829
Received: 04 May 2022; Accepted: 15 June 2022;
Published: 19 July 2022.
Edited by:
Victoria Fernandez, Polytechnic University of Madrid, SpainReviewed by:
Leo Condron, Lincoln University, New ZealandXiangnan Li, Northeast Institute of Geography and Agroecology (CAS), China
Copyright © 2022 Manzoor, Dippold, Loeppmann and Blagodatskaya. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Aamir Manzoor, YWFtaXIubWFuem9vckBzdHVkLnVuaS1nb2V0dGluZ2VuLmRl
†These authors share senior authorship