
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Plant Sci., 03 March 2021
Sec. Crop and Product Physiology
Volume 12 - 2021 | https://doi.org/10.3389/fpls.2021.627240
This article is part of the Research TopicBehind the Smoke and Mirrors: Reflections on Improving Cannabis Production and Investigating Medical PotentialView all 16 articles
 Dinesh Adhikary1
Dinesh Adhikary1 Manoj Kulkarni2Aliaa El-Mezawy3
Manoj Kulkarni2Aliaa El-Mezawy3 Saied Mobini2Mohamed Elhiti4Rale Gjuric4
Saied Mobini2Mohamed Elhiti4Rale Gjuric4 Anamika Ray2
Anamika Ray2 Patricia Polowick5
Patricia Polowick5 Jan J. Slaski3
Jan J. Slaski3 Maxwell P. Jones6
Maxwell P. Jones6 Pankaj Bhowmik5*
Pankaj Bhowmik5*In recent years high-THC (psychoactive) and low-THC (industrial hemp) type cannabis (Cannabis sativa L.) have gained immense attention in medical, food, and a plethora of other consumer product markets. Among the planting materials used for cultivation, tissue culture clones provide various advantages such as economies of scale, production of disease-free and true-to-type plants for reducing the risk of GMP-EuGMP level medical cannabis production, as well as the development and application of various technologies for genetic improvement. Various tissue culture methods have the potential application with cannabis for research, breeding, and novel trait development, as well as commercial mass propagation. Although tissue culture techniques for plant regeneration and micropropagation have been reported for different cannabis genotypes and explant sources, there are significant variations in the response of cultures and the morphogenic pathway. Methods for many high-yielding elite strains are still rudimentary, and protocols are not established. With a recent focus on sequencing and genomics in cannabis, genetic transformation systems are applied to medical cannabis and hemp for functional gene annotation via traditional and transient transformation methods to create novel phenotypes by gene expression modulation and to validate gene function. This review presents the current status of research focusing on different aspects of tissue culture, including micropropagation, transformation, and the regeneration of medicinal cannabis and industrial hemp transformants. Potential future tissue culture research strategies helping elite cannabis breeding and propagation are also presented.
Cannabis is a multipurpose crop with nutritional, medicinal, and industrial uses. Its leaves and flowers produce a spectrum of biologically active secondary metabolites, seeds are a source of nutritious oil and protein, and the stem contains two types of fiber serving as feedstock for the manufacturing of a variety of bio-based consumer goods (Small, 2004; Rodriguez-Leyva and Pierce, 2010; Wargent et al., 2013; Andre et al., 2016; Musio et al., 2018). The crop may have originated and been domesticated over 5000 years ago in Asia; since then, it has been interwoven with human history. In the South Asian regions, cannabis biotypes with elevated THC levels were commonly used for medicinal and recreational purposes, building a strong connection to social and religious rituals. While in the temperate climates, low-THC types were grown initially for fiber, and later also for food (Cheng, 1963; Li, 1974; Mechoulam, 1986; Cherney and Small, 2016; Clarke and Merlin, 2016; Jiang et al., 2016). Since the discovery of two cannabinoids [cannabidiol (CBD) in 1963) and tetrahydrocannabinol (THC) in 1964] in Dr. Raphael Mechoulam’s laboratory, more than 100 additional phytocannabinoids, flavonoids, and over 150 terpenes have been identified in the plant (Andre et al., 2016; Booth and Bohlmann, 2019; Rea et al., 2019). This high-value crop has built a strong foundation for a multi-billion-dollar global industry. Due to legal restrictions, research and development work has been slow and prevented researchers from investigating cannabis openly and making use of its full potential.
Recent cannabis legalization amendments in Canada, Europe, some parts of the United States, and other parts of the globe have helped promote research and use of this multipurpose crop. Commercial production increased in anticipation and response to the federal legalization of cannabis in Canada in October 2018 under the Cannabis Act (Government of Canada, 2018). Canada became the second nation after Uruguay (legalized December 2013) to legalize cannabis for recreational use at the federal level (Adinoff and Reiman, 2019). In the United States, 12 states have legalized cannabis for recreational use, with another 22 legalizing medical use (Adinoff and Reiman, 2019).
Inherently, cannabis is a dioecious species, with male and female flowers found on separate plants. Monoecious forms, which produce male and female flowers on the same plant, are very seldomly found in nature (Clarke and Merlin, 2016). Commercial monoecious cultivars of hemp have been bred for oilseed production and improved fiber yield and uniformity that cannot be achieved in dioecious forms exhibiting asynchronous maturation of the stems, as male plants commence an accelerated aging process soon after pollen shed. Due to the dioecious nature of most high THC-type cannabis and the lack of advanced breeding to produce true-to-type seed, they are propagated vegetatively and often grown indoors. Vegetative propagation maintains genetic purity and uniformity among the plants. Traditionally, indoor cannabis cultivators have depended on cuttings from a mother plant to produce genetically similar plants. While cannabis generally roots well (Caplan et al., 2018) and stem cuttings can produce large numbers of genetically similar plants, this method requires significant amounts of space. It has been observed that plants become less vigorous over time, the mother plants are susceptible to pests and diseases, and the resulting cuttings can harbor unwanted disease and serve as primary inoculum in production spaces.
As an alternative, in vitro techniques offer a promising approach for mass production and germplasm maintenance (Withers and Engelmann, 1997; Watt et al., 2000). Micropropagation can facilitate high throughput propagation in many species and forms the basis of disease-free plants for certified clean plant programs (Lineberger, 1983; Al-Taleb et al., 2011). Tissue culture based clean plant programs have been used in other vegetatively propagated crops such as potatoes, sweet potato, dates, sugarcane, banana, rice, tobacco, strawberry, grapes, orchids, roses, fruit trees, and some more horticulture of food and ornamental crops, helping to eradicate or prevent the spread of many plant pests, diseases, and viruses (National Clean Plant Network, 2020). Thus, developing an optimized in vitro method for propagating clean plants is a crucial strategy to produce large-scale genetically identical plants, retain genetic integrity, and maintain the long-term sustainability of the economically valuable crop (Conway, 2012). This review article aims to provide a comprehensive overview of the most updated available scientific research reported to date on tissue culture in cannabis, to contribute to our understanding of the cannabis tissue culture, and to assess potential applications of the optimized techniques in cannabis plant propagation, regeneration, and transformation.
According to Small et al. (1976), there are four groups of cannabis, ‘non-intoxicant (some C. sativa accessions),’ ‘semi-intoxicant’ (some C. sativa accessions), ‘intoxicant (C. indica),’ and ‘wild’ (C. ruderalis). Cannabis includes C. indica, C. ruderalis, and C. sativa. However, it has also been proposed that these three groups all belong to a single species (C. sativa) and the taxonomic classification among these proposed species remains a debated issue in Cannabis taxonomy (McPartland, 2018). For morphological and chemical characters (i.e., floral morphology and THC content), the earlier report considered them as different subspecies (Small and Cronquist, 1976), while another classified them as different species (Hillig, 2005).
Further complicating matters is the legal distinction between hemp and drug (narcotic) type cannabis. Any plant containing less than a defined concentration of the psychoactive THC is classified as hemp. In contrast, anything above the critical limit is classified as drug type cannabis. Depending upon the jurisdiction, the threshold THC concatenations in flowering plant parts differentiating between industrial hemp and drug type cannabis range from 0.2% of dry weight in most European counties, which is 0.3% in Canada, United States, and China and Brazil to 1% in Switzerland, Uruguay, Columbia, Mexico, and several Australian states. While this distinction is not based on taxonomy or genetic relationships, several studies have shown that most hemp cultivars are genetically distinct from drug-type cannabis (Rotherham and Harbison, 2011; Cascini et al., 2019). Mainly due to legal restrictions, artificial selection influenced by a decade’s long black market, and insufficient knowledge of the Cannabis taxonomy, these sub-types are poorly defined, especially the drug type cannabis.
Hemp is generally cultivated from seed and has named cultivars similar to most other crops. In contrast, drug type cannabis is generally propagated clonally; the clones are often referred to as ‘strains’ but are also often referred to as cultivars. As such, any given strain/cultivar can produce various clonal accessions with dramatically different phenotypes, making names unreliable. Further, many strains are offered by different seed companies, and the degree of genetic similarity or difference among providers has not been quantified; therefore, it is generally expected and accepted that there is significant variation within a single strain among seed companies and even within seed lots. Due to these factors, strain names in drug type cannabis are not reliable regarding a uniform phenotype.
Cannabis indica and Cannabis sativa are the major sources of cannabinoids, and are predominantly cultivated, while the third species, C. ruderalis is a wild and hardy species and is rarely grown by cultivators as there is no significant content of cannabinoids (Hilling and Mahlberg, 2004). In many lay literatures, distinction of ‘indica’ and ‘sativa’ have been mentioned and some of the earlier publications have also gathered some phenotypic differences (Table 1 and Figure 1); however, there is neither solid taxonomic agreement nor genetic or chemical evidence supporting the differences (Gloss, 2015; Sawler et al., 2015; Piomelli and Russo, 2016). The use of ‘indica’ and ‘sativa’ is vaguely based on the general notion that ‘sativa’ originated from European hemp, while ‘indica’ originated from the Indian subcontinent (Small, 2015), but their exact origin is still debatable.
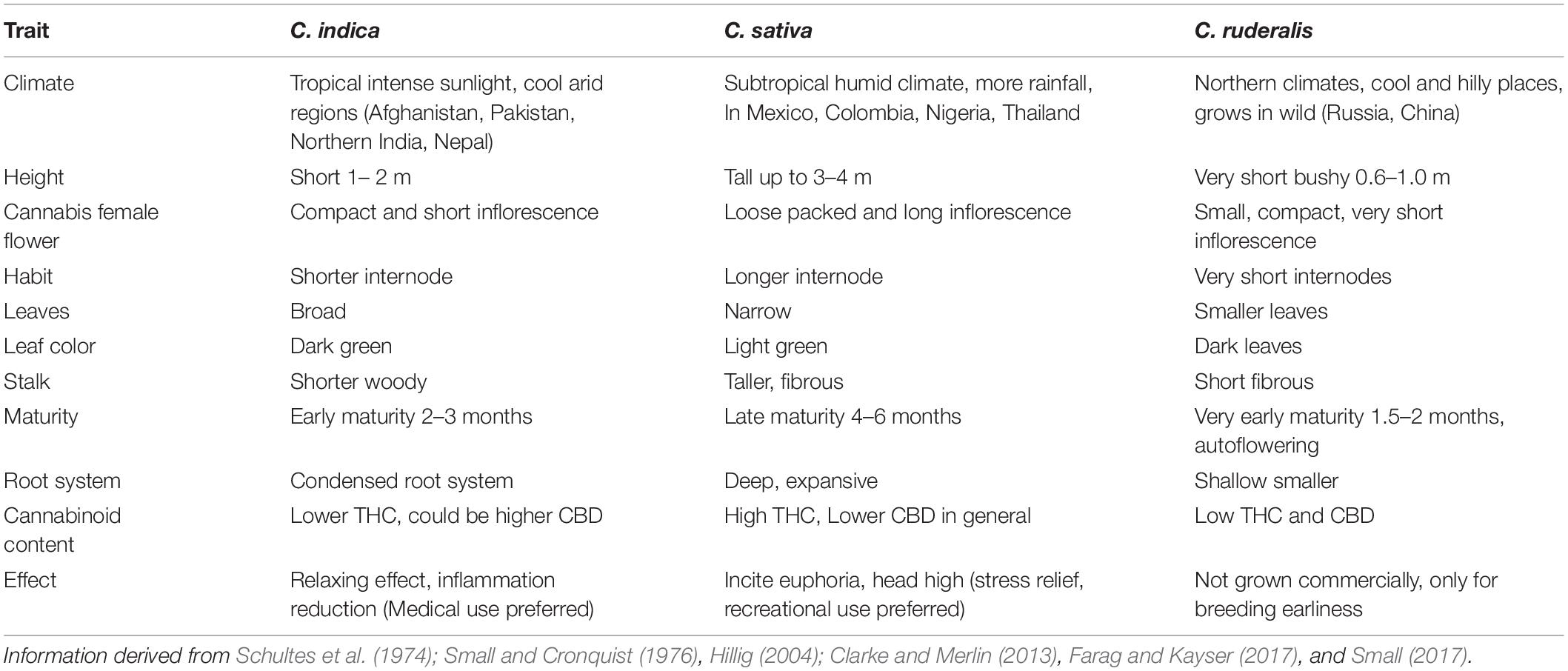
Table 1. Phenotypic differences among C. indica, C. sativa, and C. ruderalis ecotypes.

Figure 1. Cannabis leaf showing morphological differences of the three different species (C. indica, C. sativa, and C. ruderalis).
For decades, seed propagation in cannabis has supported agricultural needs and facilitated genetic improvement. However, with modern horticultural practices to the cannabis industry, stem cutting or traditional cloning, and in vitro propagation of this high-value crop has become a common practice (Lata et al., 2009a,b, 2011; Potter, 2009). Other methods of propagation are encapsulation of axillary nodes in calcium alginate beads (Lata et al., 2009a), leaf derived callus (Lata et al., 2010c), and temporary immersion bioreactor systems (Lata et al., 2010b) but these are limited in lab experiments only. Traditional cloning involves taking stem cuttings from a healthy mother plant and providing a rooting environment for the newly cut clone (Figure 2). For selection as a donor, a clear indication of alternating branches with no visible sign of insects, fungus, or any mineral deficiency in a mother plant is required. Cuttings can be taken from any part of a donor; despite some suggestions that growth in the lower half is better, no difference was observed between cuttings taken from the upper and lower part of the plant (Caplan et al., 2018). However, further research is warranted to test this across more genotypes and conditions. In general, cannabis propagates readily from stem cuttings even without rooting hormones.

Figure 2. Hemp nodal cloning. (A) Hemp plants at 6–8 leaf stage. (B) Elongated lateral branches after terminal buds removed from female plants (C) lateral branches planted in soil after excision from mother plants and. (D) Vegetative clones transferred to 7-inch pots after roots were established and grown. (E) Vegetative clone at maturity.
Stem cuttings have advantages over seed propagation, including quicker maturation, true-to-type plants, and elite genetics maintenance (Table 2). Along with the ease of propagation, the practice can limit unwanted gene flow (McKey et al., 2010), for example, between the hemp and drug-type, potentially retaining the proportions of active metabolites.
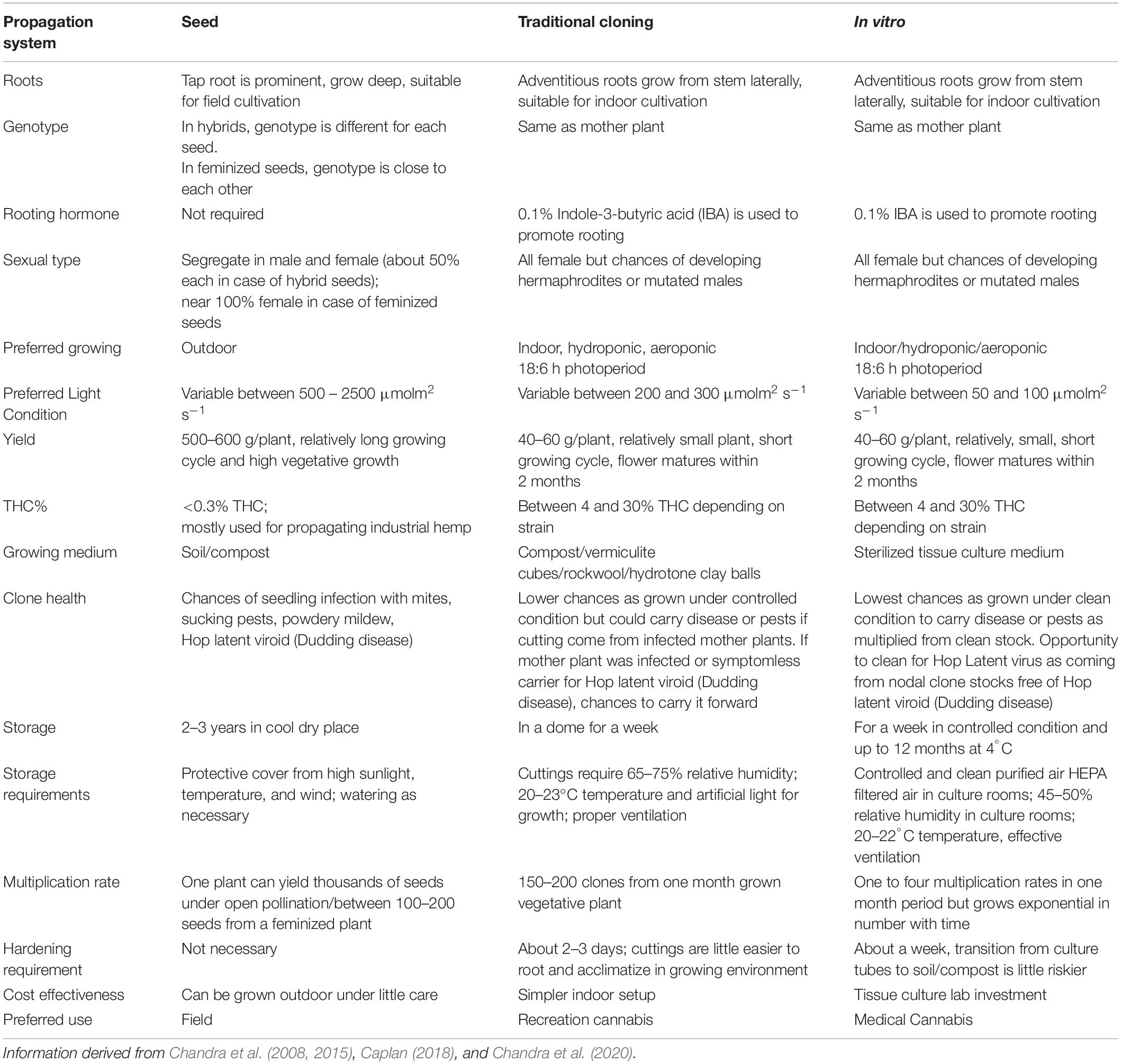
Table 2. Comparison between tissue culture cloning, manual cloning, and seed propagation in cannabis.
