 Ahmed Ismael1,2*
Ahmed Ismael1,2* Jianming Xue3
Jianming Xue3 Dean Francis Meason1
Dean Francis Meason1 Jaroslav Klápště1
Jaroslav Klápště1 Marta Gallart4
Marta Gallart4 Yongjun Li1,5
Yongjun Li1,5 Pierre Bellè1
Pierre Bellè1 Mireia Gomez-Gallego1,6
Mireia Gomez-Gallego1,6 Ki-Taurangi Bradford1
Ki-Taurangi Bradford1 Emily Telfer1
Emily Telfer1 Heidi Dungey1
Heidi Dungey1- 1Scion (New Zealand Forest Research Institute Ltd.), Rotorua, New Zealand
- 2Research and Development, Livestock Improvement Corporation, Hamilton, New Zealand
- 3Scion (New Zealand Forest Research Institute Ltd.), Christchurch, New Zealand
- 4Centre for Planetary Health and Food Security, Griffith University, Nathan, QLD, Australia
- 5Agriculture Victoria, AgriBio Center, Bundoora, VIC, Australia
- 6INRAE, IAM, Université de Lorraine, Nancy, France
The selection of drought-tolerant genotypes is globally recognized as an effective strategy to maintain the growth and survival of commercial tree species exposed to future drought periods. New genomic selection tools that reduce the time of progeny trials are required to substitute traditional tree breeding programs. We investigated the genetic variation of water stress tolerance in New Zealand-grown Pinus radiata D. Don using 622 commercially-used genotypes from 63 families. We used quantitative pedigree-based (Genomic Best Linear Unbiased Prediction or ABLUP) and genomic-based (Genomic Best Linear Unbiased Prediction or GBLUP) approaches to examine the heritability estimates associated with water stress tolerance in P. radiata. Tree seedling growth traits, foliar carbon isotope composition (δ13C), and dark-adapted chlorophyll fluorescence (Y) were monitored before, during and after 10 months of water stress. Height growth showed a constant and moderate heritability level, while the heritability estimate for diameter growth and δ13C decreased with water stress. In contrast, chlorophyll fluorescence exhibited low heritability after 5 and 10 months of water stress. The GBLUP approach provided less breeding value accuracy than ABLUP, however, the relative selection efficiency of GBLUP was greater compared with ABLUP selection techniques. Although there was no significant relationship directly between δ13C and Y, the genetic correlations were significant and stronger for GBLUP. The positive genetic correlations between δ13C and tree biomass traits under water stress indicated that intraspecific variation in δ13C was likely driven by differences in the genotype’s photosynthetic capacity. The results show that foliar δ13C can predict P. radiata genotype tolerance to water stress using ABLUP and GBLUP approaches and that such approaches can provide a faster screening and selection of drought-tolerant genotypes for forestry breeding programs.
Introduction
Global climate change scenarios predict that large areas of planted forests will be at risk in the future due to warmer temperatures and changing precipitation patterns and drought (IPCC, 2013). Drought can cause severe negative effects on plant growth and survival (Adams et al., 2009; Allen et al., 2010), such as increased susceptibility to pathogens by reducing the amount of resources available for defense (Kolb et al., 2016) and alter the forest stand dynamics by shifting species structure and composition (Rigling et al., 2013). Increases in the intensity, duration and frequency of drought are often associated with a significant increase in tree mortality in forest ecosystems across the globe (Allen et al., 2010). In New Zealand, future scenarios predicts an increase of drought frequencies in the northern and eastern regions of the country (Mullan et al., 2005).
The New Zealand commercial forestry sector is the third-largest primary sector export earner (New Zealand Forest Owners Association [NZFOA], 2019). Planted forests of Pinus radiata D. Don comprise 90% of the commercial forest land and cover 6.4% (1.7 million ha) of New Zealand’s land area (Forest Owners Association, 2016). The increased frequency and severity of drought periods are a potential major risk to the productivity and health of New Zealand forests (Dunningham et al., 2012). Previous modeling studies have estimated that drought-induced mortality in New Zealand forests could decrease the productivity of P. radiata plantations on average by16 m3 ha–1 Watt et al. (2010) and cause an equivalent of $38 M yr–1 loss in Eastern New Zealand forests alone (Xue et al., 2012). Modeling of future climates showed that a 2°C rise in global temperatures could reduce P. radiata productivity in New Zealand by at least 10% by 2080, due to low precipitation and pathogen outbreaks (Meason and Mason, 2014). One approach to mitigate the impact of climate change and drought periods on P. radiata is the selection and deployment of genotypes less susceptible to water-limiting conditions. Drought-tolerant genotypes require less water for wood biomass production and tolerate drier environments compared to more sensitive genotypes. Previous comparative studies of water-use efficiency across P. radiata genotypes have found contradictory results; and differences in water use between genotypes range from none (Whitehead et al., 1983; Hatton and Vertessy, 1990; Walcroft et al., 1996; Li et al., 2015; Espinoza et al., 2017) to large (Bennett and Rook, 1978; Rodríguez-Gamir et al., 2019) across different experimental conditions. The selection and breeding of drought-tolerant forest tree species is being investigated worldwide (e.g., Pita et al., 2005; Roussel et al., 2009; Gaspar et al., 2013; Marguerit et al., 2014; Chakhchar et al., 2017), although the genetic basis of drought tolerance is not well understood.
Drought can be mitigated by increasing water-use efficiency (WUE) at the whole plant level. This is defined as the ratio between biomass production and water consumption. At the leaf level, intrinsic water-use efficiency (iWUE) is described as the ratio of photosynthetic CO2 assimilation and stomatal conductance (Farquhar et al., 1982, 1989; Condon et al., 2004; Pita et al., 2005). Variation in iWUE among and within tree species can be estimated by the analysis of carbon isotope composition (δ13C) in plant tissues (Condon et al., 2002; Pita et al., 2005). A strong positive correlation between iWUE and δ13C has been reported for numerous crop and tree species (e.g., Sun et al., 1996; Condon et al., 2004; Roussel et al., 2009). Measuring leaf δ13C has several conceptual and logistical advantages to screening for drought tolerance based on iWUE (i.e., A/gs). Leaf δ13C is an attractive parameter that provides a spatial and temporal measure of essential traits related to carbon gain and plant water use (e.g., photosynthesis and stomatal conductance). Plant δ13C has been often used as a screening tool in breeding programs to select genotypes with greater WUE and productivity under drought conditions (Richards and Condon, 1993; Dixon and Carter, 2019; Famula et al., 2019). A rapid, accurate and non-destructive method (i.e., high-throughput phenotyping) to identify drought-tolerant woody plants for future breeding programs is also required. A promising approach is the use of chlorophyll fluorescence for the early detection of drought stress responses (Fahlgren et al., 2015; Rapacz et al., 2019). The measurement of leaf chlorophyll fluorescence is a commonly used screening tool to estimate photosynthetic activity (Zhuang et al., 2020). Plants able to maintain photosynthetic activity under water-limiting conditions are associated with the development of abiotic-stress tolerance (Su et al., 2015).
Tree growth and survival are essential traits for tree breeding. The selection of tree species and genotypes with high WUE traits may not always increase tree productivity. For instance, negative genetic correlations between growth and δ13C sometimes leads to negative trade-offs between water use and growth. In conifers, Xu et al. (2003) reported a low to moderate positive genetic correlation between δ13C and height and diameter in Araucaria cunninghamii Mudie. Conversely, strong negative genetic correlations (−0.83 to −0.96) were found between tree height and δ13C in Pinus caribaea Morelet (Xu et al., 2000). Likewise, two studies on Pinus taeda L. found negative genetic correlations between tree height and δ13C (Baltunis et al., 2008; Cumbie et al., 2011). Therefore, the assessment of the δ13C signature in addition to growth traits and yield improve the selection of drought-tolerant crops while maintaining productivity (Cregg and Zhang, 2001; Rebetzke et al., 2002; Condon et al., 2004; Gebrekirstos et al., 2011; Sinclair, 2011). To date, very few studies have reported the use of δ13C in tree improvement programs in P. radiata.
Genomic selection (GS) refers to the prediction of unobserved phenotypes using genome-wide genetic markers, such as single nucleotide polymorphisms (SNPs). Genomic information can be constructed from pedigree-based relationships among individuals to predict breeding values, known as Best Linear Unbiased Prediction (ABLUP), or from the construction of marker-based relationship matrices and mixed linear models, known as Genomic Best Linear Unbiased Prediction (GBLUP) (Wright, 1922; Meuwissen et al., 2001; VanRaden, 2008). As opposed to traditional quantitative genetics, the application of GS in breeding programs have far-reaching implications for the selection of drought-tolerance traits to reduce the time of progeny trials and improve the accuracy of selection (Grattapaglia and Resende, 2011; Thavamanikumar et al., 2013). A previous study has provided evidence that GS could shorten P. radiata breeding cycle from 17 to 9 years in New Zealand (Li and Dungey, 2018). In the present study, we investigated the use of genomic selection to study the genetic variation in drought-tolerance traits of 622 commercially-used P. radiata genotypes from 63 P. radiata families. Specifically, we applied the pedigree-based ABLUP and the marker-based GBLUP approaches to study the following objectives: (1) to determine the genetic variation (variance components and heritability estimates) of growth traits, δ13C and Y after exposure to water stress, (2) to estimate the genetic correlations of growth traits, δ13C and Y after exposure to water stress, and (3) to compare the accuracy of ABLUP and GBLUP breeding values and evaluate the relative selection efficiency of GBLUP over ABLUP in the future genomic selection of P. radiata in New Zealand.
Materials and Methods
Pinus radiata Genotypes and Measurement of Plant Traits
Plant material was sourced from a cloned elite population from the New Zealand Radiata Pine Breeding Company (RPBC). We selected a representative population of the third generation of the New Zealand P. radiata breeding program originated from the Elite germplasm collection (Dungey et al., 2009). The study included 622 genotypes from 63 families (10 genotypes per family) resulting from crossing among 55 parents, with an average of 30 clones per family. The P. radiata genotypes selected for this study displayed a wide range of variation in stem growth rate, wood density and pathogen resistance. Three ramets per genotype of similar height and basal diameter were used, with a total of 1,866 plants included in the study. Genotypes were vegetatively propagated from stool beds as bare-rooted cuttings. In August 2015, trees were transferred to 1-L pots with the industry-standard potting mix at 70% water holding capacity (WHC) and acclimated to the polyhouse conditions for over 9 months. The polyhouse was located at Scion’s nursery in Rotorua, New Zealand (Latitude −38°09′28.8′′S, Longitude −176°16′03.3E), and provided natural light but no temperature or humidity control. Plants in the polyhouse were not exposed to precipitation and thus plants were irrigated. The climate in the trial location is temperate, with the warmest months being in summer (January to March), and the coolest months being in winter (June to August). The mean daily maximum temperature is 27°C (summer), while the mean daily minimum temperature is 5°C (winter) (Chappell, 2014).
