 Xue Wu
Xue Wu Xin-Jun Zheng
Xin-Jun Zheng Xiao-Han Mu
Xiao-Han Mu Yan Li
Yan Li
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Plant Sci. , 10 November 2021
Sec. Functional Plant Ecology
Volume 12 - 2021 | https://doi.org/10.3389/fpls.2021.754887
This article is part of the Research Topic The Relationship between Climate Change and Species Distribution View all 9 articles
The allometric relationship among different functional traits is an ecological strategy for plants to promote resource utilization, which indicates the ability of plants to adapt to environmental changes coordinately. In this study, we conducted a field survey on Haloxylon ammodendron and H. persicum among different terrains (dune crest, eastern slope, western slope and inter-dune) in the Gurbantunggut Desert, obtained their quantitative and morphological characteristics, and analyzed their allometric relationships between plant height and canopy radius, plant height and basal diameter by using standardized major axis estimation. We found that: (1) The dominated terrains of H. ammodendron and H. persicum were different; (2) The individual morphology of the two Haloxylon species changed significantly with the terrains (p < 0.05), with the largest and smallest ones growing on the eastern slope and the inter-dune lowland, respectively; (3) Fixed allometric patterns were observed in the above-ground parts of the two Haloxylon species, as the growth of canopy and basal stem was preferentially to plant height; (4) These allometric relationships were significantly affected by the terrain, and exhibited discrepancy between two species, they both invested less in plant height in windy habitats, such as the dune crest and western slope, but H. ammodendron growing on the western slope and H. persicum growing on the eastern slope invested more in basal diameter for strengthening mechanical support and resources acquisition, respectively. These results indicated that both studied species adopted an ecological strategy that allocating more resources to horizontal expansion rather than vertical growth, the terrain has an important influence on the allometric relationship of their above-ground parts, and the trade-off mechanism of main components investing was different for these two species due to habitat heterogeneity and ecological adaptability.
The resources such as nutrients and water as well as carbohydrates obtained by plants through photosynthesis and root absorption determine their life processes such as survival, growth, and reproduction (Allen et al., 2008; Chen et al., 2013; Jennifer et al., 2019). However, since the resources that plants could obtain are limited, the resource distribution among various physiological/ecological functions often appears conflicting (Ogden, 1974; Liu and Ma, 2015). These conflicting resource requirements determine that plants need to balance among different functional traits, and allometric scaling is one of the most direct manifestations of such balance (Weiner, 2004). For example, with changing environment, plant biomass distributes differently in different vegetative organs (e.g., roots, stems, and leaves) or reproductive organs (e.g., flowers, fruits, and seeds), and varies between above- and below-ground parts (Cheng et al., 2014; Zhang and Peng, 2018; Chen et al., 2020; Sun et al., 2021; Zheng et al., 2021). These changes directly reflect the ecological strategies that plants adopt to cope with environmental selection pressures (Smith et al., 1985; Primack, 1987; Midgley and Bond, 1989). Allometric scaling has been proven to be an valid theory for describing the proportional relationship among plant morphological characteristics since it was first proposed, and it has been widely used because it can reveal the relationship between the size and physiological attributes of plant individuals (West et al., 1997; Lu et al., 2007; Han and Fang, 2008).
Morphological parameters such as plant height, basal diameter, and canopy radius are the most easily observed and measurable indicators in plants (Noriyuki, 2011). They partially determine the contact area of plants to external resources, and are the basis for characterizing plant growth and development (Archibald and Bond, 2003; Li S. et al., 2011). The allometric relationships among the above-ground parts of plants, that is, the relative relationship between plant height and canopy radius and that between plant height and basal diameter, has an important influence on external morphogenesis and metabolic activities of plant individuals (Zhang et al., 2009; Hou et al., 2014; Shi et al., 2015). However, the knowledge of how this allometric relationship changes along the environmental gradient is scarce, which will restrict the accurate simulation and prediction about plant growth dynamics under the background of global climate change (Lines et al., 2012).
Haloxylon ammodendron and H. persicum are typical xerophytic species with well-developed root systems, degraded leaves, and used assimilating branches to perform photosynthesis (Pyankov et al., 1999). They can resist a variety of stresses, such as barrenness, drought, salt, alkali, wind, and sand, are excellent species for wind prevention, sand fixation, and microclimate improvement in arid region (Wang et al., 2011). Within Central Asian desert ecosystems, they naturally occur and are the dominant species that widely distribute in China, Kazakhstan, Uzbekistan, Turkmenistan, and many other Central Asian countries (Buras et al., 2012). Determining the mechanisms underlying the spatial distribution of plant species is one of the central themes in biogeography and ecology (Xu et al., 2014), and the influence of terrain is one of the important research topics of this theme. The micro-habitats resulting from different terrains show strong heterogeneity in temperature, wind, soil physical and chemical properties, and directly or indirectly affecting the growth, development, and reproduction of plants, especially in the seed germination and seedling settlement stages (Li et al., 2005, Li H. et al., 2011; Xu et al., 2014; Dang et al., 2015). Exploring the effect of terrain on the two Haloxylon species’ distribution is conducive to provide better protection and management for them, and is also beneficial to sustain their role in maintaining local ecological safety. Studies have been done on their biomass allocating pattern between above- and below- ground parts, between stems and assimilated branches, and the allometric relationship between the area of assimilated branches and stems (Buras et al., 2012; Li Z. B. et al., 2013). However, the allometric relationship among basic functional traits such as plant height, basal diameter, and canopy radius of these two species, as well as the determinants that influencing their resource allocation in different terrains have not yet been clarified. Studying the allometric scaling of important plant functional traits and their determining factors under different terrain conditions will help us to further understand the ecological adaptation mechanisms of plants to environmental changes. For this purpose, we selected H. ammodendron and H. persicum in the southern Guerbantonggut Desert of Central Asia, Xinjiang, as the research material to explore the differences in their distribution, morphology, and the allometric scaling (plant height vs. canopy radius and plant height vs. basal diameter) under different terrain conditions. Our objective is to reveal the responses of desert plants’ distribution, structural function traits and allometry to different terrains, and deepen the understanding of desert plants adapting to varying environment.
The study was carried out at the vicinity of Fukang Station of Desert Ecology, Chinese Academy of Sciences (44°17′N, 87°56′E and elevation 475 m, represented by a red star in Figure 1A). The study area is located in the Gurbantunggut Desert, at the arid center of the Europe-Asia Continent. It has a typical temperate continental arid climate, with cold winters, hot summers, and large temperature differences between day and night. The annual average temperature is 6.6°C, the annual potential evapotranspiration is around 1,000 mm, and the annual precipitation is 100–150 mm. The days with snow cover are 100–160, and the maximum depth of snow cover is above 20 cm. The soil type can be divided into denser gray desert soil, salinized gray desert soil, and aeolian sandy soil with higher sand content according to the texture. The majority of the desert is covered by sand dunes and inter-dune lowland. The dunes are mainly longitudinal sand dunes, with a length up to more than 10 km, at north-south direction, and a height ranging from 10 to 50 m. There is a 10–40 m wide flowing zone on the dune crest, with slightly asymmetrical dune profiles (Wang et al., 2003). H. ammodendron and H. persicum are the dominate species in this area (Figure 1B). They are perennial shrubs or small trees belonging to the Chenopodiaceae family and are the most widely distributed species in the deserts of Central Asia.
