 Shilian Huang
Shilian Huang Dongmei Han
Dongmei Han Jing Wang
Jing Wang Jianguang Li
Jianguang Li- Key Laboratory of South Subtropical Fruit Biology and Genetic Resource Utilization (MOA), Guangdong Province Key Laboratary of Tropical and Subtropical Fruit Tree Research, Institute of Fruit Tree Research, Guangdong Academy of Agricultural Sciences, Guangzhou, China
Longan (Dimocarpus longan L.) is one of the most important tropical and subtropical fruits in the world. Longan fruit has high nutritional and medical value, and is regarded as a treasure among fruits. Since it was first reported that potassium chlorate (KClO3) could be successfully applied to promote flowering in longan, this compound has been widely used in the production of on-season and off-season longan fruits. KClO3 has thus played a great role in promoting the development of the longan industry. In this review, we summarize the application methods, influencing factors, and physiological and molecular mechanisms associated with KClO3-mediated induction of longan flowering. It can be deduced that leaves may play a crucial role in the transport of and response to KClO3. Leaves supply carbon and nitrogen nutrition, and hormone and signaling molecules needed for the differentiation of apical buds. Moreover, cytokinins may be crucial for KClO3-mediated induction of longan flowering. More effort should be focused on studying the molecular mechanisms underlying this process. This will not only help us to better understand floral induction by KClO3 in longan but also enrich our understanding of flowering regulation mechanisms in woody plants.
Introduction
Longan (Dimocarpus longan Lour.) is an important tropical and subtropical fruit tree that belongs to the Sapindaceae family (Lai et al., 2000). Native to southern China and Southeast Asia, longan is cultivated in more than 20 countries, from Sri Lanka and India to East Malaysia and Australia (Janick, 1989; Lithanatudom et al., 2017). China has the largest planting area and highest yield of longan, followed by Thailand, Vietnam, India, and South Africa. These five countries produce ninety percent longan (Food and Agriculture Organization, 2014; Wang et al., 2015). Longan can be eaten fresh or processed, and is widely consumed due to its sweet juicy taste and health benefits. These health benefits include blood metabolism promotion, memory enhancement and insomnia incidence reduction (Rangkadilok et al., 2007; Park et al., 2010). Longan have also been used as traditional Chinese medicine to treat leucorrhea, kidney disorders, allergies, and cardiovascular diseases (Jiang et al., 2009).
Floral induction (FI) is considered the first step from vegetative to reproductive growth. FI plays an important role as it determines the success of commercial orchards (Bangerth, 2009). The flowering of many plants is regulated by environmental factors, such as chilling, drought, and oxidative stresses. With the development of science and technology, it is possible to create similar environmental conditions for flowering requirement, or to find chemicals to promote off-season flowering. For example, supplementary lighting in short-day winters can induce flowering in long-day pitaya (Hylocereus undatus Britton et Rose) (Xiong et al., 2020). In addition, the use of paclobutrazol can induce off-season flowering in mango (Mangifera indica L.) (Sergent et al., 1997; Abdel Rahim et al., 2011).
Normally, a period of low temperature (usually ≤ 18°C) is required for longan to bloom (Suttitanawat et al., 2012). In warmer winter, girdling can control the growth of winter shoots and promote flowering in spring (Wu et al., 2000). In addition, low concentrations (100–200mg/L) of paclobutrazol (PP333) have been shown to significantly promote flower bud differentiation (Huang, 1996). The first report of potassium chlorate (KClO3) being used to induce flowering in longan was published by Changrui Yan. Since then, attempts have been made to achieve stable FI in longan (Yan et al., 1998; Li et al., 2006). Thus, KClO3 has now been widely used in longan production. Fresh longan fruits are currently available in local markets in Thailand all year round due to the application of KClO3 (Subhadrabandhu and Yapwattanaphun, 2000).
Thus far, FI by KClO3 has only been found to be effective for longan. Studying FI by KClO3 will provide a deeper understanding of the flowering regulation mechanism of woody plants. Furthermore, such study will provide a theoretical basis for regulating the flowering of other Sapindaceae plants, such as Litchi chinensis Sonn. In this paper, we summarize the application methods, physiological and molecular regulation mechanisms, influencing factors, and the environmental impact of KClO3-mediated FI in longan. Furthermore, the future perspectives of studying the mechanisms underlying FI by KClO3 are analyzed.
Application Methods
Year-round FI is achieved in longan with KClO3 application. The perfect time period to apply KClO3 for on-season FI is from November to January, after the maturation of the last shoot. Off-season FI can be achieved by applying KClO3 from February to October, when the leaves of the last shoot are light green. Leaves play a fundamental and essential role in this process of FI (Nunez-Elisea et al., 1996). To achieve higher FI efficiency, KClO3 should be applied when leaves are older than 60 days, as flower bud differentiation requires adequate nutrition. It has been proposed by Hegele et al. (2004) that the presence of young leaves reduces the efficiency of FI by KClO3. Furthermore, supernutrition can lead to flushes of new leaves, and thus the timing of KClO3 treatment needs to be precisely controlled (Lu et al., 2006).
Foliar spraying and soil drenching are the most common KClO3 application approaches. For foliar spraying, 0.5–3 g/L KClO3 solution is sprayed onto the leaves. For soil drenching, a circular shallow ditch with a depth of approximately 15–20 cm and a width of approximately 15–25 cm should be dug along the drip line of the longan tree crown. Then, 0.5–2 kg of solid KClO3 or water solution should be spread into the ditch. Generally, the combination of the two methods will produce better results. The specific KClO3 dosage should be determined according to the plant variety and age, and to the climate. In addition, the soil needs to be kept slightly wet for 15 days after KClO3 treatment. Furthermore, fertilization and pruning should be avoided during the period between the treatment and flowering (Huang et al., 2009).