Pots were positioned within the polyhouse in an incomplete block design. Irrigation was provided by manual watering of pots twice a week, with the irrigation adjusted to maintain potting medium at 70% WHC before the water stress was applied. The experiment was conducted on three biological replicates of each genotype of similar height and basal diameter. Before the commencement of water stress (May 2016), the basal diameter (BDI) and height (HI) of each plant were measured. In addition, four fully developed expanded fascicles were collected from under the flushed buds of each ramet. Fresh needles were oven-dried at 65°C and cryo-ground before analyzing carbon isotopic composition (δ13CI) using continuous-flow isotope ratio mass spectrometry (Europa Scientific Ltd., Crewe, United Kingdom) in the Stable Isotope Unit at the University of Waikato (Hamilton, New Zealand). The δ13C values (‰) were expressed relative to the Vienna Peedee Belemnite standard (Craig, 1957):
where Rsa and Rst are the 13C/12C ratios of the sample and the standard, respectively.
In early May 2016, three ramets per genotype of similar height and basal diameter were exposed to drought stress at 22–25% WHC. The amount of water required to reach 22–25% WHC was estimated based on the pot weight (including plant and medium). Over the course of the experiment, changes in pot weight due to tree growth were corrected using a linear relationship between pot weight and tree height. The water stress treatment was maintained by calculating the average weight change of 20–30 pots and compensating water loss every week for 11 months (late March 2017). After 6 months (November 2016) and 10 months (March 2017) of water stress, basal diameter (BD6 and BD10) and height (H6 and H10) of each plant were measured. An additional δ13C analysis in leaves was performed 10 months after water stress (March 2017, δ13C10). The maximum quantum yield of Photosystem II (PSII) in the dark-adapted state (Y) was measured after 5 months (October 2016, Y5) and 10 months (March 2017, Y10) of water stress using a pulse-amplitude modulated Portable Chlorophyll Fluorometer (Mini-PAM Photosynthesis Yield Analyzer, Heinz-Walz GmbH, Effeltrich, Germany). For each measurement, three ramets per genotype were assessed over 3 days to reduce the effect of the diurnal cycle. On the first day, maximum (Fm) and minimum (Fo) chlorophyll fluorescence emissions of dark-adapted needles were measured randomly in all ramets. During the two consecutive days, Fm and Fo measurements were repeated in a different order. The Y values for each plant were calculated as (Fm − Fo)/Fm and averaged across 3-day measurements. For each ramet, three Dark Leaf Clips (DLC-8) were simultaneously placed on mature, fully elongated needles. Needles were arranged flat across the opening of the leaf clip and the sliding shutter close 20 to 60 min before the measurement to ensure stable readings. Preliminary testing on a plant subset showed that diurnal variation was not significantly influencing the repeated Y measurements within the 3 days.
All replicate plants were carefully dissected after the 11 months of water stress into shoot and root samples. The potting medium was removed gently from the roots, and the root systems were then rinsed in running tap water. Biomass fractions, including shoot dry weight (SDW) and root dry weight (RDW) as well as total biomass or total dry weight (TDW), were determined after oven-drying the samples at 65°C for at least 72 h.
Genomic Data
The genomic information for the 622 genotypes was obtained using the exome-capture genotyping by sequencing (GBS) method (Neves et al., 2013). Further details of SNP discovery and capture probe design and testing were previously described in Telfer et al. (2018, 2019). The total number of single nucleotide polymorphisms (SNPs) markers genotyped was 1,371,123. Markers with minor allele frequency (MAF) < 0.01 and missing marker data across genotypes > 40% were discarded from the analysis. The final number of SNP markers used in this study was 61,418. The individual missing SNP genotypes were substituted with the mean genotype for that SNP as implemented in rrBLUP-R package (Endelman, 2011).
Statistical Models Used in the Genetic Analysis
Genetic analysis was computed with the Average Information Restricted Maximum Likelihood (REML) algorithm in the ASReml-R v.3 R package (Butler et al., 2009). Single-trait analysis was performed to estimate variance components and heritability for each trait separately, whereas bivariate analysis was performed to estimate genetic correlations among traits. All genetic parameters were estimated and compared using a pedigree-based model best linear unbiased prediction (ABLUP) and genomic-based unbiased prediction (GBLUP).
Preliminary analyses were used for all traits to compare the performance of a spatial first-order autoregressive mixed model and a mixed model without the autoregressive term. The preliminary analyses showed that the spatial mixed model performed better than the mixed model without the autoregressive term (non-spatial) based on Akaike’s information criterion (AIC). Therefore, only spatial mixed models were used in the current study.
The single-trait pedigree-based analysis for all traits was performed using the following model:
where y is the vector of individual tree observations; β is the vector of fixed effects containing the overall mean; a is the random additive genetic effect which was assumed to be normally distributed ∼N, where is the additive genetic variance and A is the average numerator relationship matrix; pe is the vector of the random non-additive genetic effect of the individual ramet within the genetic material ∼N, where is non-additive genetic variance and I is the identity matrix; b is the vector of random incomplete block effect ∼N, where are the incomplete block variance and I is identity matrix; X and Z are incidence matrices of the fixed and random effects of the vector of phenotypes y; e is the random residual deviations of individual trees which was partitioned into a spatial (ξ) and independent (η) residuals as [AR1 (pcol) ⊗ AR1(prow)] + I σ2 where is the spatial residual variance, ⊗ is the Kronecker product, and AR1(p) represents a first-order autoregressive correlation matrix for rows and columns where ρ is the autocorrelation parameter. The independent residual (η) was assumed to be pairwise independent as Iσ2, where I is the identity matrix, σ2 is uncorrelated residual variance.
A bivariate analysis was employed to estimate the pairwise genetic correlations between each pair of traits. The variance-covariance structure for this model is:
where icb1 and icb2 represent the incomplete block effects for 1st and 2nd trait; and incomplete block variances for the 1st and 2nd trait, a1 and a2 represent the breeding values for the 1st and 2nd trait; and are the additive genetic variances for the 1st and 2nd trait; σa1a2 is the additive genetic covariance among traits, pe1 and pe2 represent the non-additive genetic effects for the 1st and 2nd trait; and are the non-additive genetic variances for the 1st and 2nd trait, e1 and e2 represent the spatially independent residuals; and are the independent residual variances for the 1st and 2nd trait and; σe1e2 is the residual covariance among traits, and I is the identity matrix. The genetic correlations between traits (ra) were estimated as
The GBLUP model was the same as the ABLUP model, substituting the average numerator relationship matrix A by the marker-based relationship matrix G implemented based on (Forni et al., 2011) as follows:
Where M is the marker matrix with genotypes coded 0, 1, and 2 for the alternative allele homozygote, heterozygote and reference allele homozygote, respectively, and P is a vector of twice the allele frequency, trace[(M−P)(M−P)′]is a trace of the matrix defined in the nominator and n is the number of markers (Forni et al., 2011).
The individual narrow-sense heritability (h2) is the proportion of phenotypic variance that is explained by the additive genetic variance (Falconer and Mackay, 1996) and was estimated as:
where is the additive genetic variance, is non-additive genetic variance, and is the spatially independent residual variance. The two heritability estimates are reported in this study regarding implemented methods: , a narrow-sense heritability using variance components estimated in ABLUP and , a narrow-sense heritability using variance components estimated in GBLUP.
Cross-Validation and Accuracies of Estimated Breeding Values
The cross-validation was conducted by examining the accuracy of breeding values obtained by the ABLUP (ABLUP-EBV) and GBLUP (GBLUP-EBV) models. The accuracy of both models was evaluated using 10-fold cross-validation with ten random replicates. All data were randomly divided into ten equal groups. In each replication, phenotypes from one group were masked and used as a validation dataset. The accuracies of breeding values obtained by estimating the average Pearson’s correlation between predicted breeding values from masked phenotypes and the breeding values estimated using the full dataset obtained from the ten replicates. These breeding values were obtained from the single-trait analysis (Eq. 2).
Selection Response With Genomic Selection
Response to selection was calculated as the ratio between selection accuracy and breeding cycle length in years (Chen et al., 2018). Thus relative efficiency (RE) of GBLUP over ABLUP is:
Therefore, the relative efficiency of GBLUP over ABLUP per year is:
Where GBLUP-EBVV is the genomic breeding values for the trees in the validation dataset, ABLUP-EBVF is the estimated breeding values from the full dataset, ABLUP-EBVV is the estimated breeding values for the trees in the validation dataset, LTS is the length of the breeding cycle for selection based on ABLUP which is approximately 17 years, and LGS is the length of the breeding cycle for selection based on GBLUP, which is approximately 9 years (Li and Dungey, 2018).
The effective population size Ne (Wright, 1931, 1933) in the current study was 23 and was estimated based on the status number concept (Ns) of Lindgren et al. (1997) as:
where Ns is the status effective number and f is the average coancestry of the population including the coancestry of individuals with themselves.
Genotypic Variation in δ13C, Y, and Growth
One-way analyses of variance (ANOVAs) were conducted, after confirming data met assumptions for normality and homogeneous variance, to test whether means for drought-tolerant traits (i.e., δ13C and Y) differed between tree families and genotypes. A correlation was also assessed to test for the relationship between δ13C and Y after 10 months of water stress.
Results
Descriptive Statistics
Trees showed an average 39% increase in the growth of basal diameter and a 63% increase in height growth after 10 months of water stress. The mean δ13C value before water stress was −29.67‰, with a range of −26.16‰, to −32.29‰, and a standard deviation of 1.29‰ (Table 1). After 10 months of water stress, the mean δ13C value 26.50‰ with a range from −23.27‰ to −31.53‰ (Table 1). The mean Y-value for month five (Y5) was 0.79 with a range of 0.58–0.87 (Table 1). The mean Y-value at month 10 (Y10) was 0.77 with a range from 0.30 to 0.84 (Table 1).
