 Amadou Sidibé
Amadou Sidibé Marie Thérèse Charles
Marie Thérèse Charles Jean-François Lucier
Jean-François Lucier Yanqun Xu
Yanqun Xu Carole Beaulieu
Carole Beaulieu- 1Department of Biology, Université de Sherbrooke, Sherbrooke, QC, Canada
- 2Saint-Jean-sur-Richelieu Research and Development Centre, Agriculture and Agri-Food Canada, Saint-Jean-sur-Richelieu, QC, Canada
- 3College of Biosystems Engineering and Food Science, Zhejiang Key Laboratory for Agri-Food Processing, Zhejiang University, Hangzhou, China
Preharvest application of hormetic doses of ultraviolet-C (UV-C) generates beneficial effects in plants. In this study, within 1 week, four UV-C treatments of 0.4 kJ/m2 were applied to 3-week-old lettuce seedlings. The leaves were inoculated with a virulent strain of Xanthomonas campestris pv. vitians (Xcv) 48 h after the last UV-C application. The extent of the disease was tracked over time and a transcriptomic analysis was performed on lettuce leaf samples. Samples of lettuce leaves, from both control and treated groups, were taken at two different times corresponding to T2, 48 h after the last UV-C treatment and T3, 24 h after inoculation (i.e., 72 h after the last UV-C treatment). A significant decrease in disease severity between the UV-C treated lettuce and the control was observed on days 4, 8, and 14 after pathogen inoculation. Data from the transcriptomic study revealed, that in response to the effect of UV-C alone and/or UV-C + Xcv, a total of 3828 genes were differentially regulated with fold change (|log2-FC|) > 1.5 and false discovery rate (FDR) < 0.05. Among these, of the 2270 genes of known function 1556 were upregulated and 714 were downregulated. A total of 10 candidate genes were verified by qPCR and were generally consistent with the transcriptomic results. The differentially expressed genes observed in lettuce under the conditions of the present study were associated with 14 different biological processes in the plant. These genes are involved in a series of metabolic pathways associated with the ability of lettuce treated with hormetic doses of UV-C to resume normal growth and to defend themselves against potential stressors. The results indicate that the hormetic dose of UV-C applied preharvest on lettuce in this study, can be considered as an eustress that does not interfere with the ability of the treated plants to carry on a set of key physiological processes namely: homeostasis, growth and defense.
Introduction
Plants live in a complex and constantly changing environment, where they continuously interact with biotic and abiotic factors (Foyer et al., 2016). These factors can be the cause of severe economic losses in edible plants such as lettuce, one of the most widely consumed leafy vegetables in the world (Moraes et al., 2018). The susceptibility of lettuce to certain diseases such as bacterial leaf spot (BLS), caused by Xanthomonas campestris pv. vitians (Xcv) can result in 100% yield losses (Lu and Raid, 2013). The bacterium can remain viable for months in buried plant debris or in surface irrigation water (Fayette et al., 2018) and penetrates leaves through stomata or wounds, resulting in bacterial clusters in the substomatal chamber (Bull et al., 2007). Typical symptoms of BLS begin as dark, oily looking spots and then cluster into large necrotic patches in warm, moist conditions (Bull et al., 2007) making the lettuce heads unsaleable. Confine Extra (mono and di-potassium salts of phosphorous acid) is the only chemical pesticide registered for the control of BLS in 2017 (Health Canada, 2019). To obtain the expected biological response, pesticides are used excessively and repeatedly, resulting in enormous costs and posing a serious threat to the environment and human health (Konatu and Jardim, 2018). Therefore, new and effective biological approaches to improve the control of BLS are urgently needed.
Recent scientific literature indicates increased interest in the use of biotic or abiotic agents as eustressors to improve plants’ functional quality and protect them against harmful levels of stressors. A biotic or abiotic eustressor is defined as a factor of biological, chemical or physical origin that can impact the physiological status of the plants in a beneficial way (Vázquez-Hernández et al., 2019). Very little is known about the cellular and molecular responses of plants to combined stimulation by a biotic disease-causing agent and an abiotic eustressor.
In response to biotic stresses, plants have developed a series of molecular pathways to adapt using proteins involved in cellular mechanisms as key players in the maintenance of cell homeostasis. Through regulation of physiological parameters, proteins participate directly in the response to biotic stresses giving rise to new plant phenotypes with peculiar characteristics (Feussner and Polle, 2015). The sequential effect of a certain combination of abiotic and biotic stresses may have a positive or negative impact on the plant’s response in terms of pathogen resistance and tolerance to abiotic stresses (Zhang and Sonnewald, 2017), depending on the level applied. For example, the effect of drought stress on tomato plant growth has led to increased resistance to Botrytis cinerea and Oidium neolycopersici (Achuo et al., 2006).
Research has shown that following applications of abiotic stresses such as heat stress, plants activate several molecular cascades involving kinases that regulate genes, transcription factors (TFs), microRNAs, reactive oxygen species (ROS) and Hsps (Muthusamy et al., 2017). Abiotic stresses result in the activation of numerous ion channels, signaling pathways of different hormonal cascades (ethylene, salicylic acid, abscisic acid, jasmonic acid) to overcome the adverse effects caused by the stress situation (Murcia et al., 2017). A recent study showed that proteometabolomic and physiological changes in algae due to ultraviolet-C (UV-C) doses of radiation are characterized by an increase in redox homeostasis, ROS production, protein damage and repair, avoidance elements, photosynthetic electron flux, carbon fixation, and C/N metabolism (Colina et al., 2020).
A complete understanding of metabolic and signaling pathways and their interactions during stress response is critical to enabling the development of stress-tolerant plants. The use of data from transcriptomics studies can provide comprehensive information on the interactions that occur in response to a specific stress (Bagati et al., 2018) such as UV-C. While in strawberries, studies have shown that treatment with UV-C has a significant impact on gene expression and leads to the overexpression of a set of genes required for effective defense in plant-pathogen interaction (Xu et al., 2019), the impact of UV-C on gene expression in lettuce is unknown.
Numerous studies have shown the beneficial effects of postharvest UV-C hormesis in several plant species (for a review, see Duarte-Sierra et al., 2019). When applied at the preharvest stage, the beneficial effects of UV-C hormesis have been shown in strawberry (Janisiewicz et al., 2016a,b; Xie et al., 2016; Xu et al., 2019), tomato (Valencia et al., 2017), and lettuce (Vàsquez et al., 2017; Sidibé et al., 2021). Among the beneficial effects of UV-C on lettuce, Sidibé et al. (2021) showed an increase in dry matter and resistance against Xcv with no negative impact on yield. The work of Sidibé et al. (2021) suggests that the lettuce plant is able to integrate the negative effects of UV-C-induced oxidative stress signals, return to a state of homeostasis and pursue normal growth while mounting an effective defense fight against BLS. Based on these premises, using transcriptomics in the present study, we propose a hypothetical model describing the regulatory mechanisms that may underlie the beneficial effects induced by preharvest hormetic doses of UV-C radiation on lettuce.
Materials and Methods
Plant Material and Inoculation
The lettuce cultivar ‘Parris Island Cos’, which is known to be susceptible to the pathogen Xanthomonas campestris pv. vitians-B07-007 (Xcv) (Nicolas et al., 2018), was used in this study. Seeding was carried out in a 98-well tray in a germinator for 2 weeks, and seedlings were then transferred to growth chambers as described by Sidibé et al. (2021). The plants were watered daily, alternating between water and a nutrient solution containing 200 ppm nitrogen, 71 ppm potassium and 200 ppm phosphorus. Inoculation was done with a virulent Xcv strain B07-007 isolated from lettuce fields in Montérégie (QC, Canada).
The inoculation conditions were described by Nicolas et al. (2018). Briefly, Xcv B07-007 was recovered in sterile distilled water after 48 h culture on YDC agar medium at a final concentration of 108 colony forming units (CFU) per mL and then sprayed up to runoff on the adaxial and abaxial surfaces of the leaves of 18 lettuce plants, 9 control, and 9 UV-C treated (Supplementary Figure 1) using a manual sprayer.
Ultraviolet-C Treatments
Ultraviolet-C treatments began 3 weeks after sowing, corresponding to the transplanting stage of lettuce in the field as described by Sidibé et al. (2021). The growth chambers (Conviron, PGV40, MB, Canada) were modified with a set of three UV-C lamps (254 nm, 160 W, Clean Light Inc., Vineland Station, ON, Canada). The lamps were placed at 80 cm over the tops of the plants. During treatment, the intensity of the lamps was 1.1 mW/cm2 measured with a radiometer (ML1400A, Miltec UV, Stevensville, MD, United States) equipped with a SEL240 # 6090 sensor (ML1400A, Miltec UV). The time required to administer the treatment dose of 0.4 kJ/m2 was 1 min. Four UV-C doses of 0.4 kJ/m2 (Supplementary Figure 1) at intervals of 48 h between the first three doses and 72 h for the last dose was applied (Nicolas et al., 2020). In total, lettuce plants in the treated group received 1.6 kJ/m2 over a 1-week period. Forty-eight hours after the last UV-C treatment, lettuce was inoculated with Xcv B07-007 as described above and in Supplementary Figure 1.
Evaluation of the Effect of Ultraviolet-C Treatments on Bacterial Leaf Spot Symptom Expression
The method of Nicolas et al. (2018) was used to assess BLS symptom development. The severity of the disease is the proportion of the foliar surface with symptoms of disease (Nicolas et al., 2018). The area under the disease progress curve (AUDPC) was calculated using equation (1) for trapezoidal integration of disease incidence as described by Shaner and Finney (1977).
where Y is the disease severity (per unit) at the ith observation, Xi is the time (days) at the ith observation, and n is the total number of observations.
This experiment was repeated three times.
RNA Extraction, Transcriptomic and qPCR Analysis
A transcriptomic study was carried on lettuces in three independent completely randomized experiments (E1, E2, E3), each designed with three replicates of three experimental units (Supplementary Table 1 and Supplementary Figure 1). Fully extended lettuce leaves without the main vein from each replicate were sampled at two different times, T2 and T3 (Supplementary Figure 1). For each experiment, a total of 54 plants were randomly selected after transplantation and acclimatization. At this time, the plants were separated into two groups: 27 plants received the UV-C treatment and 27 plants served as control. In each group (UV-treated and control), 48 h after the last UV treatment (T2), nine plants (3 rep × 3EU) were randomly sampled as UV-48 h and C-48 h. Also at T2 inoculation was performed on nine plants from each group but there was no sampling. Later, at 24 h post-inoculation (T3), nine plants from each inoculated group were sampled to provide UVi-24 h and Ci-24 h. At the same time (T3) as an un-inoculated reference set, the nine remaining plants from each group were sampled to provide UV-72 h and C-72 h. Once the leaf samples were severed from the plants, they were immediately flash-frozen in liquid nitrogen and stored at –80°C until RNA extraction.
The total RNA of the leaves (0.1 g) was extracted using a Qiagen total RNA isolation system (RNeasy Plant Mini Kit, 74904, Qiagen, Valencia, CA, United States) according to the manufacturer’s protocol. Any contaminating genomic DNA that may have been extracted along with the total RNA was removed using the RNase free DNase kit according to the instructions provided by the manufacturer (Qiagen, Germantown, United States). The verification of RNA sample purity was performed by the Bioanalyzer Agilent 2100 system using the Nano 6000 Assay RNA kit (Bioanalyzer 2100, CA, United States). This system provides the quantity and size of the RNA strands and the evaluation of the purity of the RNA samples. The RNA integrity number (RIN) minimum accepted was 5 and the mean RIN of the samples obtained was 8.
The transcriptomic analysis was performed by Genome Quebec (Canada). The cDNA libraries were built up from RNA samples by paired-end (PE) points sequencing following the Illumina Next-Generation Sequencing (NGS) protocol. The sequencing of the PE (2 × 100 bp) was performed on the Illumina HiSeq 2000 platform at a rate of 200 million sequences per line with an error rate < 0.1% (Illumina, San Diego, CA, United States). The RNA-seq library reads were trimmed using Trimmomatic 0.36 and aligned on lettuce reference genome Lsat_Salinas_v7 (GCF_002870075.1) using the STAR aligner v2.5.3a1.
The expression profile of 10 randomly selected lettuce genes from RNA-seq data was validated using qPCR. List of primers used in this study is presented in Supplementary Table 2. For each gene, three biological replicates with two technical replicates were used. The EXP45 gene was chosen as reference genes (Borowski et al., 2014). Prior to -qPCR, cDNA biosynthesis was performed from 5 μg of each RNA sample using the Maxima Mix reverse transcription kit (Thermo Scientific) according to the manufacturer’s instructions.
qPCR reactions were performed in a final volume of 20 μl and contained 10 μl of the ROX qPCR Master Mix from Thermo Fisher Scientific, 0.5 μM of each primer, and 2 μl of cDNA, equivalent to 20 ng of total RNA. PCR thermocycling conditions were set at 95°C for 3 min, 35 cycles at 95°C for 5 s, 60°C for 30 s, in a QuantStudioTM 3. Specificity of amplification as well as the absence of primer dimers was confirmed by melting curve analysis at the end of each reaction.
To correct for technical variations in reverse transcription and qPCR reactions as well as expression data for biological variations, these were normalized to the geometric mean of the EXP45 reference gene. The fold change (±SD) is expressed as treatment versus control is calculated by the 2-ΔΔCt method (Livak and Schmittgen, 2001).
Statistical Analysis
The experiment consisted of three independent repeats in a completely randomized design with three replicates of three experimental units, an experimental unit being 1 lettuce plant in a pot. For each experiment, the lettuce seedlings were randomly assigned to two separate groups 3 weeks after sowing. One group received the UV-C treatment according to the different modalities described above (see section “Ultraviolet-C Treatments”). Another group received no treatment and served as the control group. Symptoms of BLS data were subjected to a multi-factor analysis of variance (ANOVA) with interactions on R (v3.6.2), and Python was used to create the time-based disease assessment curve. The statistical procedures for the ANOVA and the time-based disease assessment curve can be found at https://bitbucket.org/asidibe2011/bls_data_analysis and https://bitbucket.org/asidibe2011/python_curve, respectively. RNA-Seq differential transcript expression levels were computed using Cufflinks v2.2.1, and differential relative gene expression analyses were computed using DESEQ and EDGER from the Bioconductor Rtools suite v3.5.0. Heatmaps were generated with R (v3.6.2) using the “pheatmap” package to show the relative expression of genes in the different samples. The procedures are available at https://bitbucket.org/asidibe2011/heatmaps/src/66fad937f2b5?at=master. Transcripts indicating a fold change ≥ 1.5 with FDR < 0.05 were confirmed to be differentially expressed. It should also be noted that the selection of the genes presented in the results is based on their function as described in the literature (Supplementary Table 3).
