 Crispus M. Mbaluto1,2*
Crispus M. Mbaluto1,2* Esraa M. Ahmad3
Esraa M. Ahmad3 Anne Mädicke1,2Katharina Grosser1,2
Anne Mädicke1,2Katharina Grosser1,2 Nicole M. van Dam1,2
Nicole M. van Dam1,2 Ainhoa Martínez-Medina4*
Ainhoa Martínez-Medina4*- 1Molecular Interaction Ecology, German Centre for Integrative Biodiversity Research (iDiv) Halle-Jena-Leipzig, Leipzig, Germany
- 2Institute of Biodiversity, Friedrich-Schiller-Universität-Jena, Jena, Germany
- 3Department of Genetics, Faculty of Agriculture, Cairo University, Giza, Egypt
- 4Plant-Microorganism Interaction, Institute of Natural Resources and Agrobiology of Salamanca (IRNASA-CSIC), Salamanca, Spain
Plants mediate interactions between different herbivores that attack simultaneously or sequentially aboveground (AG) and belowground (BG) organs. The local and systemic activation of hormonal signaling pathways and the concomitant accumulation of defense metabolites underlie such AG-BG interactions. The main plant-mediated mechanisms regulating these reciprocal interactions via local and systemic induced responses remain poorly understood. We investigated the impact of root infection by the root-knot nematode (RKN) Meloidogyne incognita at different stages of its infection cycle, on tomato leaf defense responses triggered by the potato aphid Macrosiphum euphorbiae. In addition, we analyzed the reverse impact of aphid leaf feeding on the root responses triggered by the RKN. We focused specifically on the signaling pathways regulated by the phytohormones jasmonic acid (JA), salicylic acid (SA), abscisic acid (ABA), and indole-3-acetic acid (IAA) as well as steroidal glycoalkaloids as induced defense compounds. We found that aphid feeding did not induce AG hormonal signaling, but it repressed steroidal glycoalkaloids related responses in leaves, specifically when feeding on plants in the vegetative stage. Root infection by the RKN impeded the aphid-triggered repression of the steroidal glycoalkaloids-related response AG. In roots, the RKN triggered the SA pathway during the entire infection cycle and the ABA pathway specifically during its reproduction stage. RKN infection also elicited the steroidal glycoalkaloids related gene expression, specifically when it was in the galling stage. Aphid feeding did not systemically alter the RKN-induced defense responses in roots. Our results point to an asymmetrical interaction between M. incognita and Ma. euphorbiae when co-occurring in tomato plants. Moreover, the RKN seems to determine the root defense response regardless of a later occurring attack by the potato aphid AG.
Introduction
Plants encounter several species of insect herbivores and pathogens that reduce their fitness. To defend themselves against these attackers, plants have evolved multifaceted mechanisms to perceive and appropriately respond to the specific attackers, thus preventing or attenuating the attack (Mithöfer and Boland, 2008; War et al., 2012; Mortensen, 2013). Plant hormones regulate the plant’s immune system (Pieterse et al., 2012). Among them, jasmonic acid (JA) with its derivates (collectively called jasmonates; JAs) and salicylic acid (SA) are considered as major defense hormones (Pieterse et al., 2009, 2012; Erb et al., 2012). The activation of phytohormone related pathways occurs with considerable specificity. The JA pathway is typically (but not exclusively) activated upon the attack of chewing herbivores and necrotrophic pathogens, while piercing-sucking herbivores and biotrophic pathogens trigger the SA pathway (Walling, 2000; Zhu-Salzman et al., 2004; Howe and Jander, 2008; Diezel et al., 2009; Lemarié et al., 2015; Wasternack, 2015). While the JA and SA pathways form the backbone of the plant’s immune system, other hormones such as ethylene, abscisic acid (ABA), auxins, and cytokinins also contribute to defense signaling (Bari and Jones, 2009; Erb et al., 2012; Kammerhofer et al., 2015). These hormones can antagonistically or synergistically interact with the JA-SA backbone of the plant’s immune signaling network. This so-called hormone cross-talk provides the plant with a powerful capacity to finely regulate its immune response to the specific attacker (Pieterse et al., 2009; Li et al., 2019).
The induction of plant defense responses upon herbivory at local sites is often accompanied by systemic induced responses in distal tissues, thereby protecting undamaged plant parts from subsequent attack. Systemic signaling is not limited to the particular organ (roots or shoots) under attack, but it can cross the root-shoot interface. Several studies show that the attack by aboveground (AG) and belowground (BG) herbivores and pathogens leads to systemic responses mediated via the plant which influence organisms associated with the other organ. BG herbivores can induce systemic responses in AG plant parts that can facilitate or impede the performance of the AG insect herbivores. For example, root damage by the insect herbivore Tecia solanivora decreases the performance of the AG leaf chewers Spodoptera exigua and Spodoptera frugiperda when feeding on potato plants (Kumar et al., 2016). On the other side, root infection by the parasitic root nematode Globodera pallida increased the reproductive success of the AG-feeding aphid Myzus persicae (Hoysted et al., 2017). Although less studied, AG herbivory can also systemically influence the performance of herbivores feeding on BG plant parts. For example, simulated AG herbivory by Manduca sexta on Nicotiana attenuata enhanced the performance of the parasitic root nematode Meloidogyne incognita (Machado et al., 2018). In contrast, AG herbivory can also negatively affect BG-feeding herbivores. For example, leaf-feeding by Spodoptera littoralis on maize plants deterred larvae of Diabrotica virgifera virgifera from infesting the roots (Erb et al., 2015).
These AG-BG plant-mediated interactions are driven at least partially, by the activation of hormonal-related pathways triggered by the attacking herbivores. This leads to changes in plant primary metabolism as well as the production of compounds that are toxic or deterrent for the herbivores, at both local and systemic sites. For instance, root herbivory and root elicitation by exogenous application of JA result in the accumulation of secondary metabolites, including steroidal glycoalkaloids, glucosinolates, and nicotine in leaves of different plant species, including N. attenuata, Solanum tuberosum, and Cardamine hirsuta (Fragoso et al., 2014; Kumar et al., 2016; Bakhtiari et al., 2018). On the other hand, AG herbivory or elicitation by exogenous application of methyl jasmonate enhances JA biosynthesis and the accumulation of secondary metabolites, including steroidal glycoalkaloids, phenolic acids, and glucosinolates in roots of several plants, including tomato, potato, brussels sprouts, Brassica nigra, Solanum dulcamara, and maize (Hlywka et al., 1994; Soler et al., 2007; Abdelkareem et al., 2017; Calf et al., 2020; Karssemeijer et al., 2020; Mbaluto et al., 2020). Such systemically-triggered changes in plant defense compounds can drive the impact on the performance of herbivores in the opposite compartment (Kumar et al., 2016; Bakhtiari et al., 2018; van Dam et al., 2018; Karssemeijer et al., 2020).
Most of the studies addressing the systemic defense-responses elicited by AG and BG interacting attackers focus on insect herbivores. Moreover, the majority of these studies focus on the impact of one herbivore feeding on one organ (AG or BG) on the induced systemic responses and the effect of the herbivores feeding on the other organ (Erb et al., 2009; Kumar et al., 2016; Hoysted et al., 2017, 2018; Machado et al., 2018; van Dam et al., 2018; Karssemeijer et al., 2020). It remains less explored how plants integrate sequential BG and AG attack, and the resulting concomitant induced responses in AG and BG organs of the same plant (Kutyniok and Muller, 2012; McCarville et al., 2012; Kammerhofer et al., 2015). In this study, we analyzed the systemic induced defense responses underlying the reciprocal interaction between root-knot nematodes (RKNs) and AG-feeding aphids sharing the same host.
Root-knot nematodes are soil-inhabiting parasites that infect the roots of thousands of plants. As obligate root feeders, they spend most of their life inside roots, thereby significantly influencing root physiology. After egg hatching, the infective second-stage juveniles (J2s) penetrate their host roots and migrate in between cells to reach and settle in the vascular cylinder (Fenoll et al., 1997; Perry et al., 2009). They select several vascular cells to induce their feeding sites, commonly known as giant cells. As they feed and develop further to reach maturity, they secrete and inject effector molecules that cause hyperplasia and hypertrophy of cells surrounding the giant cells to form root galls. Throughout the development, RKNs manipulate the host’s phytohormonal signaling in order to suppress defense responses and establish a sink for nutrients (Gheysen and Mitchum, 2011, 2019). Remarkably, several studies demonstrate that root infection by RKNs also affects defense-related responses in AG plant parts (Hamamouch et al., 2011; Arce et al., 2017; van Dam et al., 2018). However, the studies dealing with the impact of RKNs on AG defenses are scarce and show contrasting results. For instance, in Arabidopsis thaliana and rice plants, root infection by different RKN species was found to both increase and decrease systemically in leaves the JA levels and the expression of marker genes in the JA and SA pathways, depending on the study systems (Hamamouch et al., 2011; Kyndt et al., 2017b).
Aphids are insect herbivores that in analogy to RKNs, feed directly on vascular content. They insert their mouthparts (stylet) in between the primary and secondary cells layers of the leaf to reach the sieve elements in the vascular tissues. Plants generally respond to aphid attack by activating the SA responsive pathway (Walling, 2000; De Vos et al., 2005), although some studies revealed the activation of the JA pathway upon aphids attack (Fidantsef et al., 1999) as well as the negative impact of JA elicitation on aphids feeding (Cooper and Goggin, 2005). Remarkably, it has been shown that aphids can trigger systemic induction of defenses in roots, and influence BG-associated biota. For example, AG herbivory by the aphid Brevicoryne brassicae triggered an increase in JA levels systemically in roots of Brassica oleracea, although this increase did not affect the development of root fly Delia radicum (Karssemeijer et al., 2020).
Because plant-parasitic root nematodes and aphids tap resources from the vascular tissues, they can affect each other via direct competition or by systemically triggering the plant’s defense system (Hol et al., 2013). In this study, we aimed to disentangle the molecular and chemical mechanisms driving the plant-mediated reciprocal interaction between RKNs and AG feeding aphids. With this aim, we established a bioassay including the important crop species tomato (Solanum lycopersicum) and addressed the effects of root infection by the RKN Ma. incognita on leaf defenses triggered by the potato aphid Macrosiphum euphorbiae; as well as the impact of leaf herbivory by Ma. euphorbiae on root defenses induced by M. incognita. Because the interactions between AG herbivores and M. incognita-induced plant defense responses are modulated by the RKN infection stages (Mbaluto et al., 2020), we studied the M. incognita-Ma. euphorbiae interaction during the different stages of the M. incognita infection cycle namely; invasion, galling, and reproduction. Our results show that M. incognita has a moderate systemic effect on defense responses triggered locally in leaves by Ma. euphorbiae. Our results further indicate that this systemic effect is modulated during the M. incognita root infection cycle. On the other hand, Ma. euphorbiae did not interfere systemically with the defense responses triggered by M. incognita locally in roots. Our findings suggest an asymmetrical interaction between M. incognita and Ma. euphorbiae when co-occurring in tomato plants, where M. incognita seems to determine the root defense response regardless of the AG Ma. euphorbiae attack.
Materials and Methods
Nematode and Aphid Cultures
We used the RKN M. incognita as the BG herbivore and the potato aphid Ma. euphorbiae as the AG herbivore. The M. incognita colony was initially obtained from Rijk Zwaan (De Lier, Netherlands) and maintained on tomato cv “Moneymaker” in a glasshouse. The colony was initiated from a single egg mass, and 8 weeks later, eggs were extracted for use in the experiments (Martínez-Medina et al., 2017). The potato aphid was kindly provided by Dr. Zeger van Herwijnen (Rijk Zwaan Breeding B.V De Lier, Netherlands). We maintained a laboratory colony using the leaf disc method (Rocca and Messelink, 2017) with slight modifications. In brief, we prepared 1% (w/v) water-agar and poured in plastic boxes 8 cm (length) × 5 cm (width) × 4 cm (height) to obtain ~0.5 cm thickness. A leaf disc from Capsicum annuum was embedded on the solidified agar with the abaxial side facing up to mimic normal aphid feeding side or position. The colony was maintained in a growth chamber (CLF PlantClimatic, CLF PlantClimatics GmbH, Wertingen, Germany) under 12-h light, 22°C: 12-h dark, 20°C, 45% relative humidity conditions. In the bioassays, we used apterous individuals.
Plant Material and Growth Conditions
We used tomato (S. lycopersicum) cultivar “Moneymaker,” as the study model plant. Tomato seeds were obtained from Intratuin B.V (Woerden, Netherlands). The seeds were sterilized, germinated, and transplanted, according to Mbaluto et al. (2020). In the glasshouse, the plants were randomly distributed and grown under 16-h light 25 ± 3°C: 8-h dark 22 ± 3, 40% relative humidity conditions. The plants were watered as required and supplemented weekly with half-strength Hoagland solution (Hoagland and Arnon, 1938). Four weeks after germination, we used the plants for the bioassays.