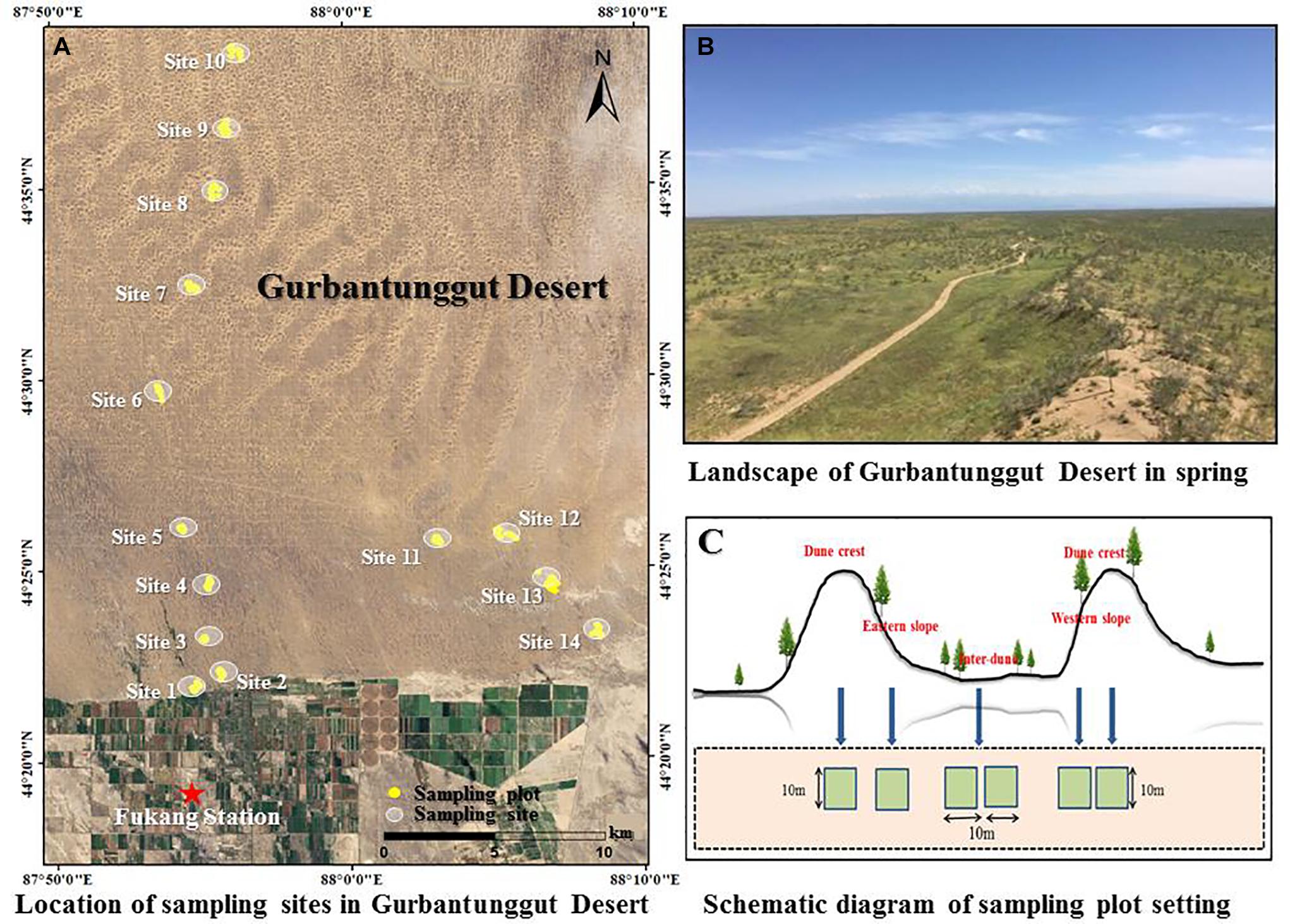
Figure 1. Landscape (A,B) and sampling diagram (A,C) of the study area.
The field survey started from August 2015, and there was no rainfall 8 days before the survey started. In order to collect more comprehensive data and provide reliable information on the distribution and morphological characteristics of H. ammodendron and H. persicum, we totally sampled 14 representative sites with well-developed natural vegetation, complete community structure, and less human disturbance. These sampling sites located from the southern edge of Gurbantunggut Desert to interior at the intervals of 3–5 km (Figure 1A and Table 1). For each sampling site, adjacent dunes with similar heights were selected, and four sample plots (10 m × 10 m) were set up on the two dune crests, eastern slope, and western slope, respectively (Figure 1C). In addition, eight plots (10 m × 10 m) were set up in the inner-dune lowland due to its larger area (Figure 1C). Finally, a total of 336 plots were sampled.
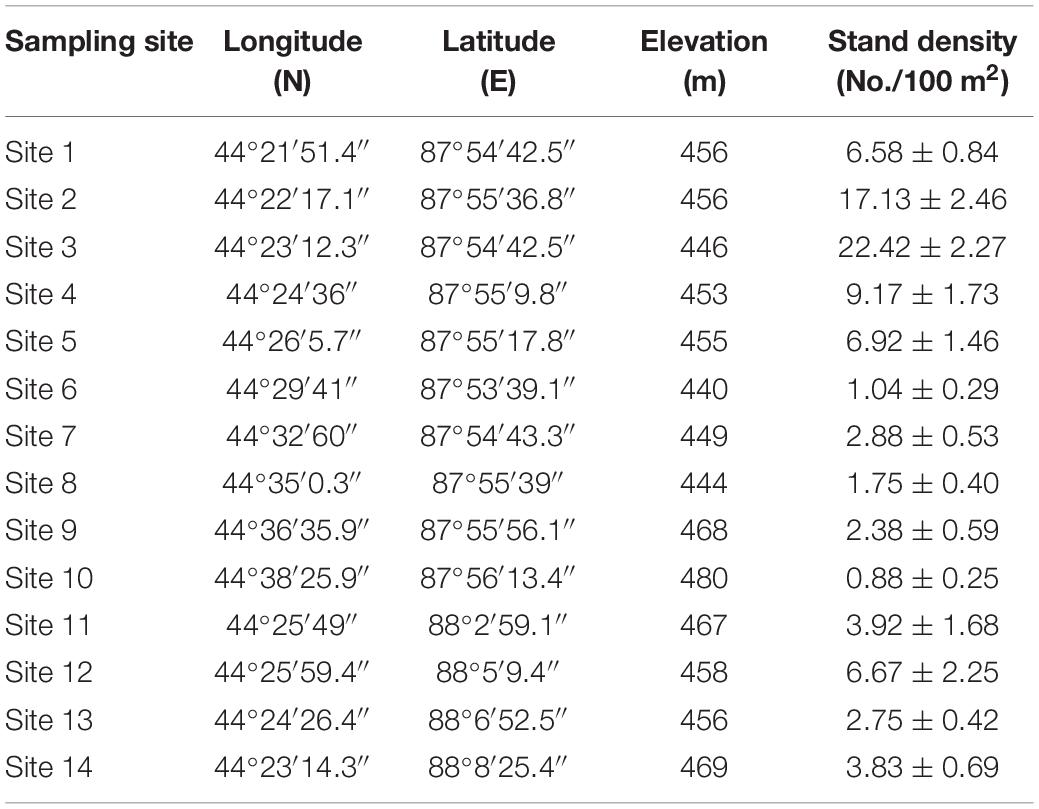
Table 1. Coordinates, elevation, and stand density of 14 sampling sites.
After removing the litter from the soil surface in each sampling plot, the five-point sampling method was used to collect a mixed soil sample from top to bottom (0–20 cm layer). The soil samples were placed in tin boxes and valve bags, then brought back to the laboratory. At the same time, using a cutting ring, a sample of the undisturbed soil was collected from the center of the sampling plot to determine soil bulk density.
Previous studies have shown that it is reasonable to model the shape of a desert shrub as a semi-triaxial ellipsoid with an elliptically shaped canopy (Li S. et al., 2011; Xu et al., 2017). The largest secant segment of the vertical projection of the individual on the ground surface is taken as the long axis of the canopy (a), the secant segment passing through the center perpendicular to the long axis is taken as the short axis (b), the distance from the highest point of the center to the ground is taken as the height (H). The diameter of the plant trunk on the ground is taken as the basal diameter (D) (Li S. et al., 2011). The height (H), basal diameter (D), long axis (a), and short axis (b) of each H. ammodendron or H. persicum in the plot were measured individually with a ruler. According to their basal diameter, plant individuals were divide into three development stages, seedling (D < 1.2 cm), young tree (1.2 cm ≤ D ≤ 6.5 cm), big tree (D > 6.5 cm), respectively (Jia and Lu, 2005).
Assuming that the canopy of H. ammodendron or H. persicum has an elliptical shape, the canopy area (CA) and canopy radius (R) were calculated according to the following equation:
where a and b are the long and short axis of the canopy, respectively.
The collected soil samples were air-dried and sieved (2 mm) to analyze physical and chemical properties, including bulk density (g/cm3), water content (%), pH, electrical conductivity (us/cm), organic carbon content (g/kg), total nitrogen content (g/kg), and total phosphorus content (g/kg). Soil bulk density was determined by the ring knife method. Soil water content was determined by the drying method, and on a mass basis, was converted to soil volumetric water content using the soil bulk density. Soil pH was determined by the potentiometric method (soil: water = 1:5). Soil electrical conductivity was determined by the electrical conductivity method (soil: water = 1:5). Soil organic carbon content was determined by the potassium dichromate external heating method. Soil total nitrogen content was determined by the Kelvin digestion method. Soil total phosphorus content was determined by the hydrochloric acid-hydrogen fluoride molybdenum antimony colorimetric method.
The relationship between plant height and basal diameter and between plant height and canopy radius of H. ammodendron or H. persicum in different terrains was determined using a linear regression model [log10(y) = log10(b) + alog10(x)] after linearization by logarithmic transformation. x and y represent the different attribute values of H. ammodendron and H. persicum. a is the slope, log10(b) is the intercept, and the value of a (Allometric index) indicates whether the relationship between the two attributes is isometric (a = 1) or allometric (a ≠ 1). A value of a > 1 indicates that the increase of y is greater than that of x, and a < 1 indicates that the increase of y is smaller than that of x. The log10(b) value does not affect the relationship. The allometric relationship between plant height (H), basal diameter (D), and canopy radius (R) were determined according to the following model (Li Y. et al., 2013):
The standardized major axis estimation was used to calculate the coefficient a and log10(b) of the regression equation. The principle of linear regression is to minimize the distance between the measured value and the fitted curve in the y-direction; thus, it is more suitable for predicting the value of one variable by another variable. However, due to the measurement errors in both x and y, it is not appropriate to only minimize the sum of squared deviations in the y direction. Unlike in linear regression models, the standardized major axis estimation determines the minimum distance between the measured value and the fitted curve, and also considers the deviation of the measured value from the fitted curve in both x and y directions, as well as the slope of the two attributes. Therefore, the standardized major axis estimation is more suitable for the slope estimation of the allometric scaling equation. Standardized major axis estimation was conducted in SMATR1, which can test for the heterogeneity of the slopes of different regression equations; p < 0.05 indicates that the difference between the slopes is significant, otherwise, a common slope is given. Minitab 17.0 was used to carry out the analysis of variance (ANOVA) and post hoc Tukey’s HSD multiple comparison of the stand density and morphological characteristics of H. ammodendron and H. persicum under different terrain conditions. Origin 8.5 (OriginLab) was used for graphical representation.