Factors Influencing KCLO3-Mediated FI
Floral induction is highly correlated with leaf age in tropical and subtropical fruit trees. Longan apical buds with high carbon content (> 50.93 mg/g) in the leaves can be induced into flowers. Meanwhile, those with leaves with low carbon content (< 37.40 mg/g) cannot be induced by KClO3 out of season (Hong et al., 2014). Previous research has suggested that mature leaves might be involved in the conversion of isopentenyladenine/isopentenyladenosine (iP/iPA) cytokinin (CK) precursors into the active zeatin/zeatin riboside (Z/ZR) CKs. Hence, leaves may be involved in the FI process (Potchanasin et al., 2009a; Tiyayon et al., 2010). Shading has been shown to inhibit the export of indoleacetic acid (IAA) out of the shoot apical buds, the leaf export of iP/iPA- and Z/ZR-type CKs, and CK accumulation in shoot apical buds. In this way, shading can prevent FI by KClO3 treatment (Sritontip et al., 2008; Sringarm et al., 2009a; Ongprasert et al., 2010). Appropriate KClO3 concentrations promote flowering, while excessive KClO3 concentrations may lead to less flowering, no flowering or leaf burn (Ongprasert et al., 2010). Weather conditions can also affect KClO3-mediated FI; the lowest flowering percentage (11.9%–50.9%) occurs in rainy seasons, and higher flowering percentage (77.5–88.6%) in cool and hot seasons (Manochai et al., 2005).
Absorption and Metabolism of Chlorate
Plant cells share the same absorption mechanism for chlorate and nitrate through nitrate transporter (Glass et al., 1999). Chlorate is generally not toxic to plants; however, chlorate becomes toxic when converted into chlorite and hypochlorite by nitrate reductase (NR) and nitrite reductase (NiR) (Hofstra, 1977). Chlorate is not harmful while plants lack NR activity (Doddema et al., 1978; Borges et al., 2004). The interactions of potassium chlorate and proteins in Arabidopsis thaliana and Populus trichocarpa were searched by STITCH 5.0 at http://stitch.embl.de/cgi/ (Szklarczyk et al., 2016). The interaction of two NRs with potassium chlorate was found in Arabidopsis thaliana and Populus trichocarpa (Figure 1). The following were also observed: one multidrug and toxic compound extrusion (MATE) transporter, one major facilitator superfamily transporter, one aspartyl protease in guard cell 1 in Arabidopsis thaliana (Figure 1). Meanwhile, four MATE transporters were found in Populus trichocarpa (Figure 1). These findings indicate that nitrate reduction systems are crucial in chlorate absorption and metabolism. MATE may pump chlorate out of cells or transfer chlorate into vacuoles for detoxification due to its implication in the membrane-mediated transport of small organic molecules, metal ions, and chloride ions (Zhang et al., 2017; Upadhyay et al., 2019).
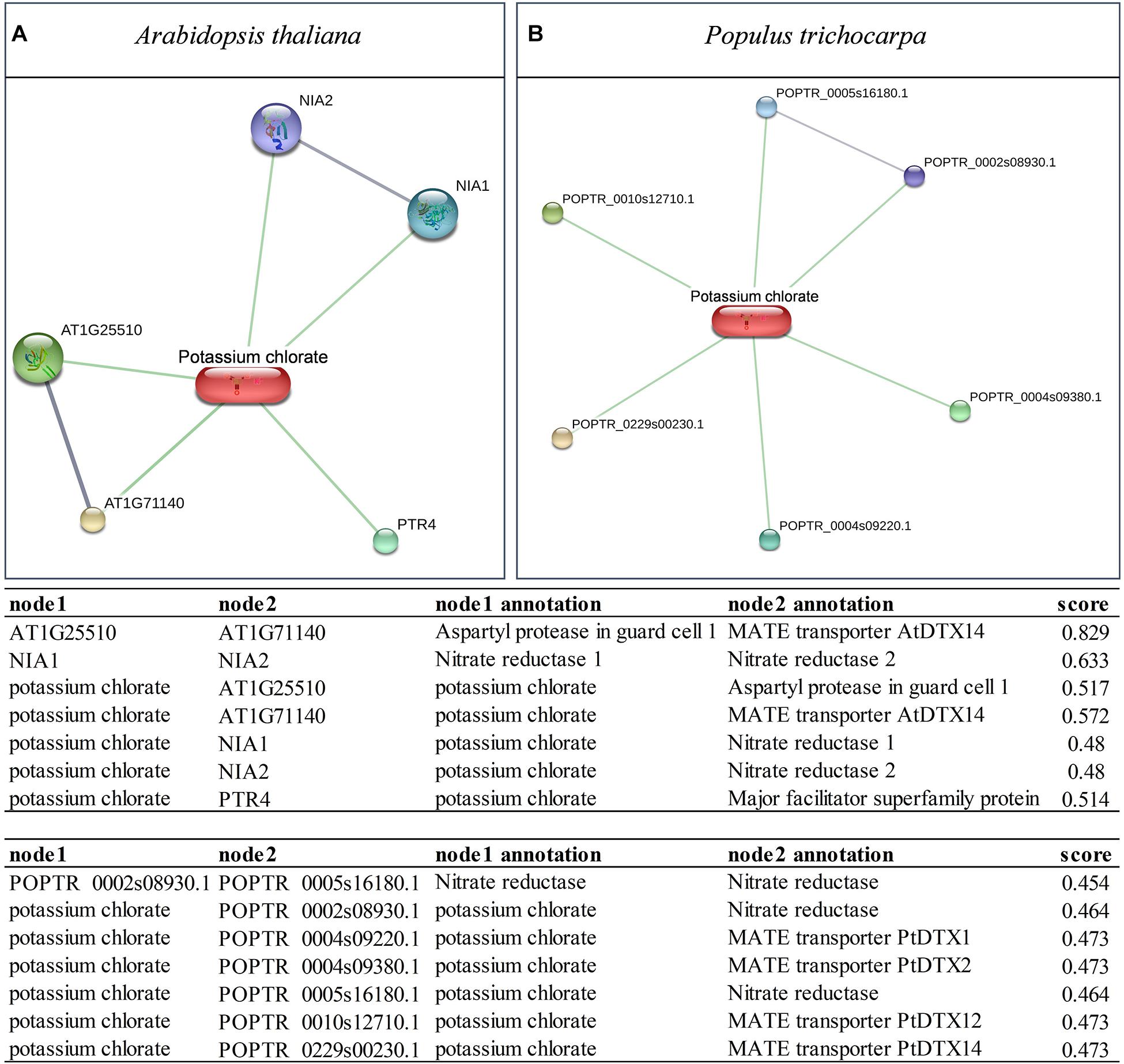
Figure 1. Interactions of potassium chlorate and proteins in Arabidopsis thaliana (A) and Populus trichocarpa (B) searched by STITCH 5.0. (A) Generated by input “Item Name: potassium chlorate, Organism: Arabidopsis thaliana”; (B) Generated by input “Item Name: potassium chlorate, Organism: Populus trichocarpa.” Stronger associations are represented by thicker lines and higher scores in the follow table. Protein-protein interactions are shown in gray and chemical-protein interactions in green.