Nematode Inoculation and Aphid Infestation
In order to mimic the natural sequence of events, we infected plants with the RKN M. incognita first. Indeed, root feeders such as plant-parasitic nematodes are among the first pests encountered by annual plants; while AG feeders such as aphids generally arrive later in the plant’s life cycle (Bezemer and van Dam, 2005; van Dam et al., 2018). In all the bioassays, the plants assigned for M. incognita inoculation received 3,000 M. incognita eggs suspended in 1 ml of tap water (Mbaluto et al., 2020). Plants not assigned for M. incognita inoculation were mock-inoculated with 1 ml of tap water. We established three-time points after the M. incognita inoculation, corresponding to the main stages of its life cycle: 5 days post nematode inoculation (dpi), corresponding to the invasion stage; 15 dpi corresponding to the galling stage, and 30 dpi corresponding with the reproduction stage (Mbaluto et al., 2020). At each specific time point after M. incognita inoculation, plants assigned to the AG herbivore were challenged with 12 Ma. euphorbiae individuals of mixed-stages (adults and nymphs). The aphids were contained on a single leaf for 24 h, using a round clip cage of 7 cm in diameter. The clip cage was mounted on one fully expanded leaf: specifically on the three leaflets close to the tip (Bandoly and Steppuhn, 2016). Similarly, we mounted an empty clip cage on similar leaves, as mentioned above, on the plants not assigned for the aphid infestation. At each study time point, i.e., invasion, galling, and reproduction stage, we established four treatment groups including; (1) controls: plants not challenged with any of the herbivores, (2) plants root-infected with M. incognita, (3) plants infested on leaves with Ma. euphorbiae, and (4) plants infected with M. incognita in roots and infested with Ma. euphorbiae on leaves. Ten biological replicates of each treatment per time point were established, giving a total of 120 plants. At 24 h after infesting the plants with aphids, we harvested the plants, starting with the leaves and followed with the roots samples. We selected this specific time point (i.e., 24 h) in accordance with previous studies (Kafle et al., 2017), and also based on a pilot experiment in which the time points 12, 24, and 48 h were tested (data not shown). For root sampling, the entire root system was harvested. For the leaves, we harvested specifically the leaves that the aphids were feeding on, or the leaves that were mounted with empty clip cages without aphids, in the case of non-infested plants. Leaf and root material was stored at −80°C until use. In addition, after washing the root systems, we counted the number of galls visible to the naked eye from the root system of M. incognita infected plants. Approximately, the number of galls visible at the galling stage (15 dpi) averaged between 120 and 130 per plant. The number of visible galls had increased to 280–300 per plant at the reproduction stage (30 dpi).
Assessment of the Impact of Nematode Root Infection on Aphids Performance
We assessed the impact of M. incognita root infection on the reproduction of Ma. euphorbiae by comparing the number of nymphs produced by the aphids on tomato plants that were root infected or not with M. incognita. For this, we established a bioassay in which we inoculated tomato plants with M. incognita eggs as described above. Plants not assigned for M. incognita inoculation were mock-inoculated with 1 ml of tap water. We established three-time points after the M. incognita inoculation, coinciding with the invasion, galling, and reproduction stages of M. incognita, as described above. At each specific time point after M. incognita inoculation, we carefully placed three apterous Ma. euphorbiae adults using a soft-bristled brush on a similar leaf to the one used in the defense response experiments. We allowed the adult female aphids to feed on the plants and reproduce for 3 days, after which we counted the number of nymphs on the third day. This experiment was conducted twice, with similar results.
Phytohormone Extraction and Analysis
We extracted and quantified phytohormones from 100 mg of leaf and root material following the protocol previously described by Machado et al. (2013), with slight modifications. The extraction solution contained deuterated form of each phytohormone as the internal standards (i.e., D6-JA, D6-JA-Ile, D6-ABA, D5-IAA, and D6-SA). At the nebulization stage, the compounds were nebulized by electron spray ionization in the negative mode using the following conditions: capillary voltage 4,500 eV, cone gas 35 arbitrary units/350°C, probe gas 60 arbitrary units/300°C, and nebulizing gas at 60 arbitrary units. Data acquisition and processing were performed using the “MS data Review” software (Bruker MS Workstation, version 8.2, Bruker, Bremen, Germany). Phytohormone levels were calculated based on the peak area of the corresponding internal standard and the amount of fresh mass of plant material (ng−1 mg−1 FW), according to Mbaluto et al. (2020).
Real-Time Quantitative qPCR
Total RNA was extracted from ~100 mg (fresh weight) of ground leaf and root material, according to Oñate-Sánchez and Vicente-Carbajosa (2008). We performed quality check both quantitative and qualitative using a NanoPhotometer® P330 (Implen, Munich Germany) and by gel electrophoresis (1% agarose). We removed traces of DNA by treating 5 μg of the extracted RNA with 2 U/μl of DNaseI (Thermo Fisher Scientific, Schwerte, Germany) and following the manufacturer’s instructions. The clean RNA was rechecked for quality as stated above. We synthesized the first-strand cDNA from 1 μg DNase free RNA by reverse transcription using 200 U/μl Revert Aid H-minus RT (Thermo Fisher Scientific Baltic UAB, Vilnius, Lithuania) following the manufacturer instructions. The amplification cycle conditions for cDNA synthesis were: at 42°C for 60 min, 50°C for 15 min, and 70°C for 15 min using a Thermal cycler (Techne, Stone, United Kingdom). Real-time quantitative qPCR reactions and relative quantification of specific mRNA levels were performed using CFX 384 Real-Time PCR system (Bio-Rad Laboratories Inc., Singapore), and with gene-specific primers described in Supplementary Table 1. The RT-qPCR cycle conditions were: 2 min at 50°C, 2 min at 95°C, and 40 cycle of 15 s at 95°C, and 60 s at 60°C (Vos et al., 2015). Melting curves analysis was done to verify the amplification of each gene transcript. Three technical replicates of each sample were included in the RT-qPCR. The gene expression levels were determined by normalizing the data to the reference gene SIEF (X14449), which encodes for the tomato elongation factor 1α (Miranda et al., 2013; Martínez-Medina et al., 2017). The stability of the SIEF gene was previously evaluated in the different tissues (roots and leaves) and under the different experimental conditions (nematode and aphids challenge) analyzed here. Normalized gene expression data were analyzed by the 2−∆∆ct method (Livak and Schmittgen, 2001).
Extraction of Metabolites and Data Processing
We extracted ~100 mg fresh leaf and root material for metabolites analysis following the method described by Mbaluto et al. (2020) with slight modifications. In brief, the modifications included, using formic acid in methanol (0.05% v/v) as solvent B in the mobile phase. The separation and characterization of secondary metabolites were done according to Mbaluto et al. (2020). The data was processed in MS-DIAL (v. 4.00, RIKEN) according to Moreno-Pedraza et al. (2019) and with modification of several parameters including retention time-end = 12.5 min, mass range end = 1,500 mass to charge ratio (m/z), and the alignment parameter setting: retention time tolerance = 0.2 min. We generated two datasets (i.e., leaves and roots datasets) from which we selected all features with mass to charge ratio (m/z) of 576.3 and 578.4 at retention time 11–12 min for each study time point. These m/z values represent fragments of the main steroidal glycoalkaloids in tomato α-dehydrotomatine and α-tomatine (Cataldi et al., 2005).
Statistical Analysis
Datasets were analyzed using R software v 3.6.1 (R Core Development Team, 2019) unless indicated otherwise. Following three-way ANOVAs with factors M. incognita (Mi), Ma. euphorbiae (Me), and time (T) as model explanatory factors; two-way ANOVAs with M. incognita (Mi) and Ma. euphorbiae (Me) as factors were performed for each time point [invasion (5 dpi), galling (15 dpi), and reproduction (30 dpi) stages] to analyze the impact of plant herbivory on the phytohormones, steroidal glycoalkaloids, and the gene expression. Before the ANOVA analysis, all data sets were pre-screened for outliers using the interquartile range (IQR) method as a function in R. The number of outlying values varied between treatment groups from 0 to 2. In cases where the ANOVA results were significant, we detected the differences between the treatment groups using Tukey’s Honest Significant Difference (HSD) for multiple comparisons at p ≤ 0.05.
Results
Impact of Meloidogyne incognita Root Infection on Leaf Hormonal Responses Triggered by Macrosiphum euphorbiae Feeding
We first analyzed the local effect of Ma. euphorbiae herbivory on leaf hormonal responses when feeding on plants not inoculated with M. incognita. Because we used the M. incognita infection cycle stages [i.e., invasion (5 dpi), galling (15 dpi), and reproduction (30 dpi)] to time the experiment, the plants had different ages over the course of the experiment. This means that plants (young) infected by M. incognita as well as their respective control plants were 33 days-old when the M. incognita were at the invasion stage, those used when M. incognita were at the galling stage were 43 days-old (medium), and by the time the M. incognita had reached the reproduction stage, the plants were 58 days-old. We found that Ma. euphorbiae feeding did not alter the concentrations of JA-Ile, SA, and ABA compared to the control plants, regardless of plant age (Figures 1A–C,E–G,I–K; black vs. yellow boxplots; Supplementary Tables 2 and 8; JA levels were below the detection threshold). In agreement with the phytohormonal responses, Ma. euphorbiae feeding did not change the expression of Proteinase inhibitor II (PI II) and Pathogenesis-related protein 1 (PR1) compared to controls (Figure 2; black vs. yellow boxplots; Supplementary Tables 3 and 9). Remarkably, Ma. euphorbiae feeding on 8 weeks old plants increased the overall levels of IAA (main effect of Me; Supplementary Table 8, p = 0.024), but there was no significant difference when compared with control plants (Figures 1D,H,L; black vs. yellow boxplots).
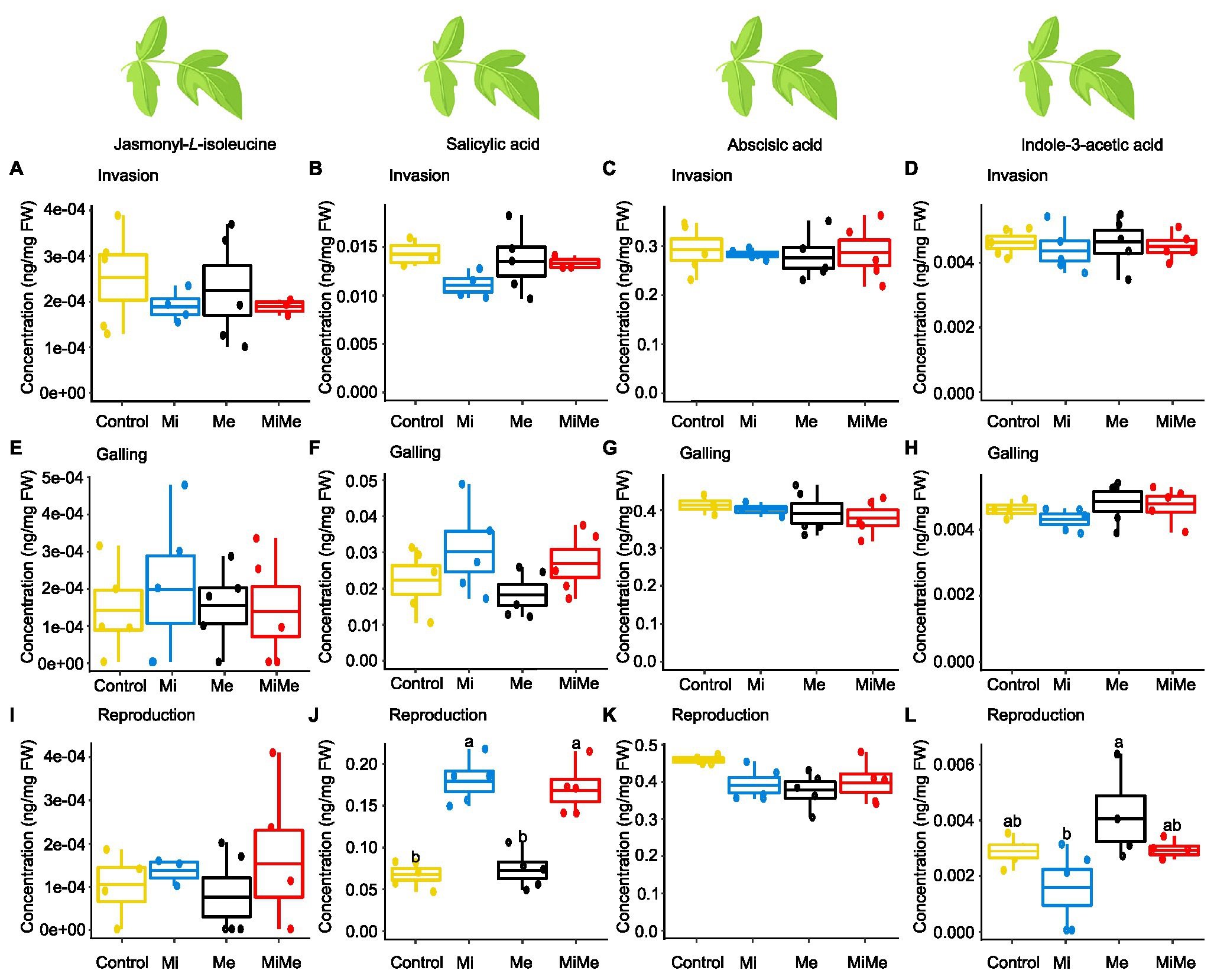
Figure 1. Phytohormones concentrations in tomato leaves upon aboveground and belowground herbivory. Mean concentrations (ng/mg fresh weight) of phytohormones in leaves of tomato plants infected belowground with Meloidogyne incognita (Mi), or infested aboveground with Macrosiphum euphorbiae (Me) or with both herbivores (MiMe). Control = plants without herbivores. Boxplots indicate the mean (±SEM, n = 5) concentrations of jasmonyl-L-isoleucine (A,E,I), salicylic acid (B,F,J), abscisic acid (C,G,K), and indole-3-acetic acid (D,H,L) measured at the nematodes’ invasion (A-D), galling (E-H), or reproduction (I-L) stages. Different letters above the boxplots indicate significant differences (p ≤ 0.05) in mean values between treatments, determined by Tukey's HSD test after ANOVA analysis.