On the downside, space for large scale production is a concern as it can take considerable physical space, representing as much as 20–25% of production space just for cloning. Also, since it is currently manually performed, there is a low multiplication rate, and it is expensive in the long run. Therefore, this technique is more suitable for small growers requiring less than 1000 plants per growth cycle. For this reason, an adaptable, scalable, and robust high throughput tissue culture system with a high multiplication rate which preserves cannabis genetics, and produces more vigorous plants than manual clones, can prove to be more cost-effective in the long run (Table 2). Even small- scale growers with a small budget to use this technique to preserve genetics and test their desired strains’ regenerative capacity as a proof-of-concept. Building a team of experts to develop and execute tissue culture protocols successfully can be expensive and time-intensive initially; however, in the long term, it is a promising tool that has benefited many industries, including horticulture and cereal crops (Brown and Thorpe, 1995; Hussain et al., 2012).
Stem cuttings or traditional cloning method is the widely used propagation system adopted by many growers. In vitro propagation is establishing in cannabis industry slowly and is expected to take over the traditional cloning method. Although stem cuttings and in vitro clones can be comparable in terms of vegetative growth and physiological performance (Lata et al., 2009a), in vitro clones provide many advantages such as faster multiplication rate, clean clones without disease or virus, cost effective etc. (Table 1). Considering these advantages in vitro propagation is expected to become method of choice for propagation as well as genetic preservation in cannabis in the near future.
The legal hemp for CBD production and the medical cannabis industry is a fast- growing market, and cultivators are turning toward advanced scientific approaches such as in vitro micropropagation, to reduce the production costs and offer scalable, healthy, and high-quality cannabis variety. In addition to a critical need for cost-effective propagation to meet demand, there is also a desire to establish and properly characterize cultivars equivalent to those of traditional agriculture with specific, consistent THC and cannabinoid content to match particular drug and therapeutic requirements. Legalization has opened up the options for accessing more mainstream research applications. This increases the demand for the application of some additional cell technologies applications to this crop.
Although a few hemp cultivars have regenerated in vitro (Figure 3), Cannabis spp. have gained a wide reputation for being recalcitrant to tissue culture. At the beginning of the 1970s, along with the conventional propagation system, in vitro cultures of cannabis were initiated. The majority of the earlier in vitro studies were focused on cannabis callus culture to produce cannabinoids (Veliky and Genest, 1972; Itokawa et al., 1975, 1977; Hemphill et al., 1978; Hartsel et al., 1983; Loh et al., 1983; Fisse and Andres, 1985). Although there are multiple reports on shoot proliferation via micropropagation (Table 3), there are fewer scientific reports showing regeneration of a full plant through de novo regeneration (Mandolino and Ranalli, 1999; Slusarkiewicz-Jarzina et al., 2005; Wielgus et al., 2008; Chaohua et al., 2016).

Figure 3. Hemp tissue culture propagation. (A) Hypocotyl explants on callus-induction media. (B) Hypocotyl explants with the callus on callus induction media. (C,D) Callus and developing shoots on shoot-induction media. (E) Developed shoots on root-induction media.
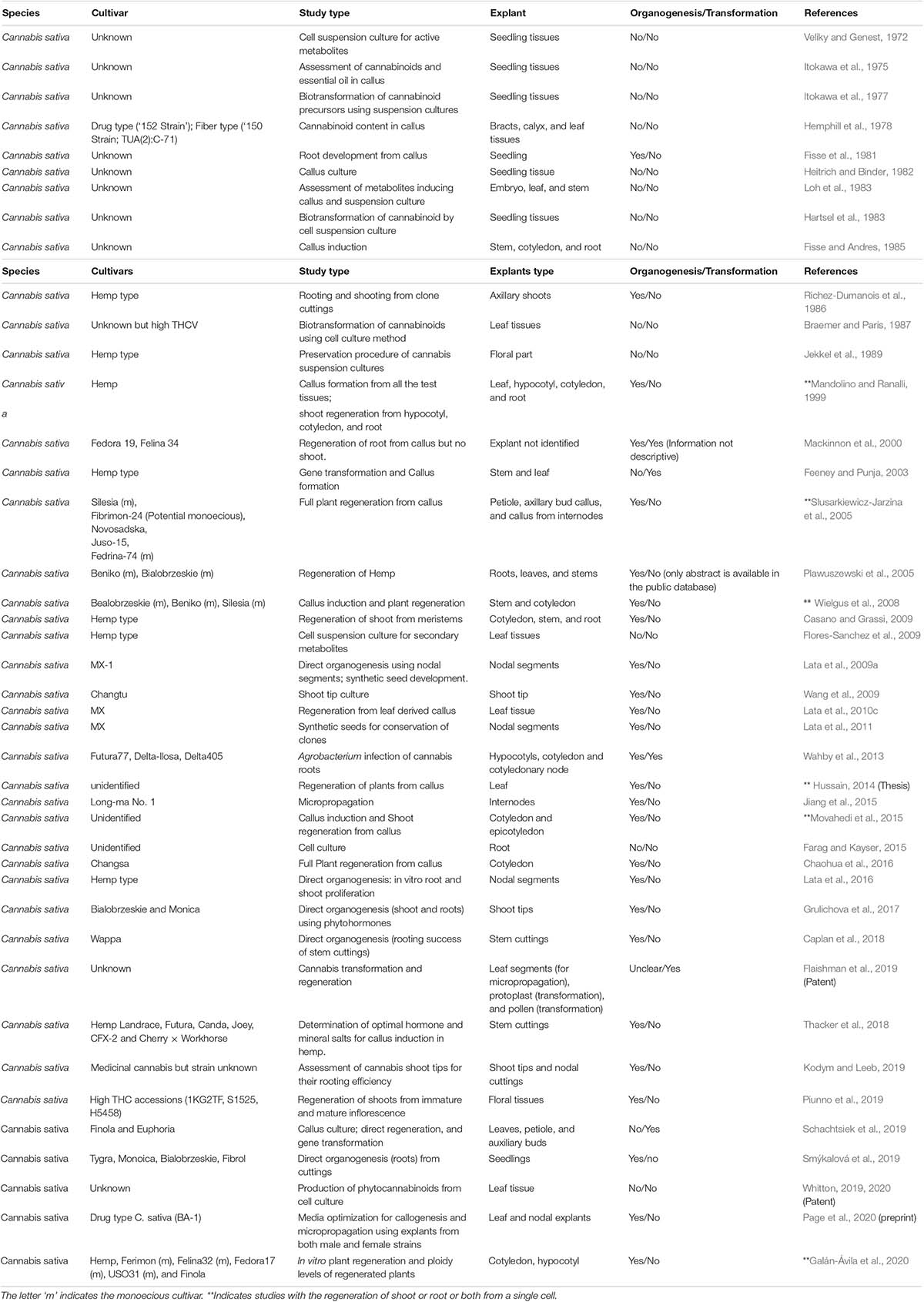
Table 3. Cannabis cell culture, transformation, and micropropagation work since 1972–2020.
The majority of regenerated strains and cultivars were monoecious, with few dioecious lines (Table 3). Recently, the optimization of a micropropagation and callogenesis protocol was reported for a few medical cannabis genotypes (Page et al., 2020). Although 48 years passed (Figure 4) since the first report of in vitro cell culture in cannabis, the available protocols are limited and inconsistent. In vitro regeneration of a cannabis plant from a single cell is still a challenge. Thus, the multi-billion-dollar cannabis industry needs an optimized tissue regeneration protocol for both industrial and medical cannabis.
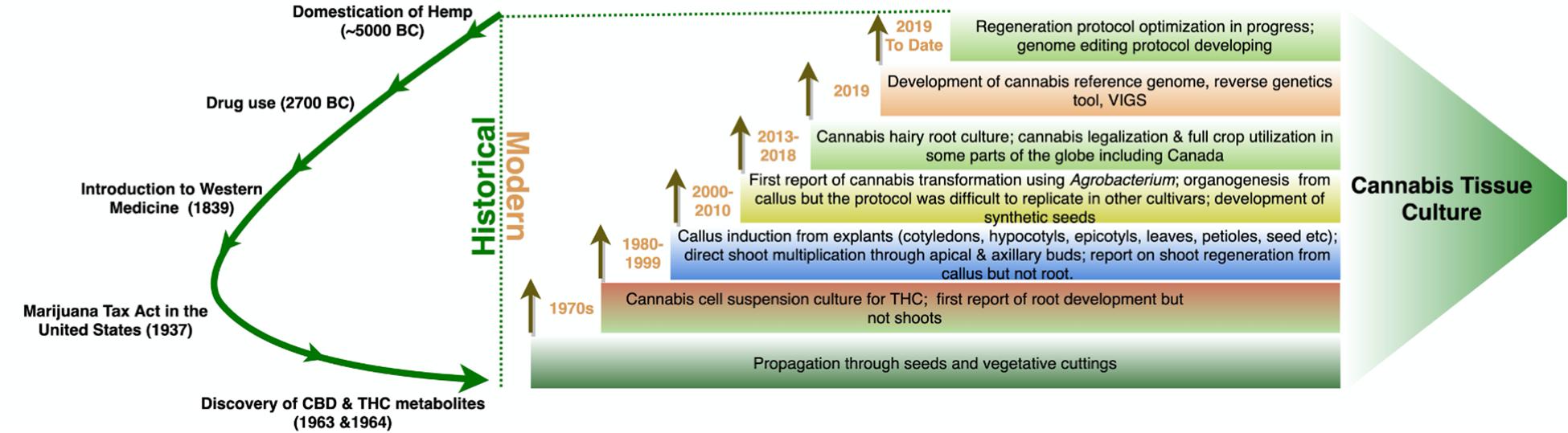
Figure 4. Evolution of cannabis tissue culture research. The green curved arrow on the left shows the key events in cannabis use. Each rectangle on the right shows the major research and development activities at different years. Each brown arrow indicates that the technology is continuously developing and research work is in progress in the particular research area.
It is generally understood that the most experienced cannabis companies have developed tissue culture and micropropagation techniques over the last two decades. However, most achievements in this in vitro field are held as a trade secret because of the competitive advantage provided within the industry. The most crucial challenges for the cannabis success micropropagation have been how to (i) reduce the length of subculture to minimize the occupied time and space, (ii) induce better root systems to increase the survival rate to >95%, (iii) optimize Plant Growth Regulators (PGRs), light (intensity and quality) and temperature required to maintain the genetically stable true-to-type clones. A generalized micropropagation workflow would require 7–8 weeks of culture transfer, 3 weeks of shoot multiplication, and 4 weeks of rooting. In terms of PGRs application, the best recommendation is optimized cytokinin and auxin for the vegetative medium and no cytokinin for the rooting medium using full MS media.
In recent years Canadian Licensed producers who are research-oriented have overcome some of these challenges. For example, the acclimatization period has been significantly reduced to less than 3 weeks. Another micropropagation challenge that the cannabis industry has recently solved is optimizing light intensity, light quality, and photoperiod in the culture room and maintaining the most effective temperature during shoot growth and root formation. Some unpublished data shows an increased propagation rate, from 3.5 to 5.8, during sub-culturing from each plantlet, through understanding and obtaining the right abiotic conditions within the culture room. As a starting point, some successful protocols are implemented with the minimum risk of somaclonal variation in cannabis (Movahedi et al., 2015; Lata et al., 2016, 2017; Grulichova et al., 2017; Page et al., 2020). These are game-changing procedures toward commercialization for cannabis micropropagation at a large-scale operation facility.
An ability to identify, characterize, and apply the genetic variability using biotechnology is the basis of molecular breeding. There are forward and reverse genetics approaches for genetic studies of an uncharacterized allele. With the improvement of sequencing technology, genetic transformation using reverse genetic tools has been an advantage in the molecular breeding program. While cannabis has gained a wide reputation of being recalcitrant to gene transformation and tissue culture, a few reports are describing the methods on gene transformation and regeneration (Feeney and Punja, 2003; Slusarkiewicz-Jarzina et al., 2005; Sirkowski, 2012; Wahby et al., 2013; Schachtsiek et al., 2019). Genome editing holds the potential to develop knockout mutants for significant cannabinoid biosynthesis genes such as THCA synthase, CBDA synthase, and CBGA synthase. Several varieties were tested; most were monoecious, although a few dioecious varieties were also used. In all cases, Agrobacterium-mediated gene transfer system was employed and exhibited successful transfer of genes, but the regeneration frequency was low to none. Feeney and Punja (2003) demonstrated the transformation success at the cellular level, but none of their treatments were successful in regeneration. Similarly, Wahby et al. (2013) applied A. rhizogenes strains (A4, AR10, C58, and IVIA251) and could induce hairy roots on the explants derived from hypocotyl and cotyledonary node; however, plantlet regeneration became a bottleneck for them as well. There is two patent information with the claim of successful genome modification and regeneration of cannabis with limited descriptions (Sirkowski, 2012). Thus, there is a need for an optimized protocol for the transformation and regeneration of cannabis replicable and reliable across different species.
There are various molecular tools developed for transient genetic transformation, including virus-induced gene silencing (VIGS). VIGSis an RNA mediated post-transcriptional gene silencing (PTGS) technique applied to study gene function in a relatively short period (Baulcombe, 1999; Liu et al., 2002; Senthil-Kumar and Mysore, 2014; Adhikary et al., 2019). Once a VIGS protocol is established in a species, it takes 3–6 weeks to see the loss-of-function phenotype of the tested gene/s in vivo (Adhikary et al., 2019). Thus, this is an ideal tool to apply, as a proof of concept, to define a target gene’s function prior to creating a stable transformation. VIGS, using the Cotton leaf crumple virus (CLCrV), was recently established in C. sativa, demonstrating the loss-of-function phenotype of phytoene desaturase (PDS) and magnesium chelatase subunit I (Chll) genes (Schachtsiek et al., 2019). Although the loss-of-function phenotype was weak, the researchers paved a clear path to explore unknown genes’ functions in the species. There are viral pathogens reported in cannabis (McPartland, 1996) and many viral vectors developed to date; tobacco rattle virus (TRV) is one of them with a broad-spectrum host range (over 400 plant species) across dicot species (Dinesh-Kumar et al., 2007). Given that TRV can also infect cannabis, potentially demonstrating a strong loss-of-phenotype than CLCrV viral vector.
Both transient and stable transformations have been incredibly beneficial for different research areas and applications in functional genomics. Stable gene transformation is preferred for many applications because once the gene modification is fixed in a plant system, it is heritable. The advantage of the altered gene function can be reaped for generations. As there are numerous reports of successful CRISPR-Cas9 mediated gene editing in many plant species, adopting this newly developed molecular tool in cannabis is vital to improving this economically important plant species. CRISPR can precisely alter a gene’s function in a genome (Jinek et al., 2012). It has great potential to benefit both basic and applied plant biology research and development. Therefore, establishing the technology in the cannabis crop is essential for functional studies of thousands of unknown genes and the development of novel varieties.
