
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Plant Sci., 18 October 2022
Sec. Plant Symbiotic Interactions
Volume 13 - 2022 | https://doi.org/10.3389/fpls.2022.932311
This article is part of the Research TopicBiostimulants in Agriculture II: Towards a Sustainable FutureView all 78 articles
 Keiji Jindo1
Keiji Jindo1 Travis L. Goron2
Travis L. Goron2 Paloma Pizarro-Tobías3
Paloma Pizarro-Tobías3 Miguel Ángel Sánchez-Monedero4
Miguel Ángel Sánchez-Monedero4 Yuki Audette5,6
Yuki Audette5,6 Ayodeji O. Deolu-Ajayi1Adrie van der Werf1
Ayodeji O. Deolu-Ajayi1Adrie van der Werf1 Misghina Goitom Teklu1
Misghina Goitom Teklu1 Moshe Shenker7
Moshe Shenker7 Cláudia Pombo Sudré8
Cláudia Pombo Sudré8 Jader Galba Busato9
Jader Galba Busato9 Raúl Ochoa-Hueso10,11
Raúl Ochoa-Hueso10,11 Marco Nocentini12Johan Rippen13
Marco Nocentini12Johan Rippen13 Ricardo Aroca14
Ricardo Aroca14 Socorro Mesa14
Socorro Mesa14 María J. Delgado14
María J. Delgado14 Germán Tortosa14*
Germán Tortosa14*Current and continuing climate change in the Anthropocene epoch requires sustainable agricultural practices. Additionally, due to changing consumer preferences, organic approaches to cultivation are gaining popularity. The global market for organic grapes, grape products, and wine is growing. Biostimulant and biocontrol products are often applied in organic vineyards and can reduce the synthetic fertilizer, pesticide, and fungicide requirements of a vineyard. Plant growth promotion following application is also observed under a variety of challenging conditions associated with global warming. This paper reviews different groups of biostimulants and their effects on viticulture, including microorganisms, protein hydrolysates, humic acids, pyrogenic materials, and seaweed extracts. Of special interest are biostimulants with utility in protecting plants against the effects of climate change, including drought and heat stress. While many beneficial effects have been reported following the application of these materials, most studies lack a mechanistic explanation, and important parameters are often undefined (e.g., soil characteristics and nutrient availability). We recommend an increased study of the underlying mechanisms of these products to enable the selection of proper biostimulants, application methods, and dosage in viticulture. A detailed understanding of processes dictating beneficial effects in vineyards following application may allow for biostimulants with increased efficacy, uptake, and sustainability.
Grape (Vitis vinifera) production has increased globally over the last decade from 65 million to 77.8 million tons. The world’s vineyards totaled 7.4 million hectares in 2018, of which ~4.5% were cultivated with organic approaches (i.e., refraining from pesticides and chemical fertilizer use and promoting sustainable cultivation practices). About 88% of organic vineyards are located in Europe, primarily in France, Italy, and Spain. There is a need, and increasing demand, for more ecologically-sustainable agricultural products as outlined by the United Nations Sustainable Development Goals (SDGs). For production to support demand despite current anthropogenic climate change (Roka, 2022), more research on sustainable food systems is urgently needed.
Higher yields are necessary for sustainable food security on a finite land basis (Godfray et al., 2010; Foley et al., 2011). According to Renaud-Gentié (Seufert et al., 2012), the yield ratio between organic and conventional agricultural production per unit area is 0.75. To close this yield gap while simultaneously satisfying consumer preference for organic products, improved techniques for organic grape farming are required.
Additionally, grape varieties often require specific environmental conditions, historically located in very narrow latitudinal zones (30–50°N and 30–40°S) with minimal temperature extremes (Nesbitt et al., 2018). Due to the high environmental sensitivity of Vitis vinifera, a strong correlation between meteorological conditions propelled by global warming (e.g., heat waves, extreme precipitation, droughts, hailstorms, and windstorms) and low grape yield and wine quality has been reported (Santos et al., 2020).
Biostimulants are substances or microorganisms applied to plants, which enhance nutrition use efficiency, abiotic stress tolerance, and crop quality traits, using mechanisms that do not directly add fertility to the soil (du Jardin, 2015), and in the EU the marketing of biostimulants is regulated by specific provisions (Regulation EU 1009, 2019). Initially, these technologies were primarily utilized in organic farming, with later uptake in both conventional and integrated systems (Rouphael and Colla, 2020). The application of biostimulant technologies is now a well-known tool, though poorly-researched strategy to enhance plant growth and prevent pests and diseases (Gutiérrez-Gamboa et al., 2019; Cataldo et al., 2022; Monteiro et al., 2022; Olavarrieta et al., 2022).
Biocontrol agents (BCAs) are organisms antagonistic to crop pests. Synthetic pesticides, mainly fungicides, are widely used in viticulture. However, pesticide residue contamination of grapes and grape products (raisins, juices, and wines) is widely studied and may be detrimental to consumer health (Walorczyk et al., 2011). An EU report (EFSA, 2021) has shown increased pesticide residue levels in primary commodity food products since 2016. Table grape products are of particular concern, in which 66% of products were contaminated, and 14.3% of products tested contained more than five different pesticide residues. In a sample of dried vine fruits, 28 pesticides were detected (Cabrera and Pastor, 2021). When testing grape leaves, 37.9% had residues above the allowed maximum residue levels (MRL). Gava et al. (2021) highlighted the importance of tracing contamination from the originating vineyard to the wine. Of additional concern is that these residues can affect alcoholic fermentation, change product flavor, and pose a toxicological risk to the consumer as well as those involved in the production chain.
This review presents a series of different biostimulant materials and biological control agents utilized in sustainable vineyards (their main effects on Vitis vinifera are summarized in Figure 1). Additionally, the origin of the materials, the application methods, and the underpinning functions/mechanisms of each material are described. The limitations of current practices and alternative approaches to aid future studies of the interaction between viticulture and global climate change are also addressed.
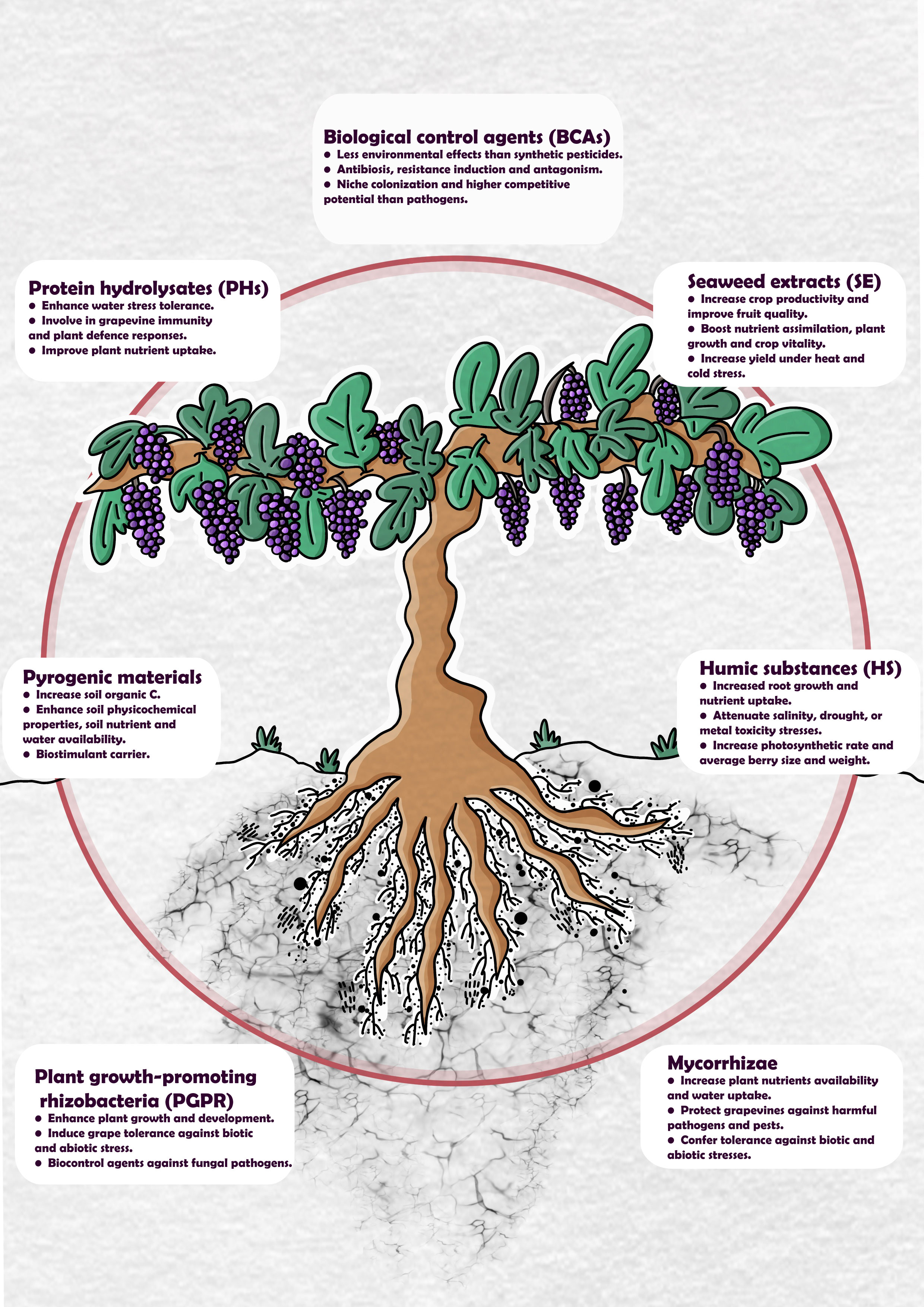
Figure 1 An overview of the main effects of biological control agents (BCAs), protein hydrolysates (PHs), seaweed extracts (SE), pyrogenic materials, humic substances (HS), plant growth-promoting rhizobacteria (PGPR), and mycorrhizae applied in viticulture.
Protein hydrolysates (PHs) refer to mixtures of polypeptides, oligopeptides, and amino acids that are manufactured from protein sources using partial hydrolysis (Schaafsma, 2009). They are produced by either acid and/or alkaline hydrolysis (Schaafsma, 2009), enzymatic hydrolysis (Colla et al., 2015), or from by-products of plant or animal origins (Meggio et al., 2020). In horticulture, more than 90% of PH utilized is derived from animal origin, while plant-derived PHs are relatively novel (Colla et al., 2015). Animal-derived PHs often contain higher amounts of total amino acids compared to plant-derived PHs (Ertani et al., 2009). Plant-derived PHs contain carbohydrates and phenols, which enhance plant oxidative stress defenses, as well as aid in energy metabolism (Colla et al., 2015). PHs can be further subdivided into two categories: (1) protein fractions with a relatively high content of specific amino acids, such as glutamine peptides and cysteine/glycine and tryptophan peptides; and (2) bioactive peptides with specific amino-acid sequences (i.e., 3 to 20 amino acids), which are usually hydrophobic (Schaafsma, 2009). In general, the effects of PHs on plant physiology are closely related to the stimulation of carbon and nitrogen metabolism, hormone activity, and nutrient supply and uptake by plants (Figure 1). Moreover, it has been demonstrated that PHs can stimulate the plant microbiome which can promote plant growth and development by enhancing water and nutrient uptake as well as their adaptation to biotic and abiotic stresses (Colla et al., 2017, and sources reviewed within).
The effects of PHs on grapevine growth are reviewed in Table 1. PHs are known to regulate gene expression involved in the transport of nutrients, and the signaling and metabolism of reactive oxygen species, thereby enhancing plant stress tolerance (Meggio et al., 2020). For example, PHs can effectively improve grapevine tolerance to water deficit (Boselli et al., 2019; Meggio et al., 2020). A study by Boselli et al. (2019) showed that PH-treated grapevines contain higher levels of anthocyanins, often observed under water stress conditions (Castellarin et al., 2007). Therefore, PHs may be able to mimic water stress conditions in grapevine and reduce water loss due to evapotranspiration (Boselli et al., 2019).
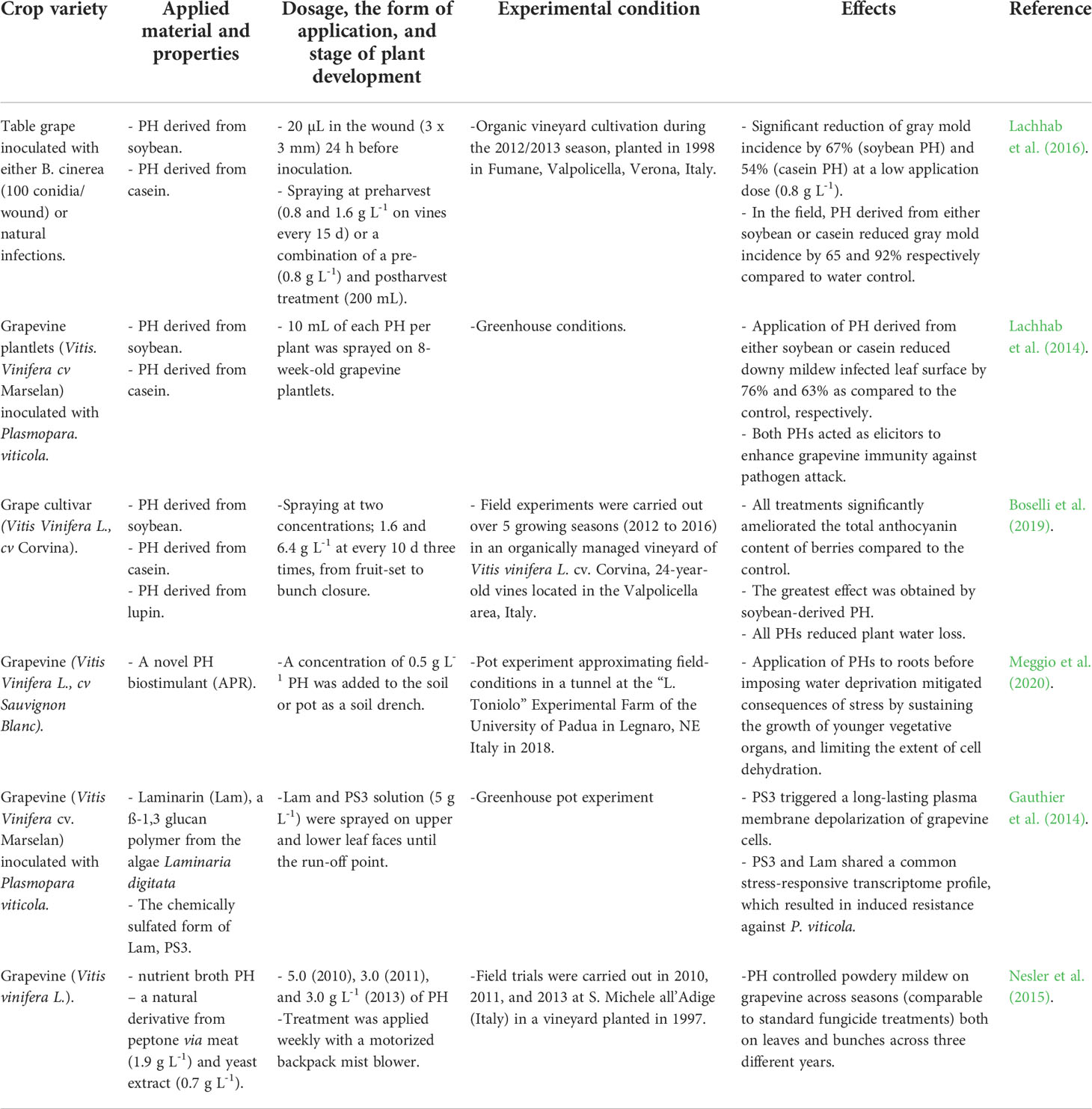
Table 1 An overview of the main effects of protein hydrolysates (PHs) applied in viticulture.
In addition, PHs act as signal compounds to trigger plant defense responses (Boller, 1995). Grapevine is highly susceptible to various pathogens (Lachhab et al., 2014). In particular, downy mildew (Plasmopara viticola) and powdery mildew (Erysiphe necator) are the most widespread and devastating grapevine diseases worldwide (Gauthier et al., 2014; Nesler et al., 2015). Frequent fungicide applications (i.e., every 7-10 days in season) are often required to control these diseases, a practice which not only dramatically increases production costs but also negatively impacts the environment and human health (Jones et al., 2014). Plant defense by-products may be an attractive alternative to chemical pesticides (van Aubel et al., 2014). Elicitors including PHs are not toxic to plants and are recognized by plant membrane receptors, meaning they are often freely able to mobilize an array of plant defenses (Boller and Felix, 2009). PHs contain a large variety of bioactive peptides, which may act as plant growth regulators, antioxidants, and biostimulants (Colla et al., 2014), as well as directly influence numerous biological processes evoking hormonal and immunological responses (Ito et al., 2006; Phelan et al., 2009). Therefore, PHs can trigger signaling events involved in grapevine immunity (Gauthier et al., 2014). Some studies (Gauthier et al., 2014; Lachhab et al., 2014) report reduced symptoms of downy mildew and gray mold (Botrytis cinerea) on grapevine following applications of PHs derived from soybean and casein. PHs may improve plant nutrient uptake through modification of root architecture, by complexation of nutrients such as Zn (Ertani et al., 2018), or stimulation of microbial and enzymatic activities (Colla et al., 2015) – e.g., increased activity of N2-fixing, P-solubilizing, and indoleacetic acid-producing bacteria was observed in lettuce (Colla et al., 2015) and tomato (Colla et al., 2014) after application of PHs. The application of PHs also increases the activity of both nitrate reductase and glutamine synthetase in roots and leaves (Ertani et al., 2009). Nitrate reductase reduces nitrate to ammonia, and glutamine synthetase catalyzes the ATP-dependent amination of glutamate to produce glutamine, thereby facilitating assimilation or re-assimilation of nitrogenous sources originating from a wide variety of anabolic or catabolic processes (Ertani et al., 2009). Applications of different PHs have been observed to increase grapevine yield, correlated with PH organic N content (Bosselli et al., 2019).