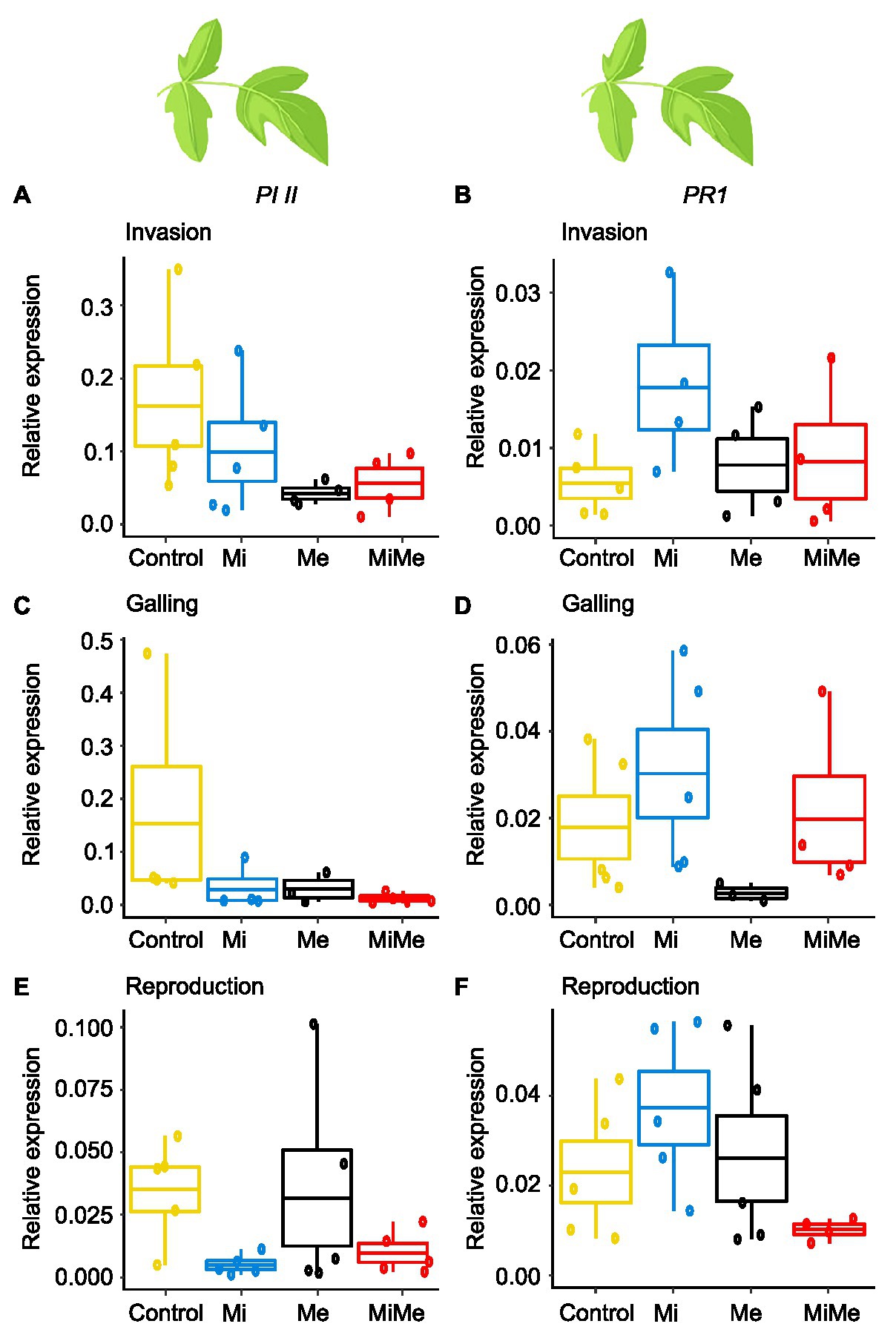
Figure 2. Expression of the jasmonic acid (JA) marker gene Proteinase inhibitor II (PI II) and the salicylic acid (SA) marker gene Pathogenesis-related protein 1 (PR1) in tomato leaves upon aboveground and belowground herbivory. Relative expression of PI II and PR1 genes were analyzed in leaves of tomato plants infected belowground with Meloidogyne incognita (Mi), or infested aboveground with Macrosiphum euphorbiae (Me) or with both herbivores (MiMe). Control = plants without herbivores. Boxplots indicate mean (±SEM, n = 5) expression values of PI II (A,C,E) and PR1 (B,D,F), measured at the nematodes’ invasion (A,B), galling (C,D), or reproduction (E,F) stages.
Next, we studied the systemic impact of M. incognita root infection, throughout its infection cycle, on leaf hormonal responses. Meloidogyne incognita root infection did not significantly affect the concentrations of JA-Ile, ABA, or IAA in tomato leaves compared to control plants at either of its infection cycle stages (Figures 1A,C–E,G–I,K,L; blue vs. yellow boxplots; Supplementary Tables 2 and 8). Despite a lack of JA-Ile response, M. incognita root infection overall downregulated the expression of the JA-responsive gene PI II compared to controls, as shown by a significant main effect (Supplementary Table 9, p = 0.028) in the reproduction stage (Figures 2A,C,E; blue vs. yellow boxplots; Supplementary Tables 3 and 9). Root infection by M. incognita significantly increased systemic SA levels compared to controls, specifically at the reproduction stage (Figure 1J; blue vs. yellow boxplot; Supplementary Tables 2 and 8). In contrast, the expression level of the SA-marker gene PR1 in M. incognita-infected plants was not significantly different from that observed in controls (Figures 2B,D,F; blue vs. yellow boxplots; Supplementary Tables 3 and 9).
To decipher the systemic effect of M. incognita root infection on AG phytohormonal-related responses triggered by Ma. euphorbiae leaf herbivory, we compared plants challenged by both M. incognita and Ma. euphorbiae to those challenged with Ma. euphorbiae alone at each stage of M. incognita root infection cycle [Figures 1, 2, red (MiMe) vs. black (Me) boxplots; Supplementary Tables 2, 3, 8, and 9]. The levels of JA-Ile, ABA, and IAA, as well as the expression levels of PI II and PR1 in leaves of co-infected plants were not significantly different from those infested with Ma. euphorbiae alone (Figures 1A,C–E,G–I,K,L, 2; red vs. black boxplots; Supplementary Tables 2, 3, 8, and 9). The SA levels were similar in Ma. euphorbiae and co-infected plants when M. incognita was at the invasion and galling stages (Figures 1B,F). Remarkably, when M. incognita was at the reproduction stage, the SA levels in co-infected plants increased compared to plants infested with Ma. euphorbiae alone (Figure 1J; red vs. black boxplots; Supplementary Tables 2 and 8).
Effect of Meloidogyne incognita Root Infection on Leaf Accumulation of Steroidal Glycoalkaloids Induced by Macrosiphum euphorbiae Feeding
Steroidal glycoalkaloids are important antiherbivore defense compounds in Solanaceae plants (Chowański et al., 2016). We first assessed the local effect of Ma. euphorbiae on leaf concentrations of the steroidal glycoalkaloids α-dehydrotomatine and α-tomatine, and the expression of the steroidal glycoalkaloid metabolism-related genes jasmonate-responsive ethylene response factor (ERF) 4 transcription factor (JRE4; encoding a master transcriptional regulator in defense-related steroidal glycoalkaloids) and glycoalkaloid metabolism 1 (GAME1; encoding a UDP-Gal:tomatidine galactosyltransferase biosynthetic gene) when feeding on plants of different ages (Figure 3). Leaf feeding by Ma. euphorbiae led to a decrease in the concentrations of α-dehydrotomatine and α-tomatine, in young plants compared to controls (Figures 3A,B; black vs. yellow boxplots; Supplementary Tables 4 and 10). In agreement, Ma. euphorbiae feeding on young plants significantly downregulated the expression of JRE4 (Figure 3C; black vs. yellow boxplot; Supplementary Tables 4 and 10). However, Ma. euphorbiae feeding did not affect GAME1 expression in young plants (Figure 3D; black vs. yellow boxplot; Supplementary Tables 4 and 10). In the medium age and old plants, infestation by Ma. euphorbiae did not significantly alter the concentrations of α-dehydrotomatine and α-tomatine nor the expression of JRE4 and GAME1 compared to controls (Figures 3E–L; black vs. yellow boxplots; Supplementary Tables 4 and 10). These findings show that Ma. euphorbiae represses the accumulation of α-dehydrotomatine and α-tomatine in tomato leaves, specifically when feeding on plants in the vegetative stage.
