
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Plant Sci. , 25 February 2019
Sec. Plant Metabolism and Chemodiversity
Volume 10 - 2019 | https://doi.org/10.3389/fpls.2019.00165
 Adam M. Lakusta1†
Adam M. Lakusta1† Moonhyuk Kwon1,2†
Moonhyuk Kwon1,2† Eun-Joo G. Kwon1
Eun-Joo G. Kwon1 Solomon Stonebloom3
Solomon Stonebloom3 Henrik V. Scheller3
Henrik V. Scheller3 Dae-Kyun Ro1*
Dae-Kyun Ro1*Guayule (Parthenium argentatum) is a perennial shrub in the Asteraceae family and synthesizes a high quality, hypoallergenic cis-1,4-polyisoprene (or natural rubber; NR). Despite its potential to be an alternative NR supplier, the enzymes for cis-polyisoprene biosynthesis have not been comprehensively studied in guayule. Recently, implications of the protein complex involving cis-prenyltransferases (CPTs) and CPT-Binding Proteins (CBPs) in NR biosynthesis were shown in lettuce and dandelion, but such protein complexes have yet to be examined in guayule. Here, we identified four guayule genes – three PaCPTs (PaCPT1-3) and one PaCBP, whose protein products organize PaCPT/PaCBP complexes. Co-expression of both PaCBP and each of the PaCPTs could complemented the dolichol (a short cis-polyisoprene)-deficient yeast, whereas the individual expressions could not. Microsomes from the PaCPT/PaCBP-expressing yeast efficiently incorporated 14C-isopentenyl diphosphate into dehydrodolichyl diphosphates; however, NR with high molecular weight could not be synthesized in in vitro assays. Furthermore, co-immunoprecipitation and split-ubiquitin yeast 2-hybrid assays using PaCPTs and PaCBP confirmed the formation of protein complexes. Of the three PaCPTs, guayule transcriptomics analysis indicated that the PaCPT3 is predominantly expressed in stem and induced by cold-stress, suggesting its involvement in NR biosynthesis. The comprehensive analyses of these PaCPTs and PaCBP here provide the foundational knowledge to generate a high NR-yielding guayule.
Natural rubber (NR) is an isoprene biopolymer, chemically defined as cis-1,4-polyisoprene, and it is used to manufacture a number of medical and industrial products (Cornish, 2001; van Beilen and Poirier, 2007). NR is best known for its prized physical properties, such as elasticity, abrasion resistance, heat dispersion, and hysteresis, which have special importance for the products used in medical and heavy-duty industries. Despite significant advances in polymer science, no material can so far fully replicate the qualities of NR (Mooibroek and Cornish, 2000). The key feature that distinguishes the NR from other synthetic polymers is that NR is a stereo-regular polymer. The isoprene monomeric units of NR are condensed entirely in cis-1,4-configuration in average Mw of ∼1 million Da. Such uniform stereo-structures are theorized to permit NR to form an extremely lengthy spring-like biomolecule, whereas synthetic rubbers are not exclusively stereo-regular (Morton, 2013). These properties, therefore, make NR a strategically important, irreplaceable commodity in modern manufacturing industries.
Natural rubber is synthesized in >7,000 plant species (Rivano et al., 2013). Nonetheless, the rubber tree (Para rubber tree or the Brazilian rubber tree; Hevea brasiliensis) from the Euphorbiaceae family is an almost exclusive NR-producing plant species. The rubber tree synthesizes NR on rubber particles in latex, the cytoplasmic content of the specialized laticifer cells (Cornish et al., 1999). The supply of NR solely depended on the rubber trees, mostly (>90%) cultivated in Southeast Asian countries (van Beilen and Poirier, 2007). During World War II, the supply route of the NR from Southeast Asia was blocked by the Japanese Empire, and in search of alternative NR-producing plants, American and Russian researchers identified guayule (Parthenium argentatum) and the Russian dandelion (Taraxacum kok-saghyz), respectively, as these plants can also synthesize high quality NR (Mooibroek and Cornish, 2000; van Beilen and Poirier, 2007). However, the research activities to develop alternative rubber plants ceased at the end of World War II.
Guayule is a perennial desert shrub native to the northern states of Mexico, but it also grows in the southern states of Texas, New Mexico, and Arizona in the United States. Guayule is unique in its ability to grow in some of North America’s most arid lands, receiving as little as 25–38 cm of rainfall annually (Rasutis et al., 2015). In addition to having little need of water, guayule is known to have dry weights ranging from 4 to 12% in NR (Cantú et al., 1997). Unlike rubber tree, guayule does not have laticifers, but instead, synthesizes NR in the parenchyma cells in stem (Kajiura et al., 2018).
The NR biosynthetic mechanism has not been fully elucidated yet, and researchers have focused on the cis-prenyltransferase (CPT) class of enzymes, as these enzymes are able to catalyze the condensation of isopentenyl diphosphate (IPP) onto a priming molecule in an exclusively cis-configuration (Teng and Liang, 2012). The prokaryotic CPT family enzyme, undecaprenyl diphosphate synthase (UPPS), catalyzes the condensations of IPP onto farnesyl diphosphate (FPP) to form primarily C55 undecaprenyl diphosphate (eight IPP molecules condensed to one FPP) (Shimizu et al., 1998). Its monophosphate form, undecaprenyl phosphate, is a vital lipid molecule that mediates sugar transfer for peptidoglycan cell-wall biosynthesis in prokaryotes. The crystal structure study demonstrated UPPS to be a soluble, homo-dimeric enzyme localized in the prokaryotic cytosol (Fujihashi et al., 2001). Similarly, a homologous eukaryotic CPT enzyme, dehydrodolichyl diphosphate synthase (DDS), catalyzes the analogous cis-condensation reaction to form C80–C120 cis-polyprenyl diphosphate, which is converted to dolichyl phosphate by a phosphatase, polyprenol reductase, and dolichol kinase (Denecke and Kranz, 2009). The resulting dolichyl phosphate serves as a sugar carrier molecule for the post-translational modification of proteins in all eukaryotes. Therefore, both UPPS and DDS are essential metabolic enzymes, and lesions on these genes are lethal (Sato et al., 1999; Zhang et al., 2008). Interestingly, the primary and tertiary structures of CPTs display no similarities to those of trans-prenyltransferases, indicating that cis- and trans-prenyltransferases do not share a common origin and have independently evolved for distinct stereo-selective catalytic reactions (Fujihashi et al., 2001; Lu et al., 2009).
Although comprehensive biochemical data for the prokaryotic CPTs have been documented in the literature, in vitro biochemical characterizations of eukaryotic CPTs has been relatively scarce in the literature until recently. This discrepancy was first resolved by the discovery of the human CPT binding partner, NogoB receptor. Human CPT forms a hetero-protein complex with the NogoB receptor and becomes active in dolichol biosynthesis (Harrison et al., 2011). Similar complexes comprised of CPT and its binding protein homolog were also found in lettuce, tomato, and Arabidopsis for dehydrodolichyl diphosphate biosynthesis (Brasher et al., 2015; Qu et al., 2015; Kwon et al., 2016). The CPT binding proteins, including the NogoB receptor, were named differently in each species. In this work, they are referred to as CBP (CPT-binding protein). CBP is only found in the eukaryotic lineage from yeast to humans, but interestingly, not in prokaryotes (Brasher et al., 2015; Qu et al., 2015). CBP displays low sequence homology to CPTs, forms a complex with CPT on the endoplasmic reticulum (ER), and lacks all conserved motifs required for CPT activity. Therefore, it is believed that CBP interacts with, activates, and tethers CPT onto the eukaryotic ER, but does not function directly in polymerization.
Intriguingly, RNA interference studies showed that analogous CPT–CBP complexes are also involved in NR biosynthesis in lettuce and dandelion (Epping et al., 2015; Qu et al., 2015). However, in vitro reconstitution solely using microsomal CPT and CBP recombinant proteins failed to produce cis-polyisoprenes longer than the short polymer, dolichol (Qu et al., 2015). It is worth noting that lettuce encodes two pairs of CPT–CBP genes – one for the dehydrodolichyl diphosphate biosynthesis within primary metabolism, and the other for NR biosynthesis for the specialized (or secondary) metabolism in latex (Qu et al., 2015). Therefore, at least in lettuce and related species, such as dandelion, it is evident that the NR biosynthesis has functionally diverged from the primary dolichol metabolic pathway.
Aside from CPT and CBP, rubber elongation factor (REF) and its homolog, small rubber particle protein (SRPP), have been suggested to play roles in NR biosynthesis (Dennis and Light, 1989; Oh et al., 1999). However, silencing REF/SRPP homologs showed no decrease in lettuce and no substantial reduction (40–50%) in dandelion (Hillebrand et al., 2012; Chakrabarty et al., 2015). These results are in contrast to the results from CPT- or CBP-silenced dandelion and lettuce where almost complete disappearance of NR was observed (Post et al., 2012; Qu et al., 2015). Thus, the REF and SRPP are not likely to be critical catalytic components in NR biosynthesis. It should be also noted that REF/SRPP homologs are also found as the lipid-droplet associated proteins in non-NR producing plants (Arabidopsis, tobacco, and avocado; Horn et al., 2013). The REF and SRPP are thus not restricted to the NR-producing plants but are likely to be common structural proteins of lipid-droplets or related monolayer structures in all plants. Recently, three new proteins were identified by using photo-affinity substrate and were proposed to be the components of NR-biosynthesizing protein complexes (Cornish et al., 2018). However, their in vivo roles and protein sequences remain unknown.
While the evidence to support the model for CPT–CBP complex in the NR biosynthesis exists in lettuce and dandelion, both species are closely related to each other. Each of these species belongs to the same Cichorieae tribe of the Cichorioideae subfamily within the Asteraceae family. Outside lettuce and dandelion (Cichorieae tribe), the rubber tree and guayule are the only plants that are commercially cultivated for NR production. Of these two, guayule (P. argentatum) is from the same Asteraceae family as lettuce and dandelion, but it comes from a different subfamily and tribe (Asteroideae and Heliantheae, respectively). On the other hand, the rubber tree (H. brasiliensis) belongs to an entirely different family, Euphorbiaceae. Recently, studies of CPT and CBP from the rubber tree suggested that its CPTs interact with its CBP (Yamashita et al., 2016).