Physiological Mechanisms of KCLO3-Mediated FI
Plant Hormones
Endogenous plant hormones participate in the entire life process of plants. Plant hormones regulate plant growth and development by forming a complex and complete signaling network that enables the transmission of exogenous or endogenous signals. Therefore, hormone signals are crucial to flower formation (Santner and Estelle, 2009; Wolters and Jürgens, 2009; Puja et al., 2015). Application of KClO3 can induce changes in endogenous hormones. The contents of two types of CK—trans-zeatin (tZ)/ZR and iP/iPA—have been shown to increase after KClO3 treatment in apical buds, sub-apical wood and bark, leaves and roots (Potchanasin et al., 2009b; Hegele et al., 2010; Suttitanawat et al., 2012). However, only ZR seemed to be involved in the FI process (Bangerth et al., 2010). Mature leaves may set the stage for the conversion of iPA into ZR, and KClO3 treatment promotes the translocation of CKs from the leaves to apical buds (Sringarm et al., 2009b; Tiyayon et al., 2010). The increased CK contents can also be detected during flowering in longan (Sringarm et al., 2009b). The contents of gibberellic acid (GA), IAA, and abscisic acid (ABA) in shoot apical buds and leaves have been shown to decrease following KClO3 application (Hegele et al., 2008, 2010; Tiyayon et al., 2010). This shows that longan FI by KClO3 may require more CKs, but less GA, IAA, and ABA. Increased production of ethylene (ETH) has also been detected in longan leaves after KClO3 treatment. It is unknown whether this increase is involved in the FI process or a stress response (Sringarm et al., 2009b). Besides ETH and CK, KClO3 treatment can also increase the amount of salicylic acid (SA). SA has also been found to be closely related to plant flower formation (Sringarm et al., 2009b; Martínez et al., 2010).
Carbon and Nitrogen Nutrition
The carbon–nitrogen (C:N) ratio is an important physiological factor influencing flowering. Carbohydrate reserves are a prerequisite for FI in tropical and subtropical trees. High carbohydrate and low nitrogen contents lead to a high C:N ratio, which is conducive to flowering. Meanwhile, a high nitrogen content results in a low C:N ratio, which is favorable for vegetative growth (Corbesier et al., 2002). It seems that KClO3 treatment does not impact the total nitrogen content, total non-structural carbohydrate content, or carbohydrate–nitrogen ratio (Charoensri et al., 2005; Wangsin and Pankasemsuk, 2005; Matsumoto et al., 2007). However, it has been shown that the content of soluble sugar, fructose, and glucose increased, the sucrose content increased significantly, and the starch content decreased significantly in longan leaves in response to KClO3 treatment (Chen and Li, 2004; Lu, 2005; Chang, 2010). KClO3 treatment has also been shown to reduce the longan leaf nitrate reductase activity. In the same study, KClO3 led to a peak in soluble amino acid accumulation in the leaves within 2 weeks of treatment; this amino acid accumulation then decreased but continued to increase in the apical buds (Lu, 2005). Generally, higher ammoniacal nitrogen contents are beneficial for flowering. Overall, it can be inferred that KClO3 treatment can change the types of carbohydrate and protein to promote reproductive development without affecting total nitrogen or total non-structural carbohydrate contents.
Stress Response
KClO3 is a type of strong oxidant that is used as a herbicide. High KClO3 concentrations will cause longan leaves to turn yellow and fall off. It is speculated that appropriate amounts of KClO3 can lead to stress responses. Reduced net carbon dioxide (CO2) assimilation, transpiration, stomatal conductance rates, and photosystem II efficiencies (Fv/Fm) have been detected after KClO3 treatment (Hegele et al., 2008; Sritontip et al., 2010, 2013). The treatment has resulted in chlorophyll degradation, the destruction of chloroplast thylakoid membrane structure, the disappearance of chloroplast starch granules, and the destruction of basal granules (Lu, 2005). The decrease in photosynthetic capacity has been shown to have been mainly caused by the inhibition of the activity of the photosynthetic apparatus (Chang, 2010). The reactive oxygen species and malondialdehyde contents, and superoxide dismutase and peroxidase activities, were found to be higher in leaves within 1 month after KClO3 treatment than in the control. Meanwhile, the leaf water potential and root activity were significantly lower than in the control (Ouyang et al., 2005).
Molecular Mechanisms of KCLO3-Mediated FI
Several genes related to the flowering of longan have been identified. Tiyayon et al. (2011) first cloned the longan flowering locus T (DlFT) gene, which shared 68% identity with the Arabidopsis thaliana gene, AtFT (Tiyayon et al., 2011). Winterhagen et al. (2013) isolated DlFT1, DlFT2, and two APETALA1-like (DlAP1-1 and DlAP1-2) sequences from longan. Transgenic analysis indicated that DlFT1 promoted flowering, while DlFT2 inhibited flowering. Ectopic overexpression of AP1 genes in Arabidopsis resulted in early or late-flowering phenotypes (Winterhagen et al., 2013). Overexpression of the longan gigentea (DlGI) and flavin-binding, kelch repeat, F-box 1 (DlFKF1) genes caused Arabidopsis to bloom early under long-day conditions (Huang et al., 2017). The early flowering 4 proteins, DlELF4-1 and DlELF4-2, were found to bind to and activate the promoter of DlGI (Waheed et al., 2020). Through transcriptome analysis of “Sijimi” longan, Zhang et al. (2016) found a large number of genes related to the four known flowering pathways and floral integrator genes. By comparing and analyzing the different expression levels of genes in the terminal tips of “Sijimi” and “Lidongbe” longan, short vegetative phase (SVP), GI, FKF1, and ELF4 were found to be involved in the continuous flowering of “Sijimi,” and ELF4 might play a key role.
Sixty-five uniquely expressed genes were identified between buds with and without KClO3 treatment, and many of them were demonstrated to be involved in shoot and floral meristem development. These genes included homologs of protodermal factor 1 (PDF1), SHEPHERD, and PISTILLATA (Matsumoto, 2006; Matsumoto et al., 2007). KClO3 treatment was also found to enhance the expression of DlFT1 in mature leaves, which was highly consistent with the increased CK content (Winterhagen et al., 2020).