Further research on the effects of PHs on viticulture should be explored, especially the impact on microbial communities living in the rhizosphere or phyllosphere (aboveground plant surfaces inhabited by microbes), which has received little study (Colla et al., 2015 and Colla et al., 2017).
Biostimulants can also be produced from marine macroalgae (i.e. seaweed extracts; SEs). In the last decade, the application of SEs in agriculture has gained traction and several seaweed species (such as brown seaweeds Ascophyllum nosdosum, Ecklonia maxima, and Laminaria spp, and red seaweeds Kappaphycus alvarezii, and Gracilaria edulis) are currently used in commercial production of biostimulants (Yakhin et al., 2017; El Boukhari et al., 2020). SEs have been successfully explored in arable cropping, including viticulture (Table 2). The most common method of applying SEs in vineyards is foliar spraying, although the extracts can be applied directly to the soil (Mancuso et al., 2006; de Carvalho et al., 2019). Like most biostimulants, SEs may increase crop productivity and improve fruit quality in both stressed and non-stressed conditions (Deolu-Ajayi et al., 2022). The vegetative shoot growth stage of the crop is frequently targeted for SE application to stimulate growth and boost crop vitality by modulating underlying biochemical processes, which are also important at the reproductive stage (Figure 1). SEs may also enhance microbial biodiversity, but information about this effect on the plant microbiome is scarce (El Boukhari et al., 2020).
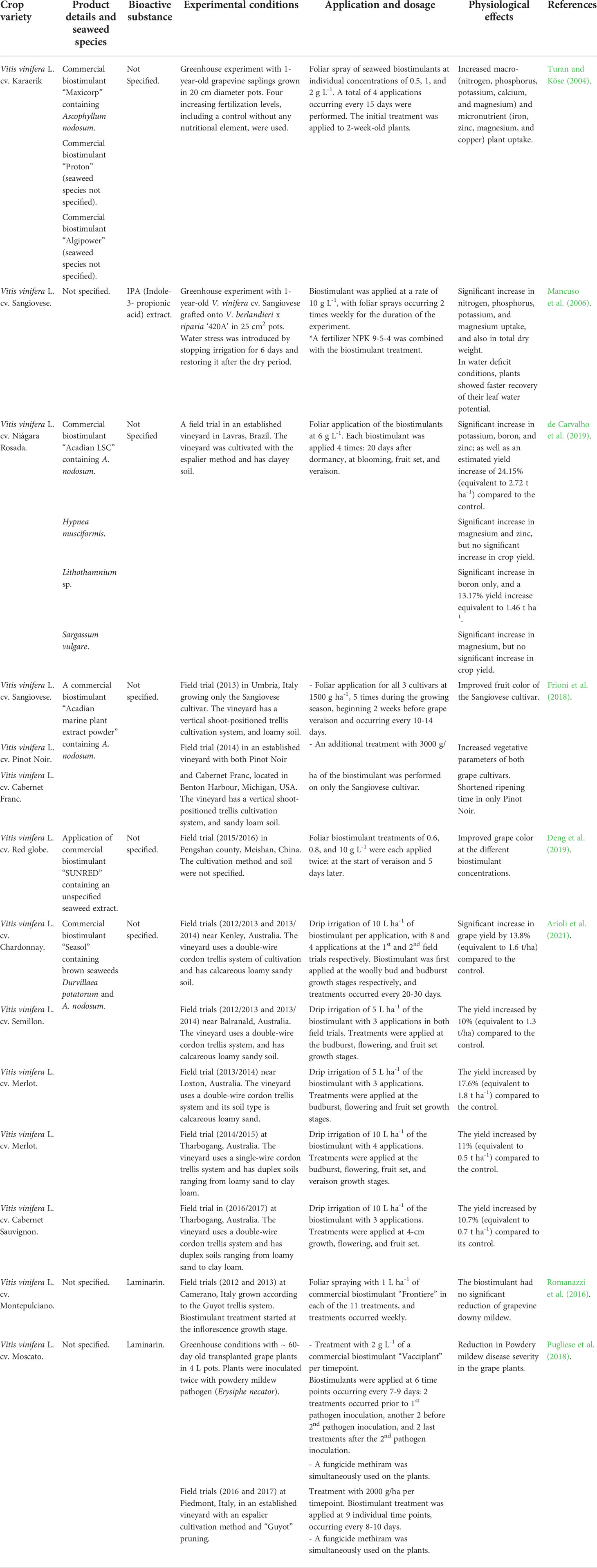
Table 2 An overview of seaweed-based (SEs) biostimulants applied in viticulture.
Physiologically, SEs boost nutrient assimilation and plant growth under non-stressed conditions. Spraying wine grapevines with any of three commercial SEs (Maxicrop, Proton, and Algipower) increased both macro- and micro-nutrient uptake from the soil (Turan and Köse, 2004; Table 2). Similar mineral uptake induction in wine grapevines was reported by Mancuso et al. (2006) and de Carvalho et al. (2019) when SEs were applied, resulting in a subsequent increase of dry weight by ~ 27% and fruit yield by ~ 24% (Table 2). Foliar application of either an Ascophyllum-derived seaweed extract or the commercially available extract “SUNRED” improved fruit quality and shortened the ripening time of wine grape (Frioni et al., 2018; Table 2) and table grape (Deng et al., 2019; Table 2) cultivars. The enhancement in fruit quality may be linked to an accumulation of anthocyanins and phenolic compounds, especially in the berry skin, which was also observed (Frioni et al., 2018; Deng et al., 2019).
During drought stress and water deprivation in controlled greenhouse experiments, foliar application of a seaweed indole-3-propionic acid (IPA)-derived extract induced faster recovery of leaf water potential and stomatal kinetics (Mancuso et al., 2006). Different wine grape cultivars treated with a commercial seaweed biostimulant “Seasol” via soil fertigation, showed an average fruit yield increase of 14.7% across five locations with varying environmental conditions (Arioli et al., 2021; Table 2). Yield increase occurred irrespective of heat and/or cold stress in a single growing season (Arioli et al., 2021). The performance of the field experiments suggests an essential soil-crop interaction for increased productivity (Arioli et al., 2021). Unfortunately, analyses of the soils and the applied extracts were not reported and therefore, cannot support this hypothesis. Moreover, variations in the SE dosage in different fields in this study prevent further conclusions regarding the direct effect soil type (and properties) may have on SE, as well as the overall impact on crop productivity in the presence of the biostimulant.
Although SEs reduce the effects of crop diseases (Shukla et al., 2019) and non-seaweed-derived biostimulants also contribute to disease prevention in viticulture (reviewed in Gutiérrez-Gamboa et al., 2019), the specific use of SEs to mitigate diseases of grapevine has not been extensively studied. Treatment with a commercial seaweed-derived biostimulant “Vacciplant” caused a reduction in the effects of powdery mildew disease in both greenhouse experiments and field trials (Table 2). Grapevine disease severity was reduced by ~ 65% and 52% in greenhouse and field experiments, respectively (Pugliese et al., 2018). Thus, there is still potential for the application of seaweed-derived biostimulants in viticulture for disease control, especially considering the need for reduced agrochemical reliance in the coming decades.
Bioactive compounds have biological activity in plants, some of which result in increased crop productivity and resilience to stress. While the presence of bioactive compounds in seaweed has long been suspected of having stimulatory effects on plants, including complex polysaccharides (absent in arable crops, e.g. alginate, laminarin, and fucoidan), phytohormones, sterols, and osmolytes (Yakhin et al., 2017), speculations remain regarding underlying mechanisms. This is perhaps due to current research focusing on growth promotion, rather than also investigating and linking the role of these bioactive compounds to specific mechanisms. Further research is needed, and will also provide essential information for biostimulant optimization. Of note is that changes in mineral concentration due to foliar application of SEs do not always enhance crop productivity. Only two of four SEs tested caused a significant increase in grapevine yield (de Carvalho et al., 2019; Table 2), indicating that SEs must be tested independently to establish their benefits. Conflicting observations may be explained by variations in biochemical composition among seaweed species and their derived extracts, linked to the environmental conditions of the cultivation sites as well as harvesting times (Khairy and El-Shafay, 2013). Overall, the application of SE is beneficial for crops in both stressed and non-stressed conditions and thus may be attractive for wider adoption in viticulture.
Humic substances (HS) are an important carbon compartment present in soils, waters, and sediments. Although researchers are not in complete agreement as to how HS are formed and structured, a growing body of evidence indicates they may serve as biostimulants of plant growth. HS biostimulants are available commercially in the form of products based on humic acids (HA), fulvic acids (FA), or a mixture of both (HA+FA). Efficacy is related to the product’s chemical composition, the form of extraction and application, the applied concentration, and the stage of development of the crop (Canellas et al., 2009; Zandonadi et al., 2013; Olivares et al., 2017; Jindo et al., 2020). Effects of HS application in plants often include the activation of plasma membrane H+-ATPase and the alteration of the primary and secondary metabolism, generally resulting in increased root growth, nutrient uptake, rate of photosynthesis, and attenuation of stress associated with salinity, drought, or metal toxicity (Zandonadi et al., 2007; Dobbss et al., 2018; Jindo et al., 2020) (Figure 1). It is suggested that some biostimulant effects of HS originate from their impact on structural and physiological modifications in roots and shoots related to nutrient assimilation and soil distribution (Canellas et al., 2015). Also, HS can simultaneously regulate the transcription and activity of some plant hormones following addition to the rhizosphere (Souza et al., 2022). Recently, it has been demonstrated that HS can alter the plant microbiome by favoring the recruitment of beneficial microorganisms after application (da Silva et al., 2021).
Despite a limited number of studies involving the specific use of HS in grape cultivation, several effects are consistently observed (Table 3). These include increases in photosynthetic rate, total soluble solids, total chlorophyll, and average berry size and weight (Ferrara and Brunetti, 2008 and Ferrara and Brunetti, 2010; Ibrahim and Ali, 2016; Popescu and Popescu, 2018; Irani et al., 2021). For example, Popescu and Popescu (2018) noted increased photosynthetic rate and total soluble fruit solids in cultivars ‘Feteasca Regala’ and ‘Riesling Italian’ planted in Romania, following foliar application of HA (40 and 50 mL L-1) previously extracted from vermicompost. The increase was attributed to a higher concentration of carotenoids in the leaves. Ferrara and Brunetti (2010) reported an improvement of parameters associated with fruit quality (e.g. increase in total soluble solids, [°Brix], and the °Brix/titratable acidity ratio, and a decrease in tartaric acid) due to the application of HA (100 mg L-1, extracted from the soil in Apulia, Italy) at full-bloom. In the cultivar ‘Italy’, six applications of HA (derived from different origins) increased chlorophyll content, °Brix value, and berry size (Ferrara and Brunetti, 2008). The authors attributed these effects to the presence of 6% nitrogen in the humic matrix. However, the increase in berry size might also be the result of the hormone-like activity of HA (i.e., auxin, gibberellin- and cytokinin-like activity), which is useful for organic seedless table grape production where the application of synthetic hormones is not permitted. In Egypt, Ibrahim and Ali (2016) applied HA (Greenhum Company, Italy) in both February and March to the cultivar ‘Superior Seedless’. Chlorophyll content, berry weight, and total fruit volume significantly increased after HA was respectively applied to leaves (5.0 g L-1) or to the soil (5 and 7.5 g L-1, 2.5 g L-1). In Spain, Sánchez-Sánchez et al. (2006) reported increased iron and phosphorus levels and reduced sodium in the leaves of cultivar ‘Italy’ following treatment with solutions containing HA+FA. Recently in Iran, Irani et al. (2021) applied FA (0.5%) via leaf spray or HA (20 g plant-1) via irrigation, and observed the alleviation of drought stress, increased nutrient absorption, and concentration of proline (an important stress signaling molecule), carbohydrates, and soluble proteins. Additionally, HA and FA application resulted in greater berry weight, productivity, and concentration of total soluble solids (Irani et al., 2021).
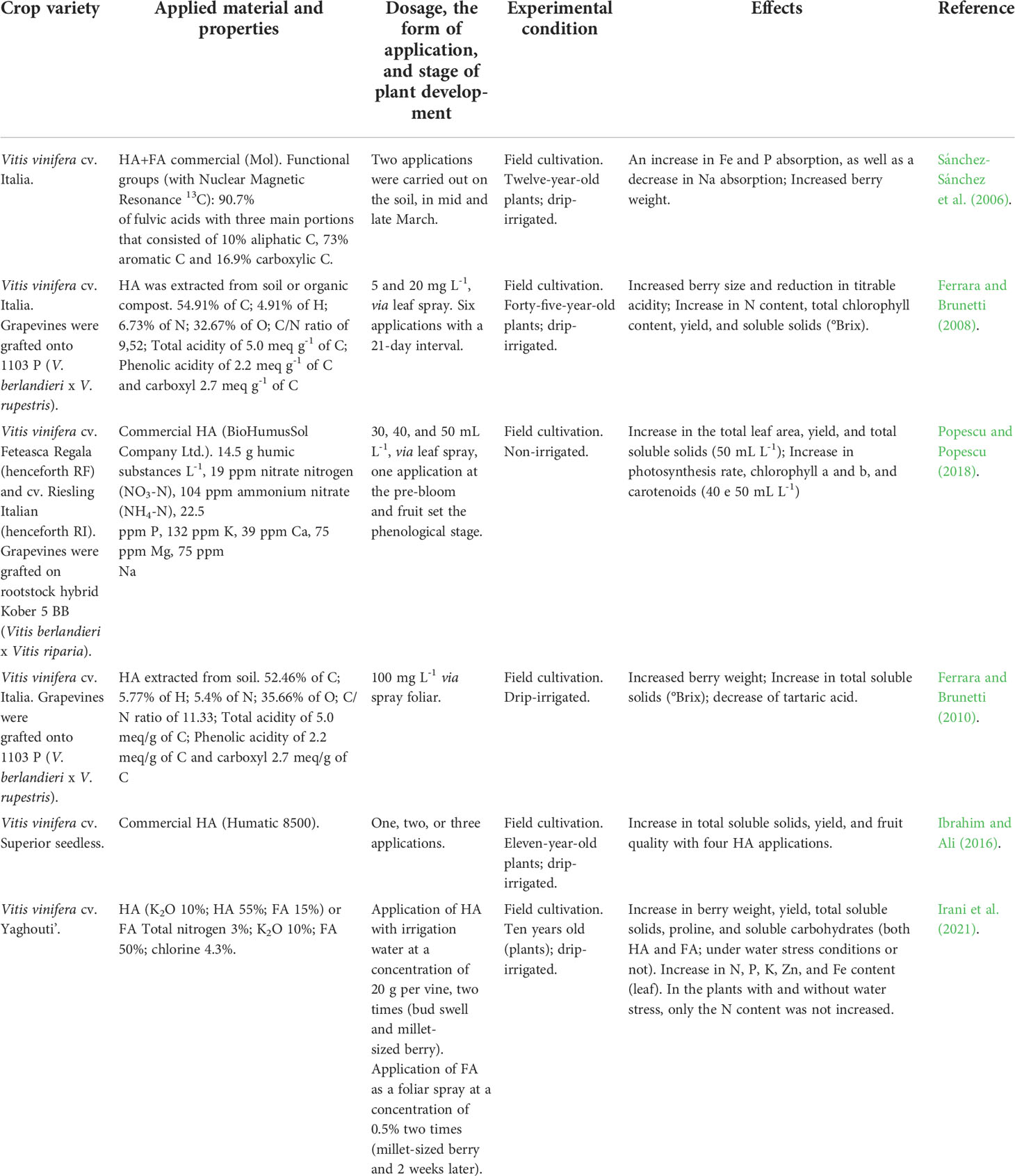
Table 3 An overview of the main effects of humic substances (HS) applied in viticulture.
Protocols outlining best practices in terms of vineyard HS application practices and optimal concentrations are yet to be developed, and research regarding potential negative structure-activity interactions has yet to be conducted. Despite these shortcomings, the beneficial effects of HS on grapevines are evident, generally resulting in increased production, increased fruit quality, and an expansion of the plant’s ability to tolerate environmental stresses.