The Heliantheae (for guayule) and the Cichorieae (for lettuce and dandelion) tribes are distantly related within the Asteraceae family as they share a common ancestor from ∼39 million years ago1. Considering such a divergence between guayule and lettuce/dandelion, along with guayule’s commercial potential to produce NR in arid regions, we conjecture that in-depth molecular studies of guayule CPTs and CBP can provide the knowledge necessary to help develop guayule as an alternative NR-producer. Guayule is transformable (Ponciano et al., 2018), and utilization of its biomass via bio-refinery process has been studied (Orts and McMahan, 2016). Isolated rubber particles from guayule have been used for biochemical studies (Cornish and Siler, 1995), but guayule CPT and CBP have not been investigated at the molecular level.
In this work, CPT and CBP homologs were identified from guayule, followed by the investigation of their biochemical activities, interactions, and expression patterns. In addition, the findings from the guayule CPT and CBP studies are discussed in the context of the current knowledge of NR biosynthesis.
Guayule plant seeds (PI478663) were obtained from the United States Department of Agriculture (USDA). Seeds were soaked in fresh water for 4 days before germination, and potted to a mixture of 35% autoclaved sands and 65% potting soil. The germinated plants were grown in a growth chamber under conditions of 16 h light, 8 h dark, at 25°C.
Two months old guayule plants were ground in liquid nitrogen using mortar and pestle, and 100 mg of powder was mixed with 1 mL of Trizol (Invitrogen), and total RNA was extracted according to the manufacturer’s protocol. First strand synthesis of cDNA was performed using 1 μg of the extracted RNA (leaf and stem) as template, combined with the M-MuLV reverse transcriptase (NEB) and anchored oligo dT22 (IDT). The synthesis of cDNA was performed according to NEB’s provided protocol. PaCPT1 was amplified from guayule cDNA by primers 1/2 and cloned into the pGEMT-easy vector (Promega) using TA/blunt ligase mix (NEB). PaCPT2/3 and PaCBP were synthesized after yeast codon optimization (SynPaCBP, MF688939; SynPaCPT2, MF688937; SynPaCPT3, MF688939). Phylogenetic and molecular evolutionary analyses were conducted using MEGA ver. 6 (Tamura et al., 2013). All sequences were retrieved from TAIR or NCBI database. The CPT sequences are from AtCPT1 (At2g23410), AtCPT2 (At2g23400), AtCPT3 (At2g17570), AtCPT4 (At5g60510), AtCPT5 (At5g60500), AtCPT6 (At5g58780), AtCPT7 (At5g58770), AtCPT8 (At5g58782), AtCPT9 (At5g58784), CeCPT (NP_001023351), EcCPT (WP_077899378), HbCPT1 (BAB92023), HbCPT2 (BAB83522), HsCPT (NP_995583), LsCPT1 (XP_023749459), LsCPT2 (XP_023743551), LsCPT3 (XP_023762616), PaCPT1 (ATD87115), PaCPT2 (ATD87118), PaCPT3 (ATD87116), RER2 (NP_009556), SaCPT (CEH25875), SlCPT1 (NP_001234633), SlCPT2 (AFW98426), SlCPT3 (AFW98427), SlCPT4 (AFW98428), SlCPT5 (AFW98429), SlCPT6 (AFW98430), SlCPT7 (AFW98431), SRT1 (NP_013819), TbCPT1 (AGE89403), TbCPT2 (AGE89404), TbCPT3 (AGE89405). The CBP sequences are from AtLew1 (At1g11755), CeCBP (NP_495928), HaCBP (OTG12218), HbCBP (XP_021659156), HtCBP (GHBN01046996), LsCBP1 (AIQ81186), LsCBP2 (AIQ81187), NogoBR (NP_612468), NUS1 (NP_010088), PaCBP (ATD87120), PtCBP (XP_002305421), SlCBP (NP_001333078), ToCBP1 (DY820019), ToCBP2 (DY832694), VvCBP (XP_003631635). The guayule sequences were deposited at NCBI: PaCBP (MF688938), PaCPT1 (MF688933), PaCPT2 (MF688936), PaCPT3 (MF688934).
Details of yeast complementation, in vitro enzyme assay, and reverse-phase thin layer chromatography were previously described (Kwon et al., 2016). All primer sequences are given in Supplementary Table S1. PaCPT1-3 were amplified using primers 3/4, 5/6 or 7/8, respectively, and the amplicons were cloned into p414-GPD using SpeI/SalI, SpeI/XhoI or BamHI/ClaI sites, respectively. PaCBP was cloned into p415-GPD using SpeI/XhoI sites. The cloned vectors were introduced to a yeast strain deficient in both copies of CPTs, rer2Δ::HIS3 srt1Δ::KanMX [pRS316-RER2], described in Kwon et al. (2016). This strain is referred to as rer2Δ srt1Δ hereafter, and Ro lab yeast stock number is ROY13. The constructed PaCBP/CPT pairs were transformed in this yeast strain. To serve as a positive control of complementation, p414-GPD ScRER2-TRP1 (previously cloned in Kwon et al., 2016) with empty p415-GPD was also transformed in the rer2Δ srt1Δ strain. The transformants were streaked on both SC-Leu-Trp-Ura-His plates (positive control) and SC-Leu-Trp-His plates containing 5FOA (5-fluoroorotic acid) and uracil to allow for counter-selection of pRS316 ScRER2-URA3. Growth on 5FOA/Ura-containing media indicates functional complementation of PaCBP/CPTs in place of yeast CPTs. Growth on both media was observed after a 7-day incubation period at 30°C.
The yeast strains, rescued from the 5FOA selection in the complementation assay, were cultured overnight and inoculated into 100 mL of SC-Leu-Trp-His media. The cells were harvested at late logarithmic phase by centrifugation at 3,000 × g for 3 min. The pelleted cells were then resuspended in 1 mL of TES-B buffer (0.6 M sorbitol, 1 mM EDTA, and 50 mM Tris-HCl, pH 7.4) and homogenized by bead-beating. The yeast lysate (1 mL) was centrifuged at 10,000 × g for 10 min to remove large cellular debris. The supernatant was diluted with 10 mL of TES-B and centrifuged again similarly. The final supernatant was centrifuged at 100,000 × g for 1 h, 4°C. The pellet was re-suspended in TG buffer [50 mM Tris-HCl and 20% (v/v) glycerol, pH 7.4] to a concentration appropriate for in vitro biochemical assays.
The in vitro biochemical assays of CPB/CPT were performed in 50 μL reaction mixtures containing 50 μg of prepared microsomal proteins, 20 μM FPP, 80 μM of IPP (14C-IPP PerkinElmer, 50–60 mCi mmol-1), 50 mM HEPES, 5 mM MgCl2, 2 mM NaF, 2 mM Na3VO4, and 2 mM DTT, pH 7.5. The reaction mixture was incubated at 30°C for 2 h. For each reaction, the extraction of polyisoprenoid products was performed in 1 mL of chloroform/methanol (2:1, v/v) and 0.4 mL of 0.9% NaCl, by vortexing for 20 s followed by centrifuged at 10,000 × g for 1 min at room temperature. The chloroform phase was transferred to a new tube and washed twice with 0.5 mL of methanol/water/chloroform mixture (48:47:3, v/v/v) by vortexing and centrifuging as above to remove unincorporated IPP. The lower chloroform phase was transferred to a glass vial and allowed to evaporate. Dephosphorylation of the product to polyprenol was carried out by incubating the dried product in 300 μL of 1 M HCl at 85°C for 1 h, followed by benzene extraction. The concentrated products in benzene were used for thin layer chromatography (TLC). The concentrated products were spotted on a C18 reverse-phase silica plate (Analtech cat. P90011, 20 × 20 cm) and separated by acetone/water (39:1, v/v) mobile phase. The TLC plate was then exposed to a BAS Storage Phosphor Screen (GE Healthcare) and the screen was scanned by a phosphorimager (Bio-Rad, Molecular Imager FX). The Rf value of known RER2 products (Brasher et al., 2015) was confirmed in our TLC system, and the Rf values were used to determine the carbon numbers.
MYTH assays were performed according to the published methods (Gisler et al., 2008; Snider et al., 2010). The bait plasmid (PaCBP-Cub-LexA-VP16) was generated by ligating the PaCBP amplified with primers 9/10 into pAMBVα using StuI/NcoI sites. For the prey plasmids, PaCPT1-3 were amplified with primers 11/12, 13/14 or 15/16, and cloned into pPR3C-NubG in SpeI/BamHI (PaCPT1) or BamHI/ClaI (PaCPT2 and 3) to yield PaCPT-HA-NubG constructs. PaCPT1-3 were also amplified with primers 17/18, 19/20 or 21/22 and cloned into pPR3N-NubG in BamHI/SalI (PaCPT1) or BamHI/ClaI (PaCPT2 and 3) to generate NubG-HA-PaCPT constructs. The bait and prey plasmid pair was co-transformed into the Y2H reporter yeast strain, THY.AP4. The control prey vectors pOST1-NubG and pOST1-NubI were also co-transformed with the bait vector. Individual transformed-yeast colonies were cultured in 30°C until OD600 reached to 3.0–3.5. The cell densities were then normalized to 0.05 (OD600 value) by dilution. The normalized cells were diluted to produce dilution of 1/10 and 1/100. Ten μL of cells was spotted on SC-Leu-Trp (SC-LT) and on SC-Leu-Trp-Ade-His (SC-LTAH) containing 3-amino-1,2,4-triazole (3-AT) at concentrations of 5, 10, or 25 mM. Spotted plates were incubated for 3 days at 30°C to observe cell growth.
The strength of protein interactions was quantified using β-galactosidase assay with O-nitrophenyl-β-D-galactoside (ONPG) as the substrate. From SC-LTAH selection plates, the positive colonies were cultured overnight in SC-LTAH medium without 3-AT. Cells from the overnight culture were inoculated to 4 mL of SC-LTAH medium and cultured for 5 h. The cells were harvested at 9,000 × g for 3 min at 4°C and resuspended in 1 mL of Z buffer (60 mM Na2HPO4, 40 mM NaH2PO4, 10 mM KCl, 1 mM MgSO4, and 50 mM β-mercaptoethanol, pH 7.0), 20 μL of chloroform and 20 μL of 0.1% SDS. After vortexing, the mixture was pre-incubated for 5 min at 28°C. The reaction was initiated by adding 0.2 mL of ONPG (4 mg mL-1 in Z buffer) and terminated by the addition of 1 M of Na2CO3. The mixture was centrifuged at 20,000 × g for 10 min, and the absorbance at 420 nm and 550 nm were measured to determine the Miller units (Miller, 1972).