Traditional genetic modification (GM) and gene editing by CRISPR method are viewed differently (Shew et al., 2018). Gene editing performed using CRISPR method is not considered to be GM organism in some regions. Conventionally, GMO crops refer to organisms that have been altered in a way that they would not have evolved naturally. Moreover, GMO involves transferring foreign DNA fragment from one species to another (transgenic) or within the same species (cisgenic). But in the case of CRISPR edited plants, the targeted mutation is created by using an enzyme and a small guide RNA. While the mutation continues to inherited, the CRISPR machinery can be eliminated in the next generation (Aliaga-Franco et al., 2019). This method is precise and faster than conventional breeding practices, and it is much less controversial than GMO techniques. Therefore, the establishment of CRISPR-Cas9 system in cannabis is another crucial aspect that needs to be explored.
Agrobacterium rhizogenes is another functional genomics tool to assess the function of a gene or developing transgenic plants. These are differentiated cultures that are induced by the infection of Agrobacterium rhizogenes, a soil bacterium. Hairy root culture has a high growth rate in a hormone-free medium and exhibits the potential to yield secondary metabolites comparable to the wildtype (Pistelli et al., 2010). It enables the use of stable and reproducible bioreactor-based production and extraction independent of weather conditions, regulatory hurdles, and a lower risk of microbial contamination. This is a safe approach for producing medicinal and active metabolites free of hormones/viruses and does not require pesticides or insecticides. It is also one of the critical avenues for cannabis genetic transformation and functional genomics research.
Calli or hypocotyls infected by A. rhizogenes respond with the emergence of hairy roots from the infected site. Hairy roots can be individually selected and tested for a higher production rate of a compound of interest and cryopreserved at –196°C as a pure culture and subculture further for commercial-scale production (Engelmann, 2004). Cannabis hairy root culture has been successfully induced by A. rhizogenes (Wahby et al., 2006, 2013). Hairy root cultures from cannabis callus were also reported using 4 mg/l NAA as a supplement to B5 medium under dark conditions at 25°C (Farag and Kayser, 2015). In the study, the level of THCA and CBDA was less than 2 μg/g dry weight indicating a very low level of cannabinoids present in the hairy root culture under the dark condition with a 28-day growth cycle.
While detectable levels of cannabinoids are not present in C. sativa hairy roots, they have been reported to contain choline, atropine, and muscarine (Wahby et al., 2006, 2017). A higher level of these compounds was observed in the A. rhizogenes transformed hairy roots compared to non-transformed control. Choline was the most significant compound ranged between 203 and 510 mg/L (control 66–153 mg/L); Atropine with 562–933 μg/L (control 532–553 μg/L); Muscarine with 231–367 μg/L (control undetectable) (Wahby et al., 2017). Additionally, the THCA synthase gene’s heterologous expression in tobacco hairy root culture has been successful (Sirikantaramas et al., 2004; Taura et al., 2009).
The culture of indeterminate organs, especially the totipotent cells in the apical dome, is a method to obtain many virus clones in a short period (Mori, 1971; Wang and Charles, 1991). The apical dome region has no vascular connection to the developing procambium, leaf primordium, and axillary buds (Wang and Charles, 1991). This lack of vascular connection provides a basis for using the meristem for pathogen elimination as viruses readily travel through the vascular system but do not efficiently transfer from cell to cell. Uninfected cells can be isolated from the meristematic dome (Wang and Charles, 1991; Wu et al., 2020). It is a robust tool for producingvirus-free clones that can then be further multiplied at a commercial scale to produce certified virus-free plants. Characteristically, a section of tissue, mostly the apical dome, is dissected either from apical or lateral buds consisting of leaf primordia (no more than 1–2 in number) and apical meristem (0.1–0.5 mm in length) and cultured in a suitable growth medium. Upon induction of the meristem cells under a favorable combination of hormones and growth environment, the cells can continue to develop into a shoot or regenerate into plants through somatic embryogenesis or shoot organogenesis. The regeneration process occasionally gives direct shoot development from the explant, and sometimes morphogenesis occurs indirectly only after the formation of the callus.
There are well-established meristem culture protocols for different model and non-model species (Mori, 1971; Mordhorst et al., 2002; Al-Taleb et al., 2011; Spanò et al., 2018), including the closest relative of cannabis, Humulus lupulus (Hops), for eliminating virus infection (Grudzinska and Solarska, 2004; Grudzinska et al., 2006; Adams, 2015; Sallie and Jones, 2015). Given the importance of cannabis as a crop, the development of meristem culture for clean plant production could be useful. Unfortunately, this technique is most effective with viral diseases and would not eliminate fungal and bacterial pathogens known to infect cannabis.
For decades, plant protoplasts have been used for genetic transformation, cell fusion, somatic mutation, and more recently, for genome editing (Lei et al., 2015). Significant progress has been made in other crop species in genetic studies using protoplasts; however, for cannabis, studies are in a development phase, with the conditions suitable for the survival of transfected protoplasts and plant regeneration are yet to be optimized. Mesophyll protoplast isolation and transformation of at least three different cannabis cultivars has been reported (Morimoto et al., 2007; Flaishman et al., 2019). Based on the recent study, only about 4% of the protoplasts survived 48 h in liquid culture and plants were not regenerated (Flaishman et al., 2019). Even in the absence of successful regeneration of a whole plant, protoplasts are of great value in confirming the effectiveness of designed guide RNA (gRNA) prior to their use for the regeneration of gene-edited plants.
Somatic embryogenesis is the regeneration of a whole plant from cultured plant cells via embryo formation, from somatic plant cells of various tissues like root, stem, leaf, hypocotyl, cotyledon or petiole (Shen et al., 2018). They morphologically resemble the zygotic embryo’s bipolar structure, bear specific embryonic organs, and go through analogous development stages with similar gene expression profiles (Shen et al., 2018). Somatic embryogenesis can occur through direct regeneration. The embryos are developed directly from explant cells, or more commonly through indirect regeneration in which callus develops first, and the development of embryos occurs from callus cells (Sharp et al., 1980).
Plant regeneration via somatic embryogenesis starts with the initiation of embryogenic cultures by culturing various explants on media supplemented with only auxins or a combination of auxins and cytokinins to control cell growth and development (Osborne and McManus, 2005). One exception to this is the use of thidiazuron (TDZ), a cytokinin-like compound that is often used alone to induce somatic embryogenesis (Murthy et al., 1995). The proliferation of embryogenic cultures can occur on solid or in liquid media supplemented with auxins and cytokinins, followed by pre-maturation of somatic embryos on lower levels of PGRs or PGR free media to stimulate somatic embryo formation and development. Maturation of somatic embryos can occur by culturing on media with reduced osmotic potential or supplemented with abscisic acid (George et al., 2007). This maturation stage is critical for synthetic seed production as it allows embryos to be desiccated, stored, encapsulated, and treated like regular seeds. However, in many somatic embryogenesis systems, the maturation phase has not been developed, and somatic embryos germinate precociously to produce plants.
Somatic embryos are used as a model system in embryology studies; however, somatic embryogenesis’s main economic applications are for developing transgenic plants and large-scale virus-free vegetative propagation of elite plant genotypes. The possibility to scale up the propagation using bioreactors has been reported (Hvoslef-Eide and Preil, 2005). Somatic embryos are also ideal for genetic manipulation purposes as they develop from a single cell, thereby reducing the chances of producing chimeric plants, common when relying on shoot organogenesis or shoot proliferation (Dhekney et al., 2016). Other less common uses of somatic embryogenesis include cryopreservation of genetic materials and synthetic seed technology (George et al., 2007).
Feeney and Punja (2003) investigated the somatic embryogenesis and tissue culture propagation of hemp. Despite testing various explants and supplements, and variations in the culture medium and changes to the culture environment, there was no successful plantlet regeneration, and a reliable protocol for somatic embryogenesis in cannabis has yet to be published.
Thin cell layer (TCL) culture utilizes a thin layer of tissue as the explant to allow close contact between wounded cells and nutrients and growth regulators supplied in the medium; this controls the morphogenesis of the cultures (Nhut et al., 2003). This is most useful where larger explants may also contain a high level of endogenous hormones, carbon sources, and other substances that influence and conflict with the effects of exogenous substances placed in the medium and, thus, interfere with development. In general, sterilized TCL explants are excised either longitudinally (0.5–1 mm wide, 5–10 mm long) or transversally (0.1–5 mm thick) prior to culturing (Nhut et al., 2003; Croom et al., 2016). Like other in vitro techniques, TCL requires an optimized protocol regarding basal media, PGRs and other added nutrients and growth conditions such as daylength, light intensity, and temperature. These conditions vary for not only the species but can be genotype-dependent. It has been widely used in different species, including bamboo, banana, citrus, tomato, rose, Lilium ledebourii, Bacopa monnieri, saffron, among others (Nhut et al., 2003; Teixeira da Silva et al., 2007; Mirmasoumi et al., 2013; Croom et al., 2016; Azadi et al., 2017). TCL’s potential is yet to be explored in Cannabis spp.; however, it may prove to have some utility in the regeneration of genetic transformants in this high value but re calcitrant regeneration crop.
Androgenesis is a biological process by which a whole plant regenerates directly from immature pollen (microspores) through the embryogenesis developmental pathway under in vitro conditions. While the resulting plant is haploid and inherently sterile, a diploid plant can arise either spontaneously or artificially (Gilles et al., 2017), usually with colchicine, which blocks cytokinesis without blocking chromosome doubling (Galazkajoa and Niemirowicz-Szczytt, 2013). This doubled haploid is homozygous at all loci. Doubled Haploid (DH) plants have been extensively used in plant breeding programs to increase the speed and efficiency with which homozygous lines can be obtained (Alisher et al., 2007). DH technology is traditionally used to genetically stabilize parental lines for F1 hybrid production. This is important for the rapid integration of new traits through backcross conversion and to develop molecular mapping populations. It is also used to fix desired traits obtained through transformation or mutagenesis and simplify genomic sequencing by eliminating heterozygosity (Ferrie and Mollers, 2011). As such, this technology would be an important tool for both forward and reverse functional genomics studies.
There are two different approaches to develop haploid plants. First, in situ methods, using particular pollination techniques such as irradiated pollen, inter-species crosses or so-called ‘inducer lines’ (Ren et al., 2017); second, in vitro methods including the culture of haploid cells (gametes) and their development to haploid embryos and consequently haploid plants through germination. The microspores, which can be harvested in large numbers (millions), are generally isolated for culture as a uniform population. Alternatively, the culture of whole anthers is used to obtain haploid plants through the androgenesis process. The main disadvantage of another culture is the potential for developing a mix of both haploid and diploid plantlets (Elhiti et al., 2010). In this review, we will focus only on the production of doubled haploids from microspores using in vitro culture.
One of the most important factors affecting DH production is the microspore developmental stage. It is a complicated factor that has a strong influence on microspore culture’s success. It has been reported that only microspores that are at a stage sufficiently immature have the ability to change their developmental fate from a gametophytic to embryogenic, leading to sporophytic development (Soriano et al., 2013). The most amenable stage is either the uni-nucleate stage of the microspore or the early binucleate stage, either at or just after the first pollen mitosis. At this developmental stage, the microspore’s transcriptional status may still be proliferative and not yet fully differentiated (Malik et al., 2007). Although all microspores within an anther would be roughly of a similar age, not all cells have embryonic competence. Therefore, the incremental differences in the stages of development of individual microspores can be considered significant. To avoid this problem, Bhowmik et al. (2011) introduced a new treatment, discontinuous Percoll gradient centrifugation, to provide a uniform population of B. napus isolated microspores at the appropriate stage of development. This approach has consistently produced high embryo yields and consistent embryo development.
In 2019, an extensive hemp breeding program was introduced at Haplotech Inc.1. As there has been no previously reported success in the area, a hemp DH project was initiated to accelerate this program. Four different Haplotech genotypes were used for this experiment. Both male racemes and pollen-induced female colas were collected, and the buds were fractionated according to size into three groups (2–3, 3–4, and 4–5 mm). Each group was surface sterilized with 15% commercial bleach and washed three times with distilled-sterilized water for 5 min each. The sterilized buds were macerated in isolation media (MS basal fortified by 13% sucrose). The isolated microspores were washed by extraction medium two times or until the supernatant became clear. The isolated microspores were subjected to fractional centrifugation using Percoll, as described by Bhowmik et al. (2011). The concentration of microspores was diluted to 4 × 104cells/ml with MS basal fortified by 10% sucrose. Five ml of this diluent (4 × 104) microspores were mixed with 5 ml of induction media (MS basal, 10% sucrose supplemented with different additives for induction) in 47 mm Petri dishes. The final concentration of the culture used was 2 × 104 cells/ml. The isolated microspores in culture were observed every 3 days using an inverted microscope and a binocular microscope.
Samples of isolated microspores were stained with 4, 6-diamidino-2-phenylindole (DAPI) and observed using a fluorescence microscope to monitor their in vitro development, once every 3 days. Monitoring of the culture samples by DAPI staining in the first 2 weeks revealed that the microspores of all four genotypes remained uninuclear (Figure 5A). This developmental stage was found to be the most responsive to embryogenesis induction in many crop plants (Soriano et al., 2013). Of the factors tested, the most crucial for further development of the microspore was the induction medium formulation. Using a relatively complex medium, a few microspores responded (0.05–0.5%) and developed further, while the remainder died within 5–10 days. Microspore derived embryos initiated by a series of random divisions within the surrounding exine wall. The nucleus of uninucleate microspores (Figure 5A) condensed and reduced in size during the first 2 days in culture (Figure 5B). They then divided symmetrically within the first 5–8 days, forming two equal-sized nuclei (Figure 5C). This developmental stage is considered the initial stage that is often referred to as sporophytic growth (Soriano et al., 2013). Within another 3–5 days, the nuclei underwent a series of divisions resulting in the formation of multinucleate structures (Figure 5D). By approximately the third week of culture, globular stage embryos were observed in culture (Figure 5E). Early in the fourth week, these globular structures developed into heart stage embryos (Figure 5F). To date, growth has not progressed past this stage of embryo development. Current experiments including adjustment of the osmoticum and removal of secondary metabolites which could inhibit (microspore-derived) embryo development are running.
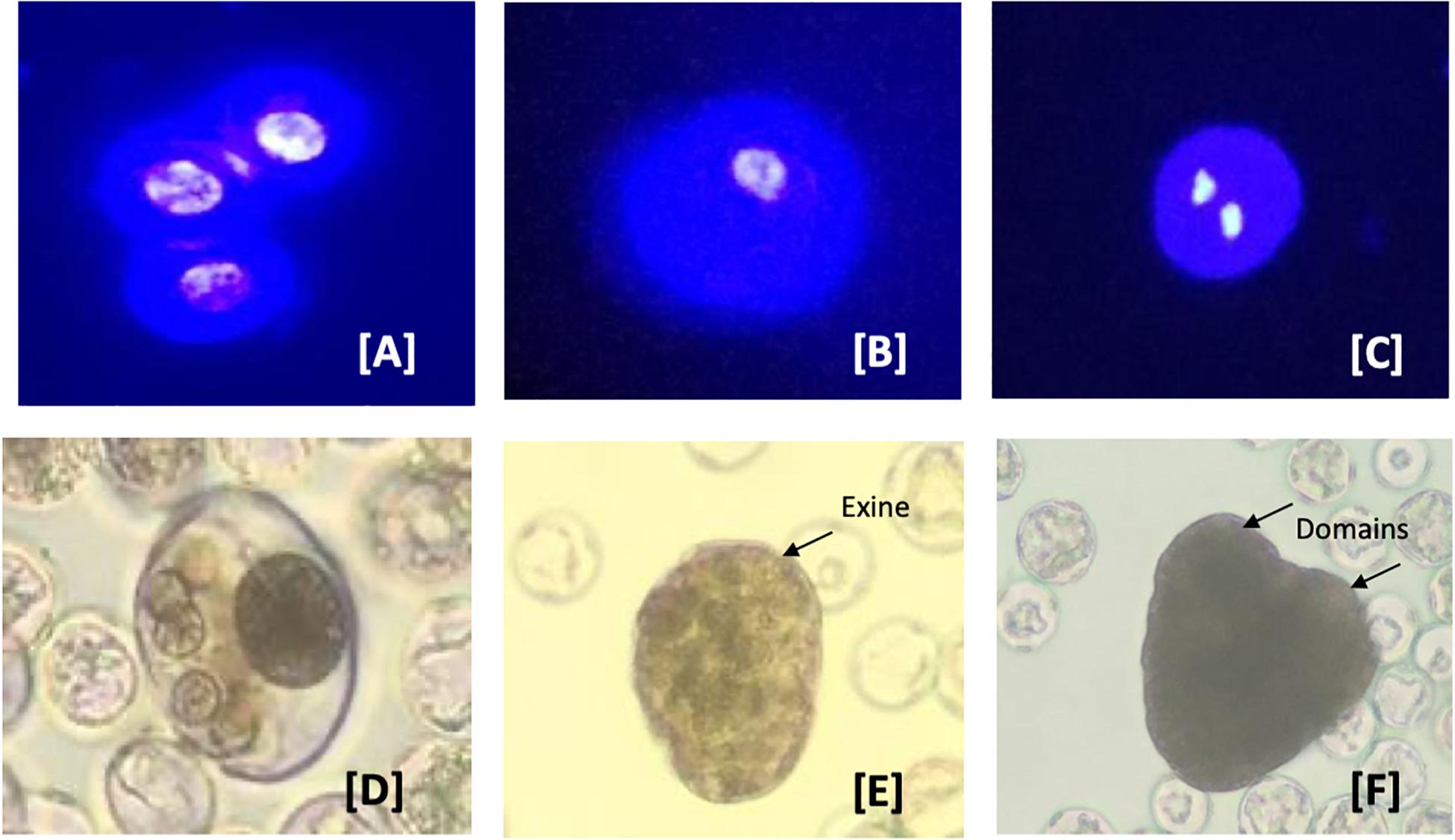
Figure 5. Developmental pathways observed in C. sativa (industrial hemp) microspore culture. (A–C) Male gametophyte development in C. sativa during in vitro culture. (A) Uninucleate microspores; (B) uninucleate microspores after 3 days in culture media; (C) symmetrically divided microspore with two equally sized nuclei; (D) multinucleate structure without organization and still enclosed in exine; (E) globular multicellular structure with developing exine; and (F) heart-shape embryo with two distinct domains. The nuclei in (A–C) are stained with the nuclear dye 4′,6-diamidino-2-phenylindole (DAPI) to indicate viability.