Results
Increased Resistance in Ultraviolet-C-Treated Lettuce to Xanthomonas campestris pv. vitians
Significant differences were observed between UV-C treated lettuce and controls in terms of Xcv symptom expression on days 4, 8, and 14 after inoculation (Figure 1). On day 4, UV-C treated lettuce leaves showed very few apparent symptoms, resulting in an AUDPC value close to 0, while on the leaves of the control lettuce, some dark and oily spots were observed, resulting in an AUDPC value of around 80. From day 8 on, the symptoms appearing on the lettuces were small necrotic spots, evolving to larger spots by day 14. This evolution was weak in UV-C treated lettuces compared to controls. As a result, AUDPC was significantly smaller in the treated group, with UV-C treatment resulting in a 62, 52, and 30% reduction in symptoms at 4, 8, and 14 days after Xcv inoculation, respectively, indicating an increase in AUDPC over time. However, this increase was significantly higher in control lettuces.
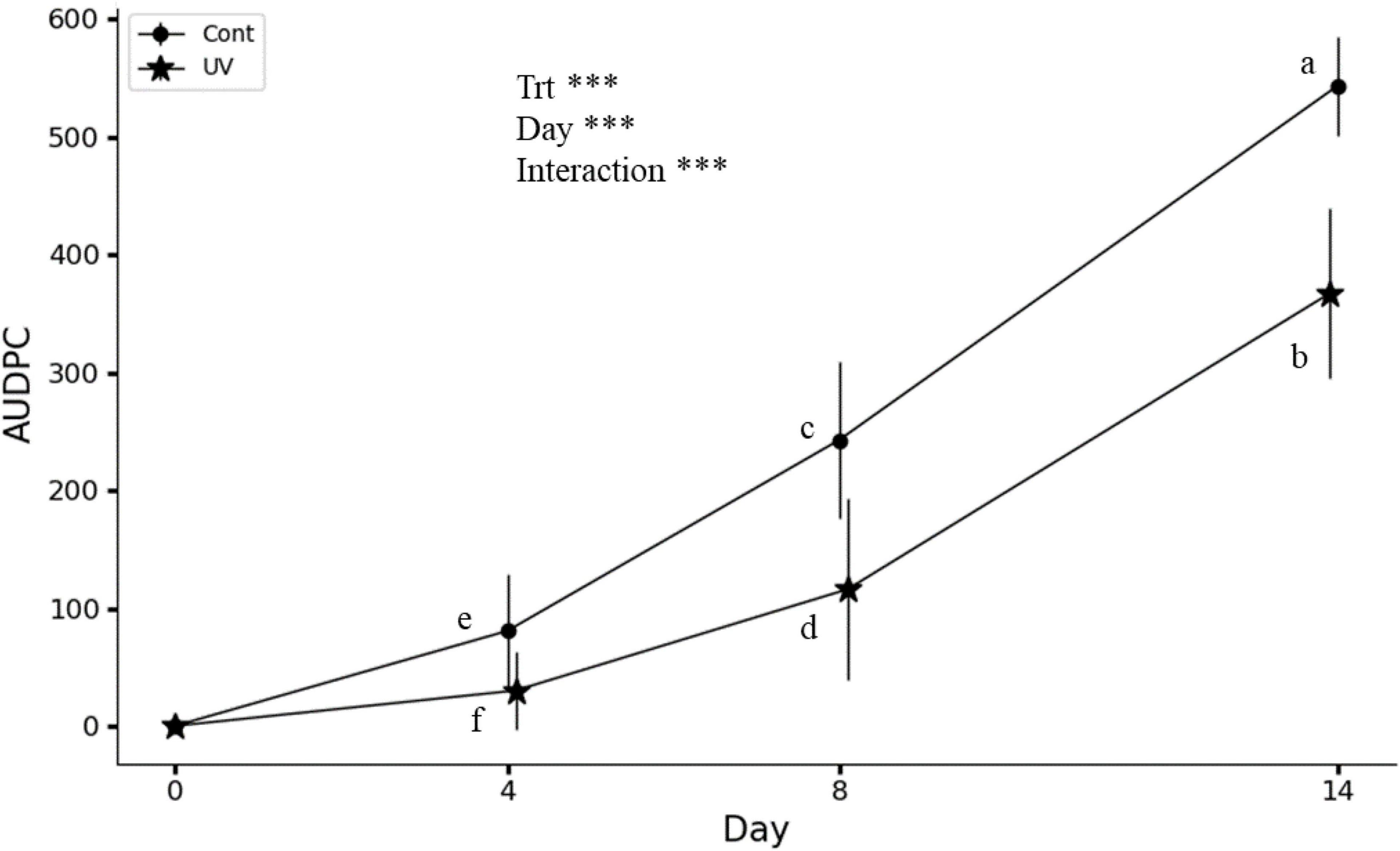
Figure 1. Effect of UV-C treatments on AUDPC (area under the disease progress curve). Treatments (Trt): Cont (control), UV (1.6 kJ/m2). Two-way ANOVA indicates a significant effect of treatments, assessment days (0, 4, 8, and 14) for BLS (bacterial leaf spot) and their interaction. AUDPC means are statistically different at P < 0.05 according to the Duncan’s multiple range test, n = 9, ***P < 0.001. Different letters indicate a significant difference between treatments at the same day or on different days (4, 8, and 14). The vertical bars show the standard deviation.
Differential Relative Expression of the RNA Sequences
RNA sequencing of the 54 samples from three biological replicas yielded a total gross readings of around 3 billion 94 million. Net readings of 3 billion 29 million, or around 97.88% of gross readings, were associated with the reference genome of Lactuca sativa L. (Supplementary Table 4). A total of 46,150 transcripts were identified. In UV-C treated lettuces, UV-48 h and UV-72 h and their controls C-48 h and C-72 h, respectively, 31538, 31371, 31370, and 31000 transcripts were detected, while in inoculated UV-C treated lettuces and the corresponding inoculated controls, 30930 and 31231 transcripts were detected, respectively (Supplementary Figure 2). The number of differentially expressed genes were 265, 2329, 2683, 2399, and 2653 in UV-48 h, UV-72 h, C-72 h, Ci-24 h and UVi-24 h respectively (Table 1 and Supplementary Figure 3).
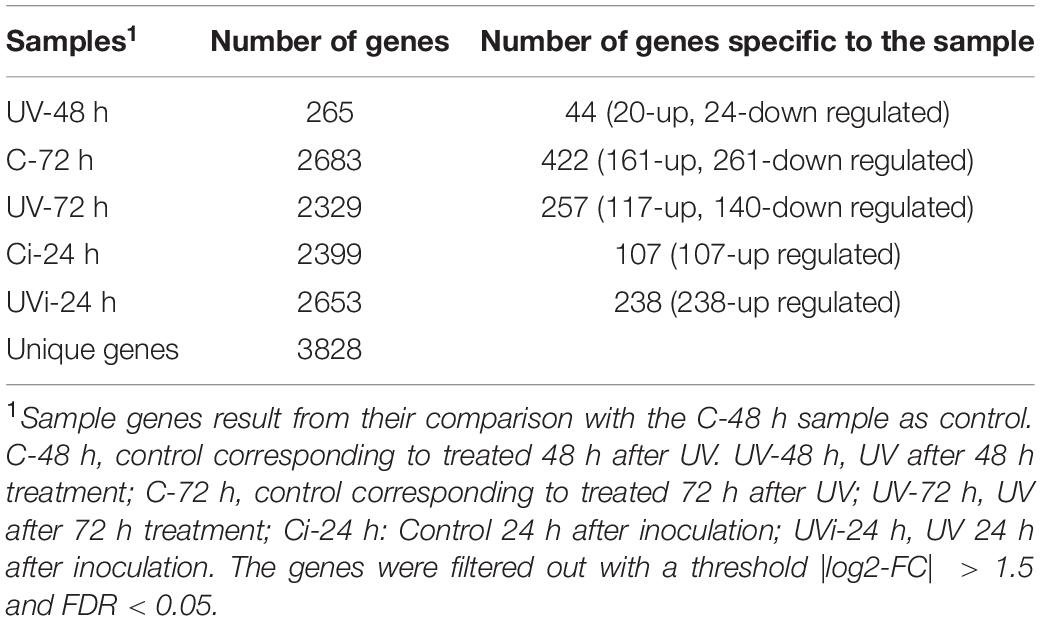
Table 1. Overall number of unique genes per sample.
Overall Functions of Genes Affected by Preharvest Ultraviolet-C and Xcv
By comparing the identified transcripts of the samples from the different groups of lettuce with the C-48 h, 3828 differentially expressed transcripts with |log2-FC| > 1.5 and false discovery rate (FDR) < 0.05 were detected. The functions of 59% of these transcripts (2270) have been defined (Supplementary Table 5). Among these 2270 genes, some were unique to the different treatments: 44 genes belonged to UV-48 h, 257 genes to UV-72 h, 422 genes to C-72 h, 107 genes to Ci-24 h, and 238 genes to UVi-24 h. These differentially expressed genes are associated with 14 biological functions including stress and antioxidant response (Table 2 and Figure 2); cell homeostasis and cell cycle (Table 3); metal transport (Figure 2); cell morphology and plant growth (Table 4); primary metabolism (Figure 3 and Supplementary Table 4); cell wall (Table 5); defense mechanisms (Figure 4); chloroplasts, phytohormones and other phenylpropanoids (Table 6). These differentially expressed genes can therefore explain the modification of the lettuce transcriptome following the effect of UV-C treatments followed by Xcv inoculation at three levels: homeostasis state, resumption of normal growth and activation of defense mechanisms.

Table 2. Relative expression of stress and antioxidant response genes.
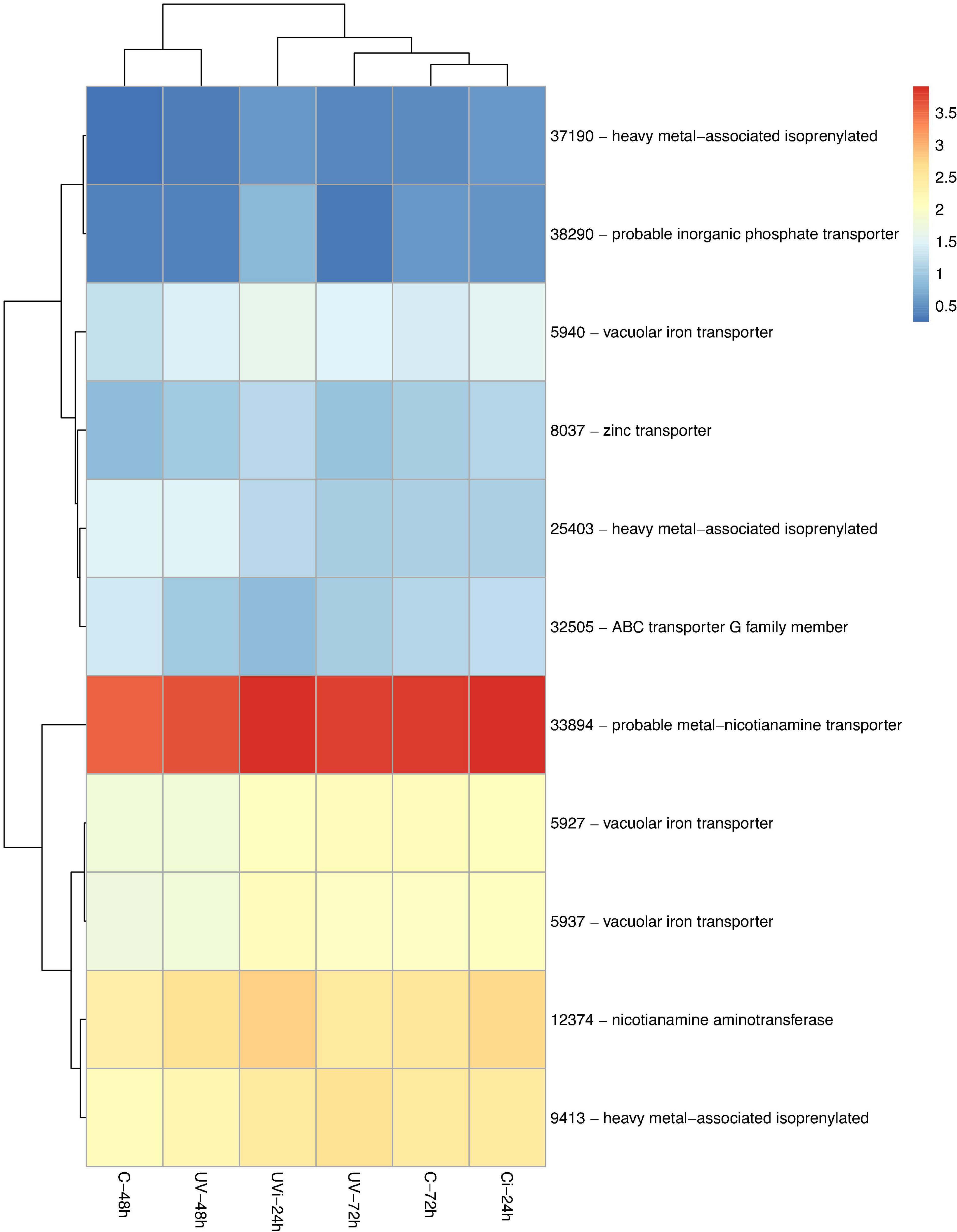
Figure 2. Genes related to metal transport differentially expressed in response to UV and Xcv inoculation. Test of hierarchical DEG (HCA) groupings. The colors reflect the level of relative gene expression. The columns and rows represent the samples and ID_genes associated with the name/function of the corresponding proteins that were grouped according to their relative expression profile, respectively. C-48 h, control corresponding to treated 48 h after UV; UV-48 h, UV after 48 h treatment; C-72 h, control corresponding to treated 72 h after UV; UV-72 h, UV after 72 h treatment; Ci-24 h, control 24 h after inoculation; UVi-24h, UV 24 h after inoculation. The genes were filtered out with a threshold |log2-FC| > 1.5 and FDR < 0.05.

Table 3. Relative expression of cell homeostasis and cell cycle arrest genes.
