 Cindy Callens
Cindy Callens José Fernandez-Goméz1
José Fernandez-Goméz1 Matthew R. Tucker
Matthew R. Tucker Zoe A. Wilson
Zoe A. Wilson- 1School of Biosciences, University of Nottingham, Loughborough, United Kingdom
- 2Waite Research Institute, School of Agriculture, Food and Wine, University of Adelaide, Urrbrae, SA, Australia
- 3Joint International Research Laboratory of Metabolic and Developmental Sciences, State Key Laboratory of Hybrid Rice, School of Life Sciences and Biotechnology, Shanghai Jiao Tong University, Shanghai, China
The Poaceae, or grasses, include many agriculturally important cereal crops such as rice (Oryza sativa), maize (Zea mays), barley (Hordeum vulgare) and bread wheat (Triticum aestivum). Barley is a widely grown cereal crop used for stock feed, malting and brewing. Abiotic stresses, particularly global warming, are the major causes of crop yield losses by affecting fertility and seed set. However, effects of heat stress on reproductive structures and fertility in barley have not been extensively investigated. In this study we examined three commercial European spring barley varieties under high temperature conditions to investigate the effects on floret development. Using a combination of fertility assays, X-ray micro computed tomography, 3-dimensional modelling, cytology and immunolabelling, we observed that male reproductive organs are severely impacted by increased temperature, while the female reproductive organs are less susceptible. Importantly, the timing of stress relative to reproductive development had a significant impact on fertility in a cultivar-dependent manner, this was most significant at pollen mitosis stage with fertility ranged from 31.6-56.0% depending on cultivar. This work provides insight into how heat stress, when applied during male pollen mother cell meiosis and pollen mitosis, affects barley fertility and seed set, and also describes complementary invasive and non-invasive techniques to investigate floret development. This information will be used to identify and study barley cultivars that are less susceptible to heat stress at specific stages of floral development.
1. Introduction
The world is facing an exponentially increasing population with associated increased demands for food, and all major climate models predict a higher average temperature globally with larger temperature fluctuations and more frequent heat waves (Christidis et al., 2014; IPCC, 2014). Abiotic stresses are, individually or in combination, one of the major reasons for crop yield loss. These stresses cause morphological, physiological, biochemical and molecular changes that impair plant development (Bita and Gerats, 2013). Lesk et al. (2016) showed that extreme heat significantly reduced national cereal production globally by 9-10%. Of particular concern is that three of the five most important cereal crops worldwide (i.e. wheat, maize and barley) exhibit the strongest negative yield impacts due to the changing climate (David and Christopher, 2007). Jacott and Boden (2020) recently reviewed the consequences of high ambient temperatures on wheat and barley, summarizing negative impacts on inflorescence development, the transition to flowering, and fertility. The increased frequency of heat stress over the last 40 years has clearly led to a greater frequency of yield anomalies in wheat (Zampieri et al., 2017), which accounts for 21% of food production. This proportion could be challenged with an increase in temperature in regions where temperatures are currently optimal (Ortiz et al., 2008; Delphine et al., 2014). Experiments in rice also showed that heat stress has an adverse effect on important yield components especially fertile spikelets and 1000 grain weight, decreasing yield significantly (Aghamolki et al., 2016). Similarly in spring barley, an increase in ambient temperature from 20°C/16°C (day/night) to 28°C/24°C reduced floret number and grains per spike (Hemming et al., 2012; Ejaz and von Korff, 2017). This serves as a warning for future global food security.
Reproductive organs are significantly more vulnerable to high temperatures than other plant organs. Planting of crops is typically timed to minimise high temperature exposure during the later reproductive stages, however the fluctuations and extremes of temperature that are now occurring mean that there is an increasing probability that peaks of high temperature will overlap with the flowering period (Teixeira et al., 2013; Kupke et al., 2022). It is proposed that “stay green” traits that enable photosynthesis to be maintained for longer, particularly under environmental stress, may enable yield losses due to abiotic stress to be minimized (Barnabas et al., 2008). Seed set requires many developmental steps to be successfully completed; for example, pollen must first be produced, viable pollen grains must be released and the pollen tubes must grow correctly to ensure functional signalling mechanisms with the style and the ovule. Concurrently, the ovule must develop within the ovary to produce and nourish the embryo sac, producing a female gamete and a suitable environment for the downstream events of seed development (Wilkinson et al., 2018). Once fertilization is complete, the embryo and endosperm must then develop normally to ensure successful seed fill (Maestri et al., 2002; Barnabas et al., 2008; Wilson and Zhang, 2009). Nevertheless, these developmental processes show variable sensitivity to heat stress (Jacott and Boden, 2020).
Prior to fertilisation, Saini et al. (1983) reported that one third of ovaries in wheat that experienced heat stress during meiosis exhibited abnormal development. Pollen, by contrast, exhibits much higher levels of abortion after heat stress at the same stage. Indeed, male reproductive development has been shown to be one of the most heat-sensitive stages in cereals (Saini et al., 1983; Stone and Basra, 2001; Prasad et al., 2008). At the pre-meiotic stage, high temperatures caused development of short anthers possessing no pollen grains in barley, while heat stress during meiosis resulted in pollen grains that possessed exine, but showed little starch accumulation (Sakata et al., 2000). Draeger and Moore (2017) also showed that exposure of wheat to high temperatures affected the progression of Pollen Mother Cell (PMC) meiosis. Disruption of synapsis is one of the most commonly reported meiotic failures under high temperatures and this can lead to unpaired univalents that segregate randomly or are lost (Bomblies et al., 2015). Elevated temperatures have also been shown to compromise mitosis 1 and 2 in both wheat and barley pollen (Saini et al., 1984; Barnabas et al., 2008). This may be due to an inability to synthesize all required Heat Shock Proteins (HSPs) necessary to survive during heat stress conditions (Cooper et al., 1984; Mascarenhas and Crone, 1996; Barnabas et al., 2008), and is consistent with causal involvement of HSPs in thermotolerance in non-cereal species (Hong and Vierling, 2000). Genotypes that express HSPs are better able to withstand heat stress as they minimise heat-induced protein aggregation and thus during the recovery period, facilitate their refolding (Nguyen et al., 1994; Feder and Hofmann, 1999; Farooq et al., 2011). In many cases, expression of HSPs is developmentally regulated and they are thus present prior to heat stress (Maestri et al., 2002).
One challenge of studying the response of florets to heat stress in cereals is determining when and where defects appear, how this differs between cultivars with different phenology and architecture, and how this might be examined in a non-destructive manner. In this study we analysed the effects of heat stress during floret development in three European spring barley varieties under controlled environmental conditions. One of the varieties, RGT Planet, represents a variety of considerable promise for high yield and malt quality in diverse conditions, including dryland environments in Australia (SeedForce, 2017). Our overall aim was to generate baseline data that might be used to develop further screening strategies and targets for reproductive heat tolerance during floret development in barley. We found that after heat stress, anthers from all varieties showed abnormal development, whilst ovules were less severely affected. The three varieties tested showed differences in sterility, confirming their varying tolerance to heat stress. RGT Planet exhibited excellent yield under control conditions, but suffered significantly after heat stress. In contrast to Moonshine and RGT Planet, Optic was the most severely affected by heat stress at pollen mitosis during floret development.
2. Materials and methods
2.1. Plant materials, growth and heat stress conditions
Seeds from three different spring barley varieties (Hordeum vulgare; Optic (derived from the cross Chad × (Corniche × Force), RAGT Moonshine and RGT Planet) were provided by the Wilson lab (University of Nottingham) and were sown in John Innes Potting Compost No.3 in 13cm diameter pots for germination and early plant establishment. All three are elite lines that have been on the AHDB Recommended List, although RAGT Moonshine and Optic were removed from the AHDB Recommended List in 2016 (AHDB, 2016). Optic is a variety created by Syngenta, while RAGT Moonshine and RGT Planet are varieties created by RAGT Seeds. RGT Planet is a relatively new variety that has been shown to be one of the highest yielding spring varieties on the AHDB Recommended List that is fully approved for brewing use (AHDB, 2017). All three are two-row varieties and are primarily grown in the United Kingdom, although RGT Planet is also popular in Australia. Temperatures of 15-20°C are considered optimal for growth and development of spring barley varieties (Kruszka et al., 2014).
After sowing, pots were placed in a controlled environment growth chamber with a continuous temperature of ~17°C and 16/8 -hour light/dark photoperiod and 80% relative humidity. After one week, the plants were transferred to CSNG (General Container Nursery Stock compost, Levington Advance) compost in 13cm diameter pots. Heat stress was applied to two stages of reproductive development (Pollen Mother Cell meiosis and pollen mitosis, Supplementary Figure S1). Staging of reproductive development was conducted according to the non-destructive staging described in Gómez and Wilson (2012), to ensure that all samples were at the appropriate and equivalent developmental stage. Pollen Mother Cell Meiosis corresponded to spike stage 4, spike one-quarter within previous last sheath; pollen mitosis corresponded to spike stage 13, spike half out (Gómez and Wilson, 2012). Stressed plants were compared to plants grown in control conditions to ensure there was a matching of the corresponding developmental stages. Male as well as female reproductive development were simultaneously targeted because the onset of the PMCs in anthers coincides approximately with meiosis in the megaspore mother cell (Saini et al., 1983).