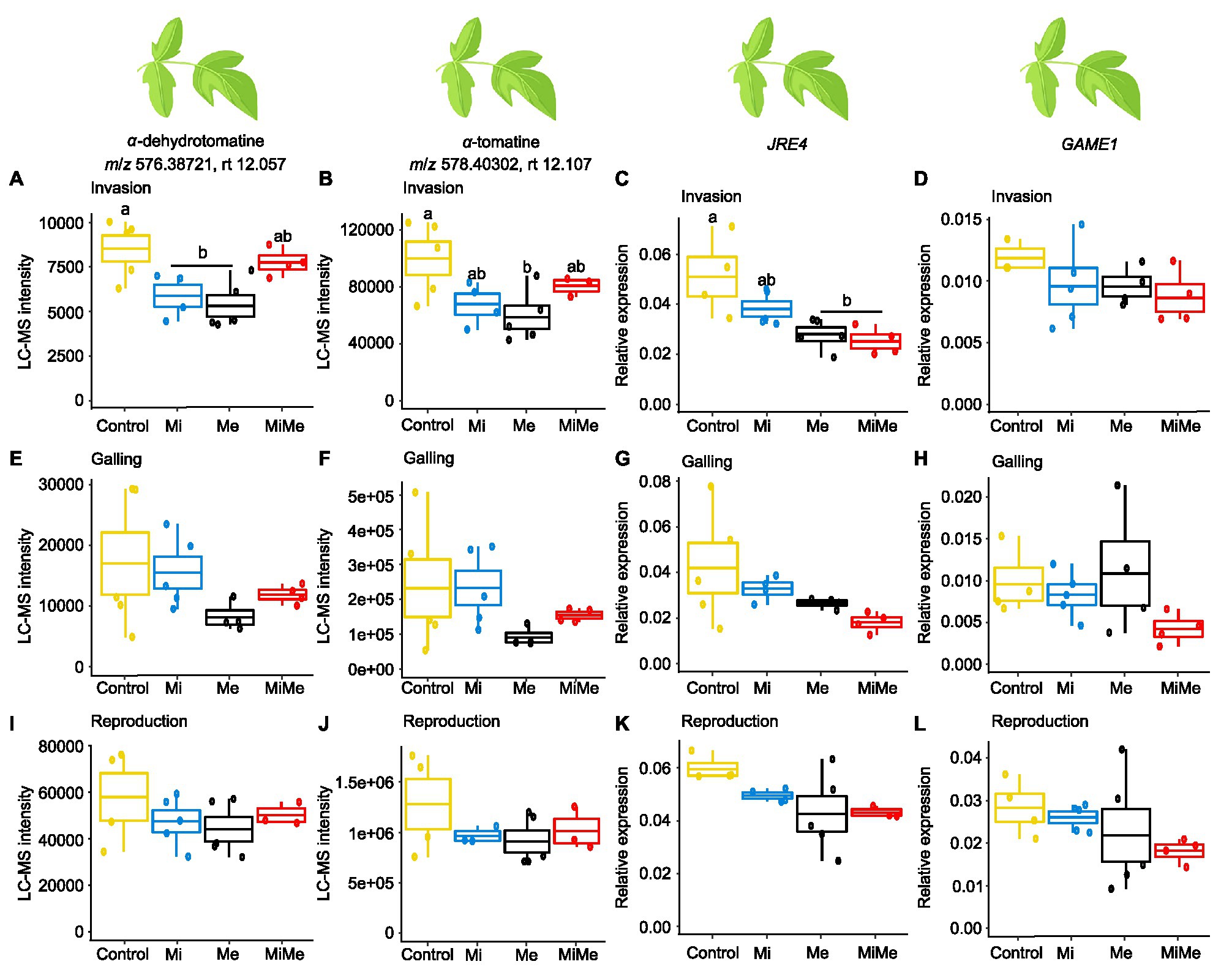
Figure 3. Relative intensities of the m/z signals of the steroidal glycoalkaloids α-dehydrotomatine and α-tomatine and relative expression of glycoalkaloid-related metabolism genes jasmonate-responsive ethylene response factor 4 (JRE4) and glycoalkaloid metabolism 1 (GAME1) in tomato leaves upon aboveground and belowground herbivory. Mean LC-MS intensities of α-dehydrotomatine (m/z 576.38721; rt. 12.057 min) and α-tomatine (m/z 578.40302; rt. 12.107 min) and expression of JRE4 and GAME1 in leaves tomato plants infected belowground with Meloidogyne incognita (Mi), or infested aboveground with Macrosiphum euphorbiae (Me) or with both herbivores (MiMe). Control = plants without herbivores. Boxplots are the mean (±SEM, n = 5) of α-dehydrotomatine (A,E,I), α-tomatine (B,F,J), JRE4 (C,G,K), and GAME1 (D,H,L) measured at the nematodes’ invasion (A-D), galling (E-H), or reproduction (I-L) stages. Different letters above the boxplots indicate significant (p ≤ 0.05) differences in mean values between treatments, determined by Tukey's HSD test after ANOVA analysis.
We next studied the systemic impact of M. incognita root infection throughout its infection cycle on the leaf concentration of α-dehydrotomatine and α-tomatine as well as on the expression levels of JRE4 and GAME1 genes. At the invasion stage, the leaf concentration of α-dehydrotomatine and α-tomatine decreased in M. incognita infected plants compared to control plants (Figures 3A,B; blue vs. yellow boxplots; Supplementary Tables 4 and 10). Although not statistically significant (Main effect of Mi in Supplementary Table 10, p = 0.089), we found a slight downregulation of JRE4 in leaves of plants that were infected with M. incognita at the invasion stage (Figure 3C; blue vs. yellow boxplot; Supplementary Tables 4 and 10). Meloidogyne incognita at the invasion stage did not affect the expression of the GAME1 gene in leaves compared to control plants (Figure 3D, blue vs. yellow boxplot; Supplementary Tables 4 and 10). At the M. incognita galling and reproduction stages, the leaf levels of α-dehydrotomatine and α-tomatine, as well as the expression of JRE4 and GAME1, were similar in M. incognita infected plants and control plants (Figures 3E–L; blue vs. yellow boxplots; Supplementary Tables 4 and 10). These results indicate that M. incognita triggers early and transient repression of the accumulation of α-dehydrotomatine and α-tomatine, specifically during the root invasion stage.
To check whether M. incognita root infection alters the repression of steroidal glycoalkaloid levels induced by Ma. euphorbiae in young (vegetative) plants, we compared plants challenged with Ma. euphorbiae alone to those co-infected with both M. incognita and Ma. euphorbiae at each of the M. incognita root infection cycle stages [Figure 3; red (MiMe) vs. black (Me) boxplots; Supplementary Tables 4 and 10]. At the invasion stage, co-infected plants had overall higher α-dehydrotomatine and α-tomatine levels than plants infested by Ma. euphorbiae alone (Figures 3A,B; red vs. black boxplots; Supplementary Tables 4 and 10 the interactive effect Mi∗Me). The expression of JRE4 and GAME1 in co-infected plants at the nematodes’ invasion stage was at a similar level to that in plants challenged with Ma. euphorbiae alone (Figures 3C,D; red vs. black boxplots; Supplementary Tables 4 and 10). At the galling and reproduction stages of M. incognita, the concentrations of α-dehydrotomatine and α-tomatine, as well as the expression of JRE4 and GAME1 in co-infected plants, were similar to those plants challenged with Ma. euphorbiae alone (Figures 3E–L; red vs. black boxplots; Supplementary Table 10). Our results indicate a moderate effect of M. incognita root infection on leaf steroidal glycoalkaloids associated with Ma. euphorbiae feeding.
Impact of Macrosiphum euphorbiae Leaf Feeding on Root Hormonal Related Responses Triggered by Meloidogyne incognita Infection
We first analyzed the local impact of M. incognita infection on root phytohormonal-related responses throughout its infection cycle. Meloidogyne incognita root infection did not significantly affect the level of JA, JA-Ile, or IAA in tomato roots compared to controls and regardless of the infection cycle stage (Figures 4A,B,E–G,J–L,O; blue vs. yellow boxplots; Supplementary Tables 5 and 11). Meloidogyne incognita infection did not affect the expression of PI II compared to controls, regardless of the infection cycle stage (Figures 5A,C,E; blue vs. yellow boxplots; Supplementary Tables 6 and 12). Meloidogyne incognita root infection triggered the root accumulation of SA at all infection stages when compared to controls (Figures 4C,H,M; blue vs. yellow boxplots; Supplementary Tables 5 and 11). However, M. incognita root infection did not alter PR1 expression compared to controls (Figures 5B,D,F; blue vs. yellow boxplots; Supplementary Tables 6 and 12). We found no differences in the levels of ABA in roots M. incognita-infected plants compared to controls when M. incognita was at the invasion and galling stages (Figures 4D,I; blue vs. yellow boxplots). However, at the reproduction stage, M. incognita significantly increased the ABA levels compared to control plants (Figure 4N; blue vs. yellow boxplot; Supplementary Tables 5 and 11).
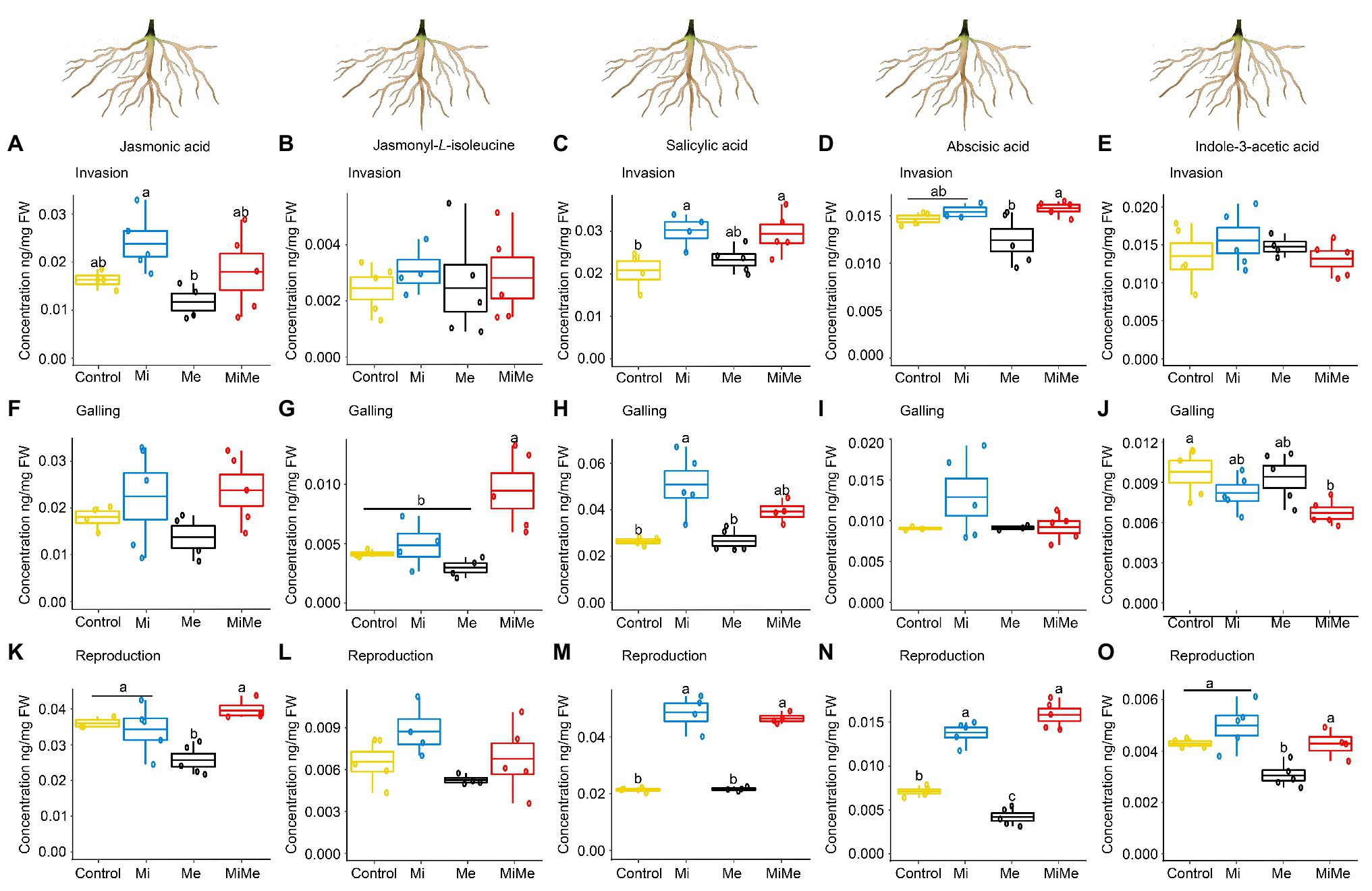
Figure 4. Phytohormones concentrations in tomato roots upon aboveground and belowground herbivory. Mean concentrations (ng/mg fresh weight) of phytohormones in roots of tomato infected belowground with Meloidogyne incognita (Mi), or infested aboveground with Macrosiphum euphorbiae (Me) or with both herbivores (MiMe). Control = plants without herbivores. Boxplots indicate the mean (±SEM, n = 5) of JA (A,F,K), jasmonyl-L-isoleucine (B,G,L), SA (C,H,M); abscisic acid (D,I,N) and indole-3-acetic acid (E,J,O) concentrations measured at the nematodes’ invasion (A-E), galling (F-J), or reproduction (K-O) stages. Different letters above the boxplots indicate significant differences (p ≤ 0.05) in mean values between treatments, determined by Tukey's HSD test after ANOVA analysis.