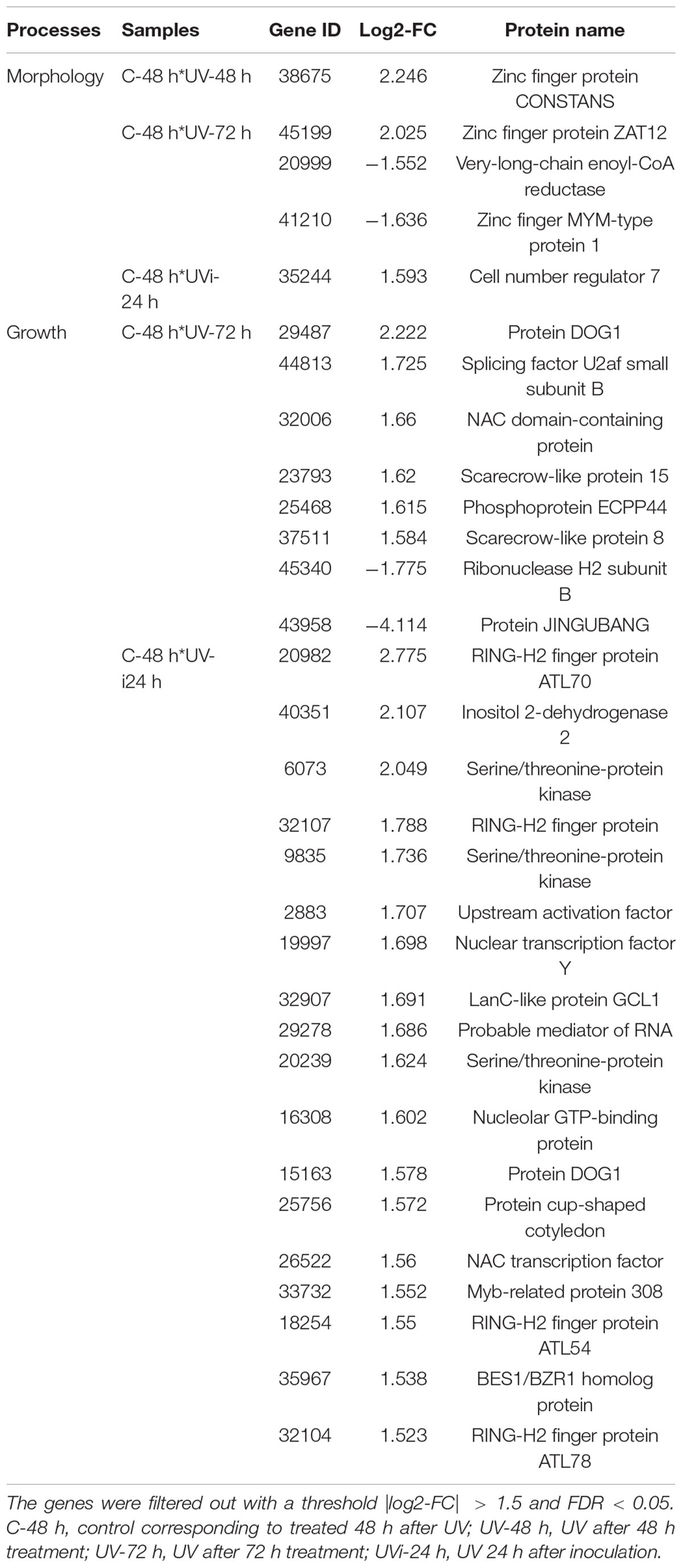
Table 4. Relative expression of cell morphology and growth genes.
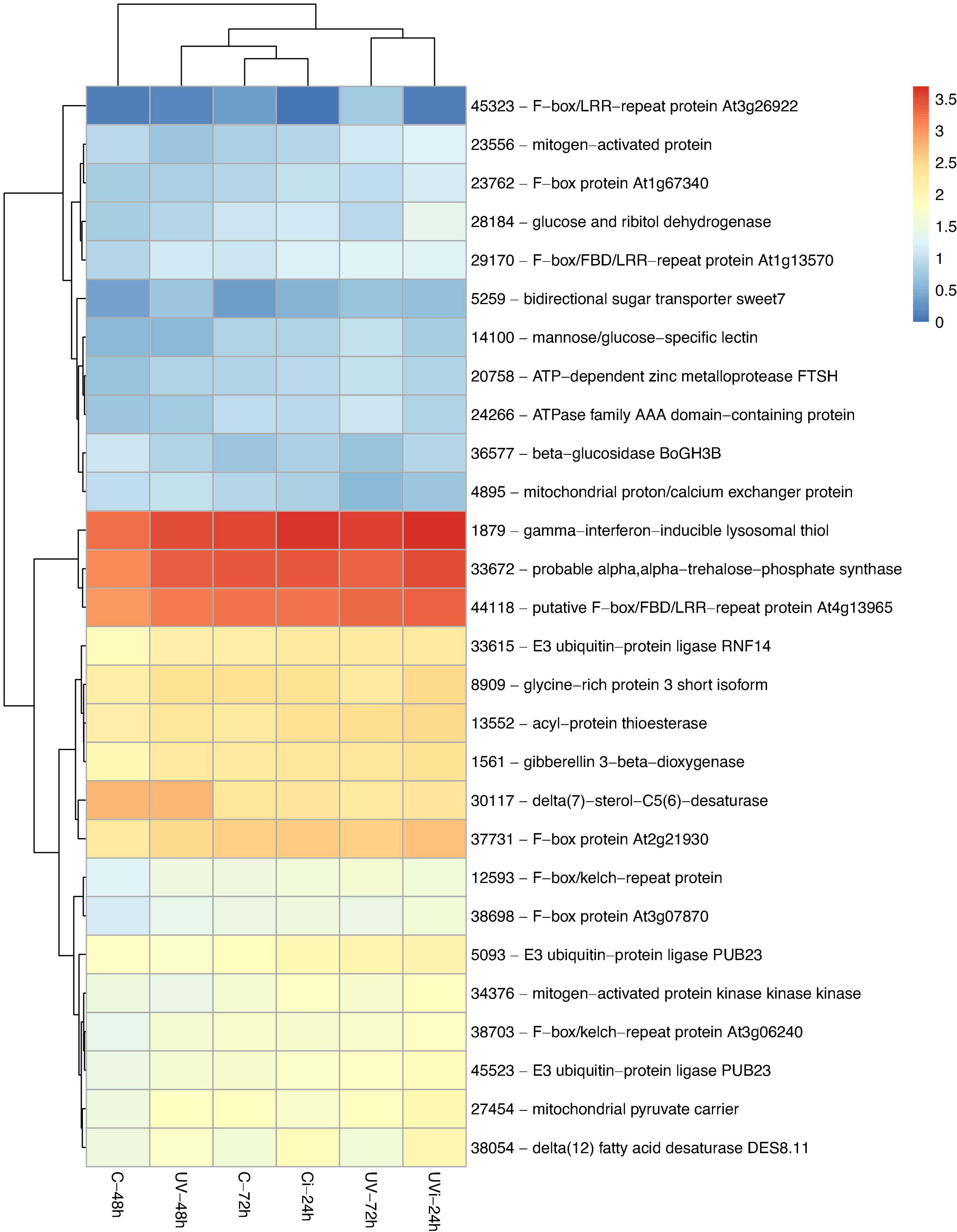
Figure 3. Primary metabolism-related genes differentially expressed in response to the influence of UV and Xcv inoculation. Test of hierarchical DEG (HCA) groupings. The colors reflect the level of relative gene expression. The columns and rows represent the samples and ID_genes associated with the name/function of the corresponding protein that were grouped according to their relative expression profile, respectively. C-48 h, control corresponding to treated 48 h after UV; UV-48 h, UV after 48 h treatment; C-72 h, control corresponding to treated 72 h after UV; UV-72 h, UV after 72 h treatment; Ci-24 h, control 24 h after inoculation; UVi-24 h, UV 24 h after inoculation. The genes were filtered out with a threshold |log2-FC| > 1.5 and FDR < 0.05.
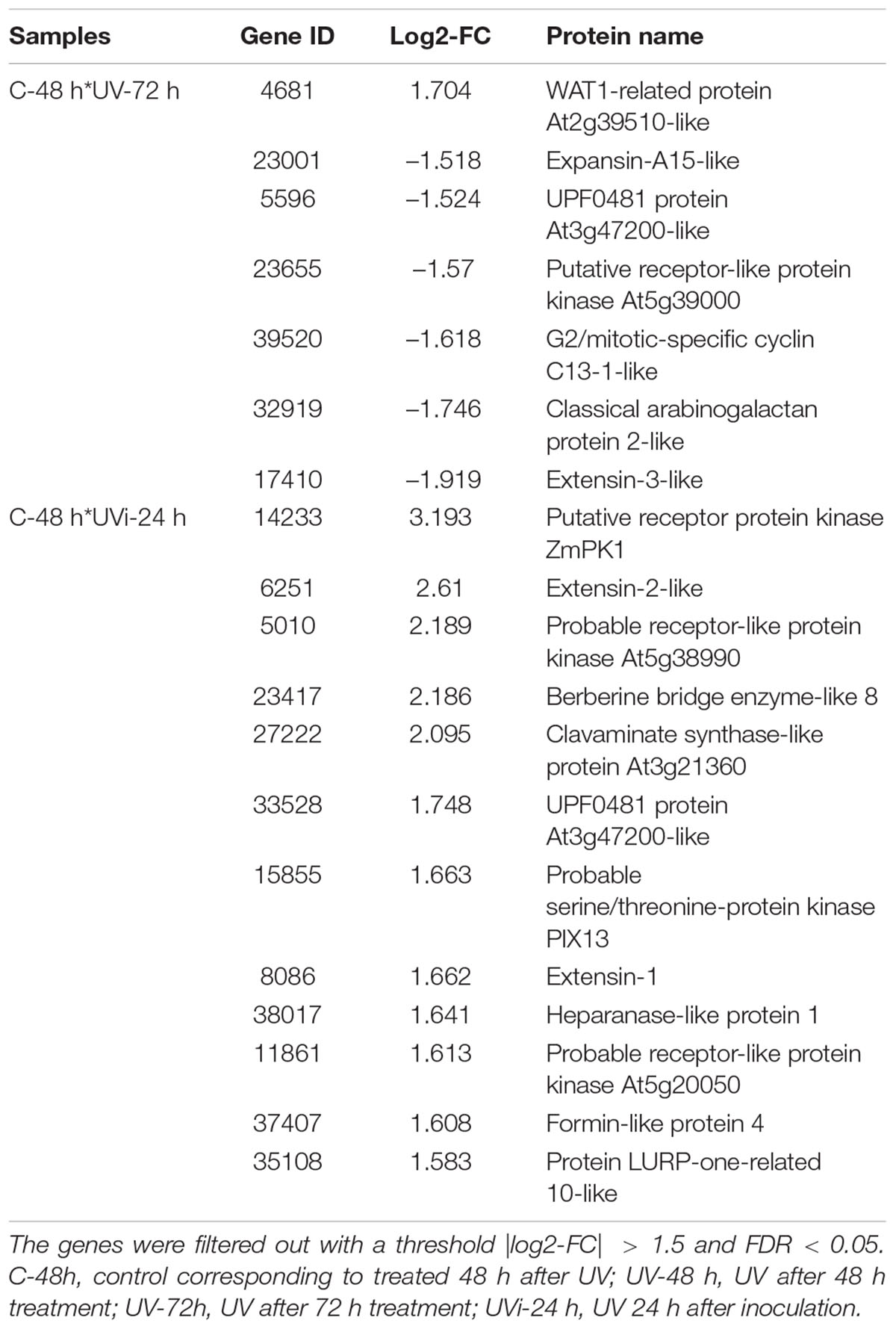
Table 5. Relative expression of cell wall genes.
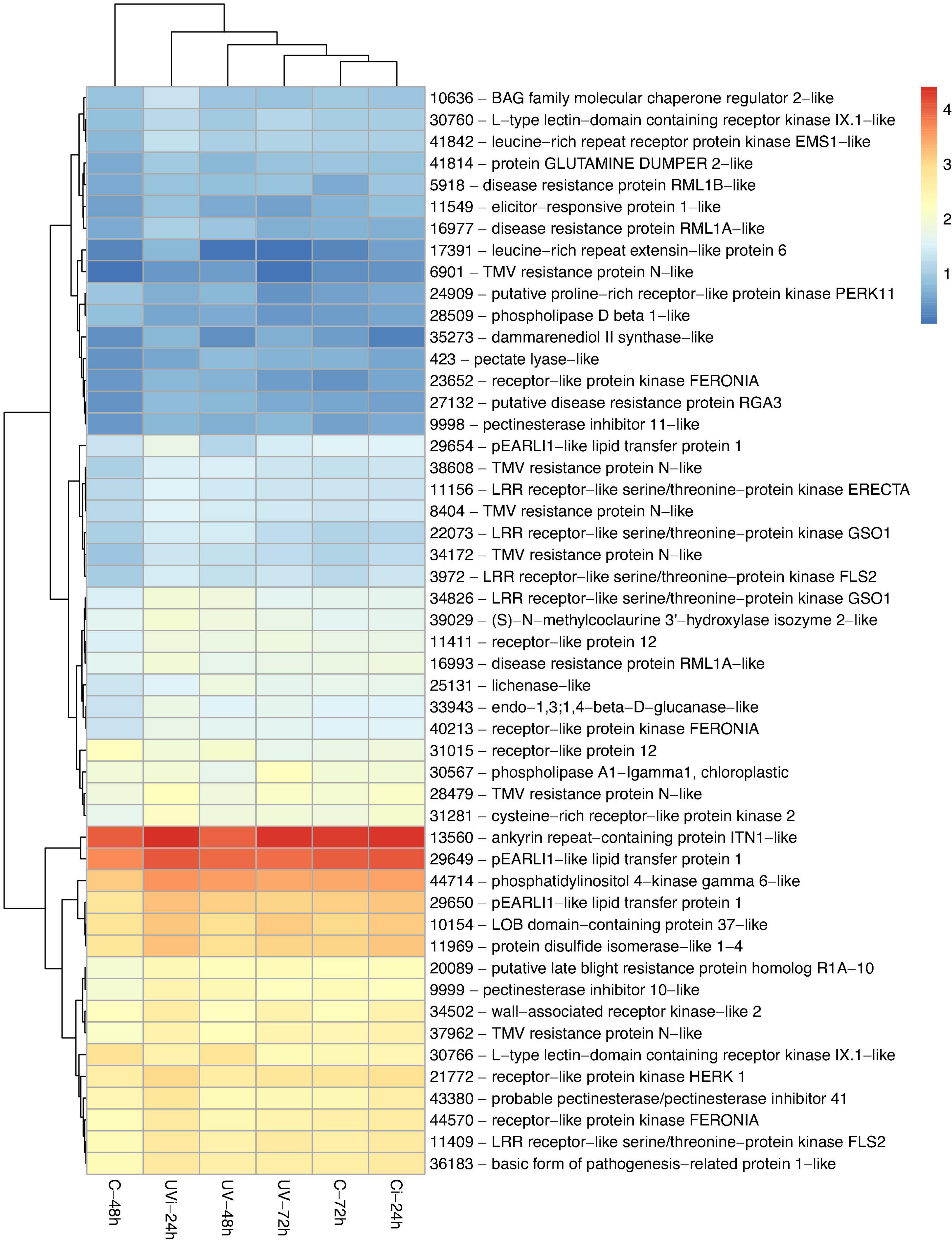
Figure 4. Genes related to defense mechanisms differentially expressed in response to UV and Xcv inoculation. Test of hierarchical DEG (HCA) groupings. The colors reflect the level of relative gene expression. The columns and rows represent the samples and ID_genes associated with the name/function of the corresponding protein that were grouped according to their relative expression profile, respectively. C-48 h, control corresponding to treated 48 h after UV; UV-48 h, UV after 48 h treatment; C-72 h, control corresponding to treated 72 h after UV; UV-72 h, UV after 72 h treatment; Ci-24 h, control 24 h after inoculation; UVi-24 h, UV 24 h after inoculation. The genes were filtered out with a threshold |log2-FC| > 1.5 and FDR < 0.05.