DNA constructs for in vitro translation were cloned into pT7CFE1-3xFLAG (constructed using pT7CFE1-NHa) for PaCPT1-3 and pT7CFE1-NHa (Thermo Fisher Scientific) for PaCBP. pT7CFE1-3xFLAG was generated by replacing HA tag in pT7CFE1-NHa with 3xFLAG. The 3xFLAG (atggactacaaagaccatgacggtgattataaagatcatgatatcgattacaaggatgacgatgacaag) was amplified using primer sets 23/24, 23/25, and 23/26 sequentially using the IRES region in pT7CFE1-NHa as a template. The final PCR product containing partial IRES sequence and 3xFLAG was digested by KpnI and BamHI and ligated to corresponding restriction sites in pT7CFE1-NHa. Synthetic PaCPT1 and PaCPT2 (GenScript, Piscataway, NJ, United States) were ligated to BamHI and XhoI sites in pT7CFE1-3xFLAG. PaCPT3 was amplified from P. argentatum leaf cDNA using a primer set, 27/28. The PaCPT3 PCR product was cloned into pT7CFE1-3xFLAG via BamHI and XhoI restriction sites. Synthetic PaCBP (GenScript, Piscataway, NJ, United States) was amplified using a primer set 29/30 and digested with BamHI and XhoI, followed by ligation to corresponding sites in pT7CFE1-NHa. A control construct, pT7CFE1-NHa-GFP for co-immunoprecipitation described below, was generated by cloning GFP sequence into pT7CFE1-NHa. GFP was amplified from pCFE-GFP (Thermo Fisher Scientific) using a primer set 31/32, containing BamHI and XhoI, respectively, followed by ligation to corresponding sites in pT7CFE1-NHa. The purified plasmid preparations of all IVT constructs were cleaned up further by ethanol precipitation, followed by resuspension in nuclease-free water to a concentration of 500 ng μL-1 prior to IVT. IVT was carried out using 1-Step Human Coupled IVT Kit (Thermo Fisher Scientific), following the manufacturer’s procedure except for reducing the reaction to half of the recommended amounts. Immediately following incubation of the IVT reaction mixture at 30°C for 4 h, 1 μL of the 12.5 μL IVT mixture was saved as input for Western blotting by adding SDS-PAGE loading buffer and storage in -20°C until use. The remaining IVT mixture was used immediately for immunoprecipitation as described below.
For pre-binding of protein G magnetic beads (NEB, ON, Canada) to monoclonal mouse anti-FLAG M2 antibody (Sigma, ON, Canada) or monoclonal rat anti-HA 3F10 antibody (Roche, Laval, QC, Canada), for each co-immunoprecipitation reaction, 10 μL of the bead suspension was washed (invert five times/vortex 5 s) in 100 μL of room temperature PBS buffer (137 mM NaCl, 2.7 mM KCl, 10 mM Na2HPO4, 1.8 mM KH2PO4, pH 7.4) four times and incubated with 0.5 μL of the antibody in 100 μL of cold PBST (PBS, pH 7.4, 0.1% Tween 20) at 4°C for 16 h on a LabquakeTM Tube Rotator. The separation of magnetic beads from solution was achieved by applying magnetic field for 30 s on a magnetic stand. To remove unbound antibody, the antibody-immobilized beads were washed (invert five times/vortex 5 s) three times, each in 100 μL of cold IP buffer (150 mM NaCl, 100 mM Tris-HCl pH 7.4, 1 mM EDTA, 10% glycerol, 1% Triton X-100, 0.5% NP-40, 1 mM PMSF). The immobilized beads were resuspended in 500 μL of cold IP buffer, followed by addition of the 11.5 μL of the completed IVT reaction mixture. Co-immunoprecipitation reaction was incubated at 4°C for 1 h on the rotator. To remove unbound proteins, the mixture was washed (invert five times/vortex 5 s) five times, each in 700 μL of cold IP buffer. Immunoprecipitated proteins were eluted from the beads by resuspending in 20 μL of 2X SDS-PAGE loading buffer [100 mM Tris-HCl pH 6.8, 4% (w/v) SDS, 20% glycerol, 0.01% (w/v) bromophenol blue, 200 mM β-mercaptoethanol] and incubating at 95°C for 5 min with intermittent gentle flicks. This elution also liberates anti-FLAG and anti-HA antibodies from the magnetic beads as the heavy (50 kDa) and light (25 kDa) chains of the IgG antibodies. After bringing the eluted mixture to room temperature, the mixture was spun down by a brief centrifugation, followed by transfer of the eluate while on a magnetic stand. Half of the eluate was used to run 12% SDS-PAGE followed by Western blotting with mouse anti-FLAG M2 antibody and the other half with rat anti-HA 3F10 antibody. The IVT reaction mixture (1 μL of 12.5 μL total IVT reaction) saved as input was also divided in half to run in SDS-PAGE and Western blotting with anti-FLAG and anti-HA antibodies. SDS-PAGE gel was transferred to PVDF membrane using a Tris-glycine-methanol buffer system, and the antibody hybridization was performed in 5% skim milk TBST (5% skim milk, 50 mM Tris-HCl pH 7.5, 150 mM NaCl, 0.05% Tween 20) at 1:2000 for both anti-FLAG and anti-HA antibodies by overnight incubation at 4°C. HRP-conjugated anti-mouse and anti-rat secondary antibodies were also hybridized in 5% skim milk TBST at 1:10,000 at room temperature for 1 h.
For de novo transcriptome assembly and differential gene expression analysis, 4-month-old greenhouse-grown guayule plants (AZ-2 cultivar) were transferred to a simulated summer condition in a growth chamber (16 h daylight, 25°C daytime and 15°C night time temperatures). At 6 months of age, plants were transferred to a simulated winter condition (13 h daylight, 25°C daytime and 5°C nighttime temperatures) for induction of rubber production. Plants were harvested for analysis at 8 months of age. RNA was extracted from three biological replicates each for leaf and stem tissues from induced and control plants using Trizol (Invitrogen) following the manufacturer’s instructions. Following Trizol extraction, RNA samples were treated with DNAase using the Turbo DNA-free kit (Ambion) and further purified using RNeasy column purification (Qiagen). Directional, bar-coded Illumina sequencing libraries were prepared using NEBNext® UltraTM Directional RNA Library Prep Kit (NEB) with the suggestions for size selection of an average insert size of 300–450 bp. Libraries were then pooled and sequenced on Illumina Miseq and HiSeq2500 platforms producing 28 million 300 bp paired end reads and 155 million 150 bp paired end reads, respectively, with an average of 2.4 million MiSeq and 12.9 million HiSeq reads per sample. RNA-seq data was quality-controlled using Trim Galore (Babraham Bioinformatics group) to eliminate adapter sequence contamination and to trim data with a quality score below Q30. Miseq and Hiseq reads were pooled for transcriptome assembly and differential gene expression analysis. The transcriptome was assembled using Trinity assembler version 2.04 set to minimum Kmer coverage of 2 (Haas et al., 2013). The initial transcriptome assembly was filtered using a utility included within the Trinity package to eliminate transcript isoforms with low abundance and low coverage. Differential gene expression analysis was performed using EdgeR (Robinson et al., 2010). The assembly data are available in the NCBI short read archive under submission number SRP107961.
Commercially cultivated guayule (Parthenium argentatum) plants for NR production are polyploids, making the analysis of gene family difficult due to gene redundancy. However, the guayule transcriptome assembly from the diploid accession PI478640 is accessible in the public NCBI database (Hodgins et al., 2014), and we used the data set from diploid guayule in this work. In this transcriptome data, 51,947 unigenes were assembled from 983,076 reads (454 GS FLX Titanium) from four different guayule tissues (root, leaf, flower, and stem). We screened this assembled transcriptome using lettuce CPT and CPT-Like (CPTL) in our previous study in lettuce (Qu et al., 2015). CPTL was renamed as CBP (CPT-Binding Protein) in the previous work (Kwon et al., 2016) and will be referred to as CBP in this work. From this analysis, one guayule CBP and three CPT cDNAs were identified, and accordingly these cDNAs were named as PaCBP and PaCPT1-3. Unlike lettuce that has two CBP isoforms, one of which displays a laticifer-specific gene expression, guayule does not have laticifer cells and has only a single copy of CBP.
To understand phylogenetic relatedness of PaCPT1-3 to other known eukaryotic and prokaryotic CPTs, a phylogenetic tree was reconstructed using the deduced amino acid sequences from PaCPT1-3 cDNAs. It has been established that two clades of CPTs (clade I and clade II) are present in plants (Figure 1A). Clade I is comprised of chloroplastic CPTs that likely originated from prokaryotic homo-dimeric CPTs, whereas clade II consists of cytosolic/ER CPTs that interact with CBPs to form hetero-protein complexes, which are then recruited to ER. The CPT clade II is further divided into two sub-clades – one type of CPT for dolichol biosynthesis in the primary metabolism and the other type for NR biosynthesis in the specialized secondary metabolism.