Pyrogenic materials are obtained by pyrolysis of plant biomass, a thermal decomposition process occurring at relatively high temperatures in the absence of oxygen. These materials represent a sustainable source of biostimulants with a long tradition in agriculture (Ogawa and Okimori, 2010). Pyrogenic materials include biochar (solid C-rich residue of pyrolysis), and pyroligneous acid (also known as wood vinegar, the aqueous fraction obtained from the condensation of pyrolysis vapors). Biochar is highly aromatic, porous, and possesses a chemically recalcitrant structure. It is generally used as a soil amendment, due to potential agronomic and environmental benefits associated with enhanced soil fertility and long-term soil C sequestration (Woolf et al., 2010; Lehmann et al., 2011). Pyroligneous acid consists of a mixture of a large number of oxygenated organic compounds (including acids, alcohols, ketones, phenols, furans, and ethers) and hydrocarbons with antioxidant and antimicrobial properties. Pyroligneous acid has potential as a plant growth and germination biostimulant, antioxidant and free-radical scavenger, pesticide, and antimicrobial agent (Mungkunkamchao et al., 2013; Grewal et al., 2018).
Biochar is more commonly used than pyroligneous acid in vineyards, either alone or in combination with compost (Figure 1 and Table 4). There is contrasting information in the literature regarding biochar’s impact on vineyard productivity. A recent meta-analysis by Payen et al. (2021) showed that biochar application increased soil organic C, at a rate of 8.96 Mg CO2-eq ha−1 yr−1. The authors also postulated biochar application might lead to enhanced vineyard productivity, but additional long-term investigations are needed to support this statement. Several authors (Baronti et al., 2014; Genesio et al., 2015; Maienza et al., 2017) have reported increased productivity in biochar-amended vineyards, associated with enhanced physicochemical properties of the amended soils, increased soil water availability, and growth of fine root biomass (Amendola et al., 2017). However, there were no significant differences in grape quality. Conversely, other reports indicate only minor effects on soil physicochemical properties following biochar application over a three-year period, which did not significantly affect vineyard fertility (Schmidt et al., 2014; Sánchez-Monedero et al., 2019). Although these authors did observe benefits following the addition of biochar-blended compost to the soil, effects were similar to those in control vineyards amended with conventional compost without biochar.
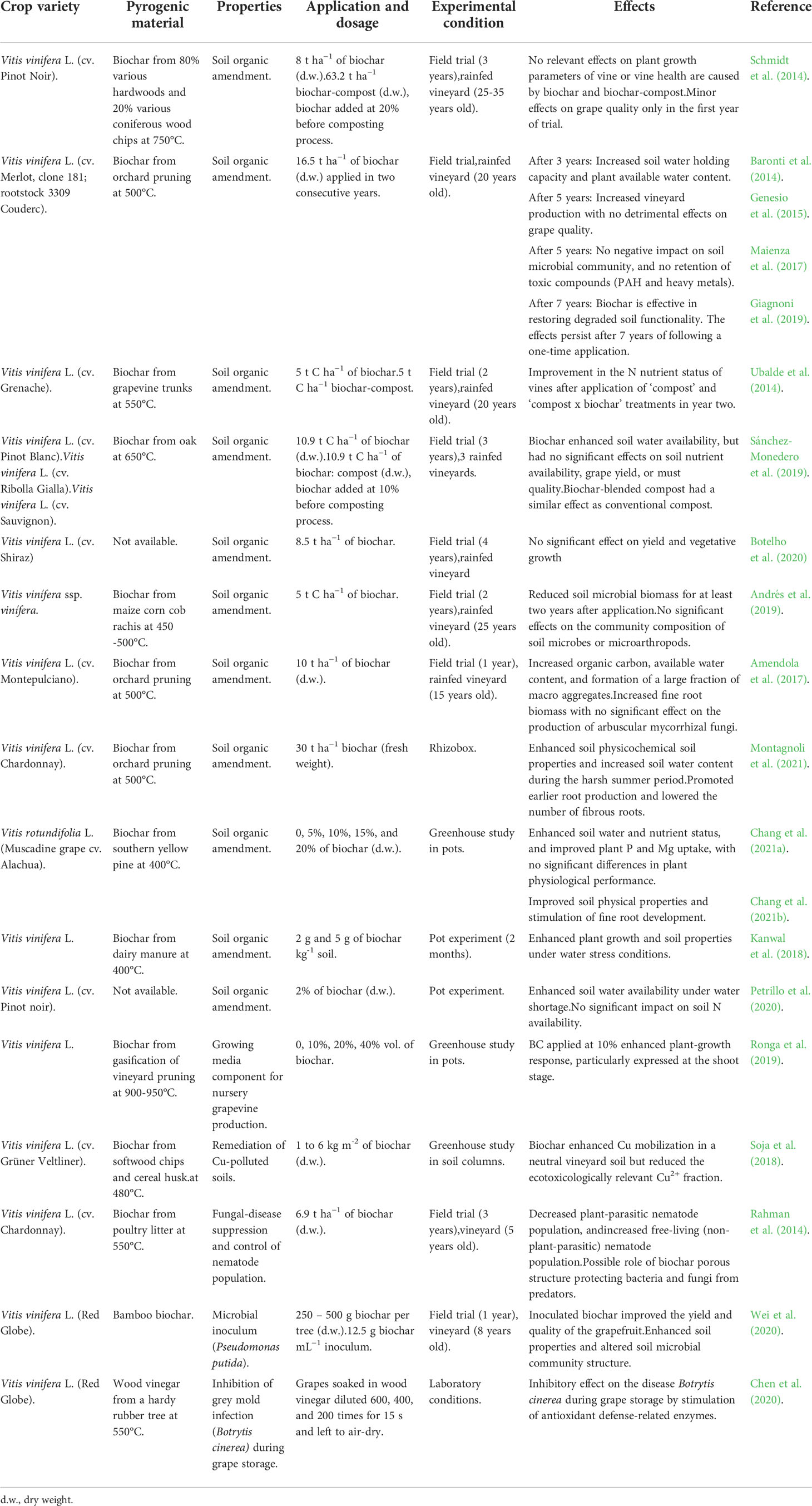
Table 4 An overview of the main effects of pyrogenic materials in viticulture.
Biochar may also act as a biostimulant when used as a carrier of inoculum for microorganisms (Hale et al., 2015; Głodowska et al., 2016; Hardy and Knight, 2021), as a suppressor of plant disease (Graber et al., 2010; Debode et al., 2020), and as a coating for novel biochar-fertilizer composites (Joseph et al., 2013) or slow-release fertilizers (An et al., 2020). However, these applications have not been fully explored in vineyards. Hale et al. (2015) demonstrated in a soil incubation experiment that biochar might support high population densities of the plant growth-promoting rhizobacteria (PGPR) Enterobacter cloacae. Interestingly, chemical properties of the biochar (particularly pH and N content) affected initial inoculum survival, but physical properties (surface area and porosity) were mainly associated with later survival after soil application (Hale et al., 2015). Biochar did not negatively affect rhizobacterium activity, which may occur due to bacterial signaling compounds or bacteria-derived plant growth hormones binding to the biochar surface. Depending on feedstock and pyrolysis conditions, certain biochars with large surface area or high pH can adsorb or hydrolyze signaling molecules, disrupting soil microbe cell-to-cell communication (Gao et al., 2016).
Similarly, Zhu et al. (2017) reported that biochar could modify communication between rhizosphere microbial communities and plant roots, affecting plant response to soil-borne pathogens (Harel et al., 2012; Akhter et al., 2015). Graber et al. (2010) proposed that induced resistance against pathogens observed in both tomato and pepper following biochar application may be due either to a shift in soil microbial populations following biochar addition, or to the release of chemicals from biochar toxic to pathogens. The utilization of beneficial microorganisms as biostimulants is discussed in greater detail below, and other mechanisms by which biochar can act as a disease-suppressing agent have been previously summarized by Bonanomi et al. (2015) and Graber et al. (2014).
Pyroligneous acid is often applied as a foliar spray or soil drench (Mungkunkamchao et al., 2013), or in combination with biochar (Zhang et al., 2020). Zhang et al. (2020) demonstrated the benefit of applying biochar and pyroligneous acid for increased blueberry yield and nutritional quality, by enhancing soil organic matter and nutrient availability. In terms of vineyard pest management, Chen et al. (2020) studied the ability of pyroligneous acid to inhibit grey mold (Botrytis cinerea) infection of table grape cultivar ‘Red Globe’. The application of diluted pyroligneous acids (200- and 400-fold dilution) improved grape resistance to grey mold, likely through stimulation of antioxidant defense-related enzyme activities, including those of superoxide dismutase, peroxidase, and ascorbate peroxidase. However, other investigations of pyroligneous acid use in vineyards are lacking.
The application of biostimulants from pyrogenic materials may help vineyards adapt to climate change, especially in a scenario of water shortage which may be particularly detrimental to viticulture (Maienza et al., 2017). Fascinatingly, the positive impacts of biochar on vineyard soil fertility are maintained in the long term (7 years), even after a single application (Giagnoni et al., 2019). This benefit has also been identified for improving the growth and symbiotic performance of other plants (lupin, Lupinus angustifolius) under drought stress conditions (Egamberdieva et al., 2017).
Certain prokaryotic soil microorganisms can establish beneficial relationships with plants. Coined ‘plant growth-promoting rhizobacteria’ (PGPR) (Kloepper and Schroth, 1979), these microbes can enhance plant growth and development through several direct and indirect processes, performed at different plant growth stages (Vorholt, 2012; Ruzzi and Aroca, 2015; Castellano-Hinojosa and Bedmar, 2017). According to Chauhan et al. (2015), the main PGPR traits are biofertilization and phytostimulation (through the excretion of phytohormones such as indole-3-acetic acid [IAA], cytokinins, and gibberellins), tolerance to biotic and abiotic stress (via 1-aminocyclopropane-1-carboxylate [ACC] deaminase activity), and biopesticide and biocontrol activity (production of antibiotics, lytic enzymes, hydrogen cyanide [HCN], and volatile organic compounds [VOCs], among others) (Chauhan et al., 2015; Kumar-Jha and Saraf, 2015; Goswami et al., 2016; Oleńska et al., 2020). Additionally, PGPRs can have synergistic interactions with the endophytic grape microbiome (Vandana et al., 2021).
Bacteria must first be isolated and cultured prior to their utilization as PGPRs. For use in viticulture, several authors have isolated viable PGPRs from the microbiome of Vitis vinifera L., which may then be re-applied as biostimulants in vineyards (Compant et al., 2011; Pinto and Gomes, 2016; Rezgui et al., 2016; Pacifico et al., 2019). The effectiveness of these PGPRs depends on several factors including environmental conditions, soil characteristics, and even crop variety (Pacifico et al., 2019). Nevertheless, PGPRs are a feasible tool for use in vineyards to effectively promote plant growth and protection (Figure 1).
Most studies of PGPRs in viticulture are focused on the initial stages of crop development in specific vine cultivars. Additionally, few studies testing strains at a field scale can be found (Table 5). Under in vitro and/or greenhouse conditions, Muganu et al. (2015); Köse et al. (2015), and Velásquez et al. (2020a) noted a wide variety of PGPR strains (Ensifer, Burkholderia, Pseudomonas, and Bacillus) were able to promote growth and root callusing percentage in four different V. vinifera cultivars: ‘Beyaz Çavus’, ‘Italia’, ‘Cabernet Sauvignon’, and ‘Chardonnay’. Köse et al. (2003) observed Bacillus strains BAI6 and OSU142 increased rooting in 41B (a hardy cross between old-word V. vinifera and new-world V. berlandieri) rootstocks. Similarly, Sabir (2013) and Sabir et al. (2017) applied P. putida BA-8 and B. simplex T7 to 41B hybrids and recorded a promotion of graft callusing, scion shoot growth, nursery survival rate, and fruitfulness. Salomon et al. (2016) tested a combination of strains B. licheniformis and P. fluorescens in order to improve the growth of V. vinifera cultivar ‘Malbec’ in greenhouse conditions. Also in a greenhouse, the cytokinin and auxin-producing B. megaterium M3 was applied in combination with Agrobacterium rubi A18 and Alcaligenes eutrophus Ca-639 to V. vinifera, grafted onto 41 rootstocks. Growth enhancement and increased grapevine pruning residue weight were noted (Salomon et al., 2017).
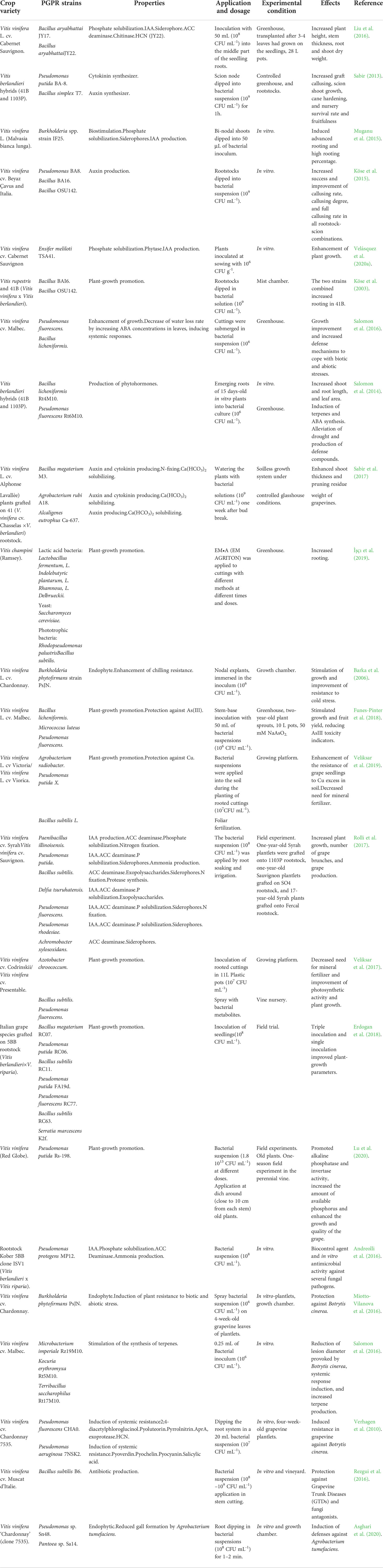
Table 5 An overview of the main effects of plant-growth-promoting rhizobacteria (PGPR) applied in viticulture.
In a rare field-scale experiment, Rolli et al. (2017) applied several bacteria from the genera Paenibacillus, Pseudomona, Bacillus, Delftia, and Achromobacter, to V. vinifera cultivars ‘Syrah’ and ‘Sauvignon’. Increased plant growth, number of grape brunches, and grape production were recorded. Veliksar et al. (2017) applied strains of Azotobacter, Pseudomonas, and Bacillus to two V. vinifera cultivars in Moldova, and observed improved photosynthetic activity, as well as reduced mineral fertilizer requirements. Erdogan et al. (2018) achieved similar improvements with combinations of a wide range of PGPR strains in grafted rootstocks of V. berlandieri and V. riparia in Turkey. In China, Lu et al. (2020) observed enhanced growth and final grape quality, following the application of P. putida Rs-198 to a perennial variety.
In addition to general growth and quality enhancement, inoculation with PGPRs may also induce grape tolerance against biotic and abiotic stresses. For example, the application of phosphate-solubilizing bacteria isolated from the Vitis rhizosphere resulted in improved plant development when grown in saline-alkaline soils (Liu et al., 2016). Salomon et al. (2014) observed both drought alleviation and production of defense compounds in the grape cultivar ‘Malbec’. Barka et al. (2006) found that Burkholderia phytofirmans PsJN could effectively protect Chardonnay grapes against cold stress.
Other studies focus on using PGPR strains to alleviate heavy metal stress. Funes-Pinter et al. (2018) identified B. licheniformis, Micrococcus luteus, and P. fluorescens as effective species for elevating growth and fruit yield in plants when grown in the presence of arsenic (As III). Veliksar et al. (2019) used strains of Agrobacterium radiobacter, P. putida, and B. subtilis to increase resistance against high soil concentrations of copper, and reduce the amount of mineral fertilizer needed.
Microorganisms may also be used as biocontrol agents against fungal pathogens in vineyard soils. For example, several studies have applied bacterial strains in order to protect vineyards from the common vine pest grey mold (Botrytis cinerea). Miotto-Vilanova et al. (2016) and Verhagen et al. (2010) found that several strains belonging to he Pseudomonas, Burkholderia, Microbacterium, Kocuria, and Terribacillus genera harbored plant growth and phytopathogen-control traits against B. cinerea. Andreolli et al. (2016) and Rezgui et al. (2016) also tested strains of P. protegens and B. subtilis against fungal pathogens, with the latter obtaining good results in field trial conditions. These studies show that a vast myriad of bacterial-borne traits can be effective for biocontrol, including the production of antibiotics, terpenes, pigments, and proteases, as well as inducing resistance against biotic stresses. As an indication of the high potential for PGPRs to combat plant biological stress, Asghari et al. (2020) applied Pseudomonas and Pantoea sp. and noted lowered gall formation produced by Agrobacterium tumefaciens.
As shown above, PGPRs have so far primarily been researched in vitro, and in greenhouse trials. Few studies exist at the field scale and any practical yield benefit from these microorganisms has yet to be quantified (Bashan et al., 2014). Other optimizations must be performed before any selected consortia can be developed into commercial products. The survivability and persistence of the applied strains must be quantified not only in the soil along with any competitive/negative effects from the native rhizosphere microbiome but also in stable storage conditions. European legislation restricting the application of chemical substances to crops has been passed relatively recently (Regulation EC 1107 (2009). It is therefore alarming that only a single reviewed study concerned the development of commercial bioinoculants; İşçı et al. (2009) applied consortium EM·A (comprised of lactic acid, phototropic bacteria, and yeasts; EM AGRITON Ltd., Belgium) to V. champini, and recorded improved rooting.