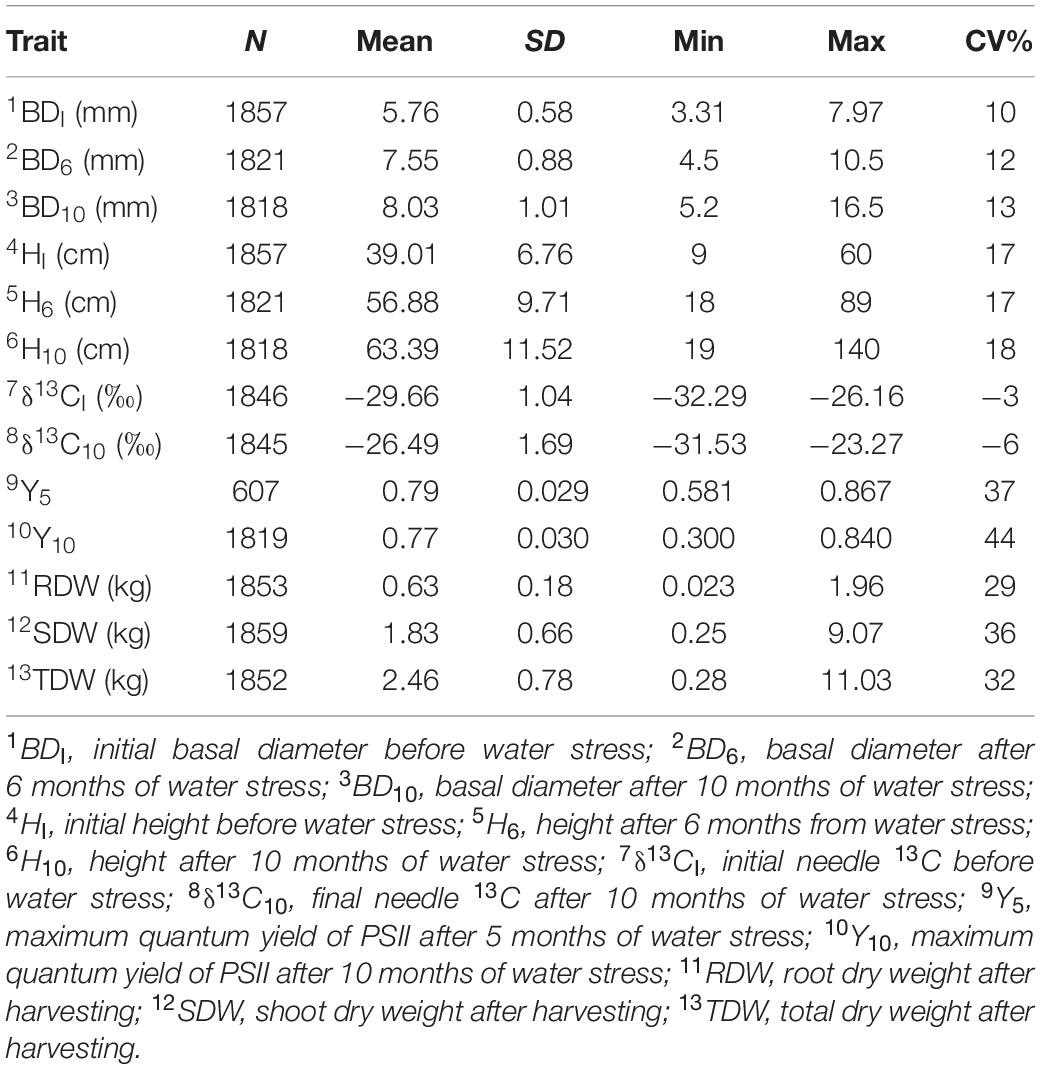
Table 1. Number of observations (N), mean standard deviation (SD), minimum (Min), maximum (Max), and coefficient of variation (CV%) for the measured growth and drought-tolerance traits before, during and after water stress in Pinus radiata genotypes.
Genetic Parameters
The measured traits showed a wide range of heritability estimates based on either ABLUP () and GBLUP ( approaches, ranging from 0.07 to 0.44 (Table 2). Using the ABLUP, for BD decreased from 0.17 (BDI) to 0.12 (BD10). In contrast, water stress increased for height from 0.38 (HI) to 0.44 (H10). The for δ13CI was 0.17 compared with 0.07 for δ13C10. The of chlorophyll fluorescence after 5 months of water stress (Y5) was close to zero due to the small number of observations recorded for this trait (N = 607). However, Y10 was 0.06. The for dry weight measures (i.e., SDW, RDW, and TDW) were similar of approximately 0.13.
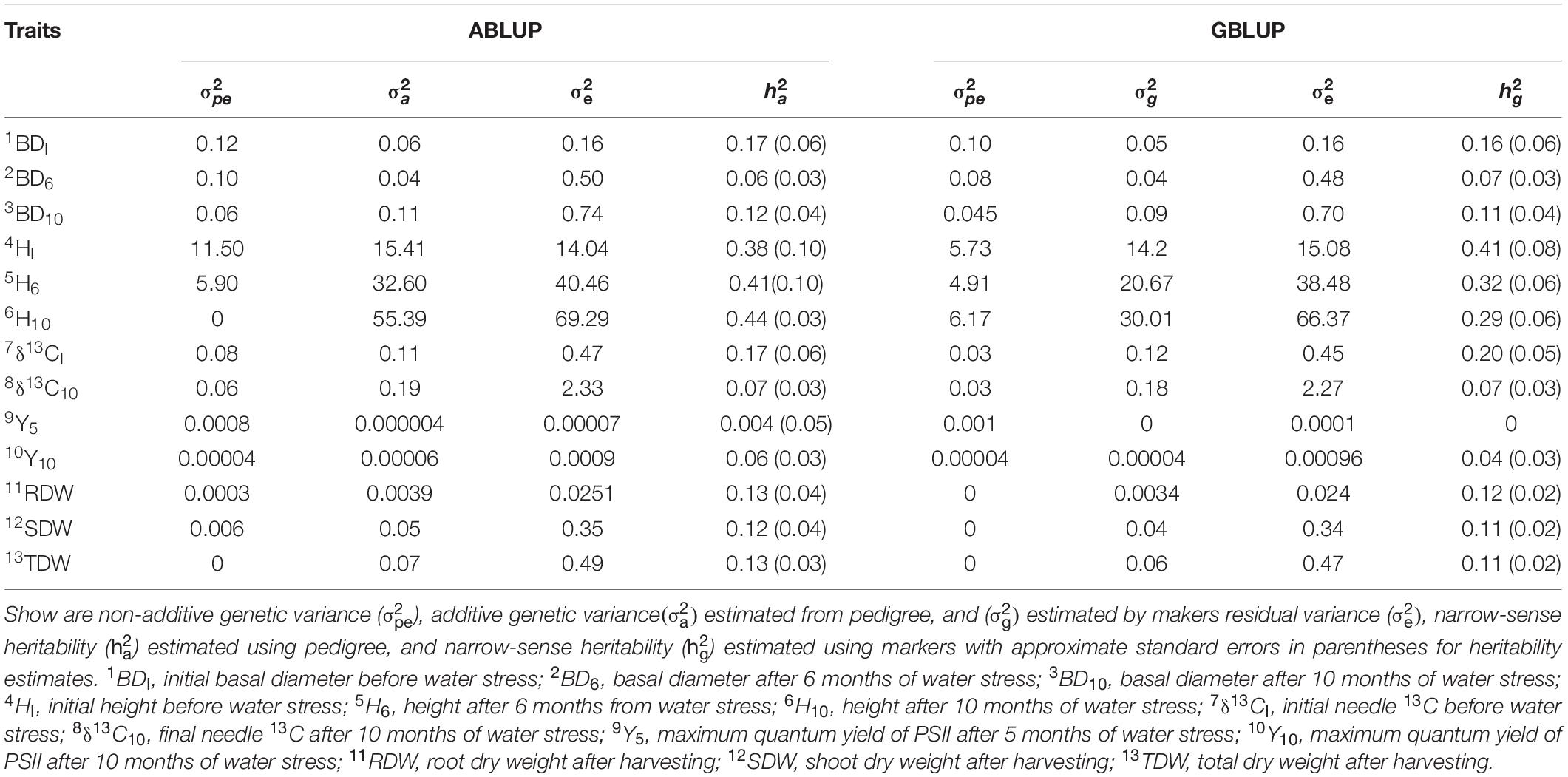
Table 2. Estimates of variance components and heritability for growth traits, carbon isotope composition and chlorophyll fluorescence.
Heritability estimates based on GBLUP were generally lower than the ABLUP estimates except for δ13CI and HI, which showed slightly higher values than . The greatest difference in after water stress was recorded for tree height, in which the estimate decreased from 0.41 (HI) to 0.29 (H10).
Genetic Correlations Between Traits
The genetic correlation between BDI and BD10 was very strong (Figure 1). Similarly, a strong genetic correlation was found between HI and H10 (r = 0.92). There were a high genetic correlations with H and BD at the beginning and end of the experiment. Genetic correlations between H and BD were moderate to large and ranged from 0.48 to 0.90. Moderate to high genetic correlations were found between biomass and tree growth traits, ranging from 0.54 to 0.99. The genetic correlation between δ13CI and δ13C10 was 0.73. In addition, genetic correlations between δ13C and growth traits were generally positive, with the strongest correlation found between δ13CI and BDI (r = 0.79). Moderate genetic correlations were observed between δ13C and biomass traits, ranging from r = 0.45 to 0.60 regardless of the point in time. The genetic correlation between Y5 and Y10 was strong (r = 0.90). Furthermore, strong and negative genetic correlations were observed between Y5 and BDI (r = −0.92), and between Y5 and HI (r = −0.88). Genetic correlations between Y10 and growth traits, and δ13C were moderate, ranging from r = −0.41 to −0.64. Genetic correlations between traits based on GBLUP showed a similar pattern but were of weaker strength compared with genetic correlations based on ABLUP (Figure 2).
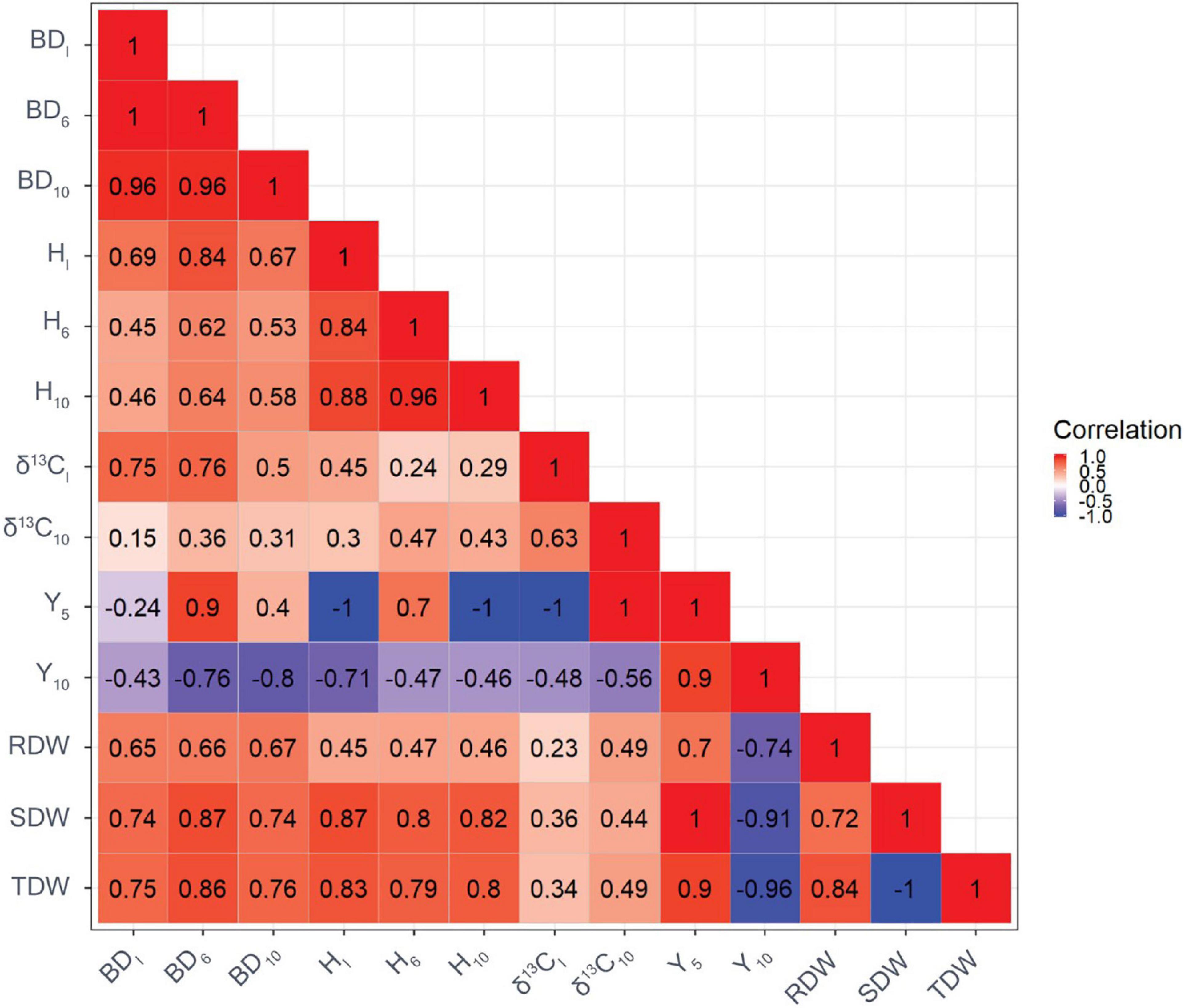
Figure 1. Genetic correlations among growth traits and carbon isotope composition δ13C based on ABLUP (the reddish the color, the greater value of genetic correlation). BDI is the initial basal diameter before water stress, BD6 is basal diameter after 6 months of water stress, BD10 is basal diameter after 10 months of water stress, HI is initial height before water stress, H6 is height after 6 months of water stress, H10 is height after 10 months of water stress, δ13CI is initial needle δ 13C before water stress, δ13C10 is final needle δ13C after 10 months of water stress, Y5 is maximum quantum yield of PSII after 5 months of water stress, Y10 is maximum quantum yield of PSII after 10 months of water stress, RDW is the root dry weight after harvesting, SDW is shoot dry weight after harvesting, and TDW is total dry weight after harvesting.