Twelve plants of each variety were used for each condition: 1) Pollen Mother Cell (PMC) meiosis heat stress, 2) Pollen mitosis heat stress and 3) control. The periods of heat stress were carried out in a controlled environment growth room with a day/night temperature of 30/25°C, 80% relative humidity and a 16/8 -hour light/dark photoperiod to mimic rapid onset heat stress conditions. The temperatures were chosen to mimic heat waves occurring more frequently in Europe. Tillers were selected at the beginning of meiosis or mitosis, and were tagged for analysis before being submitted to heat stress. All pots were monitored daily and watered to avoid concurrent drought stress. Plants undergoing the PMC meiosis heat treatment were submitted to heat stress for two days, while plants undergoing the Pollen mitosis heat treatment remained in the heat stress conditions for five days to ensure completion of pollen mitosis I and II, before being transferred back to control conditions. Control plants were maintained at a constant 17°C, 80% relative humidity with a 16/8 -hour light/dark photoperiod.
2.2. Pollen viability assay
Pollen viability in control and heat stressed plants was assessed by analysis of starch levels in pollen grains using 0.2% (w/v) potassium iodide and 1% (w/v) iodine to determine their ability to germinate and fertilize (Chang et al., 2014). Normal mature pollen grains that contain starch granules stain black, whilst immature pollen grains or deformed pollen grains appear orange, or red. Pollen viability images were taken with a Nikon Eclipse 50i microscope and a Nikon DS-Fi1 camera.
2.3. Thin sectioning and cell wall immunolabelling
Florets were harvested from the tillers tagged for heat stress during pollen meiosis immediately before anthesis, fixed in 4% (w/v) paraformaldehyde in phosphate-buffered saline (PBS) with 0.1% (v/v) Triton X-100 and 0.1% (v/v) Tween 20 and embedded in paraffin. Floret sections (8μm) were mounted on slides at 42°C, the paraffin was removed from the sections using 100% Histoclear (v/v) and tissue was rehydrated by using an ethanol series (100%, 90%, 70% and 30% (v/v)) and water. Slides were washed with 1xPBS and treated subsequently with glycine to inactivate residual aldehyde groups. They were then washed with Incubation buffer (1% (w/v) bovine serum albumin (BSA)) in 1xPBS (Burton et al., 2011). Primary antibody was added to the sections and incubated in a humidity chamber for 1h: BG1 murine monoclonal antibodies raised against barley (1,3;1,4)-β-d-glucan (diluted 1:50; Biosupplies Australia, Parkville, Vic., Australia) and LM19 monoclonal antibodies raised against homogalacturonan (Meikle et al., 1994; Verhertbruggen et al., 2009; Burton et al., 2011). After three washes with Incubation buffer the slides were dried and the secondary antibodies added. Goat anti-mouse Alexa Fluor® 488 IgG (H+L) (diluted 1:200, Invitrogen, Australia) was used for BG1 and Dylight 550 Goat anti-rat IgM (diluted 1:200, Invitrogen, Australia) was used for LM19. The slides were incubated for 2h and then washed with Incubation buffer. 0.1% (w/v) Calcofluor white was added and washed off before imaging with the Zeiss Axio Imager 2 as described in Aditya et al. (2015).
2.4. 3D X-ray computed tomography imaging
The Phoenix Nanotom S, a nanofocus Computed Tomography (CT) system was used to scan barley florets of the three varieties at the Hounsfield Facility in Nottingham as previously described (Tracy et al. (2017). Control and heat stressed florets (heat stress applied during PMC meiosis) were scanned and the scans were segmented and analysed using VG Studio Max 2.2.
2.5. Reproductive organ phenotyping
Five different, randomly chosen florets were harvested immediately before anthesis after the heat treatments and in control conditions from all varieties and dissected to observe the morphology of the anthers. Images were taken with a Zeiss Stemi SV6 microscope and an Axiocam ERc Rev. 2.0 camera.
2.6. Evaluation of sterility
All tillers from control and heat stressed plants were harvested and the numbers of seeds per spike counted (Table 1) to determine the levels of fertility. For every spike the total number of spikelets and the number of spikelets that developed into seeds were counted. The reduction of fertility due to heat exposure was expressed as the percentage of sterility per tiller, by dividing the number of seeds formed by the total number of spikelets and multiplying by 100. Tagged heads from the plants were harvested and the percentage of sterility was determined on a tiller-by-tiller basis for each heat treatment and control conditions using a Kruskal-Wallis (one-way ANOVA) test to compare between treatments and varieties.
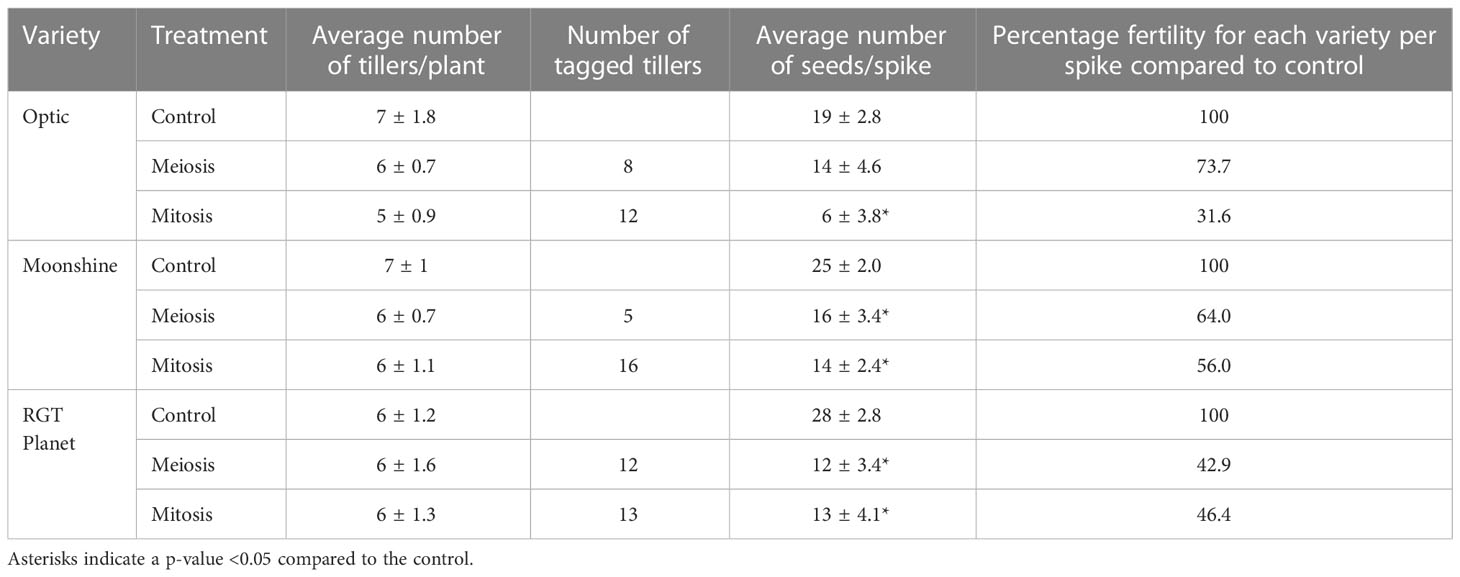
Table 1 Average number of tillers per plant and seeds per spike, and number of tagged tillers for the three different varieties in control conditions, after PMC meiosis heat treatment or after pollen mitosis heat treatment ± standard deviation.
2.7. Phylogenetic analysis of heat shock proteins in barley
To determine orthologs of AtHSP17.8 (AT1G07400.1) and AtHSP70 (AT3G12580.1) a nucleotide BLAST search was carried out in NCBI, rice database and IPK Barlex database. Orthologs were identified for rice (OsHSP17.8, LOC_Os03g16030.1 and OsHSP70, LOC_Os11g47760.1) and barley (HvHSP17.8, AK368988.1 and HvHSP70, HORVU5Hr1G021300.3 and HORVU6Hr1G081460) and verified by multiple alignment through phylogenetic tree analysis (Supplementary Figure S2). The alignment was constructed using the Geneious alignment method and the Blosum62 cost matrix in Geneious (version 8.0 by Biomatters, available from http://www.geneious.com). These proteins were chosen to give an indication of differences between varieties in the response of heat shock proteins to heat stress. Due to the high similarity between the proteins in the cytosol subgroup of the HSP70 family, we focussed only on the closest orthologues in barley.
2.8. Quantitative reverse transcription (qRT) PCR
RNA was extracted with the RNeasy® Plant Mini Kit (Qiagen, UK) from at least five florets and pooled together. cDNA was synthesised from total RNA using the SuperScript™ III reverse transcriptase (ThermoFisher Scientific, UK). qRT-PCR was performed with primers for HvHSP17.8 (HvHSP17.8 Fw 5’GAGGTGGAGGACGGCAACA3’ and HvHSP17.8 Rv 5’GATGGACTTGATCTCGGGTTT3’) and HvHSP70 (HvHSP70 Fw 5’ATCCTGAACGTGTCTGCTGA3’ and HvHSP70 Rv 5’TGGTGTTGCGCATGTTGTAA 3’) on three technical replicates and analysed using the LightCycler® 480 (Roche Life Science, UK). Expression values relative to the barley α-Tubulin gene (HvTubF 5’AGTGTCCTGTCCACCCACTC3’ and HvTubR 5’AGCATGAAGTGGATCCTTGG3’) were calculated.