More than 400 MYA, the presence of mycorrhiza in soil was essential for the colonization of terrestrial environments by plants due to their ability to provide stress tolerance and soil resources via symbiosis (Heckman et al., 2001). This adaptation continues in the majority of plants on Earth, including grapevines (Trouvelot et al., 2015). Roots of grapevines form a particular type of mycorrhizal symbiosis called arbuscular mycorrhizae, characterized by the penetration and internal colonization of plant root cells by fungal hyphae (Trouvelot et al., 2015). Arbuscules form within the plant roots and serve as the exchange site for various metabolites (Trouvelot et al., 2015). Arbuscular mycorrhizal fungi (AMF) are widely known for being able to enhance the uptake of P in the host roots, a nutrient that is typically limiting in cropping systems like vineyards (Smith et al., 2011; Van Geel et al., 2017). However, they also provide increased plant uptake of N and other limiting elements including trace metals Fe, Mn, Cu, and Zn (Clark and Zeto, 2000), which are also critical to plant health (Figure 1). This is accomplished by greatly extending the root system’s exploration and exploitation area (Smith and Read, 2008; Trouvelot et al., 2015), simultaneously increasing plant access to water sources that would be otherwise inaccessible (Al-Karaki and Clark, 1998). This feature may be particularly advantageous in rain-fed vineyards growing in nutrient-poor soils, which are widespread in most traditionally wine-producing regions such as the Mediterranean, Middle East, and Caucasian regions (Table 6).
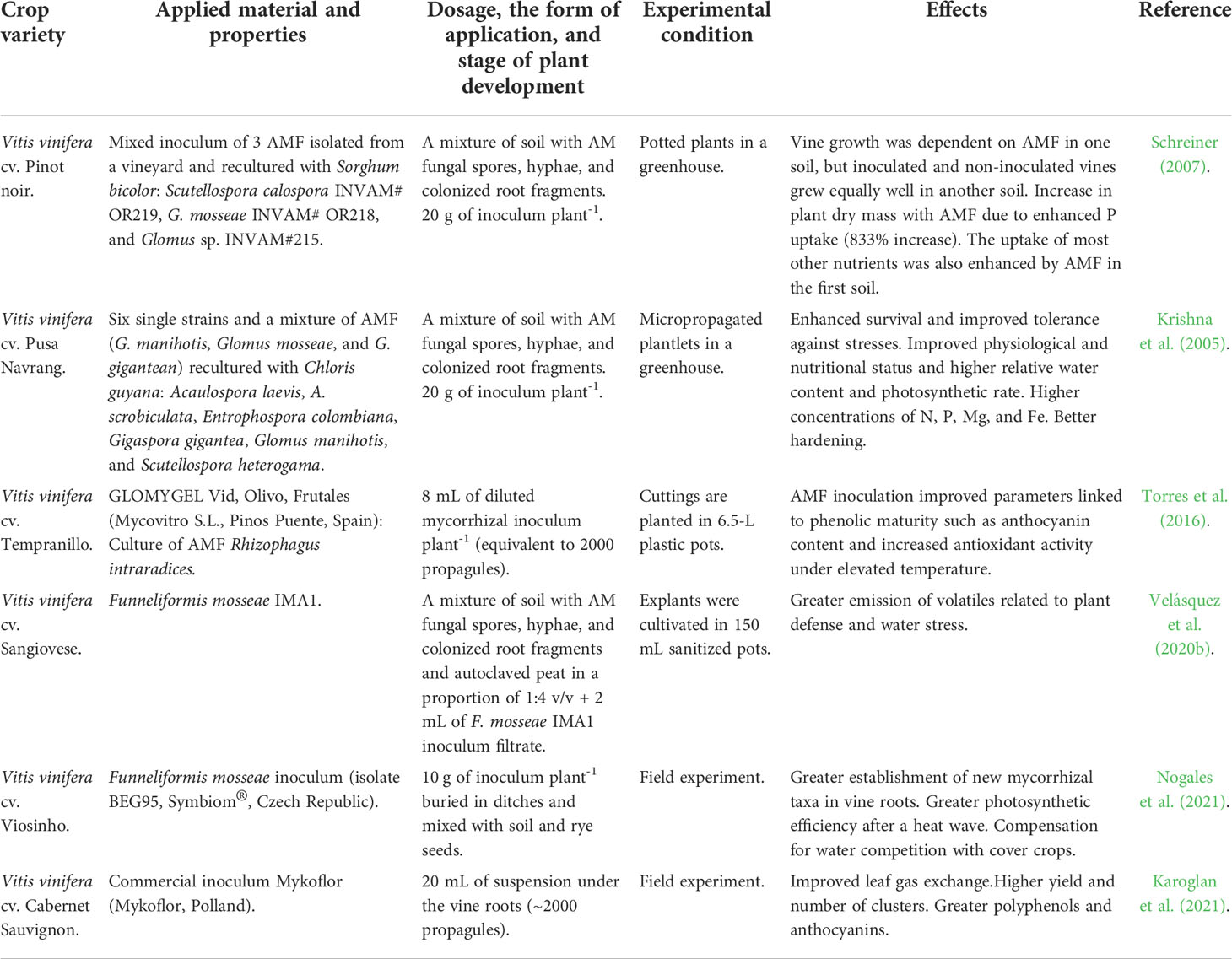
Table 6 An overview of the main effects of mycorrhizal inoculants applied in viticulture.
In addition to their role in plant water and nutrient uptake, AMF contributes to the biosynthesis of a wide range of molecules such as vitamins and hormones needed to support the metabolism and health of plants (Strzelczyk et al., 1991), although this is poorly investigated in the context of vineyards. Arbuscular mycorrhizal fungi can also protect grapevines against harmful pathogens and pests, including the root-knot forming nematode Meloidogyne incognita, through the induction of a defense response involving enzymes like chitinases (Li et al., 2006). Moreover, AMF contributes to the development of a healthy rhizosphere community (i.e., a microbially diverse community that is functionally linked to the plant), which in turn may confer tolerance/resistance against a range of biotic and abiotic stresses (reviewed above), including heat, drought, salinity, pathogenic infection, and pests (Fitter and Garbaye, 1994; Gryndler, 2000; Jeffries et al., 2003; Gupta et al., 2018). Given their importance in vine nutrition and pathogen response, AMF are a critical pillar of healthy, functioning vineyards (Trouvelot et al., 2015), especially in the current context of widespread environmental degradation and climate change. In addition to direct benefits to plant health, mycorrhizae may also provide important ecosystem services, including increasing potential carbon sequestration of vineyard soils and reducing erosion (Trouvelot et al., 2015).
Despite these benefits, mycorrhizal symbiosis is frequently disrupted in croplands due to intensive management, including excessive tilling, which breaks the orderly structure of soil aggregates and fungal networks (Gosling et al., 2006; Bowles et al., 2017; Porter and Sachs, 2020), although this has been poorly investigated in vineyards (Winter et al., 2018). Biocides can also negatively affect mycorrhizal fungi, with deleterious consequences on the establishment of symbiosis (Gosling et al., 2006; Zaller et al., 2018), while synthetic fertilizers disrupt the mycorrhizal association due to the ablation of nutritional constraints, such as N and P (Gosling et al., 2006; Van Geel et al., 2017). Regenerating proper vine-mycorrhizal balance and function in degraded vineyards is thus a priority, which may yield many benefits to both growers and the wider society.
Regeneration of the vine-AMF interaction can take place through various means, which should be considered holistically in terms of impact on total vineyard management. One such strategy is the inoculation of grapevines with one or more strains of mycorrhizal fungi previously selected for their ability to colonize vine roots (Linderman and Davis, 2001; Trouvelot et al., 2015). One aspect in which inoculation with mycorrhizal biostimulants might be particularly important is to minimize the growing threat of trunk diseases (Petit and Gubler, 2006; Holland et al., 2019), which may be linked to widespread alteration of plant-microbial associations. It has been recently suggested that declines observed in woody plants are related to microbiome modifications or imbalances (i.e., dysbiosis) (Porter and Sachs, 2020). Arbuscular mycorrhizal fungi known to help re-establish symbiosis include various strains of Glomus intraradices, such as G. intraradices BEG72 (Nogales et al., 2008) and INVAM CA501 (Petit and Gubler, 2006).
Proper timing may be critical when inoculating plants with mycorrhizal biostimulants (Sohn et al., 2003), but there is little information for vineyards. It is suggested that rootstocks be submerged in an AMF spore solution before planting, but variable success is reported. For example, a study was carried out on Vitis rupestris cv. St. George using G. intraradices and reported that pre-inoculated plants were less susceptible to black foot disease (Petit and Gubler, 2006), while (Holland et al., 2019) observed no difference between inoculated and uninoculated plants. Some studies have also reported clear effects in the greenhouse, but not under realistic field conditions (Rosa et al., 2020). Other ways to add AMF to vineyards include the direct spraying of commercial biofertilizers on adult plants, through the addition of granules containing spores, and potentially also the translocation of whole soil inoculants from previously selected locations particularly diverse in terms of AMF such as forests. This latter approach is underexplored in vineyards. Perhaps contributing to variability in inoculation success is the difficulty in altering the pre-established microbiome of adult plants. Moreover, the mixed positive and neutral effects of inoculating vines with AMF, together with the fact that some (but not all) mycorrhizal fungi show a certain degree of host specificity (Campos et al., 2018), suggest the importance of screening AMF strains against potential compatible host vine cultivars/rootstocks prior to inoculum development (Schreiner, 2007). Finally, the vine-AMF symbiosis is context dependent and may be linked to factors critical for productive vineyards including soil fertility and other properties (e.g., soil organic matter content, pH, and texture) (Trouvelot et al., 2015). For example, the presence of a well-developed community of cover crops in vineyard inter-rows may favor inoculated AMF establishment, by providing an additional host crop and continuous reservoir for supplying adjacent grapevines (Winter et al., 2018).
Climate change simulations predict that the fitness of crop insect pests will increase beyond 30° latitude North and South of the equator (Santos et al., 2020). Indeed, a three-decade study observed shifts in the phenology of grape berry moths (Eupoecilia ambiguella, E. viteana, and Lobesia botrana), distribution ranges of leafhoppers (Scaphoideus titanus Ball, a common vector of grapevine diseases), and range expansion of grapevine mealybugs (Planococcus ficus) (Reineke and Thiéry, 2016).
The effective control of crop pests is a continuous process. For native and newly introduced pests, the adoption of novel biological control agents (BCAs) is needed for sustainable alternative pest management (Figure 1). BCAs are living organisms antagonistic against pests. By definition, biological control must involve: 1) a biocontrol agent, 2) a pest to be controlled, and 3) a farmer or a stakeholder benefitting from the pest control (Stenberg et al., 2021). Organisms such as insects or mites, bacteria, fungi, and nematodes are used to control weeds or pests and diseases of cultivated plants (Ehlers, 2011). Viruses are not living entities, but contain structural biological components such as nucleic acids and proteins and are therefore also considered BCAs (Stenberg et al., 2021). Semiochemicals (chemical molecules produced by organisms that modify the behavior of other living beings, i.e., bio-communication) and plant extracts that can act directly on a pathogen or pest are other options for BCAs (Ehlers, 2011). Biological control is an interaction between at least two organisms, and success is therefore influenced by many factors including climate, reproduction mode and rate, food availability, and others. Holistically considering the total impact of all dynamic factors together when designing a balanced pest management plan is referred to as integrated pest management (IPM). BCAs are a main component of IPM, and together, are becoming increasingly utilized in agriculture.
Biocontrol agents may be naturally present in the agroecosystem (e.g. a native population of soil microbes antagonistic to plant-parasitic nematodes), or first grown in vitro and then released (Sharma et al., 2009; Stenberg et al., 2021). Grapevine is a perennial crop and harbors a large microbiome in both the rhizosphere and stem tissues. Additional microorganisms which act to balance the microbiota are also found in the phyllosphere and fructosphere (Ranade et al., 2021). Many of these microorganisms act as BCAs and are a promising ecological strategy for disease control (Carro-Huerga et al., 2021).
The cropping system (e.g. conventional vs. organic farming) also influences the potential number of biocontrol agents which may be isolated for application. Cordero-Bueso et al. (2017) evaluated the biocontrol potential of 230 grape yeast isolates from different cultivation systems. The fractions with the most candidates were isolated from wild vines (62.7%) and biodynamic vineyards (17.7%). The least number of candidates were isolated from organic (6.2%) and conventional systems (7.2%). Wild vine species may therefore serve as a valuable resource for bioprospecting future BCAs. Additional potential sources of candidates include other plants, insects, animal intestinal tracts, soils, and marine and freshwater environments (Kurtzman et al., 2011).
BCAs can have several mechanisms of action against pathogens. During resource limitation, BCAs may outcompete detrimental microbes in terms of space (niches), nutrients, water, and/or light (Wang et al., 2018; Almeida et al., 2020). Other mechanisms include iron depletion (Sipiczki, 2006; Cordero-Bueso et al., 2017); production of lytic enzymes (Cordero-Bueso et al., 2017; Cabañas et al., 2020); production of volatile organic and semio-compounds antagonistic against pests (Cabañas et al., 2020; Don et al., 2021); resistance induction of the host plant (Arras, 1996; Jeandet et al., 2002; Maachia et al., 2015; Haidar et al., 2016); direct parasitism; tolerance of reactive oxygen species (ROS) (Aziz et al., 2003; Jamalizadeh et al., 2011); biofilm formation (Cabañas et al., 2020); synthesis of pathogenesis-related proteins (Chan and Tian, 2006); and antibiotic production (Maachia et al., 2015; Cordero-Bueso et al., 2017).
Production of extracellular mucilage produced by microbial antagonists throughout host cells may be linked to cell adhesion, and contain biochemical elicitors to signal defense responses (El-Ghaouth et al., 1998). Fragments of yeast cell wall oligosaccharides have also been noted to possess elicitor potential.
In viticulture, most BCAs are applied to control fungal diseases and pests associated with insect and mite vectors. However, other biological controls have also been researched and suggested for other pests. For example, harmful mollusks (snails) in vineyards in cool and wet climates may be effectively controlled by the nematode Phasmarhabditis hermaphrodite (Schneider), as there are restrictions on the use of synthetic molluscicides (Egleton et al., 2021). BCAs are also being developed for the control of weed species prevalent in vineyards (Samad et al., 2017).
In a grapevine host, antibiosis and resistance induction were observed, attributed to the antagonism of Bacillus subtilis B29 against Uncinula necator (powdery mildew) and Botrytis cinerea (grey mold) (Maachia et al., 2015). An n-hexane extraction of the cell-free supernatant of B. subtilis B29 revealed the presence of 17 fractions through HPLC. Two fractions were considered antibiotics against M. ramannianus and M. luteus, based on their antimicrobial activity (Sihem et al., 2011). B. subtilis B29 and B27 have also been described as inducing host resistance through the high production of phenolic compounds, with a significant increase in hydroxycinnamic acid (Maachia et al., 2015).
Antagonism against pests alone does not necessarily make an organism a BCA. The applied fungi, yeasts, and bacteria must possess other characteristics to allow for practical use in the field. For example, a BCA with greater adaptability than the pest may allow for widespread niche colonization and higher competitive potential (El-Ghaouth et al., 2004). Additionally, prior to application, the BCA candidate should be extensively screened for the production of metabolites harmful to non-pest organisms, especially humans. In the specific case of biological control of molds on grapes following harvest, the BCA should not leave residue on the berries. After identifying the mechanism of antagonism, and confirming BCA adaptability and safety, appropriate tests following the guidelines of national regulatory bodies must be conducted before registration as a commercial inoculant.
The use of BCAs in agriculture began in the second half of the 20th century, and several candidates have been identified. Although many researchers have since evaluated their pest control potential, few inoculants reach commercialization. Bacillus subtilis is one of the most widely applied fungicidal BCAs (Garrido and Botton, 2020). During the 2020/2021 harvest in Brazil, 182 commercial brands of synthetic fungicides were available for application to vineyards, compared to 28 biological fungicides (Table 7). Only two genera were represented, Bacillus and Paecilomyces, with a predominance of Bacillus (mostly B. subtilis). Other species of Bacillus were B. amyloliquefaciens; B. licheniformis; B. methylotrophicus; and B. pumilus. Paecilomyces lilacinus has been cited as an active ingredient in seven commercial brands. Most are composed of a single species and strain.