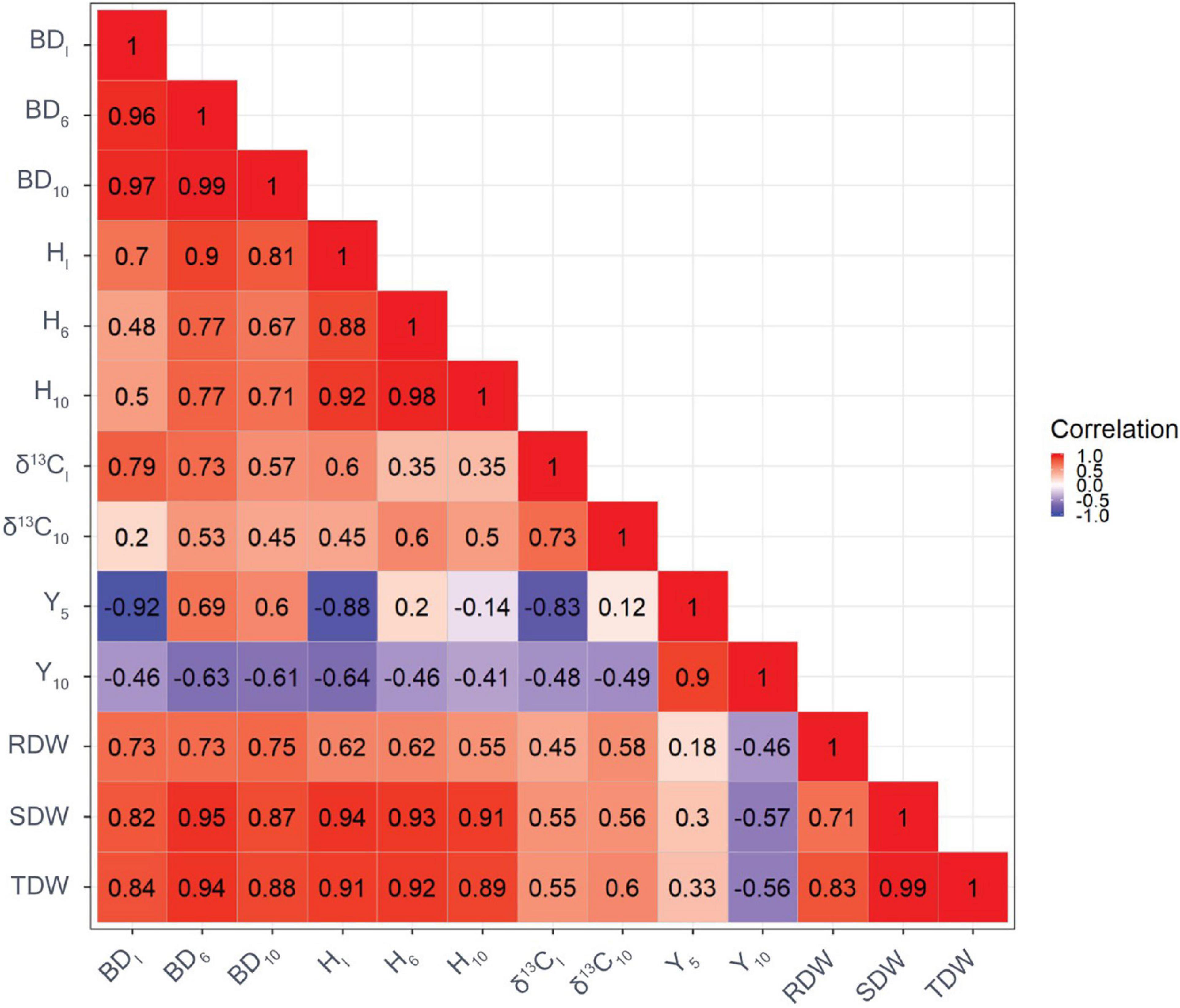
Figure 2. Genetic correlations among growth traits, carbon isotope composition δ13C based on GBLUP (the reddish the color, the greater value of genetic correlation). BDI is the initial basal diameter before water stress, BD6 is basal diameter after 6 months of water stress, BD10 is basal diameter after 10 months of water stress, HI is initial height before water stress, H6 is height after 6 months of water stress, H10 is height after 10 months of water stress, δ13CI is initial needle δ13C before water stress, δ13C10 is final needle δ13C after 10 months of water stress, Y5 is maximum quantum yield of PSII after 5 months of water stress, Y10 is maximum quantum yield of PSII after 10 months of water stress, RDW is the root dry weight after harvesting, SDW is shoot dry weight after harvesting, and TDW is total dry weight after harvesting.
Cross-Validation and Accuracies of Estimated Breeding Values
The cross-validation estimated moderate to large accuracies of breeding values for all traits using all models (Table 3). The accuracy ABLUP-EBV for diameter ranged from 0.77 for BDI to 0.83 for BD6, and height ranged from 0.67 for H10 to 0.70 for both HI, and H6. The accuracy for biomass traits reached 0.80. The accuracies for carbon isotope δ13C were 0.80 for δ13C10 and 0.78 for δ13CI, and for Y10 was 0.79. The accuracy of GBLUP-EBV was 7–26% lower than ABLUP-EBV. The accuracy of GBLUP-EBV for diameter ranged from 0.61 for BDI to 0.65 for BD10, and height ranged from 0.59 for H10 to 0.65 for H6. The accuracy for biomass traits ranged from 0.66 for RDW to 0.69 for TDW. The accuracies for carbon isotope δ13C traits were 0.66 for δ13CI and δ13C10, and for Y10 was 0.52.
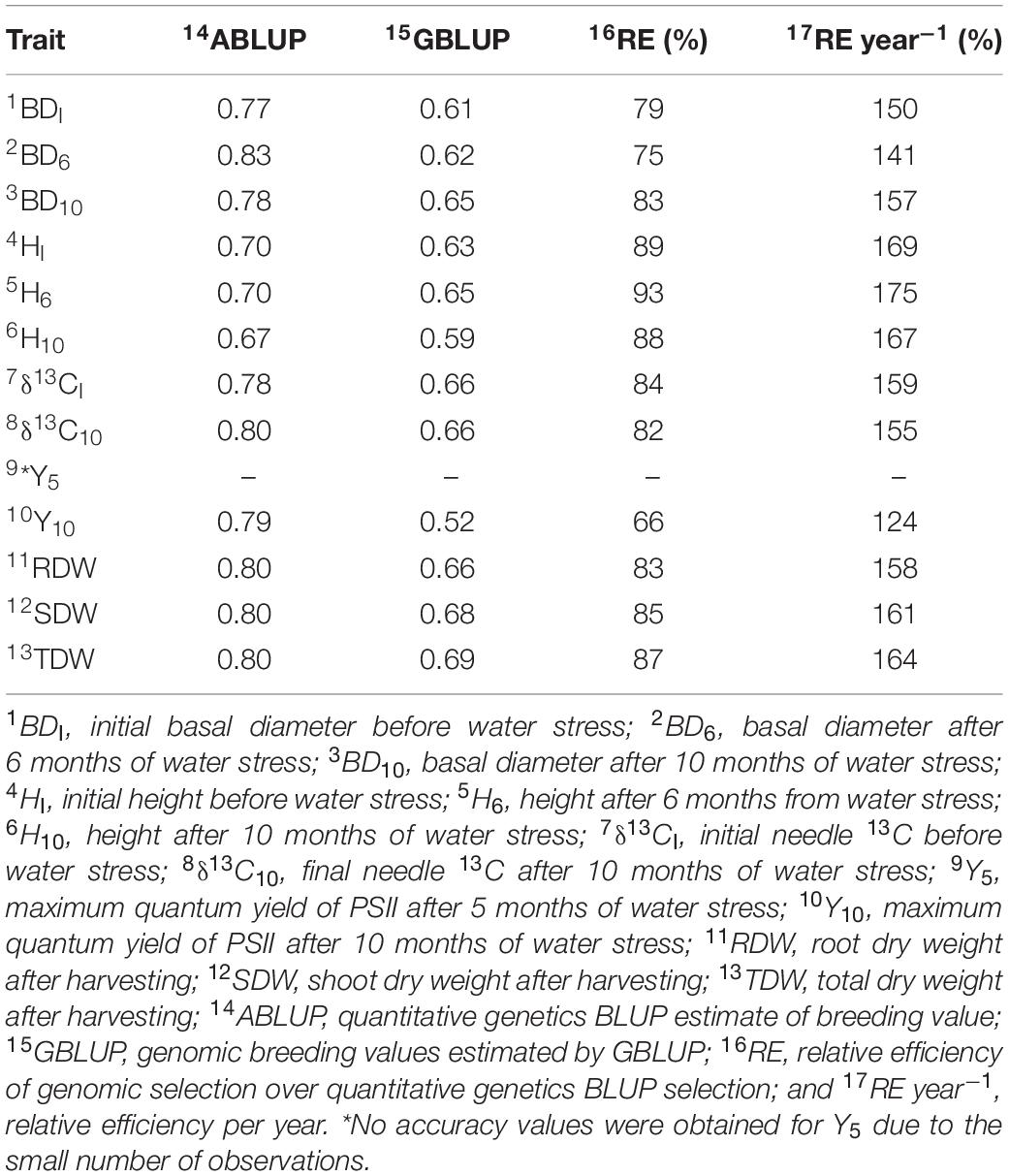
Table 3. Accuracy of breeding values from single trait pedigree-based model, single trait genomic based model (10-fold cross-validation), relative efficiency of genomic selection (GBLUP) over quantitative genetic (ABULP) selection, and the relative efficiency per year.