Conclusion
By analyzing the results of previous studies, it can be inferred that in longan, leaves are the main plant organs that respond to KClO3 treatment. KClO3 treatment can induce stress responses in leaves. These stress responses include reduced leaf water potential, net CO2 assimilation, transpiration, stomatal conductance rates and Fv/Fm, increased destruction of photosynthetic apparatus, malondialdehyde and reactive oxygen species contents, and superoxide dismutase and peroxidase activities (Figure 2; Ouyang et al., 2005; Sritontip et al., 2013). KClO3 treatment can also cause changes in carbon and nitrogen nutrition in longan leaves. It reduces the starch content and increases the soluble sugar, fructose, glucose, sucrose, and soluble amino acid contents (Figure 2; Lu, 2005; Chang, 2010). Furthermore, KClO3 treatment can lead to changes in leaf hormone contents. The treatment slightly reduces the contents of GA, IAA, and ABA and increases the contents of ETH, SA, and CKs (Figure 2; Sringarm et al., 2009b; Martínez et al., 2010). CKs may play a particularly vital role in FI by KClO3. KClO3 treatment can also slightly reduce the contents of GA, IAA, and ABA, and increase the contents of iPA- and ZR-type CKs in the apical bud, which may be due to transport from the leaves (Figure 2; Sringarm et al., 2009b; Tiyayon et al., 2010). The enhanced soluble sugar and soluble amino acid contents provide nutrition for flower bud differentiation. In addition, the H2O2 generated in the stress response process may act as an important signal molecule in off-season FI of longan as it can promote the expression of DlAP1 and DlFT (Hong et al., 2015; Yang et al., 2016).
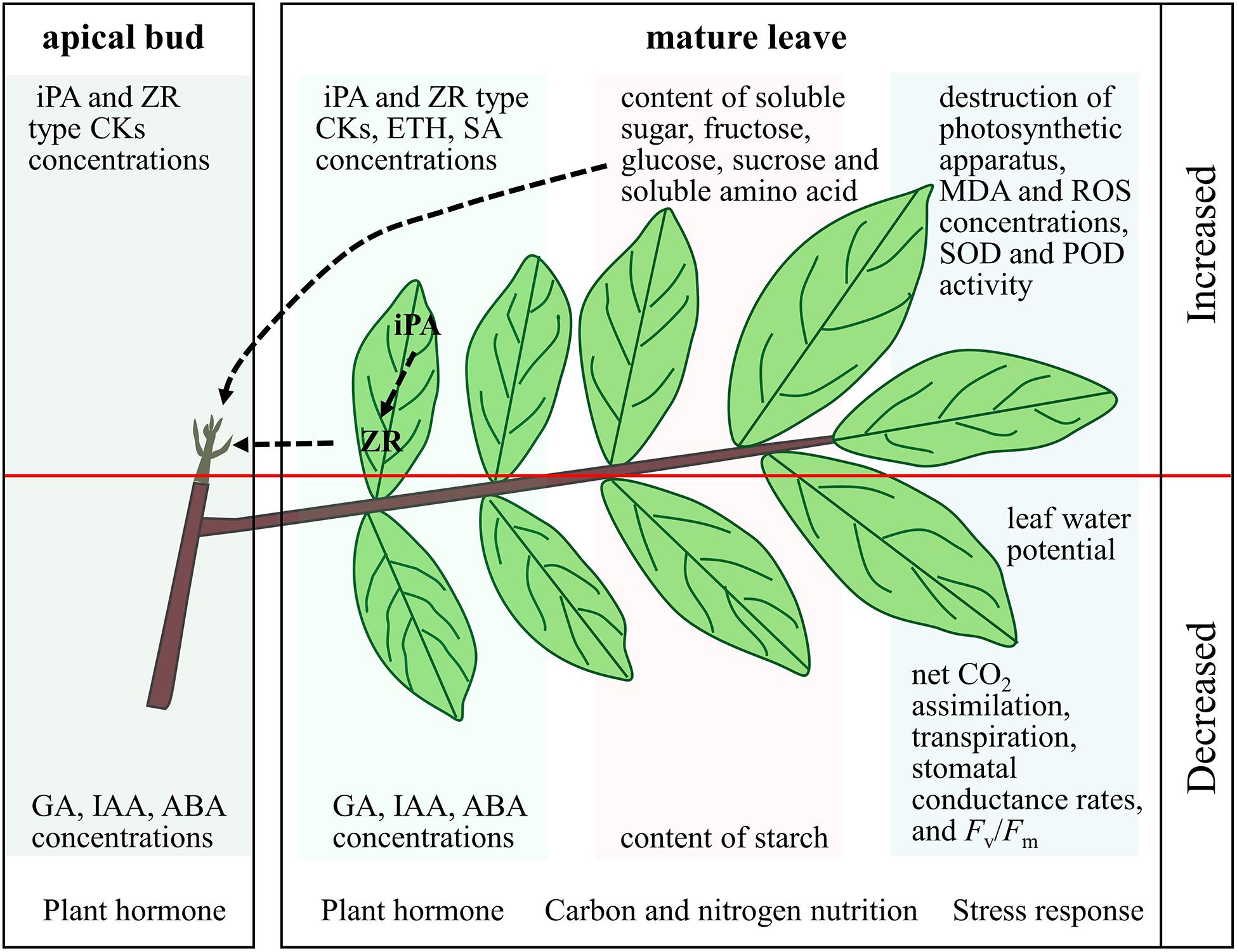
Figure 2. Physiological mechanism of floral induction by KClO3 in longan. The physiological indexes above the red line increased after KClO3 treatment, while the indexes under the red line decreased.