Soil physical and chemical properties of different terrains are shown in Figure 2. The results of ANOVA showed significant differences in soil volumetric water content, pH, electrical conductivity, organic carbon content, total nitrogen and total phosphorus content among different terrains (F = 3.06, p < 0.05; F = 3.80, p < 0.05; F = 3.46, p < 0.05; F = 72.08, p < 0.05; F = 12.59, p < 0.05; F = 21.14, p < 0.05). The soil volumetric water content of the eastern slope was the highest; the soil pH, electrical conductivity, organic carbon content, total nitrogen and total phosphorus content of the inter-dune lowland were significantly higher than those of the other terrains (p < 0.05). Moreover, the soil volumetric water content of the inter-dune lowland was also relatively high, indicating that the water and nutrient conditions of the inter-dune lowland habitat are superior. On the contrary, soil physical and chemical properties of the dune crest were the lowest.
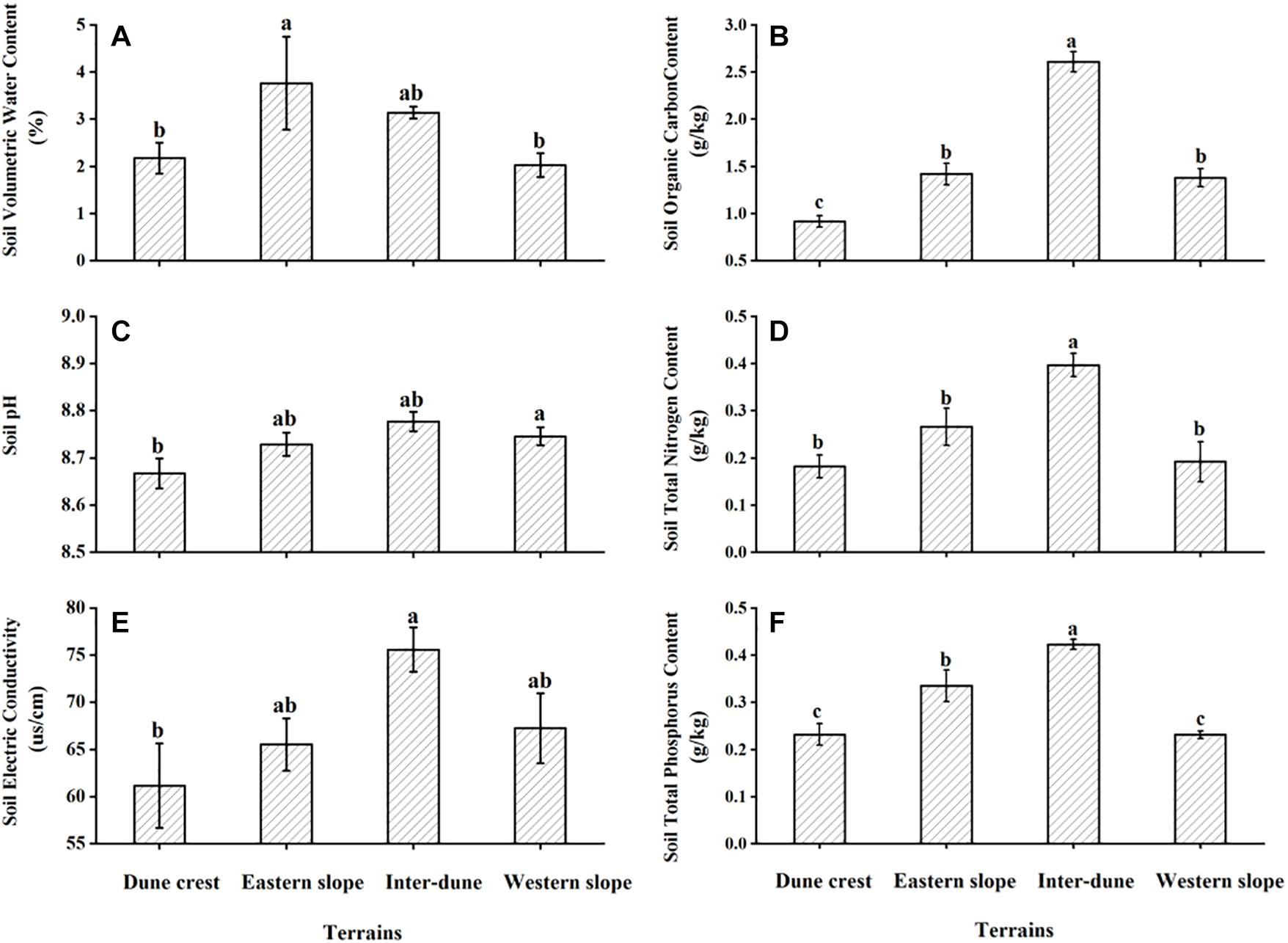
Figure 2. Soil physical (A) and chemical properties (B–F) of different terrains. Different letters represent significant differences (p < 0.05). Data are presented as the mean ± 1 standard error.
Among the 336 sampling plots, there were 112 sampling plots located on the dune crest and the inter-dune lowland, respectively, and 56 sampling plots located on the eastern and western slope, respectively. In this study, a total of 2,114 plants were investigated, including 1,131 H. ammodendron individuals and 983 H. persicum individuals. There were significant differences in stand density of H. ammodendron among different terrains (F = 27.90, p < 0.05), which were in the order of inter-dune lowland > western slope > eastern slope > dune crest. The density of H. ammodendron growing in the inter-dune lowland was the highest (8.12 ± 1.04 plants/100 m2) (Figure 3), most of them were seedlings and young individuals (Figure 4). Different from H. ammodendron, the densities of H. persicum were in the order of dune crest > western slope > eastern slope > inter-dune lowland. Only the density difference of H. persicum between the dune crest and inter-dune lowland was significant (p < 0.05), there was no difference among the other terrains, and the young tree always occupied a larger proportion at each terrain (Figure 4).
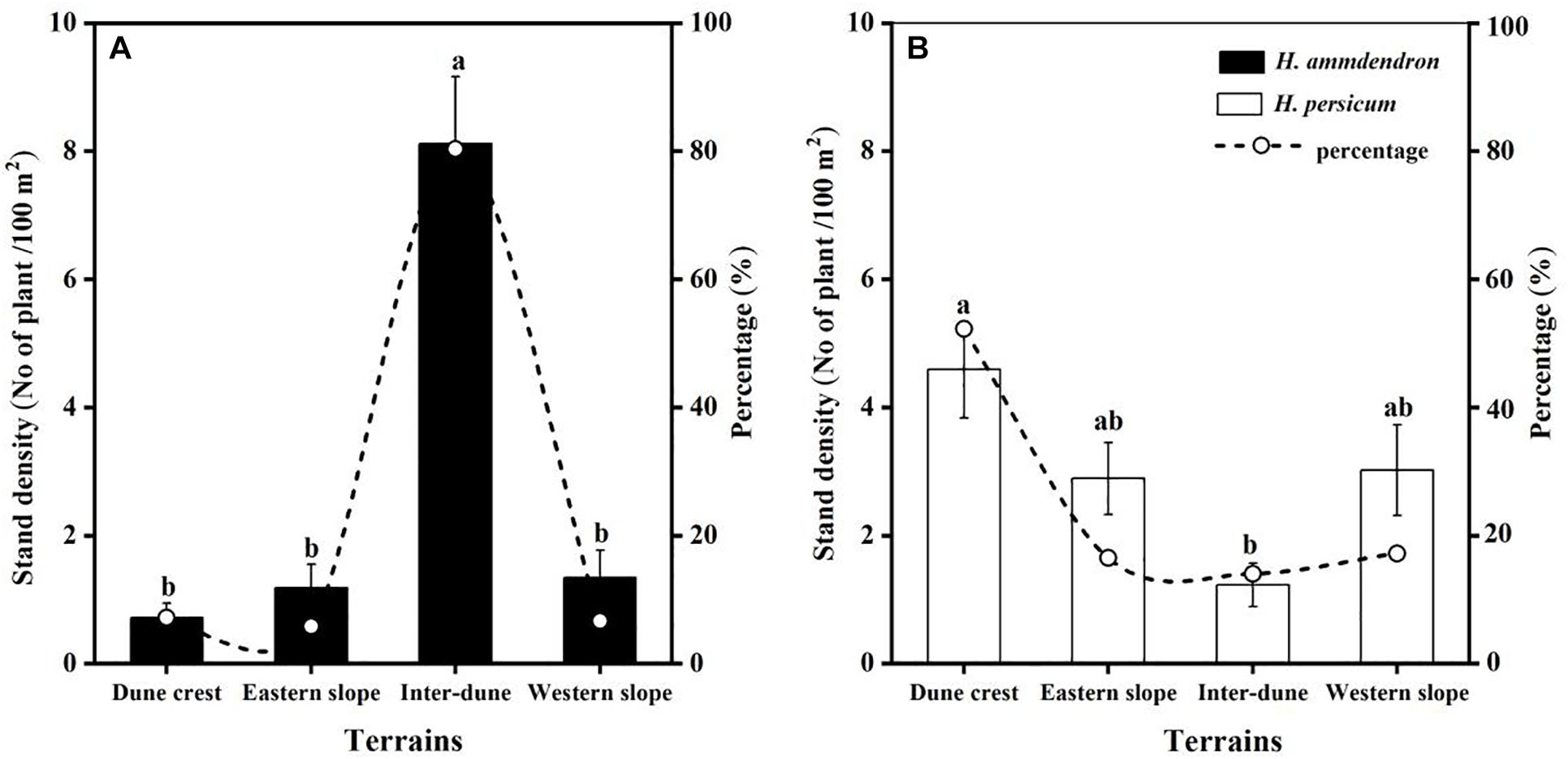
Figure 3. Quantitative characteristic of Haloxylon ammodendron (A) and H. persicum (B) at different terrains. Different letters represent significant differences (p < 0.05). Data are presented as the mean ± 1 standard error.