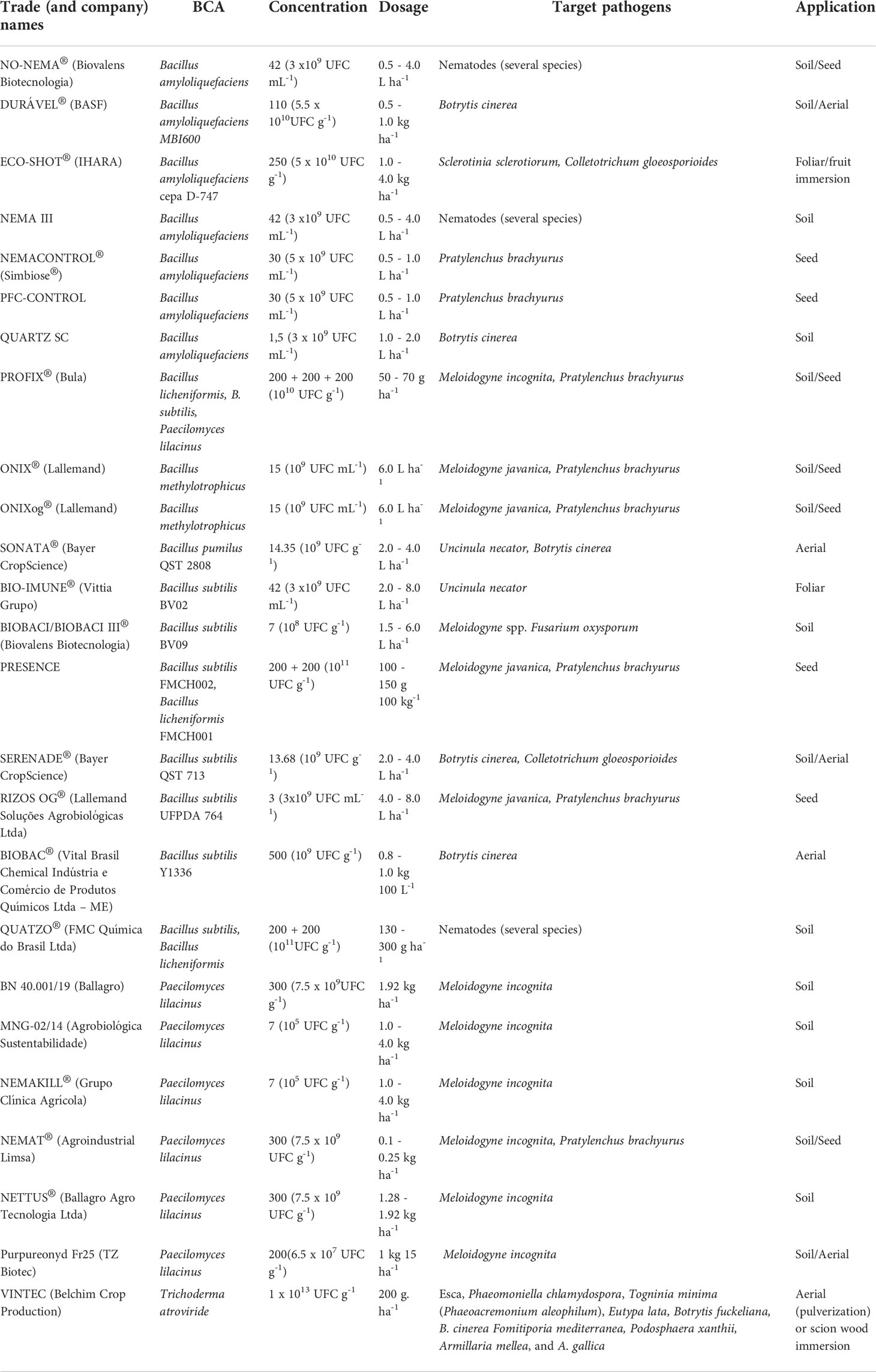
Table 7 Principal biological control agents (BCAs) applied in viticulture in Brazil.
Esca is caused by the association of the fungi Phaeoacremonium minimum and Phaeomoniella chlamydospore and is one of the main Grapevine trunk diseases (GTDs). Control of Esca is achieved mainly through BCAs, in which common biological agents are Trichoderma fungi (Chervin et al., 2022). Trichoderma atroviride-based products (CS1 by Vintec® - Belchim Crop Production and I-1237 by Esquive® - Agrauxine by Lesaffre) are often effective because they have multiple mechanisms of action: substrate competition, antibiosis, and mycoparasitism (Pertot et al., 2015; Belchim, 2022). For the control of powdery mildew (Uncinula necator), AQ10® (Ampelomyces quisqualis) is used commercially (Benuzzi and Baldoni, 2000). Another biologically-controlled disease is downy mildew (Plasmopara viticola). Trichoderma harzianum (known as Trichodex®), acts against the oomycete by increasing lignin, callose, and hydrogen peroxide, in addition to upregulating the defense enzymes phenylalanine ammonia-lyase, peroxidase, and 1,3-glucanase (Kamble et al., 2021).
The choice of BCA is of paramount importance. Simultaneous with disease control, some BCAs can influence grape productivity and quality with variable effects on acidity, soluble solids content, and berry size. Malviya et al. (2022) cite significant differences in the acidity of vine fruits treated with three BCAs ranging from 3.7 to 4.2 and 4° Brix in the SST. Production per plant doubled when treated with BCAs and almost tripled when combined with BCA + sulfur.
As for insecticides and acaricides, 55 synthetic products and 38 biological commercial products have been authorized for grapevines. Twenty-six contain Beauveria bassiana as an active ingredient and three with B. thuringiensis. For the control of insects and mites, recommendations for predatory insects (one Orius insidiosus and one sterile male pupil of Ceratitis capitata), predatory mites (four Neoselulus californicus and two Phytoselulus macropilis with AcMNPV virus), and one viral compound (AcMNPV virus, ChinNPV Virus, HearNPV Virus, SfMNPV Virus) are available.
Many antagonists with BCA potential are studied with increasingly efficient and cost-effective techniques. In addition to bioprospecting for biopesticides, valuable antibiotics may also be obtained from these organisms. Decreased BCA production costs and improved management techniques that increase and prolong BCA effects will allow for scaling their use in viticulture. Thus, biological products with multiple benefits (controlling disease(s), promoting plant growth, and increasing grape quality) are compliant with the sustainable development objectives of the 2030 agenda of the United Nations (UN). The reduction of synthetic chemical inputs in this manner can simultaneously reduce contamination throughout the production chain and ecosystem.
Future studies should investigate other applications of BCAs, such as the use of parasitic nematodes (Phasmarhabditis hermaphrodita) or earthworms (Lumbricidae) for pest control against snails and slugs in vineyards.
The application of biostimulants and BCAs may allow for improved sustainable viticulture and may serve as alternatives for chemically synthesized agronomic inputs, thereby reducing the negative environmental impact of pesticides and fungicides. The positive effects on plant growth are summarized in Figure 1 and vary widely depending on the type of biostimulant applied to the crop. The optimization of these materials is necessary for a successful application, by considering biostimulant concentration and dosage effects, plant developmental stage, climatic/environmental conditions, and experimental setup.
Therefore, the main limitations and areas for further investigation for future biostimulant and BCA optimization in sustainable viticulture are listed below:
● A) Manufacturing and commercialization
● Harmonization of legislation and lack of regulations on product quality
● Competitive commercialization costs
● Availability of raw materials used for their manufacturing, especially the lack of quality material (i.e. heterogeneous composition)
● Storage and effectivity duration (especially for biological products)
● B) Application
● Lack of information at field-scale, and very few reports on the application in uncontrolled realistic conditions with positive results
● Effect variability on the unknown plant microbiome
● Effect variability on plant growth, depending on product dosage (concentration and number of applications)
● Effect variability depending on the mode of product application (foliar spraying, or via soil irrigation/fertigation)
● Effect variability depending on the stage of crop development
● Speculation remains as to underlying mechanisms associated with the biostimulant
● Comparative studies with agrochemicals as control
● Synergetic effects with other products and potential negative effects
● Lack of standard protocols depending on grape cultivar
● C) Environmental and practical issues
● Soil management
● Environmental and practical issues
● Soil management for effect optimization
● Integration with agronomical management (e.g. avoiding tillage for AMF)
● Short and long-term environmental impact (e.g. contamination of soil and watersheds)
KJ: Conceptualization, Methodology, Writing - Original Draft; TLG: Writing - Review and Editing; PP-T, MS-M, YA, AD-A, AW, MG, MS, CP, JB, RO-H, MN, JR, and RA: Writing - Original Draft; SM and MD: Funding acquisition; GT: Writing - Original Draft, Writing - Review and Editing, Supervision and Funding acquisition. All authors approved the manuscript.
KJ wishes to acknowledge financial support (3710473400); MS-M thanks to RTI2018-099417-B-I00 (Spanish Ministry of Science, Innovation and Universities cofunded with EU FEDER funds); JB wish to acknowledge the Conselho Nacional de Desenvolvimento Científico e Tecnológico/Brasil (CNPQ process number 309477/2021-2); RO-H is supported by the Ramón y Cajal program from the MICINN (RYC-2017 22032), PAIDI 2020 (Ref. 20_00323), AEI GGOO 2020 (GOPC-CA-20-0001), “José Castillejo” program from the “Ministerio de Universidades” (CAS21/00125) and PID2019-106004RA-I00/AEI/10.13039/501100011033. SM and GT thanks to Ministerio de Ciencia e Innovación (grant PID2020-114330GB-100). PAIDI2020 from Junta de Andalucía, grant P18-RT-1401 to SM, MD, and GT is also acknowledged. GT acknowledge the support of the publication fee by the CSIC Open Access Publication Support Initiative through its Unit of Information Resources for Research (URICI).
Author YA was employed by Chitose Laboratory Corp.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Akhter, A., Hage-Ahmed, K., Soja, G., Steinkellner, S. (2015). Compost and biochar alter mycorrhization, tomato root exudation, and development of Fusarium oxysporum f. sp. lycopersici. Front. Plant Sci. 6. doi: 10.3389/fpls.2015.00529
Al-Karaki, G. N., Clark, R. B. (1998). Growth, mineral acquisition, and water use by mycorrhizal wheat grown under water stress. J. Plant Nutr. 21 (2), 263–276. doi: 10.1080/01904169809365401
Almeida, A. B. D., Concas, J., Campos, M. D., Materatski, P., Varanda, C., Patanita, M., et al. (2020). Endophytic fungi as potential biological control agents against grapevine trunk diseases in Alentejo region. Biology 9 (12), 420. doi: 10.3390/biology9120420
Amendola, C., Montagnoli, A., Terzaghi, M., Trupiano, D., Oliva, F., Baronti, S., et al. (2017). Short-term effects of biochar on grapevine fine root dynamics and arbuscular mycorrhizae production. Agric. Ecosyst. Environ. 239 236–245. doi: 10.1016/j.agee.2017.01.025
Andreolli, M., Lampis, S., Zapparoli, G., Angelini, E., Vallini, G. (2016). Diversity of bacterial endophytes in 3 and 15 year-old grapevines of Vitis vinifera cv. corvina and their potential for plant growth promotion and phytopathogen control. Microbiol. Res. 183, 42–52. doi: 10.1016/j.micres.2015.11.009
Andrés, P., Rosell-Melé, A., Colomer-Ventura, F., Denef, K., Cotrufo, M. F., Riba, M., et al. (2019). Belowground biota responses to maize biochar addition to the soil of a Mediterranean vineyard. Sci. Total Environ. 660, 1522–1532. doi: 10.1016/j.scitotenv.2019.01.101
An, X., Wu, Z., Yu, J., Cravotto, G., Liu, X., Li, Q., et al. (2020). Copyrolysis of biomass, bentonite, and nutrients as a new strategy for the synthesis of improved biochar-based slow-release fertilizers. ACS Sustain. Chem. Eng. 8 (8), 3181–3190. doi: 10.1021/acssuschemeng.9b06483
Arioli, T., Mattner, S. W., Hepworth, G., McClintock, D., McClinock, R. (2021). Effect of seaweed extract application on wine grape yield in Australia. J. Appl. Phycol. 33, 1883–1891. doi: 10.1007/s10811-021-02423-1
Arras, G. (1996). Mode of action of an isolate of Candida famata in biological control of Penicillium digitatum in orange fruits. Postharvest Biol. Tec. 8 (3), 191–198. doi: 10.1016/0925-5214(95)00071-2
Asghari, S., Harighi, B., Ashengroph, M., Clement, C., Aziz, A., Esmaeel, Q., et al. (2020). Induction of systemic resistance to Agrobacterium tumefaciens by endophytic bacteria in grapevine. Plant Pathol. 69 (5), 827–837. doi: 10.1111/ppa.13175
Aziz, A., Poinssot, B., Daire, X., Adrian, M., Bézier, A., Lambert, B., et al. (2003). Laminarin elicits defense responses in grapevine and induces protection against Botrytis cinerea and Plasmopara viticola. Mol. Plant-Microbe Interact. 16 (12), 1118–1128. doi: 10.1094/MPMI.2003.16.12.1118
Barka, E., Nowak, J., Clément, C. (2006). Enhancement of chilling resistance of inoculated grapevine plantlets with a plant growth-promoting rhizobacterium, Burkholderia phytofirmans strain PsJN. Appl. Environ. Microb. 72 (11), 7246–7252. doi: 10.1128/AEM.01047-06
Baronti, S., Vaccari, F. P., Miglietta, F., Calzolari, C., Lugato, E., Orlandini, S., et al. (2014). Impact of biochar application on plant water relations in Vitis vinifera (L.). Eur. J. Agron. 53, 38–44. doi: 10.1016/j.eja.2013.11.003
Bashan, Y., de-Bashan, L. E., Prabhu, S. R., Hernández, J. P. (2014). Advances in plant growth-promoting bacterial inoculant technology: Formulations and practical perspectives, (1998–2013). Plant Soil 378, 1–33. doi: 10.1007/s11104-013-1956-x
Belchim (2022) Vintec: A natureza trabalhando por si. Available at: https://belchim.pt/producten/vintec/ (Accessed July 30, 2022).
Benuzzi, M., Baldoni, G. (2000). AQ 10, a new biofungicide based on Ampelomyces quisqualis for powdery mildew control on grapes. Informatore Fitopatologico 50 (5), 33–36.
Boller, T. (1995). Chemoperception of microbial signals in plant cells. Annu Rev. Physical. Plant Mol. Biol. 46, 189–214. doi: 10.1146/annurev.pp.46.060195.001201
Boller, T., Felix, G. (2009). A ranaissance of elicitors: peception of microbe-associated molecular patterns and danger signals by pattern-recognition receptors. Annu. Rev. Plant Biol. 60 379–406. doi: 10.1146/annurev.arplant.57.032905.105346
Bonanomi, G., Ippolito, F., Scala, F. (2015). A “black” future for plant pathology? Biochar as a new soil amendment for controlling plant diseases. J. Plant Pathol. 97, 223–234. doi: 10.4454/jpp.v97i2.3381
Boselli, M., Bahouaoui, M. A., Lachhab, N., Sanzani, S. M., Ferrara, G., Ippolito, A. (2019). Protein hydrolysates effects on grapevine (Vitis vinifera L., cv. corvina) performance and water stress tolerance. Sci. Hortic-Amsterdam 258, 108784. doi: 10.1016/j.scienta.2019.108784
Botelho, M., Cruz, A., Ricardo- Da-Silva, J., De Castro, R., Ribeiro, H. (2020). Mechanical pruning and soil fertilization with distinct organic amendments in vineyards of syrah: Effects on vegetative and reproductive growth. Agronomy 10 (8), 10081090. doi: 10.3390/agronomy10081090
Bowles, T. M., Jackson, L. E., Loeher, M., Cavagnaro, T. R. (2017). Ecological intensification and arbuscular mycorrhizas: A meta-analysis of tillage and cover crop effects. J. Appl. Ecol. 54 (6), 1785–1793. doi: 10.1111/1365-2664.12815
Cabañas, C. M., Hernández, A., Martínez, A., Tejero, P., Vázquez-Hernández, M., Martín, A., et al. (2020). Control of Penicillium glabrum by indigenous antagonistic yeast from vineyards. Foods 9 (12), 1864. doi: 10.3390/foods9121864
Cabrera, L. C., Pastor, P. M. (2021). The 2019 European union report on pesticide residues in food. EFSA J. 19 (4), 6491. doi: 10.2903/j.efsa.2021.6491
Campos, C., Carvalho, M., Brígido, C., Goss, M. J., Nobre, T. (2018). Symbiosis specificity of the preceding host plant can dominate but not obliterate the association between wheat and its arbuscular mycorrhizal fungal partners. Front. Microbiol. 9 2920. doi: 10.3389/fmicb.2018.02920
Canellas, L. P., Olivares, F. L., Aguiar, N. O., Jones, D. L., Nebbioso, A., Mazzei, P. (2015). Humic and fulvic acids as biostimulants in horticulture. Sci. Hortic. 196, 15–27. doi: 10.1016/j.scienta.2015.09.013
Canellas, L. P., Spaccini, R., Piccolo, A., Dobbss, L. B., Okorokova-Façanha, A. L., Santos, G. A. (2009). Relationships between chemical characteristics and root growth promotion of humic acids isolated from Brazilian oxisols. Soil Sci. 174, 611–620. doi: 10.1097/SS.0b013e3181bf1e03
Carro-Huerga, G., Mayo-Prieto, S., Rodríguez-González, Á., González-López, Ó., Gutiérrez, S., Casquero, P. A. (2021). Influence of fungicide application and vine age on Trichoderma diversity as source of biological control agents. Agronomy 11 (3), 446. doi: 10.3390/agronomy11030446
Castellano-Hinojosa, A., Bedmar, E. J. (2017). “Methods for evaluating plant growth-promoting rhizobacteria traits”, in Advances in PGPR research. Eds. Harikesh, B., Singh, Birinchi, Sarma, K., Chetan, K. (CABI, USA), ISBN: 978-1-78639-032-5. ePDF.