3. Results
3.1. Anther morphology in barley varieties after heat stress
Anthers were dissected from at least three randomly selected florets for the different varieties (Optic, Moonshine and RGT Planet) that had been exposed to heat treatment at PMC meiosis and Pollen mitosis, and compared to those from the control conditions. Staging of anther development was conducted according to the non-destructive reproductive staging approach of Gómez and Wilson (2012), this ensured that heat stress was delivered at the appropriate developmental stage and that comparisons between samples were consistent based upon developmental stage. All of the heat-treated lines showed effects on anther morphology, with alterations in anther shape and a change in colour from yellow to white (Figure 1). In all three varieties, the effect on anther morphology was more severe after heat stress during PMC meiosis compared to Pollen mitosis, but both treatments showed impacts on anther shape and colour. In particular, Optic and RGT Planet showed a severe change in PMC meiosis heat stressed anthers (Figure 1). Anther morphology was investigated further using 3D images obtained from X-ray micro-CT scans of a representative floret from Optic and RGT Planet (Figure 2) and RAGT Moonshine (Supplementary Figure S3). Anther volume varied significantly; control non-stressed anthers were approximately 0.9mm3 (4.33mm length, 1.98mm width, 1.43mm depth) in Optic and 1.1 mm3 (4.49 mm length. 2.83 mm width, 1.37 mm depth) in RGT Planet. Whereas the PMC meiosis heat stressed anthers were smaller: 0.29 mm3 (3.91 mm length, 3.01 mm width, 1.82 mm depth) in Optic and 0.71 mm3 (3.1 mm length, 2.35 mm width, 1.01 mm depth) in RGT Planet (Figure 2). The septum in the heat-stressed anthers could be observed as broken in the micro-CT scans, while this was not the case for the control anthers (Figure 2A).
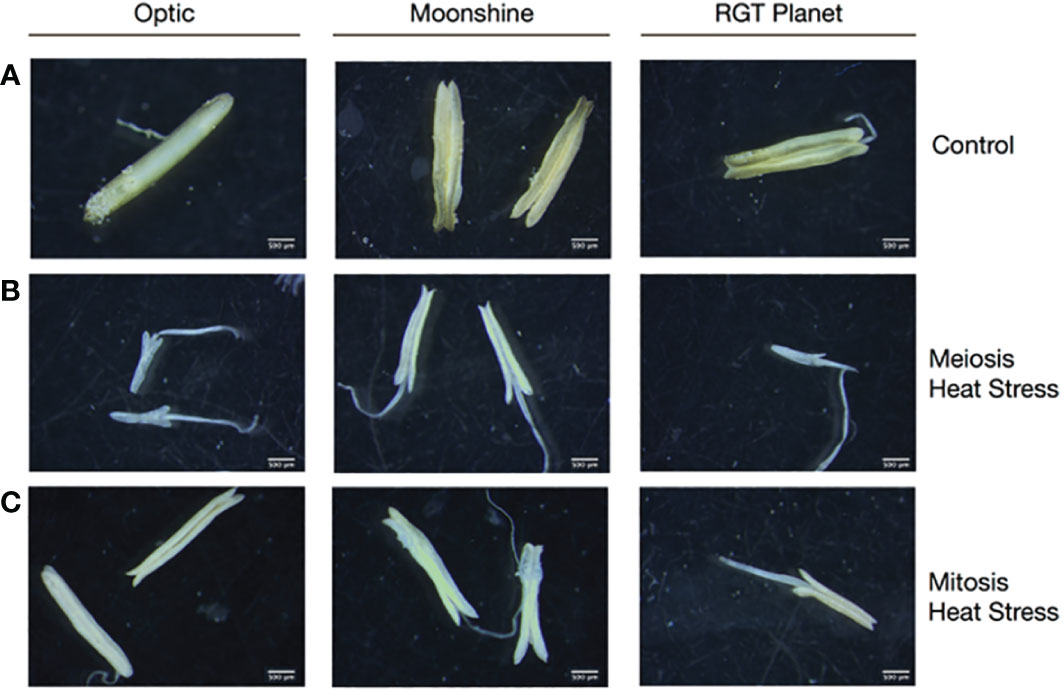
Figure 1 Anther development in barley cultivars Optic, Moonshine and RGT Planet. Representative anthers dissected from a minimum of three florets from Control (A) and heat stressed florets, (B) Heat Stress during Pollen Mother Cell meiosis and (C) Pollen Mitosis show differences in anther development under the different conditions.
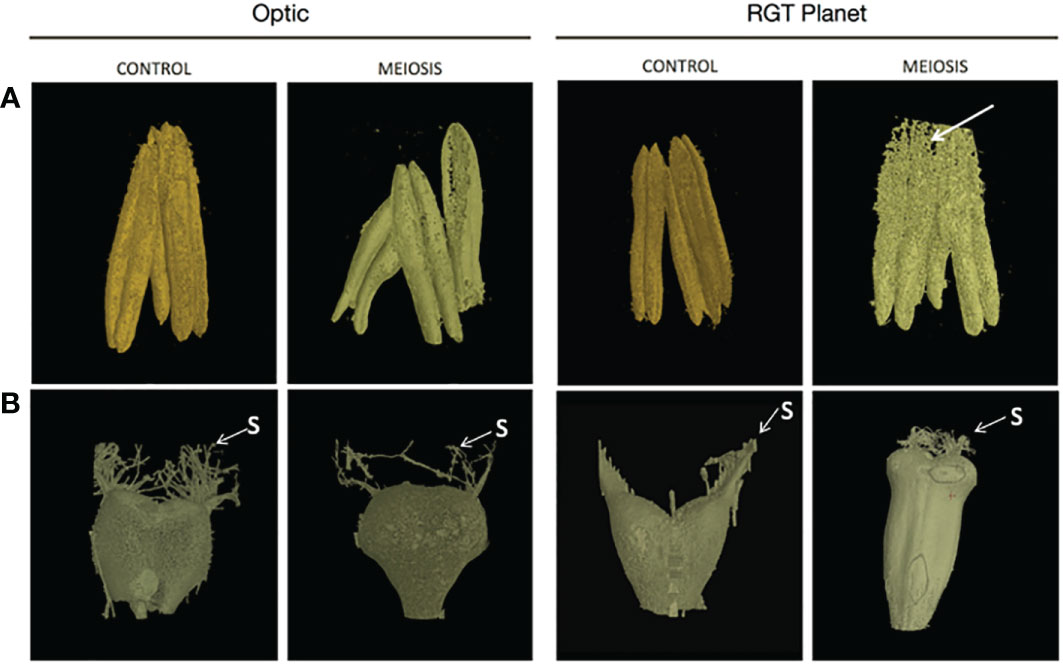
Figure 2 X-ray CT images of (A) anther and (B) carpel development from a representative floret from control and heat stress conditions for barley cultivars Optic and Planet. X-ray images of the anthers and carpels obtained by X-ray CT were compared for control environment and heat stress environment (heat stress during meiosis of pollen development) immediately before anthesis. Arrow indicates the broken septum in RGT Planet heat stressed anther.
Immunolabelling was subsequently used to observe sections of the anthers and highlight cell wall components such as 1,3;1,4-β-glucan, de-esterified homogalacturonan and cellulose to determine if the cell wall composition was impacted as a consequence of the heat stress (Figure 3; Supplementary Figures S4-6). Calcofluor white stained β-glycan polysaccharides in the outer layers of the anther, the filament vascular tissue and the pollen grains (Figure 3A). Some autofluorescence was detected in the pollen grains, particularly in the 1,3;1,4-β-glucan channel (Figure 3B), but positive controls confirmed 1,3;1,4-β-glucan and HG labelling (Figure 3C). The results clearly indicated that the meiosis heat-stressed anthers of all the lines were significantly impacted by the stress and labelling of HG in the anther walls appeared to be more intense compared to control anthers (Figures 3E, G, I). In Optic (Figures 3D, E), pollen was visible in the control anthers, but not in the PMC meiosis heat-stressed anthers. The epidermal cells of the anther wall were intact, but the endothecium had degraded. Both tissue layers were intact and visible in the Optic control anthers (Supplementary Figure S4). In Moonshine (Figures 3F, G), no pollen was visible in the PMC meiosis heat-stressed anthers despite it being observed in the micro-CT scanned images (Supplementary Figure S3). The epidermis was also intact, but the endothecium had degraded in the heat stressed anthers (Supplementary Figure S5). Similar to the other two varieties, while pollen grains were visible in the control anthers, none were visible in the heat stressed anthers in RGT Planet (Figures 3H, I). The anther wall in the heat stressed anthers seemed to have collapsed, which agrees with the observations from the micro-CT scan (Figure 2; Supplementary Figure S5).