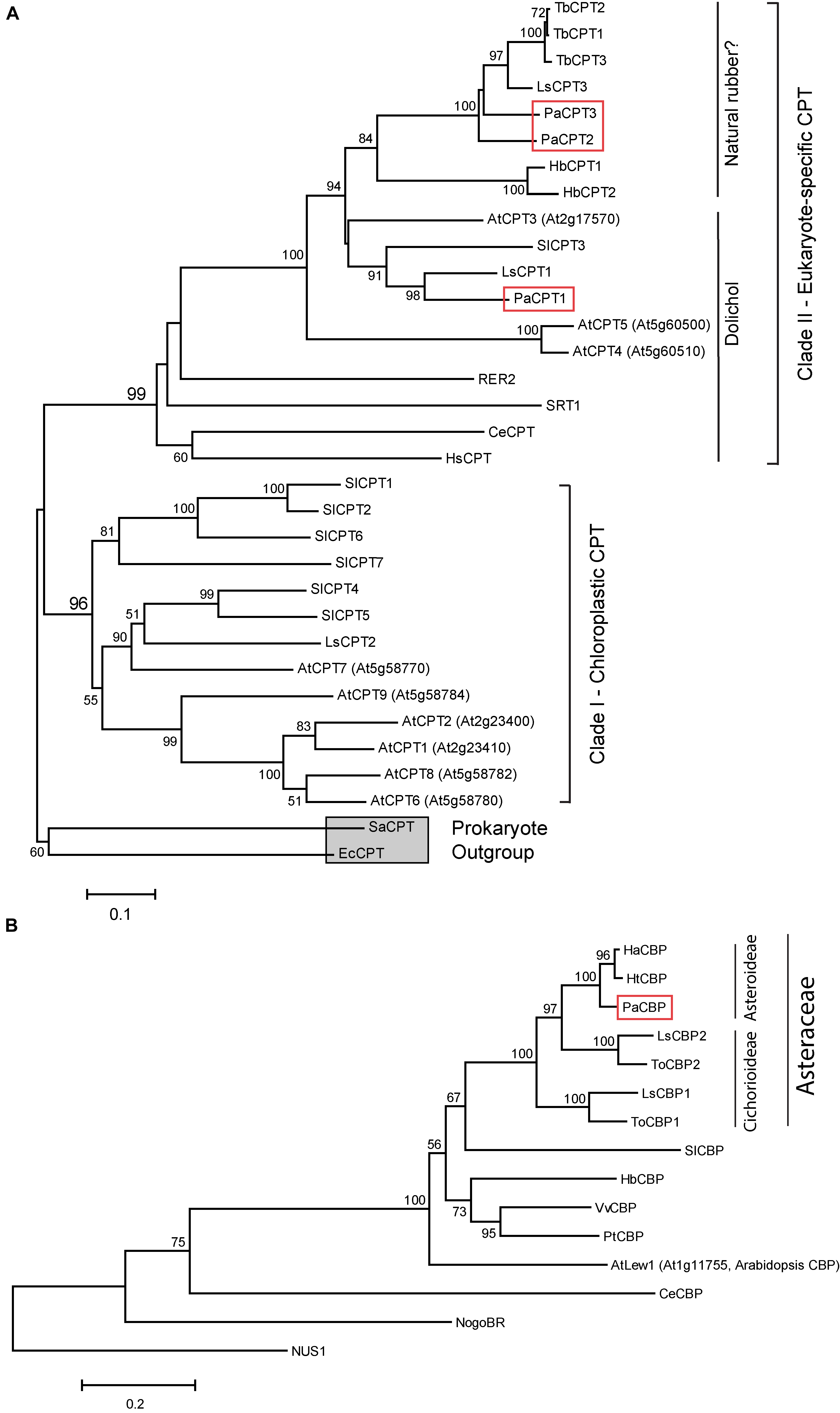
Figure 1. Phylogenetic analysis of CPT (A) and CBP proteins (B). Phylogenetic trees were created based on the protein sequence similarities between CPTs and CBPs from various prokaryotic and eukaryotic species. Bootstrap values from 1,000 replicates were calculated, and the percentages of replicates are shown in each node only when higher than 50%. Two prokaryotic CPTs (EcCPT and SaCPT) are used as the outgroup and indicated by a gray square. PaCPT1-3 and PaCBP are marked by red boxes. Abbreviations used are: At, Arabidopsis thaliana; Ce, Caenorhabditis elegans; Ec, Escherichia coli; Ha, Helianthus annuus; Hb, Hevea brasiliensis; Hs, Homo sapiens; Ht, Helianthus tuberosus; Ls, Lactuca sativa; Pa, Parthenium argentatum; Pt, Populus tremuloides; Sa, Staphylococcus aureus; Sl, Solanum lycopersicum; Tb, Taraxacum brevicorniculatum; To, Taraxacum officinale; Vv, Vitis vinifera; RER2 and SRT1 are yeast CPTs. NUS1 is yeast homolog of CBP. Accession numbers of the sequences used to construct the phylogeny are given in “Materials and Methods.” Arabidopsis CPTs (AtCPT1-9) and tomato CPTs (SlCPT1-7) were numbered according to the published articles (Surmacz and Swiezewska, 2011; Akhtar et al., 2013). Note that Kera et al. (2012) assigned different numbering of the nine Arabidopsis CPTs, and different nomenclature of AtCPTs can be listed in other publications.
In the phylogenetic analysis, all three PaCPTs belong to the cytosolic/ER clade II CPT. At the sub-clade level, with strong statistical supports, PaCPT1 is clustered with LsCPT1 (lettuce), SlCPT3 (tomato), and AtCPT1 (Arabidopsis), all of which are known to synthesize dehydrodolichyl diphosphate (Brasher et al., 2015; Qu et al., 2015; Kwon et al., 2016). On the other hand, PaCPT2 and PaCPT3 form a close cluster with LsCPT3 and TbCPT1-3 (dandelion), known to have implications in NR biosynthesis (Post et al., 2012; Qu et al., 2015). This phylogenetic tree suggested that guayule possesses two sub-clades of diverged CPTs within the CPT clade II. It can be inferred from these results that PaCPT1 is likely to be involved in the dolichol metabolism, while PaCPT2 and PaCPT3 are involved in NR metabolism.
The phylogenetic tree for CBPs was also constructed. CBP is exclusively found in eukaryotes, and thus the phylogenetic structure of CBP shows a monophyletic group (Figure 1B). The duplications of CBP (i.e., two CBP isoforms) are restricted to lettuce and dandelion in the Cichorieae tribe (within Cichorioideae sub-family under Asteraceae family), and other plants with high quality genome sequences encode a single copy of CBP. Interestingly, PaCBP and other single copy CBPs from the Asteroideae sub-family (sunflower HaCBP and Jerusalem artichoke HtCBP) are tightly clustered with the laticifer-specific CBPs (lettuce LsCBP2 and common dandelion ToCBP2), even though there is no developed laticifer in sunflower and artichoke. Thus, the CBP gene duplication event likely occurred prior to the divergence of the two sub-families and the advent of laticifer in Cichorioideae subfamily.
In eukaryotes, dolichol is an indispensable carrier molecule of sugar moieties for post-translational protein modifications. Therefore, all eukaryotes have at least one set of CPT and CBP to synthesize dehydrodolichyl diphosphate (i.e., dolichol precursor), and the absence of either CPT or CBP is lethal. Yeast (Saccharomyces cerevisiae) has one homolog of CBP (NUS1) and two homologs of CPTs (RER2 and SRT1). We previously developed a yeast strain that has double deletions in RER2 and SRT1, in which its lethality is rescued by expressing a plasmid copy of RER2 (Kwon et al., 2016). To examine the ability of PaCPT and/or PaCBP to rescue the dolichol-deficiency of rer2Δ srt1Δ, these guayule genes were first transformed to this strain individually or in PaCPT/PaCBP pairs, and then the URA-selective plasmid containing RER2 was removed by 5-fluoroorotic acid (5FOA) selection (Figure 2). This enabled us to assess the PaCPT activity in the absence of both of the yeast CPTs.

Figure 2. Complementation of rer2Δ srt1Δ yeast by PaCPT1-3 and PaCBP. The yeast strain, rer2Δ srt1Δ is lethal but is maintained by expressing RER2 in URA-selectable plasmid. This strain was used to transform plasmids expressing each PaCPT1-3 and PaCBP. The successful transformants were streaked on 5FOA selection plates to remove RER2 containing URA-plasmid. Yeast growth in 5FOA selection was observed only by PaCPT/PaCBP pairs or by retransformed RER2 in TRP-plasmid. No growth was observed when PaCPT alone or PaCBP alone was expressed.
In the absence of CPT enzymatic activity, the rer2Δ srt1Δ lethal phenotype will not be rescued. In all cases tested, the rer2Δ srt1Δ strain transformed with empty plasmids or a single plasmid that contains either PaCBP or one of the PaCPTs failed to grow upon removal of the RER2-URA plasmid by 5FOA selection. In contrast, co-expression of PaCBP and one of the three PaCPTs could effectively complement the lethality of the yeast CPT-knockouts upon removal of the RER2-URA plasmid. Our results indicate that each PaCPT can produce isoprenoid molecules analogous to dehydrodolichyl diphosphate only in the presence of PaCBP but not together with endogenous yeast CBP, Nus1.
The surviving yeast strains after 5FOA selection should be able to bring about necessary CPT activities for the yeast survival in the rer2Δsrt1Δ yeast. To assess the guayule CPT activities, microsomes were prepared from the rescued yeast strains from the complementation assay, and radiolabeled 14C-isopentenyl diphosphate (14C-IPP) and the priming molecule, FPP, were co-incubated with the isolated microsomes. When the incorporations of 14C-IPP to hydrophobic polymers were measured, significant IPP-incorporation activities ranging from 14.4 to 50.8% incorporation were measured in 2-h assays (Table 1). Their specific activities were comparable to (in the cases of PaCPT1/PaCBP1or PaCPT3/PaCBP activity) or exceeded (in the case of PaCPT2/PaCBP activity) the CPT activities previously reported from lettuce CPT and CBP (Qu et al., 2015). The CPT activity of the microsomes from the rescued yeast strain transformed with RER2 as a complementation control was substantially lower (4.5- to 15.6-fold) than those from guayule PaCPT/PaCBP co-expression. The lower CPT activity of RER2 control strain is attributable to the imbalance between overexpressed RER2 and its binding partner NUS1, expressed at a native level in yeast.

Table 1. Specific activities of PaCPTs and PaCBP in isolated microsomes.
The resulting 14C-labeled products were separated by the reverse-phase thin layer chromatography (RP-TLC) optimized for the separation of dolichol-sized polymers. In the RP-TLC analysis, we detected no obvious differences among the radiolabeled isoprene polymers produced from any of the PaCPT and PaCBP pairs (Figure 3A), and their polymer size was slightly smaller than, but still comparable to, the dehydrodolichols produced by RER2 (Figure 3B). We could not find evidence that the cis-isoprene polymer, significantly longer than dolichol, is synthesized by PaCPT and PaCBP in yeast microsomes.

Figure 3. Isoprene product separations by reverse phase thin layer chromatography. Microsomes were prepared from the yeast strains selected on 5FOA in Figure 2, and in vitro enzyme assays were performed using the microsomes and 14C-isopentenyl diphosphate, followed by TLC (C18 reverse-phase silica) of extracted cis-polyisoprene products using a solvent mixed with acetone and water (39:1), and phosphorimager analysis. (A) The cis-polyisoprene product profiles from yeast RER2 as a size calibration standard. (B) The cis-polyisoprene products from PaCPT/PaCBPs are identical to one another and slightly shorter than those from the RER2-containing microsomes. S indicates the starting point of separation and SF indicates solvent front. Rf values and carbon numbers are shown.
The complementation of rer2Δ srt1Δ yeast strain by co-expression of PaCPT and PaCBP, but not by a single PaCPT or PaCBP expression, implied that both proteins are required for dolichol metabolism. Thus, possible interactions between each PaCPT and PaCBP were examined by split-ubiquitin yeast 2-hybrid assay developed for protein interactions of membrane-bound proteins (Stagljar et al., 1998). In this assay, the restoration of ubiquitin by N-terminal ubiquitin (Nub) and C-terminal ubiquitin (Cub) allows a release of transcriptional regulator to nucleus to activate the auxotrophic marker genes and the β-galactosidase reporter gene. A single residue mutation (I to G) of natural Nub protein (i.e., NubI to NubG) prevents the auto-assembly of the ubiquitin, unless NubG and Cub binding is mediated by the interaction of the proteins of interest each fused to NubG and Cub. Therefore, interactions between PaCPT and PaCBP proteins translationally fused to Cub and NubG can be assessed by yeast growth on selective medium and β-galactosidase activity.