A mutation occurs in DNA, naturally or it can also be induced artificially. The majority of the genetic variation existing in a gene pool has occurred naturally. These genetic variations can be recombined through conventional breeding practices to develop a novel variety with desired gene traits. Although these spontaneous mutations are frequent, the desired mutation in the desired gene segment altering its biological role is extremely rare. Therefore, mutation induction tools are used in the rapid development of genetic variability in crops. For the last few decades, there were several scientific reports published assessing the impact of an induced mutation in the improvement of crops (Brock, 1971; Broertjes and Van Harten, 1988; Micke, 1999; Oladosu et al., 2016). However, in cannabis research and development is rapidly flourishing, but there are only a few reports on targeted mutation through genetic transformation (Feeney and Punja, 2003; Slusarkiewicz-Jarzina et al., 2005; Sirkowski, 2012; Wahby et al., 2013) and there is no mutant variety introduced at the commercial level. In vitro culture techniques, coupled with mutagenesis, has simplified the crop improvement work for both seeds and vegetatively propagated plants (Hussain et al., 2012). Little efforts have been made and published to establish DH production in cannabis, but once streamlined will open up exciting opportunities for DH mutagenesis as it has been successfully employed in canola (Szarejko, 2003).
Synthetic seeds usually refer to artificially encapsulated somatic embryos (Murashige, 1977) but have also been used in reference to encapsulated vegetative tissues that have the potential to develop into a whole plant (auxiliary buds, cell aggregates, shoot buds). Somatic embryos provide the ideal approach to developing synthetic seeds as they often have the ability to survive desiccation and can be treated in much the same way as true seeds. At the same time, other tissues lack this capacity and are less useful (Rihan et al., 2017). As shown in Figure 6, synthetic seeds can be successfully developed by using various explants, media, and encapsulation protocols (Bapat et al., 1987; Corrie and Tandon, 1993; Nyende et al., 2003; Chand and Singh, 2004; Rai et al., 2008; Lata et al., 2009a).
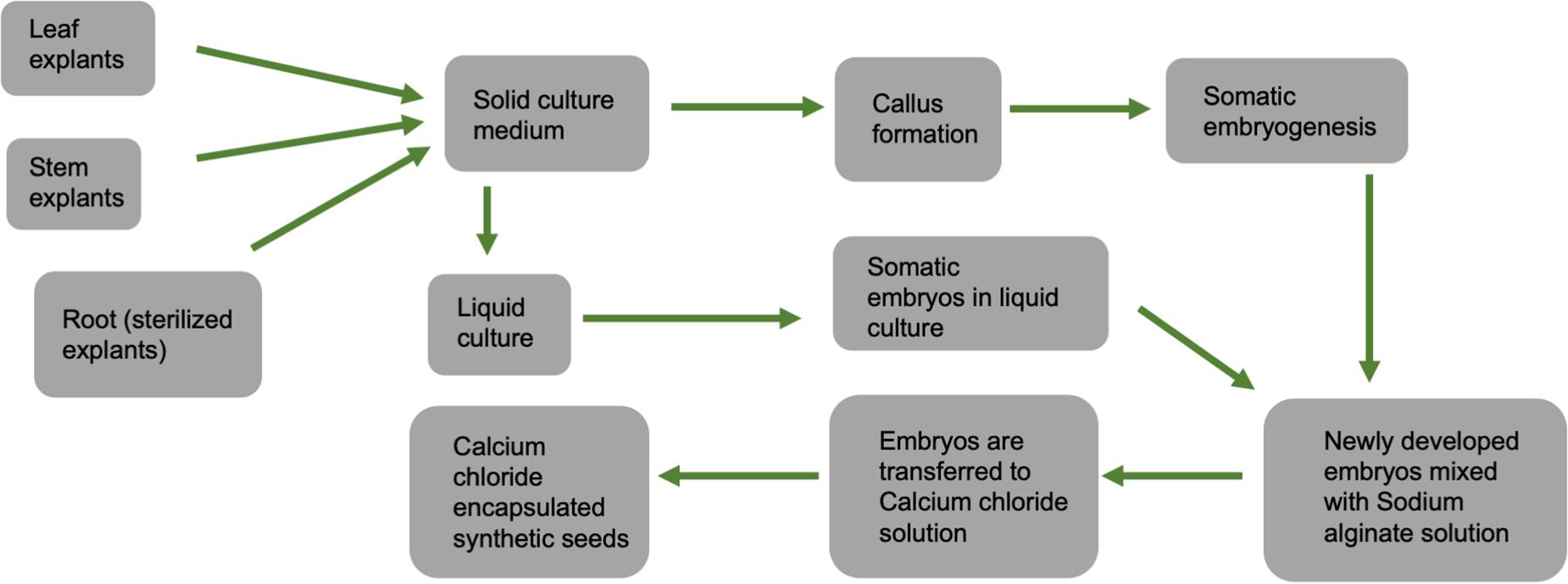
Figure 6. General schematic diagram showing steps for calcium chloride encapsulated synthetic seed production.
Cannabis is generally a cross-pollinating crop, and due to its allogamous nature, it is difficult to maintain existing elite varieties by seed. Typically, a minimum isolation distance of 5 km between breeding nurseries and hemp production fields is required to minimize the occurrence of nuisance pollen. Such separation is often difficult to achieve in areas with high hemp production intensity. Therefore, in vitro propagation using synthetic seed technology is an alternative method for large-scale clonal propagation and germplasm preservation. As the cannabis industry grows, this method may be cheaper and faster than traditional tissue culture methods. Along with the preservation of genetic uniformity, clones produced through this technique are pathogen-free, easy to handle, and transport.
Moreover, in other species, this approach has resulted in increased quality of planting material (Rihan et al., 2017). While cannabis tissue culture methods are still being optimized, Lata et al. (2009a) developed a high-frequency propagation of axillary buds of C. sativa encapsulated in calcium alginate gel. Calcium alginate is a hydrogel that contains nutrients, growth regulators, and sometimes antibiotics.
When directly sown on a substrate, encapsulation aids in the physical protection and establishment and growth of the explant. According to Lata et al. (2009a), gel capsule consisted of 5% sodium alginate with 50 mM CaCl2.2H2O, and full-strength MS medium supplied with 0.5 μM TDZ, and 0.075% plant preservative mixture (PPM). The optimal regrowth and conversion were achieved in MS medium supplemented with antimicrobial components, PPM (0.075%) and TDZ (0.5 μM) under in vitro conditions. Under in vivo condition, the optimal conversion and regrowth were exhibited on 1:1 potting mix-fertilome with coco natural growth medium supplied with MS medium containing 3% sucrose, 0.5% PPM. Clones regenerated from the explants were successfully hardened and transferred to the soil (Lata et al., 2009a).
Another hurdle to in vitro propagation is transporting requested strains from the tissue culture facility to the growers in a timely manner. These transportation issues become incredibly challenging for maintaining crop schedules because cannabis crops can take more than 2 months to reach hardening stages, then spend 4 weeks in vegetative growth, then 7 or 8 weeks in flower. Greenhouse or indoor growers require a consistent supply demand to receive a high volume of plantlets every week to start over a new grow room at a very tight on-time delivery schedule, which is the most important metric in their operations. An established and cost-effective synthetic seed encapsulation technique would provide an opportunity to eliminate the transportation challenge.
Cryopreservation refers to the storage of diverse living materials at below –130°C (Engelmann, 2004). It serves as an alternative conservation approach to the conventional field and in vitro (i.e., slow growth) germplasm conservation and is cost-effective over extended periods with minimal space and routine maintenance requirements (Pence, 2011; Engelmann, 2014; Popova et al., 2015). It also assists current breeding programs by providing long-term storage and an easy long-distance exchange of genetic materials (e.g., pollen and meristematic apices and buds). Cryopreservation has been implemented for various plant species using different methods, the most popular and widely applicable, including controlled freezing, vitrification, encapsulation-dehydration, encapsulation-vitrification, and droplet-vitrification (Sakai and Engelmann, 2007; Popova et al., 2015). These methods follow distinct approaches to dehydrate cryopreserving living materials by converting liquid water to a glassy state to avoid the lethal formation of intracellular ice. The selection of methods and the scales of conservation using this approach are strongly determined by genotypes and tissue materials used, which contain different responses to pre- and post-cryopreservation treatments.
Conventional and in vitro conservation of cannabis require considerable amounts of space and routine maintenance, have genetic mutations accumulate in the plants. Conventional conservation may expose plants to virulence pathogens. The plants may eventually become susceptible to diseases. The application of cryopreservation can serve as an essential tool for the conservation of various valuable C. sativa genotypes with unique attributes and trading the genotypes nationally and internationally in sterile conditions. The first study on applying cryopreservation techniques in C. sativa was reported in 1989 using cell suspension cultures (Jekkel et al., 1989). The suspension cultures were preserved using 10% dimethyl sulfoxide (DMSO) cryoprotectant and a controlled cooling rate of 2°C/min and transfer temperature of –10°C, with a 58% survival rate after cryopreservation of the cultures. A cryopreservation protocol for C. sativa shoot tips was recently developed using a droplet-vitrification in liquid nitrogen for long-term conservation of this crop (Uchendu et al., 2019). The report showed that vitrified shoot tips using a cryoprotectant solution of 30% glycerol, 15% ethylene glycol, 15% DMSO in liquid MS medium with 0.4 M sucrose, pH 5.8 had 63% re-growth efficiency. Despite the promising progress made, more studies need to be done on selecting appropriate cryopreservation methods with respect to the tissue types and genotypes, increasing re-growth and survival efficiency of preserved samples, and genetic stability of regenerated plants after using different cryopreservation tools, among others.
The in vitro condition also raises some issues for concern, primarily when the material is maintained over a long period of time.
In vitro mass-propagation and maintenance of elite germplasm requires genetically stable true-to-type clones. Several factors, such as the number of subcultures, changes in the relationship of auxin/cytokinin, explant type, and a high concentration of growth regulators, may influence the genetic stability of a clone under in vitro conditions (Joyce et al., 2003; Sato et al., 2011; Smulders and de Klerk, 2011; Nwauzoma and Jaja, 2013). While carefully selecting explant types and optimizing the conditions above, but depending on the plant species, clonal stability can be obtained during in vitro mass-propagation and germplasm conservation of the desired elite genotypes maintained. To date, C. sativa plants regenerated from nodal culture, and in vitro conserved synthetic seeds (‘Encapsulated’ nodal segments) have shown no evidence of genetic mutations; however, this has only been evaluated using low numbers of markers (Lata et al., 2010a, 2011). Despite optimizing and using properly in vitro conditions that limit somaclonal variations, assessment of clonal stability is required to ensure the regenerated clones are the true-to-type of the donor plants.
Although clonal propagation and maintenance of elite germplasm require a substantial genetic uniformity among in vitro regenerated plantlets, there may be a large possibility of genetic variations, called “somaclonal variation” among these plants and/or relative to the donor plants. Somaclonal variation is commonly a result of genetic alterations and changes in the new in vitro plants’ epigenetics compared to the original source plants (Miguel and Marum, 2011; Abreu et al., 2014). The frequency and nature of somaclonal variation in vitro culture can be influenced by different factors, such as explant source, genotype, in vitro techniques, in vitro growth conditions, length of the culture period, and the number of subcultures. The use of de novo regeneration from highly differentiated tissues (i.e., roots, leaves, stems, hypocotyls, cotyledons, etc.) is generally considered to produce more somaclonal variation compared to explants with developed meristems (i.e., axillary buds and shoot tips) (Pijut et al., 2012). Most of these factors generate oxidative stress during culture initiation and subsequent subculturing. The explants and the subsequent regenerated plants exposed to the stress may retain genetic changes. For example, protoplast and callus based plant regeneration impose a high degree of oxidative stress; thus, the stress promotes a high mutation rate, whereas plants regenerated through auxiliary branching (e.g., nodes, shoot tip) experience very low oxidative stress, normally resulting in no genetic variation (Zayova et al., 2010; Smulders and de Klerk, 2011; Krishna et al., 2016). Genetic variation can also arise from somatic mutations already present in the explants collected from the donor plant (Karp, 1994). In vitro regeneration of plants can also be genotype-specific, in which genotypes have different degrees of mutation risks and thus strongly determine the formation of somaclonal variation (Alizadeh et al., 2010; Eftekhari et al., 2012; Nwauzoma and Jaja, 2013). The genetic alterations strongly depend on the in vitro techniques used to regenerate in vitro plants. Additionally, despite differences across plant species, cultures maintained for a long period tend to generate high somaclonal variation, and vice versa (Farahani et al., 2011; Jevremovic et al., 2012; Sun et al., 2013). When cultures are getting old and continuously subcultured, the chance of generating genetically less uniform plants is increased (Zayova et al., 2010), but depends upon the plant species. For example, any more than eight subculture cycles increased somaclonal variation in banana (Khan et al., 2011), whereas over 30 subcultures did not cause any detectable somaclonal variations in C. sativa (Lata et al., 2010a).
Although the molecular mechanism of how somaclonal variations generated from a single plant genotype under the same in vitro conditions is not fully explored, several potential mechanisms causing genetic alternations and epigenetics have been proposed in different plant species. These mechanisms include changes in chromosome number, point mutations, somatic crossing over and sister chromatid exchange, chromosome breakage and rearrangement, somatic gene rearrangement, DNA replication, changes in organelle DNA, insertion or excision of transposable elements, segregation of pre-existing chimeral tissues, DNA methylation, epigenetic variation, and histone modifications and RNA interference (Sato et al., 2011; Krishna et al., 2016; references therein).
The occurrence of somaclonal variations in regenerated in vitro plants may be advantageous or disadvantageous, depending on in vitro propagation goals. If in vitro propagation aims to generate new variants, obtaining variations among in vitro plants can be advantageous that increases genetic diversity for a genotype used. It provides an alternative tool to the breeders for obtaining genetic variability in different plant species, which are either difficult to breed or have narrow genetic bases. On the flip side, when in vitro propagation targets to produce multiple true-to-type in vitro plants and maintain elite germplasm, the occurrence of subtle somaclonal variations is a severe problem.
Nature has deftly adorned cannabis species with a spectrum of phytocannabinoids or monoterpenoids that are chemically designed with para-oriented isoprenyl and aralkyl groups (Hanus et al., 2016). Since the discovery of tetrahydrocannabinol (THC) and cannabidiol (CBD) in the early 1960s, there are over 120 cannabinoids that has been reported, and the biosynthesis pathway of these compounds has been greatly improved (Taura et al., 1995; Sirikantaramas et al., 2004; Taura et al., 2007b, 2009; Gagne et al., 2012; Stout et al., 2012; Laverty et al., 2019). Presumably, cannabigerolic acid (CBGA), the product formed by the alkylation of geranyl diphosphate and olivetol, is the key precursor compound in the synthesis of cannabinoids (Fellermeier and Zenk, 1998). The cyclization event of prenyl components of CBGA, catalyzed by three enzymes – tetrahydrocannabinolic acid synthase (THCAS) (genebank accession: AB057805), cannabidiolic acid synthase (CBDAS) (genebank accession: AB292682), and cannabichromenic acid synthase (CBCAS), lead to the formation of three major cannabinoids, THCA, CBDA, and CBCA, respectively (Sirikantaramas et al., 2004; Taura et al., 2007a). Biochemical characterization of the enzymes, THCAS and CBDAS, have demonstrated that the enzymes follow a similar reaction mechanism. In the presence of molecular oxygen, the enzymes use flavin adenine dinucleotide (FAD) cofactor to catalyze CBGA forming THCA and CBDA, and hydrogen peroxide as its chemical biproduct (Sirikantaramas et al., 2004; Taura et al., 2007b). Although it is a bit unclear, the chemical reaction for CBCAS also believed to use FAD as cofactor and molecular oxygen to complete the enzymatic activity on CBGA. The genes that encode for CBCAS and THCAS are highly similar in the nucleotide level, indicating that CBCAS is also flavoproteins, like the other two enzymes, requiring oxygen to catalyze CBGA to CBCA (Laverty et al., 2019). THCA, CBDA, and CBCA are the major cannabinoids in acidic forms that are synthesized in cannabis plant; upon decarboxylation, these compounds convert into neutral forms, THC, CBD, and CBC respectively (Wang et al., 2016).