We have a general understanding of the physiological mechanism underlying longan FI by KClO3. Though little is known about the corresponding molecular mechanism, a speculated regulation model of KClO3-mediated FI was proposed based on above information. Chlorate can be reduced to chlorite and hypochlorite by nitrate reductase (NR) and nitrite reductase (NiR) (Figure 3; Borges et al., 2004). Chlorite and hypochlorite may directly cause stress response, CK content increasement and expression of flowering-related genes (Figure 3, blue dotted lines). It is reported that stress could induce the cytokinin synthesis (Reguera et al., 2013) and flowering (Cho et al., 2017). There is another possibility that the stress response caused by chlorite and hypochlorite may contribute to the CK content increasement and expression of flowering-related genes (Figure 3, red dotted lines). Also, the enhanced CK content may induce the expression of flowering related genes as indicated by Winterhagen et al. (2020) (Figure 3, black dotted lines). Excess chlorate in plant cell may be transferred out of cell or into vacuole by MATE transporters (Figure 3, blue dotted lines).
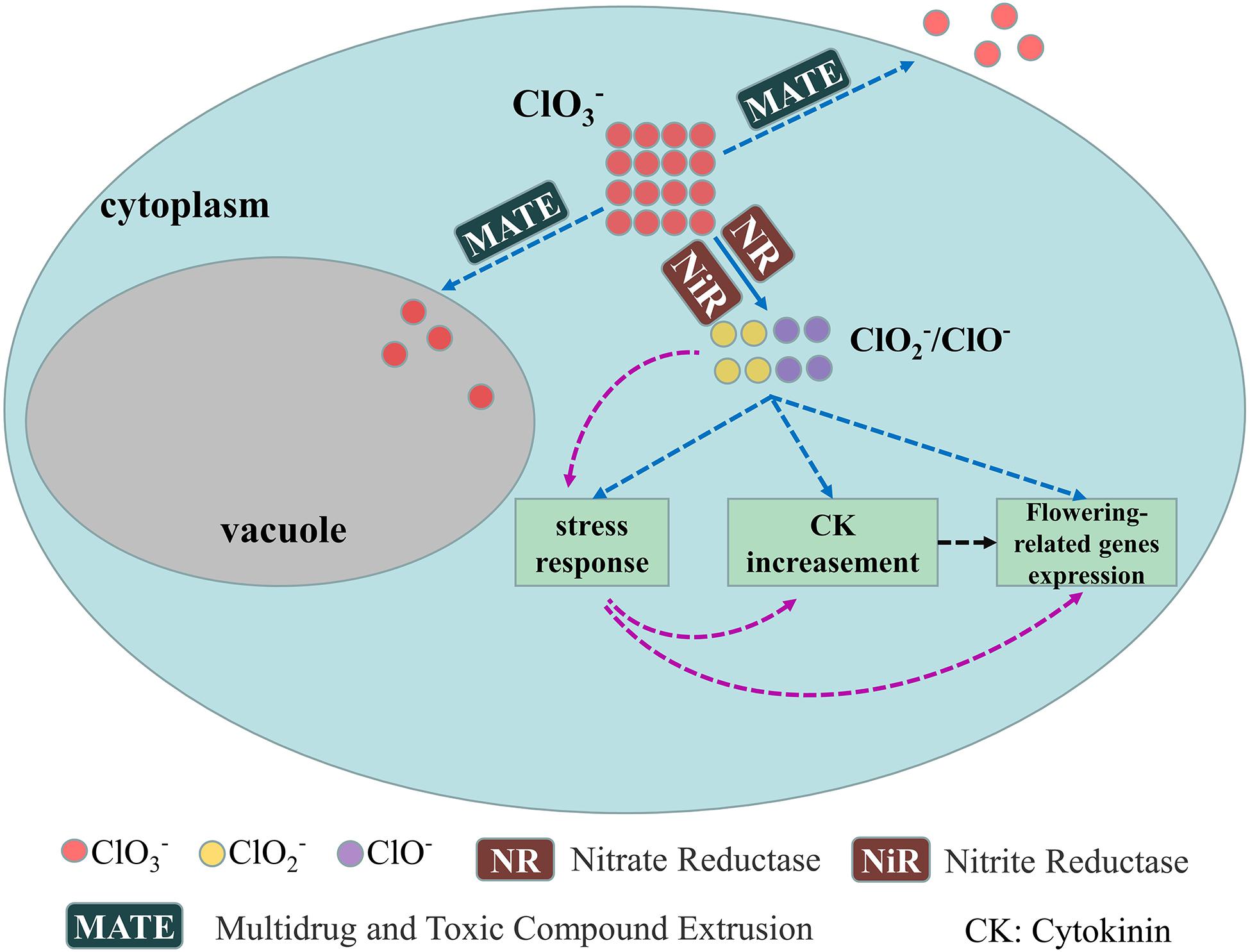
Figure 3. Proposed model of the molecular regulation of KClO3 mediated FI. The solid line shows confirmed result while dotted lines indicate speculated. Blue dotted lines (after ClO2–/ClO–): ClO2–/ClO– may directly induce stress response, CK content increasement and flowering-related genes expression; Red dotted lines: ClO2–/ClO– induce stress response, then stress response induce CK content increasement and flowering-related genes expression; Black dotted line: CK content increasement may also induce flowering-related genes expression.
It remains to be understood how KClO3 is transferred from the root to leaf or from the leaf to root. Furthermore, it is unclear how KClO3 causes various physiological changes and why it is possible to use KClO3 in place of cold treatment in FI. In particular, it will be important to understand the relationship between KClO3 treatment and the contents of CKs, and what role CKs play in FI by KClO3. Addressing these issues will not only provide a full understanding of the mechanism underlying FI by KClO3 in longan but also enrich our theoretical understanding of flowering regulation in woody plants. Although the genome of longan has been published (Lin et al., 2017), a genetic transformation system, or a highly efficient virus-induced gene silencing system for longan, will be required to undertake the necessary further research.
Author Contributions
SH, DH, and JW wrote the manuscript. SH initiated the idea of the review. DG and JL revised the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This research was supported by the Guangdong Provincial Key Research and Development Program (2020B020220006), and the Guangdong Province Modern Agricultural Industrial Technology System Innovation Team Building Project (2019KJ123).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank LetPub (www.letpub.com) for its linguistic assistance during the preparation of this manuscript.
References
Abdel Rahim, A. O. S., Elamin, O. M., and Bangerth, F. K. (2011). Effects of paclobutrazol (PBZ) on floral induction and associated hormonal and metabolic changes of biennially bearing mango (Mangifera indica L.) cultivars during off year. J. Agr. Bio. Sci. 6, 55–67.