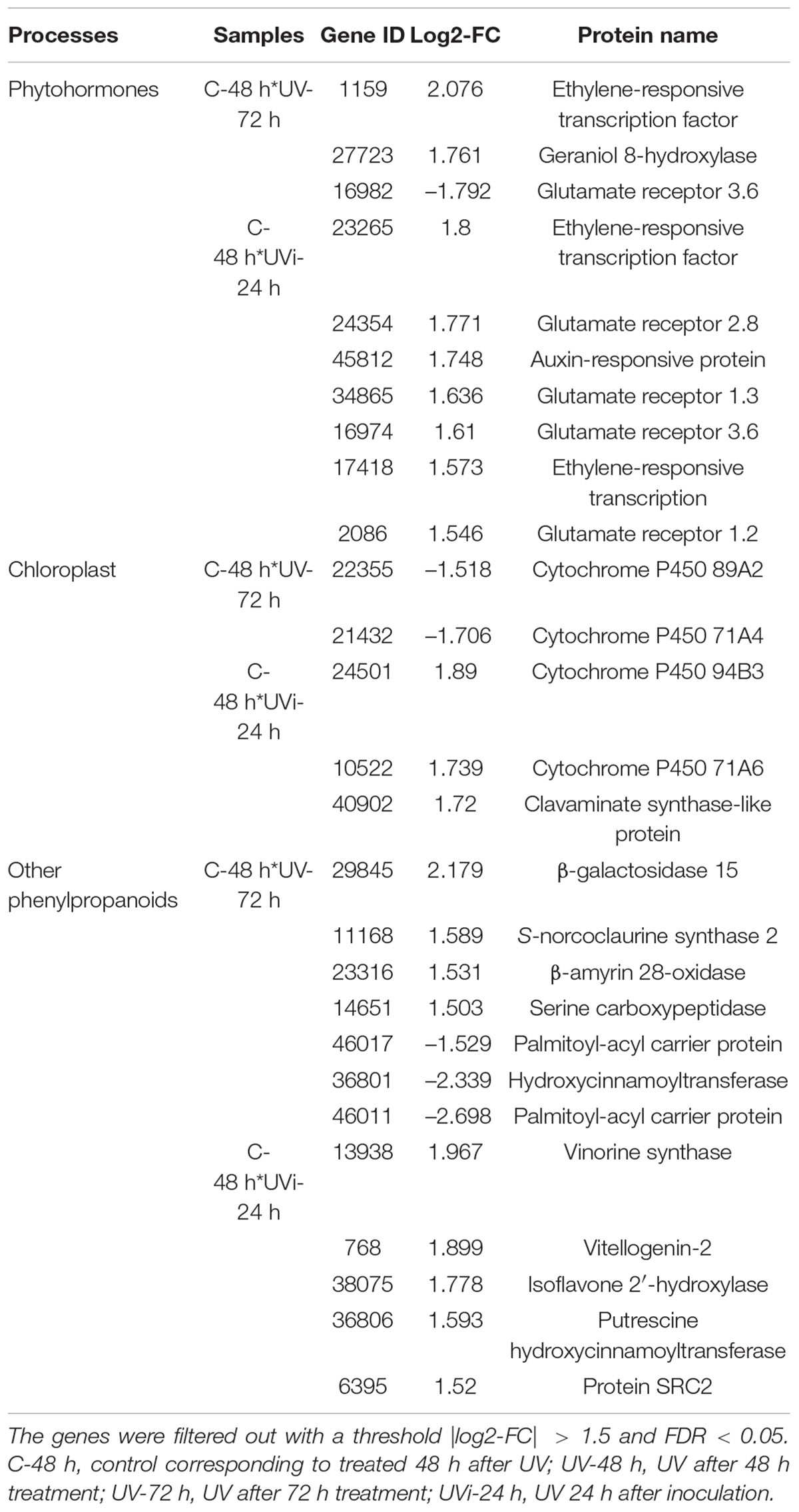
Table 6. Relative expression of phytohormones, chloroplast, and phenylpropanoids biosynthesis genes.
Genes Linked to Homeostasis State
Table 2 and Figure 2 show that the combined effect of UV-C and inoculation resulted in the relative overexpression of 13 genes, including those of the WRKY TFs, while the single effect of UV-C 72 h after treatment resulted in the relative overexpression of four genes and the downregulation of a single gene. At the level of genes associated with WRKY TFs and upregulated, there was only one gene (40915) induced by UV-C and two genes (25732 and 23563) induced by the combined effect of UV-C and inoculation. In addition, the combined effect of UV-C and inoculation resulted in positive regulation of the genes 22470, 44623, 31324, and 1939 associated with homogentisate phytyltransferase (HPT), peroxidase N1-like, probable carboxylesterase and epidermis-specific secreted glycoprotein (EP1), respectively.
A total of six genes (19989, 27642, 39875, 4352, 11830, 33035) linked to cell homeostasis calcium-binding protein PBP1-like, calcium-binding protein CML38-like, inorganic phosphate transporter 1-4-like, AAA-ATPase At5g57480-like, protein early flowering 3 and calcium-dependent protein kinase 7-like were differentially expressed (Table 3). The differential expression of these genes is due to the combined effect of UV-C and Xcv, with the exception of the genes encoding calcium-binding protein PBP1-like, calcium-binding protein CML38-like, inorganic phosphate transporter 1-4-like, which are due respectively to the effect of UV-C 48 h and 72 h after the last treatment. All these genes were upregulated with the exception of gene 19989 encoding a calcium-binding protein PBP1-like.
The gene 27642 linked to the calcium binding protein CML38-like (calmodulin-like proteins) and the gene 33035 linked to the calcium-dependent protein kinase 7-like were positively regulated in samples UV-72h and UVi-24h, respectively. In the UV-48h samples, gene 38678, associated with the U-box domain-containing protein 1-like, was observed to be downregulated. In the UV-72h samples, the genes (14274, 7029) linked to cyclin D5-3-like and 19-like protein containing the U-box domain were downregulated and upregulated, respectively.
A treatment-dependent regulation of three heavy-metal-binding protein biosynthesis genes (25403, 37190, and 9413) was observed (Figure 2). Genes 25403 and 9413 are respectively downexpressed and overexpressed in response to UV-72h, while lettuce in response to UVi-24 h resulted in positive regulation of gene 37190. In addition, UVi-24 h caused the relative overexpression of three genes (5927, 5937, and 5940) linked to the vacuolar iron transporter1 (VIT1) and one gene (12374) linked to nicotianamine aminotransferase A-like (NA).
Genes Linked to Resumption of Normal Growth
Among the genes involved in cell morphology, five were differentially expressed (Table 4) in UVi-24 h samples, one gene, 35244, which is associated with the cell number regulator 7-like (CNR), is overexpressed. In samples UV-48 h and UV-72 h, a positive regulation is observed with gene 38675, associated with zinc finger protein, Constans-like 10-like, and gene 45199, associated with zinc finger protein ZAT12-like, respectively.
Table 4 also indicates the upregulation of four genes (18254, 20982, 32107, and 32104) associated with ATL (RING-H2 finger protein) and of gene 35967 linked to BRI1-EMS suppressor1/brassinazole-resistant1 (BES1/BZR1) homolog protein 4-like in response to the effect of UV-C and Xcv (Table 2). The effect of UV-C alone 72 h after treatment resulted in the upregulation of gene 25468 linked to phosphoprotein ECPP44-like.
Concerning photosynthesis, gene 7707 associated with ruBisco (ribulose-1,5-bisphosphate carboxylase/oxygenase) was positively regulated following UV-C 48 and 72 h after the last treatment and also in response to the combined effect of UV-C and Xcv with respective FDR of 0.006, 1.4E-13, 7.2E-14 and log2-FC of 1.42, 2.84, and 2.96. Similarly, gene 35073 linked to bisphosphoglycerate was overexpressed by the combined effect of UV-C and Xcv, with a FDR of 1.5E-14 and log2-FC of 0.7.
The heatmap presented in Figure 3 show the relative expression of the genes mainly involved in primary metabolism. They include a group of five highly induced genes (38054, 1879, 33672, 28184, and 23004) encoding delta (12) fatty acid desaturase (DES8.11), gamma-interferon-inducible lysosomal thiol reductase, probable alpha alpha-trehalose phosphate synthase (TPS10) and glucose ribitol dehydrogenase-like. There were also two groups of moderately expressed genes linked to the F-box family. Of the F-box family, the gene associated with the repeat protein F-box/LRR At3g26922 is strongly overexpressed in response to UV-C, 72 h after the last treatment, with a fold change > 3 (Supplementary Table 5).
Several genes in Figure 3 and Table 3 are involved in protein ubiquitination. These are the genes associated with E3 ubiquitin-protein ligase PUB23-like (gene 45523), E3 ubiquitin-protein ligase PUB23-like (gene 5093), and E3 ubiquitin-protein ligase RNF14-like (gene 33615), with respective fold changes of 1.863, 1.539, and 1.503 (Figure 3 and Supplementary Table 5). In the treated and inoculated lettuce, three upregulated genes (9509, 44951, and 26329) were differentially expressed: cyclin-dependent kinase inhibitor 3-like, U-box domain-containing protein 13-like and U-box domain-containing protein 29-like. Two protein-associated genes (20758 and 24266) of the protease subfamily, namely the ATP-dependent zinc metalloprotease FtsH 6, located in the chloroplast, and At1g05910-like, a member of the ATPase family AAA (ATPase Associated with various cellular Activities), were positively regulated, with a respective fold change of 1.6 and 1.5 (Figure 3 and Supplementary Table 5) in response to the UV-C effect 72 h after treatment.
Our results indicate a positive upregulation of two genes (34376 and 23556) linked to mitogen-activated protein kinase 18-like, one gene (27454) related to mitochondrial pyruvate carrier (Figure 3 and Supplementary Table 5), and one gene (41814) linked to protein glutamine dumper 2-like (Figure 4) following the combined effect of UV-C and the pathogen Xcv.
In UV-72h samples, a single gene 4681 linked to WAT1-related protein At2g39510-like was overexpressed. However, in response to the combined effect of UV-C and inoculation, all 12 genes observed in Table 5 (14233, 6251, 5010, 23417, 27222, 33528, 15855, 8086, 38017, 11861, 37407, and 35108) were positively regulated and were associated with 12 different cell wall-related proteins. Among the proteins positively regulated in response to the combined effect of UV-C and inoculation are two clavaminate synthase-like proteins (Tables 2, 3). The combined effect of UV-C and inoculation also resulted in the relative overexpression of five genes (Figure 4 and Tables 4, 5) coding for proteins related to the maintenance of the cell wall: BES1/BZR1 homolog protein, leucine-rich repeat extensin protein (LRX), leucine-rich repeat receptor protein kinase and two extensin proteins.
Genes Linked to Activation of Defense Mechanisms
Our results (Figure 4) show a relative overexpression of five genes (11156, 3972, 11409, 34826, and 22073) linked to the receptor-like kinase (RLK) family protein biosynthesis and one ankyrin repeat-containing protein ITN1-like following the combined effect of UV-C and Xcv inoculation. The coding sequence of gene 28479, a precursor of the CRK proteins, was positively regulated in response to the combined effect of UV-C and inoculation.
As indicated in Figure 4, positive regulation was observed in response to the effect of UV-C and inoculation of three AZI1 genes (29649, 29650, and 29654), gene 10636, which is associated with the biosynthesis of Bcl-2-associated athanogene (BAG) family, and gene 36183, which is associated with the biosynthesis of PR-1 (Pathogenesis Related) proteins. The pEARLI1-like lipid transfer protein 1 biosynthesis genes are known as AZI1.
The effect of UV-C alone 72 h after the last treatment resulted in downregulation of gene 31015 involved in the biosynthesis of receptor-like proteins (RLPs) and gene 30766 associated with L-type lectin receptor kinase (LECRKs). However, the combined effect of UV-C and inoculation resulted in the positive regulation of five RLP genes (11411, 40213, 23652, 44570, and 21772), one LECRKs gene (30760), and one wall-associated receptor kinase-like 2 (WAK) protein biosynthesis gene (34502) (Figure 2).
The data from Figure 4 show relative overexpression in response to UVi-24h of 9 genes (6901, 38608, 28479, 34172, 37962, 8404, 20089, 17391, and 41842). Gene 20089 is a presumed homologue of the mildew resistance protein R1A-10, and genes 17391 and 41842 are associated with the biosynthesis of proteins of the leucine-rich repeated extensin (LRX) family. With respect to the cell wall, the effect of UVi-24 h resulted in the relative overexpression of two genes (9999 and 9998) associated with the inhibition of pectin methylesterase proteins (PME) and three genes (43380, 9999, and 9998) linked to the biosynthesis of cell wall proteins (CWPs) and pectin. Gene 33943, a precursor of β-1,3:1,4-Glucan, and gene 25131, associated with glycan metabolism, are highly overexpressed as a result of UV-C and inoculation.
Figure 4 also shows positive regulation of four other disease resistance genes. The relative overexpression of genes 16977, 16993, and 27132, which are related to disease resistance proteins RML1A-like, RML1B-like and RGA3, is due to the combined effect of UV-C and inoculation with Xcv. The upregulation of gene 5918 associated with disease resistance is due solely to the effect of UV-C 72 h after the last treatment. Genes 24266 and 20758, linked to the AAA ATPase family (Figure 3) overexpressed in response to the effect of UV-C, are also involved in plant defense.
The results presented in Table 6 indicate that two genes (21432 and 22355) associated with cytochrome P450 were downexpressed in UV-72h. However, the combined effect of UV-C and inoculation with Xcv resulted in a positive regulation of two other genes (10522 and 24501) linked to cytochrome P450 and two genes (40902 and 27222) linked to clavaminate synthase-like protein.
Our study showed 10 differentially expressed hormone-related genes (Tables 4, 6). The effect of UV light 72 h after the last treatment resulted in relative overexpression of gene 1159, which is associated with the ethylene-responsive transcription factor ERF017-like. In addition, the combined effect of UV-C and inoculation resulted in the relative overexpression of two genes (23265 and 17418) involved in the biosynthesis of ERF family proteins and of four genes (24354, 34865, 16974, 2086) associated with the glutamate receptor GLRs.
Table 6 shows the relative overexpression of five genes (29845, 11168, 23316, 14651, and 27723) in UV-72h samples. These genes are related to biosynthesis of several enzymes, namely, β-galactosidase 15-like, S-norcoclaurine synthase 2-like, β-amyrin 28-oxidase-like, serine carboxypeptidase-like and geraniol 8-hydroxylase-like (G8H).