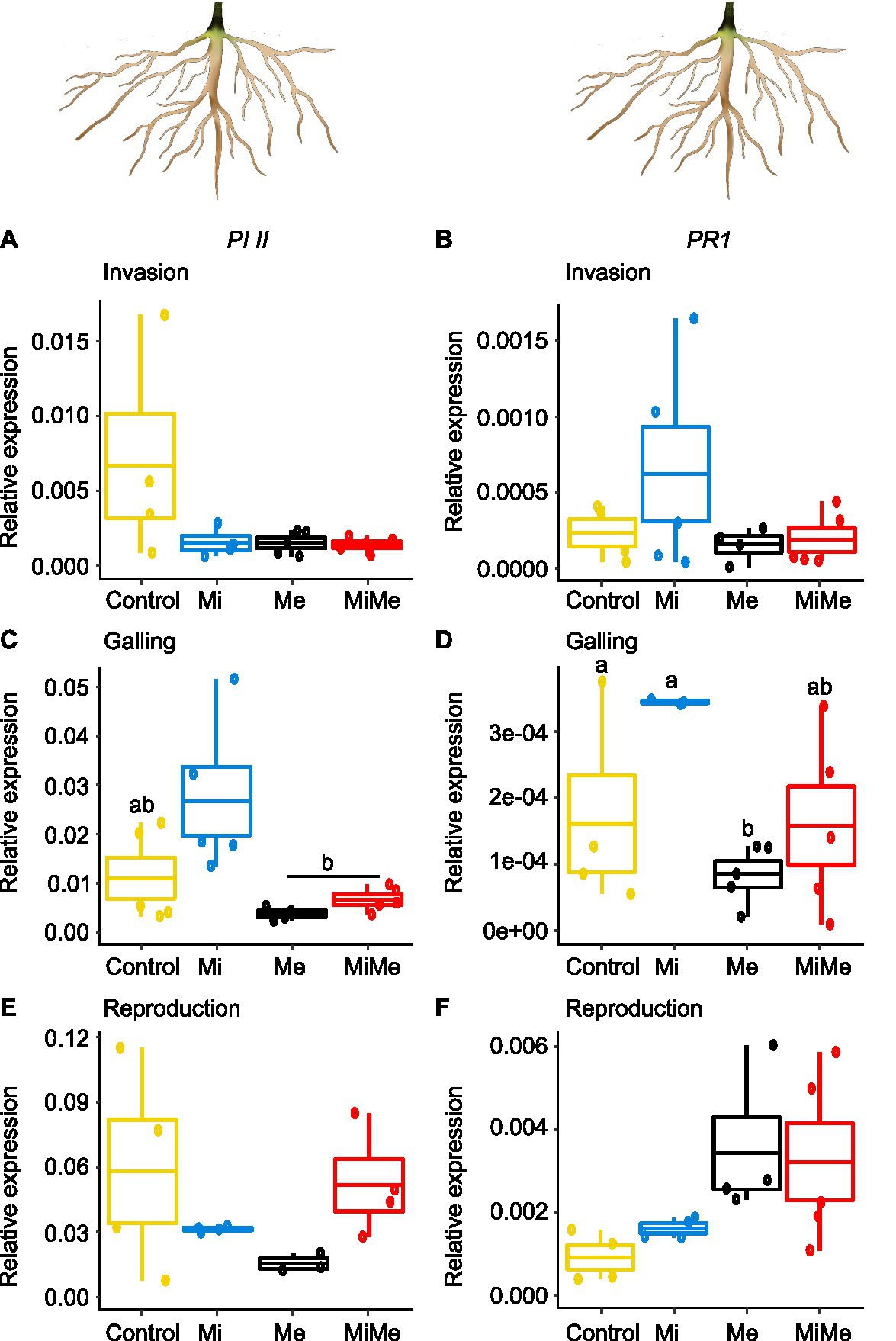
Figure 5. Expression of the jasmonic acid (JA) marker gene Proteinase inhibitor II (PI II) and the salicylic acid (SA) marker gene Pathogenesis-related protein 1 (PR1) in tomato roots upon aboveground and belowground herbivory. Relative expression of PI II and PR1 genes were analyzed in roots of tomato plants infected belowground with Meloidogyne incognita (Mi), infested aboveground with Macrosiphum euphorbiae (Me), or with both herbivores (MiMe). Control = plants without herbivores. Boxplots indicate mean (±SEM, n = 5) expression values of PI II (A,C,E) and PR1 (B,D,F) measured at the nematodes’ invasion (A,B), galling (C,D), or reproduction (E,F) stages. Different letters above the boxplots indicate significant differences (p ≤ 0.05) in mean expression among treatments, determined by Tukey's HSD test after ANOVA analysis.
Leaf herbivory by Ma. euphorbiae did not systemically affect root levels of JA-Ile or SA regardless of the plant age (Figures 4B,C,G,H,L,M; black vs. yellow boxplots; Supplementary Tables 5 and 11). We observed that JA, ABA, or IAA also did not systemically change when Ma. euphorbiae was feeding on young or medium-age (vegetative) plants (Figures 4A,D–F,I,J; black vs. yellow boxplots; Supplementary Tables 5 and 11). However, in old plants, Ma. euphorbiae feeding led to a significant decrease in the root levels of JA, ABA, and IAA (Figures 4K,N,O; black vs. yellow boxplots; Supplementary Tables 5 and 11). Macrosiphum euphorbiae feeding, in general, did not affect the expression level of PI II and PR1, regardless of plant age (Figure 5). Only in medium-aged plants, the expression levels of PR1 decreased in the roots of plants challenged with Ma. euphorbiae (Figure 5D; black vs. yellow boxplot; Supplementary Tables 6 and 12).
We finally assessed whether M. euphoribae feeding affected the phytohormonal root responses associated with M. incognita root infection [Figures 4, 5; red (MiMe) vs. blue (Mi) boxplots; Supplementary Tables 5, 6, 11, and 12]. Roots infected by M. incognita alone had similar levels of JA, SA, ABA, and IAA as roots of plants co-infected with M. incognita and Ma. euphorbiae (Figures 4A,C–F,H–K,M–O; red vs. blue boxplots; Supplementary Tables 5 and 11). Similar to the SA levels, the expression of PR1 in M. incognita-infected roots did not differ from that in roots of co-infected plants (Figures 5B,D,F; red vs. blue boxplots; Supplementary Tables 6 and 12). The levels of JA-Ile in the roots of co-infected plants were similar to that on roots of plants infected with M. incognita alone at the invasion and reproduction stages (Figures 4B,L). However, when M. incognita was at the galling stage, a higher level of JA-Ile was observed in the root of co-infected plants compared to roots of plants infected with M. incognita alone (Figure 4G; red vs. blue boxplot; Supplementary Tables 5 and 11). By contrast, a higher expression level of PI II was found in the roots of M. incognita-infected plants compared to expression in co-infected plants at the galling stage. At the invasion and reproduction stages, PI II expression was similar in M. incognita and co-infected plants (Figures 5A,E; red vs. blue boxplots; Supplementary Tables 6 and 12).
Effect of Macrosiphum euphorbiae Leaf Feeding on Root Steroidal Glycoalkaloids Induced by Meloidogyne incognita Infection
We first analyzed the impact of M. incognita on the root concentration of α-dehydrotomatine and α-tomatine as well as on the expression of JRE4 and GAME1 genes throughout its infection cycle. Meloidogyne incognita root infection at the invasion and reproduction stages did not significantly affect the root level of α-dehydrotomatine and α-tomatine or the expression of JRE4 and GAME1 compared to controls (Figures 6A–D,I–L; blue vs. yellow boxplots; Supplementary Tables 7 and 13). When M. incognita was in the galling stage, its feeding increased the level of α-dehydrotomatine and α-tomatine and the expression of JRE4 and GAME1 compared to controls (Figures 6E–H; blue vs. yellow boxplots; Supplementary Tables 7 and 13).
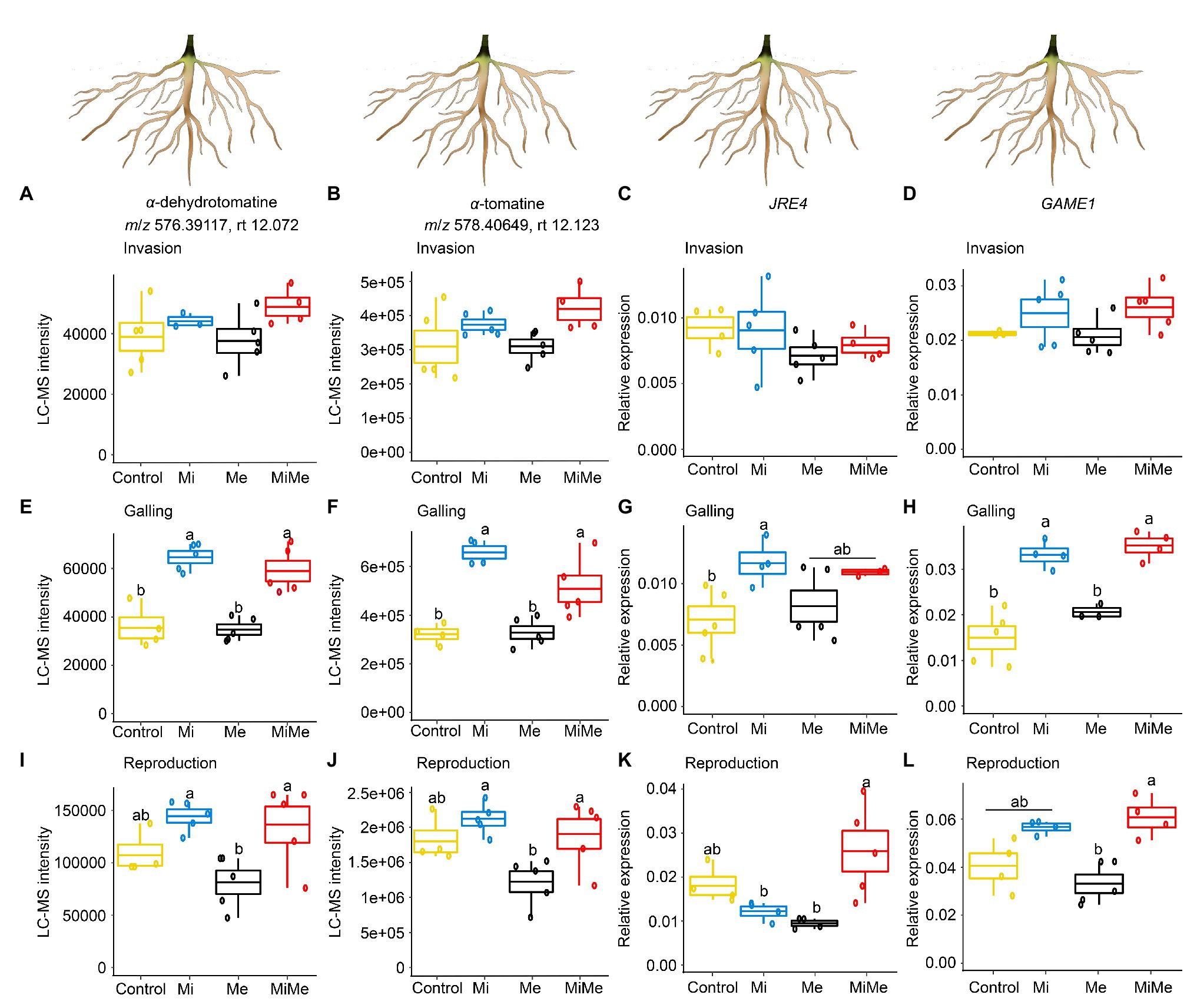
Figure 6. Relative intensities of the m/z signals of the steroidal glycoalkaloids α-dehydrotomatine and α-tomatine and relative expression of glycoalkaloid-related metabolism genes jasmonate-responsive ethylene response factor (ERF) 4 transcription factor (JRE4) and glycoalkaloid metabolism 1 (GAME1) in tomato roots upon aboveground and belowground herbivory. Mean LC-MS intensities of α-dehydrotomatine (m/z 576.39117; rt. 12.072 min) and α-tomatine (m/z 578.40649; rt. 12.123 min) and expression of JRE4 and GAME1 in roots of tomato plants infected belowground with Meloidogyne incognita (Mi), infested aboveground with Macrosiphum euphorbiae (Me), or with both herbivores (MiMe). Control = plants without herbivores. Boxplots indicate the mean (±SEM, n = 5) of α-dehydrotomatine (A,E,I), α-tomatine (B,F,J) m/z intensities, JRE4 (C,G,K), and GAME1 (D,H,L) measured at the nematodes’ invasion (A-D), galling (E-H), or reproduction (I-L) stages. Different letters above the boxplots indicate significant differences (p ≤ 0.05) in mean values between treatments, determined by Tukey's HSD test after ANOVA analysis.
We then assessed the systemic impact of Ma. euphorbiae leaf herbivory on the root defense expression. Leaf herbivory by Ma. euphorbiae did not affect the level of α-dehydrotomatine and α-tomatine, nor the expression of JRE4 and GAME1, regardless of plant age (Figure 6; black vs. yellow boxplots; Supplementary Tables 7 and 13). These results indicate that leaf feeding by Ma. euphorbiae does not systemically alter the steroidal glycoalkaloids metabolism pathway in tomato roots.
Finally, we analyzed whether Ma. euphorbiae feeding systemically affects the root levels of steroidal glycoalkaloids and the expression patterns of GAME genes associated with M. incognita root infection. In general, the levels of α-dehydrotomatine and α-tomatine, and the expression of JRE4 and GAME1 of M. incognita-infected roots were similar to those in co-infected plants, regardless of the M. incognita infection cycle stage [Figure 6; red (MiMe) vs. blue (Mi) boxplots; Supplementary Tables 7 and 13]. Only in the case of JRE4 expression, a higher expression level was found in the roots of co-infected plants compared to the roots of plants infected with M. incognita at the reproduction stage (Figure 6K; red vs. blue boxplot; Supplementary Tables 7 and 13). Overall, these results show that Ma. euphorbiae leaf herbivory has only a minor effect on root steroidal glycoalkaloid induction associated with M. incognita root infection.