Variations between regenerated and donor plants can be exhibited at phenotypic, cytological, biochemical, and genetic/epigenetic levels (Hillig, 2005; Miguel and Marum, 2011; Smulders and de Klerk, 2011; Abreu et al., 2014). These variations can be determined through different approaches, such as morphological, cytological, biochemical, and molecular analyses (Figure 7). For morphological traits, changes are not always observed at early developmental stages or may not entirely display the variations. By contrast, the use of cytological and molecular detection approaches determines differences at chromosomal and DNA levels, respectively, regardless of the developmental stages in various plant species (Clarindo et al., 2012; Pathak and Dhawan, 2012; Currais et al., 2013; Abreu et al., 2014; Bello-Bello et al., 2014). To date, several studies have been reported on the use of different molecular markers in Cannabis spp. genetic diversity, fingerprinting, etc. These markers include random amplified polymorphic DNA (RAPD), restriction fragment length polymorphisms (RFLP), amplified fragment length polymorphism (AFLP), microsatellites, inter simple sequence repeat (ISSR), short tandem repeat (STR) multiplex, and single nucleotide polymorphisms (SNPs) and PCR Allele Competitive Extension (PACE) assay (Faeti et al., 1996; Kojoma et al., 2002; Alghanim and Almirall, 2003; Gilmore and Peakall, 2003; Hakki et al., 2003; Datwyler and Weiblen, 2006; Mendoza et al., 2009; Lata et al., 2010a; Gao et al., 2014; Dufresnes et al., 2017; Henry et al., 2018). These molecular markers coupled with cytological and morphological analyses (Abreu et al., 2014) are valuable techniques to ensure the genetic stability of in vitro regenerated plants or in vitro conserved germplasm of C. sativa. To date, only ISSR markers have been used to confirm the genetic stability of C. sativa synthetic seeds during in vitro multiplication and storage for 6 months under different growth conditions, and in vitro propagated plants over 30 nodal subcultures in culture and hardening in soil for 8 months, compared to the corresponding donor plants (Lata et al., 2010a, 2011).
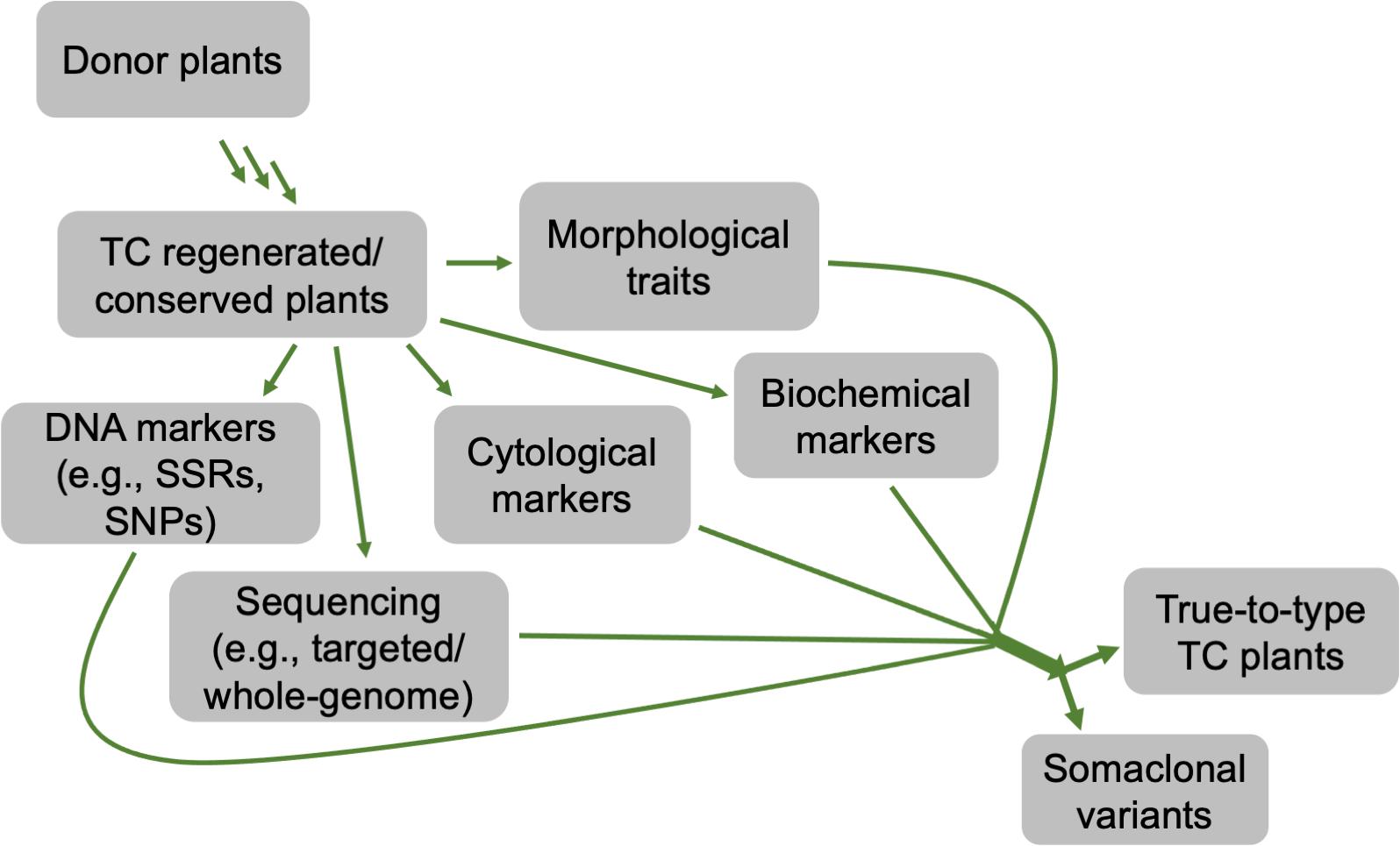
Figure 7. A flow chart depicting different approaches that can be used to determine the genetic stability of in vitro regenerated or conserved cannabis plants, compared to its donor counterparts.
The present global cannabis market is worth $340 B2. To supply cannabis (medical and recreational) to global consumers, a stable supply chain of quality production and value-added product development still needs to be established. Considering the average annual weighted usage base of 110 g per customer (Canaccord Genuity), the global cannabis demand currently could be around 19-20 M kg per year. Major cannabis consumers are in Europe, North America, South America, Asia, and Oceanic parts of the world, with an estimate of 263 million people using the drug in the previous year (European Consumer Stables Report, 2018; World Drug Report, 2019). To produce 20 M kg of cannabis every year, considering a 40-gm yield per plant, would require 500 M clones/seeds a year. An average price of $10, as, then, the overall present global expected market size for tissue culture clones/manual clones could be predicted around $5B. With intensive indoor cultivation, tissue culture clonal planting material can also reduce the risk of fungal and viral diseases, substantially reducing production cost to under $0.5 per gram to maintain a profitable cannabis production (Table 4). Considering these global demand scenarios, the supply of clean cannabis clones (pest free, and true to type tested) is an important supply chain component essential for the success and future growth of cannabis industry. To sustain and support the industry growth and make the production cost-effective, optimization in the cannabis tissue culture technology is vital.
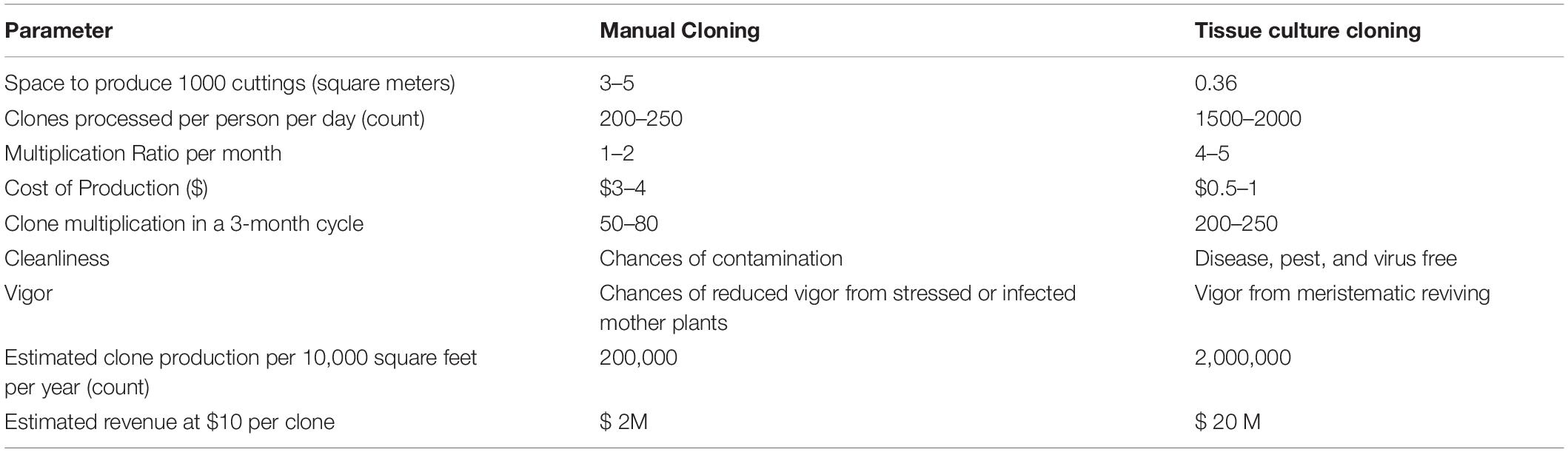
Table 4. Comparison between tissue culture cloning and manual cloning in cannabis.
The in vitro propagation of cannabis is superior to conventional methods because of disease-free elite plants’ production and a high multiplication rate. The cannabis industry is keen to invest in in vitro propagation due to (i) saving footprint/production area by shifting a mother room to a tissue culture lab that will be almost 10% the size of the space needed same number of clones.
The main hurdle of in vitro propagation is the capital cost for the tissue culture lab setup. Setting up a massive large-scale production facility can involve a multimillion-dollar investment. Industry and technology will need to continue to improve and reduce costs so that in vitro propagation can be affordable for all growers.
In other plants, under a laminar flow hood setting, on an average of 100 plants per hour with 2000 working hours, 200,000 plants can be produced in a year. With an hourly labor cost of $35 per hour will cost around $0.35 per tissue culture plant (Sluis, 2005). This is around 60% of the production cost, adding another $0.15 for other costs (including electricity, resources, and marketing) makes it a baseline cost of $0.50 per plant. Scale also makes some impact on the cost of production being larger facilities can reduce the cost per plant significantly. These production costs can be as low as $0.15 per plant if the plants are produced in India, Singapore, China, or Africa where labor costs are comparatively low.
A few biotech companies recently added robotic sub-culturing technology for their cannabis plantlets and developed a fully automated micropropagation system to reduce large-scale operation costs. However, the capital investment to purchase this kind of robotic system is incredibly high at this time. Automated technologies for media preparation and dispensing, photoautotrophic bioreactor systems, robotic explant handling, and cutting, transfer laser dissected explants into fresh culture media, and automated acclimatized and hardened plant packaging in future will make cannabis tissue culture industry high throughput and extremely cost-effective for assured “Just In Time” supply of pest free, true-to-type cannabis clones. A conceptual model for high throughput automated cannabis in vitro clonal mass propagation is depicted in Figure 8. Robotics has the potential to bring tissue culture cost down by 25% (as low as $0.15 per plant to compete with low-cost production in some parts of the world). Tissue culture automation technology is slowly progressing, and it will not only bring high-level consistent output but also reduce the cost of production as low as 20 cents per plant.
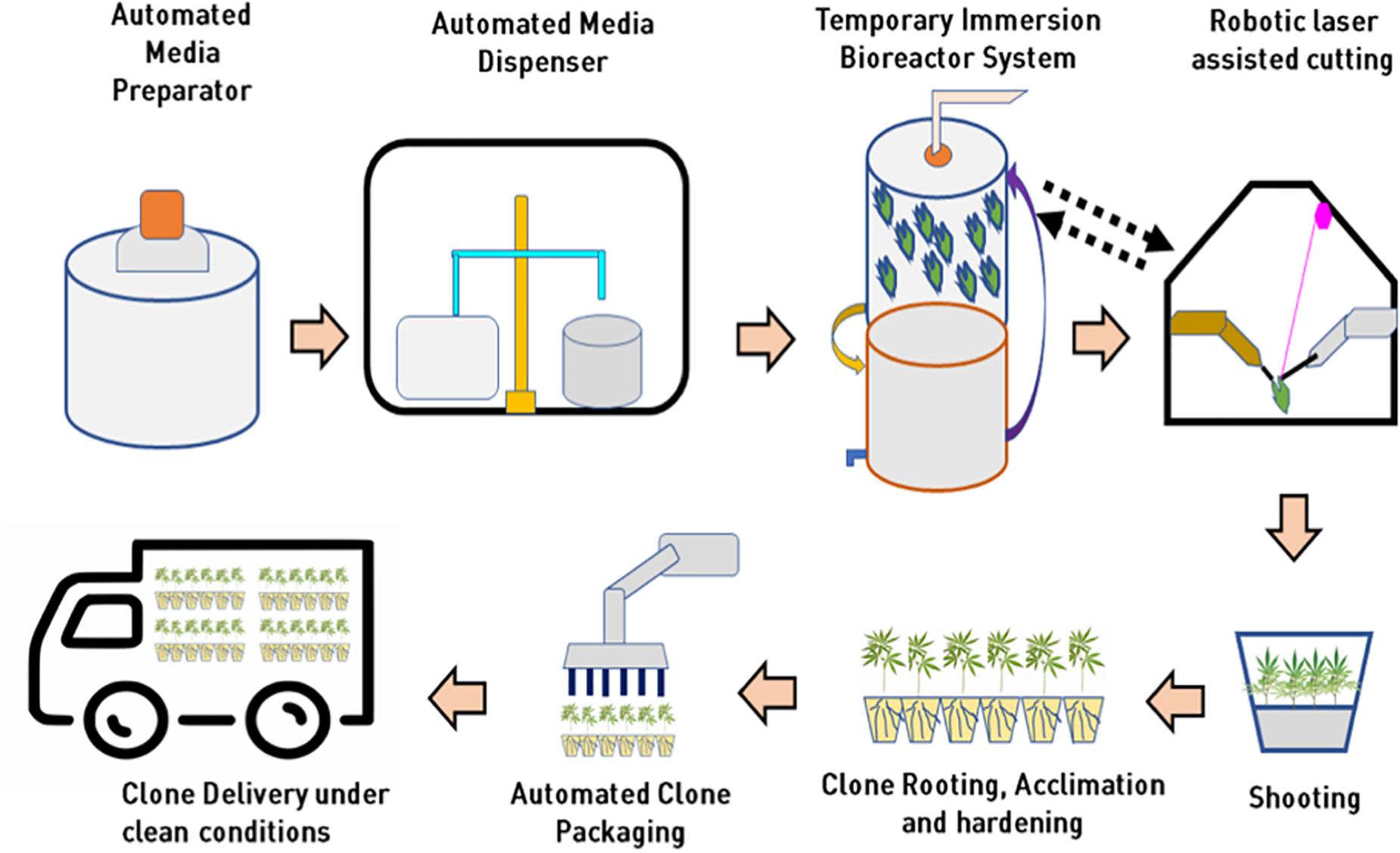
Figure 8. Integration of automation and bioreactor technologies for mass propagation in cannabis for low cost clonal multiplication at in vitro level.