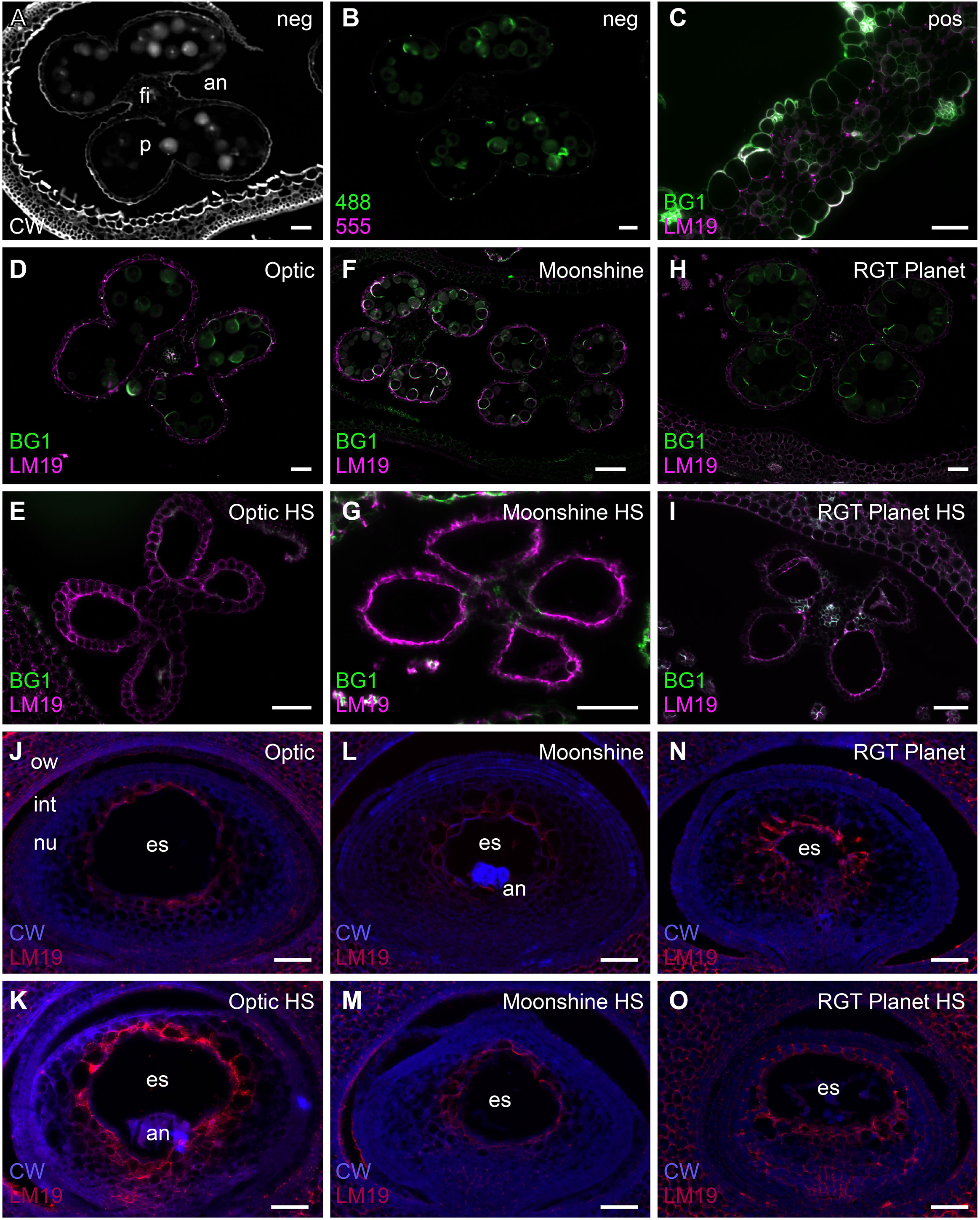
Figure 3 Immunolabelling of cell wall components in anthers and ovules in Optic, Moonshine and Planet. (A) Calcofluor white staining (CW, white colour) of a control anther (an). The filament (fi) and pollen (p) are indicated. (B) The same anther shown in A, showing autofluorescence (green) in the Alexafluor-488 channel. (C) A barley leaf sample showing positive labelling of (1,3;1,4)-β-glucan (BG1, green) and de-esterified pectin (LM19, magenta). (D–O) Comparison of control and heat stressed anthers and ovules in barley florets of the three varieties. Merged images of the antibody labelling patterns are shown. (D) Optic control anther. (E) Optic heat-stressed anther. (F) Moonshine control anther. (G) Moonshine heat-stressed anther. (H) Planet control anther. (I) Planet heat-stressed anther. (J) Optic control ovule. The ovary wall (ow), integuments (int), nucellus (nu) and embryo sac (es) are indicated. (K) Optic heat-stressed ovule. In this ovule the antipodals (an) are also evident. (L) Moonshine control ovule. (M) Moonshine heat-stressed ovule. (N) Planet control ovule. (O) Planet heat-stressed ovule. Bar = 50µm in all images. Images are representatives from at least 3 different florets per variety/heat stress treatment.
3.2. Carpel morphology in barley varieties after heat stress
The 3D images of the carpels from Optic showed no significant changes in morphology between the PMC meiosis heat-stressed and control florets, but there was a difference in size (Figure 2; Supplementary Figure S4). The control carpel measured 1.31mm3 (2.71mm length, 2.35mm width, 1.45mm depth), which is smaller than the heat stressed carpel, which measured 2.58mm3 (2.86 length, 2.77mm width, 1.34 depth). The X-ray imaging also showed a significantly enlarged carpel in the RGT Planet heat-stressed floret with a volume of 4.46mm3 (5.37mm length, 2.51mm width, 1.57mm depth) compared to the control which had a volume of 1.37mm3 (2.42mm length, 2.22mm width, 1.13mm depth) (Figure 2).
The immunolabelling of ovule sections in Optic only showed slight differences between control and heat stressed florets (Figures 3J, K): the embryo sac was present in all cases, but there seemed to be less cell layers between the embryo sac and the integument, and nucellus cells appeared larger. This could be due to regional nucellar degeneration and/or faster development of heat-stressed ovules. Similarly, embryo sac morphology in the heat-stressed ovules of Moonshine did not show any significant difference compared to control ovules (Figures 3L, M). The ovules in the heat-stressed florets of RGT Planet seemed to be further developed than the controls (Figures 3N, O), but no obvious irregularities in the morphology of the embryo sacs could be identified.
3.3. Male fertility after heat stress
PMC meiosis and pollen mitosis stages are known to be vulnerable to heat stress during pollen development, therefore pollen viability was assessed using potassium iodide/iodine solution in anthers from all three varieties with and without heat treatment. Using this method, viable pollen stains black due to starch deposition whilst non-viable pollen stains orange-red. Despite the lack of pollen identified in thin sectioning, pollen was still present in anthers from some florets after heat stress at PMC meiosis and pollen mitosis, but there was a lack of starch staining indicating a high level of sterility (Supplementary Figure S1B). Pollen could be observed in the Optic and RGT Planet anthers, after both PMC meiosis and pollen mitosis heat stress, but was far less abundant than in the control plants, and poor staining suggested that little if any was viable. Moonshine was even more strongly affected with no pollen present after the PMC meiosis heat stress. After heat stress during pollen mitosis, viable pollen was observed in Moonshine but at a lower amount than in the control anthers. The impact of heat stress varied depending on developmental stage and cultivar, with increased sterility seen for heat stress during the pollen mitosis stage; with 26-57% sterility observed for heat stress at the PMC meiosis stage, whilst 44-68% sterility was seen when heat stress occurred at pollen mitosis stage (Table 1).
3.4. Seed set after heat stress
The number of tillers and seeds per spike was assessed collectively for the three varieties after both treatments and in control conditions (Table 1). In Optic, there was no significant difference between the average number of seeds per spike after heat stress during PMC meiosis (~14 seeds) and the control (~19 seeds). However, there was a significant reduction of average seed number per spike after pollen mitosis heat stress compared to the control, from 19 to 6 seeds per spike. In Moonshine there was a significant decrease in seed set after both PMC meiosis and pollen mitosis heat treatments, from 25 in the control conditions to 16 and 14 respectively, after stress. The same could be observed in RGT Planet where a significant decrease was identified after both heat treatments (Table 1). In control conditions, there was an average of 28 seeds per spike, while after PMC meiosis heat stress this was reduced to 12 and after pollen mitosis heat stress to 13. These results contrasted with the lack of pollen and severe defects in pollen viability described above. However, it likely reflects variation in the response of different florets on each spike (depending on position) and different tillers depending on their developmental stage
To partially address this variability, all tagged heads from the plants were harvested and the percentage of sterility was determined on a tiller-by-tiller basis for each heat treatment and in control conditions for all varieties. A Kruskal-Wallis test was used to compare between treatments and between varieties (Figures 4, 5). Even though some spikes showed significant levels of sterility in Optic, consistent with the overall analysis, there was no significant difference in the percentage of sterility between PMC meiosis heat treatment and control conditions. There was however a difference between pollen mitosis heat treatment and control conditions, showing a significant increase in sterility (Figure 4A). In Moonshine there was no significant difference between both heat treatments and the control conditions, despite the majority of PMC meiosis heat treated tillers exhibiting 100% sterility. In this case, overall significance was likely confounded by a small sample size and a single tiller that showed relatively normal fertility (Figure 4B). In contrast, Moonshine did appear to be particularly tolerant to heat treatment at the pollen mitosis stage. In RGT Planet the sterility was significantly different between both heat treatments and the control (Figure 4C). Comparison between varieties showed that there was no significant difference after PMC meiosis heat stress (Figure 4B). After pollen mitosis heat treatment there was no significant difference between Moonshine and RGT Planet, but there was a significant difference between Moonshine or RGT Planet and Optic, the latter having increased sterility compared to the other two varieties. In control conditions Moonshine and Optic showed similar yields, while RGT Planet had a significantly higher yield.
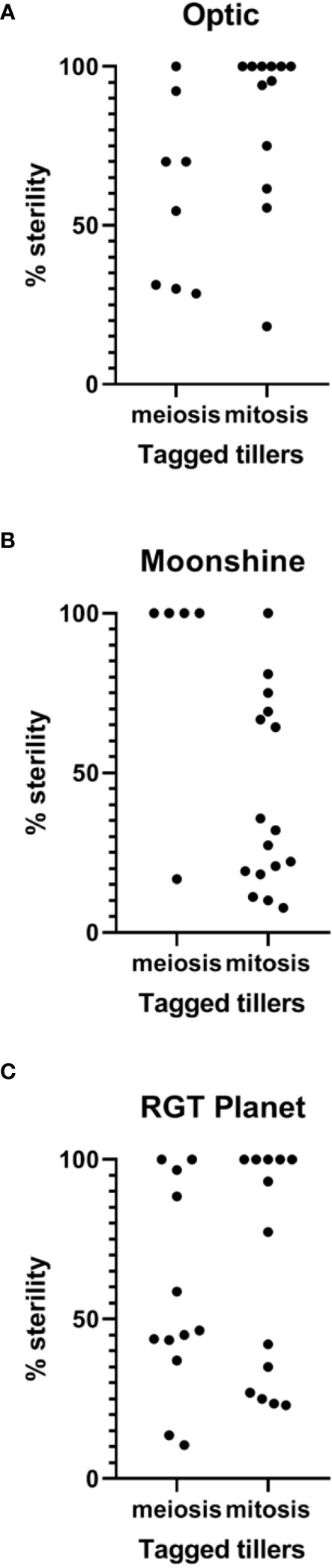
Figure 4 Sterility in heat stressed and control conditions in three European barley varieties. Percentage of sterility was calculated for all tillers for all three varieties after PMC meiosis and pollen mitosis heat treatments for (A) Optic (B) Moonshine and (C) RGT Planet. Kruskal-Wallis test was used to determine significance of difference between varieties and between treatments. All analysis was based upon based upon at least three biological replicates.