The RE % of the models showed increased efficiency of GBLUP over ABLUP models (Table 3). For diameter traits, GBLUP showed an increased efficiency that ranged from 75% for BD6 to 83% for BD10. For height traits, the efficiency ranged from 88% for H10 to 93% for H6. For carbon isotope δ13C traits, the efficiency was 84% for δ13CI and 82% for δ13C10, and for biomass traits, the efficiency ranged from 83% for RDW to 87% for TDW. The lowest relative selection efficiency using GBLUP was obtained for Y10 (66%). On the other hand, the relative efficiency in selection per year (RE year−1) resulted in the overall superiority of GBLUP over ABLUP. When assuming the reduction of time for progeny trials from 17 to 9 years by the genomic selection, the lowest relative selection efficiency per year using GBLUP was obtained for Y10.
Relationship Between δ13C and Y in Response to Water Stress
Significant differences in δ13CI were only observed among families (p < 0.001), but not among genotypes (p = 0.193). In contrast, differences in δ13C10 were significantly influenced by both family (p = 0.002) and genotype (p = 0.005). On the other hand, Y5 differed among families and genotypes (p < 0.001 and p < 0.001, respectively), as well as Y10 (p < 0.001 and p < 0.001, respectively). Nonetheless, no significant linear relationship was found between δ13C10 and Y10 (p > 0.05) by family (Figure 3A) or genotype (Figure 3B).
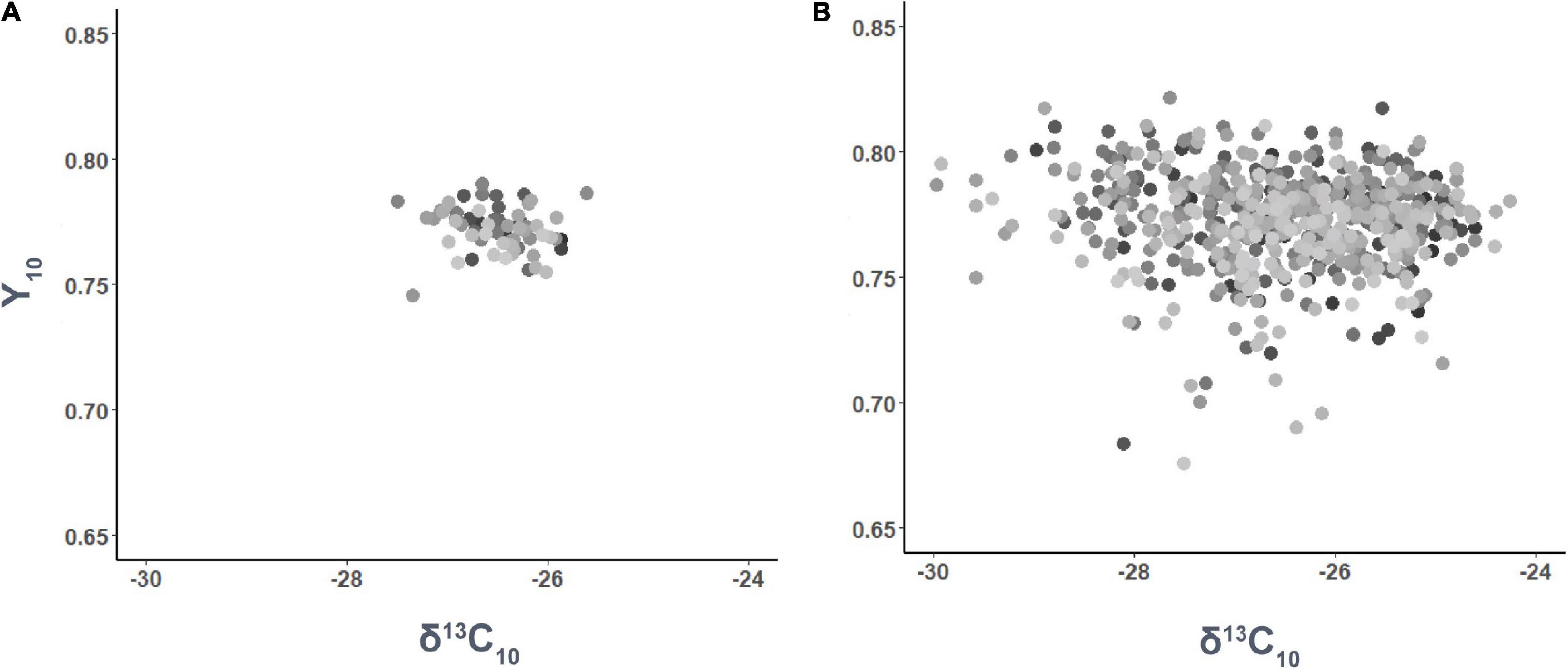
Figure 3. Phenotypic relationship between needle carbon isotope composition (δ13C10) and maximum quantum yield of PSII (Y10) after 10 months of water stress by family (A) and genotype (B).
Discussion
Genetic Parameters for Growth Traits and Foliar Carbon Isotope Composition
In this study, growth traits such as tree diameter and height presented moderate heritability after water stress. In case of tree diameter, heritability estimates decreased with time after water stress (0.17–0.12). This decrease was mainly due to the increasing residual variance from BDI to BD10, even though the additive variance increased during the experiment. In contrast, heritability for height increased with time after water stress (0.38–0.44), likely due to increasing the additive genetic variance in balance with increasing residual variance. The observed heritability estimates for BDI and HI were in agreement with previous in situ field genetic studies in P. radiata across Australia and New Zealand sites (Kumar et al., 2008; Gapare et al., 2012), which reported heritability estimates between 0.10 and 0.48 for tree diameter, and 0.34 and 0.55 for height. Another study using two populations of water-stressed P. radiata seedlings under glasshouse conditions reported similar initial height estimates but lower initial diameter estimates (Espinoza et al., 2014).
We observed that average δ13C increased after water stress and the magnitude of the shift is similar to previous studies with water-stressed P. radiata (Walcroft et al., 1997; Korol et al., 1999; Barbour et al., 2002; Waghorn et al., 2015). The increase in δ13C under water stress had overall low to moderate effect on heritability estimates of growth traits before and after water stress, however, the heritability estimate for δ13CI was twice than that for δ13C10. This is presumably due to the greater residual variance obtained for δ13C10 which was close to fivefold the residual variance for δ13CI. To our knowledge, no published studies investigated estimates for the δ13C in genotypes with narrow-sense heritability, such as P. radiata in New Zealand. In other conifers, low to moderate heritability estimates for δ13C have been reported; for instance 0.17 for P. pinaster (Brendel et al., 2002) and 0.09 for P. taeda (Baltunis et al., 2008). The decrease in δ13C heritability after the application of water stress has been observed in other studies across different species. Xu et al. (2003) found a reduction of between 0.33 and 0.20 in A. cunninghamii growing in wet sites compared to dry sites. Similarly, Ehdaie and Waines (1994) reported a lower δ13C heritability estimate in wheat-growing under drought conditions (0.12) compared to wet conditions (0.57). The authors justified this decrease due to the reduced additive genetic variance estimated under water stress conditions. Additionally, Klisz et al. (2019) found loss of any genetic signal in productivity among different Norway spruce provenances when these were growing at marginal climate conditions. Although it is difficult to make direct heritability comparisons across plant species, the results from these and other studies suggest that the induction of moderate to severe water stress may result in the loss of genetic variance in traits.
The estimates of additive genetic variance and narrow-sense heritability showed greater precision when GBLUP was implemented compared to ABLUP. Such a trend was also found in previous studies across different tree species (Gamal El-Dien et al., 2015, 2016; Chen et al., 2018). In summary, when using the marker-based genomic approach, heritability estimates decreased in most of the measured traits compared with the pedigree-based approach. These differences between the two types of correlations may be due to the fundamental differences between genomic and pedigree relationship matrices. For instance, the relationships in the pedigree-based relationship matrix are based on identity by descent (the probability that 2 alleles come from a common ancestor), whereas, the relationships in the genomic matrix reflect the identity by state probabilities (the probability that 2 alleles are the same) (Powell et al., 2010). Furthermore, differences between pedigree- and genomic-based correlations could also be driven by the source of information used. The former uses expected genetic covariation while the latter uses genetic covariation captured by SNPs (Veerkamp et al., 2011; Momen et al., 2017). Another explanation could be that unknown environmental effects were confounding the pedigree relationship-matrix and inflated the genetic variance (Lee et al., 2010).
Genetic Correlations Between Plant Growth Traits and Water-Use Efficiency Measured as δ13C
This study recorded positive genetic correlations among δ13C, chlorophyll fluorescence, basal diameter and height growth, and biomass. The moderate to large genetic correlations between δ13C and growth traits, including tree height, diameter and biomass, suggest that fast-growing genotypes under water stress could display greater WUE. However, further studies of genetic correlations between δ13C and growth traits in P. radiata are warranted to confirm these results.
The findings of this study confirm previous studies with other species that found moderate to strong genetic correlations (Johnsen et al., 1999; Xu et al., 2000; Baltunis et al., 2008). The moderate genetic correlation obtained between the intital diameter or dimater measured after 6 months with initial δ13C suggests that diameter might be an indicator trait for greater WUE of P. radiata breeding populations. The positive genetic correlation between growth and δ13CI or δ13C10 provides evidence that the variation in δ13C (a surrogate of WUE) between genotypes may be possibly due to the differences in the carboxylation efficiency of photosynthesis, rather than changes in stomatal conductance. Further, more drought-tolerant genotypes (i.e., more negative δ13C values) may be less sensitive to water stress and able to maintain photosynthetic capacity required for growth with longer periods of open stomata compared with less drought-tolerant genotypes. Rodríguez-Gamir et al. (2019) found that more water-efficient genotypes may be able to maintain a higher whole-plant hydraulic conductance under water stress, thus maintaining leaf water potential and able to keep the stomata opened for longer for photosynthesis. The same study also found that the down-regulation of root aquaporins is an important factor driving intraspecific changes in hydraulic conductance on P. radiata under water stress. Katul et al. (2003) and Rodriguez-Gamir et al. (2020) hypothesized that an equilibrium exists between maximum carbon demand by photosynthesis and maximum water supply, which might result in constant long-term mean intercellular CO2 concentration. This may be due to the co-evolution of hydraulic and biochemical plant attributes at the genotype level.
The genetic correlations between diameter or height before and after water stress were strong (0.88–0.99), indicating that genotypes were ranked consistently before and after imposing water stress. These strong genetic correlations indicate that the level of water stress in the current experiment was not strong enough to change the ranking of genotypes. This may be due to the experiment not applying severe enough water stress. Indeed, it is possible that genotype rankings could have changed using lower WHC, which would place the genotypes under more severe water stress deficit. Another theory is that only a few genotypes were sensitive to the water stress. However, results indicate that the best performing genotypes for growth traits will remain the best under water stress conditions, which is important for P. radiata breeding programs.