The process of developing new varieties through conventional breeding can take 7–12 years, depending on crop species. The progress of cannabis breeding programs is limited due to the difficulty in maintaining selected high yielding cross-pollinated elite genotypes under field or greenhouse conditions. Therefore, tissue culture techniques are advantageous for cannabis improvement because they can facilitate high multiplication rate and production of disease-free elite plants by overcoming the problems of heterozygosity from cross-pollination. The development of new industrial hemp and medical cannabis cultivars with improved traits could be further advanced using genome editing and other precision breeding tools, combined with in vitro techniques for regeneration. Unfortunately, hemp and cannabis plants’ dioecious nature complicates the efforts toward the improvement of specific traits, such as resistance to pests and diseases. Therefore, with the recent legalization, calls for serious targeted efforts are required to advance the regeneration and transformation protocols aiming to enhance the quality and safety of the plants and end products.
All authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
ME and RG were employed by the company Haplotech Inc.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We would like to thank Dr. Ayelign M. Adal from the University of British Columbia for his contribution and suggestion on composing this article.
Abreu, I. S., Carvalho, C. R., and Clarindo, W. R. (2014). Massal induction of Carica papaya L. ‘Golden’ somatic embryos and somaclone screening by flow cytometry and cytogenetic analysis. Cytologia 79, 475–484. doi: 10.1508/cytologia.79.475
Adams, A. N. (2015). Elimination of viruses from the Hop (Humulus lupulus) by heat therapy and meristem culture. J. Hortic. Sci. 50, 152–160. doi: 10.1080/00221589.1975.11514616
Adhikary, D., Khatri-Chhetri, U., Tymm, F. J. M., Murch, S. J., and Deyholos, M. K. (2019). Virus induced gene-silencing (VIGS) system for functional genomics in betalainic species, Amaranthus tricolor. Appl. Plant Sci. 7:1221. doi: 10.1002/aps3.1221
Adinoff, B., and Reiman, A. (2019). Implementing social justice in thetransition from illicit to legal cannabis. Am. J. Drug Alcohol. Abuse 6, 673–688. doi: 10.1080/00952990.2019.1674862
Alghanim, H. J., and Almirall, J. R. (2003). Development of microsatellite markers in Cannabis sativa for DNA typing and genetic relatedness analyses. Anal. Bioanal. Chem. 376, 1225–1233. doi: 10.1007/s00216-003-1984-0
Aliaga-Franco, N., Zhang, C., Presa, S., Srivastava, A. K., Granell, A., Blazquez, M. A., et al. (2019). Identification of transgene-free CRISPR-edited plants of rice, tomato, and arabidopsis by monitoring DsRED fluorescence in dry seeds. Front. Plant Sci. 10:1150. doi: 10.3389/fpls.2019.01150
Alizadeh, M., Singh, S. K., and Patel, V. B. (2010). Comparative performance of in vitro multiplication in four grape (Vitis spp.) rootstock genotypes. Int. J. Plant Prod. 4, 41–50. doi: 10.22069/IJPP.2012.680
Al-Taleb, M. M., Hassawi, D. S., and Abu-Romman, S. M. (2011). Production of virus free potato plants using meristem culture from cultivars grown under Jordanian environment. J. Agric. Environ. Sci. 11, 467–472.
Andre, C. M., Hausman, J. F., and Guerriero, G. (2016). Cannabis sativa: The plant of the thousand and one molecules. Front. Plant Sci. 7:1–17. doi: 10.3389/fpls.2016.00019
Azadi, P., Bagheri, K., Gholami, M., Mirmasoumi, M., Moradi, A., and Sharafi, A. (2017). Thin cell layer, a suitable explant for in vitro regeneration of saffron (Crocus sativus L.). J. Agric. Sci. Technol. 19, 1429–1435.
Bapat, V. A., Mhatre, M., and Rao, P. S. (1987). Propagation of Morus indica L. (Mulberry) by encapsulated shoot buds. Plant Cell Rep. 6, 393–395. doi: 10.1007/BF00269570
Baulcombe, D. C. (1999). Fast forward genetics based on virus-induced gene silencing. Curr. Opin. Plant Biol. 2, 109–113. doi: 10.1016/S1369-5266(99)80022-3
Bello-Bello, J. J., Iglesias-Andreu, L. G., Aviles-Vinas, S. A., Gomez-Uc, E., Canto-Flick, A., and Santana-Buzzy, N. (2014). Somaclonal variation in habanero pepper (Capsicum chinense Jacq.) as assessed ISSR molecular markers. HortScience 49, 481–485. doi: 10.21273/HORTSCI.49.4.481
Bhowmik, P., Dirpaul, J., Polowick, P., and Ferrie, A. M. R. (2011). A high throughput Brassica napus microspore culture system: influence of percoll gradient separation and bud selection on embryogenesis. Plant Cell Tiss Organ. Cult. 106, 359–362. doi: 10.1007/s11240-010-9913-3
Booth, J. K., and Bohlmann, J. (2019). Terpenes in Cannabis sativa – From plant genome to humans. Plant Sci. J. 284, 67–72. doi: 10.1016/j.plantsci.2019.03.022
Braemer, R., and Paris, M. (1987). Biotransformation of cannabinoids by a cell suspension culture of Cannabis sativa L. Plant Cell Rep. 6, 150–152. doi: 10.1007/BF00276675
Brock, R. D. (1971). The role of induced mutations in plant improvement. Environ. Exp. Bot. 11, 181–196. doi: 10.1016/S0033-7560(71)90273-0
Broertjes, C., and Van Harten, A. M. (1988). Applied mutation breeding for vegetatively propagated crops. Developments in crop science. The Netherlands: Elsevier, 197–204. doi: 10.1007/BF00024972
Brown, D. C., and Thorpe, T. A. (1995). Crop improvement through tissue culture. World J. Microbiol. Biotechnol. 11, 409–415. doi: 10.1007/BF00364616
Caplan, D. M. (2018). Propagation and root zone management for controlled environment Cannabis production. Ph.D. thesis, Guelph: University of Guelph.
Caplan, D., Stemeroff, J., Dixon, M., and Zheng, Y. (2018). Vegetative propagation of cannabis by stem cuttings: effects of leaf number, cutting position, rooting hormone, and leaf tip removal. Can. J. Plant Sci. 98, 1126–1132. doi: 10.1139/cjps-2018-0038
Casano, S., and Grassi, G. (2009). Evaluation of media for hemp (Cannabis sativa L.) in vitro propagation. Italus Hortus 16, 109–112.
Cascini, F., Farcomeni, A., Migliorini, D., Baldassarri, L., Boschi, I., Martello, S., et al. (2019). Highly predictive genetic markers distinguish drug type from fiber type Cannabis sativa L. Plants 8:496. doi: 10.3390/plants8110496
Chand, S., and Singh, A. K. (2004). Plant regeneration from encapsulated nodal segments of Dalbergia sissoo Roxb, a timber-yielding leguminous tree species. J. Plant Physiol. 161, 237–243. doi: 10.1078/0176-1617-01036
Chandra, S., Lata, H., and ElSohly, M. A. (2020). Propagation of Cannabis for Clinical Research: An Approach Towards a Modern Herbal Medicinal Products Development. Front. Plant Sci. 11:958. doi: 10.3389/fpls.2020.00958
Chandra, S., Lata, H., Khan, I. A., and ElSohly, M. A. (2008). Photosynthetic response of Cannabis sativa L.@to variations in photosynthetic photon flux densities, temperature and CO2 conditions. Physiol. Mol. Biol. Plants 14, 299–306. doi: 10.1007/s12298-008-0027-x
Chandra, S., Lata, H., Khan, I. A., Mehmedic, Z., and ElSohly, M. A. (2015). Light Dependence of Photosynthesis and Water Vapour Exchange Characteristics in Different High Δ9-THC Yielding Varieties of Cannabis sativa L. J. Appl. Res. Med. Aromat. Plants 2, 39–47. doi: 10.1016/j.jarmap.2015.03.002
Chaohua, C., Gonggu, Z., Lining, Z., Chunsheng, G., Qing, T., Jianhua, C., et al. (2016). A rapid shoot regeneration protocol from the cotyledons of hemp (Cannabis sativa L.). Ind. Crop Prod. 83, 61–65. doi: 10.1016/j.indcrop.2015.12.035
Cheng, T. K. (1963). Archeology in China, Vol. 3 Chou China. Cambridge: W. Heffer and Sons Ltd, 36–37.
Cherney, J. H., and Small, E. (2016). Industrial Hemp in North America: Production. Polit. Potent. Agronomy 6:58. doi: 10.3390/agronomy6040058
Clarindo, W. R., Carvalho, C. R., and Mendonça, M. A. C. (2012). Ploidy instability in long-term in vitro cultures of Coffea arabica L. monitored by flow cytometry. Plant Growth Regul. 68, 533–538. doi: 10.1007/s10725-012-9740-0
Clarke, R. C., and Merlin, M. D. (2013). Cannabis: evolution and ethnobotany. California: Univ of California Press.
Clarke, R. C., and Merlin, M. D. (2016). Cannabis domestication, breeding history, present-day genetic diversity, and future prospects. Critic. Rev. Plant Sci. 6, 293–327. doi: 10.1080/07352689.2016.1267498
Conway, G. (2012). One billion hungry: can we feed the world?. New York, NY: Cornell University Press.
Corrie, S., and Tandon, P. (1993). Propagation of Cymbidium giganteum wall through high frequency conversion of encapsulated protocorms under in vivo and in vitro conditions. Ind. J. Exp. Biol. 31, 61–64.
Croom, L., Jackson, C., Vaidya, B., Parajuli, P., and Joshee, N. (2016). Thin cell layer (TCL) culture system for herbal biomass production and genetic transformation of Bacopa monnieri L. Wettst. Am. J. Plant Sci. 7, 1232–1245. doi: 10.4236/ajps.2016.78119
Currais, L., Loureiro, J., Santos, C., and Canhoto, J. M. (2013). Ploidy stability in embryogenic cultures and regenerated plantlets of tamarillo. Plant Cell Tiss. Org. 114, 149–159. doi: 10.1007/s11240-013-0311-5
Datwyler, S. L., and Weiblen, G. D. (2006). Genetic variation in hemp and marijuana (Cannabis sativa L.) according to amplified fragment length polymorphisms. J. Forensic Sci. 51, 371–375. doi: 10.1111/j.1556-4029.2006.00061.x
Dhekney, S. A., Li, Z. T., Grant, T. N. L., and Gray, D. J. (2016). “Somatic embryogenesis and genetic modification of Vitis,” in In Vitro Embryogenesis in higher plants. Methods in Molecular Biology, Vol. 1359, eds M. Germana and M. Lambardi (New York, NY: Humana Press), doi: 10.1007/978-1-4939-3061-6
Dinesh-Kumar, S., Liu, Y., and Schiff, M. (2007). Tobacco rattle virus vectors and related compositions and methods. United States Patent US7229829B2. Alexandria, VA: United States Patent and Trademark Office.
Dufresnes, C., Jan, C., Bienert, F., Goudet, J., and Fumagalli, L. (2017). Broad-scale genetic diversity of Cannabis for forensic applications. PLoS One 12:170522. doi: 10.1371/journal.pone.0170522
Eftekhari, M., Alizadeh, M., Mashayekhi, K., and Asghari, H. R. (2012). In vitro propagation of four Iranian grape varieties: influence of genotype and pretreatment with arbuscular mycorrhiza. Vitis 51, 175–182.
Elhiti, M., Tahir, M., Gulden, R. H., Khamiss, K., and Stasolla, C. (2010). Modulation of embryo-forming capacity in culture through the expression of Brassica genes involved in the regulation of the shoot apical meristem. J. Exp. Bot. 61, 4069–4085. doi: 10.1093/jxb/erq222
Engelmann, F. (2004). Plant cryopreservation: progress and prospects. In Vitro Cell. Dev. 40, 427–433. doi: 10.1079/IVP2004541
Engelmann, F. (2014). Cryopreservation of clonal crops: a review of key parameters. Acta Hort. 1039, 31–39. doi: 10.17660/ActaHortic.2014.1039.2
Faeti, V., Mandolino, G., and Ranalli, P. (1996). Genetic diversity of Cannabis sativa germplasm based on RAPD markers. Plant Breed. 115, 367–370. doi: 10.1111/j.1439-0523.1996.tb00935.x
Farag, S., and Kayser, O. (2015). Cannabinoids production by hairy root cultures of Cannabis sativa L. Am. J. Plant Sci. 6:1874. doi: 10.4236/ajps.2015.611188
Farag, S., and Kayser, O. (2017). The cannabis plant: botanical aspects. In Handbook of Cannabis and Related Pathologies, ed. V. Preedy (Cambridge: Academic Press), 3–12.
Farahani, F., Yari, R., and Masoud, S. (2011). Somaclonal variation in Dezful cultivar of olive (Olea europaea subsp. europaea). Gene Conserve. 10, 216–221.
Feeney, M., and Punja, Z. K. (2003). Tissue culture and Agrobacterium-mediated transformation of hemp (Cannabis sativa L.). In Vitro Cell. Dev. Biol. 39, 578–585. doi: 10.1079/IVP2003454
Fellermeier, M., and Zenk, M. H. (1998). Prenylation of olivetolate by a hemp transferase yields cannabigerolic acid, the precursor of tetrahydrocannabinol. FEBS Lett. 427, 283–285.
Ferrie, A. M., and Mollers, C. (2011). Haploids and doubled haploids in Brassica spp. for genetic and genomic research. Plant Cell Tiss. Org. Cult. 104, 375–386. doi: 10.1007/s11240-010-9831-4
Fisse, J., and Andres, J. (1985). Organogenesis and biosynthesis in an in vitro culture of Cannabis sativa L. Boletin da Academia Galega de Ciencias 4, 57–67.
Fisse, J., Braut, F., Cosson, L., and Paris, M. (1981). Étude in vitro des capacités organogénétiques de tissus de Cannabis sativa L.; Effet de différentes substances de croissance. Plantes Médicinales et Phytotherapi 15, 217–223.
Flaishman, M. A., Cohen, P. R., Cohen, O., and Bocobza, S. (2019). Method of regenerating and transforming cannabis. WIPO/PCT Patent No WO 2019/234750 A1. Geneva: WIPO.
Flores-Sanchez, I. J., Pec, J., Fei, J., Hae-Choi, Y., Dusek, J., and Verpoorte, R. (2009). Elicitation studies in cell suspension cultures of Cannabis sativa L. J. Biotechnol. 143, 157–168. doi: 10.1016/j.jbiotec.2009.05.006
Alisher, T., Forster, B. P., and Jain, S. M. (2007). Advances in haploid production in higher plants. Dordrecht: Springer, 35–46.
Gagne, S. J., Stout, J. M., Liu, E., Boubakir, Z., Clark, S. M., and Page, J. E. (2012). Identification of olivetolic acid cyclase from Cannabis sativa reveals a unique catalytic route to plant polyketides. Proc. Natl. Acad. Sci. 109, 12811–12816. doi: 10.1073/pnas.1200330109
Galán-Ávila, A., García-Fortea, E., Prohens, J., and Herraiz, F. J. (2020). Development of a Direct in vitro plant regeneration protocol From Cannabis sativa L. seedling explants: developmental morphology of shoot regeneration and ploidy level of regenerated plants. Front. Plant Sci. 11:645. doi: 10.3389/fpls.2020.00645
Galazkajoa, J., and Niemirowicz-Szczytt, K. (2013). Review of research on haploid production in cucumber and other cucurbits. Folia Hort. 25, 67–78. doi: 10.2478/fhort-2013-0008
Gao, C., Xin, P., Cheng, C., Tang, Q., Chen, P., Wang, C., et al. (2014). Diversity Analysis in Cannabis sativa Based on Large-Scale Development of Expressed Sequence Tag-Derived Simple Sequence Repeat Markers. PLoS One 9:e110638. doi: 10.1371/journal.pone.0110638
George, E. F., Hall, M. A., and De Klerk, G. J. (eds) (2007). Plant propagation by tissue culture, 3rd Edn. Dordrecht: Springer, doi: 10.1007/978-1-4020-5005-3
Gilles, L. M., Khaled, A., Laffaire, J. B., Chaignon, S., Gendrot, G., Laplaige, J., et al. (2017). Loss of pollen-specific phospholipase NOT LIKE DAD triggers gynogenesis in maize. EMBO J. 36, 707–717. doi: 10.15252/embj.201796603
Gilmore, S., and Peakall, R. (2003). Isolation of microsatellite markers in Cannabis sativa L. (marijuana). Mol. Ecol. Notes 3, 105–107.
Gloss, D. (2015). An overview of products and bias in research. Neurotherapeutics 12, 731–734. doi: 10.1007/s13311-015-0370-x
Grudzinska, M., and Solarska, E. (2004). The elimination of virus and Hop Latent Viroid from Hop (Humulus Lupulus L.) in Poland. Acta Hortic. 668, 149–152. doi: 10.17660/ActaHortic.2005.668.19
Grudzinska, M., Solarska, E., Czubacka, A., Przybys, M., and Fajbus, A. (2006). Elimination of Hop Latent Viroid from Hop plants by cold treatment and meristem tip culture. Phytopathol. Pol. 40, 21–30.