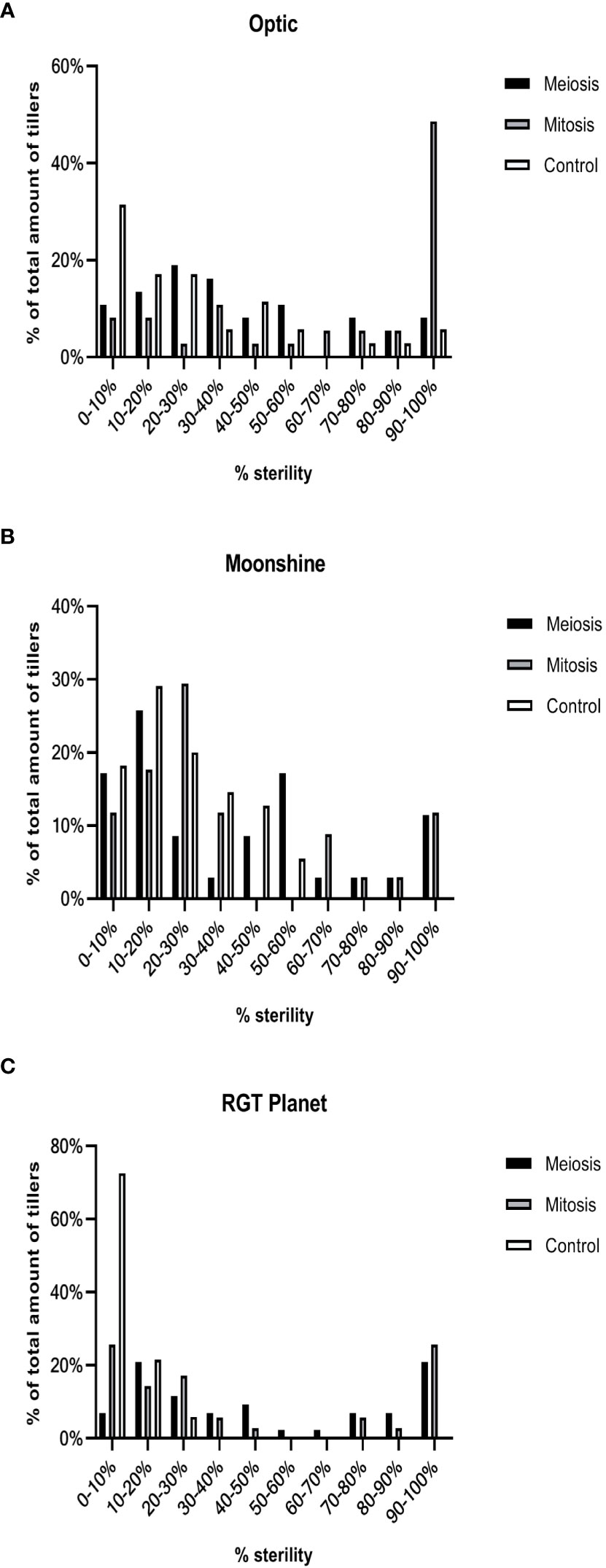
Figure 5 Frequency distribution of sterility in heat stressed and control conditions in three European barley varieties. The percentage sterility (florets that did not produce seed) was calculated for all three varieties in control conditions and after meiosis or mitosis I & II heat treatment for all pots (A–C).
3.5. Genes encoding heat stress proteins, HvHSP17.8 and HvHSP70, are induced after heat stress applied during mitosis
To determine the impact of heat stress on gene expression, two well characterized Arabidopsis heat stress proteins were investigated in barley; HvHSP17.8 is a small heat shock protein (sHSP) involved in protein stability, while HvHSP70 is a chaperone involved in protein folding (Hartl, 1996; Wang et al., 2004). The putative barley orthologues of the two Arabidopsis heat stress genes [HvHSP17.8 and HvHSP70, identified by BLAST and multiple alignment analysis (Supplementary Figure S2)] were analysed in florets as exemplars of gene expression changes after heat stress. The relative expression of HvHSP17.8 and HvHSP70 was examined on the first and last day (day 5) of pollen mitosis heat stress for all three varieties and control plants (Figure 6). HvHSP17.8 was significantly upregulated after one day of heat stress in Optic and RGT Planet, but not in Moonshine. Expression of HvHSP17.8 had returned to the minimal background level after five days of heat stress. HvHSP70 was upregulated in all three varieties after one day of heat stress, most significantly in RGT Planet. Although HvHSP70 expression decreased after five days of heat stress, it was still upregulated in all three cultivars compared to the controls.
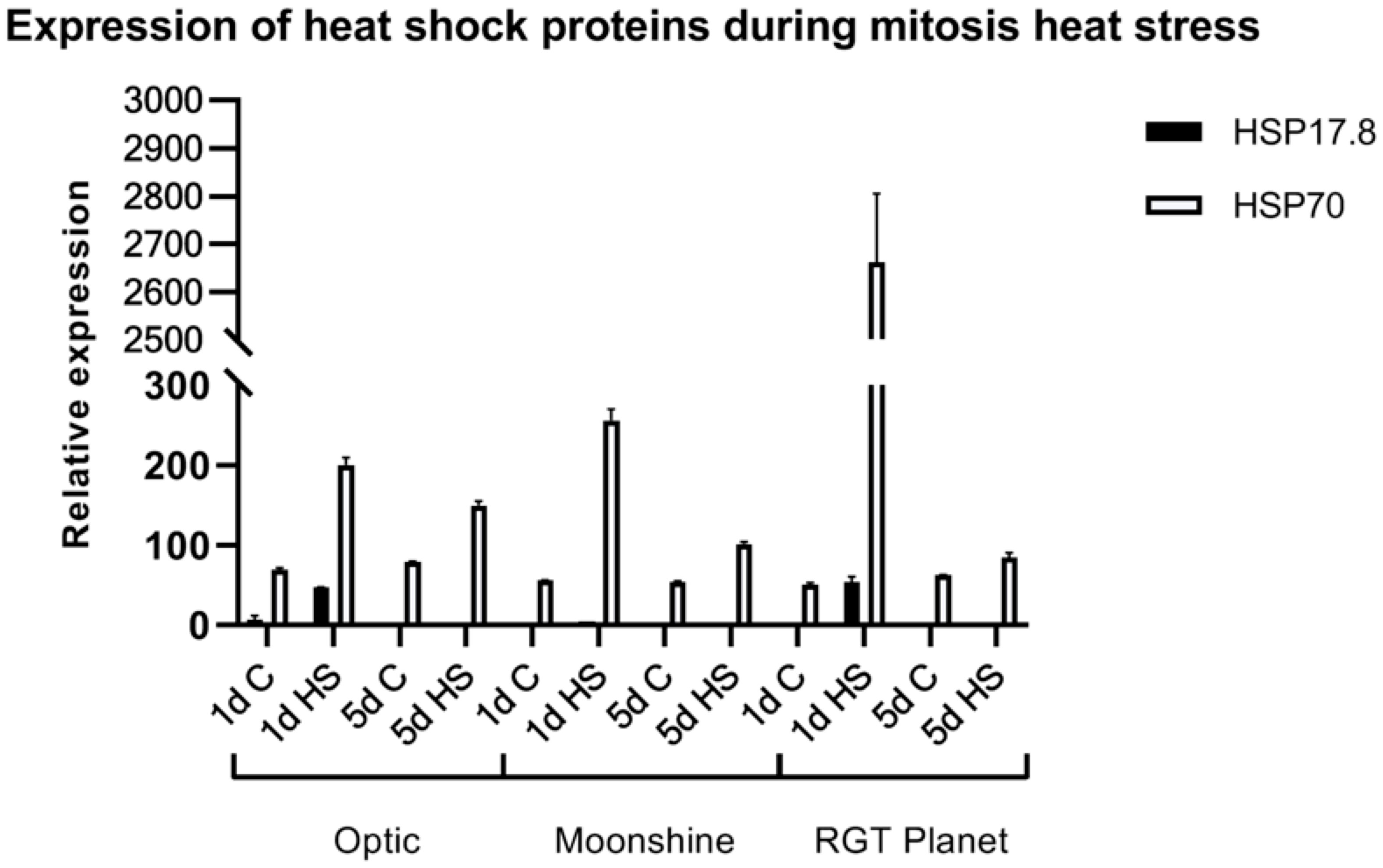
Figure 6 Expression of Heat Stress Proteins, HvHSP17.8 and HvHSP70, in florets of three European barley varieties, Optic, Moonshine and RGT Planet, during heat stress at the pollen mitosis I & II stages. C: control no heat stress; 1 and 5 days heat stress. Error bars indicate standard error, based upon at least three biological replicates.