Castellarin, S. D., Pfeiffer, A., Sivilotti, P., Degan, M., Peterlunger, E., Gaspero, G. (2007). Transcriptional regulation of anthocyanin biosynthesis in ripening fruits of grapevine under seasonal water deficit. Plant Cell Environ. 30 (11), 1381–1399. doi: 10.1111/j.1365-3040.2007.01716.x
Cataldo, E., Fucile, M., Mattii, G. B. (2022). Biostimulants in viticulture: A sustainable approach against biotic and abiotic stresses. Plants 11 (2), 162. doi: 10.3390/plants11020162
Chang, Y., Rossi, L., Zotarelli, L., Gao, B., Sarkhosh, A. (2021a). Greenhouse evaluation of pinewood biochar effects on nutrient status and physiological performance in muscadine grape (Vitis rotundifolia L.). HortScience 56 (2), 277–285. doi: 10.21273/HORTSCI15428-20
Chang, Y., Rossi, L., Zotarelli, L., Gao, B., Shahid, M. A., Sarkhosh, A. (2021b). Biochar improves soil physical characteristics and strengthens root architecture in muscadine grape (Vitis rotundifolia L.). Chem. Biol. Technol. Agric. 8, 7. doi: 10.1186/s40538-020-00204-5
Chan, Z., Tian, S. (2006). Induction of H2O2-metabolizing enzymes and total protein synthesis by antagonistic yeast and salicylic acid in harvested sweet cherry fruit. Postharvest Biol. Tec. 39 (3), 314–320. doi: 10.1016/j.postharvbio.2005.10.009
Chauhan, H., Bagyaraj, D. J., Selvakumar, G., Sundaram, S. P. (2015). Novel plant growth promoting rhizobacteria: Prospects and potential. Appl. Soil Ecol. 95, 38–53. doi: 10.1016/j.apsoil.2015.05.011
Chen, Y. H., Li, Y. F., Wei, H., Li, X. X., Zheng, H. T., Dong, X. Y., et al. (2020). Inhibition efficiency of wood vinegar on grey mould of table grapes. Food Biosci. 38, 100755. doi: 10.1016/j.fbio.2020.100755
Chervin, J., Romeo-Oliván, A., Fournier, S., Puech-Pages, V., Dumas, B., Jacques, A., et al. (2022). Modification of early response of Vitis vinifera to pathogens relating to esca disease and biocontrol agent Vintec® revealed by untargeted metabolomics on woody tissues. Front. Microbiol. 2. doi: 10.3389/fmicb.2022.835463
Clark, R. B., Zeto, S. K. (2000). Mineral acquisition by arbuscular mycorrhizal plants. J. Plant Nutr. 23 (7), 867–902. doi: 10.1080/01904160009382068
Colla, G., Hoagland, L., Ruzzi, M., Cardarelli, M., Bonini, P., Canaguier, R., et al. (2017). Biostimulant action of protein hydrolysates: Unraveling their effects on plant physiology and microbiome. Front. Plant Sci. 8. doi: 10.3389/fpls.2017.02202
Colla, G., Nardi, S., Cardarelli, M., Ertani, A., Lucini, L., Canaguier, R., et al. (2015). Protein hydrolysates as biostimulants in horticulture. Sci. Hortic-Amsterdam 196, 28–38. doi: 10.1016/j.scienta.2015.08.037
Colla, G., Rouphael, Y., Canaguier, R., Svecova, E., Cardarelli., M. (2014). Biostimulant action of a plant-derived protein hydrolysate produced through enzymatic hydrolysis. Front. Plant Sci., 5, 448. doi: 10.3389/fpls.2014.00448
Compant, S., Mitter, B., Colli-Mull, J., Gangl, H., Sessitsch, A. (2011). Endophytes of grapevine flowers, berries, and seeds: Identification of cultivable bacteria, comparison with other plant parts, and visualization of niches of colonization. Microb. Ecol. 62, 188–197. doi: 10.1007/s00248-011-9883-y
Cordero-Bueso, G., Mangieri, N., Maghradze, D., Foschino, R., Valdetara, F., Cantoral, J. M., et al. (2017). Wild grape-associated yeasts as promising biocontrol agents against Vitis vinifera fungal pathogens. Front. Microbiol. 8. doi: 10.3389/fmicb.2017.02025
da Silva, M. S. R. A., Huertas Tavares, O. C., Gonçalves Ribeiro, T., da Silva, C. S. R. A., da Silva, C. S. R. A., García-Mina, J. M., et al. (2021). Humic acids enrich the plant microbiota with bacterial candidates for the suppression of pathogens. Appl. Soil Ecol. 168, 104146. doi: 10.1016/j.apsoil.2021.104146
Debode, J., Ebrahimi, N., D'Hose, T., Cremelie, P., Viaene, N., Vandecasteele, B. (2020). Has compost with biochar added during the process added value over biochar or compost to increase disease suppression? Appl. Soil Ecol. 153, 103571. doi: 10.1016/j.apsoil.2020.103571
de Carvalho, R. P., Pasqual, M., de Oliveira Silveira, H. R., de Melo, P. C., Bispo, D. F. A., Laredo, R. R., et al. (2019). “Niágara rosada” table grape cultivated with seaweed extracts: Physiological, nutritional, and yielding behavior. J. Appl. Phycol. 31, 2053–2064. doi: 10.1007/s10811-018-1724-7
Deng, Q., Xia, H., Lin, L., Wang, J., Yuan, L., Li, K., et al. (2019). SUNRED, a natural extract-based biostimulant, application stimulates anthocyanin production in the skins of grapes. Sci. Rep. 9, 2590. doi: 10.1038/s41598-019-39455-0
Deolu-Ajayi, A. O., van der Meer, I. M., van der Werf, A., Karlova, R. (2022). The power of seaweeds as plant biostimulants to boost crop production under abiotic stress. Plant Cell Environ. 45, 2537–2553. doi: 10.1111/pce.14391
Dobbss, L. B., Santos, T. C., Pittarello, M., Souza, S. B., Ramos, A. C., Busato, J. G. (2018). Alleviation of iron toxicity in Schinus terebinthifolius raddi (Anacardiaceae) by humic substances. Environ. Sci. Poll. Res. 25, 9416–9425. doi: 10.1007/s11356-018-1193-1
Don, S. M. Y., Schmidtke, L. M., Gambetta, J. M., Steel, C. C. (2021). Volatile organic compounds produced by Aureobasidium pullulans induce electrolyte loss and oxidative stress in Botrytis cinerea and Alternaria alternata. Res. Microbiol. 172 (1), 103788. doi: 10.1016/j.resmic.2020.10.003
du Jardin, P. (2015). Plant biostimulants: Definition, concept, main categories and regulation. Sci. Horti-Amsterdam 196, 3–14. doi: 10.1016/j.scienta.2015.09.021
EFSA (2021). Consolidated Annual Activity Report 2021. European Food Safety Authority (EFSA). Available in: https://www.efsa.europa.eu/sites/default/files/2022-03/ar2021.pdf (last access in 29 09 22).
Egamberdieva, D., Reckling, M., Wirth, S. (2017). Biochar-based Bradyrhizobium inoculum improves growth of lupin (Lupinus angustifolius L.) under drought stress. Eu. J. Soil Biol. 78, 38–42. doi: 10.1016/j.ejsobi.2016.11.007
Egleton, M., Erdos, Z., Raymond, B., Matthews, A. C. (2021). Relative efficacy of biological control and cultural management for control of mollusc pests in cool climate vineyards. Biocontrol Sci. Techn. 31 (7), 725–738. doi: 10.1080/09583157.2021.1882387
Ehlers, R. U. (2011). “Regulation of biological control agents and the EU policy support action REBECA”, in Regulation of biological control agents (Dordrecht: Springer), 3–23. doi: 10.1007/978-90-481-3664-3
El Boukhari, M. E. M., Barakate, M., Bouhia, Y., Lyamlouli, K. (2020). Trends in seaweed extract based biostimulants: Manufacturing process and beneficial effect on soil-plant systems. Plants 9, 359. doi: 10.3390/plants9030359
El-Ghaouth, A., Wilson, C. L., Wisniewski, M. (1998). Ultrastructural and cytochemical aspects of the biological control of Botrytis cinerea by Candida saitoana in apple fruit. Phytopathology 88 (4), 282–291. doi: 10.1094/PHYTO.1998.88.4.282
El-Ghaouth, A., Wilson, C., Wisniewski, M. (2004). “Biologically-based alternatives to synthetic fungicides for the control of postharvest diseases of fruit and vegetables”, in Diseases of fruits and vegetables: Volume II (Dordrecht: Springer), 511–535.
Erdogan, U., Turan, M., Ates, F., Kotan, R., Çakmakçi, R., Erdogan, Y., et al. (2018). Effects of root plant growth promoting rhizobacteria inoculations on the growth and nutrient content of grapevine. Commun. Soil Sci. Plan. 49 (14), 1731–1738. doi: 10.1080/00103624.2018.1474910
Ertani, A., Cavani, L., Pizzeghello, D., Brandellero, E., Altissimo, A., Ciavatta, C., et al. (2009). Biostimulant activity of two protein hydrolyzates in the growth and nitrogen metabolism of maize seedlings. J. Plant Nutr. Soil Sci. 172, 237–244. doi: 10.1002/jpln.200800174
Ertani, A., Francisco, O., Ferrari, E., Schiavon, M., Nardi, S. (2018). Spectroscopic-chemical fingerprint and biostimulant activity of a protein-based product in solid form. Molecules 23, 1031. doi: 10.3390/molecules23051031
Ferrara, G., Brunetti, G. (2008). Influence of foliar applications of humic acids in Vitis vinifera L. cv italia. J. Int. Sci. Vigne Vin 42, 79–87. doi: 10.20870/oeno-one.2008.42.2.822
Ferrara, G., Brunetti, G. (2010). Effects of the times of application of a soil humic acid on berry quality of table grape (Vitis vinifera L.) cv italia. Spa. J. Agric. Res. 8, 817–822. doi: 10.5424/1283
Fitter, A. H., Garbaye, J. (1994). Interactions between mycorrhizal fungi and other soil organisms. Plant Soil 159 (1), 123–132. doi: 10.1007/BF00000101
Foley, J., Ramankutty, N., Brauman, K., Cassidy, E. S., Gerber, J. S., Johnston, M., et al. (2011). Solutions for a cultivated planet. Nature 478, 337–342. doi: 10.1038/nature10452
Frioni, T., Sabbatini, P., Tombesi, S., Norrie, J., Poni, S., Gatti, M., et al. (2018). Effects of a biostimulant derived from the brown seaweed Ascophyllum nodosum on ripening dynamics and fruit quality of grapevines. Sci. Hortic-Amsterdam 232, 97–106. doi: 10.1016/j.scienta.2017.12.054
Funes-Pinter, M. I., Salomon, M. V., Berli, F., Gil, R., Bottini, R., Piccoli, P. (2018). Plant growth promoting rhizobacteria alleviate stress by AsIII in grapevine. Agr. Ecosyst. Environ. 267, 100–108. doi: 10.1016/j.agee.2018.08.015
Gao, X., Cheng, H. Y., Del Valle, I., Liu, S., Masiello, C. A., Silberg, J. J. (2016). Charcoal disrupts soil microbial communication through a combination of signal sorption and hydrolysis. ACS Omega 1, 226–233. doi: 10.1021/acsomega.6b00085
Garrido, L. D. R., Botton, M. (2020). Agrotóxicos registrados para a cultura da Videira-Safra 945 2020/21. Embrapa Uva e Vinho-Comunicado Técnico (INFOTECA-E). Available at: https://www.embrapa.br/busca-de-publicacoes/-/publicacao/1124869/agrotoxicos-registrados-para-a-cultura-da-videira—safra-202021.
Gauthier, A., Trouvelot, S., Kelloniemi, J., Grettinger, P., Wendehenne, D., Daire, X., et al. (2014). The sulfated laminarin triggers a stress transcriptome before priming the SA- and ROS-dependent defenses during grapevine’s induced resistance against Plasmppara viticola. PLoS One 9, e88145. doi: 10.1371/journal.pone.0194327
Gava, A., Emer, C. D., Ficagna, E., Fernandes de Andrade, S., Fuentefria, A. M. (2021). Occurrence and impact of fungicides residues on fermentation during wine production–a review. Food Addit. Contam, 38(6), 1–19. doi: 10.1080/19440049.2021.1894357
Genesio, L., Miglietta, F., Baronti, S., Vaccari, F. P. (2015). Biochar increases vineyard productivity without affecting grape quality: Results from a four years field experiment in Tuscany. Agric. Ecosyst. Environ. 201, 20–25. doi: 10.1016/j.agee.2014.11.021
Giagnoni, L., Maienza, A., Baronti, S., Vaccari, F. P., Genesio, L., Taiti, C., et al. (2019). Long-term soil biological fertility, volatile organic compounds and chemical properties in a vineyard soil after biochar amendment. Geoderma 344, 127–136. doi: 10.1016/j.geoderma.2019.03.011
Głodowska, M., Husk, B., Schwinghamer, T., Smith, D. (2016). Biochar is a growth-promoting alternative to peat moss for the inoculation of corn with a pseudomonad. Agron. Sustain. Dev. 36 (1), 1–10. doi: 10.1007/s13593-016-0356-z
Godfray, H. C., Beddington, J. R., Crute, I. R., Haddad, L., Lawrence, D., Muir, J. F., et al. (2010). Food security: the challenge of feeding 9 billion people. Science 327 (5967), 812–818. doi: 10.1126/science.1185383
Gosling, P., Hodge, A., Goodlass, G., Bending, G. D. (2006). Arbuscular mycorrhizal fungi and organic farming. Agr. Ecosyst. Environ. 113 (1), 17–35. doi: 10.1016/j.agee.2005.09.009
Goswami, D., Thakker, J. N., Dhandhukia, P. C. (2016). Portraying mechanics of plant growth promoting rhizobacteria (PGPR): A review. Cogent Food Agric. 2, 1127500. doi: 10.1080/23311932.2015.1127500
Graber, E. R., Frenkel, O., Jaiswal, A. K., Elad, Y. (2014). How may biochar influence severity of diseases caused by soilborne pathogens? Carbon Manage. 5 (2), 169–183. doi: 10.1080/17583004.2014.913360
Graber, E. R., Harel, Y. M., Kolton, M., Cytryn, E., Silber, A., David, D. R., et al. (2010). Biochar impact on development and productivity of pepper and tomato grown in fertigated soilless media. Plant Soil 337, 481–496. doi: 10.1007/s11104-010-0544-6
Grewal, A., Abbey, L. E., Gunupuru, L. R. (2018). Production, prospects and potential application of pyroligneous acid in agriculture. J. Ana. Appl. Pyrol. 135, 152–159. doi: 10.1016/j.jaap.2018.09.008
Gryndler, M. (2000). Interactions of arbuscular mycorrhizal fungi with other soil organisms BT - arbuscular mycorrhizas: Physiology and function. Springer (USA) Eds. Kapulnik, Y., Douds, D. D. doi: 10.1007/978-94-017-0776-3_11
Gupta, M. M., Aggarwal, A., Asha (2018). From mycorrhizosphere to rhizosphere microbiome: The paradigm shift BT- root biology. Springer (USA) Eds. Giri, R., Prasad, Varma, A. doi: 10.1007/978-3-319-75910-4_20enta.2017.12.054
Gutiérrez-Gamboa, G., Romanazzi, G., Garde-Cerdán, T., Pérez-Álvarez, E. P. (2019). A review of the use of biostimulants in the vineyard for improved grape and wine quality: Effects on prevention of grapevine diseases. J. Sci. Food Agric. 99, 1001–1009. doi: 10.1002/jsfa.9353
Haidar, R., Fermaud, M., Calvo-Garrido, C., Roudet, J., Deschamps, A. (2016). Modes of action for biological control of Botrytis cinerea by antagonistic bacteria. Phytopathol. Mediterr, 55(3), 301–322. doi: 10.14601/Phytopathol_Mediterr-18079
Hale, L., Luth, M., Crowley, D. (2015). Biochar characteristics relate to its utility as an alternative soil inoculum carrier to peat and vermiculite. Soil Biol. Biochem. 81, 228–235. doi: 10.1016/j.soilbio.2014.11.023
Hardy, K., Knight, J. D. (2021). Evaluation of biochars as carriers for Rhizobium leguminosarum. Can. J. Microbiol. 67 (1), 53–63. doi: 10.1139/cjm-2020-0416
Harel, Y. M., Elad, Y., Rav-David, D., Borenstein, M., Shulchani, R., Lew, B., et al. (2012). Biochar mediates systemic response of strawberry to foliar fungal pathogens. Plant Soil 357, 245–257. doi: 10.1007/s11104-012-1129-3
Heckman, D. S., Geiser, D. M., Eidell, B. R., Stauffer, R. L., Kardos, N. L., Hedges, S. B. (2001). Molecular evidence for the early colonization of land by fungi and plants. Science 293 (5532), 1129–1133. doi: 10.1126/science.1061457
Holland, T., Bowen, P., Kokkoris, V., Urbez-Torres, J. R., Hart, M. (2019). Does inoculation with arbuscular mycorrhizal fungi reduce trunk disease in grapevine rootstocks? Horticulturae 5 (3), 61. doi: 10.3390/horticulturae5030061
Ibrahim, M. M., Ali, A. A. (2016). Effect of humic acid on productivity and quality of superior seedless grape cultivar. Middle East J. Agric. Res. 5, 239–246.