Negative genetic correlations were obtained in this study between δ13C and Y. That is, genotypes that performed better under water stress presented more negative foliar δ13C values and larger Y values. Physiologically, these results suggest that more drought-tolerant genotypes are more efficient at photosynthesising and have more intact chloroplasts and chlorophyll (i.e., Photosystem II) under water stress, as suggested by recent studies in P. radiata genotypes (Rodríguez-Gamir et al., 2019; Rodriguez-Gamir et al., 2020). Further, Gallart et al. (2019) suggested that the superior growth performance of some P. radiata genotypes under dry field conditions could be due to genotype-specific responses to water stress. However, future research is required to test whether greater drought tolerance also involves greater photosynthetic rates over a wide range of genotypes (Maxwell and Johnson, 2000), to determine the strength of the relationship between Y and δ13C.
We did not observe a significant linear relationship between δ13C and Y traits. Although previous studies reported δ13C to be negatively associated with Y (Mena-Petite et al., 2000, 2005; Dias and Brüggemann, 2010), Y was a poor predictor of water stress tolerance across the 63 families we tested. The poor predictability of Y in our study could be due to several reasons. First, plants were under moderate rather than severe water stress. Unlike other studies, this study maintained the potting medium to a constant 22–25% WHC. The strong relationship between δ13C and Y determined by Mena-Petite et al. (2005) was obtained from Y measurements collected over a wide range of foliar relative water potential (∼25–85% range) after withholding irrigation over a short-term period. However, a small number of Y values in this study were below 0.70. Interestingly, our study showed a range of relative water content (%) in needles between 70 and 90% (data not shown), which suggests that the application of more severe and/or more prolonged water stress treatments could have led to stronger Y differences among genotypes. Second, Y is a measure of maximum potential quantum yield of chloroplasts under Photosystem II, which may differ of the actual achievable quantum yield and photosynthesis (e.g., electron transfer rate) under ambient light and water stress conditions (e.g., Subrahmanyam et al., 2006; Baker, 2008; Wu et al., 2008). Although the use of light-adapted chlorophyll fluorescence may be more appropriate to measure quantum yield, the time required to conduct the measurements was considered unsuitable for this study.
Cross-Validation and Accuracies of Estimated Breeding Values
The accuracies of GBLUP-EBV for growth traits under the conditions used in this experiment were consistent or greater than the prediction accuracies obtained for growth traits in other tree species, such as Picea glauca (Moench) Voss (Beaulieu et al., 2014; Gamal El-Dien et al., 2016), P. pinaster (Bartholomé et al., 2016; Isik et al., 2016); and P. tadea (Resende et al., 2012). Accuracies of δ13CI and δ13C10 could not be compared with literature because this is the first study that evaluates the accuracy of genomic prediction for WUE assessed by δ13C for P. radiata. Although the accuracy of estimated GBLUP-EBV was lower than the ABLUP-EBV, the relative efficiency of selection per unit of time (RE year–1) was greater, reaching half of the breeding cycle (Isik, 2014; Li and Dungey, 2018). This is consistent with the previous studies for height and diameter in P. taeda and Picea abies Karst, in which the relative efficiency of genomic selection per unit of time ranged from 53 to 181% (Resende et al., 2012; Chen et al., 2018).
In the current study, we used the status number concept (Ns) to estimate the effective population size (Ne). Tambarussi et al., 2019 found that the status number is equivalent to the effective population size only when the size of the population is constant over generations.
Conclusion
We’ve demonstrated large genetic variation of P. radiata drought tolerance assessed by plant growth and needle δ13C (a surrogate trait for WUE). The needle δ13C had low to moderate heritability estimates, however, there were moderate to high genetic correlations with plant growth or biomass traits. Thus, genotypes of P. radiata with a narrow-genetic background can have significant physiological differences related to water-use efficiency traits. Our results suggest that the variation of needle δ13C could be primarily controlled by the photosynthetic capacity and photosynthesis, being the main process driving tree growth under water stress conditions. The results of this study have important practical implications for managing commercial Pinus forests across the globe: First, selection of family and genotype with both greater WUE and better plant growth appears to be practically possible without trade-off. This means that breeding programs using these traits should ensure that Pinus forest productivity is maintained or even improved under some degree of drought stress. Second, the genomic selection for WUE using δ13C is possible with a good degree of accuracy. Carbon stable isotopes are a useful tool to increase the agility of global breeding programs to respond to the challenges of climate change.
Data Availability Statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: https://doi.org/10.5281/zenodo.5768774 (Ismael et al., 2021).
Author Contributions
AI analyzed the data and wrote the manuscript. HD, JX, DM, and YL conceived and supervised the study, made substantial contributions to the interpretation of the results, and contributed to the revision of the manuscript. K-TB, PB, and MG-G collected the data. MG and JK made substantial contributions to the interpretation of the results and contributed to the revision of the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This study was funded by MBIE Strategic Science Investment Funding (no. C04X1703), leveraging the investment of the Radiata Pine Breeding Company (RPBC) through the RPBC MBIE Genomic Selection Partnership (no. RPBC1301), and the RPBC MBIE breeding partnership (no. C04X1808).
Conflict of Interest
AI was employed by Livestock Improvement Corporation (LIC), Hamilton, New Zealand, and Scion, New Zealand. DM, JX, JK, YL, PB, MG, K-TB, ET, and HD were employed by Scion, New Zealand.
The remaining author declare that this study received funding from NZ Ministry of Business, Innovation and Employment and the Radiata Pine Breeding Company. The funders were not involved in the study design, analysis, interpretation of data, the writing of this article or the decision to submit it for publication.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
The authors are grateful for and acknowledge the access to genetic material and genotypes. The authors would also like to acknowledge Beena Poi, Trish Moke-Pouwhare, Debbie Shann, and Raniera Mana for sample collection and processing.
References
Adams, H. D., Guardiola-Claramonte, M., Barron-Gafford, G. A., Villegas, J. C., Breshears, D. D., Zou, C. B., et al. (2009). Temperature sensitivity of drought-induced tree mortality portends increased regional die-off under global-change-type drought. Proc. Natl. Acad. Sci. U.S.A. 106, 7063–7066. doi: 10.1073/pnas.0901438106
Allen, C. D., Macalady, A. K., Chenchouni, H., Bachelet, D., Mcdowell, N., Vennetier, M., et al. (2010). A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 259, 660–684. doi: 10.1016/j.foreco.2009.09.001
Baker, N. R. (2008). Chlorophyll fluorescence: a probe of photosynthesis in vivo. Annu. Rev. Plant Biol. 59, 89–113. doi: 10.1146/annurev.arplant.59.032607.092759
Baltunis, B. S., Martin, T. A., Huber, D. A., and Davis, J. M. (2008). Inheritance of foliar stable carbon isotope discrimination and third-year height in Pinus taeda clones on contrasting sites in Florida and Georgia. Tree Genet. Genomes 4, 797–807. doi: 10.1007/s11295-008-0152-2
Barbour, M. M., Walcroft, A. S., and Farquhar, G. D. (2002). Seasonal variation in δ13C and δ18O of cellulose from growth rings of Pinus radiata. Plant Cell Environ. 25, 1483–1499. doi: 10.1046/j.0016-8025.2002.00931.x
Bartholomé, J., Van Heerwaarden, J., Isik, F., Boury, C., Vidal, M., Plomion, C., et al. (2016). Performance of genomic prediction within and across generations in maritime pine. BMC Genomics 17:604. doi: 10.1186/s12864-016-2879-8
Beaulieu, J., Doerksen, T. K., Mackay, J., Rainville, A., and Bousquet, J. (2014). Genomic selection accuracies within and between environments and small breeding groups in white spruce. BMC Genomics 15:1048. doi: 10.1186/1471-2164-15-1048
Bennett, K. J., and Rook, D. A. (1978). Stomatal and mesophyll resistances in two clones of Pinus radiata D. Don known to differ in transpiration and survival rate. Funct. Plant Biol. 5, 231–238.
Brendel, O., Pot, D., Plomion, C., Rozenberg, P., and Guehl, J.-M. (2002). Genetic parameters and QTL analysis of δ13C and ring width in maritime pine. Plant Cell Environ. 25, 945–953.
Butler, D. G., Cullis, B. R., Gilmour, A. R., and Gogel, B. J. (2009). ASReml-R Reference Manual. Brisbane, QLD: The State of Queensland, Department of Primary Industries and Fisheries.
Chakhchar, A., Haworth, M., El Modafar, C., Lauteri, M., Mattioni, C., Wahbi, S., et al. (2017). An assessment of genetic diversity and drought tolerance in argan tree (Argania spinosa) populations: potential for the development of improved drought tolerance. Front. Plant Sci. 8:276. doi: 10.3389/fpls.2017.00276
Chappell, P. R. (2014). The Climate and Weather of Bay of Plenty, New Zealand. NIWA Science and Technology Series No. 62. Wellington: NIWA.
Chen, Z.-Q., Baison, J., Pan, J., Karlsson, B., Andersson, B. G., Westin, J., et al. (2018). Accuracy of genomic selection for growth and wood quality traits in two control-pollinated progeny trials using exome capture as genotyping platform in Norway spruce. BMC Genomics 19:946. doi: 10.1186/s12864-018-5256-y
Condon, A. G., Richards, R. A., Rebetzke, G. J., and Farquhar, G. D. (2002). Improving intrinsic water-use efficiency and crop yield presented at the 1999 CSSA symposium on water use efficiency, organized by Div. C-2 chair, Dr. Tom Gerik. Crop Sci. 42, 122–131. doi: 10.2135/cropsci2002.1220
Condon, A. G., Richards, R. A., Rebetzke, G. J., and Farquhar, G. D. (2004). Breeding for high water-use efficiency. J. Exp. Bot. 55, 2447–2460. doi: 10.1093/jxb/erh277
Craig, H. (1957). Isotopic standards for carbon and oxygen and correction factors for mass-spectrometric analysis of carbon dioxide. Geochim. Cosmochim. Acta 12, 133–149. doi: 10.1016/0016-7037(57)90024-8
Cregg, B. M., and Zhang, J. W. (2001). Physiology and morphology of Pinus sylvestris seedlings from diverse sources under cyclic drought stress. For. Ecol. Manag. 154, 131–139.
Cumbie, W. P., Eckert, A., Wegrzyn, J., Whetten, R., Neale, D., and Goldfarb, B. (2011). Association genetics of carbon isotope discrimination, height and foliar nitrogen in a natural population of Pinus taeda L. Heredity 107, 105–114. doi: 10.1038/hdy.2010.168
Dias, M. C., and Brüggemann, W. (2010). Limitations of photosynthesis of Phaseolus vulgaris under srought stress: gas exchange, chlorophyll fluorescence and Calvin cycle enzymes. Photosynthetica 48, 96–102. doi: 10.1007/s11099-010-0013-8
Dixon, L., and Carter, A. H. (2019). Toward a new use for carbon isotope discrimination in wheat breeding. Agronomy 9:385. doi: 10.3390/agronomy9070385
Dungey, H. S., Brawner, J. T., Burger, F., Carson, M., Henson, M., Jefferson, P., et al. (2009). A new breeding strategy for Pinus radiata in New Zealand and New South Wales. Silvae Genet. 58, 28–38. doi: 10.1515/sg-2009-0004
Dunningham, A., Kirschbaum, M., Payn, T., and Meason, D. (2012). “Chapter 7. Forestry: long-term adaptation of productive forests in a changing climatic environment. impacts of climate change on land-based sectors and adaptation options,” in Technical Report to the Sustainable Land Management and Climate Change Adaptation Technical Working Group. MPI Technical Paper, (2012/33) (Wellington: Ministry for Primary Industries), 293–346.