Impact of Meloidogyne incognita Root Infection on the Reproduction of Macrosiphum euphorbiae
We found an overall stronger effect of M. incognita root infection on the plant responses triggered by Ma. euphorbiae in leaves, compared to the reverse interaction (i.e., the reciprocal effect of Ma. euphorbiae feeding on root induced responses by M. incognita infection). Because of this, we next aimed to assess the impact of M. incognita root infection on the reproduction rate of Ma. euphorbiae. Similar to the defense response bioassays, we used the M. incognita infection cycle stages to time the performance bioassay. On plants without M. incognita root infection, we found that the numbers of nymphs produced on young and medium-age plants was significantly higher compared to those found on old (flowering) plants (Table 1). In plants that were challenged with M. incognita, we found a similar number of nymphs, at every nematode root infection stage compared to those observed in plants without M. incognita (Table 1).
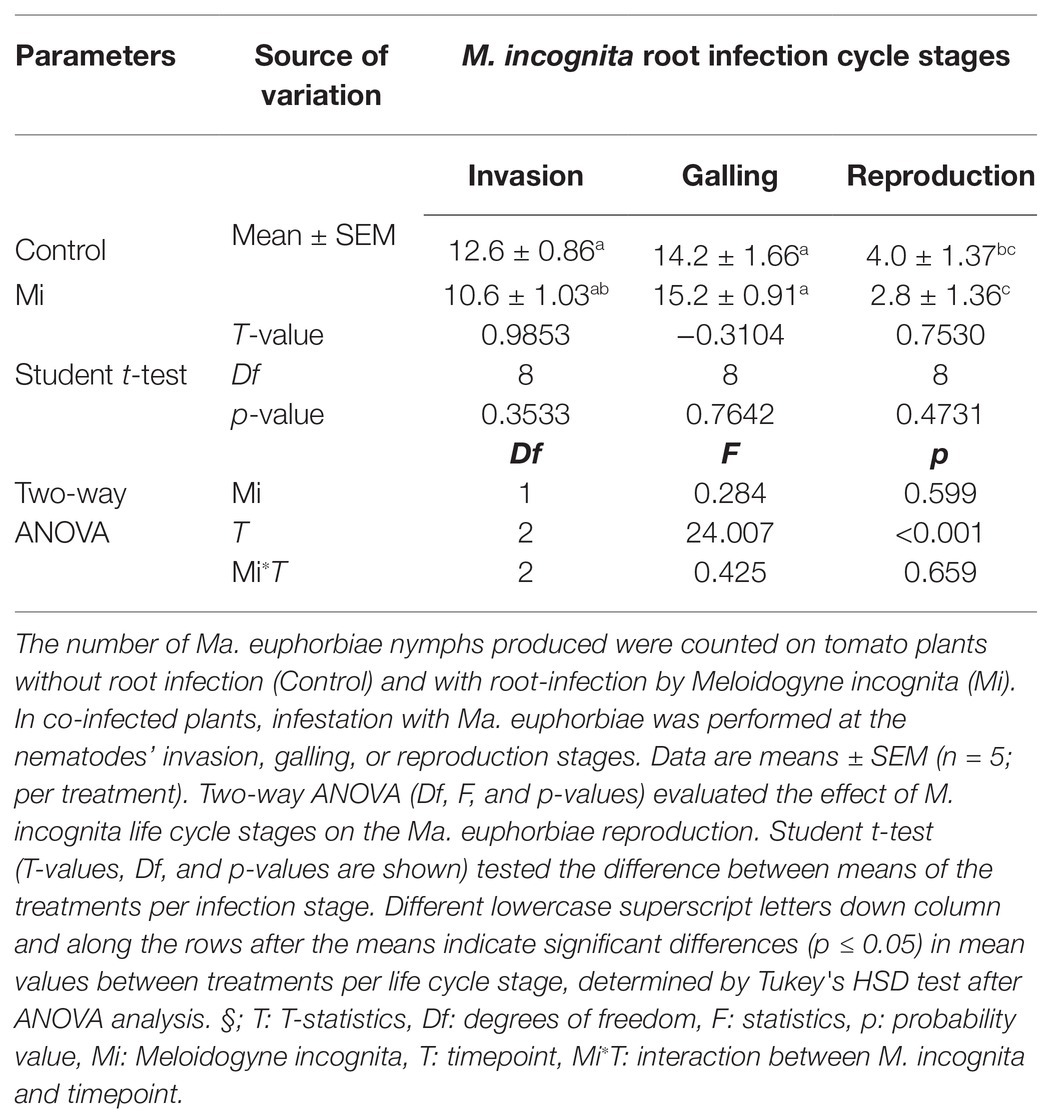
Table 1. Number of nymphs produced by Macrosiphum euphorbiae adults on tomato plants.
Discussion
Here, we used tomato as a model plant, to explore how root infection by M. incognita affects the leaf responses triggered by Ma. euphorbiae, and the reciprocal impact of leaf herbivory by Ma. euphorbiae on root responses induced by M. incognita infection in roots. Because root responses to M. incognita infection are tightly modulated during its infection cycle stages (Mbaluto et al., 2020), we studied the dynamics of the interaction between the induced plant responses by the two herbivores during the entire M. incognita root infection cycle. We show that root infection by M. incognita had mild systemic effects on phytohormones and steroidal glycoalkaloid responses triggered by Ma. euphorbiae locally on leaves. On the reverse, leaf-feeding by Ma. euphorbiae did not interfere systemically with the defense responses triggered by M. incognita locally in roots. In both interaction directions, the induction of defense responses occurred depending on the M. incognita root infection cycle stages. Collectively, our results indicate that root infection by M. incognita induces a strong effect in roots that is not overruled by AG Ma. euphorbiae feeding. They also demonstrate that the root infection cycle of M. incognita is an important factor influencing the dynamics of the interaction between the two herbivores.
We found that feeding by Ma. euphorbiae did not significantly affect phytohormonal signaling locally in leaves. In contrast to our results, several studies revealed that plants can activate the SA pathway upon attack by aphids, including Ma. euphorbiae (Mohase and van der Westhuizen, 2002; Chaman et al., 2003; Kuśnierczyk et al., 2008; Coppola et al., 2013). For instance, an increase in the expression of SA-responsive genes has been reported in A. thaliana upon the attack by M. persicae (Moran and Thompson, 2001) and by Schizaphis graminum on aphid-susceptible barley (Chaman et al., 2003). Moreover, the attack by M. euphobiae, B. brassicae, or M. persicae triggered the expression of both SA- and JA-responsive genes in A. thaliana and tomato plants (de Ilarduya et al., 2003; Kuśnierczyk et al., 2008; Coppola et al., 2013). Although we do not have a clear explanation for such apparent discrepancies with our results, the different outcomes may be partly explained by differences in the experimental set-ups, including the number and different stages of aphids or the duration of the experiments. Moreover, the differences in the studies can be due to the fact that piercing-sucking herbivores may antagonize defense responses to make the plant a more suitable host, depending on the system under investigation (Walling, 2008). Aphid salivary secretions contain effector proteins that may suppress defense responses (Hogenhout and Bos, 2011; Kettles and Kaloshian, 2016). In line with this, we found that Ma. euphorbiae infestation reduced the levels of the steroidal glycoalkaloids α-dehydrotomatine and α-tomatine and the expression of steroidal glycoalkaloid-related gene JRE4 in leaves. Solanum alkaloids have a broad range of biological activity against insect herbivores, including aphids (Chowański et al., 2016). Thus, our results suggest the ability of Ma. euphorbiae to manipulate the secondary chemistry of the host plant to its benefit. Previous studies showed that aphids, including Ma. euphorbiae and M. perciase, can decrease secondary metabolites as well as trigger the downregulation of a set of alkaloid biosynthesis genes in tomato and A. thaliana (Mewis et al., 2012; Coppola et al., 2013). Interestingly, in this study, the aphid-triggered decrease in steroidal glycoalkaloids was specifically observed when the aphid fed on plants at the vegetative stage (young and medium-aged plants). By contrast, Ma. euphorbiae failed to counteract the steroidal glycoalkaloid-related responses in plants at the flowering stage (old plants). This indicates that plant age and ontogeny are important factors determining the ability of Ma. euphorbiae to modulate defense responses in tomato plants. In accordance to the inability of Ma. euphorbiae to suppress steroidal glycoalkaloid-related responses in flowering plants, we found that it performed worse when feeding on plants at the flowering stage (old plants), compared to plants in the vegetative stage. This is further evidence that the suppression of steroidal glycoalkaloid-related responses in local tissues can be important for aphid’s performance.
Whereas Ma. euphorbiae feeding did not induce phytohormonal responses locally in leaves, it systemically decreased the levels of JA, ABA, and IAA in roots, suggesting that this aphid might alter the allocation of defenses between roots and leaves. It has been previously demonstrated that aphids can reduce aliphatic glucosinolates in the roots. This led to a shift in the ratio of aliphatic and indole glucosinolates in systemic tissues, indicating that plants alter the allocation of defense compounds upon aphid attack (Kutyniok and Muller, 2012). Remarkably, the systemic impact of Ma. euphorbiae on root phytohormonal responses was only observed when it fed on flowering plants (old plants). In the same plants, we observed a trend for reduced levels of steroidal glycoalkaloids in roots. This suggests that plant age and ontogeny are also important factors influencing the systemic effect of Ma. euphorbiae leaf herbivory on root responses. Possibly, plants prioritize the allocation of defenses to reproductive tissues over vegetative tissues after herbivory (Chrétien et al., 2018). However, the ecological consequences of the decrease in the levels of phytohormones and glycoalkaloids triggered systemically by the aphids in the roots of the flowering tomato plants remain unclear.
Root infection by M. incognita triggered an increase of SA levels locally in roots throughout the entire infection cycle. Local accumulation of SA in roots upon the infection by different RKN species was found in several plant species, including A. thaliana, rice, and tomato (Branch et al., 2004; Hamamouch et al., 2011; Kumari et al., 2016; Guo and Ge, 2017). Meloidogyne incognita infection further led to an increase in ABA levels, specifically when it was at the reproduction stage. Increases in ABA are associated with increasing the susceptibility to Meloidogyne infection (Kyndt et al., 2017a). Therefore, we speculate that the increase in ABA levels triggered by M. incognita at the reproduction stage might be related to an enhancement of host susceptibility to the next generation of infective juveniles.
Besides the changes in phytohormone levels M. incognita infection also altered the steroidal glycoalkaloid response locally in roots. Indeed, specifically at the galling stage, M. incognita triggered an increase in the levels of the steroidal glycoalkaloids α-dehydrotomatine and α-tomatine and upregulated the expression of the steroidal glycoalkaloid related genes JRE4 and GAME1. In agreement with our results, an increase in α-tomatine levels has been reported in tomato plants infected by M. incognita at the galling stage (Elliger et al., 1988). The induction of steroidal glycoalkaloids is associated with enhanced resistance to root infecting plant-parasitic nematodes (Wang et al., 2012; Jang et al., 2015). Therefore, the relevance of the increased steroidal glycoalkaloid levels in the present study remains unclear.
We further found that M. incognita root infection had just mild effects on the hormonally regulated pathways systemically in the leaves. Such a mild effect of root infecting plant-parasitic nematodes on systemic phytohormone signaling in AG tissues has been previously observed (Hamamouch et al., 2011; Kutyniok and Muller, 2012; Hoysted et al., 2017). On the other hand, M. incognita root infection reduced the levels of steroidal glycoalkaloids systemically in leaves, although this effect was significant just for α-dehydrotomatine in the invasion stage. Although it is unclear how systemic repression of host plant defenses can benefit nematode parasitism, some nematode effectors can suppress systemic signaling of defense responses in AG (Kyndt et al., 2014). These results indicate that M. incognita can cause subtle systemic changes in major defense compounds of tomato.
In co-infected plants, infection with M. incognita, in general, did not affect the leaf phytohormonal profile associated with Ma. euphorbiae leaf-feeding. However, the increase in SA levels triggered systemically in leaves by M. incognita infection at the reproduction stage was also evident in leaves of plants that were co-infected with both herbivores. This indicates that Ma. euphorbiae was unable to counteract SA signaling triggered systemically by M. incognita root infection. M. incognita infection further affected the steroidal glycoalkaloid-related responses triggered by Ma. euphorbiae feeding on plants at the vegetative stage. Indeed, the levels of α-dehydrotomatine and α-tomatine in leaves of co-infected plants were in between the levels found in controls and Ma. euphorbiae plants. This indicates that M. incognita infection counteracted, at least partially, the decrease in the levels of steroidal glycoalkaloids triggered by Ma. euphorbiae. Remarkably, these interactions did not affect the performance of the aphids. Previous studies show that SA levels can increase AG after root infection by plant-parasitic nematodes, but these changes differentially affect AG piercing-sucking insect herbivores. Guo and Ge (2017) reported an increase in SA levels in leaves of tomato plants infected by M. incognita in roots, which was concomitant with a reduction in the performance of whiteflies (Bemisia tabaci). On the other hand, Hoysted et al. (2017) found an increase in SA level in leaves of potato plants that were infected with G. pallida in roots, which correlated with the greater reproductive success of M. persicae. Taking together these studies, the variations in the findings can be attributed to differences in herbivores specialization and the plant system under investigation.