Irani, H., Kaji, B. V., Naeini, M. R. (2021). Biostimulant-induced drought tolerance in grapevine is associated with physiological and biochemical changes. Chem. Biol. Technol. Agric. 8, 5. doi: 10.1186/s40538-020-00200-9
İşçı, B., Kacar, E., Altindışlı, A. (2019). Effects of IBA and plant growth-promoting rhizobacteria (PGPR) on rooting of ramsey American grapevine rootstock. Appl. Ecol. Environ. Res. 17 (2), 4693–4705. doi: 10.15666/aeer/1702_46934705
Ito, Y., Nakanomyo, I., Motose, H., Iwamoto, K., Sawa, S., Dohmae, N., et al. (2006). Dodeca-CLE peptides as suppressors of plant stem cell differentiation. Science 313 313(5788), 842–845. doi: 10.1126/science.1128436
Jamalizadeh, M., Etebarian, H. R., Aminian, H., Alizadeh, A. (2011). A review of mechanisms of action of biological control organisms against post harvest fruit spoilage. Eppo Bull. 41 (1), 65–71. doi: 10.1111/j.1365-2338.2011.02438.x
Jeandet, P., Douillet-Breuil, A. C., Bessis, R., Debord, S., Sbaghi, M., Adrian, M. (2002). Phytoalexins from the vitaceae: Biosynthesis, phytoalexin gene expression in transgenic plants, antifungal activity, and metabolism. J. Agr. Food Chem. 50 (10), 2731–2741. doi: 10.1021/jf011429s
Jeffries, P., Gianinazzi, S., Perotto, S., Turnau, K., Barea, J. M. (2003). The contribution of arbuscular mycorrhizal fungi in sustainable maintenance of plant health and soil fertility. Biol. Fert. Soils 37 (1), 1–16. doi: 10.1007/s00374-002-0546-5
Jindo, K., Olivares, F. L., Malcher, D. J. P., Sánchez-Monedero, M. A., Kempenaar, C., Canellas, L. P. (2020). From lab to field: Role of humic substances under open-field and greenhouse conditions as biostimulant and biocontrol agent. Front. Plant Sci. 11. doi: 10.3389/fpls.2020.00426
Jones, L., Riaz, S., Morales-Cruz, A., Amrine, K. C. H., McGuire, B., Gubler, W. D., et al. (2014). Adaptive genomic strucltural variation in the grape powdery mildew pathogen, Erysiphe necator. BMC Genomics 15 1081. doi: 10.1186/1471-2164-15-1081
Joseph, S., Graber, E. R., Chia, C., Munroe, P., Donne, S., Thomas, T., et al. (2013). Shifting paradigms: Development of high-efficiency biochar fertilizers based on nano-structures and soluble components. Carbon Manage. 4 (3), 323–343. doi: 10.4155/cmt.13.23
Kamble, M. V., Joshi, S. M., Hadimani, S., Jogaiah, S. (2021). O Biopriming com Trichoderma harzianum da rizosfera promove proteção contra o míldio da videira, desencadeando respostas de defesa histopatológicas e bioquímicas. Rhizosphere 19, 100398. doi: 10.1016/j.rhisph.2021.100398
Kanwal, S., Batool, A., Ghufran, M. A., Khalid, A. (2018). Effect of dairy manure derived biochar on microbial biomass carbon, soil carbon and Vitis vinifera under water stress conditions. Pak. J. Bot. 50 (5), 1713–1718.
Karoglan, M., Radić, T., Anić, M., Andabaka, Ž., Stupić, D., Tomaz, I., et al. (2021). Mycorrhizal fungi enhance yield and berry chemical composition of in field grown “Cabernet Sauvignon” grapevines (V. vinifera L.). Agriculture 11 (7), 615. doi: 10.3390/agriculture11070615
Khairy, H. M., El-Shafay, S. M. (2013). Seasonal variations in the biochemical composition of some common seaweed species from the coast of Abu qir bay, Alexandria, Egypt. Oceanologia 55, 435–452. doi: 10.5697/oc.55-2.435
Kloepper, J. W., Schroth, M. N. (1979). Plant growth promoting rhizobacteria: evidence that the mode of action involves root microflora interactions. Phytophatology 69, 1034.
Köse, C., Güleryüz, M., Şahin, F., Demirtaş, I. (2003). Effects of some plant growth promoting rhizobacteria (PGPR) on rooting of grapevine rootstocks. Acta Agrobot. 56 (1-2), 47–52. doi: 10.5586/aa.2003.005
Köse, C., Güleryüz, M., Şahin, F., Demirtaş, I. (2015). Effects of some plant growth promoting rhizobacteria (PGPR) on graft union of grapevine. J. Sustain. Agr. 26 (2), 139–147. doi: 10.1300/J064v26n02_10
Krishna, H., Singh, S. K., Sharma, R. R., Khawale, R. N., Grover, M., Patel, V. B. (2005). Biochemical changes in micropropagated grape (Vitis vinifera L.) plantlets due to arbuscular-mycorrhizal fungi (AMF) inoculation during ex vitro acclimatization. Sci. Hortic-Amsterdam 106, 554–567. doi: 10.1016/j.scienta.2005.05.009
Kumar Jha, C., Saraf, M. (2015). Plant growth promoting rhizobacteria (PGPR): a review. E3 J. Agric. Res. Dev. 5 (2), 0108–0119. doi: 10.13140/RG.2.1.5171.2164
Kurtzman, C. P., Fell, J. W., Boekhout, T. (2011). The yeasts: A taxonomic study. Burlington: Elsevier science. Elsevier, Amsterdam, Nederland.
Lachhab, N., Sanzani, S. M., Adrian, M., Chiltz, A., Balacey, S., Boselli, M., et al. (2014). Soybean and casein hydrolysates induce grapevine immune responses and resistance against Plasmopara viticola. Front. Plant Sci. 5 716. doi: 10.3389/fpls.2014.00716
Lachhab, N., Sanzani, S. M., Bahouaoui, M. A., Boselli, M., Ippolito, A. (2016). Effect of some protein hydrolysates against gray mould of table and wine grapes. Eur. J. Plant Pathol. 144 (4), 821–830. doi: 10.1007/s10658-015-0749-x
Lehmann, J., Rillig, M. C., Thies, J., Masiello, C. A., Hockaday, W. C., Crowley, D. (2011). Biochar effects on soil biota - a review. Soil Biol. Biochem. 43 (9), 1812–1836. doi: 10.1016/j.soilbio.2011.04.022
Linderman, R. G., Davis, E. A. (2001). Comparative response of selected grapevine rootstocks and cultivars to inoculation with different mycorrhizal fungi. Am. J. Enol. Viticult. 52 (1), 8–11.
Liu, M., Liu, X., Cheng, B., Ma, X. L., Lyu, X. T., Zhao, X. F., et al. (2016). Selection and evaluation of phosphate-solubilizing bacteria from grapevine rhizospheres for use as biofertilizers. Span. J. Agric. Res. 14 (4), e1106. doi: 10.5424/sjar/2016144-9714
Li, H. Y., Yang, G. D., Shu, H. R., Yang, Y. T., Ye, B. X., Nishida, I., et al. (2006). Colonization by the arbuscular mycorrhizal fungus Glomus versiforme induces a defense response against the root-knot nematode Meloidogyne incognita in the grapevine (Vitis amurensis rupr.), which includes transcriptional activation of the class III chitin. Plant Cell Physiol. 47 (1), 154–163. doi: 10.1093/pcp/pci231
Lu, H., Wu, Z., Wang, W., Xu, X., Liu, X. (2020). Rs-198 liquid biofertilizers affect microbial community diversity and enzyme activities and promote Vitis vinifera L. growth. BioMed. Res. Int., 8321462. doi: 10.1155/2020/8321462
Maachia, B. S., Rafik, E., Chérif, M., Nandal, P., Mohapatra, T., Bernard, P. (2015). Biological control of the grapevine diseases ‘grey mold’and ‘powdery mildew’by Bacillus B27 and B29 strains. Indian J. Exp. Biol. 53 (2), 109–115.
Maienza, A., Baronti, S., Cincinelli, A., Martellini, T., Grisolia, A., Miglietta, F., et al. (2017). Biochar improves the fertility of a Mediterranean vineyard without toxic impact on the microbial community. Agron. Sustain. Dev. 37, 47. doi: 10.1007/s13593-017-0458-2
Malviya, D., Thosar, R., Kokare, N., Pawar, S., Singh, U. B., Saha, S., et al. (2022). A comparative analysis of microbe-based technologies developed at ICAR-NBAIM against Erysiphe necator causing powdery mildew disease in grapes (Vitis vinifera l.). Front. Microbiol. 13, 871901. doi: 10.1400/53262
Mancuso, S., Azzarello, E., Mugnai, S., Briand, X.. (2006). Marine bioactive substances (IPA extract) improve foliar ion uptake and water stress tolerance in potted Vitis vinifera plants. Adv. Hortic. Sci. 20, 156–161. doi: 10.1400/53262
Meggio, F., Trevisan, S., Manoli, A., Ruperti, B., Quaggiotti, S. (2020). Systematic investigation of the effects of a novel protein hydrolysate on the growth, physiological parameters, fruit development and yield of grapevine (Vitis vinifera L., cv Sauvignon Blanc) under water stress conditions. Agronomy 10, 1785. doi: 10.3390/agronomy10111785
Miotto-Vilanova, L., Jacquard, C., Courteaux, B., Wortham, L., Michel, J., Clément, C., et al. (2016). Burkholderia phytofirmans PsJN confers grapevine resistance against Botrytis cinerea via a direct antimicrobial effect combined with a better resource mobilization. Front. Plant Sci. 7. doi: 10.3389/fpls.2016.01236
Montagnoli, A., Baronti, S., Alberto, D., Chiatante, D., Scippa, G. S., Terzaghi, M. (2021). Pioneer and fibrous root seasonal dynamics of Vitis vinifera L. are affected by biochar application to a low fertility soil: A rhizobox approach. Sci. Total Environ. 751, 141455. doi: 10.1016/j.scitotenv.2020.141455
Monteiro, E., Gonçalves, B., Cortez, I., Castro, I. (2022). The role of biostimulants as alleviators of biotic and abiotic stresses in grapevine: A review. Plants 31, 11(3):396. doi: 10.3390/plants11030396
Muganu, M., Paolocci, M., Bignami, C., Di Mattia, E. (2015). Enhancement of adventitious root differentiation and growth of in vitro grapevine shoots inoculated with plant growth promoting rhizobacteria. Vitis 54, 73–77. doi: 10.5073/vitis.2015.54.73-77
Mungkunkamchao, T., Kesmala, T., Pimratch, S., Toomsan, B., Jothityangkoon, D. (2013). Wood vinegar and fermented bioextracts: Natural products to enhance growth and yield of tomato (Solanum lycopersicum L.). Sci. Horti-Amsterdam 154, 66–72. doi: 10.1016/j.scienta.2013.02.020
Nesbitt, A., Dorling, S., Lovett, A. (2018). A suitability model for viticulture in England and Wales: Opportunities for investment, sector growth and increased climate resilience. J. Land Use Sci., 13(4), 414–438. doi: 10.1080/1747423X.2018.1537312
Nesler, A., Perazzolli, M., Puopolo, G., Giovannini, O., Elad, Y., Pertot, I. (2015). A complex protein derivative acts as biogenic elicitor of grapevine resistance against powdery mildew under filed conditions. Front. Plant Sci. 6. doi: 10.3389/fpls.2015.00715
Nogales, A., Aguirreolea, J., Santa María, E., Camprubí, A., Calvet, C. (2008). Response of mycorrhizal grapevine to Armillaria mellea inoculation: Disease development and polyamines. Plant Soil 317 (1), 177. doi: 10.1007/s11104-008-9799-6
Nogales, A., Rottier, E., Campos, C., Victorino, G., Costa, J. M., Coito, J. L., et al. (2021). The effects of field inoculation of arbuscular mycorrhizal fungi through rye donor plants on grapevine performance and soil properties. Agric. Ecosyst. Environ. 313, 107369. doi: 10.1016/j.agee.2021.107369
Ogawa, M., Okimori, Y. (2010). Pioneering works in biochar research, Japan. Aust. J. Soil Res. 48 (6-7), 489–500. doi: 10.1071/SR10006
Olavarrieta, C. E., Sampedro, M. C., Vallejom, A., Štefelovám, N., Barriom, R. J., De Diego, N. (2022). Biostimulants as an alternative to improve the wine quality from Vitis vinifera (cv. tempranillo) in La Rioja. Plants 11 (12), 1594. doi: 10.3390/plants11121594
Oleńska, E., Małek, W., Wójcik, M., Swiecicka, I., Thijs, S., Vangronsveld, J. (2020). Beneficial features of plant growth-promoting rhizobacteria for improving plant growth and health in challenging conditions: A methodical review. Sci. Total Environ. 743, 140682. doi: 10.1016/j.scitotenv.2020.140682
Olivares, F. L., Busato, J. G., de Paula, A. M., Silva, L. L., Aguiar, N. O., Canellas, L. P. (2017). Plant growth promoting bacteria and humic substances: Crop promotion and mechanisms of action. Chem. Biol. Technol. Agric. 4, 30. doi: 10.1186/s40538-017-0112-x
Pacifico, D., Squartini, A., Crucitti, D., Barizza, E., Lo Schiavo, F., Muresu, R., et al. (2019). The role of the endophytic microbiome in the grapevine response to environmental triggers. Front. Plant Sci. 10. doi: 10.3389/fpls.2019.01256
Payen, F. T., Sykes, A., Aitkenhead, M., Alexander, P., Moran, D., MacLeod, M. (2021). Soil organic carbon sequestration rates in vineyard agroecosystems under different soil management practices: A meta-analysis. J. Clea. Prod. 290, 125736. doi: 10.1016/j.jclepro.2020.125736
Pertot, I., Longa, C. M. O., Savazzini, F., Prodorutti, D., Michelon, L. (2015). Trichoderma atroviride SC1 for biocontrol of fungal diseases in plants, International Patent Application US 8,431,120B2.
Petit, E., Gubler, W. D. (2006). Influence of Glomus intraradices on black foot disease caused by Cylindrocarpon macrodidymum on Vitis rupestris under controlled conditions. Plant Dis. 90 (12), 1481–1484. doi: 10.1094/PD-90-1481
Petrillo, M., Zanotelli, D., Lucchetta, V., Aguzzoni, A., Tagliavini, M., Andreotti, C. (2020). The use of biochar as soil amendment: Effects on nitrogen and water availability for potted grapevines. Italus Hortus 27 (2), 28–40. doi: 10.26353/j.itahort/2020.2.2840
Phelan, M., Aherne, A., FitzGerald, R. J., O’Brien, N. M. (2009). Casein-derived bioactive peptides: Biological effects, industrial uses, safety aspects and regulatroy status. Int. Dairy J. 19, 643–654. doi: 10.1016/j.idairyj.2009.06.001
Pinto, C., Gomes, A. C. (2016). Vitis vinifera microbiome: from basic research to technological development. BioControl 61, 243–256. doi: 10.1007/s10526-016-9725-4
Popescu, G. G. C., Popescu, M. (2018). Yield, berry quality and physiological response of grapevine to foliar humic acid application. Bragantia 77, 273–282. doi: 10.1590/1678-4499.2017030
Porter, S. S., Sachs, J. L. (2020). Agriculture and the disruption of plant–microbial symbiosis. Trends Ecol. Evol. 35 (5), 426–439. doi: 10.1016/j.tree.2020.01.006
Pugliese, M., Monchiero, M., Gullino, M. L., Garibaldi, A. (2018). Application of laminarin and calcium oxide for the control of grape powdery mildew on Vitis vinifera cv. moscato. J. Plant Dis. Prot. 125, 477–482. doi: 10.1007/s41348-018-0162-8
Rahman, L., Whitelaw-Weckert, M. A., Orchard, B. (2014). Impact of organic soil amendments, including poultry-litter biochar, on nematodes in a riverina, new south Wales, vineyard. Soil Res. 52 (6), 604–619. doi: 10.1071/SR14041
Ranade, Y., Sawant, I., Saha, S., Chandrashekar, M., Pathak, P. (2021). Epiphytic microbial diversity of Vitis vinifera fructosphere: Present status and potential applications. Curr. Microbiol., 78, 1–13. doi: 10.1007/s00284-021-02385-0
Regulation EC 1107 (2009) Regulation (EC) no 1107/2009 of the European parliament and of the council of 21 October 2009 concerning the placing of plant protection products on the market and repealing council directives 79/117/EEC and 91/414/EEC. Available at: https://eur-lex.europa.eu/legal-content/EN/ALL/?uri=CELEX:32009R1107.
Regulation EU 1009 (2019) Regulation EU 2019/1009 of the European parliament and of the council of 5 June 2019 laying down rules on the making available on the market of EU fertilising products and amending regulations (EC) no 1069/2009 and (EC) no 1107/2009 and repealing regulation (EC) no 2003/2003. Available at: https://eur-lex.europa.eu/legal-content/EN/TXT/HTML/?uri=CELEX:32019R1009&from=ES.