As a control, co-expression of PaCBP-Cub fusion and OST1 (α-subunit of the oligosaccharyltransferase complex localized on the ER)-NubI fusion allows restoration of ubiquitin, thereby presenting efficient yeast growth and strong β-galactosidase activity (Figure 4A). In contrast, changing NubI to NubG in the same experiment did not permit yeast growth, suggesting PaCBP and OST1 do not interact with each other and cannot bring the two ubiquitin subunits together as expected as a negative control (Figure 4A). Similarly, PaCPTs 1–3 were individually fused to NubG, and each fusion construct was co-expressed with PaCBP-Cub to assess whether PaCPT-PaCBP interaction occurs to restore ubiquitin. The interaction between each of the PaCPTs and PaCBP could be clearly inferred by the efficient yeast growth and the by β-galactosidase activities. Interestingly, the interactions were only observed in one orientation of the PaCPT fusion protein (Figure 4B). The NubG-PaCPT orientation allowed activations of the growth markers and reporter gene, whereas the PaCPT-NubG orientation did not show evidence of protein–protein interactions. Activations of marker genes only when NubG is located in the N-terminus of prey protein have been previously reported (Yao et al., 2017) and have occasionally happened in split-ubiquitin yeast 2-hybrid assays (personal communication with Igor Stagljar, University of Toronto). Wherever interactions were observed, the HIS3 marker gene was activated very efficiently as yeast could grow at up to 25 mM of AT (3-amino-1,2,4-triazole), a competitive inhibitor of HIS3. In addition, β-galactosidase activities were comparable to that from the positive control. Therefore, the split-ubiquitin Y2H provided molecular evidence that each PaCPT and PaCBP interact with each other to form a hetero-protein complex in yeast.
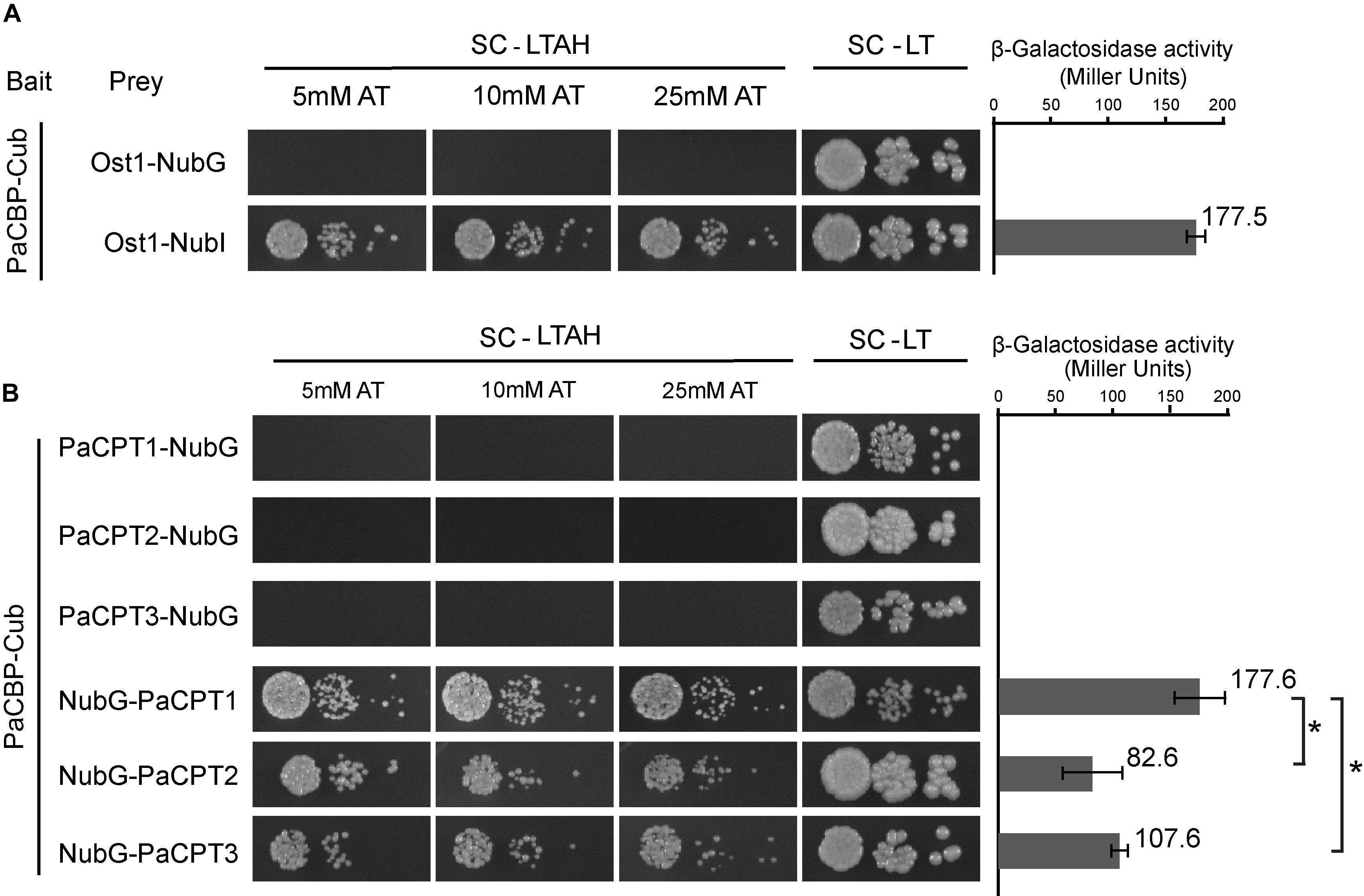
Figure 4. Split ubiquitin yeast 2-hybrid assays to assess the protein interactions between PaCPT1-3 and PaCBP. The bait protein, Cub, was fused at the C-terminus of PaCBP in all assays. Under the selective growth condition, SC-LTAH, yeast growth was observed only when the bait and prey proteins interacted either through NubI-Cub auto-assembly (A: Ost1-NubI, positive control) or through the PaCPT/PaCBP interaction-mediated NubG-Cub assembly (B: NubG-PaCPT3, NubG-PaCPT2, NubG-PaCPT3). PaCPT/PaCBP interactions occurred only when NubG was fused on the N-terminus of PaCPTs. Under the non-selective growth condition, SC-LT, all yeast strains grew regardless of the protein interaction. As an important negative control, NubG-fused Ost1 protein did not interact with Cub-fused PaCBP (A), demonstrating a specific interaction of PaCBP to PaCPT, but not to Ost1. Yeast growth was observed in a wide range of 5–25 mM AT, indicating a strong activation of the HIS3 reporter gene by the ubiquitin-activated transcription factor. The β-galactosidase activity was measured in the corresponding yeast strains that showed protein interactions in the absence of AT. PaCPT1/PaCBP interaction showed the same level of β-galactosidase activity as the Ost1-NubI control while PaCPT2/PaCBP and PaCT3/PaCBP showed lower activities. Ten μL of OD600 yeast culture was placed in the first spot, and yeast cells with 10- and 100-times dilution were placed in the subsequent spots. Asterisks indicate statistically significant differences (p-value < 0.05).
In addition to the split-ubiquitin Y2H approach, PaCPT1-3 and PaCBP interaction were independently examined by co-immunoprecipitation assays (Co-IP). PaCPT1-3 and PaCBP were tagged by FLAG- and HA-epitope, respectively, and their proteins were prepared by in vitro transcription and translation. Green fluorescent protein (GFP), which has a similar size to PaCBP and the PaCPTs, was used as a negative control in Co-IP experiments. All in vitro co-transcribed and co-translated PaCPT/PaCBP proteins were detectable in input by immunoblotting with either FLAG or HA antibody (Figures 5A,B input; Supplementary Figure 1). Incubation of FLAG-antibody immobilized magnetic beads resulted in the co-IP of FLAG-PaCPTs with HA-PaCBP as detected by FLAG-antibody and HA-antibody immunoblotting, respectively, while HA-GFP control protein could not be pulled down by FLAG-PaCPT2 (Figure 5A and Supplementary Figure 1). Similarly, immunoprecipitation with HA-antibody immobilized magnetic beads resulted in the co-IP of HA-PaCBP with FLAG-PaCPTs, while FLAG-PaCPT2 control protein could not be pulled down by HA-GFP (Figure 5B). These results showed that each of the PaCPT1-3 and PaCBP interact with each other to organize hetero-protein complexes when co-expressed in vitro.