Grulichova, M., Mendel, P., Lalge, A. B., Slamova, N., Trojan, V., Vyhnanek, T., et al. (2017). Effect of different phytohormones on growth and development of micropropagated Cannabis sativa L. Mendel Net. 2017, 618–623.
Hakki, E. E., Uz, E., Sag, A., Atasoy, S., and Akkaya, S. M. (2003). Genotyping of Cannabis sativa L. accessions from Turkey using RAPD and AFLP markers. Forensic Sci. Int. 136:31.
Hanus, L. O., Meyer, S. M., Munoz, E., Taglialatela-Scafati, O., and Appendino, G. (2016). Phytocannabinoids: a unified critical inventory. Nat. Prod. Rep. 33, 1357–1392. doi: 10.1039/c6np00074f
Hartsel, S. C., Loh, W. H. T., and Robertson, L. W. (1983). Biotransformation of cannabidiol to Cannabielsoin by suspension cultures of Cannabis sativa and Saccharum officinarum. Planta Med. 48, 17–19. doi: 10.1055/s-2007-969870
Heitrich, A., and Binder, M. (1982). Identification of (3R,4R)-gD as an isolation artefact of cannabinoid acids formed by callus cultures of Cannabis sativa L. Experientia 38, 898–899. doi: 10.1007/BF01953640
Hemphill, J. K., Turner, J. C., and Mahlberg, P. G. (1978). Studies on growth and cannabinoid composition of callus derived from different strains of Cannabis sativa. Lloydia 41, 453–462.
Henry, P., Hilyard, A., Johnson, S., and Orser, C. (2018). Predicting chemovar cluster and variety verification in vegetative cannabis accessions using targeted single nucleotide polymorphisms. PeerJ Prepr. 6:e27442v1. doi: 10.7287/peerj.preprints.27442v1
Hillig, K. W. (2004). A multivariate analysis of allozyme variation in 93 Cannabis accessions from the VIR germplasm collection. J. Ind. Hemp. 9, 5–22. doi: 10.1300/J237v09n02_02
Hillig, K. W. (2005). Genetic evidence for speciation in Cannabis (Cannabaceae). Genet. Resour. Crop Evol. 52, 161–180. doi: 10.1007/s10722-003-4452-y
Hilling, K. W., and Mahlberg, P. G. (2004). A chemotaxonomic analysis of cannabinoid variationin Cannabis (Cannabaceae). Am. J. Bot. 91, 966–975.
Hussain, A., Qarshi, I. A., Nazir, H., and Ullah, I. (2012). Plant Tissue Culture: current status and opportunities. InTech Open 28:50568. doi: 10.5772/50568
Hussain, S. H. F. (2014). Cannabinoids production in Cannabis sativa L. An in vitro approach. Ph.D. thesis, Dortmund: Technischen Universität Dortmund, 1–138.
Hvoslef-Eide, A. K., and Preil, W. (2005). Liquid culture systems for in vitro plant propagation. Dordrecht: Springer, doi: 10.1007/1-4020-3200-5
Itokawa, H., Takeya, K., and Akasu, M. (1975). Studies on the constituents isolated from the callus of Cannabis sativa L. Shoyakugaku Zasshi 29, 106–112.
Itokawa, H., Takeya, K., and Mihashi, S. (1977). Biotransformation of Cannabinoid precursor and related alcohols by suspension cultures of callus induced from Cannabis sativa L. Chem. Pharm. Bull. 25, 1941–1946. doi: 10.1248/cpb.25.1941
Jekkel, Z., Heszky, L. E., and Ali, A. H. (1989). Effect of different cryoprotectants and transfer temperatures on the survival rate of hemp (Cannabis sativa L.) cell suspension in deep freezing. Acta Biol. Hung. 40, 127–136.
Jevremovic, S., Subotic, A., Miljkovic, D., Trifunovic, M., Petric, M., and Cingel, A. (2012). Clonal fidelity of Chrysanthemum cultivars after long term micropropagation by stem segment culture. Acta Hortic. 961, 211–216. doi: 10.17660/ActaHortic.2012.961.25
Jiang, H., Wang, L., Merlin, M. D., Clarke, R. C., Pan, Y., Zhang, Y., et al. (2016). Ancient Cannabis burial shroud in a Central Eurasian cemetery. Econ. Bot. 70, 213–221. doi: 10.1007/s12231-016-9351-1
Jiang, Y., Zunmin, X. I. A., Tang, Y., Han, Q., and Han, C. (2015). Preliminary studies on the tissue culture of Cannabis sativa L. (Industrial hemp). Agricult. Sci Technol. 16, 923–925.
Jinek, M., Chylinski, K., Fonfara, I., Hauer, M., Doudna, J. A., and Charpentier, E. (2012). A programmable dual-RNA–guided DNA endonuclease in adaptive bacterial immunity. Science 337, 816–821. doi: 10.1126/science.1225829
Joyce, S. M., Cassells, A. C., and Jain, S. M. (2003). Stress and aberrant phenotypes in vitro culture. Plant Cell Tiss. Org. 74, 103–121. doi: 10.1023/A:1023911927116
Karp, A. (1994). “Origins, causes and uses of variation in plant tissue cultures,” in Plant cell and tissue culture, eds I. K. Vasil and T. A. Thorpe (Dordrecht: Kluwer Academic Publishers), 139–152. doi: 10.1007/978-94-017-2681-8
Khan, S., Saeed, B., and Kauser, N. (2011). Establishment of genetic fidelity of in vitro raised banana plantlets. Pak. J. Bot. 43, 233–242.
Kodym, A., and Leeb, C. J. (2019). Back to the roots: protocol for the photoautotrophic micropropagation of medicinal cannabis. Plant Cell Tiss. Org. 138, 399–402. doi: 10.1007/s11240-019-01635
Kojoma, M., Iida, O., Makino, Y., Sekita, S., and Satake, M. (2002). DNA fingerprinting of Cannabis sativa using inter-simple sequence repeat (ISSR) amplification. Planta Med. 68, 60–63. doi: 10.1055/s-2002-19875
Krishna, H., Alizadeh, M., Singh, D., Singh, U., Chauhan, N., Eftekhari, M., et al. (2016). Somaclonal variations and their applications in horticultural crops improvement. Biotech 6, 1–18. doi: 10.1007/s13205-016-0389-7
Lata, H., Chandra, S., Khan, I. A., and ElSohly, M. A. (2010b). Cannabis sativa L. Micropropagation in temporary immersion bioreactor system. Planta Med. 76:9. doi: 10.1055/s-0030-1251771
Lata, H., Chandra, S., Khan, I., and ElSohly, M. A. (2009a). Propagation through alginate encapsulation of axillary buds of Cannabis sativa L. an important medicinal plant. Physiol. Mol. Biol. Plants 15, 79–86. doi: 10.1007/s12298-009-0008-8
Lata, H., Chandra, S., Khan, I., and ElSohly, M. A. (2009b). Thidiazuron-induced high-frequency direct shoot organogenesis of Cannabis sativa L. Vitr. Cell. Dev. Biol. Plant 45, 12–19. doi: 10.1007/s11627-008-9167-5
Lata, H., Chandra, S., Khan, I., and ElSohly, M. A. (2010a). Assessment of genetic stability of micropropagated Cannabis sativa plants by ISSR markers. Planta Med. 76, 97–100. doi: 10.1055/s-0029-1185945
Lata, H., Chandra, S., Khan, I., and ElSohly, M. A. (2010c). High frequency plant regeneration from leaf derived callus of high D9-tetrahydrocannabinol yielding Cannabis sativa L. Planta Med. 76, 1629–1633. doi: 10.1055/s-0030-1249773
Lata, H., Chandra, S., Khan, I., and Elsohly, M. A. (2017). Micropropagation of Cannabis sativa L.—an update. Cannabis sativa L. Bot. Biotechnol. 2017, 285–297. doi: 10.1007/978-3-319-54564-6_13
Lata, H., Chandra, S., Mehmedic, Z., Khan, I., and ElSohly, M. A. (2011). In vitro germplasm conservation of high Tetrahydocannab- inol yielding elite clones of Cannabis sativa L. under slow growth conditions. Acta Physiol. Plant. 34, 743–750. doi: 10.1007/s11738-011-0874-x
Lata, H., Chandra, S., Techen, N., Khan, I. A., and ElSohly, M. A. (2016). In vitro mass propagation of Cannabis sativa L.: A protocol refinement using novel aromatic cytokinin meta-topolin and the assessment of eco-physiological, biochemical and genetic fidelity of micropropagated plants. J. Appl. Res. Med. Aromat. Plants. 3, 18–26. doi: 10.1016/j.jarmap.2015.12.001
Laverty, K. U., Stout, J. M., Sullivan, M. J., Shah, H., Gill, N., Holbrook, L., et al. (2019). A physical and genetic map of Cannabis sativa identifies extensive rearrangements at the THC/CBD acid synthase loci. Genome Res. 29, 146–156. doi: 10.1101/gr.242594.118
Lei, R., Qiao, W., Fan, H., and Jiang, H. (2015). A simple and effective method to encapsulate tobacco mesophyll protoplasts to maintain cell viability. MethodX 2, 24–32. doi: 10.1016/j.mex.2014.11.004
Li, H. (1974). An archaeological and historical account of cannabis in China. Econ. Bot. 28, 437–448. doi: 10.1007/BF02862859
Lineberger, R. D. (1983). Shoot proliferation, rooting, and transplant survival of tissue-culture ‘Hally-Jolivette’ cherry. HortScience 18, 182–185. doi: 10.1007/BF00044251
Liu, Y., Schiff, M., and Dinesh-Kumar, S. (2002). Virus-induced gene silencing in tomato. Plant J. 31, 777–786. doi: 10.1046/j.1365-313x.2002.01394.x
Loh, W. H. T., Hartsel, S. C., and Robertson, L. W. (1983). Tissue culture of Cannabis sativa L. and in vitro biotransformation of phenolics. Zeitschrift fuer P?anzenphysiol. 111, 395–400. doi: 10.1016/S0044-328X(83)80003
Mackinnon, L., Mcdougall, G., Aziz, N., and Millam, S. (2000). Progress towards transformation of fibre hemp. Scottish Crop Research Institute Annual Report 2000/2001. Dundee: Scottish Crop Research Institute.
Malik, M. R., Wang, F., Dirpaul, J. M., Zhou, N., Polowick, P. L., Ferrie, A. M., et al. (2007). Transcript profiling and identification of molecular markers for early microspore embryogenesis in Brassica napus. Plant Physiol. 144, 134–154.
Mandolino, G., and Ranalli, P. (1999). Advances in biotechnological approaches for hemp breeding and industry. Haworth: Advances in hemp research, doi: 10.1201/9781498705820
McKey, D., Elias, M., Pujol, M. E., and Duputié, A. (2010). The evolutionary ecology of clonally propagated domesticated plants. N. Phytol. 186, 318–332. doi: 10.1111/j.1469-8137.2010.03210.x
McPartland, J. M. (2018). Cannabis systematics at the levels of family, genus, and species. Cannabis Cannabinoid Res. 3, 203–212.
Mechoulam, R. (1986). “The pharmacohistory of Cannabis sativa,” in Cannabinoids as Therapeutic Agents, ed. R. Mechoulam (Boca Raton, FL: CRC Press), 1–19. doi: 10.1201/9780429260667-1
Mendoza, M. A., Mills, D. K., Lata, H., Chandra, S., ElSohly, M. A., and Almirall, J. R. (2009). Genetic individualization of Cannabis sativa by short tandem repeat multiplex system. Anal. Bioanal. Chem. 393, 719–726. doi: 10.1007/s00216-008-2500-3
Micke, A. (1999). “Mutation in plant breeding,” in Breeding in crop plants – mutations and in vitro mutation breeding, eds B. A. Siddiqui and S. Khan (New Delhi: Kalyani Publication), 1–19. doi: 10.1007/978-1-4614-7028-1
Miguel, C., and Marum, L. (2011). An epigenetic view of plant cells cultured in vitro: somaclonal variation and beyond. J. Exp. Bot. 62, 3713–3725. doi: 10.1093/jxb/err155
Mirmasoumi, M., Azadi, P., Sharafi, A., Ntui, V. O., and Mii, M. (2013). Simple protocol for plant regeneration of Lilium ledebouriiusing transverse thin cell layer. Prog. Biol. Sci. 3, 117–122. doi: 10.22059/PBS.2013.35828
Mordhorst, A. P., Hartog, M. V., El-Tamer, M. K., Laux, T., and De Vries, S. C. (2002). Somatic embryogenesis from Arabidopsis shoot apical meristem mutants. Planta 214, 829–836. doi: 10.1007/s00425-001-0700-6
Mori, K. (1971). Production of virus-free carnations by means of meristem culture. Netherl. Plant Pathol. J. 6, 1–7. doi: 10.1007/BF02102383
Morimoto, S., Tanaka, Y., Sasaki, K., Tanaka, H., Fukamizu, T., Shoyama, Y., et al. (2007). Identification and characterization of cannabinoids that induce cell death through mitochondrial permeability transition in Cannabis leaf cells. J. Biol. Chem. 282, 20739–20751. doi: 10.1074/jbc.M700133200
Movahedi, M., Ghasemi-Omran, V. O., and Torabi, S. (2015). The effect of different concentrations of TDZ and BA on in vitro regeneration of Iranian Cannabis (Cannabis sativa) using cotyledon and epicotyl explants. J. Plant Mol. Breed. 3, 20–27. doi: 10.22058/JPMB.2015.15371
Murashige, T. (1977). “Plant cell and organ cultures as horticultural practices,” in Proceedings of the Symposium on Tissue Culture for Horticultural Purposes, (Ghent: ISHS), doi: 10.17660/ActaHortic.1977.78.1
Murthy, B. N. S., Murch, S. J., and Saxena, P. K. (1995). Thidiazuron-induced somatic embryogenesis in intact seedlings of peanut (Arachis hypogaeay). Endogenous growth regulator levels and significance of cotyledons. Physiol. Plant. 94, 268–276.
Musio, S., Müssig, J., and Amaducci, S. (2018). Optimizing hemp fiber production for high performance composite applications. Front. Plant Sci. 9:1702. doi: 10.3389/fpls.2018.01702
National Clean Plant Network (2020). NCPN Factsheets, Brochures and Posters. Auburn, AL: National Clean Plant Network.
Nhut, D. T., Teixeira, da Silva, J. A., Van Le, B., and Thanh Van, K. T. (2003). “Thin cell Layer (TCL) Morphogenesis as a powerful tool in woody plant and fruit crop micropropagation and biotechnology, floral genetics and genetic transformation,” in Micropropagation of woody trees and fruits. Forestry Sciences, Vol. 75, eds S. M. Jain and K. Ishii (Dordrecht: Springer), 783–814. doi: 10.1007/978-94-010-0125-0_27
Nwauzoma, A. B., and Jaja, E. T. (2013). A review of somaclonal variation in plantain (Musa spp): mechanisms and applications. J. Appl. Biosci. 67, 5252–5260. doi: 10.4314/jab.v67i0.95046
Nyende, A. B., Schittenhelm, S., Mix-Wagner, G., and Greef, J. M. (2003). Production, storability, and regeneration of shoot tips of potato (Solanum tuberosum L.) encapsulated in calcuim alginate hollow beads. In Vitro Cell. Dev. Biol. Plant. 39, 540–544. doi: 10.1079/IVP2003442
Oladosu, Y., Rafii, M. Y., Abdullah, N., Hussin, G., Ramli, A., Rahim, H. A., et al. (2016). Principle and application of plant mutagenesis in crop improvement: A review. Biotechnol. Biotechnol. 30, 1–16. doi: 10.1080/13102818.2015.1087333
Osborne, D. J., and McManus, M. T. (2005). Hormones signals and target cells in plant development. Cambridge: Cambridge University Press, doi: 10.1017/CBO9780511546228
Page, L. S. R. G., Monthony, A. S., and Jones, A. M. P. (2020). Basal media optimization for the micropropagation and callogenesis of Cannabis sativa. bioRxiv preprint. doi: 10.1101/2020.02.07.939181
Pathak, H., and Dhawan, V. (2012). ISSR assay for ascertaining genetic fidelity of micropropagated plants of apple rootstock Merton 793. In Vitro Cell. Dev. Biol. Plant. 48, 137–143. doi: 10.1007/s11627-011-9385-0
Pence, V. C. (2011). Evaluating costs for the in vitro propagation and preservation of endangered plants. In Vitro Cell. Dev. Biol. Plant. 47, 176–187. doi: 10.1007/s11627-010-9323-6
Pijut, P. M., Beasley, R. R., Lawson, S. S., Palla, K. J., Stevens, M. E., and Wang, Y. (2012). In vitro propagation of tropical hardwood tree species—a review (2001–2011). Propag. Ornam. Plants 12, 25–51.