Reineke, A., Thiéry, D. (2016). Grapevine insect pests and their natural enemies in the age of global warming. J. Pest Sci. 89 (2), 313–328. doi: 10.1007/s10340-016-0761-8
Rezgui, A., Ben Ghnaya-Chakroun, A., Vallance, J., Bruez, E., Hajlaoui, M. R., Sadfi-Zouaoui, N., et al. (2016). Endophytic bacteria with antagonistic traits inhabit the wood tissues of grapevines from Tunisian vineyards. Biol. Control 99, 28–37. doi: 10.1016/j.biocontrol.2016.04.005
Roka, K. (2022). “Anthropocene and climate change,” in Climate action, encyclopedia of the UN sustainable development goals. Ed. Leal Filho, W., et al (Nature Switzerland: Springer), 2020. doi: 10.1007/978-3-319-95885-9
Rolli, E., Marasco, R., Saderi, S., Corretto, E., Mapelli, F., Cherif, A., et al. (2017). Root-associated bacteria promote grapevine growth: from the laboratory to the field. Plant Soil 410 (1–2), 369–382. doi: 10.1007/s11104-016-3019-6
Romanazzi, G., Mancini, V., Feliziani, E., Andrea, S., Endeshaw, S., Neri, D. (2016). Impact of alternative fungicides on grape downy mildew control and vine growth and development. Plant Dis. 100 (4), 739–748. doi: 10.1094/PDIS-05-15-0564-RE
Ronga, D., Francia, E., Allesina, G., Pedrazzi, S., Zaccardelli, M., Pane, C., et al. (2019). Valorization of vineyard by-products to obtain composted digestate and biochar suitable for nursery grapevine (Vitis vinifera L.) production. Agronomy 9 (8), 420. doi: 10.3390/agronomy9080420
Rosa, D., Pogiatzis, A., Bowen, P., Kokkoris, V., Richards, A., Holland, T., et al. (2020). Performance and establishment of a commercial mycorrhizal inoculant in viticulture. Agriculture 10 (11), 539. doi: 10.3390/agriculture10110539
Rouphael, Y., Colla, G. (2020). Editorial: Biostimulants in agriculture. Front. Plant Sci. 11. doi: 10.3389/fpls.2020.00040
Ruzzi, M., Aroca, R. (2015). Plant growth-promoting rhizobacteria act as biostimulants in horticulture. Sci. Hortic-Amsterdam 196, 124–134. doi: 10.1016/j.scienta.2015.08.042
Sabir, A. (2013). Improvement of grafting efficiency in hard grafting grape berlandieri hybrid rootstocks by plant growth-promoting rhizobacteria (PGPR). Sci. Hortic-Amsterdam 164, 24–29. doi: 10.1016/j.scienta.2013.08.035
Sabir, A., Karaca, U., Yazar, K., Sabir, F. K., Yazici, M. A., Dogan, O., et al. (2017). Vine growth and yield response of alphonse lavallée (V. vinifera L.) grapevines to plant growth promoting rhizobacteria under alkaline condition in soilless culture. Acta Sci. Pol-Hortoru. 16 (4), 25–32. doi: 10.24326/asphc.2017.4.3
Salomon, M. V., Bottini, R., de Souza Filho, G. A., Cohen, A. C., Moreno, D., Gil, M., et al. (2014). Bacteria isolated from roots and rhizosphere of Vitis vinifera retard water losses, induce abscisic acid accumulation and synthesis of defense-related terpenes in in vitro cultured grapevine. Physiol. Plantarum 151 (4), 359–374. doi: 10.1111/ppl.12117
Salomon, M. V., Piccoli, P., Funes-Pinter, I., Stirk, W. A., Kulkarni, M., van Staden, J., et al. (2017). Bacteria and smoke-water extract improve growth and induce the synthesis of volatile defense mechanisms in Vitis vinifera l. Plant Physiol. Biochem. 120, 1–9. doi: 10.1016/j.plaphy.2017.09.013
Salomon, M. V., Purpora, R., Bottini, R., Piccoli, P. (2016). Rhizosphere associated bacteria trigger accumulation of terpenes in leaves of Vitis vinifera L. cv. Malbec that protect cells against reactive oxygen species. Plant Physiol. Biochem. 106, 295–304. doi: 10.1016/j.plaphy.2016.05.007
Samad, A., Antonielli, L., Sessitsch, A., Compant, S., Trognitz, F. (2017). Comparative genome analysis of the vineyard weed endophyte Pseudomonas viridiflava CDRTc14 showing selective herbicidal activity. Sci. Rep. 7 (1), 1–15. doi: 10.1038/s41598-017-16495-y
Sánchez-Monedero, M. A., Cayuela, M. L., Sánchez-García, M., Vandecasteele, B., D’Hose, T., López, G., et al. (2019). Agronomic evaluation of biochar, compost and biochar-blended compost across different cropping systems: Perspective from the European project FERTIPLUS. Agronomy 9 (5), 225. doi: 10.3390/agronomy9050225
Sánchez-Sánchez, A., Sánchez-Andreu, J., Juárez, M., Jordá, J., Bermúdez, D. (2006). Improvement of iron uptake in table grape by addition of humic substances. J. Plant Nutr. 29, 259–272. doi: 10.1080/01904160500476087
Santos, J. A., Fraga, H., Malheiro, A. C., Moutinho-Pereira, J., Dinis, L. T., Correia, C., et al. (2020). A review of the potential climate change impacts and adaptation options for European viticulture. App. Sci. 10 (9), 3092. doi: 10.3390/app10093092
Schaafsma, G. (2009). Safety of protein hydrolysates, fractions thereof and bioactive peptides in human nutrition. Eur. J. Clin. Nutr. 63, 1161–1168. doi: 10.1038/ejcn.2009.56
Schmidt, H. P., Kammann, C., Niggli, C., Evangelou, M. W. H., Mackie, K. A., Abiven, S. (2014). Biochar and biochar-compost as soil amendments to a vineyard soil influences on plant growth, nutrient uptake, plant health and grape quality. Agric. Ecosyst. Environ. 191, 117–123. doi: 10.1016/j.agee.2014.04.001
Schreiner, P. R. (2007). Effects of native and nonnative arbuscular mycorrhizal fungi on growth and nutrient uptake of ‘Pinot noir’ (Vitis vinifera l.) in two soils with contrasting levels of phosphorus. Appl. Soil Ecol. 36, 205–215. doi: 10.1016/j.apsoil.2007.03.002
Seufert, V., Ramankutty, N., Foley, J. (2012). Comparing the yields of organic and conventional agriculture. Nature 485, 229–232. doi: 10.1038/nature11069
Sharma, R. R., Singh, D., Singh, R. (2009). Biological control of postharvest diseases of fruits and vegetables by microbial antagonists: A review. Biol. Control 50 (3), 205–221. doi: 10.1016/j.biocontrol.2009.05.001
Shukla, P. S., Mantin, E. G., Adil, M., Bajpai, S., Critchley, A. T., Prithiviraj, B. (2019). Ascophyllum nodosum-based biostimulants: Sustainable applications in agriculture for the stimulation of plant growth, stress tolerance, and disease management. Front. Plant Sci. 10. doi: 10.3389/fpls.2019.00655
Sihem, B. M., Rafik, E., Florence, M., Mohamed, C., Ahmed, L. (2011). Identification and partial characterization of antifungal and antibacterial activities of two Bacillus sp. strains isolated from salt soil in Tunisia. Afr. J. Microbiol. Res. 5 (13), 1599–1608. doi: 10.5897/AJMR11.0
Sipiczki, M. (2006). Metschnikowia strains isolated from botrytized grapes antagonize fungal and bacterial growth by iron depletion. App. Environ. Microbiol. 72 (10), 6716–6724. doi: 10.1128/AEM.01275-06
Smith, S. E., Jakobsen, I., Grønlund, M., Smith, F. A. (2011). Roles of arbuscular mycorrhizas in plant phosphorus nutrition: Interactions between pathways of phosphorus uptake in arbuscular mycorrhizal roots have important implications for understanding and manipulating plant phosphorus acquisition. Plant Physiol. 156 (3), 1050–1057. doi: 10.1104/pp.111.174581
Smith, S. E., Read, D. (2008). Mycorrhizal symbiosis. 3rd ed. (Academic Press, Elsevier, Amsterdam, Neterland). doi: 10.1016/B978-0-12-370526-6.X5001-6
Sohn, B. K., Kim, K. Y., Chung, S. J., Kim, W. S., Park, S. M., Kang, J. G., et al. (2003). Effect of the different timing of AMF inoculation on plant growth and flower quality of chrysanthemum. Sci. Hortic.Amsterdam 98 (2), 173–183. doi: 10.1016/S0304-4238(02)00210-8
Soja, G., Wimmer, B., Rosner, F., Faber, F., Dersch, G., von Chamier, J., et al. (2018). Compost and biochar interactions with copper immobilisation in copper-enriched vineyard soils. App. Geochem. 88, 40–48. doi: 10.1016/j.apgeochem.2017.06.004
Souza, A. C., Olivares, F. L., Peres, L. E. P., Piccolo, A., Canellas, L. P. (2022). Plant hormone crosstalk mediated by humic acids. Chem. Biol. Technol. Agric. 9, 29. doi: 10.1186/s40538-022-00295-2
Stenberg, J. A., Sundh, I., Becher, P. G., Björkman, C., Dubey, M., Egan, P. A., et al. (2021). When is it biological control? A framework of definitions, mechanisms, and classifications. J. Pest Sci. 94 (3), 665–676. doi: 10.1007/s10340-021-01354-7
Strzelczyk, E., Dahm, H., Pachlewski, R. (1991). B-group vitamins production by mycorrhizal fungi in response to pH (in vitro studies). Plant Soil 137 (2), 237–241. doi: 10.1007/BF00011202
Torres, N., Goicoechea, N., Morales, F., Antolín, M. C. (2016). Berry quality and antioxidant properties in Vitis vinifera cv. tempranillo as affected by clonal variability, mycorrhizal inoculation and temperature. Crop Pasture Sci. 67, 961–977. doi: 10.1071/CP16038
Trouvelot, S., Bonneau, L., Redecker, D., van Tuinen, D., Adrian, M., Wipf, D. (2015). Arbuscular mycorrhiza symbiosis in viticulture: A review. Agron. Sustain. Dev. 35 (4), 1449–1467. doi: 10.1007/s13593-015-0329-7
Turan, M., Köse, C. (2004). Seaweed extracts improve copper uptake of grapevine. Acta Agric. Scand. Sect. B Soil Plant Sci. 54, 213–220. doi: 10.1080/09064710410030311
Ubalde, J. M., Payan, E., Sort, X., Rosas, J. G., Gómez, N. (2014). Application of biochar amendments to Mediterranean soils Effects on vine growth and grape quality. Geophys Res. Abstr. 16, 440000.
van Aubel, G., Buonatesta, R., Van Cutsem, P. (2014). COS-OGA: A novel oligosaccharidic elicitor that protects grapes and cucumbers against powdery mildew. Crop Prot. 65, 129–137. doi: 10.1016/j.cropro.2014.07.015
Vandana, U. K., Rajkumari, J., Singha, L. P., Satish, L., Alavilli, H., Sudheer, P. D. V. N., et al. (2021). The endophytic microbiome as a hotspot of synergistic interactions, with prospects of plant growth promotion. Biology 10, 101. doi: 10.3390/biology10020101h
Van Geel, M., Verbruggen, E., De Beenhouwer, M., van Rennes, G., Lievens, B., Honnay, O. (2017). High soil phosphorus levels overrule the potential benefits of organic farming on arbuscular mycorrhizal diversity in northern vineyards. Agr. Ecosyst. Environ. 248, 144–152. doi: 10.1016/j.agee.2017.07.017
Velásquez, A., Valenzuela, M., Carvajal, M., Fiaschi, G., Avio, L., Giovannetti, M., et al. (2020b). The arbuscular mycorrhizal fungus Funneliformis mosseae induces changes and increases the concentration of volatile organic compounds in Vitis vinifera cv. sangiovese leaf tissue. Plant Physiol. Biochem. 155, 437–443. doi: 10.1016/j.plaphy.2020.06.048
Velásquez, A., Vega-Celedón, P., Fiaschi, G., Agnolucci, M., Avio, L., Giovannetti, M., et al. (2020a). Responses of Vitis vinifera cv. Cabernet Sauvignon roots to the arbuscular mycorrhizal fungus Funneliformis mosseae and the plant growth-promoting rhizobacterium Ensifer meliloti include changes in volatile organic compounds. Mycorrhiza 30 (1), 161–170. doi: 10.1007/s00572-020-00933-3
Veliksar, S., Lemanova, N., Bratco, D., Gladei, M. (2017). The influence of trace elements and PGPR on growth and photosynthetic activity of grape seedlings. Lucrări ştiinţifice seria horticultură 60 (1), 109–114.
Veliksar, S., Lemanova, N., Gladei, M., Tatiana, D. (2022a). The impact of trace elements applied with PGPB on the Vitis vinifera L seedlings resistance to the copper excess in soil. SSRG Int. J. Agric. Environ. Sci. (SSRG-IJAES) 6 (4), 43–47.
Verhagen, B. W. M., Trotel-Aziz, P., Couderchet, M., Höfte, M., Aziz, A. (2010). Pseudomonas spp.-induced systemic resistance to Botrytis cinerea is associated with induction and priming of defence responses in grapevine. J. Exp. Bot. 61 (1), 249–260. doi: 10.1093/jxb/erp295
Vorholt, J. (2012). Microbial life in the phyllosphere. Nat. Rev. Microbiol. 10, 828–840. doi: 10.1038/nrmicro2910
Walorczyk, S., Drożdżyński, D., Gnusowski, B. (2011). Multiresidue determination of 160 pesticides in wines employing mixed-mode dispersive-solid phase extraction and gas chromatography-tandem mass spectrometry. Talanta 85 (4), 1856–1870. doi: 10.1016/j.talanta.2011.07.029
Wang, X., Glawe, D. A., Kramer, E., Weller, D., Okubara, P. A. (2018). Biological control of Botrytis cinerea: Interactions with native vineyard yeasts from Washington state. Phytopathology 108 (6), 691–701. doi: 10.1094/PHYTO-09-17-0306-R
Wei, M., Liu, X., He, Y., Xu, X., Wu, Z., Yu, K., et al. (2020). Biochar inoculated with Pseudomonas putida improves grape (Vitis vinifera L.) fruit quality and alters bacterial diversity. Rhizosphere 16, 100261. doi: 10.1016/j.rhisph.2020.100261
Winter, S., Bauer, T., Strauss, P., Kratschmer, S., Paredes, D., Popescu, D., et al. (2018). Effects of vegetation management intensity on biodiversity and ecosystem services in vineyards: A meta-analysis. J. App. Ecol. 55 (5), 2484–2495. doi: 10.1111/1365-2664.13124
Woolf, D., Amonette, J. E., Street-Perrott, F. A., Lehmann, J., Joseph, S. (2010). Sustainable biochar to mitigate global climate change. Nat. Commun. 1 (5), 56. doi: 10.1038/ncomms1053
Yakhin, O. I., Lubyanov, A. A., Yakhin, I. A., Brown, P. H. (2017). Biostimulants in plant science: A global perspective. Front. Plant Sci. 7. doi: 10.3389/fpls.2016.02049
Zaller, J. G., Cantelmo, C., Dos Santos, G., Muther, S., Gruber, E., Pallua, P., et al. (2018). Herbicides in vineyards reduce grapevine root mycorrhization and alter soil microorganisms and the nutrient composition in grapevine roots, leaves, xylem sap and grape juice. Environ. Sci. Pollut. Res. Int. 25 (23), 23215–23226. doi: 10.1007/s11356-018-2422-3
Zandonadi, D. B., Canellas, L. P., Façanha, A. R. (2007). Indolacetic and humic acids induce lateral root development through a concerted plasmalemma and tonoplast H+ pumps activation. Planta 225, 1583–1595. doi: 10.1007/s00425-006-0454-452
Zandonadi, D. B., Santos, M. P., Busato, J. G., Peres, L. E. P., Façanha, A. R. (2013). Plant physiology as affected by humified organic matter. Theor. Exp. Plant Physiol. 25, 13–25. doi: 10.1590/s2197-002520130001000
Zhang, Y., Wang, X., Liu, B., Liu, Q., Zheng, H., You, X., et al. (2020). Comparative study of individual and co-application of biochar and wood vinegar on blueberry fruit yield and nutritional quality. Chemosphere 246, 125699. doi: 10.1016/j.chemosphere.2019.125699
Keywords: biological agents, biostimulants, grapevine, organic, vineyard, viticulture, wine
Citation: Jindo K, Goron TL, Pizarro-Tobías P, Sánchez-Monedero MÁ, Audette Y, Deolu-Ajayi AO, van der Werf A, Goitom Teklu M, Shenker M, Pombo Sudré C, Busato JG, Ochoa-Hueso R, Nocentini M, Rippen J, Aroca R, Mesa S, Delgado MJ and Tortosa G (2022) Application of biostimulant products and biological control agents in sustainable viticulture: A review. Front. Plant Sci. 13:932311. doi: 10.3389/fpls.2022.932311
Received: 29 April 2022; Accepted: 05 September 2022;
Published: 18 October 2022.
Edited by:
Youssef Rouphael, University of Naples Federico II, ItalyReviewed by:
Marco Nuti, Sant’Anna School of Advanced Studies, ItalyCopyright © 2022 Jindo, Goron, Pizarro-Tobías, Sánchez-Monedero, Audette, Deolu-Ajayi, van der Werf, Goitom Teklu, Shenker, Pombo Sudré, Busato, Ochoa-Hueso, Nocentini, Rippen, Aroca, Mesa, Delgado and Tortosa. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Germán Tortosa, Z2VybWFuLnRvcnRvc2FAZWV6LmNzaWMuZXM=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.