Table 6 show that the combined effect of UV-C and inoculation with Xcv resulted in the positive regulation of six genes (13938, 768, 38075, 36806, 6395, and 39029) associated with the biosynthesis of vinorin synthase-like, vitellogenin-2-like, isoflavone 2′-hydroxylase-like, putrescine hydroxycinnamoyltransferase 1-like (PHT), protein SRC2-like, and (S)-N-methylcoclaurine 3′-hydroxylase isozyme (CYP80B1). Two other genes (16310 and 25705) linked to shikimate O-hydroxycinnamoyltransferase-like (CST) and caffeoylshikimate esterase-like (CSE), with respectively FDR of 2.5E-9, 7.2E-47 and log2-FC of 1.719, 5.196, were overexpressed in response to UV-C treatment and Xcv.
Comparison of RNA-seq Analysis to qPCR
Real-time quantitative polymerase chain reaction (qPCR) was performed on ten selected genes that were induced by the different treatments in our experimental conditions. Among these ten genes, ATL5 and ERF are associated with defense mechanisms, BGAL15, DES8.11, and FBXL22 are involved in plant growth recovery, FtsH, GLR1.2, MAPKK18, MAPKK19, and WAK2 play a role in the return of homeostasis state following UV effects. All treatments were included in this validation, and all ten genes showed consistent trend of fold change expression between RNA-seq and qPCR (Figure 5).
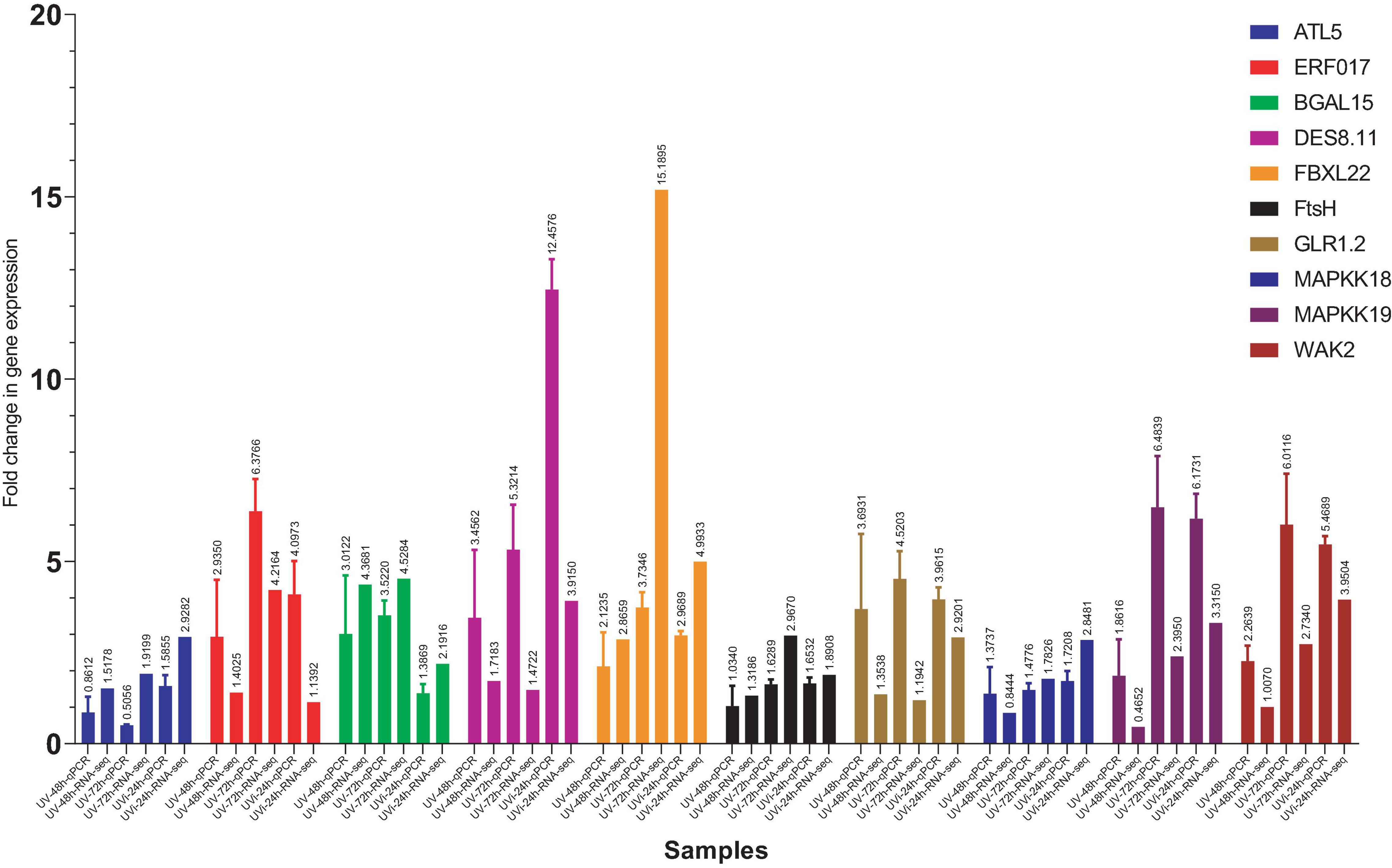
Figure 5. Comparison of gene expression levels between RNA-seq and qPCR. On the X-axis, we have qPCR data coupled to RNA-seq data for each gene and for each treatment: UV-48 h, UV after 48 h treatment; UV-72 h, UV after 72 h treatment; UVi-24 h, UV 24 h after inoculation. C-48h (Control corresponding to treated 48 h after UV) was used as a control.
Although the trend is similar for all tested genes (overexpression) for both PCR and RNSeq data, discrepancy in the amplitude of FCs can be observed for several genes. This is illustrated for example for ALT54 at UV-72 h (qPCR < RNA-seq), ERF017 at UV-48 h (qPCR > RNA-seq), ERF017 at UVi-24 h (qPCR > RNA-seq) and WAK2 at UV-72 h (qPCR > RNA-seq). At this point, we can only postulate that these apparent discrepancies are related to differences between the two techniques and handling error bias.
Discussion
This work confirms that a UV-C treatment of 1.6 kJ/m2 applied as described by Nicolas et al. (2020) falls within the hormetic dose range conferring a protective effect on lettuce against BLS. This is consistent with previous studies that have reported the efficacy of hermetic treatment of UV-C radiation in reducing the symptoms observed in a variety of plant-pathogen interactions (Janisiewicz et al., 2016a,b; Xu et al., 2019; Aarrouf and Urban, 2020; Nicolas et al., 2020; Vàsquez et al., 2020). Figure 6, which served as a framework for the discussion below, was developed using original data from the present study and available data from the literature. This figure shows that the early responses of lettuce treated with UV-C followed by Xcv inoculation led to a change in the transcriptome, which likely translated into a return to homeostasis, resumption of normal growth and the activation of defense mechanisms.
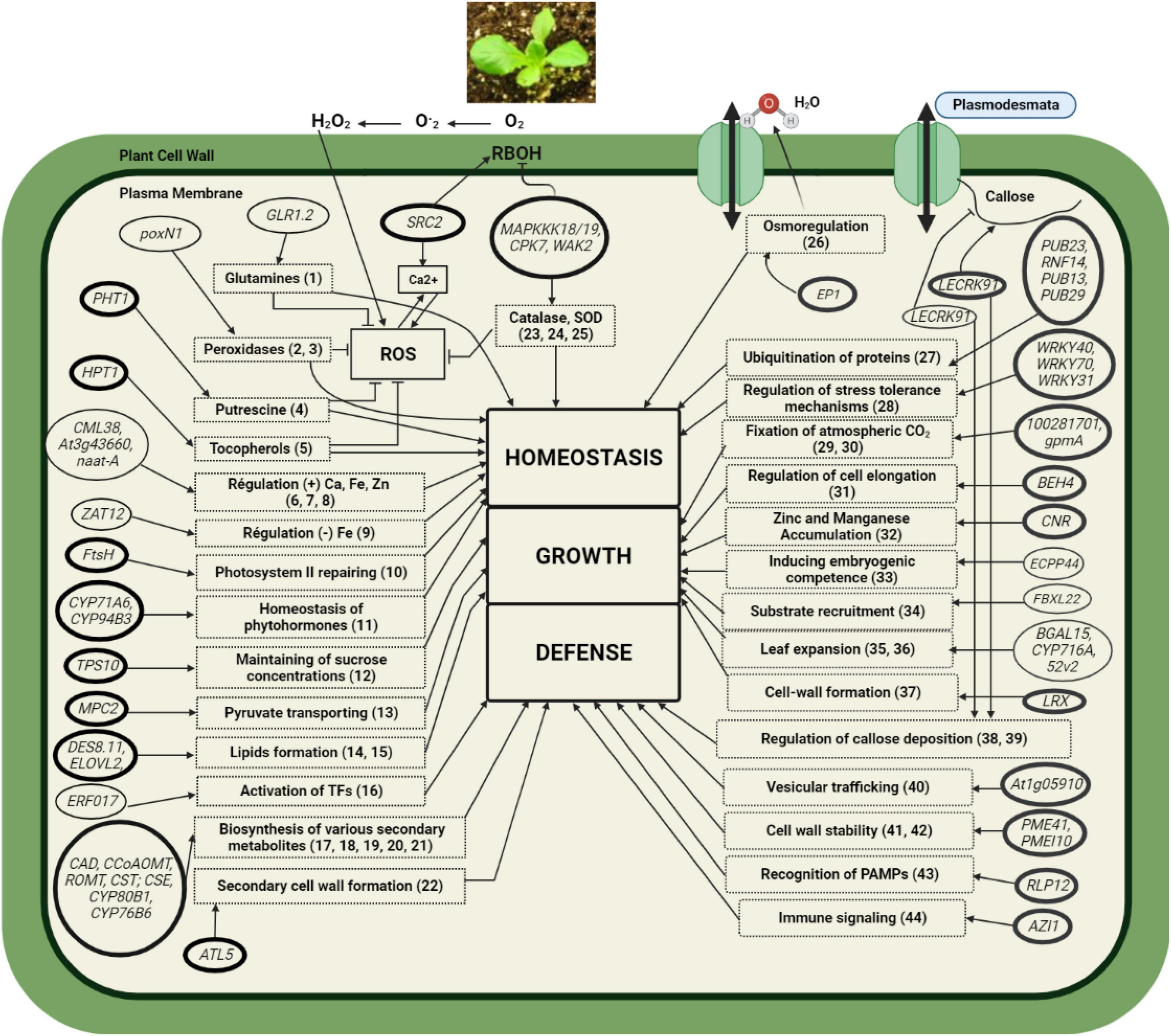
Figure 6. Representative model of the regulation mechanisms induced in lettuce by the preharvest application of hormetic doses of UV-C and inoculation with the pathogen Xcv. Thick-bordered rectangular boxes: Physiological (or biological) processes; dashed rectangular boxes: Function or molecules associated with genes as reported in the literature. Thin-bordered ellipses: genes affected by UV-C alone; thick-bordered ellipses: genes affected by UV-C + Xcv infection. The pointed arrows indicate activation reactions and the blunt arrows indicate inhibition reactions. The numbers in brackets in the rectangular boxes represent the reference source(s). (1) Cheng et al. (2018); (2) Kim et al. (2007); (3) Wang et al. (2016); (4) Pushpa et al. (2014); (5) Savidge et al. (2002); (6) Lokdarshi et al. (2016); (7) Gollhofer et al. (2014); (8) Cassin et al. (2009); (9) Le et al. (2016); (10) Kato and Sakamoto (2018); (11) Yan et al. (2016); (12) Lunn et al. (2014); (13) Shen et al. (2017); (14) Doan et al. (2009); (15) Nayeri and Yarizade (2014); (16) Dey and Vlot (2015); (17) Zhang and Liu (2015); (18) Ma et al. (2005); (19) Ziegler and Facchini (2008); (20) Sung et al. (2011); (21) Höfer et al. (2013); (22) Noda et al. (2013); (23) Zhou et al. (2014); (24) Bundó and Coca (2017); (25) Li et al. (2020); (26) van Engelen et al. (1993); (27) Kimura and Tanaka (2010); (28) Ashrafi-Dehkordi et al. (2018); (29) Hauser et al. (2015); (30) Li et al. (2018); (31) Chaiwanon et al. (2016); (32) Qiao et al. (2019); (33) Tan and Kamada (2000); (34) Skaar et al. (2013); (35) Ban et al. (2020); (36) Han et al. (2013); (37) Zhao et al. (2018); (38) Lannoo and Van Damme (2014); (39) De Storme and Geelen (2014); (40) Zhu et al. (2016); (41) Lionetti et al. (2017); (42) Niu and Wang (2020); (43) Jamieson et al. (2018); (44) Dutton et al. (2019).
Return to Homeostasis
The change in lettuce transcriptome demonstrated a return to the homeostasis state revealed by the relative overexpression of several genes. These genes act directly or indirectly on the production or removal of ROS to ensure the return and/or maintenance of homeostasis (Mittler et al., 2004).
Among those genes, the relative overexpression of SRC2 genes and protein kinase biosynthesis genes (MAPKK18/19, CPK7, WAK2) was found. The SRC2 genes induce the expression of NADPH oxidases, which are homologous proteins to respiratory burst oxidase homologs (RBOHs) (Kawarazaki et al., 2013). The latter are enzymes that contribute to the generation of ROS in response to hormonal and environmental signals (Suzuki et al., 2011). The SRC2 genes encoding SRC2 proteins are involved in the activation of Ca2+ influx-dependent ROS generation in Arabidopsis thaliana (Kawarazaki et al., 2013). The relative overexpression of SRC2 genes therefore suggests (Figure 6) ROS production. This would be in agreement with the work of Xu et al. (2014, 2019), who showed induction of ROS production in response to UV-C and bacterial flagellin, respectively.
In plants, RBOHs have an N-terminal cytosolic extension consisting of two Ca2+-binding EF-hand motifs and phosphorylation target sites that are important for their activity (Drerup et al., 2013). Considering the relative overexpression of SRC2, CML38 (calmodulin-like proteins), an increase in Ca2+ concentration can be presumed. Ca2+ is known to be essential to many plants cellular processes, including regulation of cellular ROS levels (Hu et al., 2018), division, differentiation, and programmed cell death (López-Fernández and Maldonado, 2015).
In contrast to SRC2, protein kinases negatively regulate RBOHD-dependent ROS production by, for instance, limiting the phosphorylation cascades that drive ROS production in response to stress (Escudero et al., 2019). Studies have shown that plants possess a comprehensive antioxidant system, consisting of enzymatic and non-enzymatic antioxidants, that can scavenge ROS (Baxter et al., 2014). Following the effect of UV-C treatment alone or in combination with Xcv inoculation, the relative overexpression of genes associated with the production of enzymatic antioxidants, such as SOD, CAT, and peroxidases (POD), and non-enzymatic antioxidants, such as tocopherols was found (Savidge et al., 2002). Tocopherols limit the oxidation of lipids to protect the thylakoid membrane from photo-oxidation. They also have antioxidant properties against oxidative damage caused by ROS. Our results also indicate an upregulation of metals such as Fe and Zn. Metal ions are cofactors of ROS detoxification enzymes (Ravet and Pilon, 2013). FtsH, an ATP-dependent metalloprotease with zinc as a cofactor, has been found to play a role in thylakoid membrane biogenesis, quality control in photosystem II repair, and the assembly of several protein complexes in photosynthetic electron transport pathways in A. thaliana (Kato and Sakamoto, 2018).