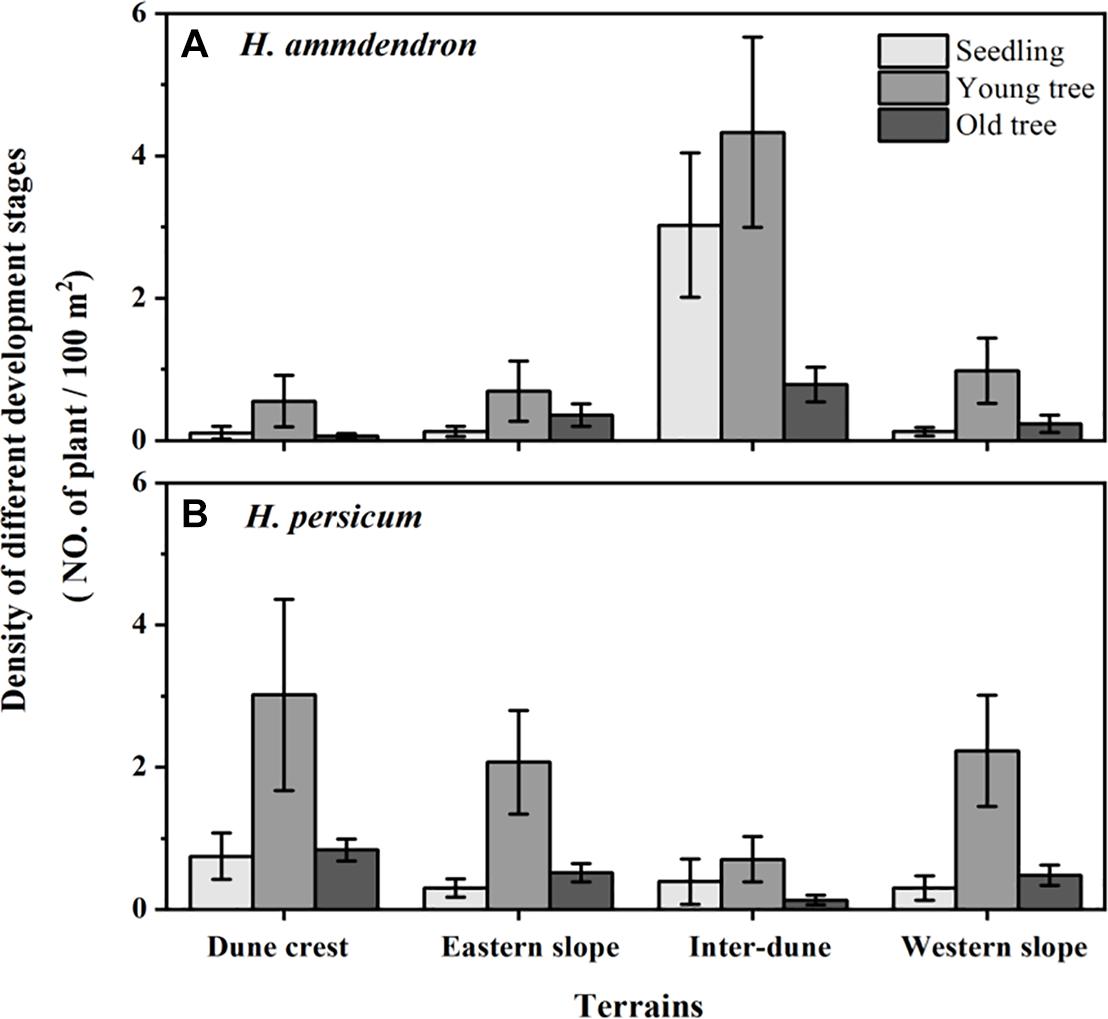
Figure 4. Density of Haloxylon ammodendron (A) and H. persicum (B) of different development stage at different terrains. Data are presented as the mean ± 1 standard error.
As shown in Figure 5, the patterns of plant morphological characteristics were consistent for H. ammodendron and H. persicum under different terrain conditions, both were in the order of eastern slope > western slope > dune crest > inter-dune lowland. The plant height, basal diameter, and canopy radius of H. ammodendron all showed significant differences among various terrains (F = 34.18, p < 0.05; F = 12.85, p < 0.05; F = 13.76, p < 0.05). The minimum values all appeared in the inter-dune lowland, which were 0.81 ± 0.02, 0.03 ± 0.001, and 0.37 ± 0.01 m, respectively. The plant height and canopy radius of H. persicum showed significant differences among various terrains (F = 11.40, p < 0.05; F = 3.89, p < 0.05); the maximum values (1.63 ± 0.07 and 0.53 ± 0.03 m) were observed on the eastern slope, while the minimum values (1.09 ± 0.08 and 0.38 ± 0.03 m) were in the inter-dune lowland. The basal diameter exhibited no significant differences (F = 1.58, p > 0.05).
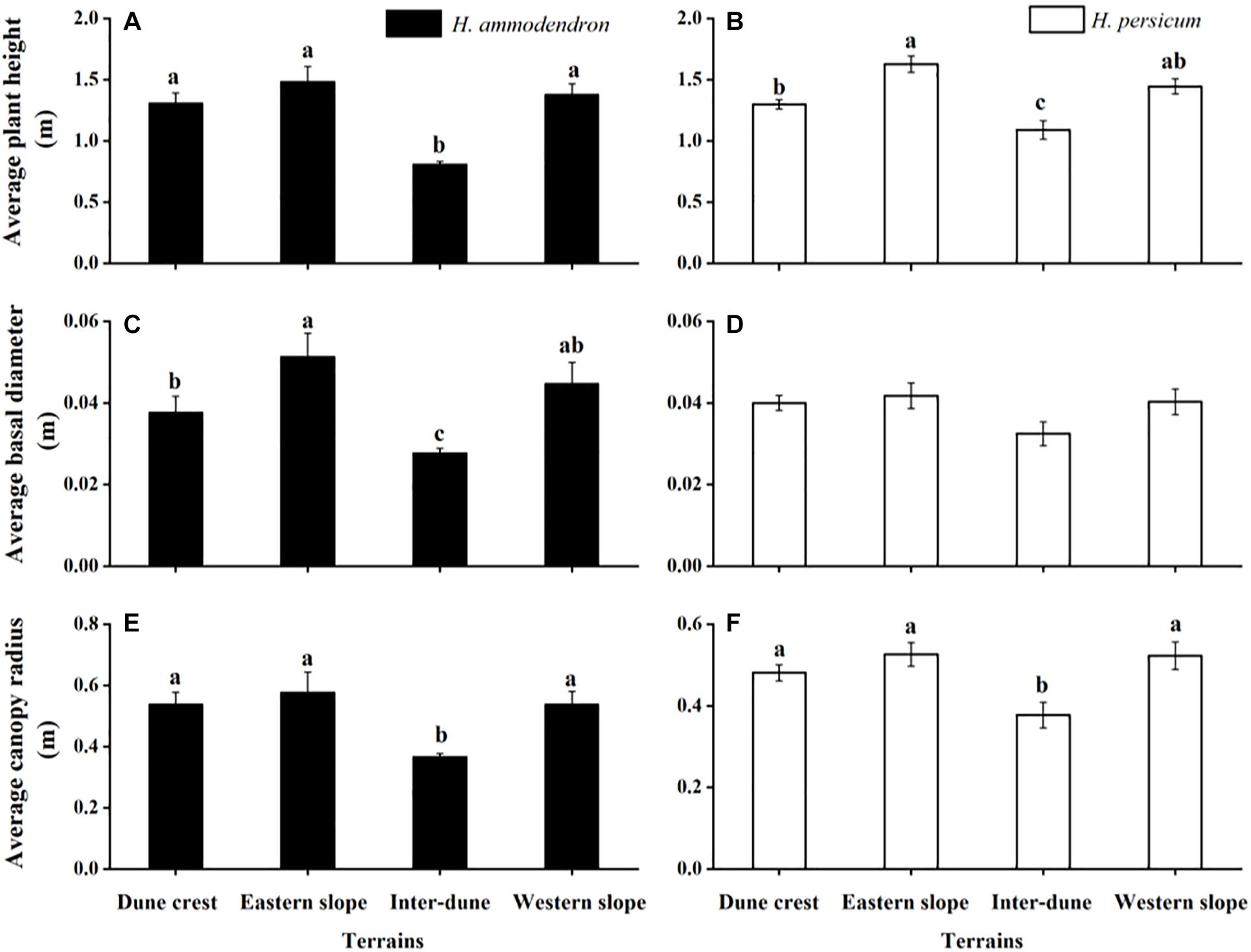
Figure 5. Morphological characteristics of Haloxylon ammodendron (A,C,E) and H. persicum (B,D,F) at different terrains. Different letters represent significant differences (p < 0.05). Data are presented as the mean ± 1 standard error.