4. Discussion
Three elite barley varieties were assessed for their response to heat stress during floret development. In all cases, plants that experienced heat stress at two key stages (PMC meiosis and pollen mitosis) showed severe morphological deficiencies in anther development, and these are likely to be a leading cause of sterility due to high temperature stress. A short period of heat stress during early floret development resulted in significantly smaller and deformed anthers. Although the anthers were harvested at the same time from heat stressed and control plants, the septa in the heat stressed anthers had already broken, suggesting a more rapid developmental progression and/or a change in the physical properties of the anther walls. It has been suggested that thick locule walls and well-developed cavities in the septa may be responsible for heat tolerance in rice (Matsui et al., 2001). The cavities are hypothesised to enable easy rupture of the septa in response to the swelling of the pollen grains, while the thick locule walls promote the swelling of the pollen grains by retaining water in the locules (Matsui et al., 2001). In the anthers examined here, the locule walls and cavities seemed less developed in the heat stressed plants, which might be another reason why the pollen grains did not develop correctly.
Saini et al. (1984) described two types of anther defects after heat stress in wheat. Type 1 typically had premature tapetal degeneration which resulted in periplasmodial invasion of the locule at meiosis and ultimately led to sterility. Degeneration of the outer layers of the anther wall was also observed. The second type was characterised by microspores that completed pollen grain mitosis I (PGM1), but a proportion of which became disoriented from the tapetum and developed no further. The breakage of the septa identified in heat-stressed barley might also be attributed to the fragility of the anther walls, as we observed some degeneration of anther wall layers in the immunolabelled images from RGT Planet.
In addition to defects in anther morphology, pollen development was severely affected by heat stress during both PMC meiosis and pollen mitosis, with decreased viability, pollen count, and accumulation of starch. However, as shown by the X-ray micro CT images of Moonshine florets after PMC meiosis heat stress, and by the presence of viable pollen in Optic after meiosis heat stress, pollen is still formed in some heat-stressed florets and, based on KI staining and seed counts, some may be viable. The absence of pollen detected in anthers during immunolabelling may be due to early anther rupture and loss of pollen during fixation and embedding. Alternatively, many florets may lack pollen entirely, and pollen is only released from some florets that avoid stress based on their position within the spike and their stage of development, which varies between the central and terminal regions. This highlights the importance of optimising non-destructive X-ray CT to assess anther development along the spike in vivo after heat stress.
Despite apparent contradictions between the immunolabelling experiments, KI staining assays, and CT data in regard to pollen grain number and viability after heat stress, seed set was used as a functional output of floret fertility. This varied between the treatments and cultivars. In Moonshine, overall seed set was significantly reduced across many tillers after heat treatment at PMC meiosis or pollen mitosis, but analysis of individual spikes suggested that Moonshine was more resilient against heat stress at the pollen mitosis stage. Optic, on the other hand, showed no significant response to heat stress at PMC meiosis, but was strongly sensitive at pollen mitosis. In contrast, RGT Planet was sensitive to heat stress at both PMC meiosis and mitosis stages of pollen development. These results potentially reveal different temporal sensitivities to heat stress in three elite barley cultivars.
The reduction in floret fertility and seed number may result from multiple defects in the pollen after heat stress. This includes reduced starch accumulation, which was observed in Optic pollen after heat stress at pollen mitosis. It has been suggested that stresses such as water stress can inhibit starch deposition in rice and wheat pollen, either by decreasing the availability of assimilates or by impairing the activities of enzymes involved in starch biosynthesis (Sheoran and Saini, 1996; Ji et al., 2010). Pressman et al. (2002) found that continuous high temperatures prevented the transient increase in starch concentration in tomato pollen grains which led to decreases in the concentrations of soluble sugars in the anther walls and the pollen. They concluded that this might contribute to decreased pollen viability in tomato after heat stress. This has been confirmed in other species, such as sorghum (Jain et al., 2007), and has been supported by evidence that barley grains from heat stressed plants accumulated less starch than grains from control plants due to reduced conversion of sucrose to starch (Wallwork et al., 1998). Heat stress has been reported to have a negative effect on the activities of enzymes involved in the sucrose-to-starch metabolism in cereals which might explain the reduction in starch content (Duke and Doehlert, 1996; Wilhelm et al., 1999; Hurkman et al., 2003).
The female reproductive organs did not show any significant differences in embryo sac phenotypes after heat stress. However, the carpels were bigger, possibly as a result of swelling of the unfertilized ovaries similar to that observed in wheat (Okada et al., 2018). Moreover, the carpels and ovules appeared to have developed faster than the controls. This suggests that in the three varieties under examination, the female reproductive organs are not as severely impacted by heat stress as anthers at the PMC meiotic and pollen mitotic stages of development, but advance more quickly through development. The broken septa in the anthers in the heat stressed plants also indicate that plants exposed to short pulse of heat stress around PMC meiosis might respond by speeding up reproductive organ development. A similar phenomenon was previously reported where heat stress hastened spike development and reduced spike number, thus impacting the number of grains per spike (Halse and Weir, 1974; Saini and Aspinall, 1982; Johnson and Kanemasu, 1983).
Several studies indicate that both pre- and post-fertilisation stages of ovary development are sensitive to stress, but this varies depending on the species and genotype (Sun et al., 2004; Zinn et al., 2010; Bac-Molenaar et al., 2015; Onyemaobi et al., 2017). In wheat, plants exposed to severe heat stress at the start of meiosis experienced disrupted nucellus and integument development, and complete ovule abortion at a frequency of 30% (Saini et al., 1983). In contrast, little is known about the effects of abiotic stress on cellular morphology and the regulatory network of the carpel and ovule in barley. The nucellar cells in all three barley cultivars examined here, particularly Optic, appeared to be disorganised and enlarged after heat stress relative to controls. However, in all cases, similar immunolabelling patterns for de-esterified homogalacturonan were observed around what appeared to be an intact embryo sac. Barley only has one ovule per floret and therefore needs to safeguard its one chance of survival. The multi-layered nucellus might be one of the reasons the developmental program of the ovule is more robust than that of the anther (Wilkinson et al., 2018).
The expression of two heat shock protein genes, HvHSP17.8 and HvHSP70, were upregulated after heat stress. HvHSP17.8 is a small heat shock protein (sHSP) that assists in preventing aggregation and stabilizing proteins while HvHSP70 is a chaperone that assists in protein folding processes (Hartl, 1996; Wang et al., 2004). Both genes have been shown to be upregulated under stress conditions and to confer heat stress tolerance (Sung et al., 2001; Guo et al., 2009; Montero-Barrientos et al., 2010). HvHSP70 was upregulated in all three varieties after one day and also after five days of heat stress (Figure 6). HvHSP70 expression showed a peak after one day of pollen mitosis heat stress in RGT Planet, with much higher expression than in the other two varieties. Optic showed the least amount of upregulation of HvHSP70, which correlated with the elevated level of sterility in this cultivar after pollen mitosis heat stress. In contrast, HvHSP17.8 was upregulated in Optic and RGT Planet after one day of heat stress at the pollen mitosis stage, but levels were indistinguishable from controls after five days of heat stress. This might indicate that HvHSP17.8 is involved in the early response to heat stress. Curiously, a significant increase in HvHSP17.8 was not observed in Moonshine despite this cultivar appearing to be less susceptible to heat-stress at the pollen mitosis stage. One possibility is that the timing of HvHSP17.8 induction differs in Moonshine; alternatively, other HSPs may be induced and contribute to the higher levels of stress tolerance observed in this cultivar.
5. Conclusions
A short period of heat stress during the reproductive phase in barley is detrimental for the development of the male reproductive organs, but less so for the female reproductive organs. Results indicate that after heat stress, floret development was generally hastened, which might reflect an overall stress response from the plant to ensure seed set, albeit with a smaller amount of seeds. Prolonged heat stress at a later stage in development (pollen mitosis) showed more severe effects in the male reproductive organs in Optic than in the other two varieties, with no viable pollen and significantly more sterility. The female reproductive organs of the three cultivars showed no severe effects after heat stress, at least in terms of embryo sac expansion. Overall, RGT Planet performed worst compared to the control conditions, whilst Moonshine performed best. However, there were less florets per spike in the heat stressed plants across all three varieties compared to control plants. Further investigation will be required to identify the traits that contribute to Moonshine being more heat tolerant than the other two varieties. The data provided here provides a basis for further studies into differential heat stress responses during key stages of barley reproductive development. Moreover, the use of X-ray imaging is a promising new method to visualise the morphology of floret organs without dissection and manipulation with chemicals, giving a more representative image and accurate volume measurements. Further work is needed to determine how the impact of the heat stress observed here equates to that under field conditions and the resultant potential consequences for yield loss. Nevertheless, significant yield reductions have been observed (Jacott and Boden, 2020), so it seems likely that equivalent impacts will be seen. The lines used here our elite cultivars that have not been particularly developed for heat stress resilience. However, the impact of the heat stress observed here can provide a basis for trait targeting for increasing future crop resilience.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Author contributions
CC, ZW, and DZ conceived the study. CC carried out the majority of experiments and data analysis. JFG assisted with plant maintenance and spike analysis. MT assisted with immunolabelling. All authors contributed to the article and approved the submitted version.
Funding
CC was supported by a Dual PhD scholarship from The University of Nottingham and University of Adelaide. This work was supported by the Australian Research Council (DZ, DP170103352; MT, FT140100780), an Australia-China Science and Research Fund Joint Research Centre grant ACSRF48187, the National Key Technologies Research and Development Program of China, Ministry of Science and Technology (grant no. 2016YFD 0100804; 2016YFE0101000), and the National Natural Science Foundation of China (NSFC) (31230051).