Ehdaie, B., and Waines, J. G. (1994). Genetic analysis of carbon isotope discrimination and agronomic characters in a bread wheat cross. Theor. Appl. Genet. 88, 1023–1028. doi: 10.1007/BF00220811
Endelman, J. B. (2011). Ridge regression and other kernels for genomic selection with R package rrBLUP. Plant Genome 4, 250–255.
Espinoza, S. E., Magni, C. R., Rubilar, R. A., Yañez, M. A., Santelices, R. E., Cabrera, A. M., et al. (2017). Field performance of various Pinus radiata breeding families established on a drought-prone site in central Chile. N. Z. J. For. Sci. 47:12. doi: 10.4206/bosque.1977.v2n1-03
Espinoza, S. E., Martínez, V. A., Magni, C. R., Ivković, M., Santelices, R. E., Guerra, F. P., et al. (2014). Genetic control of growth, biomass allocation, and survival under drought stress in Pinus radiata D. Don seedlings. Tree Genet. Genomes 10, 1045–1054. doi: 10.1007/s11295-014-0741-1
Fahlgren, N., Feldman, M., Gehan, M. A., Wilson, M. S., Shyu, C., Bryant, D. W., et al. (2015). A versatile phenotyping system and analytics platform reveals diverse temporal responses to water availability in Setaria. Mol. Plant 8, 1520–1535. doi: 10.1016/j.molp.2015.06.005
Falconer, D. S., and Mackay, T. F. C. (1996). Introduction to Quantitative Genetics. Essex: Longman Group.
Famula, R. A., Richards, J. H., Famula, T. R., and Neale, D. B. (2019). Association genetics of carbon isotope discrimination and leaf morphology in a breeding population of Juglans regia L. Tree Genet. Genomes 15:6.
Farquhar, G. D., Hubick, K. T., Condon, A. G., and Richards, R. A. (1989). “Carbon isotope fractionation and plant water-use efficiency,” in Stable Isotopes in Ecological Research. Ecological Studies (Analysis and Synthesis), eds P. W. Rundel, J. R. Ehleringer, and K. A. Nagy (New York, NY: Springer), 21–40. doi: 10.1007/978-1-4612-3498-2_2
Farquhar, G. D., O’leary, M. H., and Berry, J. A. (1982). On the relationship between carbon isotope discrimination and the intercellular carbon dioxide concentration in leaves. Aust. J. Plant Physiol. 9, 121–137. doi: 10.1111/j.1469-8137.2008.02518.x
Forest Owners Association (2016). Facts and Figures New Zealand Plantation Forest Industry. Wellington: Forest Owners Association.
Forni, S., Aguilar, I., and Misztal, I. (2011). Different genomic relationship matrices for single-step analysis using phenotypic, pedigree and genomic information. Genet. Sel. Evol. 43:1. doi: 10.1186/1297-9686-43-1
Gallart, M., Love, J., Meason, D. F., Coker, G., Clinton, P. W., Xue, J., et al. (2019). Field-scale variability in site conditions explain phenotypic plasticity in response to nitrogen source in Pinus radiata D. Don. Plant Soil 443, 353–368. doi: 10.1007/s11104-019-04237-0
Gamal El-Dien, O., Ratcliffe, B., Klápště, J., Chen, C., Porth, I., and El-Kassaby, Y. A. (2015). Prediction accuracies for growth and wood attributes of interior spruce in space using genotyping-by-sequencing. BMC Genomics 16:370. doi: 10.1186/s12864-015-1597-y
Gamal El-Dien, O., Ratcliffe, B., Klápště, J., Porth, I., Chen, C., and El-Kassaby, Y. A. (2016). Implementation of the realized genomic relationship matrix to open-pollinated white spruce family testing for disentangling additive from nonadditive genetic effects. G3 6, 743–753. doi: 10.1534/g3.115.025957
Gapare, W. J., Ivković, M., Dutkowski, G. W., Spencer, D. J., Buxton, P., and Wu, H. X. (2012). Genetic parameters and provenance variation of Pinus radiata D. Don. ‘Eldridge collection’ in Australia 1: growth and form traits. Tree Genet. Genomes 8, 391–407. doi: 10.1007/s11295-011-0449-4
Gaspar, M. J., Velasco, T., Feito, I., Alía, R., and Majada, J. (2013). Genetic variation of drought tolerance in Pinus pinaster at three hierarchical levels: a comparison of induced osmotic stress and field testing. PLoS One 8:e79094. doi: 10.1371/journal.pone.0079094
Gebrekirstos, A., Van Noordwijk, M., Neufeldt, H., and Mitlöhner, R. (2011). Relationships of stable carbon isotopes, plant water potential and growth: an approach to asses water use efficiency and growth strategies of dry land agroforestry species. Trees 25, 95–102. doi: 10.1007/s00468-010-0467-0
Grattapaglia, D., and Resende, M. D. V. (2011). Genomic selection in forest tree breeding. Tree Genet. Genomes 7, 241–255. doi: 10.1007/s11295-010-0328-4
Hatton, T. J., and Vertessy, R. A. (1990). Transpiration of plantation Pinus radiata estimated by the heat pulse method and the Bowen ratio. Hydrol. Process. 4, 289–298. doi: 10.1002/hyp.3360040309
IPCC (2013). “Working group I contribution to the IPCC fifth assessment report (AR5), climate change 2013: the physical science basis approved summary for policymakers,” in Summary for Policymakers, eds T. F. Stocker, D. Qin, G.-K. Plattner, M. Tignor, S. K. Allen, J. Boschung, et al. (Geneva: IPCC).
Isik, F. (2014). Genomic selection in forest tree breeding: the concept and an outlook to the future. New For. 45, 379–401. doi: 10.1007/s11056-014-9422-z
Isik, F., Bartholomé, J., Farjat, A., Chancerel, E., Raffin, A., Sanchez, L., et al. (2016). Genomic selection in maritime pine. Plant Sci. 242, 108–119. doi: 10.1016/j.plantsci.2015.08.006
Ismael, A., Xue, J., Meason, D. Klápšte, J., Gallart, M., Li, Y., et al. (2021). Data for Genetic Variation in Drought-Tolerance Traits and Their Relationships to Growth in Pinus radiata D. Don Under Water Stress. doi: 10.5281/zenodo.5768774
Johnsen, K. H., Flanagan, L. B., Huber, D. A., and Major, J. E. (1999). Genetic variation in growth, carbon isotope discrimination, and foliar N concentration in Picea mariana: analyses from a half-diallel mating design using field-grown trees. Can. J. For. Res. 29, 1727–1735. doi: 10.1139/x99-144
Katul, G., Leuning, R., and Oren, R. (2003). Relationship between plant hydraulic and biochemical properties derived from a steady-state coupled water and carbon transport model. Plant Cell Environ. 26, 339–350. doi: 10.1046/j.1365-3040.2003.00965.x
Klisz, M., Buras, A., Sass-Klaassen, U., Puchalka, R., Koprowski, M., and Ukalska, J. (2019). Limitations at the limits? Diminishing of genetic effects in Norway spruce provenance trials. Front. Plant Sci. 10:306. doi: 10.3389/fpls.2019.00306
Kolb, T. E., Fettig, C. J., Bentz, B. J., Stewart, J. E., Weed, A. S., Hicke, J. A., et al. (2016). “Forest insect and fungal pathogen responses to drought,” in Effects of Drought on Forests and Rangelands in the United States: A Comprehensive Science Synthesis. Gen. Tech. Rep. WO-93b (Washington, DC: U.S, Department of Agriculture, Forest Service, Washington Office), 113–133.
Korol, R. L., Kirschbaum, M. U., Farquhar, G. D., and Jeffreys, M. (1999). Effects of water status and soil fertility on the C-isotope signature in Pinus radiata. Tree Physiol. 19, 551–562. doi: 10.1093/treephys/19.9.551
Kumar, S., Burdon, R. D., Stovold, G. T., and Gea, L. D. (2008). Implications of selection history on genetic architecture of growth, form, and wood-quality traits in Pinus radiata. Can. J. For. Res. 38, 2372–2381. doi: 10.1139/x08-086
Lee, S. H., Goddard, M. E., Visscher, P. M., and Van Der Werf, J. H. (2010). Using the realized relationship matrix to disentangle confounding factors for the estimation of genetic variance components of complex traits. Genet. Sel. Evol. 42:22. doi: 10.1186/1297-9686-42-22
Li, Y., and Dungey, H. S. (2018). Expected benefit of genomic selection over forward selection in conifer breeding and deployment. PLoS One 13:e0208232. doi: 10.1371/journal.pone.0208232
Li, Y., Xue, J., Clinton, P. W., and Dungey, H. S. (2015). Genetic parameters and clone by environment interactions for growth and foliar nutrient concentrations in radiata pine on 14 widely diverse New Zealand sites. Tree Genet. Genomes 11:10.
Lindgren, D., Gea, L. D., and Jefferson, P. A. (1997). Status number for measuring genetic diversity. For. Genet. 4, 762–764.
Marguerit, E., Bouffier, L., Chancerel, E., Costa, P., Lagane, F., Guehl, J.-M., et al. (2014). The genetics of water-use efficiency and its relation to growth in maritime pine. J. Exp. Bot. 65, 4757–4768. doi: 10.1093/jxb/eru226
Maxwell, K., and Johnson, G. N. (2000). Chlorophyll fluorescence-a practical guide. J. Exp. Bot. 51, 659–668. doi: 10.1093/jxb/51.345.659
Meason, D. F., and Mason, W. L. (2014). Evaluating the deployment of alternative species in planted conifer forests as a means of adaptation to climate change—case studies in New Zealand and Scotland. Ann. For. Sci. 71, 239–253. doi: 10.1007/s13595-013-0300-1
Mena-Petite, A., González-Moro, C., González-Murua, M. L., and Rueda, A. M. (2000). Sequential effects of acidic precipitation and drought on photosynthesis and chlorophyll fluorescence parameters of Pinus radiata D. Don seedlings. J. Plant Physiol. 156, 84–92. doi: 10.1016/s0176-1617(00)80276-x
Mena-Petite, A., Munoz-Rueda, A., and Lacuesta, M. (2005). Effect of cold storage treatments and transplanting stress on gas exchange, chlorophyll fluorecence and survival under water limiting conditions of Pinus radiata stock-types. Eur. J. For. Res. 124, 73–82. doi: 10.1007/s10342-005-0056-8
Meuwissen, T. H. E., Hayes, B. J., and Goddard, M. E. (2001). Prediction of total genetic value using genome-wide dense marker maps. Genetics 157, 1819–1829. doi: 10.1093/genetics/157.4.1819
Momen, M., Mehrgardi, A. A., Sheikhy, A., Esmailizadeh, A., Fozi, M. A., Kranis, A., et al. (2017). A predictive assessment of genetic correlations between traits in chickens using markers. Genet. Sel. Evol. 49:16. doi: 10.1186/s12711-017-0290-9
Mullan, B., Porteous, A., Wratt, D., and Hollis, M. (2005). Changes in Drought Risk with Climate Change. NIWA Client Report: WLG2005-23, Prepared for Ministry for the Environment (NZ Climate Change Office) and Ministry of Agriculture and Forestry. Wellington: National Institute of Water & Atmospheric Research Ltd.