Supplementary Figure 1 and Table 2 showed the scatter plot and the quantitative relationship of logarithm-transformed plant height (H) and canopy radius (R) of H. ammodendron and H. persicum. The log(H)∼log(R) linear regression analysis resulted in the coefficient of determination (R2) was greater than 0.7 (p < 0.05), indicating a significant positive correlation between H and R of H. ammodendron and H. persicum under different terrain conditions. Isometric tests showed that only the relationship between the H and R of H. ammodendron on the dune crest and inter-dune lowland, and H. persicum in the inter-dune lowland were isometric (F = 3.64, p = 0.06; F = 5.05, p = 0.08; F = 0.01, p = 0.93), the relationships among other terrains were allometric (p < 0.05). In addition, there was a significant difference in the allometric index (namely the slope) between H and R for H. ammodendron and H. persicum among various terrain conditions (F = 16.07, p < 0.05; F = 19.23, p < 0.05). The allometric indices between H and R of H. ammodendron were 1.10, 1.21, 1.03, and 1.20 on the dune crest, eastern slope, inter-dune lowland, and western slope, respectively; while those of H. persicum were 1.22, 1.15, 1.00, and 1.15, respectively. The smallest allometric indices between H and R for H. ammodendron and H. persicum (1.03 and 1.00) both appeared in the inter-dune lowland, and there was no significant difference among other terrains (p > 0.05).
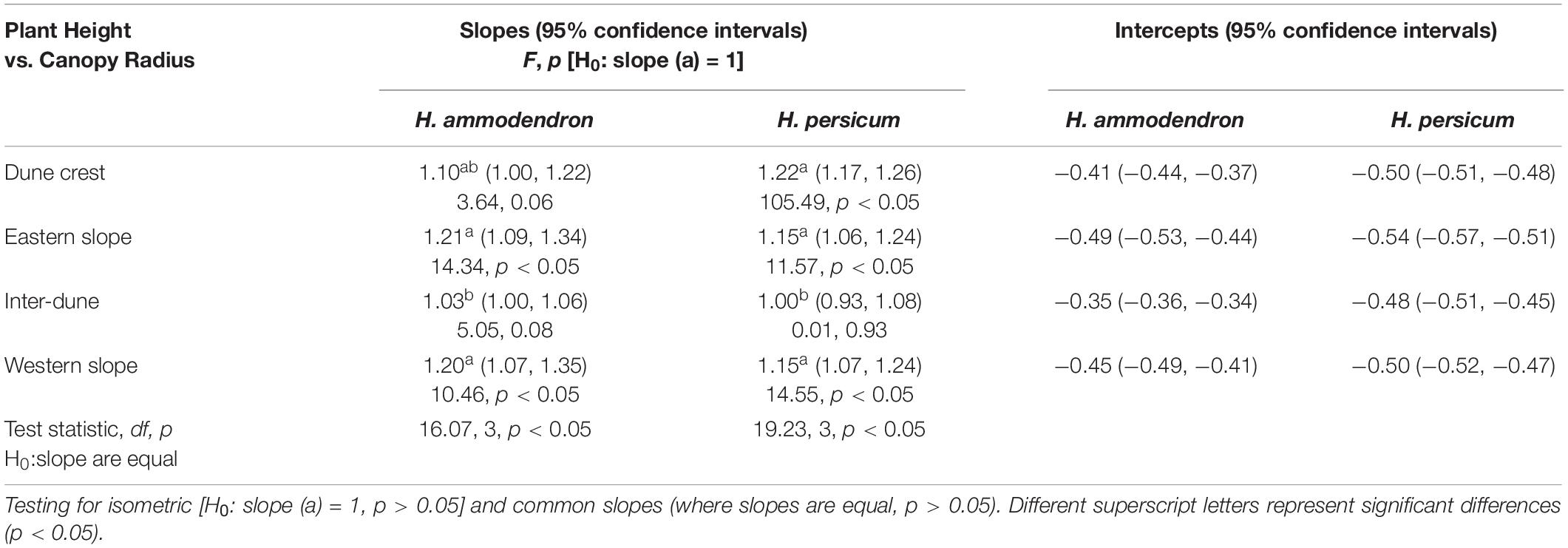
Table 2. Results for SMA fitted slopes between plant height and canopy radius of Haloxylon ammodendron and H. persicum at different terrains.
Similar to the relationship between H and R, the log(H)∼log(D) linear regression analysis indicated a significant positive correlation between the H and D of H. ammodendron and H. persicum under various terrain conditions (p < 0.05). Isometric tests showed that the relationship between H and D of H. ammodendron and H. persicum under various terrain conditions was allometric (p < 0.05), and the allometric indices were significantly different (F = 12.15, p < 0.05; F = 15.01, p < 0.05). The allometric indices between H and D of H. ammodendron growing on the dune crest, eastern slope, inter-dune lowland, and western slope were 1.30, 1.12, 1.14, and 1.39, respectively, whereas those of H. persicum were 1.30, 1.38, 1.15, and 1.15, respectively. The largest allometric indices between H and D for H. ammodendron and H. persicum (1.39 and 1.38) appeared in the western and eastern slope, respectively.
In the current study, we found that H. ammodendron and H. persicum were distributed at all terrains, but the dominant distribution areas of the two species were different. Therefore, the statement “Haloxylon persicum grows on sand dunes while H. ammodendron grows on inter-dune lowlands” is inaccurate (Xu et al., 2014). As shown in Figure 3, H. ammodendron was concentrated in the inter-dune lowland, accounting for more than 80% of the total, with a density of 8.12 ± 1.04 plants/100 m2 (Figure 3). However, H. persicum was least distributed in the inter-dune lowland and mostly distributed on the dune crest (accounting for about 50% of the total), with a density of 4.59 ± 0.76 plants/100 m2 (Figure 3). The stand density of H. persicum on the eastern and western slopes were similar, which were both greater than those of H. ammodendron. These results indicate that the distribution patterns of H. ammodendron and H. persicum are significantly different, the distribution of H. persicum is more scattered comparing to H. ammodendron.
The difference in the distribution patterns of H. ammodendron and H. persicum among various terrains is closely related to soil physical and chemical properties of their habitats. Soil is the growth substrate for terrestrial plants, and it is the results of climate, biology, parent material, topography, and time, and is also profoundly affected by modern anthropogenic activities (Weil and Brady, 2016). In desert region, aeolian transport of soil particles greatly affects the spatial distribution of resources, which in turn, exerts significant influence on the growth and recruitment of desert plants (Alvarez et al., 2012; Niu et al., 2021). Studies have shown that the inter-dune lowland and the middle-lower part of the dune in the Gurbantonggut Desert are mainly composed of fixed aeolian sandy soil. In contrast, the middle-upper part and the crest of the dune are mainly composed of semi-fixed aeolian sandy soil (Wang et al., 2003; Zhu et al., 2021). The soil pH, electrical conductivity, water content, and nutrient content of the inter-dune lowland were significantly higher than those of dune crest, as shown in Figure 2, which was in line with other studies (Li C. J. et al., 2011; Xu et al., 2014). To better adapt to the significantly different soil conditions, H. ammodendron and H. persicum developed different adaptabilities. As proved by habitat soil substrate exchange experiments, H. persicum had stronger adaptability and settlement ability on the nutrient-poor dune crest due to its excellent resistance to salt and alkalinity, whereas H. ammodendron was more suitable for the inter-dune lowland with a higher soil salt content (Li H. et al., 2011). In addition, sand burial caused by aeolian soil is another important factor controlling the distribution and establishment of desert plant communities (Jia et al., 2011). Previous studies have showed that the ability of H. ammodendron to resist wind erosion was weak, but the ability of H. persicum to tolerate sand burial and grow in sandy environments was stronger (Hu, 1984; Li H. et al., 2011). Therefore, the differences in soil characteristic of micro-habitats among various terrains and the consequent differences in biological adaptability of the plants together determine the general distribution patterns of H. ammodendron and H. persicum (Li H. et al., 2011; Xu et al., 2014), and the current study demonstrated that their difference in distribution is not absolute (Figure 3).