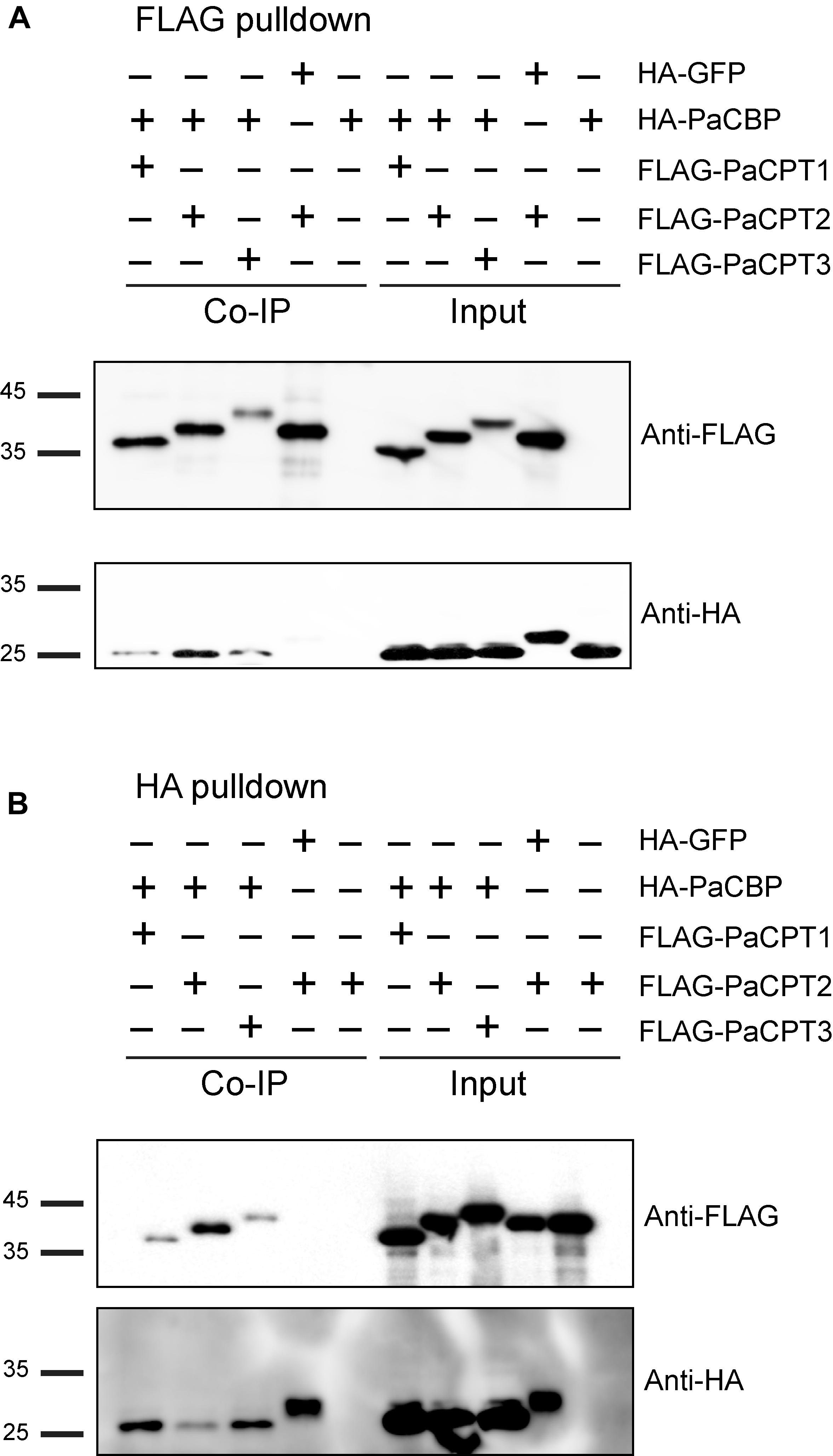
Figure 5. Co-immunoprecipitation of recombinant PaCPT/PaCBP proteins. (A) HA-PaCBP could be co-immunoprecipitated by FLAG-PaCPT1-3, and (B) FLAG-PaCPT1-3 could be co-immunoprecipitated by HA-PaCBP. PaCPT1-3 and PaCBP proteins tagged with FLAG- and HA-epitopes, respectively, and were prepared by in vitro transcription and translation. GFP tagged with an HA-epitope was used as a negative control. The information of the protein mixture is detailed on the top panel with the appropriate epitope-tag. The antibodies used in the immunoblot are indicated beside blots (FLAG, FLAG antibody or HA, HA antibody).
Cold temperature is known to induce NR biosynthesis in guayule (Madhavan et al., 1989). Based on the positive effect of cold-stress on rubber synthesis, Illumina sequencing of guayule stem and leaf tissues after cold-stress was recently performed, and the sequencing data are publicly accessible at NCBI (short read archive number: SRP107961). Using these data, we conducted a RNA-seq analysis to examine the transcript abundance of PaCPT1-3 and PaCBP (Figure 6).
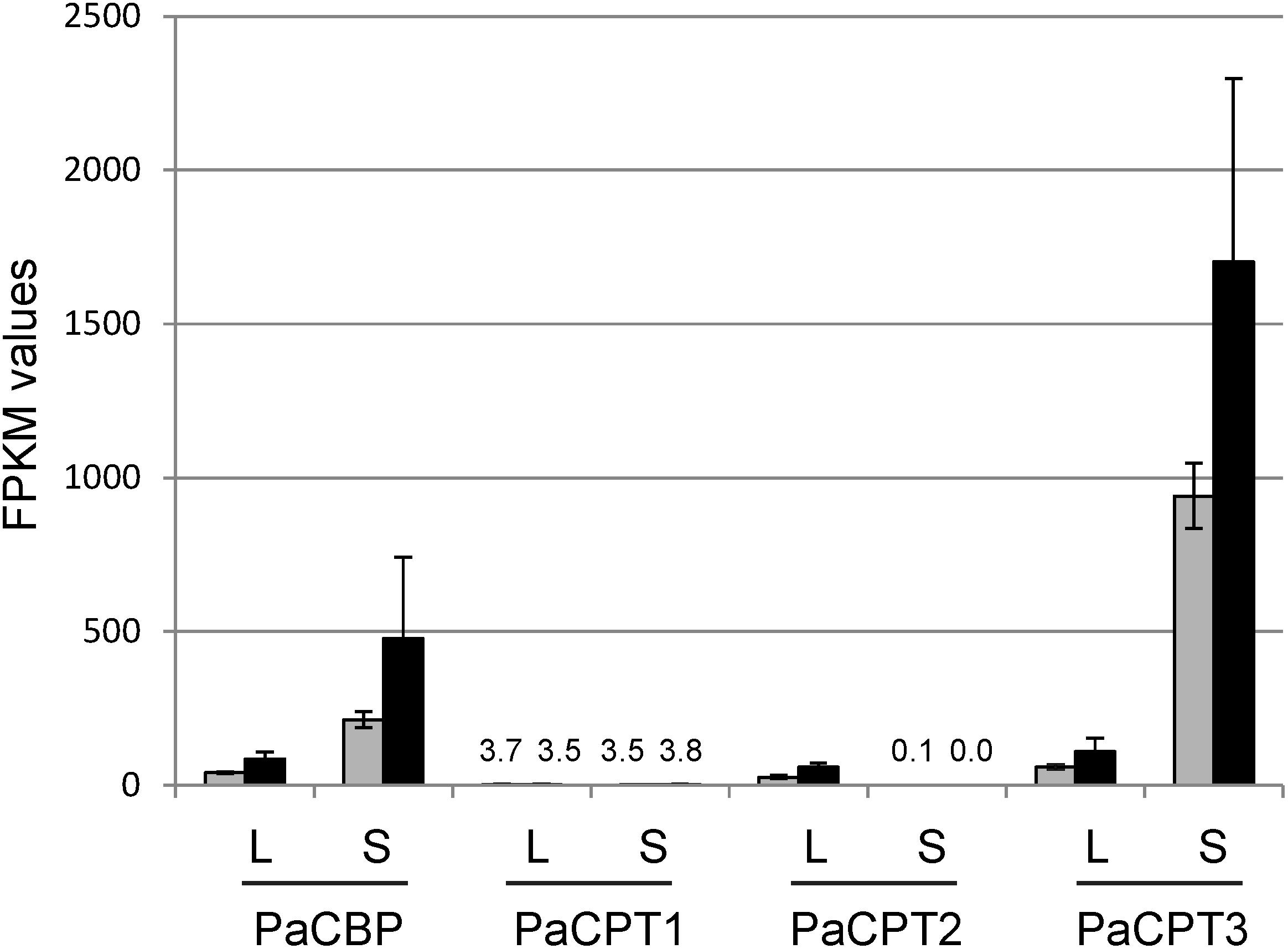
Figure 6. RNA-seq analysis of PaCBP and PaCPT1-3. FPKM (fragments per kilobase million) values for PaCBP and PaCPT1-3 were calculated from the publicly available data (short read archive number: SRP107961). Gray bars, control; Black bars, cold treatment. L, leaf sample; S, stem sample. Data are means ± SD (n = 3).
PaCPT1 transcripts were maintained at a relatively constant basal level in leaves and stems under both normal and cold-treated conditions. These data suggest the basal metabolic role of PaCPT1 in dolichol metabolism and are congruent to the phylogenetic analysis, in which PaCPT1 is clustered with other CPTs involved in the dolichol metabolism. PaCPT2 transcripts were hardly detectable in stem tissues while detectable at moderate levels in leaves. Of the three PaCPTs examined, PaCPT3 showed the highest level of expression in stem (2- to 3-orders of magnitude higher levels than PaCPT1/2), and its transcript abundance increased 1.8-fold by cold-treatment in stem. As NR is primarily synthesized in the parenchyma cells of the guayule stem (Kajiura et al., 2018), these data suggest that PaCPT3 is the primary CPT that contributes to the NR biosynthesis in guayule stem. On the other hand, transcript levels of PaCBP showed a lower degree of fluctuation, compared to PaCPT1-3 gene family. Its expression levels closely matched those of PaCPT2 and PaCPT3 in leaves. Noticeably, the PaCBP transcript abundance is highest in stem and is increased by 2.2-fold by cold-treatment, precisely mirroring the expression pattern of PaCPT3 in stem.
Average values of PaCBP and PaCPT3 expression increased by cold treatment between 1.8- and 2.2-fold in leaves and stem, but only PaCBP1 induction in leave was statistically significant (p-value < 0.05) in the triplicate Illumina transcript data. Systematic statistical analyses further showed that expression levels of PaCPT3 in comparison to PaCPT1 and PaCPT2 are significant in stem (p-value < 0.01) and leaves (p-value < 0.05). Preferential expression of PaCPT3 in stem in comparison to leaves was also statistically significant (p-value < 0.05).
Natural rubber is an irreplaceable biomaterial for hundreds of medical and industrial products, but its supply depends entirely on the rubber tree. In nature, more than 7,000 plants are known to produce NR (Rivano et al., 2013), but the quality of NR in most plants do not meet the NR polymer properties required for industrial uses. The exceptions are Russian dandelion (Taraxacum kok-saghyz), lettuce (Lactuca sativa), and guayule (Parthenium argentatum) as they can synthesize ∼1 million average Mw NR. Biochemical and reverse genetic data for NR biosynthesis in lettuce and dandelion have been reported (Post et al., 2012; Epping et al., 2015; Qu et al., 2015); however, little is known about NR biosynthesis in guayule at the level of individual cDNA and enzyme, except for the activity data from the isolated guayule rubber particles (Cornish and Siler, 1995). In this work, we identified and characterized three PaCPT isoforms and one PaCBP. While the results from this work confirmed some known facts, such as the formation of CPT/CBP protein complex and the lack of NR biosynthesis by CPT/CBP in vitro, they also provided additional new insights of NR biosynthesis in guayule.