Neves, L. G., Davis, J. M., Barbazuk, W. B., and Kirst, M. (2013). Whole-exome targeted sequencing of the uncharacterized pine genome. Plant J. 75, 146–156. doi: 10.1111/tpj.12193
New Zealand Forest Owners Association [NZFOA] (2019). New Zealand Forest Industry Facts & Figures 2018/2019. Wellington: Forest Owners Association.
Pita, P., Cañas, I., Soria, F., Ruiz, F., and Toval, G. (2005). Use of physiological traits in tree breeding for improved yield in drought-prone environments. The case of Eucalyptus globulus. 14, 383–393. doi: 10.5424/srf/2005143-00931
Powell, J. E., Visscher, P. M., and Goddard, M. E. (2010). Reconciling the analysis of IBD and IBS in complex trait studies. Nat. Rev. Genet. 11, 800–805. doi: 10.1038/nrg2865
Rapacz, M., Wójcik-Jagła, M., Fiust, A., Kalaji, H. M., and Kościelniak, J. (2019). Genome-wide associations of chlorophyll fluorescence OJIP transient parameters connected with soil drought response in barley. Front. Plant Sci. 10:78. doi: 10.3389/fpls.2019.00078
Rebetzke, G. J., Condon, A. G., Richards, R. A., and Farquhar, G. D. (2002). Selection for reduced carbon isotope discrimination increases aerial biomass and grain yield of rainfed bread wheat. Crop Sci. 42, 739–745. doi: 10.2135/cropsci2002.0739
Resende, M. F. Jr., Munoz, P., Acosta, J. J., Peter, G. F., Davis, J. M., Grattapaglia, D., et al. (2012). Accelerating the domestication of trees using genomic selection: accuracy of prediction models across ages and environments. New Phytol. 193, 617–624. doi: 10.1111/j.1469-8137.2011.03895.x
Richards, R. A., and Condon, A. G. (1993). “Challenges ahead in using carbon isotope discrimination in plant-breeding programs,” in Stable Isotopes and Plant Carbon-Water Relations, eds J. R. Ehleringer, A. E. Hall, and G. D. Farquhar (San Diego, CA: Academic Press), 451–462. doi: 10.1016/b978-0-08-091801-3.50038-6
Rigling, A., Bigler, C., Eilmann, B., Feldmeyer-Christe, E., Gimmi, U., Ginzler, C., et al. (2013). Driving factors of a vegetation shift from Scots pine to pubescent oak in dry Alpine forests. Glob. Change Biol. 19, 229–240. doi: 10.1111/gcb.12038
Rodríguez-Gamir, J., Xue, J., Clearwater, M. J., Meason, D. F., Clinton, P. W., and Domec, J.-C. (2019). Aquaporin regulation in roots controls plant hydraulic conductance, stomatal conductance, and leaf water potential in Pinus radiata under water stress. Plant Cell Environ. 42, 717–729. doi: 10.1111/pce.13460
Rodriguez-Gamir, J., Xue, J., Meason, D. F., Clearwater, M., Clinton, P. W., and Domec, J.-C. (2020). Inter-clonal variation, coordination and trade-offs between hydraulic conductance and gas exchange in Pinus radiata. J. Exp. Bot. 72, 2419–2433. doi: 10.1093/jxb/eraa587
Roussel, M., Dreyer, E., Montpied, P., Le-Provost, G., Guehl, J.-M., and Brendel, O. (2009). The diversity of 13C isotope discrimination in a Quercus robur full-sib family is associated with differences in intrinsic water use efficiency, transpiration efficiency, and stomatal conductance. J. Exp. Bot. 60, 2419–2431. doi: 10.1093/jxb/erp100
Sinclair, T. R. (2011). Challenges in breeding for yield increase for drought. Trends Plant Sci. 16, 289–293. doi: 10.1016/j.tplants.2011.02.008
Su, L., Dai, Z., Li, S., and Xin, H. (2015). A novel system for evaluating drought–cold tolerance of grapevines using chlorophyll fluorescence. BMC Plant Biol. 15:82. doi: 10.1186/s12870-015-0459-8
Subrahmanyam, D., Subash, N., Haris, A., and Sikka, A. K. (2006). Influence of water stress on leaf photosynthetic characteristics in wheat cultivars differing in their susceptibility to drought. Photosynthetica 44:125. doi: 10.1007/s11099-005-0167-y
Sun, Z. J., Livingston, N. J., Guy, R. D., and Ethier, G. J. (1996). Stable carbon isotopes as indicators of increased water use efficiency and productivity in white spruce (Picea glauca (Moench) Voss) seedlings. Plant Cell Environ. 19, 887–894. doi: 10.1111/j.1365-3040.1996.tb00425.x
Tambarussi, E. V., Pereira, F. B., Acosta, J. J., and Vencovsky, R. (2019). “Status (Ns) and variance effective (Ne) numbers as useful parameters for breeding and conservation genetics programs,” in Proceedings of the IUFRO Tree Biotechnology 2019 Meeting, Raleigh, NC. doi: 10.13140/RG.2.2.16409.26729
Telfer, E., Graham, N., Macdonald, L., Li, Y., Klápště, J., and Resende, M. Jr. (2019). A high-density exome capture genotype-by-sequencing panel for forestry breeding in Pinus radiata. PLoS One 14:e0222640. doi: 10.1371/journal.pone.0222640
Telfer, E., Graham, N., Macdonald, L., Sturrock, S., Wilcox, P., and Stanbra, L. (2018). Approaches to variant discovery for conifer transcriptome sequencing. PLoS One 13:e0205835. doi: 10.1371/journal.pone.0205835
Thavamanikumar, S., Southerton, S. G., Bossinger, G., and Thumma, B. R. (2013). Dissection of complex traits in forest trees — opportunities for marker-assisted selection. Tree Genet. Genomes 9, 627–639. doi: 10.3389/fpls.2018.01693
VanRaden, P. (2008). Efficient methods to compute genomic predictions. J. Dairy Sci. 91, 4414–4423. doi: 10.3168/jds.2007-0980
Veerkamp, R. F., Mulder, H. A., Thompson, R., and Calus, M. P. L. (2011). Genomic and pedigree-based genetic parameters for scarcely recorded traits when some animals are genotyped. J. Dairy Sci. 94, 4189–4197. doi: 10.3168/jds.2011-4223
Waghorn, M. J., Whitehead, D., Watt, M. S., Mason, E. G., and Harrington, J. J. (2015). Growth, biomass, leaf area and water-use efficiency of juvenile Pinus radiata in response to water deficits. N. Z. J. For. Sci. 45:3.
Walcroft, A. S., Silvester, W. B., Grace, J. C., Carson, S. D., and Waring, R. H. (1996). Effects of branch length on carbon isotope discrimination in Pinus radiata. Tree Physiol. 16, 281–286. doi: 10.1093/treephys/16.1-2.281
Walcroft, A. S., Silvester, W. B., Whitehead, D., and Kelliher, F. M. (1997). Seasonal changes in stable carbon isotope ratios within annual rings of Pinus radiata reflect environmental regulation of growth processes. Funct. Plant Biol. 24, 57–68. doi: 10.1071/pp96025
Watt, M. S., Palmer, D. J., Kimberley, M. O., Höck, B. K., Payn, T. W., and Lowe, D. J. (2010). Development of models to predict Pinus radiata productivity throughout New Zealand. Can. J. For. Res. 40, 488–499. doi: 10.1139/x09-207
Whitehead, D., Sheriff, D. W., and Greer, D. H. (1983). The relationship between stomatal conductance, transpiration rate and tracheid structure in Pinus radiata clones grown at different water vapour saturation deficits. Plant Cell Environ. 6, 703–710. doi: 10.1111/1365-3040.ep11589331
Wright, S. (1922). Coefficients of inbreeding and relationship. Am. Nat. 56, 330–338. doi: 10.1086/279872
Wright, S. (1931). Evolution in Mendelian populations. Genetics 16, 97–159. doi: 10.1093/genetics/16.2.97
Wright, S. (1933). Inbreeding and homozygosis. Proc. Natl. Acad. Sci. U.S.A. 19, 411–420. doi: 10.1073/pnas.19.4.411
Wu, F. Z., Bao, W. K., Li, F. L., and Wu, N. (2008). Effects of water stress and nitrogen supply on leaf gas exchange and fluorescence parameters of Sophora davidii seedlings. Photosynthetica 46, 40–48. doi: 10.1007/s11099-008-0008-x
Xu, Z., Prasolova, N., Lundkvist, K., Beadle, C., and Leaman, T. (2003). Genetic variation in branchlet carbon and nitrogen isotope composition and nutrient concentration of 11-year-old hoop pine families in relation to tree growth in subtropical Australia. For. Ecol. Manag. 186, 359–371. doi: 10.1093/treephys/20.15.1049
Xu, Z. H., Saffigna, P. G., Farquhar, G. D., Simpson, J. A., Haines, R. J., Walker, S., et al. (2000). Carbon isotope discrimination and oxygen isotope composition in clones of the F1 hybrid between slash pine and Caribbean pine in relation to tree growth, water-use efficiency and foliar nutrient concentration. Tree Physiol. 20, 1209–1217. doi: 10.1093/treephys/20.18.1209
Xue, J., Dungey, H., Clinton, P., Henley, D., Niollet, S., and Leckie, A. (2012). The Potential for Using Foliar Carbon Isotopic Signature to Screen Drought Tolerant Radiata Pine Genotypes for Dryland Plantation Forests in New Zealand. MPI Technical Paper No: 2013/38. Wellington: New Zealand Forest Research Institute Limited.
Keywords: drought tolerance, water stress, heritability, genetic correlation, genomic selection, carbon isotope composition, Pinus radiata, chlorophyll fluorescence
Citation: Ismael A, Xue J, Meason DF, Klápště J, Gallart M, Li Y, Bellè P, Gomez-Gallego M, Bradford K-T, Telfer E and Dungey H (2022) Genetic Variation in Drought-Tolerance Traits and Their Relationships to Growth in Pinus radiata D. Don Under Water Stress. Front. Plant Sci. 12:766803. doi: 10.3389/fpls.2021.766803
Received: 30 August 2021; Accepted: 29 November 2021;
Published: 04 January 2022.
Edited by:
Jianjun Chen, University of Florida, United StatesReviewed by:
Evandro Vagner Tambarussi, Midwestern Paraná State University, BrazilSergio Espinoza, Catholic University of the Maule, Chile
Copyright © 2022 Ismael, Xue, Meason, Klápště, Gallart, Li, Bellè, Gomez-Gallego, Bradford, Telfer and Dungey. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ahmed Ismael, YWhtZWQuaXNtYWVsQGxpYy5jby5ueg==, YWhtZWRpc21hZWxzYXllZEBnbWFpbC5jb20=