Acknowledgments
The authors wish to thank members of the Wilson, Zhang and Tucker laboratories for fruitful discussions and suggestions regarding experiments. In particular we thank Dr Laura Wilkinson for assisting with immunolabelling experiments.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2022.918730/full#supplementary-material
Supplementary Figure 1 | Schematic of heat stress treatments during pollen development for barley varieties and pollen viability in barley cultivars Optic, Moonshine and RGT Planet. (A) Two pots with four plants each of the three varieties were submitted to either meiosis heat stress during meiosis I and II of pollen development, or mitosis heat stress during mitosis I and II of pollen development. (B) Pollen viability is shown from a randomly selected floret right before anthesis from heat stressed plants and control plants. Black pollen is viable, while yellow/brown pollen is not viable; low numbers of pollen reflect the lack of pollen in the mature anthers.
Supplementary Figure 2 | Phylogenetic analysis of rice, Arabidopsis and barley HSP17.8 (A) and HSP70 orthologs (B). The alignments were constructed using the Geneious alignment method and the Blosum62 cost matrix in Geneious. (C D) Phylogenetic tree of Arabidopsis BLAST analysis HSP17.8 (C) and HSP70 (D).
Supplementary Figure 3 | X-ray CT images of anther and carpel development in control and heat stress conditions for Moonshine. X-ray images of the anthers and carpels obtained by X-ray CT are compared for control environment and heat stress environment (heat stress during meiosis of pollen development) right before anthesis.
Supplementary Figure 4 | Immunolabelling of cell wall components in anthers and ovules in Optic. Comparison of control and heat stressed anthers and ovules in barley florets of the variety RGT Planet. Antibodies include BG1 (1,3;1,4-β-glucan, green) and LM19 (low methylesterified pectin, red). Calcofluor White counterstain was used to detect cellulose and mixed-linkage glucan (blue).
Supplementary Figure 5 | Immunolabelling of cell wall components in anthers and ovules in Moonshine. Comparison of control and heat stressed anthers and ovules in barley florets of the variety RGT Planet. Antibodies include BG1 (1,3;1,4-β-glucan, green) and LM19 (low methylesterified pectin, red). Calcofluor White counterstain was used to detect cellulose and mixed-linkage glucan (blue).
Supplementary Figure 6 | Immunolabelling of cell wall components in anthers and ovules in RGT Planet. Comparison of control and heat stressed anthers and ovules in barley florets of the variety RGT Planet. Antibodies include BG1 (1,3;1,4-β-glucan, green) and LM19 (low methylesterified pectin, red). Calcofluor White counterstain was used to detect cellulose and mixed-linkage glucan (blue).
References
Aditya, J., Lewis, J., Shirley, N., Tan, H.-T., Henderson, M., Fincher, G., et al (2015). The dynamics of cereal cyst nematode infection differ between susceptible and resistant barley cultivars and lead to changes in (1,3;1,4)-β-glucan levels and HvCslF gene transcript abundance. New Phytol. 207, 135–147. doi: 10.1111/nph.13349
Aghamolki, M. T. K., Yusop, M. K., Oad, F. C., Jaafar, H. Z., Khalatbari, A. M., Kharidah, S., et al. (2016). Impact of heat stress on growth and yield of rice (Oryza sativa l.) cultivars. J. Food Agric. Environ. 14, 111–116.
AHDB (2016). AHDB recommended lists 2016-2017. Available at: https://ahdb.org.uk/knowledge-library/recommended-lists-for-cereals-and-oilseeds-rl.
AHDB (2017). Spring barley harvest results 2017. Available at: https://ahdb.org.uk/knowledge-library/recommended-lists-for-cereals-and-oilseeds-rl.
Bac-Molenaar, J. A., Fradin, E. F., Becker, F. F. M., Rienstra, J. A., van der Schoot, J., Vreugdenhil, D., et al. (2015). Genome-wide association mapping of fertility reduction upon heat stress reveals developmental stage-specific QTLs in arabidopsis thaliana. Plant Cell 27, 1857–1874. doi: 10.1105/tpc.15.00248
Barnabas, B., Jager, K., Feher, A. (2008). The effect of drought and heat stress on reproductive processes in cereals. Plant Cell Environ. 31, 11–38.
Bita, C. E., Gerats, T. (2013). Plant tolerance to high temperature in a changing environment: scientific fundamentals and production of heat stress-tolerant crops. Front. Plant Sci. 4, 273. doi: 10.3389/fpls.2013.00273
Bomblies, K., Higgins, J. D., Yant, L. (2015). Meiosis evolves: adaptation to external and internal environments. New Phytol. 208, 306–323. doi: 10.1111/nph.13499
Burton, R. A., Collins, H. M., Kibble, N. A., Smith, J. A., Shirley, N. J., Jobling, S. A., et al. (2011). Over-expression of specific HvCslF cellulose synthase-like genes in transgenic barley increases the levels of cell wall (1,3;1,4)-beta-d-glucans and alters their fine structure. Plant Biotechnol. J. 9, 117–135. doi: 10.1111/j.1467-7652.2010.00532.x
Chang, F., Zhang, Z., Jin, Y., Ma, H. (2014). “Cell biological analyses of anther morphogenesis and pollen viability in arabidopsis and rice,” in Flower development: Methods and protocols. Eds. Riechmann, J. L., Wellmer, F. (New York, NY: Springer New York) 203–216.
Christidis, N., Jones, G. S., Stott, P. A. (2014). Dramatically increasing chance of extremely hot summers since the 2003 European heatwave. Nat. Climate Change 5, 46.
Cooper, P., Ho, T.-H. D., Hauptmann, R. M. (1984). Tissue specificity of the heat-shock response in maize. Plant Physiol. 75, 431. doi: 10.1104/pp.75.2.431
David, B. L., Christopher, B. F. (2007). Global scale climate–crop yield relationships and the impacts of recent warming. Environ. Res. Lett. 2, 014002. doi: 10.1088/1748-9326/2/1/014002
Delphine, D., Declan, C., Navin, R., Jeff, P., Rachel, W. (2014). Global crop yield response to extreme heat stress under multiple climate change futures. Environ. Res. Lett. 9, 034011. doi: 10.1088/1748-9326/9/3/034011
Draeger, T., Moore, G. (2017). Short periods of high temperature during meiosis prevent normal meiotic progression and reduce grain number in hexaploid wheat (Triticum aestivum l.). Theor. Appl. Genet. 130, 1785–1800. doi: 10.1007/s00122-017-2925-1
Duke, E. R., Doehlert, D. C. (1996). Effects of heat stress on enzyme activities and transcript levels in developing maize kernels grown in culture. Environ. Exp. Bot. 36, 199–208. doi: 10.1016/0098-8472(96)01004-0
Ejaz, M., von Korff, M. (2017). The genetic control of reproductive development under high ambient temperature. Plant Physiol. 173, 294–306. doi: 10.1104/pp.16.01275
Farooq, M., Bramley, H., Palta, J. A., Siddique, K. H. M. (2011). Heat stress in wheat during reproductive and grain-filling phases. Crit. Rev. Plant Sci. 30, 491–507. doi: 10.1080/07352689.2011.615687
Feder, M. E., Hofmann, G. E. (1999). HEAT-SHOCK PROTEINS, MOLECULAR CHAPERONES, AND THE STRESS RESPONSE: Evolutionary and ecological physiology. Annu. Rev. Physiol. 61, 243–282. doi: 10.1146/annurev.physiol.61.1.243
Gómez, J. F., Wilson, Z. A. (2012). Non-destructive staging of barley reproductive development for molecular analysis based upon external morphology. J. Exp. Bot. 63, 4085–4094. doi: 10.1093/jxb/ers092
Guo, P., Baum, M., Grando, S., Ceccarelli, S., Bai, G., Li, R., et al. (2009). Differentially expressed genes between drought-tolerant and drought-sensitive barley genotypes in response to drought stress during the reproductive stage. J. Exp. Bot. 60, 3531–3544. doi: 10.1093/jxb/erp194
Halse, N. J., Weir, R. N. (1974). Effects of temperature on spikelet number of wheat. Aust. J. Agric. Res. 25, 687–695. doi: 10.1071/AR9740687
Hartl, F. U. (1996). Molecular chaperones in cellular protein folding. Nature 381, 571. doi: 10.1038/381571a0
Hemming, M. N., Walford, S. A., Fieg, S., Dennis, E. S., Trevaskis, B. (2012). Identification of high-temperature-responsive genes in cereals. Plant Physiol. 158, 1439–1450. doi: 10.1104/pp.111.192013
Hong, S.-W., Vierling, E. (2000). Mutants of arabidopsis thaliana defective in the acquisition of tolerance to high temperature stress. Proc. Natl. Acad. Sci. United States America 97, 4392–4397.
Hurkman, W. J., McCue, K. F., Altenbach, S. B., Korn, A., Tanaka, C. K., Kothari, K. M., et al. (2003). Effect of temperature on expression of genes encoding enzymes for starch biosynthesis in developing wheat endosperm. Plant Sci. 164, 873–881. doi: 10.1016/S0168-9452(03)00076-1
IPCC (2014). “Climate change 2014: Mitigation of climate change. contribution of working group III to the fifth assessment report of the intergovernmental panel on climate change,”. Eds. Edenhofer, O., Pichs-Madruga, R., Sokona, Y., Farahani, E., Kadner, S., Seyboth, K., Adler, A., Baum, I., Brunner, S., Eickemeier, P., Kriemann, B., Savolainen, J., Schlömer, S., von Stechow, C., Zwickel, T., Minx, J. C. (Cambridge, United Kingdom and New York, NY, USA: Cambridge University Press).