Although the whole genome sequence has not been completed in guayule, we identified one PaCBP from 983,076 reads. Considering the read depth, it is unlikely that any expressed, sufficiently diverged PaCBP isoform is present in major tissues of guayule. Thus, PaCBP appears to be encoded as a single copy gene in guayule. Similarly, a single copy of CBP has been identified from the whole genome sequences and transcriptomics of the rubber tree, Hevea brasiliensis (Tang et al., 2016). Outside of NR-producing plants, the sequence analyses of high quality plant genomes (e.g., Arabidopsis, tomato, poplar, grapevine, and sunflower) also indicate that a single copy of CBP is present in these species. On the contrary, lettuce and dandelion, both originating from the same phylogenetic lineage to the tribe level (Cichorieae tribe), have two isoforms of CBP – one being general for dehydrodolichyl diphosphate biosynthesis in all cells and the second isoform being specific for NR biosynthesis in the laticifer. This CBP gene duplication appears to be restricted to the laticiferous plants from the Cichorieae tribe but is not common in other plants, even including NR-producing plants. Taken together, it is apparent that neo-functionalization and recruitment of a specialized CBP isoform are not required for quality NR biosynthesis because regardless of the presence of two isoforms of CBP, guayule and rubber tree can all synthesize high quality NR. It can be further inferred that a single CBP in the rubber tree and guayule should simultaneously support both primary dolichol and secondary NR metabolism. In such a scenario, we envision a challenge in finding the in planta evidence (i.e., RNAi or mutant) showing the necessity of CBP for NR biosynthesis in rubber tree and guayule, as severe knock-down by silencing or knock-out of CBP will be lethal in these plants.
Among the three PaCPTs in guayule, a few lines of evidence support that PaCPT3 is the key isoform involved in guayule NR biosynthesis. (1) Phylogenetically, PaCPT2 and PaCPT3 closely clustered with lettuce LsCPT3 and dandelion TbCPT1-3, whose expressions are specific to laticifer (Post et al., 2012; Qu et al., 2015), and RNA interference of TbCPT1-3 resulted in significant NR reduction (Post et al., 2012). (2) In our RNA-Seq analysis, the PaCPT3 transcript level was three orders of magnitude higher than that of PaCPT1 in stem where NR is known to accumulate. (3) The expression patterns of PaCPT3 closely coincide with PaCBP in two tissues under normal or the induced (cold-stress) condition. Despite clustering closely with LsCPT3, PaCPT2 had a negligible level of expression in stem. Similarly, PaCPT1 showed a basal level of expression and clustered phylogenetically with other CPTs implicated in dehydrodolichyl diphosphate biosynthesis. Collectively, these data indicate that PaCPT3 supports NR biosynthesis in guayule. To further substantiate this, our future efforts will be focused on silencing or overexpression of PaCPT3 to observe NR reduction or increase in guayule, respectively.
In addition to identifying the candidate genes necessary for NR biosynthesis in guayule, a suite of molecular and biochemical experiments (i.e., complementation in rer2Δ srt1Δ yeast, in vitro enzyme assays, Y2H, and Co-IP) reliably demonstrated that each of the PaCPT1-3 and PaCBP form hetero-protein complexes to synthesize dehydrodolichyl diphosphate. Despite the formation of protein complexes, we were not able to find biochemical evidence that cis-polyisoprenes, longer than dehydrodolichol (dephosphorylated form of dehydrodolichyl diphosphate), can be synthesized using PaCPT3 and PaCBP – the prime candidates for NR biosynthesis in guayule. The polyisoprene products synthesized in vitro by recombinant PaCPT3/PaCBP were essentially identical to those from PaCPT1/PaCBP or PaCPT2/PaCBP (Figure 3A). Plants may have more favorable environments (e.g., ER-associated protein, lipid compositions, and post-translational modification) than yeast for NR production by PaCBP and PaCPTs, and it will be worth expressing PaCBP and PaCPTs in plant and testing their enzyme activities in the future.
Although speculative, some other proteins or cofactors, energy sources for continued polymerizations, or sophisticated protein nano-structures may be necessary to complete sufficiently lengthy NR biosynthesis in vitro.
It was reported that a recombinant rubber tree CPT (HRT1) or a lettuce CPT (LsCPT3) alone without the respective recombinant CBP was sufficient to synthesize NR with average ∼1 million Da Mw, when expressed in, or reconstituted with, the washed rubber particles isolated from the rubber tree (Asawatreratanakul et al., 2003; Yamashita et al., 2016). To date, these two reports, which use washed rubber particles, are the only studies demonstrating in vitro reconstitutions of ∼1 million kDa NR using CPT recombinant proteins. It is attractive to include the washed rubber particles (mixtures of all proteins and lipid mixtures) as structural components in vitro, but this complicates the interpretation of the in vitro data. In addition, both reports showed no need of externally supplied rubber tree CBP or lettuce CBP for NR biosynthesis despite of their in planta interaction (Qu et al., 2015; Yamashita et al., 2016). Furthermore, these results contradict the in vivo CBP silencing evidence in lettuce and dandelion, which independently demonstrated the absolute necessity of CBP for NR biosynthesis (Epping et al., 2015; Qu et al., 2015). The discrepancy of these data cannot be easily resolved unless the rubber tree and lettuce/dandelion have different mechanisms for NR biosynthesis. Further investigation is necessary to address whether NR formation is catalyzed by the same or distinct mechanism in the rubber tree (Euphorbiaceae family) and other NR-producing plants (e.g., Asteraceae family; lettuce, dandelion, guayule, and sunflower).
In summary, three PaCPTs and one PaCBP cDNAs were identified from guayule in this work, and protein–protein interactions between PaCPTs and PaCBP have been substantiated by rer2Δ srt1Δ yeast complementation assays, split ubiquitin yeast 2-hybrid assays, and co-immunoprecipitation. The co-expression of each PaCPT and PaCBP in yeast allowed for the formation of the PaCPT–PaCBP hetero-protein complex for the efficient enzymatic synthesis of dehydrodolichyl diphosphate in microsomal in vitro enzyme assays. The RNA-Seq and phylogenetic analyses indicated PaCPT3 to be the most likely CPT involved in guayule NR biosynthesis. Collectively, these data suggest that any future bio-engineering efforts should be directed toward PaCPT3 and its binding partner, PaCBP to increase NR yield in guayule.
D-KR designed the research, constructed the phylogenetic trees, and wrote the manuscript with AL. AL conducted the gene cloning, yeast complementation, and co-immunoprecipitation. MK conducted yeast complementation and 2-hybrid. E-JK and AL conducted in vitro enzyme assays. SS and HS generated RNA-Seq data.
This work was supported by the National Science and Engineering Research Council of Canada (NSERC) and Canada Research Chair (CRC) program to D-KR. This work was also supported by a grant from and Basic Science Research Program by the National Research Foundation of Korea (NRF) funded by the Ministry of Education (2017R1A6A3A03003409) and the Next-Generation BioGreen 21 Program (SSAC, Grant No. PJ01326501), RDA, South Korea to MK. This work was supported by a cooperative research and development agreement (CRADA10138) between Lawrence Berkeley National Laboratory and Bridgestone Research, LLC. The work made use of facilities provided by the Joint Bioenergy Institute supported by the United States Department of Energy, Office of Science, Office of Biological and Environmental Research, through contract DE-AC02-05CH11231 between Lawrence Berkeley National Lab and the United States Department of Energy. The pre-print version of this article was released to the bioRxiv repository and referenced in this article (Lakusta et al., 2018).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We thank Professor Igor Stagljar (University of Toronto, Canada) for providing the split ubiquitin membrane yeast 2-hybrid system.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2019.00165/full#supplementary-material
Akhtar, T. A., Matsuba, Y., Schauvinhold, I., Yu, G., Lees, H. A., Klein, S. E., et al. (2013). The tomato cis–prenyltransferase gene family. Plant J. 73, 640–652. doi: 10.1111/tpj.12063
Asawatreratanakul, K., Zhang, Y. W., Wititsuwannakul, D., Wititsuwannakul, R., Takahashi, S., Rattanapittayaporn, A., et al. (2003). Molecular cloning, expression, and characterization of cDNA encoding cis-prenyltransferases from Hevea brasiliensis. Eur. J. Biochem. 270, 4671–4680. doi: 10.1046/j.1432-1033.2003.03863.x
Brasher, M. I., Surmacz, L., Leong, B., Pitcher, J., Swiezewska, E., Pichersky, E., et al. (2015). A two-component enzyme complex is required for dolichol biosynthesis in tomato. Plant J. 82, 903–914. doi: 10.1111/tpj.12859
Cantú, D. J., Angulo-Sánchez, J. L., Rodríguez-García, R., and Kuruvadi, S. (1997). Seasonal growth, rubber and resin yield characteristics of guayule under natural environmental conditions. Ind. Crops Prod. 6, 131–137. doi: 10.1016/S0926-6690(96)00211-7
Chakrabarty, R., Qu, Y., and Ro, D. K. (2015). Silencing the lettuce homologs of small rubber particle protein does not influence natural rubber biosynthesis in lettuce (Lactuca sativa). Phytochemistry 113, 121–129. doi: 10.1016/j.phytochem.2014.12.003
Cornish, K. (2001). Biochemistry of natural rubber, a vital raw material, emphasizing biosynthetic rate, molecular weight and compartmentalization, in evolutionarily divergent plant species. Nat. Prod. Rep. 18, 182–189. doi: 10.1039/A902191D
Cornish, K., Scott, D. J., Xie, W., Mau, C. J. D., Zheng, Y. F., Liu, H. H., et al. (2018). Unusual subunits are directly involved in binding substrates for natural rubber biosynthesis in multiple plant species. Phytochemistry 156, 55–72. doi: 10.1016/j.phytochem.2018.08.014
Cornish, K., and Siler, D. J. (1995). Effect of different allylic diphosphates on the initiation of new rubber molecules and on cis-1,4-polyisoprene biosynthesis in guayule (Parthenium argentatum Gray). J. Plant Physiol. 147, 301–305. doi: 10.1016/S0176-1617(11)82157-7
Cornish, K., Wood, D. F., and Windle, J. J. (1999). Rubber particles from four different species, examined by transmission electron microscopy and electron-paramagnetic-resonance spin labeling, are found to consist of a homogeneous rubber core enclosed by a contiguous, monolayer biomembrane. Planta 210, 85–96. doi: 10.1007/s004250050657
Denecke, J., and Kranz, C. (2009). Hypoglycosylation due to dolichol metabolism defects. Biochim. Biophys. Acta Mol. Basis Dis. 1792, 888–895. doi: 10.1016/j.bbadis.2009.01.013
Dennis, M. S., and Light, D. R. (1989). Rubber elongation factor from Hevea brasiliensis. Identification, characterization, and role in rubber biosynthesis. J. Biol. Chem. 264, 18608–18617.