In roots, Ma. euphorbiae did not affect the phytohormonal profile associated with M. incognita root infection. The increased SA levels triggered by M. incognita throughout its infection cycle were still evident in the roots of co-infected plants. In analogy, Ma. euphorbiae did not interfere with the increased ABA levels triggered by M. incognita at the reproduction stage, even when Ma. euphorbiae infestation alone decreased the JA, ABA, and IAA levels systemically in roots. In line with this, Ma. euphorbiae feeding did not interfere with the increased levels of steroidal glycoalkaloids triggered by M. incognita at the galling stage. This further corroborates that the local effect of M. incognita determined the plant responses regardless of the later arriving herbivore Ma. euphorbiae. However, in our experimental design, the aphids were feeding for a limited time (24 h). Therefore, we cannot rule out a possible effect of Ma. euphorbiae on M. incognita-triggered plant responses nor on the performance of M. incognita at a later time points after aphid infestation.
In conclusion, we found that both M. incognita and Ma. euphorbiae triggered different local and systemic defense responses in tomato plants. When both herbivores co-occurred, M. incognita caused mild systemic effects on the induced plant responses to Ma. euphorbiae herbivory in leaves, which were not associated with changes in aphid’s performance. On the other hand, M. incognita-induced local root responses were not overruled by the systemic effect caused by Ma. euphorbiae leaf feeding, suggesting an asymmetrical interaction between M. incognita and Ma. euphorbiae when co-occurring in tomato plants.
Data Availability Statement
The data underlying this study are published as open access at the iDiv Data Repository (https://idata.idiv.de/ddm/Data/ShowData/1876?version=0). The data can be cited as CMM, AMM, and NMvD (2021). Above and belowground systemic induced plant responses to root infection by root-knot nematode and leaf feeding by the potato aphid. iDiv Data Repository (https://doi.org/10.25829/idiv.1876-13-8394).
Author Contributions
CMM, NMvD, and AMM conceived the research idea and designed the experiment. CMM and EMA performed the experiments. CMM, EMA, AMM, and KG processed the samples and analyzed the data. CMM conducted the literature search and wrote the initial manuscript with support of AMM and NMvD. All authors contributed to writing, reviewing, and approval of the final manuscript for submission.
Funding
All authors gratefully acknowledge the support of German Centre for Integrative Biodiversity Research (iDiv) Halle-Jena-Leipzig funded by the German Research Foundation (DFG – FZT 118, 202548816). CMM acknowledges the Deutscher Akademischer Austauschdienst (DAAD) for a doctoral research grant (research grant No: 91607343). EMA acknowledges the DAAD Short-Term Scholarship Program (GERSS) scholarship awarded to her to conduct part of her Master of Science Research at iDiv. AMM further acknowledges funding from the program for attracting talent to Salamanca from Fundación Salamanca Ciudad de Cultura y Saberes and Ayuntamiento de Salamanca.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Dr. Zeger van Herwijnen of Rijk-Zwaan Breeding B.V (De Lier, Netherlands), for providing us with the potato aphids Macrosiphum euphorbiae. We acknowledge Dr. Fredd Vergara for his technical support in chemical analysis. Dr. Anahita Kazem and the iDiv Data & Code Unit for support in data preparation, curation and archiving. Lastly, we thank the two reviewers Dr. Mark G. Sterken and Dr. Krzysztof Wieczorek, as well as section editor Dr. Colette Broekgaarden for their helpful and constructive comments on the earlier versions of the manuscript.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2021.632212/full#supplementary-material
References
Abdelkareem, A., Thagun, C., Nakayasu, M., Mizutani, M., Hashimoto, T., and Shoji, T. (2017). Jasmonate-induced biosynthesis of steroidal glycoalkaloids depends on COI1 proteins in tomato. Biochem. Biophys. Res. Commun. 489, 206–210. doi: 10.1016/j.bbrc.2017.05.132
Arce, C. C. M., Machado, R. A. R., Ribas, N. S., Cristaldo, P. F., Ataíde, L. M. S., Pallini, Â., et al. (2017). Nematode root herbivory in tomato increases leaf defenses and reduces leaf miner oviposition and performance. J. Chem. Ecol. 43, 120–128. doi: 10.1007/s10886-016-0810-z
Bakhtiari, M., Glauser, G., and Rasmann, S. (2018). Root JA induction modifies glucosinolate profiles and increases subsequent aboveground resistance to herbivore attack in Cardamine hirsuta. Front. Plant Sci. 9:1230. doi: 10.3389/fpls.2018.01230
Bandoly, M., and Steppuhn, A. (2016). Bioassays to investigate the effects of insect oviposition on a plant’s resistance to herbivores. Bio Proto. 6:e1823. doi: 10.21769/BioProtoc.1823
Bari, R., and Jones, J. D. G. (2009). Role of plant hormones in plant defence responses. Plant Mol. Biol. 69, 473–488. doi: 10.1007/s11103-008-9435-0
Bezemer, T. M., and van Dam, N. M. (2005). Linking aboveground and belowground interactions via induced plant defenses. Trends Ecol. Evol. 20, 617–624. doi: 10.1016/j.tree.2005.08.006
Branch, C., Hwang, C.-F., Navarre, D. A., and Williamson, V. M. (2004). Salicylic acid is part of the Mi-1-mediated defense response to root-knot nematode in tomato. Mol. Plant-Microbe Interact. 17, 351–356. doi: 10.1094/MPMI.2004.17.4.351
Calf, O. W., Lortzing, T., Weinhold, A., Poeschl, Y., Peters, J. L., Huber, H., et al. (2020). Slug feeding triggers dynamic metabolomic and transcriptomic responses leading to induced resistance in Solanum dulcamara. Front. Plant Sci. 11:803. doi: 10.3389/fpls.2020.00803
Cataldi, T. R. I., Lelario, F., and Bufo, S. A. (2005). Analysis of tomato glycoalkaloids by liquid chromatography coupled with electrospray ionization tandem mass spectrometry. Rapid Commun. Mass Spectrom. 19, 3103–3110. doi: 10.1002/rcm.2176
Chaman, M. E., Copaja, S. V., and Argandoña, V. H. (2003). Relationships between salicylic acid content, phenylalanine ammonia-lyase (PAL) activity, and resistance of barley to aphid infestation. J. Agric. Food Chem. 51, 2227–2231. doi: 10.1021/jf020953b
Chowański, S., Adamski, Z., Marciniak, P., Rosiński, G., Büyükgüzel, E., Büyükgüzel, K., et al. (2016). A review of bioinsecticidal activity of solanaceae alkaloids. Toxins 8:60. doi: 10.3390/toxins8030060
Chrétien, L. T. S., David, A., Daikou, E., Boland, W., Gershenzon, J., Giron, D., et al. (2018). Caterpillars induce jasmonates in flowers and alter plant responses to a second attacker. New Phytol. 217, 1279–1291. doi: 10.1111/nph.14904
Cooper, W. R., and Goggin, F. L. (2005). Effects of jasmonate-induced defenses in tomato on the potato aphid, Macrosiphum euphorbiae. Entomol. Exp. Appl. 115, 107–115. doi: 10.1111/j.1570-7458.2005.00289.x
Coppola, V., Coppola, M., Rocco, M., Digilio, M. C., D’Ambrosio, C., Renzone, G., et al. (2013). Transcriptomic and proteomic analysis of a compatible tomato-aphid interaction reveals a predominant salicylic acid-dependent plant response. BMC Genomics 14:515. doi: 10.1186/1471-2164-14-515
de Ilarduya, O. M., Xie, Q., and Kaloshian, I. (2003). Aphid-induced defense responses in Mi-1-mediated compatible and incompatible tomato interactions. Mol. Plant-Microbe Interact. 16, 699–708. doi: 10.1094/MPMI.2003.16.8.699
de Miranda, V., Coelho, R., Viana, A. A., de Oliveira Neto, O., Carneiro, R. M. D., Rocha, T., et al. (2013). Validation of reference genes aiming accurate normalization of qPCR data in soybean upon nematode parasitism and insect attack. BMC. Res. Notes 6:196. doi: 10.1186/1756-0500-6-196
De Vos, M., Van Oosten, V. R., Van Poecke, R. M. P., Van Pelt, J. A., Pozo, M. J., Mueller, M. J., et al. (2005). Signal signature and transcriptome changes of Arabidopsis during pathogen and insect attack. Mol. Plant-Microbe Interact. 18, 923–937. doi: 10.1094/MPMI-18-0923
Diezel, C., von Dahl, C. C., Gaquerel, E., and Baldwin, I. T. (2009). Different lepidopteran elicitors account for cross-talk in herbivory-induced phytohormone signaling. Plant Physiol. 150, 1576–1586. doi: 10.1104/pp.109.139550
Elliger, C. A., Waiss, A. C., Dutton, H. L., and Rose, M. F. (1988). α-Tomatine and resistance of tomato cultivars toward the nematode, Meloidogyne incognita. J. Chem. Ecol. 14, 1253–1259. doi: 10.1007/BF01019350
Erb, M., Flors, V., Karlen, D., de Lange, E., Planchamp, C., D’Alessandro, M., et al. (2009). Signal signature of aboveground-induced resistance upon belowground herbivory in maize. Plant J. 59, 292–302. doi: 10.1111/j.1365-313X.2009.03868.x
Erb, M., Meldau, S., and Howe, G. A. (2012). Role of phytohormones in insect-specific plant reactions. Trends Plant Sci. 17, 250–259. doi: 10.1016/j.tplants.2012.01.003
Erb, M., Robert, C. A. M., Marti, G., Lu, J., Doyen, G., Villard, N., et al. (2015). A physiological and behavioral mechanism for leaf-herbivore induced systemic root resistance. Plant Physiol. 169, 2884–2894. doi: 10.1104/pp.15.00759
Fenoll, C., Grundler, F. M. W., and Ohl, S. A. (1997). Cellular and molecular aspects of plant-nematode interactions. 1st Edn. Dordrecht: Kluwer Academic Publishers.
Fidantsef, A. L., Stout, M. J., Thaler, J. S., Duffey, S. S., and Bostock, R. M. (1999). Signal interactions in pathogen and insect attack: expression of lipoxygenase, proteinase inhibitor II, and pathogenesis-related protein P4 in the tomato, Lycopersicon esculentum. Physiol. Mol. Plant Pathol. 54, 97–114. doi: 10.1006/pmpp.1998.0192
Fragoso, V., Rothe, E., Baldwin, I. T., and Kim, S.-G. (2014). Root jasmonic acid synthesis and perception regulate folivore-induced shoot metabolites and increase Nicotiana attenuata resistance. New Phytol. 202, 1335–1345. doi: 10.1111/nph.12747
Gheysen, G., and Mitchum, M. G. (2011). How nematodes manipulate plant development pathways for infection. Curr. Opin. Plant Biol. 14, 415–421. doi: 10.1016/j.pbi.2011.03.012
Gheysen, G., and Mitchum, M. G. (2019). Phytoparasitic nematode control of plant hormone pathways. Plant Physiol. 179, 1212–1226. doi: 10.1104/pp.18.01067
Guo, H., and Ge, F. (2017). Root nematode infection enhances leaf defense against whitefly in tomato. Arthropod Plant Interact. 11, 23–33. doi: 10.1007/s11829-016-9462-8
Hamamouch, N., Li, C., Seo, P. J., Park, C. M., and Davis, E. L. (2011). Expression of Arabidopsis pathogenesis-related genes during nematode infection. Mol. Plant Pathol. 12, 355–364. doi: 10.1111/j.1364-3703.2010.00675.x
Hlywka, J. J., Stephenson, G. R., Sears, M. K., and Yada, R. Y. (1994). Effects of insect damage on glycoalkaloid content in potatoes (Solanum tuberosum). J. Agric. Food Chem. 42, 2545–2550. doi: 10.1021/jf00047a032
Hoagland, D., and Arnon, D. (1938). The water-culture method for growing plants without soil, vol. Circular 347. CA: UC College of Agriculture, Ag. Exp. Station, 1–39. Available at: https://archive.org/details/watercultureme3450hoag (Accessed September 06, 2019).