Our results indicate the relative overexpression of PUB, WRKY, and EP1 genes with UV-C treatments alone or in combination with Xcv inoculation. PUB genes in several plant species are involved in the regulation of intracellular trafficking and ROS production (Tian et al., 2015) and in cell homeostasis (Kimura and Tanaka, 2010; Proietti et al., 2014). Genes associated with the hypothetical WRKY transcription factors participate in plant response to biotic (Scarpeci et al., 2013) and abiotic stresses (Scarpeci et al., 2013; Buti et al., 2019), by enabling dynamic cellular homeostatic reprogramming (Phukan et al., 2016). Studies have hypothesized that EP1 genes may be involved in the regulation of water flow through the cell wall (van Engelen et al., 1993).
Resumption of Normal Growth
In addition to the return and/or maintenance of homeostasis, the effect of UV-C treatments alone or in combination with Xcv inoculation was followed by a resumption of normal growth likely corresponding to an increase in photosynthetic activity. This increase in photosynthetic activity could generate an increase in dry matter synthesis (Farahbakhsh et al., 2017). Our recent work has also indicated an increase in dry matter in lettuce following preharvest treatments with hormetic doses of UV-C (Sidibé et al., 2021). In the present study, the effect of UV-C treatments alone or in combination with Xcv resulted in the relative overexpression of 13 genes directly or indirectly associated with the resumption of normal growth. Four of these genes (ECPP44, BGAL15, FBXL22, CYP716A52v2) were induced in response to the effect of UV-C alone. During somatic embryogenesis in plants, the phosphoprotein ECPP44 induces embryogenic competence in plants (Tan and Kamada, 2000). The induction of ECPP44 in response to the single effect of UV-C is therefore consistent with the overexpression of genes associated with embryogenic processes in cassava in response to drought (Wu et al., 2018). The β-galactosidases, such as BGAL15, are involved in polysaccharide metabolism (Ban et al., 2020) and in the separation of cellulose microfibrils during wall extension and remodeling (Sampedro et al., 2012). The released galactose fragments can serve as energy sources and substrates, which would explain the observed relative overexpression of the FBXL22 gene. FBXL22 is associated with F-box family proteins and is known to be involved in substrate recruitment (Skaar et al., 2013), plant growth, signal transduction (Huo et al., 2014), and disease tolerance (Kumar et al., 2020). The CYP716A52v2 gene encodes the β-amyrin 28-oxidase enzyme, which catalyzes the triterpene hydroxylation responsible for the biosynthesis of oleic-type saponins (Han et al., 2013). Studies have shown that saponins and their biosynthetic intermediates are involved in plant growth and development (Moses et al., 2014) by stimulating chlorophyll biosynthesis, as observed in radish and pea (Boiteau and Ratsimamanga, 1956).
Seven genes (DES8.11, TPS10, BEH4, MPC2, CNR, and LRX) were induced in response to the joint effect of UV-C and Xcv. These seven genes code for proteins known to be involved in plant growth and development. The DES8.11 gene is associated with fatty acid desaturase proteins, which at the plasma membrane site produce unsaturated fatty acids by introducing double bonds into fatty acid hydrocarbon chains (Nayeri and Yarizade, 2014). DES8.11 gene induction is in agreement with the work of Gierz et al. (2017), who showed overexpression of fatty acid-encoding genes in algae (Symbiodinium sp.) in response to heat stress.
The TPS genes encode for a trehalose-6-phosphate synthase, which is a key enzyme for trehalose biosynthesis in yeast (Yeo et al., 2000). In beans (Phaseolus vulgaris), induction of trehalose-6-phosphate synthase biosynthesis has been shown to increase yield and improve drought tolerance (Suárez et al., 2008). BEH4 proteins of the BES1/BZR1 BES1 family integrate a variety of plant signaling pathways (Yin et al., 2005), such as brassinosteroid (BR) signaling. BRs are phytohormones involved in the regulation of cell elongation and lateral organ development (Chaiwanon et al., 2016). Pyruvate, as the end product of glycolysis, is derived from sources in the cell cytoplasm, and much of the pyruvate is transported into mitochondria by MPC2 proteins for oxidative metabolism of the Krebs cycle (Shen et al., 2017). Studies have shown that the CNR gene, which is relatively overexpressed in rice, has increased tolerance to otherwise toxic levels of Zn and Mn (Qiao et al., 2019). Mn has been shown to be an essential micronutrient for all stages of plant development, including as a catalytic center for photosystem II (Yang et al., 2007). The LRX gene is responsible for maintaining cell wall integrity through cross-linking with other cell wall components, such as pectin (Zhao et al., 2018). Overexpression of these genes (TPS10, BEH4, MPC2, CNR, and LRX) associated with cell tissue expansion is consistent with the increase in dry matter observed in lettuce following preharvest UV-C applications (Sidibé et al., 2021).
Activation of Defense Mechanisms
The model in Figure 6 proposes an activation of defense mechanisms in lettuce via the relative overexpression of 15 genes following the effect of UV-C alone or in combination with Xcv. The effect of UV-C treatments alone resulted in the induction of the ERF017 gene and the inhibition of the LECRK91 gene. Genes associated with the structural reinforcement of the cell wall (LECRK91, RLP12, ATL5, PMEI10, PME41, AZI1, At1g05910, CYP80B1, CYP76B6, PAL, CST, and CSE) were induced by the effect of UV-C and Xcv. The induction of these genes suggests a “priming effect” of UV-C treatments in lettuce against the pathogen.
The inhibition of the LECRK91 gene in response to the single effect of UV-C and its activation following the combined effect of UV-C and Xcv observed in the present study is not surprising. In A. thaliana, inhibition of LECRK gene expression has been established under various oxidative stresses (UV-B, drought, heat) (Mondal and Das, 2020). Conversely, overexpression of LECRK family genes was observed in response to infection by bacterial pathogens (Balagué et al., 2017). Receptor-like proteins (RLPs) often associate with receptor-like kinases (RLKs) and are involved in the development of immunity and growth (Jamieson et al., 2018). RLK proteins (Quezada et al., 2019) and Bcl-2-associated athanogenes (BAGs) (Williams et al., 2010) have been shown to cause cell death associated with a hypersensitivity reaction. For their part, the lectin receptor kinase (LECRK) and L-type lectin domain proteins (LLP) proteins (Hofberger et al., 2015) are pathogen recognition receptors (PRRs) involved in the recognition of carbohydrate structures of microbial organisms (Lannoo and Van Damme, 2014). The ERF gene induces the Ethylene-Responsive Factor (ERF) family of proteins, which in turn induce physiological responses to stress in plants (Dey and Vlot, 2015). Secondary wall biosynthesis in plants is induced by the ATL5 gene (Noda et al., 2013). Also, it should be noted that the RING domain of ATL proteins is essential for E3 ubiquitin ligase (PUB) function in response to pathogen invasion (Deng et al., 2017). Therefore, it would be logical to assume that some complementarity between ATL and PUB proteins enabled the control of BLS observed in the present study.
Pectin methylesterase proteins can contribute to the generation of pectic fragments, some of which are reported to induce plant defense mechanisms (Wormit and Usadel, 2018). As PME activity is modulated by pectin methylesterase inhibitors (PMEI) (Wormit and Usadel, 2018), it is therefore appropriate to speculate that in this study, in conjunction with the relative overexpression of PME, the increased activity of PMEI10 contributes to protecting the lettuce cell wall from degradation by Xcv-secreted pectinases. It is well known that Xcv pectinases are important factors of virulence (Kazemi-Pour et al., 2004).
AZI1 genes inducing pEARLI1-like lipid transfer protein biosynthesis link to glycerol-3-phosphate accumulation required for systemic acquired resistance conferred by azelaic acid (Yu et al., 2013). In rice plants, the AAA ATPase family protein-associated gene At1g05910 is involved with the PUB genes in the induction of multivesicular body (MVB) trafficking of the plant defense system (Zhu et al., 2016). This result therefore suggests a complex feedback regulatory loop between AZI1, AAA ATPases, PUB, and glycerol-3-phosphate proteins.
In A. thaliana, the Geraniol 8-hydroxylase (G8H) (CYP76B6) biosynthetic gene have been shown to be involved in the flavonoid (Sung et al., 2011), alkaloid, and monoterpenoid biosynthetic pathway (Höfer et al., 2013). The precursors of G8H enzymes are derived from the deamination of L-phenylalanine (Baucher et al., 1998) which is catalyzed by PAL, the main entry point in the biosynthetic pathway of phenylpropanoids, including flavonoids, which play a role in pathogen defense and UV protection (Buer et al., 2010). Other enzymes, such as Caffeoyl shikimate esterase (CSE) and Shikimate O-hydroxycinnamoyltransferase-like (CST), which are involved in the lignin biosynthetic pathway (Vanholme et al., 2013), are relatively overexpressed in response to the effect of UV-C and Xcv. In addition to the overexpression of the latter two genes (CSE and CST), the overexpression of ATL5 and LECRK91 genes suggests that the protection of UV-C-treated lettuce would be partly due to the reinforcement of structural barriers. This UV-C effect was previously observed in tomato treated at the postharvest stage (Charles et al., 2008).
Conclusion
The control of BLS following the application of the total UV-C treatment of 1.6 kJ/m2 observed in this study can be explained by a change in several physiological processes in lettuce. Transcriptome analysis of lettuce in response to UV-C alone or in combination with Xcv showed that the differentially expressed genes are associated with homeostasis, growth and defense. These results allow us to conclude that UV-C hormesis applied under the conditions described in the present study is an effective eustress that does not interfere with the ability of treated plants to resume normal growth or to defend themselves against potential stressors.
Data Availability Statement
The original contributions presented in the study are publicly available in NCBI under accession number SUB10516369.
Author Contributions
AS, MTC, and CB conceived and designed the study. AS performed the experiments. AS, MTC, J-FL, and YX analyzed the data. AS, MTC, and CB wrote the manuscript with valuable contributions from J-FL and YX. All authors read and approved the final manuscript.
Funding
This work was financially supported by Agriculture and Agri-Food Canada (Grant # J-001300) and the AgroPhytoSciences program at Centre SÈVE, Sherbrooke University.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
The authors would like to thank Karine Fréchette and Bertrand Riendeau for maintenance of lettuce plants; Daniel Rolland, Dominique Roussel, Pierre-Yves Véronneau, Marie Ciotola, and Mélanie Cadieux for their assistance in monitoring the experiments; Benjamin Mimee, Etienne Lord, Martin Laforest, and Joel Lafond-Lapalme for their assistance in bioinformatics; as well as Sylvain Lerat and Mamadou Lamine Fall for critical review of the manuscript.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2021.793989/full#supplementary-material
Footnotes
References
Aarrouf, J., and Urban, L. (2020). Flashes of UV-C light: an innovative method for stimulating plant defences. PLoS One 15:e0235918. doi: 10.1371/journal.pone.0235918
Achuo, E. A., Prinsen, E., and Höfte, M. (2006). Influence of drought, salt stress and abscisic acid on the resistance of tomato to Botrytis cinerea and Oidium neolycopersici. Plant Pathol. 55, 178–186. doi: 10.1111/j.1365-3059.2006.01340.x
Ashrafi-Dehkordi, E., Alemzadeh, A., Tanaka, N., and Razi, H. (2018). Meta-analysis of transcriptomic responses to biotic and abiotic stress in tomato. PeerJ 6:e4631. doi: 10.7717/peerj.4631
Bagati, S., Mahajan, R., Nazir, M., Dar, A. A., and Zargar, S. M. (2018). “‘Omics’: a gateway towards abiotic stress tolerance,” in Abiotic Stress-Mediated Sensing and Signaling in Plants: An Omics Perspective, eds S. Zargar and M. Zargar (Singapore: Springer), 1–45. doi: 10.1007/978-981-10-7479-0_1
Balagué, C., Gouget, A., Bouchez, O., Souriac, C., Haget, N., Boutet-Mercey, S., et al. (2017). The Arabidopsis thaliana lectin receptor kinase LecRK-I. 9 is required for full resistance to Pseudomonas syringae and affects jasmonate signalling. Mol. Plant Pathol. 18, 937–948. doi: 10.1111/mpp.12457
Ban, Q., Jiao, J., He, Y., Jin, M., and Rao, J. (2020). Ectopic expression of the persimmon β-galactosidase gene DkGAL2 promotes leaf growth, delays dark-induced senescence and enhances tolerance to abiotic stress in Arabidopsis. Sci. Hortic. 265:109232. doi: 10.1016/j.scienta.2020.109232
Baucher, M., Monties, B., Montagu, M. V., and Boerjan, W. (1998). Biosynthesis and genetic engineering of lignin. Crit. Rev. Plant Sci. 17, 125–197. doi: 10.1080/07352689891304203
Baxter, A., Mittler, R., and Suzuki, N. (2014). ROS as key players in plant stress signalling. J. Exp. Bot. 65, 1229–1240. doi: 10.1093/jxb/ert375
Boiteau, P., and Ratsimamanga, A. R. (1956). Asiaticoside extracted from Centella asiatica and its therapeutic uses in cicatrization of experimental and refractory wounds (leprosy, cutaneous tuberculosis and lupus). Therapie 11, 125–149.