As previously demonstrated, soil physical and chemical properties determine the distribution patterns of plants, meanwhile, they play important roles in the morphogenesis of plants, especially the soil water supply conditions (Li et al., 2005; Niu et al., 2021). This effect is more prominent for desert plants in arid regions where water supply is limited. Figure 5 showed that the plant height, canopy radius, and basal diameter of H. ammodendron and H. persicum in the inter-dune lowland were all the smallest, while those on the eastern slope were the largest. This phenomenon may result from the special soil characteristics of inter-dune lowland, including poorly sorted soil sediments, well-developed herb layers which consisting of a large number of ephemeral, ephemeroid and annual plants (as shown in Figure 1B), as well as diverse biological crusts covering the soil which could form an effective permeability barrier: the above factors finally resulted in higher water content in the shallow soil layers (Wang et al., 2003). The superior water and nutrient supplies of the shallow soil layers of the inter-dune lowland (Figure 2) provided suitable conditions for the seed germination of H. ammodendron, and triggered a boom of new seedlings (Figures 3, 4). However, due to the limited growth time of newly emerged seedlings, these individuals generally owned smaller height, canopy radius, and basal diameter (Figure 5). On account of the large quantity of new seedling and young tree (Figure 4), the individual morphology of H. ammodendron in the inter-dune lowland exhibited a relatively lower stature. On the other hand, the root depth of new seedling and young tree is limited, mainly concentrating at the range of 0–100 cm soil layer (Dan et al., 2007; Huang and Guo, 2009), therefore they principally depended on shallow soil water to survive (Li et al., 2020). Although the overall water status of the inter-dune habitat was relatively better, the water resources competition was fierce considering the large number of H. ammodendron seedlings and other desert plants. Under this circumstance, water resources became precious and scarce, which could greatly restricting the survival and development of H. ammodendron population in subsequent stages. As the drought intensified, most of the H. ammodendron seedlings died (personal observations). Different from H. ammodendron, the small individual morphology of H. persicum exhibited in the inter-dune lowland may be related to its poor adaptability to high salinity, which limiting its growth and development in this habitat (Lu and Ming, 2012).
The eastern slope of the study area is steep and leeward, with short sunshine time and smaller evaporation, thus the soil water condition is better than the other terrains except the inter-dune lowland (Figure 2). During the field survey, we found relatively few H. ammodendron and H. persicum seedlings appearing on the eastern slope, instead, there were more big trees existing (Figure 4). These big individuals were usually the ones that surviving from adversities, such as drought and cold, they have developed huge and deep root systems to cope with environmental stresses. According to previous study, the root depth of H. ammodendron and H. persicum could be up to 13.5 and 18 m, respectively (Sheng et al., 2004; Xu et al., 2017). This allowed them to absorb water from deep soil layers for survival and to be more adaptive to environmental changes (Wu et al., 2019a,b). Therefore, they are advantaged in plant height, canopy radius, and basal diameter (Figure 5). In turn, these advantages help them to take up more light, water and nutrients for life activities, and to occupy an advantageous niche in the competition.
The morphogenesis of desert trees and shrubs, namely the horizontal expansion (canopy and stem) and vertical growth (height), highlights the coordinative relationship between individual plant growth and environment (Archibald and Bond, 2003; Li S. et al., 2011). In this study, the canopy radius was used to represent the canopy size, so that the characterizing of the allometric relationship was more direct and clear. The results showed that both H. ammodendron and H. persicum preferred to adopt allometric patterns under various terrain conditions, as the allometric indices between H and R, and H and D were all greater than 1 (Tables 2, 3), which suggested that the relative growth rate of the canopy radius and basal diameter both surpassed that of plant height. Previous drought treatment study has shown that between above- and below-ground, H. ammdendron preferentially allocated biomass to the below-ground part (namely roots) along a fixed allometric trajectory, which could benefit the survival and the post-drought recovery of the plants (Zhang et al., 2016). The current study further revealed that H. ammodendron also adopted a special allometric strategy for above-ground parts: to allocate more resources to horizontal expansion rather than vertical growth.
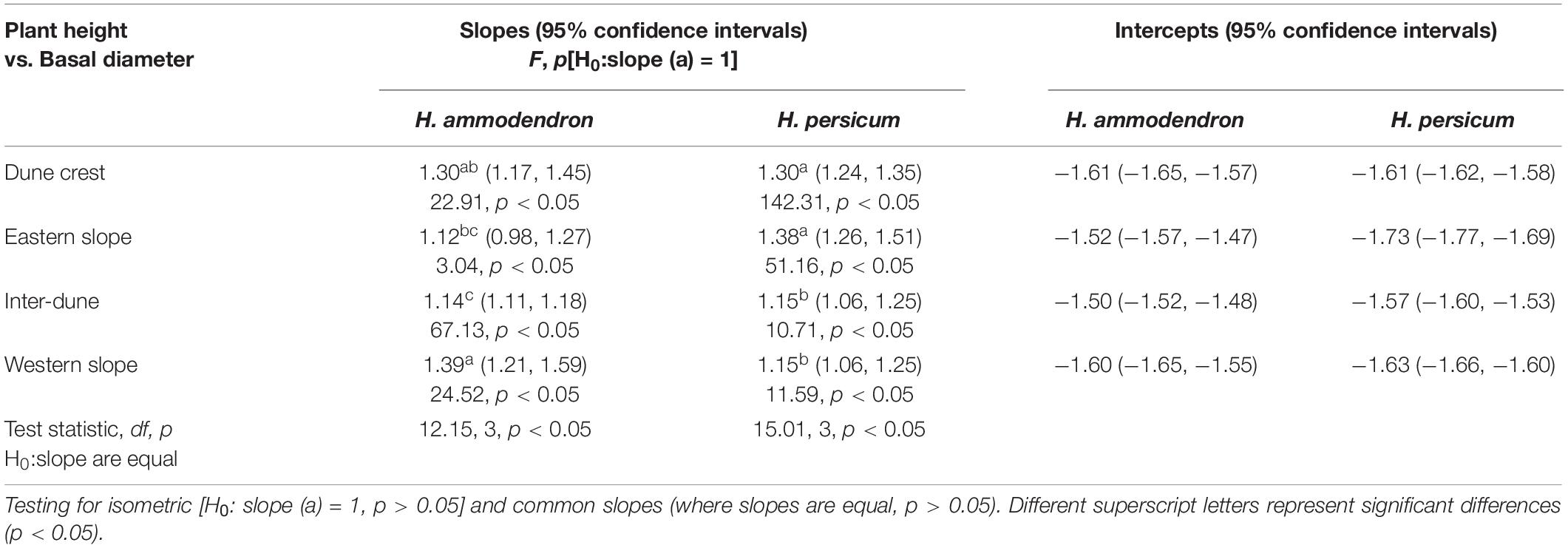
Table 3. Results for SMA fitted slopes between plant height and basal diameter of Haloxylon ammodendron and H. persicum at different terrains.
The allometric relationship between H and R of studied species might resulted from the increased xylem water transport resistance: as the plant height increases would result in the decrease of water transported to the top of plant. In response, plants lowered the stomatal conductance through the physiological adjustment to reduce water loss, meanwhile, the absorption of CO2 as the raw material for photosynthesis was inevitably affected. This led to a decreased ability of plants to assimilate carbon, which ultimately restricted the heights of plants (Shi et al., 2015). This has been confirmed in the study of Zhang et al. (2009), in which they showed that the number of dead branches and the individual mortality rate increased as the height of a tree increased in a savanna, because along with this process, their water transport system and water physiology developed into the direction that were more prone to water shortage. In addition, the study area is a typical desert ecosystem with abundant light resource and sparse plant cover, the above-ground parts of vegetation distribute intermittently, thus unlike some dense forest ecosystems, light is not an important limiting factor in the intraspecific and interspecific competition (Li S. et al., 2011). This might be the other reason why the growth of plant height became slow: they don’t need to grow higher to get light. Similarly, the trade-off allocation of resources reflected in the allometric relationship between H and D of studied species was also an indicator of plant response to numerous selection pressures and constraints. Therefore, the allometric relationships between H and R, H and D could be regarded as an important feature of plants adaptability to heterogeneous environments, which enables them to optimally allocate various resources to different parts and achieve maximum benefit for survival and growth (Jacob, 2004; Nicotra et al., 2010; Hou et al., 2014; Turcotte and Levine, 2016).