Hogenhout, S. A., and Bos, J. I. B. (2011). Effector proteins that modulate plant--insect interactions. Curr. Opin. Plant Biol. 14, 422–428. doi: 10.1016/j.pbi.2011.05.003
Hol, W. H. G., De Boer, W., Termorshuizen, A. J., Meyer, K. M., Schneider, J. H. M., van Der Putten, W. H., et al. (2013). Heterodera schachtii nematodes interfere with aphid-plant relations on Brassica oleracea. J. Chem. Ecol. 39, 1193–1203. doi: 10.1007/s10886-013-0338-4
Howe, G. A., and Jander, G. (2008). Plant immunity to insect herbivores. Annu. Rev. Plant Biol. 59, 41–66. doi: 10.1146/annurev.arplant.59.032607.092825
Hoysted, G. A., Bell, C. A., Lilley, C. J., and Urwin, P. E. (2018). Aphid colonization affects potato root exudate composition and the hatching of a soil borne pathogen. Front. Plant Sci. 9:1278. doi: 10.3389/fpls.2018.01278
Hoysted, G. A., Lilley, C. J., Field, K. J., Dickinson, M., Hartley, S. E., and Urwin, P. E. (2017). A plant-feeding nematode indirectly increases the fitness of an aphid. Front. Plant Sci. 8:1897. doi: 10.3389/fpls.2017.01897
Jang, J. Y., Le Dang, Q., Choi, Y. H., Choi, G. J., Jang, K. S., Cha, B., et al. (2015). Nematicidal activities of 4-quinolone alkaloids isolated from the aerial part of Triumfetta grandidens against Meloidogyne incognita. J. Agric. Food Chem. 63, 68–74. doi: 10.1021/jf504572h
Kafle, D., Hänel, A., Lortzing, T., Steppuhn, A., and Wurst, S. (2017). Sequential above- and belowground herbivory modifies plant responses depending on herbivore identity. BMC Ecol. 17:5. doi: 10.1186/s12898-017-0115-2
Kammerhofer, N., Egger, B., Dobrev, P., Vankova, R., Hofmann, J., Schausberger, P., et al. (2015). Systemic above- and belowground cross talk: hormone-based responses triggered by Heterodera schachtii and shoot herbivores in Arabidopsis thaliana. J. Exp. Bot. 66, 7005–7017. doi: 10.1093/jxb/erv398
Karssemeijer, P. N., Reichelt, M., Gershenzon, J., van Loon, J., and Dicke, M. (2020). Foliar herbivory by caterpillars and aphids differentially affects phytohormonal signalling in roots and plant defence to a root herbivore. Plant Cell Environ. 43, 775–786. doi: 10.1111/pce.13707
Kettles, G. J., and Kaloshian, I. (2016). The potato aphid salivary effector Me47 is a glutathione-S-Transferase involved in modifying plant responses to aphid infestation. Front. Plant Sci. 7:1142. doi: 10.3389/fpls.2016.01142
Kumar, P., Ortiz, E. V., Garrido, E., Poveda, K., and Jander, G. (2016). Potato tuber herbivory increases resistance to aboveground lepidopteran herbivores. Oecologia 182, 177–187. doi: 10.1007/s00442-016-3633-2
Kumari, C., Dutta, T. K., Banakar, P., and Rao, U. (2016). Comparing the defence-related gene expression changes upon root-knot nematode attack in susceptible versus resistant cultivars of rice. Sci. Rep. 6:22846. doi: 10.1038/srep22846
Kuśnierczyk, A., Winge, P. E. R., Jørstad, T. S., Troczyńska, J., Rossiter, J. T., and Bones, A. M. (2008). Towards global understanding of plant defence against aphids–timing and dynamics of early Arabidopsis defence responses to cabbage aphid (Brevicoryne brassicae) attack. Plant Cell Environ. 31, 1097–1115. doi: 10.1111/j.1365-3040.2008.01823.x
Kutyniok, M., and Muller, C. (2012). Crosstalk between above- and belowground herbivores is mediated by minute metabolic responses of the host Arabidopsis thaliana. J. Exp. Bot. 63, 6199–6210. doi: 10.1093/jxb/ers274
Kyndt, T., Denil, S., Bauters, L., Van Criekinge, W., and De Meyer, T. (2014). Systemic suppression of the shoot metabolism upon rice root nematode infection. PLoS One 9:e106858. doi: 10.1371/journal.pone.0106858
Kyndt, T., Nahar, K., Haeck, A., Verbeek, R., Demeestere, K., and Gheysen, G. (2017a). Interplay between carotenoids, abscisic acid and jasmonate guides the compatible rice-Meloidogyne graminicola interaction. Front. Plant Sci. 8:951. doi: 10.3389/fpls.2017.00951
Kyndt, T., Zemene, H. Y., Haeck, A., Singh, R., De Vleesschauwer, D., Denil, S., et al. (2017b). Below-ground attack by the root knot nematode Meloidogyne graminicola predisposes rice to blast disease. Mol. Plant-Microbe Interact. 30, 255–266. doi: 10.1094/MPMI-11-16-0225-R
Lemarié, S., Robert-Seilaniantz, A., Lariagon, C., Lemoine, J., Marnet, N., Jubault, M., et al. (2015). Both the jasmonic acid and the salicylic acid pathways contribute to resistance to the biotrophic clubroot agent Plasmodiophora brassicae in Arabidopsis. Plant Cell Physiol. 56, 2158–2168. doi: 10.1093/pcp/pcv127
Li, N., Han, X., Feng, D., Yuan, D., and Huang, L.-J. (2019). Signaling crosstalk between salicylic acid and ethylene/jasmonate in plant defense: do we understand what they are whispering? Int. J. Mol. Sci. 20:671. doi: 10.3390/ijms20030671
Livak, K. J., and Schmittgen, T. D. (2001). Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 25, 402–408. doi: 10.1006/meth.2001.1262
Machado, R. A. R., Ferrier, A. P., Robert, C. A. M., Glauser, G., Kallenbach, M., Baldwin, I., et al. (2013). Leaf-herbivore attack reduces carbon reserves and regrowth from the roots via jasmonate and auxin signaling. New Phytol. 200, 1234–1246. doi: 10.1111/nph.12438
Martínez-Medina, A., Fernandez, I., Lok, G. B., Pozo, M. J., Pieterse, C. M. J., and Van Wees, S. C. M. (2017). Shifting from priming of salicylic acid- to jasmonic acid-regulated defences by Trichoderma protects tomato against the root knot nematode Meloidogyne incognita. New Phytol. 213, 1363–1377. doi: 10.1111/nph.14251
Mbaluto, C. M., Ahmad, E. M., Fu, M., Martínez-Medina, A., and van Dam, N. M. (2020). The impact of Spodoptera exigua herbivory on Meloidogyne incognita induced root responses depends on the nematodes’ life cycle stages. AoB Plants 12:plaa029. doi: 10.1093/aobpla/plaa029
McCarville, M. T., O’Neal, M., Tylka, G. L., Kanobe, C., and MacIntosh, G. C. (2012). A nematode, fungus, and aphid interact via a shared host plant: implications for soybean management. Entomol. Exp. Appl. 143, 55–66. doi: 10.1111/j.1570-7458.2012.01227.x
Mewis, I., Khan, M. A. M., Glawischnig, E., Schreiner, M., and Ulrichs, C. (2012). Water stress and aphid feeding differentially influence metabolite composition in Arabidopsis thaliana (L.). PLoS One 7:e48661. doi: 10.1371/journal.pone.0048661
Mithöfer, A., and Boland, W. (2008). Recognition of herbivory-associated molecular patterns. Plant Physiol. 146, 825–831. doi: 10.1104/pp.107.113118
Mohase, L., and van der Westhuizen, A. J. (2002). Salicylic acid is involved in resistance responses in the Russian wheat aphid-wheat interaction. J. Plant Physiol. 159, 585–590. doi: 10.1078/0176-1617-0633
Moran, P. J., and Thompson, G. A. (2001). Molecular responses to aphid feeding in Arabidopsis in relation to plant defense pathways. Plant Physiol. 125, 1074–1085. doi: 10.1104/pp.125.2.1074
Moreno-Pedraza, A., Gabriel, J., Treutler, H., Winkler, R., and Vergara, F. (2019). Effects of water availability in the soil on tropane alkaloid production in cultivated Datura stramonium. Meta 9:131. doi: 10.3390/metabo9070131
Oñate-Sánchez, L., and Vicente-Carbajosa, J. (2008). DNA-free RNA isolation protocols for Arabidopsis thaliana, including seeds and siliques. BMC. Res. Notes 1:93. doi: 10.1186/1756-0500-1-93
Pieterse, C. M. J., Leon-Reyes, A., Van der Ent, S., and Van Wees, S. C. M. (2009). Networking by small-molecule hormones in plant immunity. Nat. Chem. Biol. 5, 308–316. doi: 10.1038/nchembio.164
Pieterse, C. M. J., Van der Does, D., Zamioudis, C., Leon-Reyes, A., and Van Wees, S. C. M. (2012). Hormonal modulation of plant immunity. Annu. Rev. Cell Dev. Biol. 28, 489–521. doi: 10.1146/annurev-cellbio-092910-154055
R Core development Team (2019). A language and environment for statistical computing. Available at: http://www.r-project.org/index.html(Accessed October 22, 2019).
Rocca, M., and Messelink, G. J. (2017). Combining lacewings and parasitoids for biological control of foxglove aphids in sweet pepper. J. Appl. Entomol. 141, 402–410. doi: 10.1111/jen.12355
Soler, R., Bezemer, T. M., Cortesero, A. M., Van der Putten, W. H., Vet, L. E. M., and Harvey, J. A. (2007). Impact of foliar herbivory on the development of a root-feeding insect and its parasitoid. Oecologia 152, 257–264. doi: 10.1007/s00442-006-0649-z
van Dam, N. M., Wondafrash, M., Mathur, V., and Tytgat, T. O. G. (2018). Differences in hormonal signaling triggered by two root-feeding nematode species result in contrasting effects on aphid population growth. Front. Ecol. Evol. 6:88. doi: 10.3389/fevo.2018.00088
Vos, I. A., Moritz, L., Pieterse, C. M. J., and Van Wees, S. C. M. (2015). Impact of hormonal crosstalk on plant resistance and fitness under multi-attacker conditions. Front. Plant Sci. 6:639. doi: 10.3389/fpls.2015.00639
Walling, L. L. (2000). The myriad plant responses to herbivores. J. Plant Growth Regul. 19, 195–216. doi: 10.1007/s003440000026
Walling, L. L. (2008). Avoiding effective defenses: strategies employed by phloem-feeding insects. Plant Physiol. 146, 859–866. doi: 10.1104/pp.107.113142
Wang, K., Luo, C., Liu, H., Xu, J., Sun, W., and Zhou, L. (2012). Nematicidal activity of the alkaloids from Macleaya cordata against certain nematodes. African J. Agric. Res. 7, 5925–5929. doi: 10.5897/AJAR11.1940
War, A. R., Paulraj, M. G., Ahmad, T., Buhroo, A. A., Hussain, B., Ignacimuthu, S., et al. (2012). Mechanisms of plant defense against insect herbivores. Plant Signal. Behav. 7, 1306–1320. doi: 10.4161/psb.21663
Wasternack, C. (2015). How jasmonates earned their laurels: past and present. J. Plant Growth Regul. 34, 761–794. doi: 10.1007/s00344-015-9526-5
Keywords: aboveground-belowground interactions, phytohormones, plant-mediated interactions, potato aphid (Macrosiphum euphorbiae), root-knot nematodes (Meloidogyne incognita), steroidal glycoalkaloids, systemic responses, local responses
Citation: Mbaluto CM, Ahmad EM, Mädicke A, Grosser K, van Dam NM and Martínez-Medina A (2021) Induced Local and Systemic Defense Responses in Tomato Underlying Interactions Between the Root-Knot Nematode Meloidogyne incognita and the Potato Aphid Macrosiphum euphorbiae. Front. Plant Sci. 12:632212. doi: 10.3389/fpls.2021.632212
Edited by:
Colette Broekgaarden, KeyGene, NetherlandsReviewed by:
Mark G. Sterken, Wageningen University and Research, NetherlandsKrzysztof Wieczorek, University of Natural Resources and Life Sciences Vienna, Austria
Copyright © 2021 Mbaluto, Ahmad, Mädicke, Grosser, van Dam and Martínez-Medina. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Crispus M. Mbaluto, Y3Jpc3B1cy5tYmFsdXRvQGlkaXYuZGU=; Y3Jpc21iYWx1dG9AZ21haWwuY29t; Ainhoa Martínez-Medina, YWluaG9hLm1hcnRpbmV6QGlybmFzYS5jc2ljLmVz