Borowski, J. M., Galli, V., da Silva Messias, R., Perin, E. C., Buss, J. H., e Silva, S. D. D. A., et al. (2014). Selection of candidate reference genes for real-time PCR studies in lettuce under abiotic stresses. Planta 239, 1187–1200. doi: 10.1007/s00425-014-2041-2
Buer, C. S., Imin, N., and Djordjevic, M. A. (2010). Flavonoids: new roles for old molecules. J. Integr. Plant Biol. 52, 98–111. doi: 10.1111/j.1744-7909.2010.00905.x
Bull, C. T., Goldman, P. H., Hayes, R., Madden, L. V., Koike, S. T., and Ryder, E. (2007). Genetic diversity of lettuce for resistance to bacterial leaf spot caused by Xanthomonas campestris pv. vitians. Plant Health Prog. 10, 1094–2007. doi: 10.1094/PHP-2007-0917-02-RS
Bundó, M., and Coca, M. (2017). Calcium-dependent protein kinase OsCPK10 mediates both drought tolerance and blast disease resistance in rice plants. J. Exp. Bot. 68, 2963–2975. doi: 10.1093/jxb/erx145
Buti, M., Baldoni, E., Formentin, E., Milc, J., Frugis, G., Lo Schiavo, F., et al. (2019). A meta-analysis of comparative transcriptomic data reveals a set of key genes involved in the tolerance to abiotic stresses in rice. Int. J. Mol. Sci. 20:5662. doi: 10.3390/ijms20225662
Cassin, G., Mari, S., Curie, C., Briat, J. F., and Czernic, P. (2009). Increased sensitivity to iron deficiency in Arabidopsis thaliana overaccumulating nicotianamine. J. Exp. Bot. 60, 1249–1259. doi: 10.1093/jxb/erp007
Chaiwanon, J., Wang, W., Zhu, J. Y., Oh, E., and Wang, Z. Y. (2016). Information integration and communication in plant growth regulation. Cell 164, 1257–1268. doi: 10.1016/j.cell.2016.01.044
Charles, M. T., Goulet, A., and Arul, J. (2008). Physiological basis of UV-C induced resistance to Botrytis cinerea in tomato fruit: IV. Biochemical modification of structural barriers. Postharvest Biol. Technol. 47, 41–53. doi: 10.1016/j.postharvbio.2007.05.019
Cheng, Y., Zhang, X., Sun, T., Tian, Q., and Zhang, W. H. (2018). Glutamate receptor homolog3.4 is involved in regulation of seed germination under salt stress in Arabidopsis. Plant Cell Physiol. 59, 978–988. doi: 10.1093/pcp/pcy034
Colina, F., Carbó, M., Meijón, M., Cañal, M. J., and Valledor, L. (2020). Low UV-C stress modulates Chlamydomonas reinhardtii biomass composition and oxidative stress response through proteomic and metabolomic changes involving novel signalers and effectors. Biotechnol. Biofuels 13:110. doi: 10.1186/s13068-020-01750-8
De Storme, N., and Geelen, D. (2014). Callose homeostasis at plasmodesmata: molecular regulators and developmental relevance. Front. Plant Sci. 5:138. doi: 10.3389/fpls.2014.00138
Deng, F., Guo, T., Lefebvre, M., Scaglione, S., Antico, C. J., Jing, T., et al. (2017). Expression and regulation of ATL9, an E3 ubiquitin ligase involved in plant defense. PLoS One 12:e0188458. doi: 10.1371/journal.pone.0188458
Dey, S., and Vlot, A. C. (2015). Ethylene responsive factors in the orchestration of stress responses in monocotyledonous plants. Front. Plant Sci. 6:640. doi: 10.3389/fpls.2015.00640
Doan, T. T., Carlsson, A. S., Hamberg, M., Bülow, L., Stymne, S., and Olsson, P. (2009). Functional expression of five Arabidopsis fatty acyl-CoA reductase genes in Escherichia coli. J. Plant Physiol. 166, 787–796. doi: 10.1016/j.jplph.2008.10.003
Drerup, M. M., Schlücking, K., Hashimoto, K., Manishankar, P., Steinhorst, L., Kuchitsu, K., et al. (2013). The calcineurin B-like calcium sensors CBL1 and CBL9 together with their interacting protein kinase CIPK26 regulate the Arabidopsis NADPH oxidase RBOHF. Mol. Plant 6, 559–569. doi: 10.1093/mp/sst009
Duarte-Sierra, A., Charles, M. T., and Arul, J. (2019). “UV-C hormesis: a means of controlling diseases and delaying senescence in fresh fruits and vegetables during storage,” in Postharvest Pathology of Fresh Horticultural Produce, 1st Edn. eds L. Palou and J. Smilanick (Boca Raton, FL: CRC Press), 539–594.
Dutton, C., Hõrak, H., Hepworth, C., Mitchell, A., Ton, J., Hunt, L., et al. (2019). Bacterial infection systemically suppresses stomatal density. Plant Cell Environ. 42, 2411–2421. doi: 10.1111/pce.13570
Escudero, V., Torres, M. Á, Delgado, M., Sopeña-Torres, S., Swami, S., Morales, J., et al. (2019). Mitogen-activated protein kinase phosphatase 1 (MKP1) negatively regulates the production of reactive oxygen species during Arabidopsis immune responses. Mol. Plant Microbe Interact. 32, 464–478. doi: 10.1094/MPMI-08-18-0217-FI
Farahbakhsh, H., Pasandi Pour, A., and Reiahi, N. (2017). Physiological response of henna (Lawsonia inermise L.) to salicylic acid and salinity. Plant Prod. Sci. 20, 237–247. doi: 10.1080/1343943X.2017.1299581
Fayette, J., Jones, J. B., Pernezny, K., Roberts, P. D., and Raid, R. (2018). Survival of Xanthomonas campestris pv. vitians on lettuce in crop debris, irrigation water, and weeds in south Florida. Eur. J. Plant Pathol. 151, 341–353. doi: 10.1007/s10658-017-1377-4
Feussner, I., and Polle, A. (2015). What the transcriptome does not tell—proteomics and metabolomics are closer to the plants’ patho-phenotype. Curr. Opin. Plant Biol. 26, 26–31. doi: 10.1016/j.pbi.2015.05.023
Foyer, C. H., Rasool, B., Davey, J. W., and Hancock, R. D. (2016). Cross-tolerance to biotic and abiotic stresses in plants: a focus on resistance to aphid infestation. J. Exp. Bot. 67, 2025–2037. doi: 10.1093/jxb/erw079
Gierz, S. L., Forêt, S., and Leggat, W. (2017). Transcriptomic analysis of thermally stressed Symbiodinium reveals differential expression of stress and metabolism genes. Front. Plant Sci. 8:271. doi: 10.3389/fpls.2017.00271
Gollhofer, J., Timofeev, R., Lan, P., Schmidt, W., and Buckhout, T. J. (2014). Vacuolar-iron-transporter1-like proteins mediate iron homeostasis in Arabidopsis. PLoS One 9:e110468. doi: 10.1371/journal.pone.0110468
Han, J. Y., Kim, M. J., Ban, Y. W., Hwang, H. S., and Choi, Y. E. (2013). The involvement of β-amyrin 28-oxidase (CYP716A52v2) in oleanane-type ginsenoside biosynthesis in Panax ginseng. Plant Cell Physiol. 54, 2034–2046. doi: 10.1093/pcp/pct141
Hauser, T., Popilka, L., Hartl, F. U., and Hayer-Hartl, M. (2015). Role of auxiliary proteins in Rubisco biogenesis and function. Nat. Plants 1, 1–11 doi: 10.1038/nplants.2015.65
Health Canada (2019). Search Product Label, Pest Management Regulatory Agency. Available online at: http://pr-rp.hc-sc.gc.ca/ls-re/index-eng.php (accessed November 1, 2020).
Hofberger, J. A., Nsibo, D. L., Govers, F., Bouwmeester, K., and Schranz, M. E. (2015). A complex interplay of tandem-and whole-genome duplication drives expansion of the L-type lectin receptor kinase gene family in the Brassicaceae. Genome Biol. Evol. 7, 720–734. doi: 10.1093/gbe/evv020
Höfer, R., Dong, L., André, F., Ginglinger, J. F., Lugan, R., Gavira, C., et al. (2013). Geraniol hydroxylase and hydroxygeraniol oxidase activities of the CYP76 family of cytochrome P450 enzymes and potential for engineering the early steps of the (seco) iridoid pathway. Metab. Eng. 20, 221–232. doi: 10.1016/j.ymben.2013.08.001
Hu, W., Yan, Y., Tie, W., Ding, Z., Wu, C., Ding, X., et al. (2018). Genome-wide analyses of calcium sensors reveal their involvement in drought stress response and storage roots deterioration after harvest in cassava. Genes 9:221. doi: 10.3390/genes9040221
Huo, D., Zheng, W., Li, P., Xu, Z., Zhou, Y., Chen, M., et al. (2014). Identification, classification, and drought response of F-box gene family in foxtail millet. Acta Agron. Sin. 40, 1585–1594. doi: 10.3724/SP.J.1006.2014.01585
Jamieson, P. A., Shan, L., and He, P. (2018). Plant cell surface molecular cypher: receptor-like proteins and their roles in immunity and development. Plant Sci. 274, 242–251. doi: 10.1016/j.plantsci.2018.05.030
Janisiewicz, W. J., Takeda, F., Glenn, D. M., Camp, M. J., and Jurick, W. M. (2016a). Dark period following UV-C treatment enhances killing of Botrytis cinerea conidia and controls gray mold of strawberries. J. Phytopathol. 106, 386–394. doi: 10.1094/PHYTO-09-15-0240-R
Janisiewicz, W. J., Takeda, F., Nichols, B., Glenn, D. M., Jurick, W. M. II, and Camp, M. J. (2016b). Use of low-dose UV-C irradiation to control powdery mildew caused by Podosphaera aphanis on strawberry plants. Can. J. Plant Pathol. 38, 430–439. doi: 10.1080/07060661.2016.1263807
Kato, Y., and Sakamoto, W. (2018). FtsH protease in the thylakoid membrane: physiological functions and the regulation of protease activity. Front. Plant Sci. 9:855. doi: 10.3389/fpls.2018.00855
Kawarazaki, T., Kimura, S., Iizuka, A., Hanamata, S., Nibori, H., Michikawa et al. (2013). A low temperature-inducible protein AtSRC2 enhances the ROS-producing activity of NADPH oxidase AtRbohF. Biochim. Biophys. Acta 1833, 2775–2780. doi: 10.1016/j.bbamcr.2013.06.024
Kazemi-Pour, N., Condemine, G., and Hugouvieux-Cotte-Pattat, N. (2004). The secretome of the plant pathogenic bacterium Erwinia chrysanthemi. Proteomics 4, 3177–3186. doi: 10.1002/pmic.200300814
Kim, Y. H., Lim, S., Han, S. H., Lee, J. C., Song, W. K., Bang, J. W., et al. (2007). Differential expression of 10 sweetpotato peroxidases in response to sulfur dioxide, ozone, and ultraviolet radiation. Plant Physiol. Biochem. 45, 908–914. doi: 10.1016/j.plaphy.2007.07.019
Kimura, Y., and Tanaka, K. (2010). Regulatory mechanisms involved in the control of ubiquitin homeostasis. J. Biochem. 147, 793–798. doi: 10.1093/jb/mvq044
Konatu, F. R. B., and Jardim, I. C. S. F. (2018). Development and validation of an analytical method for multiresidue determination of pesticides in lettuce using QuEChERS–UHPLC–MS/MS. J. Sep. Sci. 41, 1726–1733. doi: 10.1002/jssc.201701038
Kumar, D., Kumar, A., Chhokar, V., Gangwar, O. P., Bhardwaj, S. C., Sivasamy, M., et al. (2020). Genome-wide association studies in diverse spring wheat panel for stripe, stem and leaf rust resistance. Front. Plant Sci. 11:748. doi: 10.3389/fpls.2020.00748
Lannoo, N., and Van Damme, E. J. (2014). Lectin domains at the frontiers of plant defense. Front. Plant Sci. 5:397. doi: 10.3389/fpls.2014.00397
Le, C. T. T., Brumbarova, T., Ivanov, R., Stoof, C., Weber, E., Mohrbacher, J., et al. (2016). Zinc finger of Arabidopsis thaliana12 (ZAT12) interacts with FER-like iron deficiency-induced transcription factor (FIT) linking iron deficiency and oxidative stress responses. J. Plant Physiol. 170, 540–557. doi: 10.1104/pp.15.01589
Li, Q., Hu, A., Qi, J., Dou, W., Qin, X., Zou, X., et al. (2020). CsWAKL08, a pathogen-induced wall-associated RLK in sweet orange, confers resistance to citrus bacterial canker via ROS control and JA signaling. Hortic. Res. 7:42. doi: 10.1038/s41438-020-0263-y
Li, Y., Chang, D., Yang, H., Wang, J., and Shi, H. (2018). Metabolome and molecular basis for carbohydrate increase and nitrate reduction in burley tobacco seedlings by glycerol through upregulating carbon and nitrogen metabolism. Sci. Rep. 8:13300. doi: 10.1038/s41598-018-31432-3
Lionetti, V., Fabri, E., De Caroli, M., Hansen, A. R., Willats, W. G., Piro, G., et al. (2017). Three pectin methylesterase inhibitors protect cell wall integrity for Arabidopsis immunity to Botrytis. Plant Physiol. 173, 1844–1863. doi: 10.1104/pp.16.01185
Livak, K. J., and Schmittgen, T. D. (2001). Analysis of relative gene expression data using real-time quantitative PCR and the 2– ΔΔCT method. Methods 25, 402–408.
Lokdarshi, A., Conner, W. C., McClintock, C., Li, T., and Roberts, D. M. (2016). Arabidopsis CML38, a calcium sensor that localizes to ribonucleoprotein complexes under hypoxia stress. J. Plant Physiol. 170, 1046–1059. doi: 10.1104/pp.15.01407
López-Fernández, M. P., and Maldonado, S. (2015). Programmed cell death in seeds of angiosperms. J. Integr. Plant Biol. 57, 996–1002. doi: 10.1111/jipb.12367
Lu, H., and Raid, R. (2013). A novel screening method for evaluation of lettuce germplasm for bacterial leaf spot resistance. Hortscience 48, 171–174. doi: 10.21273/HORTSCI.48.2.171
Lunn, J. E., Delorge, I., Figueroa, C. M., Van Dijck, P., and Stitt, M. (2014). Trehalose metabolism in plants. Plant J. 79, 544–567. doi: 10.1111/tpj.12509
Ma, X., Koepke, J., Panjikar, S., Fritzsch, G., and Stöckigt, J. (2005). Crystal structure of vinorine synthase, the first representative of the BAHD superfamily. J. Biol. Chem. 280, 13576–13583. doi: 10.1074/jbc.M414508200
Mittler, R., Vanderauwera, S., Gollery, M., and Van Breusegem, F. (2004). Reactive oxygen gene network of plants. Trends Plant Sci. 9, 490–498. doi: 10.1016/j.tplants.2004.08.009
Mondal, R., and Das, P. (2020). Data-mining bioinformatics: suggesting Arabidopsis thaliana L-type lectin receptor kinase IX. 2 (LecRK-IX. 2) modulate metabolites and abiotic stress responses. Plant Signal. Behav. 15:1818031. doi: 10.1080/15592324.2020.1818031
Moraes, R. M., Carvalho, M., Nery, F. C., Filho, P. R. S., Nogueira, M. L., and Barbosa, S. (2018). Effects of hydrogen peroxide on initial growth and enzymatic antioxidant system of Lactuca sativa L (Asteraceae). Pak. J. Bot. 50, 1769–1774.