Jacott, C. N., Boden, S. A. (2020). Feeling the heat: developmental and molecular responses of wheat and barley to high ambient temperatures. J. Exp. Bot. 71, 5740–5751. doi: 10.1093/jxb/eraa326
Jain, M., Prasad, P. V., Boote, K. J., Hartwell, A. L., Jr., Chourey, P. S. (2007). Effects of season-long high temperature growth conditions on sugar-to-starch metabolism in developing microspores of grain sorghum (Sorghum bicolor l. moench). Planta 227, 67–79. doi: 10.1007/s00425-007-0595-y
Ji, X., Shiran, B., Wan, J., Lewis, D. C., Jenkins, C. L. D., Condon, A. G., et al. (2010). Importance of pre-anthesis anther sink strength for maintenance of grain number during reproductive stage water stress in wheat. Plant Cell Environ. 33, 926–942. doi: 10.1111/j.1365-3040.2010.02130.x
Johnson, R. C., Kanemasu, E. T. (1983). Yield and development of winter wheat at elevated Temperatures1. Agron. J. 75, 561–565. doi: 10.2134/agronj1983.00021962007500030033x
Kruszka, K., Pacak, A., Swida-Barteczka, A., Nuc, P., Alaba, S., Wroblewska, Z., et al. (2014). Transcriptionally and post-transcriptionally regulated microRNAs in heat stress response in barley. J. Exp. Bot. 65, 6123–6135. doi: 10.1093/jxb/eru353
Kupke, B. M., Tucker, M. R., Able, J. A., Porker, K. D. (2022). Manipulation of barley development and flowering time by exogenous application of plant growth regulators. Front. Plant Sci. 12, 694424. doi: 10.3389/fpls.2021.694424
Lesk, C., Rowhani, P., Ramankutty, N. (2016). Influence of extreme weather disasters on global crop production. Nature 529, 84. doi: 10.1038/nature16467
Maestri, E., Klueva, N., Perrotta, C., Gulli, M., Nguyen, H. T., Marmiroli, N. (2002). Molecular genetics of heat tolerance and heat shock proteins in cereals. Plant Mol. Biol. 48, 667–681. doi: 10.1023/A:1014826730024
Mascarenhas, J. P., Crone, D. E. (1996). Pollen and the heat shock response. Sexual Plant Reprod. 9, 370–374. doi: 10.1007/BF02441959
Matsui, T., Omasa, K., Horie, T. (2001). The difference in sterility due to high temperatures during the flowering period among japonica-rice varieties. Plant Product. Sci. 4, 90–93. doi: 10.1626/pps.4.90
Meikle, P. J., Hoogenraad, N. J., Bonig, I., Clarke, A. E., Stone, B. A. (1994). A (1–>3,1–>4)-beta-glucan-specific monoclonal antibody and its use in the quantitation and immunocytochemical location of (1–>3,1–>4)-beta-glucans. Plant J. 5, 1–9. doi: 10.1046/j.1365-313X.1994.5010001.x
Montero-Barrientos, M., Hermosa, R., Cardoza, R. E., Gutiérrez, S., Nicolás, C., Monte, E. (2010). Transgenic expression of the trichoderma harzianum hsp70 gene increases arabidopsis resistance to heat and other abiotic stresses. J. Plant Physiol. 167, 659–665. doi: 10.1016/j.jplph.2009.11.012
Nguyen, H. T., Joshi, C. P., Klueva, N., Weng, J., Hendershot, K. L., Blum, A. (1994). The heat-shock response and expression of heat-shock proteins in wheat under diurnal heat stress and field conditions. Funct. Plant Biol. 21, 857–867. doi: 10.1071/PP9940857
Okada, T., Jayasinghe, JEARM, Nansamba, M., Baes, M., Warner, P., Kouidri, A., et al. (2018). Unfertilized ovary pushes wheat flower open for cross-pollination. J. Exp. Bot. 69, 399–412. doi: 10.1093/jxb/erx410
Onyemaobi, I., Liu, H., Siddique, K. H. M., Yan, G. (2017). Both Male and female malfunction contributes to yield reduction under water stress during meiosis in bread wheat. Front. Plant Sci. 7. doi: 10.3389/fpls.2016.02071
Ortiz, R., Sayre, K. D., Govaerts, B., Gupta, R., Subbarao, G. V., Ban, T., et al. (2008). Climate change: Can wheat beat the heat? Agricult. Ecosyst. Environ. 126, 46–58. doi: 10.1016/j.agee.2008.01.019
Prasad, P. V. V., Pisipati, S. R., Mutava, R. N., Tuinstra, M. R. (2008). Sensitivity of grain sorghum to high temperature stress during reproductive development. Crop Sci. 48, 1911–1917. doi: 10.2135/cropsci2008.01.0036
Pressman, E., Peet, M. M., Pharr, D. M. (2002). The effect of heat stress on tomato pollen characteristics is associated with changes in carbohydrate concentration in the developing anthers. Ann. Bot. 90, 631–636. doi: 10.1093/aob/mcf240
Saini, H. S., Aspinall, D. (1982). Abnormal sporogenesis in wheat (Triticum aestivum l.) induced by short periods of high temperature. Ann. Bot. 49, 835–846. doi: 10.1093/oxfordjournals.aob.a086310
Saini, H., Sedgley, M., Aspinall, D. (1983). Effect of heat stress during floral development on pollen tube growth and ovary anatomy in wheat (<I>Triticum aestivum</I> l.). Funct. Plant Biol. 10, 137–144. doi: 10.1071/PP9830137
Saini, H. S., Sedgley, M., Aspinall, D. (1984). Development anatomy in wheat of Male sterility induced by heat stress, water deficit or abscisic acid. Funct. Plant Biol. 11, 243–253. doi: 10.1071/PP9840243
Sakata, T., Takahashi, H., Nishiyama, I., Higashitani, A. (2000). Effects of high temperature on the development of pollen mother cells and microspores in barley hordeum vulgare l. J. Plant Res. 113. doi: 10.1007/PL00013947
Sheoran, I. S., Saini, H. S. (1996). Drought-induced male sterility in rice: Changes in carbohydrate levels and enzyme activities associated with the inhibition of starch accumulation in pollen. Sexual Plant Reprod. 9, 161–169. doi: 10.1007/BF02221396
Stone, P., Basra, A. (2001). “The effects of heat stress on cereal yield and quality,” in Crop responses and adaptations to temperature stress (Binghampton: NY Food Products Press), 179–187.
Sung, D. Y., Vierling, E., Guy, C. L. (2001). Comprehensive expression profile analysis of the arabidopsis Hsp70 gene family. Plant Physiol. 126, 789–800. doi: 10.1104/pp.126.2.789
Sun, K., Hunt, K., Hauser, B. A. (2004). Ovule abortion in arabidopsis triggered by stress. Plant Physiol. 135, 2358–2367. doi: 10.1104/pp.104.043091
Teixeira, E. I., Fischer, G., van Velthuizen, H., Walter, C., Ewert, F. (2013). Global hot-spots of heat stress on agricultural crops due to climate change. Agric. For. Meteorol. 170, 206–215. doi: 10.1016/j.agrformet.2011.09.002
Tracy, S. R., Gómez, J. F., Sturrock, C. J., Wilson, Z. A., Ferguson, A. C. (2017). Non-destructive determination of floral staging in cereals using X-ray micro computed tomography (µCT). Plant Methods 13, 9. doi: 10.1186/s13007-017-0162-x
Verhertbruggen, Y., Marcus, S. E., Haeger, A., Ordaz-Ortiz, J. J., Knox, J. P. (2009). An extended set of monoclonal antibodies to pectic homogalacturonan. Carbohydr. Res. 344, 1858–1862. doi: 10.1016/j.carres.2008.11.010
Wallwork, M. A. B., Logue, S. J., MacLeod, L. C., Jenner, C. F. (1998). Effect of high temperature during grain filling on starch synthesis in the developing barley grain. Funct. Plant Biol. 25, 173–181. doi: 10.1071/PP97084
Wang, W., Vinocur, B., Shoseyov, O., Altman, A. (2004). Role of plant heat-shock proteins and molecular chaperones in the abiotic stress response. Trends Plant Sci. 9, 244–252. doi: 10.1016/j.tplants.2004.03.006
Wilhelm, E. P., Mullen, R. E., Keeling, P. L., Singletary, G. W. (1999). Heat stress during grain filling in maize: Effects on kernel growth and metabolism. Crop Sci. 39, 1733–1741. doi: 10.2135/cropsci1999.3961733x
Wilkinson, L. G., Bird, D. C., Tucker, M. R. (2018). Exploring the role of the ovule in cereal grain development and reproductive stress tolerance. Annu. Plant Rev. online. doi: 10.1002/9781119312994.apr0609
Wilson, Z. A., Zhang, D.-B. (2009). From arabidopsis to rice: pathways in pollen development. J. Exp. Bot. 60, 1479–1492. doi: 10.1093/jxb/erp095
Zampieri, M., Ceglar, A., Dentener, F., Toreti, A. (2017). Wheat yield loss attributable to heat waves, drought and water excess at the global, national and subnational scales. Environ. Res. Lett. 12, 064008. doi: 10.1088/1748-9326/aa723b
Keywords: barley, heat stress, floret development, meiosis, mitosis, pollen, male sterility, breeding
Citation: Callens C, Fernandez-Goméz J, Tucker MR, Zhang D and Wilson ZA (2023) Heat stress responses vary during floret development in European spring barley cultivars. Front. Plant Sci. 13:918730. doi: 10.3389/fpls.2022.918730
Received: 12 April 2022; Accepted: 20 December 2022;
Published: 03 February 2023.
Edited by:
Marta Adelina Mendes, University of Milan, ItalyReviewed by:
Umar Masood Quraishi, Quaid-i-Azam University, PakistanCara Griffiths, Rothamsted Research, United Kingdom
Sivakumar Sukumaran, The University of Queensland, Australia
Copyright © 2023 Callens, Fernandez-Goméz, Tucker, Zhang and Wilson. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Zoe A. Wilson, em9lLndpbHNvbkBub3R0aW5naGFtLmFjLnVr