Bangerth, K. F. (2009). Floral induction in mature, perennial angiosperm fruit trees: similarities and discrepancies with annual/biennial plants and the involvement of plant hormones. Sci. Hortic. 122, 153–163. doi: 10.1016/j.scienta.2009.06.014
Bangerth, K. F., Potchanasin, P., Sringarm, K., Dongliang, Q., Mitra, S. K., and Diczbalis, Y. (2010). Hormonal regulation of the regular and ‘off season’ floral induction process of longan (Dimocarpus longan) trees. Acta Hortic. 863, 215–224. doi: 10.17660/ActaHortic.2010.863.28
Borges, R., Miguel, E. C., Dias, J. M. D., da Cunha, M., Bressan-Smith, R. E., de Oliveira, J. G., et al. (2004). Ultrastructural, physiological and biochemical analyses of chlorate toxicity on rice seedlings. Plant Sci. 166, 1057–1062. doi: 10.1016/j.plantsci.2003.12.023
Chang, Q. (2010). Study on relationship between off-season floral induction in longan and nutrition of the carbon and nitrogen, Master’s thesis, Fuzhou: Fujian Agriculture & Forestry University.
Charoensri, P., Jutamanee, K., Tawatpun, J., Tongumpai, P., and Krisanapook, K. (2005). Effects of potassium chlorate and girdling on flowering of ‘Phet Sakhon’ longan. Acta Hortic. 665, 259–268. doi: 10.17660/ActaHortic.2005.665.30
Chen, Q. X., and Li, S. G. (2004). KClO3 induced longan to form floral bud and bloom and its changes of carbohydrates and protein in leaves. J. Fujian Agr. Forestry U 33, 182–185.
Cho, L. H., Yoon, J., and An, G. (2017). The control of flowering time by environmental factors. Plant J. 90, 708–719. doi: 10.1111/tpj.13461
Corbesier, L., Bernier, G., and Périlleux, C. (2002). C:N ratio increases in the phloem sap during floral transition of the long-day plants Sinapis alba and Arabidopsis thailiana. Plant Cell Physiol. 43, 684–688. doi: 10.1093/pcp/pcf071
Doddema, H., Hofstra, J. J., and Feenstra, W. J. (1978). Uptake of nitrate by mutants of Arabidopsis thaliana, disturbed in uptake or reduction of nitrate: I. effect of nitrogen source during growth on uptake of nitrate and chlorate. Physiol. Plant. 43, 343–350. doi: 10.1111/j.1399-3054.1978.tb01592.x
Food and Agriculture Organization (2014). Tropical fruits compendium. Available online at: http://www.fao.org/docrep/meeting/022/am481t.pdf (accessed September 20, 2014).
Glass, A. D. M., Erner, Y., Hunt, T., Kronzucker, H. J., Okamoto, M., Rawat, S., et al. (1999). “Inorganic nitrogen absorption by plant roots,” in Plant Nutrition-Molecular Biology and Genetics, Gissel-Nielsen, G., Jensen, A (Dordrecht: Springer), 1–16. doi: 10.1007/978-94-017-2685-6_1
Hegele, M., Manochai, P., Naphrom, D., Sruamsiri, P., and Wünsche, J. (2008). Flowering in longan (Dimocarpus longan L.) induced by hormonal changes following KClO3 applications. Europ. J. Hort. Sci. 73, 49–54.
Hegele, M., Naphrom, D., Manochai, P., Chattrakul, A., Sruamsiri, P., and Bangerth, F. (2004). Effect of leaf age on the response of flower induction and related hormonal changes in longan trees after KClO3 treatment. Acta Hortic. 653, 41–49. doi: 10.17660/ActaHortic.2004.653.4
Hegele, M., Sritontip, C., Chattrakul, A., Tiyayon, P., and Wünsche, J. N. (2010). Hormonal control of flower induction in litchi and longan. Acta Hortic. 863, 305–314. doi: 10.17660/ActaHortic.2010.863.40
Hong, J., Chen, X., Li, S., Zhang, L., and Yang, Z. (2015). The impact of H2O2 and NO on FT gene expression and floral bud formation in longan. Chin. J. Trop. Crop. 36, 2002–2006.
Hong, J. W., Li, S. G., Zhang, L., and Yang, Z. Q. (2014). Carbon analysis on flowering differentiation in off-season longan. Guangdong Agr. Sci. 151, 921–928. doi: 10.16768/j.issn.1004-874x.2014.16.030
Hofstra, J. J. (1977). Chlorate toxicity and nitrate reductase activity in tomato plants. Physiol. Plant 41, 65–69. doi: 10.1111/j.1399-3054.1977.tb01524.x
Huang, F., Fu, Z., Zeng, L., and Morley-Bunker, M. (2017). Isolation and characterization of GI and FKF1 homologous genes in the subtropical fruit tree Dimocarpus longan. Mol. Breeding 37:90. doi: 10.1007/s11032-017-0691-z
Huang, J. Y., Pang, X. H., and Zhou, Q. G. (2009). Research on the technology of forcing logan. J. Guangxi Vocational. Tech. Coll. 2, 5–6.
Huang, Q. W. (1996). Effects of plant growth regulators on endogenous hormones and bud differentiation of longan. Acta Bot. Yunnanica 18, 145–150.
Jiang, G., Jiang, Y., Yang, B., Yu, C., Tsao, R., Zhang, H., et al. (2009). Structural characteristics and antioxidant activities of oligosaccharides from longan fruit pericarp. J. Agric. Food Chem. 57, 9293–9298. doi: 10.1021/jf902534v
Lai, Z., Chen, C., Zeng, L., and Chen, Z. (2000). “Somatic embryogenesis in longan [Dimocarpus longan Lour.],” in Somatic Embryogenesis in Woody Plants, eds S. M. Jain, P. Gupta, and R. Newton (Netherlands: Springer), 415–431. doi: 10.1007/978-94-017-3030-3_13
Li, M., Ren, L., and Wang, Y. (2006). Review on applications and mechanism of KClO3 on inducing flower formation of Longan (Dimocarpus longan Lour.) trees. J. South. Agr. 37, 293–297.