Piomelli, D., and Russo, E. B. (2016). The Cannabis sativa versus Cannabis indica debate: An interview with Ethan Russo, MD. Cannabis Cannabinoid Res. 1, 44–46. doi: 10.1089/can.2015.29003.ebr
Pistelli, L., Giovannini, A., Ruffoni, B., Bertoli, A., and Pistelli, L. (2010). “Hairy root cultures for secondary metabolites production,” in Bio-Farms for Nutraceuticals, eds G. Rea, M. T. Giardi, and B. Berra (Boston, MA: Springer), 167–184. doi: 10.1007/978-1-4419-7347-4
Piunno, K. F., Golenia, G., Boudka, E. A., Downey, C., and Jones, A. M. (2019). Regeneration of shoots from immature and mature inflorescences of Cannabis sativa. Can. J. Plant Sci. 99, 556–559. doi: 10.1139/cjps-2018-0308
Plawuszewski, M., Lassocinski, W., and Wieglus, K. (2005). “Regeneration of polish cultivars of monoecious hemp (Cannabis sativa L.) grown in vitro,” in Renewable resources and plant biotechnology, eds R. Kozlowski, E. Gennady, and F. Pudel (New York, NY: Nova Science Publishers Inc.), 149–154.
Popova, E., Shukla, M., and Kim, H. H. (2015). “Plant cryopreservation for biotechnology and breeding,” in Advances in plant breeding strategies: breeding, biotechnology and molecular Tools, eds J. M. AI-Khayri, S. M. Jain, and D. V. Johnson (Berlin: Springer International Publishing).
Potter, D. J. (2009). The propagation, characterisation and optimisation of Cannabis sativa L. as a phytopharmaceutical. Ph.D. thesis, London: King’s College London, 255.
Rai, M. K., Jaiswal, V. S., and Jaiswal, U. (2008). Encapsulation of shoot tips of guava (Psidium guajava L.) for short-term storage and germplasm exchange. Sci. Hortic. 118, 33–38. doi: 10.1016/j.scienta.2008.05.017
Rea, K. A., Casaretto, J. A., Al-Abdul-Wahid, M. S., Sukumaran, A., Geddes-McAlister, J., Rothstein, S. J., et al. (2019). Biosynthesis of cannflavins A and B from Cannabis sativa L. Phytochem 164, 162–171. doi: 10.1016/j.phytochem.2019.05.009
Ren, J., Wu, P., Trampe, B., Tian, X., Lübberstedt, T., and Chen, S. (2017). Novel technologies in doubled haploid line development. Plant Biotechnol. J. 15, 1361–1370. doi: 10.1111/pbi.12805
Richez-Dumanois, C., Braut-Boucher, F., Cosson, L., and Paris, M. (1986). Multiplication vegetative in vitro du chanvre (Cannabis sativa L.) application a la conservation des clones selectionnes. Agronomie 6, 487–495.
Rihan, H. Z., Kareem, F., El-Mahrouk, M. E. E., and Fuller, M. P. (2017). Artificial seeds principle, aspect, and applications. Agronomy 7:7040071. doi: 10.3390/agronomy7040071
Rodriguez-Leyva, D., and Pierce, G. N. (2010). The cardiac and hemostatic effects of dietary hempseed. Nutr. Metab. 7, 1–9. doi: 10.1186/1743-7075-7-32
Rotherham, D., and Harbison, S. A. (2011). Differentiation of drug and non-drug Cannabis using single nucleotide polymorphism (SNP) assay. Forensic Sci. Int. 207, 193–197. doi: 10.1016/j.forsciint.2010.10.006
Sakai, A., and Engelmann, F. (2007). Vitrification, encapsulation-vitrification and droplet-vitrification: a review. Cryo. Lett. 28, 151–172.
Sallie, J. V., and Jones, O. P. (2015). The culture of shoot tips of hop (Humulus lupulus L.) to eliminate viruses. J. Hortic. Sci. 44, 281–284. doi: 10.1080/00221589.1969.11514310
Sato, M., Hosokawa, M., and Doi, M. (2011). Somaclonal variation is induced de novo via the tissue culture process: a study quantifying mutated cells in Saintpaulia. PLoS One 6:23541. doi: 10.1371/journal.pone.0023541
Sawler, J., Stout, J. M., Gardner, K. M., Hudson, D., Vidmar, J., Butler, L., et al. (2015). The Genetic structure of Marijuana and Hemp. PLoS One 26:e0133292. doi: 10.1371/journal.pone.0133292
Schachtsiek, J., Hussain, T., Azzouhri, K., Kayser, O., and Stehle, F. (2019). Virus-induced gene silencing (VIGS) Cannabis sativa L. Plant Methods 15:157. doi: 10.1186/s13007-019-0542-5
Schultes, R. E., Klein, W. M., Plowman, T., and Lockwood, T. E. (1974). Cannabis: an example of taxonomic neglect. Botanic. Museum Leaflets 23, 337–367.
Senthil-Kumar, M., and Mysore, K. S. (2014). Tobacco rattle virus–based virus-induced gene silencing in Nicotiana benthamiana. Nat. Protoc. 9, 1549–1562. doi: 10.1038/nprot.2014.092
Sharp, W. R., Sondahl, M. R., Caldas, L. S., and Maraffa, S. B. (1980). The physiology of in vitro asexual embryogenesis. Hortic. Rev. 2, 268–310. doi: 10.1002/9781118060759.ch6
Shen, H. J., Chen, J. T., Chung, H. H., and Chang, W. C. (2018). Plant regeneration via direct somatic embryogenesis from leaf explants of Tolumnia Luise Elmore ‘Elsa.’. Bot. Stud. 59:4. doi: 10.1186/s40529-018-0220-3
Shew, A. M., Nalley, L. L., Snell, H. A., Nayga, R. M., and Dixon, B. L. (2018). CRISPR versus GMOs: Public acceptance and valuation. Glob. Food Secur. 19, 71–80. doi: 10.1016/j.gfs.2018.10.005
Sirikantaramas, S., Morimoto, S., Shoyama, Y., Ishikawa, Y., Wada, Y., Shoyama, Y., et al. (2004). The gene controlling marijuana psychoactivity: molecular cloning and heterologous expression of Delta1-tetrahydrocannabinolic acid synthase from Cannabis sativa L. J. Biol. Chem. 279, 39767–39774. doi: 10.1074/jbc.M403693200
Sirkowski, E. (2012). Marked Cannabis for indicating medical Marijuana. US Patent 20120311744. Alexandria, VA: United States Patent and Trademark Office.
Sluis, C. J. (2005). “Integrating automation technologies with commercial micropropagation,” in Plant Tissue Culture Engineering, eds S. D. Gupta and Y. Ibaraki (Berlin: Springer), 231–251. doi: 10.1007/978-1-4020-3694-1
Slusarkiewicz-Jarzina, A., Ponitka, A., and Kaczmarek, Z. (2005). Influence of cultivar, explant source and plant growth regulator on callus induction and plant regeneration of Cannabis sativa L. Acta Biol. Craco. Ser. Bot. 47, 145–151.
Small, E. (2004). “Narcotic Plants as sources of medicinal, nutraceuticals, and functional foods,”. In Proceedings of the International Symposium on the Development of Medicinal Plants. Canada: Agriculture and Agri-Food, 11–67.
Small, E. (2015). Evolution and Classification of Cannabis sativa (Marijuana, Hemp) in Relation to Human Utilization. Bot. Rev. 81, 189–294.
Small, E. (2017). “Classification of Cannabis sativa L. in relation to agricultural, biotechnological, medical and recreational utilization,” in Cannabis sativa L. - Botany and Biotechnology, eds S. Chandra, H. Lata, and M. ElSohly (Cham: Springer).
Small, E., and Cronquist, A. (1976). A practical and natural taxonomy for cannabis. Taxon 25, 405–435. doi: 10.2307/1220524
Small, E., Jul, P. Y., and Lefkovitch, L. P. (1976). A numerical taxonomic analysis of Cannabis with special reference to species. Syst. Bot. 1, 67–84. doi: 10.2307/2418840
Smulders, M., and de Klerk, G. (2011). Epigenetics in plant tissue culture. Plant Growth Regul. 63, 137–146. doi: 10.1007/s10725-010-9531-4
Smýkalová, I., Vrbová, M., Cvečková, M., Plackova, L., Zukauskaite, A., Zatloukal, M., et al. (2019). The effects of novel synthetic cytokinin derivatives and endogenous cytokinins on the in vitro growth responses of hemp (Cannabis sativa L.) explants. Plant Cell Tiss. Org. 139, 381–394. doi: 10.1007/s11240-019-01693
Soriano, M., Li, H., and Boutilier, K. (2013). Microspore embryogenesis: establishment of embryo identity and pattern in culture. Plant Reprod. 26, 181–196. doi: 10.1007/s00497-013-0226-7
Spanò, R., Bottalico, G., Corrado, A., Campanale, A., Di Franco, A., and Mascia, T. (2018). A protocol for producing virus-free artichoke genetic resources for conservation, breeding, and production. Agriculture 8:36. doi: 10.3390/agriculture8030036
Stout, J. M., Boubakir, Z., Ambrose, S. J., Purves, R. W., and Page, J. E. (2012). The hexanoyl-CoA precursor for cannabinoid biosynthesis is formed by an acyl-activating enzyme in Cannabis sativa trichomes. Plant J. 71, 353–365. doi: 10.1111/j.1365-313X.2012.04949.x
Sun, S., Zhong, J., Li, S., and Wang, X. (2013). Tissue culture-induced somaclonal variation of decreased pollen viability in torenia (Torenia fournieri Lind.). Bot. Stud. 54:36. doi: 10.1186/1999-3110-54-36
Szarejko, I. (2003). “Doubled haploid mutant production,” in Doubled haploid production in crop plants, eds M. Maluszynski, K. J. Kasha, B. P. Forster, and I. Szarejko (Dordrecht: Springer), doi: 10.1007/978-94-017-1293-4_48
Taura, F., Morimoto, S., and Shoyama, Y. (1995). First direct evidence for the mechanism of Δ1-tetrahydrocannabinolic acid biosynthesis. J. Am. Chem. Soc. 117, 9766–9767. doi: 10.1021/ja00143a024
Taura, F., Sirikantaramas, S., Shoyama, Y., Shoyama, Y., and Morimoto, S. (2007a). Phytocannabinoids in Cannabis sativa: recent studies on biosynthetic enzymes. Chem. Biodivers. 4, 1649–1663. doi: 10.1002/cbdv.200790145
Taura, F., Sirikantaramas, S., Shoyama, Y., Yoshikai, K., Shoyama, Y., and Morimoto, S. (2007b). Cannabidiolic-acid synthase, the chemotype- determining enzyme in the fiber-type Cannabis sativa. FEBS Lett. 581, 2929–2934. doi: 10.1016/j.febslet.2007.05.043
Taura, F., Tanaka, S., Taguchi, C., Fukamizu, T., Tanaka, H., Shoyama, Y., et al. (2009). Characterization of olivetol synthase, a polyketide synthase putatively involved in cannabinoid biosynthetic pathway. FEBS Lett. 583, 2061–2066. doi: 10.1016/j.febslet.2009.05.024
Teixeira da Silva, J. A., Tran Thanh, Van, K., Biondi, S., Nhut, D. T., et al. (2007). Thin cell layers: developmental building blocks in ornamental biotechnology. Floricult. Ornamental Biotech. 1, 1–13.
Thacker, X., Thomas, K., Fuller, M., Smith, S., and DuBois, J. (2018). Determination of optimal hormone and mineral salts levels in tissue culture media for callus induction and growth of industrial hemp (Cannabis sativa L.). J. Agric. Sci. 9, 1250–1268. doi: 10.4236/as.2018.910088
Uchendu, E., Lata, H., Chandra, S., Khan, I., and ElSohly, M. A. (2019). Cryopreservation of shoot tips of elite cultivars of Cannabis sativa L. by droplet vitrification. Med. Cannabis cannabinoids 2, 29–34. doi: 10.1159/000496869
Veliky, I. A., and Genest, K. (1972). Growth and metabolites of Cannabis sativa cell suspension cultures. Lloydia 35, 450–456.
Wahby, I., Arráez-Román, D., Segura-Carretero, A., Ligero, F., Caba, J. M., and Fernández-Gutiérrez, A. (2006). Analysis of choline and atropine in hairy root cultures of Cannabis sativa L. by capillary electrophoresis-electrospray mass spectrometry. Electrophoresis 27, 2208–2215. doi: 10.1002/elps.200500792
Wahby, I., Caba, J. M., and Ligero, F. (2013). Agrobacterium infection of hemp (Cannabis sativa L.): establishment of hairy root cultures. J. Plant Interact. 8, 312–320. doi: 10.1080/17429145.2012.746399
Wahby, I., Caba, J. M., and Ligero, F. (2017). “Hairy root culture as a biotechnological tool in C. sativa,” in Cannabis sativa L. Botany and Biotechnology, eds M. ElSohly, H. Lata, and C. Suman (Cham: Springer), 299–317. doi: 10.1007/978-3-319-54564-6
Wang, M., Wang, Y. H., Avula, B., Radwan, M. M., Wanas, A. S., van Antwerp, J., et al. (2016). Decarboxylation study of acidic cannabinoids: A novel approach using ultra-high-performance supercritical fluid chromatography/photodiode array-mass spectrometry. Cannabis Cannabinoid Res. 1, 262–271. doi: 10.1089/can.2016.0020
Wang, P. J., and Charles, A. (1991). “Micropropagation through meristem culture,” in High-Tech and Micropropagation I. Biotechnology in Agriculture and Forestry, Vol. 17, ed. Y. P. S. Bajaj (Berlin: Springer), doi: 10.1007/978-3-642-76415-8
Wang, R., He, L. S., Xia, B., Tong, J. F., Li, N., and Peng, F. (2009). A micropropagation system for cloning of hemp (Cannabis Sativa L.) by shoot tip culture. Pak. J. Bot. 41, 603–608.
Wargent, E. T., Zaibi, M. S., Silvestri, C., Hislop, D. C., Stocker, C. J., Stott, C. G., et al. (2013). The cannabinoid Δ9-tetrahydrocannabivarin (THCV) ameliorates insulin sensitivity in two mouse models of obesity. Nutr. Diabetes 3:e68. doi: 10.1038/nutd.2013.9
Watt, M. P., Thokoane, N. L., Mycock, D., and Blakeway, F. (2000). In vitro storage of Eucalyptus grandis germplasm under minimal growth conditions. Plant Cell Tiss. Org. 61, 161–164. doi: 10.1023/A:1006447506869
Whitton, P. A. (2019). Method of production of phytocannabinoids for use in medical treatments. US Patent 10,477,791 B2. Alexandria, VA: USPTO.
Whitton, P. A. (2020). Method of production of phytocannabinoids for use in medical treatments. US Patent US2020/0060111 A1. Alexandria, VA: USPTO.
Wielgus, K., Luwanska, A., Lassocinski, W., and Kaczmarek, Z. (2008). Estimation of Cannabis sativa L. tissue culture conditions essential for callus induction and plant regeneration. J. Nat. Fibers 5, 199–207. doi: 10.1080/15440470801976045
Withers, L. A., and Engelmann, F. (1997). In vitro conservation of plant genetic resources. The Netherlands: Springer.
Wu, H., Qu, X., Dong, Z., Luo, L., Shao, C., Forner, J., et al. (2020). WUSCHEL triggers innate antiviral immunity in plant stem cells. Science 370, 227–231. doi: 10.1126/Science.abb7360
Keywords: Cannabis sativa, micropropagation, tissue culture, hemp, in vitro
Citation: Adhikary D, Kulkarni M, El-Mezawy A, Mobini S, Elhiti M, Gjuric R, Ray A, Polowick P, Slaski JJ, Jones MP and Bhowmik P (2021) Medical Cannabis and Industrial Hemp Tissue Culture: Present Status and Future Potential. Front. Plant Sci. 12:627240. doi: 10.3389/fpls.2021.627240
Received: 16 November 2020; Accepted: 04 February 2021;
Published: 03 March 2021.
Edited by:
Derek Stewart, The James Hutton Institute, United KingdomReviewed by:
Daniel A. Jacobo-Velázquez, Monterrey Institute of Technology and Higher Education (ITESM), MexicoCopyright © 2021 Adhikary, Kulkarni, El-Mezawy, Mobini, Elhiti, Gjuric, Ray, Polowick, Slaski, Jones and Bhowmik. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Pankaj Bhowmik, UGFua2FqLkJob3dtaWtAbnJjLWNucmMuZ2MuY2E=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.