Within a limited spatial scale, topography indirectly affects plant growth dynamics and species distribution through redistribution of soil water, nutrients and light resources (Zhao et al., 2019; Tang et al., 2020). As shown in Tables 2, 3, the allometric indices between H and R, H and D of two Haloxylon species were significantly different among various terrains, this might own to the habitat heterogeneity at different terrains, and also might be related to the ecological adaptability of two studied species in heterogeneous habitats. In the study area, western wind prevails all year around, the wind speed gradually increases from the bottom of the dune along the slope, and reaches the maximum on the dune crest (Wang et al., 2003). The eastern, western slope and the dune crest habitats have a windy environment with complicated water conditions (Figure 2). The allometric indices between H and R of studied species on these three terrains were greater than that of inter-dune lowland (Table 2), suggesting that H. ammodendron and H. persicum preferentially invested more photosynthetic products into the canopy growth instead of plant height. This might because taller plants face unnecessary pressures in a windy environment, like being broken off. However, in the inter-dune lowland with less wind, most of individuals were seedlings that did not reached their growth limit (Figure 4), they adopted an isometric allocating mode between H and R (Table 2). As an important link of above- and below-ground part, the basal stem of plant connects the trunk upward and the root downward. Although the overall allometric relationship between H and D confirmed that studied species preferentially meet the requirements of water and nutrient transport as well as mechanical support (Brüchert and Gardiner, 2006; Shi et al., 2015), the terrain had a significant effect on it, and the effect was quite different between these two species. According to earlier studies, the basal diameter would increase when plants were mechanically stimulated (Goodman and Ennos, 1996), and stems with a larger basal diameter could transport water under a low tension of xylem vessels (Zhao et al., 2013). As shown in Table 3, the largest allometric index between H and D (1.39) for H. ammodendron appeared in western slope, where was windward and water-limited (Figure 2). It suggested that H. ammodendron was inclined to invest more into the basal diameter growth, so as to resist the mechanical stimulation caused by long-term unidirectional wind and to maintain a high water transport capacity. In contrast, H. persicum growing on the eastern slope, where the water supply is relatively sufficient (Figure 2), promoted investment into its basal diameter, as confirmed by the largest allometric index between H and D (1.38). A larger basal diameter corresponded to a bigger cross-sectional area of the xylem, meaning a higher water and nutrient transport capacity, large stem could help plants to access to resources efficiently. The differences in the allometric relationship between H and R, H and D of H. ammodendron and H. persicum among various terrain conditions indicated their ecological adaptability to heterogeneous environments, and also revealed the diversification of plants to adapt to various environments.
In the current study, through investigating two dominant species in the Gurbantunggut Desert of Central Asia, we found that the above-ground parts of Haloxylon ammodendron and H. persicum both adopted a fixed allometric pattern, as they preferentially invested into plant canopy radius and basal stem instead of plant height. This indicated that the above-ground parts of desert plants were more in favor of horizontal expansion rather than vertical growth, which might be closely related to the environmental characteristics of resource-deficient but light-rich. The stand densities and individual morphology of the two Haloxylon species exhibited distinct differences among different terrains, suggesting that the topography could significantly affect the plant growth dynamics. In addition, the allometric indices between H and R, H and D of studied species also significantly varied among terrains although they along a fixed allometric trajectory, indicating that they could finely control the investment in main functional traits in changing environment. These findings contributed to the understanding of the adaptation strategies employed by desert plants, and how changes in terrains within in a limited spatial scale influenced them.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
YL conceived the study and critically reviewed and edited the manuscript. XW, X-JZ, and X-HM conducted the experiments. XW analyzed the data and drafted the manuscript. All authors have contributed to the study conceptualization and read and approved the final version of this manuscript.
This work was financially supported by the Natural Sciences Foundation of Xinjiang Uygur Autonomous Region (No. 2020D01C053) and National Natural Sciences Foundation of China (Nos. 41730638 and 32001145).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The reviewers DZ and LT declared a shared affiliation, with no collaboration, with several of the authors X-JZ and X-HM to the handling editor at the time of the review.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
We would like to thank all the staff at the Fukang Station of Desert Ecology for their excellent assistance. We would especially like to thank Ren Wen, Chern, Zhang Yan-Yan, Yin Xin-wei, Yue Yue-meng, and Li Rui-Xi for their help in the laboratory analysis and manuscript writing.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2021.754887/full#supplementary-material
Allen, A. P., Pockman, W. T., Restrepo, C., and Milne, B. T. (2008). Allometry, growth and population regulation of the desert shrub Larrea tridentata. Funct. Ecol. 22, 197–204. doi: 10.1111/j.1365-2435.2007.01376.x
Alvarez, L. J., Epstein, H. E., Li, J. R., and Okin, G. S. (2012). Aeolian process effects on vegetation communities in an arid grassland ecosystem. Ecol. Evol. 2, 809–821. doi: 10.1002/ece3.205
Archibald, S., and Bond, W. J. (2003). Growing tall vs growing wide, tree architecture and allometry of Acacia karroo in forest, savanna, and arid environments. Oikos 102, 3–14.
Brüchert, F., and Gardiner, B. (2006). The effect of wind exposure on the tree aerial architecture and biomechanics of Sitka spruce (Picea Sitchensis, Pinaceae). Am. J. Botnay 93, 1512–1521. doi: 10.3732/ajb.93.10.1512
Buras, A., Wucherer, W., Zerbe, S., Noviskiy, Z., Muchitdinov, N., Shimshikov, S., et al. (2012). Allometric variability of Haloxylon species in central Asia. Forest Ecol. Manag. 274, 1–9.
Chen, G. P., Yang, K. T., Wang, L., Wang, F., Cao, X. W., and Chen, L. S. (2020). Allometric relations for biomass partitioning of seven alpine Rhododendron species in South of Gansu. Chinese J. Plant Ecol. 44, 1040–1049. doi: 10.17521/cjpe.2020.0119
Chen, D. L., Ma, Y. Z., Zhong, Q. L., and Xu, W. F. (2013). Allocation of gross primary production in forest ecosystems, allometric constraints and environmental responses. New Phytol. 200, 1176–1186. doi: 10.1111/nph.12426
Cheng, D., Ma, Y., Zhong, Q., and Xu, W. (2014). Allometric scaling relationship between above- and below- ground biomass within and across five woody seedings. Ecol. Evol. 4, 3968–3977. doi: 10.1002/ece3.1184
Dan, L. S., Zhang, X. M., Hua, Y. H., Wei, J., Yan, H. L., and Xie, T. T. (2007). Response of root distribution of Haloxylon ammodenderon seedings to iirrigation amounts in the taklimakan desert, China. J. Plant Ecol. 31, 769–776.
Dang, J. J., Zhao, C. Z., Li, Y., Hou, Z. J., and Dong, X. G. (2015). Relationship between leaf traits of Melica przewalskyi and slope aspects in alpine grassland of Qilian Mountains, China. Chinese J. Plant Ecol. 39, 23–31. doi: 10.17521/cjpe.2015.0003
Goodman, A. M., and Ennos, A. R. (1996). A comparative study of the response of the roots and shoots of sunflower and maize to mechanical stimulation. J. Exp. Bot. 47, 1499–1507. doi: 10.1093/jxb/47.10.1499
Han, W. X., and Fang, J. Y. (2008). Review on the mechanism models of allometric scaling laws:3/4 VS. 2/3Power. Chinses J. Plant Ecol. 32, 951–960.
Hou, Z. J., Zhao, C. Z., Li, W., Zhang, Q., and Ma, X. L. (2014). Trade-off between height and branch numbers in Stellera chamaejasme on slopes of different aspects in a degraded alpine grassland. Chinese J. Plant Ecol. 38, 281–288. doi: 10.3724/sp.j.1258.2014.00025
Hu, W. K. (1984). The type, characteristics and dynamics of Haloxylon ammodendron desert in the Southern Junggar Basin. Arid Zone Res. 2, 28–38.
Huang, Y., and Guo, Y. H. (2009). Root distribution characteristics of Haloxylon ammodendron (Mey.) Bunge Plantation. Acta Agrestia Sinica 17, 84–87.
Jacob, W. (2004). Allocation, plasticity and allometry in plants. Perspect. Plant Ecol. Evol. Systemati. 6, 207–215. doi: 10.1078/1433-8319-00083
Jennifer, A. R., Alesia, H., Stephanie, R. B., Lauren, B., Kristofer, M. H., Marcy, E. L., et al. (2019). Sensitivity of dryland plant allometry to climate. Funct. Ecol. 33, 2290–2303. doi: 10.1111/1365-2435.13463
Jia, X. H., Li, X. R., Li, Y. S., and Zhou, Y. Y. (2011). Responses of Nitraria tangutorum population characteristics to sand burial at Southeastern Fringe of Tengger Desert. Chinese J. Ecol. 30, 1851–1857.
Li, C. J., Ma, J., Li, Y., and Li, H. (2011). Nutrient gradient distribution in soil around taproots of Haloxylon ammodendron and Haloxylon persicum. J. Desert Res. 31, 1174–1180.
Li, H., Li, Y., and Fan, L. L. (2011). Response of seedling emergence of two Haloxylon species to the interchanging of soil matrix in habitants and sand burial depth. Arid Zone Res. 28, 780–788.
Li, S., Zheng, X. J., Tang, L. S., and Li, Y. (2011). Morphological investigation of desert shrubs of China’s junggar basin based on allometric theory. Chinese J. Plant Ecol. 35, 471–479.
Li, T., Peng, L. P., Shi, Q. D., Marhaba, N., and Dai, Y. (2020). Water sources of Haloxylon ammodendron and H. Persicum of different diameter classes in Junggar Basin, Xinjiang. Acta Ecol. Sinica 40, 2099–2110.
Li, Y., Zhao, C. Z., Dong, X. G., Hou, Z. J., Ma, X. L., and Zhang, Q. (2013). Twig and leaf trait differences in Stellera chamaejasme with slope in alpine grassland. Chinese J. Plant Ecol. 37, 709–717. doi: 10.3724/sp.j.1258.2013.00074
Li, Z. B., Li, P. H., Wang, Q., and Xu, L. (2013). Allometric characteristics of terminal twigs of Tamarix ramosissima and Haloxylon ammodendron. Bull. Botan. Res. 33, 274–281.