Moses, T., Papadopoulou, K. K., and Osbourn, A. (2014). Metabolic and functional diversity of saponins, biosynthetic intermediates and semi-synthetic derivatives. Crit. Rev. Biochem. Mol. 49, 439–462. doi: 10.3109/10409238.2014.953628
Murcia, G., Fontana, A., Pontin, M., Baraldi, R., Bertazza, G., and Piccoli, P. N. (2017). ABA and GA3 regulate the synthesis of primary and secondary metabolites related to alleviation from biotic and abiotic stresses in grapevine. Phytochemistry 135, 34–52. doi: 10.1016/j.phytochem.2016.12.007
Muthusamy, S. K., Dalal, M., Chinnusamy, V., and Bansal, K. C. (2017). Genome-wide identification and analysis of biotic and abiotic stress regulation of small heat shock protein (HSP20) family genes in bread wheat. J. Plant Physiol. 211, 100–113. doi: 10.1016/j.jplph.2017.01.004
Nayeri, F. D., and Yarizade, K. (2014). Bioinformatics study of delta-12 fatty acid desaturase 2 (FAD2) gene in oilseeds. Mol. Biol. Rep. 41, 5077–5087. doi: 10.1007/s11033-014-3373-5
Nicolas, O., Charles, M. T., Chabot, D., Aarrouf, J., Jenni, S., Toussaint, V., et al. (2020). Preliminary evaluation of the impact of preharvest UV-C on lettuce: potential for the control of Xanthomonas campestris. Acta Hortic. 1271, 387–394. doi: 10.17660/ActaHortic.2020.1271.53
Nicolas, O., Charles, M. T., Jenni, S., Toussaint, V., and Beaulieu, C. (2018). Relationships between Xanthomonas campestris pv. vitians population sizes, stomatal density and lettuce resistance to bacterial leaf spot. Can. J. Plant. Pathol. 40, 399–407. doi: 10.1080/07060661.2018.1495269
Niu, L., and Wang, W. (2020). Defensive forwards: stress-responsive proteins in cell walls of crop plants. bioRxiv [Preprint]. doi: 10.1101/2020.02.15.950535v1
Noda, S., Takahashi, Y., Tsurumaki, Y., Yamamura, M., Nishikubo, N., Yamaguchi, M., et al. (2013). ATL54, a RING-H2 domain protein selected by a gene co-expression network analysis, is associated with secondary cell wall formation in Arabidopsis. Plant Biotechnol. J. 30, 169–177. doi: 10.5511/plantbiotechnology.13.0304a
Phukan, U. J., Jeena, G. S., and Shukla, R. K. (2016). WRKY transcription factors: molecular regulation and stress responses in plants. Front. Plant Sci. 7:760. doi: 10.3389/fpls.2016.00760
Proietti, S., Giangrande, C., Amoresano, A., Pucci, P., Molinaro, A., Bertini, L., et al. (2014). Xanthomonas campestris lipooligosaccharides trigger innate immunity and oxidative burst in Arabidopsis. Plant Physiol. Biochem. 85, 51–62. doi: 10.1016/j.plaphy.2014.10.011
Pushpa, D., Yogendra, K. N., Gunnaiah, R., Kushalappa, A. C., and Murphy, A. (2014). Identification of late blight resistance-related metabolites and genes in potato through nontargeted metabolomics. Plant Mol. Biol. Rep. 32, 584–595. doi: 10.1007/s11105-013-0665-1
Qiao, K., Wang, F., Liang, S., Wang, H., Hu, Z., and Chai, T. (2019). New biofortification tool: wheat TaCNR5 enhances zinc and manganese tolerance and increases zinc and manganese accumulation in rice grains. J. Agric. Food Chem. 67, 9877–9884. doi: 10.1021/acs.jafc.9b04210
Quezada, E. H., García, G. X., Arthikala, M. K., Melappa, G., Lara, M., and Nanjareddy, K. (2019). Cysteine-rich receptor-like kinase gene family identification in the Phaseolus genome and comparative analysis of their expression profiles specific to mycorrhizal and rhizobial symbiosis. Genes 10:59. doi: 10.3390/genes10010059
Ravet, K., and Pilon, M. (2013). Copper and iron homeostasis in plants: the challenges of oxidative stress. Antioxid. Redox Signal. 19, 919–932. doi: 10.1089/ars.2012.5084
Sampedro, J., Gianzo, C., Iglesias, N., Guitián, E., Revilla, G., and Zarra, I. (2012). AtBGAL10 is the main xyloglucan β-galactosidase in Arabidopsis, and its absence results in unusual xyloglucan subunits and growth defects. Plant Physiol. 158, 1146–1157. doi: 10.1104/pp.111.192195
Savidge, B., Weiss, J. D., Wong, Y. H. H., Lassner, M. W., Mitsky, T. A., Shewmaker, C. K., et al. (2002). Isolation and characterization of homogentisate phytyltransferase genes from Synechocystis sp. PCC 6803 and Arabidopsis. J. Plant Physiol. 129, 321–332. doi: 10.1104/pp.010747
Scarpeci, T. E., Zanor, M. I., Mueller-Roeber, B., and Valle, E. M. (2013). Overexpression of AtWRKY30 enhances abiotic stress tolerance during early growth stages in Arabidopsis thaliana. Plant Mol. Biol. 83, 265–277. doi: 10.1007/s11103-013-0090-8
Shaner, G., and Finney, R. E. (1977). The effect of nitrogen fertilization on the expression of slow-mildewing resistance in knox wheat. J. Phytopathol. 67, 1051–1056.
Shen, J. L., Li, C. L., Wang, M., He, L. L., Lin, M. Y., Chen, D. H., et al. (2017). Mitochondrial pyruvate carrier 1 mediates abscisic acid-regulated stomatal closure and the drought response by affecting cellular pyruvate content in Arabidopsis thaliana. BMC Plant Biol. 17:217. doi: 10.1186/s12870-017-1175-3
Sidibé, A., Charles, M. T., Nicolas, O., and Beaulieu, C. (2021). Preharvest UV-C affects lettuce resistance to Xanthomonas campestris pv. vitians and quality. Sci. Hortic. 285:110094. doi: 10.1016/j.scienta.2021.110094
Skaar, J. R., Pagan, J. K., and Pagano, M. (2013). Mechanisms and function of substrate recruitment by F-box proteins. Nat. Rev. Mol. Cell Biol. 14, 369–381. doi: 10.1038/nrm3582
Suárez, R., Wong, A., Ramírez, M., Barraza, A., Orozco, M. D. C., Cevallos, M. A., et al. (2008). Improvement of drought tolerance and grain yield in common bean by overexpressing trehalose-6-phosphate synthase in rhizobia. Mol. Plant Microbe Interact. 21, 958–966. doi: 10.1094/MPMI-21-7-0958
Sung, P. H., Huang, F. C., Do, Y. Y., and Huang, P. L. (2011). Functional expression of geraniol 10-hydroxylase reveals its dual function in the biosynthesis of terpenoid and phenylpropanoid. J. Agric. Food Chem. 59, 4637–4643. doi: 10.1021/jf200259n
Suzuki, N., Miller, G., Morales, J., Shulaev, V., Torres, M. A., and Mittler, R. (2011). Respiratory burst oxidases: the engines of ROS signaling. Curr. Opin. Plant Biol. 14, 691–699. doi: 10.1016/j.pbi.2011.07.014
Tan, S. K., and Kamada, H. (2000). Initial identification of a phosphoprotein that appears to be involved in the induction of somatic embryogenesis in carrot. Plant Cell Rep. 19, 739–747. doi: 10.1007/s002999900186
Tian, M., Lou, L., Liu, L., Yu, F., Zhao, Q., Zhang, H., et al. (2015). The RING finger E3 ligase STRF1 is involved in membrane trafficking and modulates salt-stress response in Arabidopsis thaliana. Plant J. 82, 81–92. doi: 10.1111/tpj.12797
Valencia, M. A., Hoyos, L. F. P., Ramírez, J. H., Sánchez, D. A. C., Gómez, J. A., and Quijano, J. C. (2017). Using UV-C radiation and image analysis for fungus control in tomato plants. Opt. Pura Apl. 50, 369–378.
van Engelen, F. A., Hartog, M. V., Thomas, T. L., Taylor, B., Sturm, A., van Kammen, A., et al. (1993). The carrot secreted glycoprotein gene EP1 is expressed in the epidermis and has sequence homology to Brassica S-locus glycoproteins. Plant J. 4, 855–862. doi: 10.1046/j.1365-313X.1993.04050855.x
Vanholme, R., Cesarino, I., Rataj, K., Xiao, Y., Sundin, L., Goeminne et al. (2013). Caffeoyl shikimate esterase (CSE) is an enzyme in the lignin biosynthetic pathway in Arabidopsis. Science 341, 1103–1106. doi: 10.1126/science.1241602
Vàsquez, H., Ouhibi, C., Forges, M., Lizzi, Y., Urban, L., and Aarrouf, J. (2020). Hormetic doses of UV-C light decrease the susceptibility of tomato plants to Botrytis cinerea infection. J. Phytopathol. 168, 524–532. doi: 10.1111/jph.12930
Vàsquez, H., Ouhibi, C., Lizzi, Y., Azzouz, N., Forges, M., Bardin, M., et al. (2017). Pre-harvest hormetic doses of UV-C radiation can decrease susceptibility of lettuce leaves (Lactuca sativa L.) to Botrytis cinerea L. Sci. Hortic. 222, 32–39. doi: 10.1016/j.scienta.2017.04.017
Vázquez-Hernández, M. C., Parola-Contreras, I., Montoya-Gómez, L. M., Torres-Pacheco, I., Schwarz, D., and Guevara-González, R. G. (2019). Eustressors: chemical and physical stress factors used to enhance vegetables production. Sci. Hortic. 250, 223–229. doi: 10.1016/j.scienta.2019.02.053
Wang, X. Y., Li, D. Z., Li, Q., Ma, Y. Q., Yao, J. W., Huang, X., et al. (2016). Metabolomic analysis reveals the relationship between AZI1 and sugar signaling in systemic acquired resistance of Arabidopsis. Plant Physiol. Biochem. 107, 273–287. doi: 10.1016/j.plaphy.2016.06.016
Williams, B., Kabbage, M., Britt, R., and Dickman, M. B. (2010). AtBAG7, an Arabidopsis Bcl-2–associated athanogene, resides in the endoplasmic reticulum and is involved in the unfolded protein response. Proc. Natl. Acad. Sci. U.S.A. 107, 6088–6093. doi: 10.1073/pnas.0912670107
Wormit, A., and Usadel, B. (2018). The multifaceted role of pectin methylesterase inhibitors (PMEIs). Int. J. Mol. Sci. 19:2878. doi: 10.3390/ijms19102878
Wu, C., Hu, W., Yan, Y., Tie, W., Ding, Z., Guo, J., et al. (2018). The late embryogenesis abundant protein family in cassava (Manihot esculenta Crantz): genome-wide characterization and expression during abiotic stress. Molecules 23:1196. doi: 10.3390/molecules23051196
Xie, Z., Fan, J., Charles, M. T., Charlebois, D., Khanizadeh, S., Rolland, D., et al. (2016). Preharvest ultraviolet-C irradiation: influence on physicochemical parameters associated with strawberry fruit quality. Plant Physiol. Biochem. 108, 337–343. doi: 10.1016/j.plaphy.2016.07.026
Xu, J., Xie, J., Yan, C., Zou, X., Ren, D., and Zhang, S. (2014). A chemical genetic approach demonstrates that MPK 3/MPK 6 activation and NADPH oxidase-mediated oxidative burst are two independent signaling events in plant immunity. Plant J. 77, 222–234. doi: 10.1111/tpj.12382
Xu, Y., Charles, M. T., Luo, Z., Mimee, B., Veronneau, P. Y., Rolland, D., et al. (2019). Ultraviolet-C priming of strawberry leaves against subsequent Mycosphaerella fragariae infection involves the action of reactive oxygen species, plant hormones, and terpenes. Plant Cell Environ. 42, 815–831. doi: 10.1111/pce.13491
Yan, Q., Cui, X., Lin, S., Gan, S., Xing, H., and Dou, D. (2016). GmCYP82A3, a soybean cytochrome P450 family gene involved in the jasmonic acid and ethylene signaling pathway, enhances plant resistance to biotic and abiotic stresses. PLoS One 11:e0162253. doi: 10.1371/journal.pone.0162253
Yang, Z. B., You, J. F., and Yang, Z. M. (2007). Manganese uptake and transportation as well as antioxidant response to excess manganese in plants. J. Plant Physiol. Mol. Biol. 33, 480–488.
Yeo, E. T., Kwon, H. B., Han, S. E., Lee, J. T., Ryu, J. C., and Byu, M. O. (2000). Genetic engineering of drought resistant potato plants by introduction of the trehalose-6-phosphate synthase (TPS1) gene from Saccharomyces cerevisiae. Mol. Cells 10, 263–268.
Yin, Y., Vafeados, D., Tao, Y., Yoshida, S., Asami, T., and Chory, J. (2005). A new class of transcription factors mediates brassinosteroid-regulated gene expression in Arabidopsis. Cell 120, 249–259. doi: 10.1016/j.cell.2004.11.044
Yu, K., Soares, J. M., Mandal, M. K., Wang, C., Chanda, B., Gifford, A. N., et al. (2013). A feedback regulatory loop between G3P and lipid transfer proteins DIR1 and AZI1 mediates azelaic-acid-induced systemic immunity. Cell Rep. 3, 1266–1278. doi: 10.1016/j.celrep.2013.03.030
Zhang, X., and Liu, C. J. (2015). Multifaceted regulations of gateway enzyme phenylalanine ammonia-lyase in the biosynthesis of phenylpropanoids. Mol. Plant 8, 17–27. doi: 10.1016/j.molp.2014.11.001
Zhang, H., and Sonnewald, U. (2017). Differences and commonalities of plant responses to single and combined stresses. Plant J. 90, 839–855. doi: 10.1111/tpj.13557
Zhao, C., Zayed, O., Yu, Z., Jiang, W., Zhu, P., Hsu et al. (2018). Leucine-rich repeat extesin proteins regulate plant salt tolerance in Arabidopsis. Proc. Natl. Acad. Sci. U.S.A. 115, 13123–13128. doi: 10.1073/pnas.1816991115
Zhou, J., Xia, X. J., Zhou, Y. H., Shi, K., Chen, Z., and Yu, J. Q. (2014). RBOH1-dependent H2O2 production and subsequent activation of MPK1/2 play an important role in acclimation-induced cross-tolerance in tomato. J. Exp. Bot. 65, 595–607. doi: 10.1093/jxb/ert404
Zhu, X., Yin, J., Liang, S., Liang, R., Zhou, X., Chen, Z., et al. (2016). The multivesicular bodies (MVBs)-localized AAA ATPase LRD6-6 inhibits immunity and cell death likely through regulating MVBs-mediated vesicular trafficking in rice. PLoS Genet. 12:e1006311. doi: 10.1371/journal.pgen.1006311
Keywords: bacterial disease, cell homeostasis, defense mechanisms, eustress, leaf spot disease, lettuce, physiological processes, plant growth
Citation: Sidibé A, Charles MT, Lucier J-F, Xu Y and Beaulieu C (2022) Preharvest UV-C Hormesis Induces Key Genes Associated With Homeostasis, Growth and Defense in Lettuce Inoculated With Xanthomonas campestris pv. vitians. Front. Plant Sci. 12:793989. doi: 10.3389/fpls.2021.793989
Received: 12 October 2021; Accepted: 22 December 2021;
Published: 17 January 2022.
Edited by:
Shabir Hussain Wani, Sher-e-Kashmir University of Agricultural Sciences and Technology of Kashmir, IndiaCopyright © 2022 Sidibé, Charles, Lucier, Xu and Beaulieu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Marie Thérèse Charles, bWFyaWV0aGVyZXNlLmNoYXJsZXNAYWdyLmdjLmNh