Lin, Y. L., Min, J. M., Lai, R. L., Wu, Z. Y., Chen, Y. K., Yu, L. L., et al. (2017). Genome-wide sequencing of longan (Dimocarpus longan lour.) provides insights into molecular basis of its polyphenol-rich characteristics. GigaScience 6, 1–14. doi: 10.1093/gigascience/gix023
Lithanatudom, S. K., Chaowasku, T., Nantarat, N., Jaroenkit, T., Smith, D. R., and Lithanatudom, P. (2017). A first phylogeny of the genus Dimocarpus and suggestions for revision of some taxa based on molecular and morphological evidence. Sci. Rep. 7:6716. doi: 10.1038/s41598-017-07045-7
Lu, J. M. (2005). A physiological study on biological effects of potassium chlorate on longan (Dimocarpus longan Lour.), master’s thesis, Guangzhou: South China Agricultural University.
Lu, J. M., Huang, X. M., Wang, H. C., Zhang, C. L., Yang, R. T., and Xie, L. (2006). Current situation in the research and application of chlorate in longan production and approaches to understanding the mechanisms of the biological effects of chlorate. Plant Physiol. J. 42, 567–572.
Manochai, P., Sruamsiri, P., Wiriya-Alongkorn, W., Naphrom, D., Hegele, M., and Bangerth, F. (2005). Year around off-season flower induction in longan (Dimocarpus longan Lour.) trees by KClO3 applications: potentials and problems. Sci. Hortic. 104, 379–390. doi: 10.1016/j.scienta.2005.01.004
Martínez, C., Pons, E., Prats, G., and León, J. (2010). Salicylic acid regulates flowering time and links defence responses and reproductive development. Plant J. 37, 209–217. doi: 10.1046/j.1365-313X.2003.01954.x
Matsumoto, T. K. (2006). Genes uniquely expressed in vegetative and potassium chlorate induced floral buds of Dimocarpus longan. Plant Sci. 170, 500–510. doi: 10.1016/j.plantsci.2005.09.016
Matsumoto, T. K., Tsumura, T., and Zee, F. (2007). Exploring the mechanism of potassium chlorate-induced flowering in Dimocarpus longan. Acta Hortic. 738, 451–458. doi: 10.17660/ActaHortic.2007.738.56
Nunez-Elisea, E. R., Davenport, T. L., and Carderia, M. L. (1996). Control of bud morphogenesis in mango (Mangifera indica L.) by girdling, defoliation and temperature modification. J. Hort. Sci. 71, 25–39. doi: 10.1080/14620316.1996.11515379
Ongprasert, S., Wiriya-Alongkorn, W., and Spreer, W. (2010). The factors affecting longan flower induction by chlorate. Acta horticulturae 863, 375–380. doi: 10.17660/actahortic.2010.863.50
Ouyang, R., Liu, H. P., Li, P., Wang, H. C., and Hu, G. B. (2005). Physiological responses of longan to KClO3 stress. Acta Agr. U. Jiangxiensi 27, 34–38.
Park, S. J., Dong, H. P., Dong, H. K., Lee, S., Yoon, B. H., Jung, W. Y., et al. (2010). The memory-enhancing effects of Euphoria longan fruit extract in mice. J. Ethnopharmacol. 128, 160–165. doi: 10.1016/j.jep.2010.01.001
Potchanasin, P., Sringarm, K., Naphrom, D., and Bangerth, K. F. (2009a). Floral induction in longan (Dimocarpus longan, Lour.) trees: iv. the essentiality of mature leaves for potassium chlorate induced floral induction and associated hormonal changes. Sci. Hortic. 122, 312–317. doi: 10.1016/j.scienta.2009.06.007
Potchanasin, P., Sringarm, K., Sruamsiri, P., and Bangerth, K. F. (2009b). Floral induction (FI) in longan (Dimocarpus longan Lour.) trees: part i. low temperature and potassium chlorate effects on fi and hormonal changes exerted in terminal buds and sub-apical tissue. Sci. Hortic. 122, 288–294. doi: 10.1016/j.scienta.2009.06.008
Puja, O., Renu, B., Shagun, B., Ravinderjit, K., Shivam, J., Anjali, K., et al. (2015). The Common Molecular Players in Plant Hormone Crosstalk and Signaling. Curr. Protein Pept. Sc. 16, 369–388. doi: 10.2174/1389203716666150330141922
Rangkadilok, N., Sitthimonchai, S., Worasuttayangkurn, L., Mahidol, C., Ruchirawat, M., and Satayavivad, J. (2007). Evaluation of free radical scavenging and antityrosinase activities of standardized longan fruit extract. Food Chem. Toxicol. 45, 328–336. doi: 10.1016/j.fct.2006.08.022
Reguera, M., Peleg, Z., Abdel-Tawab, Y. M., Tumimbang, E. B., Delatorre, C. A., and Blumwald, E. (2013). Stress-induced cytokinin synthesis increases drought tolerance through the coordinated regulation of carbon and nitrogen assimilation in rice. Plant Physiol. 163, 1609–1622. doi: 10.1104/pp.113.227702
Santner, A., and Estelle, M. (2009). Recent advances and emerging trends in plant hormone signalling. Nature 459, 1071–1078. doi: 10.1038/nature08122
Sergent, E., Ferrari, D., and Leal, F. (1997). Effects of potasium nitrate and paclobutrazol on flowering induction and yield of mango (Mangifera indica L.) CV. Haden. Acta Hortic. 455, 180–187. doi: 10.17660/ActaHortic.1997.455.25
Sringarm, K., Potchanasin, P., Naphrom, D., and Bangerth, K. F. (2009a). Floral induction (FI) in longan (Dimocarpus longan Lour.) trees. iii: effect of shading the trees on potassium chlorate induced FI and resulting hormonal changes in leaves and shoots. Sci. Hortic. 122, 301–311. doi: 10.1016/j.scienta.2009.06.006
Sringarm, K., Potchanasin, P., Sruamsiri, P., and Bangerth, K. F. (2009b). Floral induction (FI) in longan (Dimocrapus longan Lour.) trees – The possible participation of endogenous hormones: II. Low temperature and potassium chlorate effects on hormone concentration in and their export out of leaves. Sci. Hortic. 122, 295–300. doi: 10.1016/j.scienta.2008.11.031
Sritontip, C., Khaosumain, Y., Changjeraja, S., and Changjeraja, R. (2008). Effects of light intensity and potassium chlorate on photosynthesis and flowering in ‘Do’ Longan. Acta Hortic. 787, 285–288. doi: 10.17660/actahortic.2008.787.33
Sritontip, C., Tiyayon, P., Hegele, M., Sruamsiri, P., and Wünsche, J. N. (2010). Effects of temperature and potassium chlorate on leaf gas exchange and flowering in longan. Acta Hortic. 663, 323–328. doi: 10.17660/actahortic.2010.863.42
Sritontip, C., Tiyayon, P., Sringam, K., Pantachod, S., Naphrom, D., Ruamrungsri, S., et al. (2013). Influence of water regimes and potassium chlorate on floral induction, leaf photosynthesis and leaf water potential in longan. J. Agr. Sci. 5, 211–220. doi: 10.5539/jas.v5n6p211
Subhadrabandhu, S., and Yapwattanaphun, C. (2000). Regulation of flowering time for longan (Dimorcarpus longan) production in Thailand. J. Appl. Horticult. 2, 102–105. doi: 10.37855/jah.2000.v02i02.31
Suttitanawat, P., Sruamsiri, P., and Sringarm, K. (2012). Changes in cytokinins concentrations during induction period of longan cv. Daw in sand culture. J. Agr. Tech. 8, 2353–2362.