Li, L., Zhang, X. M., and He, X. Y. (2005). Effects of salinity and water potential on seed germination and radicle growth of riparian Populus euphratica. Arid Zone Res. 4, 104–109.
Lines, E. R., Zavala, M. A., Purves, D. W., and Coomes, D. A. (2012). Predictable changes in aboveground allometry of trees along gradients of temperature, aridity and competition. Global Ecol. Biogeography 21, 1017–1028.
Liu, X. J., and Ma, K. P. (2015). Plant functional traits-concepts, applications and future directions. Sci. Snica 45, 325–339. doi: 10.1360/n052014-00244
Lu, H. Y., and Ming, Z. Y. (2012). Response of growth and biomass allocation of Haloxylon persicum seedlings to different salt treatments. Arid Zone Res. 29, 194–202.
Lu, X. M., Zhou, C. F., An, S. Q., Fang, C., Zhao, W., Yang, Q., et al. (2007). Phenotypic plasticity, allometry and invasiveness of plants. Chinese J. Ecol. 26, 1438–1444.
Midgley, J., and Bond, W. (1989). Leaf size and inflorescence size may be allometrically related traits. Oecologia 78, 427–429. doi: 10.1007/bf00379120
Nicotra, A. B., Atkin, O. K., Bonser, S. P., Davidson, A. M., Finnegan, E. J., Mathesius, U., et al. (2010). Plant phenotypic plasticity in a changing climate. Trends Plant Sci. 15, 684–692. doi: 10.1016/j.tplants.2010.09.008
Niu, F. R., Pierce, N. A., Archer, S. R., and Okin, G. S. (2021). Germination and early establishment of dryland grasses and shrubs on intact and wind-eroded soils under greenhouse conditions. Plant Soil 465, 245–260. doi: 10.1007/s11104-021-05005-9
Noriyuki, O. (2011). Height-dependent changes in shoot structure and tree allometry in relation to maximum height in four deciduous tree species. Funct. Ecol. 25, 777–786.
Ogden, J. (1974). The reproductive strategy of higher plants: ii. the reproductive strategy of Tussilago Farfara L. J. Ecol. 62, 291–324. doi: 10.2307/2258894
Primack, R. B. (1987). Relationships among flowers, fruits, and seeds. Ann. Rev. Ecol. Systemat. 18, 409–430. doi: 10.1146/annurev.es.18.110187.002205
Pyankov, V. I., Black, C. C., Artyusheva, E. G., Elena, V. V., Maurice, S. B. K., and Gerald, E. E. (1999). Features of photosynthesis in Haloxylon species of Chenopodiaceae that are dominant plants in central Asian Deserts. Plant Cell Physiol. 40, 125–134. doi: 10.1007/BF00346994
Sheng, J. H., Qiao, Y. X., Liu, H. Y., Zhai, Z. X., and Guo, Y. H. (2004). A Study on the root system of Haloxylon ammodendron (C. A. Mey.) Bunge. Acta Agrestia Sinica 12, 91–94.
Shi, Y. C., Zhao, C. Z., Song, Q. H., Du, J., and Wang, J. W. (2015). Allometric relationship between height and crown width or diameter of Platycladus orientalis on different slope aspects of lanzhou Northern Mountains. Chinese J. Ecol. 34, 1879–1885.
Smith, J. M. B., Burian, R., Kauffman, S., Alberch, P., Campbell, J., Goodwin, B., et al. (1985). Developmental constraints and evolution: a perspective from the mountain lake conference on development and evolution. Quar. Rev. Biol. 60, 265–287. doi: 10.1086/414425
Sun, J., Xia, J. B., Dong, B. T., Gao, F. L., Chen, P., Zhao, W. L., et al. (2021). Root morphology and growth characteristics of Tamarix chinensis with different densities on the beach of the Yellow River Delta. Acta Ecol. Sinica 41, 3775–3783.
Tang, Z. H., Yu, Q. S., Liu, H. J., Jiang, S. X., He, F. L., Zhang, Y. H., et al. (2020). Characteristics of alpine vegetation community and its relationship to topographic climate factors in the eastern Qilian mountain. Acta Ecol. Sinica 40, 223–232.
Turcotte, M. M., and Levine, J. M. (2016). Phenotypic plasticity and species coexistence. Trends Ecol. Evol. 31, 803–813. doi: 10.1016/j.tree.2016.07.013
Wang, L. J., Sun, D. Y., Zhao, C. Y., Li, J. Y., and Sheng, Y. (2011). Plant architecture characteristics of Haloxylon ammodendron and Haloxylon persicum in Zhungar Basin. Acta Ecol. Sinica 31, 4952–4960.
Wang, X. Q., Li, B. W., and Zhang, Y. M. (2003). Stabilization of dune surface and formation of mobile belt at the top of longitudinal dunes in Gurbantunggut Desert, Xinjiang, China. J. Desert Res. 23, 126–131.
Weil, R. R., and Brady, N. C. (2016). The Nature and Properties of Soils, 15th Edn. London: Pearson Education Inc.
Weiner, J. (2004). Allocation, plasticity and allometry in plants. Perspect. Plant Ecol. Evol. Systemat. 6, 207–215.
West, G. B., Brown, J. H., and Enquist, B. J. (1997). A general model for the origin of allometric scaling laws in biology. Science 276, 122–126. doi: 10.1126/science.276.5309.122
Wu, X., Zheng, X.-J., Li, Y., and Xu, G. Q. (2019a). Varying responses of two Haloxylon species to extreme drought and groundwater depth. Environ. Exp. Bot. 158, 63–72.
Wu, X., Zheng, X. J., Yin, X. W., Yue, Y. M., Liu, R., Xu, G. Q., et al. (2019b). Seasonal variation in groundwater dependency of two domaint woody species in the desert region of Central Aisa. Plant Soil 444, 39–55. doi: 10.1007/s11104-019-04251-2
Xu, G. Q., Yu, D. D., and Li, Y. (2017). Patterns of biomass allocation in Haloxylon persicum woodlands and their understory herbaceous layer along a groundwater depth gradient. Forest Ecol. Manag. 395, 37–47. doi: 10.1016/j.foreco.2017.03.037
Xu, G. Q., Yu, D. D., Xie, J. B., Tang, L. S., and Li, Y. (2014). What makes Haloxylon persicum grow on sand dunes while H. Ammodendron grow on interdune lowlands: a proof from reciprocal transplant experiments. J. Arid Land 6, 581–591.
Zhang, Q. Y., and Peng, S. L. (2018). Effects of warming on the biomass allocation and allometric growth of the invasive shrub Lantana camara. Acta Ecol. Sinica 38, 6670–6676.
Zhang, Y., Li, Y., and Xie, J. B. (2016). Fixed allocation patterns, rather than plasticity, benefit recruitment and recovery from drought in seedlings of a desert shrub. Ann. Bot. 8:lw020. doi: 10.1093/aobpla/plw020
Zhang, Y. J., Meinzer, F. C., Hao, G. Y., Scholz, F. G., Bucci, S. J., Takahashi, F. S. C., et al. (2009). Size-dependent mortality in a neotropical savanna tree: the role of height-related adjustments in hydraulic architecture and carbon allocation. Plant Cell Environ. 32, 1456–1466. doi: 10.1111/j.1365-3040.2009.02012.x
Zhao, P., Qu, J. J., Xu, X. Y., Tang, J. N., Han, Q. J., Xie, S. B., et al. (2019). Study on the characteristics of sandy alpine grasslands and its relationship between plant distribution and microtopography in the source regions of Yangtze River. Acta Ecol. Sinica 39, 1030–1040.
Zhao, P., Sun, G. C., Ni, G. Y., and Zeng, X. P. (2013). Seasonal differences in the leaf hydraulic conductance of mature Acacia mangium in response to its leaf water use and photosynthesis. Chinese J. Appl. Ecol. 24, 49–56.
Zheng, W., Fan, G. H., Huang, Y. X., Wang, T., Yu, P. J., and Wang, H. Q. (2021). Allometric relationships between the morphological traits and biomass allocation strategies of Salsola collina under different population density. Acta Ecol. Sinica 41, 2845–2854.
Keywords: plant height, canopy radius, basal diameter, allometric scaling, terrain
Citation: Wu X, Zheng X-J, Mu X-H and Li Y (2021) Differences in Allometric Relationship of Two Dominant Woody Species Among Various Terrains in a Desert Region of Central Asia. Front. Plant Sci. 12:754887. doi: 10.3389/fpls.2021.754887
Received: 10 August 2021; Accepted: 26 October 2021;
Published: 10 November 2021.
Edited by:
Ravi Kant Chaturvedi, Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences (CAS), ChinaReviewed by:
Yunpeng Nie, Institute of Subtropical Agriculture, Chinese Academy of Sciences (CAS), ChinaCopyright © 2021 Wu, Zheng, Mu and Li. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yan Li, bGl5YW4yMDE2QHphZnUuZWR1LmNu
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.