Szklarczyk, D., Santos, A., von Mering, C., Jensen, L. J., Bork, P., and Kuhn, M. (2016). STITCH 5: augmenting protein-chemical interaction networks with tissue and affinity data. Nucleic Acids Res. 44, D380–D384. doi: 10.1093/nar/gkv1277
Tiyayon, P., Hegele, M., Wünsche, J. N., Pongsriwat, K., Sruamsiri, P., and Samach, A. (2011). Studies on the molecular basis of flowering in longan (Dimocarpus longan). Acta Hortic. 903, 979–985. doi: 10.17660/actahortic.2011.903.137
Tiyayon, P., Sritontip, C., Hegele, M., Manochai, P., Sruamsiri, P., and Wünsche, J. N. (2010). Effects of girdling and defoliation on hormonal changes during flower induction in longan (Dimocarpus longan lour.). Acta Hortic. 863, 329–334. doi: 10.17660/actahortic.2010.863.43
Upadhyay, N., Kar, D., Mahajan, D. B., Nanda, S., Rahiman, R., Panchakshari, N., et al. (2019). The multitasking abilities of MATE transporters in plants. J. Exp. Bot. 70, 4643–4656. doi: 10.1093/jxb/erz246
Waheed, S., Peng, Y., and Zeng, L. (2020). Identification and characterization of DlGI promoter involved in photoperiod, light intensity, hormone, and DlELF4 response from longan. J. Am. Soc. Hortic. Sci. 145, 340–348. doi: 10.21273/JASHS04946-20
Wang, B., Tan, H. W., Fang, W., Meinhardt, L. W., Mischke, S., Matsumoto, T., et al. (2015). Developing single nucleotide polymorphism (SNP) markers from transcriptome sequences for identification of longan (Dimocarpus longan) germplasm. Hortic. Res. 2:14065. doi: 10.1038/hortres.2014.65
Wangsin, N., and Pankasemsuk, T. (2005). Effect of potassium chlorate on flowering, total nitrogen, total nonstructural carbohydrate, C/N ratio, and contents of cytokinin-like and gibberellin-like substances in stem apex of ‘do’ longan. Acta Hortic. 665, 255–258. doi: 10.17660/ActaHortic.2005.665.29
Winterhagen, P., Hegele, M., Tiyayon, P., and Wünsche, J. N. (2020). Cytokinin accumulation and flowering gene expression are orchestrated for floral meristem development in longan (Dimocarpus longan Lour.) after chemical flower induction. Sci. Hortic. 270:109467. doi: 10.1016/j.scienta.2020.109467
Winterhagen, P., Tiyayon, P., Samach, A., Hegele, M., and Wünsche, J. N. (2013). Isolation and characterization of FLOWERING LOCUS T subforms and APETALA1 of the subtropical fruit tree Dimocarpus longan. Plant Physiol. Biochem. 71, 184–190. doi: 10.1016/j.plaphy.2013.07.013
Wolters, H., and Jürgens, G. (2009). Survival of the flexible: hormonal growth control and adaptation in plant development. Nat. Rev. Genet. 10, 305–317. doi: 10.1038/nrg2558
Wu, D. Y., Qiu, J. D., Zhang, H. L., and Luo, X. Z. (2000). A study on flowering promotion by ringing in longan (Dimocarpus longan Lour.). Sci. Agr. Sinica 33, 40–43.
Xiong, R., Liu, C., Xu, M., Wei, S. S., and Huang, J. Q. (2020). Transcriptomic analysis of flower induction for long-day pitaya by supplementary lighting in short-day winter season. BMC Genomics 21:329. doi: 10.1186/s12864-020-6726-6
Yan, C. R., Zhao, Z. N., and Zhang, Z. W. (1998). Effects of chemical on introducing blossom of longan. J. Chinese Soc. Hort. Sci. 44, 517–518.
Yang, Z., Xu, Z., Zhang, L., Hong, J., Chen, Y., and Li, S. (2016). Effect of H2O2 and NO on AP1 gene expression and out of season flower bud formation in longan. South China Fruits 45, 13–17.
Zhang, H., Zhao, F. G., Tang, R. J., Yu, Y., Song, J., Wang, Y., et al. (2017). Two tonoplast MATE proteins function as turgor-regulating chloride channels in Arabidopsis. Proc. Natl. Acad. Sci. USA 114, E2036–E2045. doi: 10.1073/pnas.1616203114
Keywords: Dimocarpus longan, floral induction, KClO3, stress response, plant hormone
Citation: Huang S, Han D, Wang J, Guo D and Li J (2021) Floral Induction of Longan (Dimocarpus longan) by Potassium Chlorate: Application, Mechanism, and Future Perspectives. Front. Plant Sci. 12:670587. doi: 10.3389/fpls.2021.670587
Received: 15 March 2021; Accepted: 10 May 2021;
Published: 07 June 2021.
Edited by:
María Serrano, Miguel Hernández University of Elche, SpainReviewed by:
Hui-Cong Wang, College of Horticulture, South China Agricultural University, ChinaDhananjay K. Pandey, Amity University, Jharkhand, India
Copyright © 2021 Huang, Han, Wang, Guo and Li. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jianguang Li, bGpnLmNqLmpyQDE2My5jb20=; Dongliang Guo, Z3VvZG9uZ2xpYW5nQGdkYWFzLmNu