Epping, J., van Deenen, N., Niephaus, E., Stolze, A., Fricke, J., Huber, C., et al. (2015). A rubber transferase activator is necessary for natural rubber biosynthesis in dandelion. Nat. Plants 1:15048. doi: 10.1038/nplants.2015.48
Fujihashi, M., Zhang, Y. W., Higuchi, Y., Li, X. Y., Koyama, T., and Miki, K. (2001). Crystal structure of cis-prenyl chain elongating enzyme, undecaprenyl diphosphate synthase. Proc. Natl. Acad. Sci. U.S.A. 98, 4337–4342. doi: 10.1073/pnas.071514398
Gisler, S. M., Kittanakom, S., Fuster, D., Wong, V., Bertic, M., Radanovic, T., et al. (2008). Monitoring protein-protein interactions between the mammalian integral membrane transporters and PDZ-interacting partners using a modified split-ubiquitin membrane yeast two-hybrid system. Mol. Cell. Proteomics 7, 1362–1377. doi: 10.1074/mcp.M800079-MCP200
Haas, B. J., Papanicolaou, A., Yassour, M., Grabherr, M., Blood, P. D., Bowden, J., et al. (2013). De novo transcript sequence reconstruction from RNA-seq using the Trinity platform for reference generation and analysis. Nat. Protoc. 8, 1494–1512. doi: 10.1038/nprot.2013.084
Harrison, K. D., Park, E. J., Gao, N., Kuo, A., Rush, J. S., Waechter, C. J., et al. (2011). Nogo-B receptor is necessary for cellular dolichol biosynthesis and protein N-glycosylation. EMBO J. 30, 2490–2500. doi: 10.1038/emboj.2011.147
Hillebrand, A., Post, J. J., Wurbs, D., Wahler, D., Lenders, M., Krzyzanek, V., et al. (2012). Down-regulation of small rubber particle protein expression affects integrity of rubber particles and rubber content in Taraxacum brevicorniculatum. PLoS One 7:e41874. doi: 10.1371/journal.pone.0041874
Hodgins, K. A., Lai, Z., Oliveira, L. O., Still, D. W., Scascitelli, M., Barker, M. S., et al. (2014). Genomics of Compositae crops: reference transcriptome assemblies and evidence of hybridization with wild relatives. Mol. Ecol. Resour. 14, 166–177. doi: 10.1111/1755-0998.12163
Horn, P. J., James, C. N., Gidda, S. K., Kilaru, A., Dyer, J. M., Mullen, R. T., et al. (2013). Identification of a new class of lipid droplet-associated proteins in plants. Plant Physiol. 162, 1926–1936. doi: 10.1104/pp.113.222455
Kajiura, H., Suzuki, N., Mouri, H., Watanabe, N., and Nakazawa, Y. (2018). Elucidation of rubber biosynthesis and accumulation in the rubber producing shrub, guayule (Parthenium argentatum Gray). Planta 247, 513–526. doi: 10.1007/s00425-017-2804-7
Kera, K., Takahashi, S., Sutoh, T., Koyama, T., and Nakayama, T. (2012). Identification and characterization of a cis, trans-mixed heptaprenyl diphosphate synthase from Arabidopsis thaliana. FEBS J. 279, 3813–3827. doi: 10.1111/j.1742-4658.2012.08742.x
Kwon, M., Kwon, E. J. G., and Ro, D. K. (2016). cis-Prenyltransferase and Polymer Analysis from a Natural Rubber Perspective. Methods Enzymol. 576, 121–145. doi: 10.1016/bs.mie.2016.02.026
Lakusta, A. M., Kwon, M., Kwon, E. J. G., Stonebloom, S., Scheller, H. V., and Ro, D. K. (2018). Characterization of cis-prenyltransferase complexes in guayule (Parthenium argentatum), an alternative natural rubber-producing plant. bioRxiv [Preprint]. doi: 10.1101/384149
Lu, Y.-P., Liu, H.-G., and Liang, P.-H. (2009). Different reaction mechanisms for cis- and trans-prenyltransferases. Biochem. Biophys. Res. Commun. 379, 351–355. doi: 10.1016/j.bbrc.2008.12.061
Madhavan, S., Greenblatt, G. A., Foster, M. A., and Benedict, C. R. (1989). Stimulation of isopentenyl pyrophosphate incorporation into polyisoprene in extracts from guayule plants (Parthenium argentatum Gray) by low temperature and 2-(3,4-dichlorophenoxy) triethylamine. Plant Physiol. 89, 506–511. doi: 10.1104/pp.89.2.506
Miller, J. H. (1972). Experiments in Molecular Genetics. New York, NY: Cold Spring Harbor Laboratory, 352–355.
Mooibroek, H., and Cornish, K. (2000). Alternative sources of natural rubber. Appl. Microbiol. Biotechnol. 53, 355–365. doi: 10.1007/s002530051627
Morton, M. (2013). “Introduction to polymer science,” in Rubber Technology, ed. M. Morton (Berlin: Springer Science Business Media), 1–19.
Oh, S. K., Kang, H., Shin, D. H., Yang, J., Chow, K. S., Yeang, H. Y., et al. (1999). Isolation, characterization, and functional analysis of a novel cDNA clone encoding a small rubber particle protein from Hevea brasiliensis. J. Biol. Chem. 274, 17132–17138. doi: 10.1074/jbc.274.24.17132
Orts, W. J., and McMahan, C. M. (2016). Biorefinery developments for advanced biofuels from a sustainable array of biomass feedstocks: survey of recent biomass conversion research from agricultural research service. Bioenergy Res. 9, 430–446. doi: 10.1007/s12155-016-9732-4
Ponciano, G., Dong, N., Chen, G., and McMahan, C. M. (2018). A bicistronic transgene system for genetic modification of Parthenium argentatum. Plant Biotechnol. Rep. 12, 149–155. doi: 10.1007/s11816-018-0478-7
Post, J., Van Deenen, N., Fricke, J., Kowalski, N., Wurbs, D., Schaller, H., et al. (2012). Laticifer-specific cis-prenyltransferase silencing affects the rubber, triterpene, and inulin content of Taraxacum brevicorniculatum. Plant Physiol. 158, 1406–1417. doi: 10.1104/pp.111.187880
Qu, Y., Chakrabarty, R., Tran, H. T., Kwon, E. J., Kwon, M., Nguyen, T. D., et al. (2015). A lettuce (Lactuca sativa) homolog of human Nogo-B receptor interacts with cis-prenyltransferase and is necessary for natural rubber biosynthesis. J. Biol. Chem. 290, 1898–1914. doi: 10.1074/jbc.M114.616920
Rasutis, D., Soratana, K., Mcmahan, C., and Landis, A. E. (2015). A sustainability review of domestic rubber from the guayule plant. Ind. Crops Prod. 70, 383–394. doi: 10.1016/j.indcrop.2015.03.042
Rivano, F., Mattos, C. R. R., Cardoso, S. E. A., Martinez, M., Cevallos, V., Le Guen, V., et al. (2013). Breeding Hevea brasiliensis for yield, growth and SALB resistance for high disease environments. Ind. Crops Prod. 44, 659–670. doi: 10.1016/j.indcrop.2012.09.005
Robinson, M. D., Mccarthy, D. J., and Smyth, G. K. (2010). EdgeR: a Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 26, 139–140. doi: 10.1093/bioinformatics/btp616
Sato, M., Sato, K., Nishikawa, S., Hirata, A., Kato, J., and Nakano, A. (1999). The yeast RER2 gene, identified by endoplasmic reticulum protein localization mutations, encodes cis-prenyltransferase, a key enzyme in dolichol synthesis. Mol. Cell. Biol. 19, 471–483. doi: 10.1128/MCB.19.1.471
Shimizu, N., Koyama, T., and Ogura, K. (1998). Molecular cloning, expression, and purification of undecaprenyl diphosphate synthase: no sequence similarity between E- and Z-prenyl diphosphate synthases. J. Biol. Chem. 273, 19476–19481. doi: 10.1074/jbc.273.31.19476
Snider, J., Kittanakom, S., Curak, J., and Stagljar, I. (2010). Split-ubiquitin based membrane yeast two-hybrid (MYTH) system: a powerful tool for identifying protein-protein interactions. J Vis Exp. 36:e1698. doi: 10.3791/1698
Stagljar, I., Korostensky, C., Johnsson, N., and Te Heesen, S. (1998). A genetic system based on split-ubiquitin for the analysis of interactions between membrane proteins in vivo. Proc. Natl. Acad. Sci. U.S.A. 95, 5187–5192. doi: 10.1073/pnas.95.9.5187
Surmacz, L., and Swiezewska, E. (2011). Polyisoprenoids – secondary metabolites or physiologically important superlipids? Biochem. Biophys. Res. Commun. 407, 627–632. doi: 10.1016/j.bbrc.2011.03.059
Tamura, K., Stecher, G., Peterson, D., Filipski, A., and Kumar, S. (2013). MEGA6: molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 30, 2725–2729. doi: 10.1093/molbev/mst197
Tang, C., Yang, M., Fang, Y., Luo, Y., Gao, S., Xiao, X., et al. (2016). The rubber tree genome reveals new insights into rubber production and species adaptation. Nat. Plants 2:16073. doi: 10.1038/nplants.2016.73
Teng, K. H., and Liang, P. H. (2012). Structures, mechanisms and inhibitors of undecaprenyl diphosphate synthase: a cis-prenyltransferase for bacterial peptidoglycan biosynthesis. Bioorg. Chem. 43, 51–57. doi: 10.1016/j.bioorg.2011.09.004
van Beilen, J. B., and Poirier, Y. (2007). Guayule and russian dandelion as alternative sources of natural rubber. Crit. Rev. Biotechnol. 27, 217–231. doi: 10.1080/07388550701775927
Yamashita, S., Yamaguchi, H., Waki, T., Aoki, Y., Mizuno, M., Yanbe, F., et al. (2016). Identification and reconstitution of the rubber biosynthetic machinery on rubber particles from Hevea brasiliensis. eLife 5:e19022. doi: 10.7554/eLife.19022.001
Yao, Z., Darowski, K., St-Denis, N., Wong, V., Offensperger, F., Villedieu, A., et al. (2017). A global analysis of the receptor tyrosine kinase-protein phosphatase interactome. Mol. Cell. 65, 347–360. doi: 10.1016/j.molcel.2016.12.004
Keywords: terpenoid, cis-prenyltransferase, guayule (Parthenium argentatum), polyisoprene, protein complex
Citation: Lakusta AM, Kwon M, Kwon E-JG, Stonebloom S, Scheller HV and Ro D-K (2019) Molecular Studies of the Protein Complexes Involving Cis-Prenyltransferase in Guayule (Parthenium argentatum), an Alternative Rubber-Producing Plant. Front. Plant Sci. 10:165. doi: 10.3389/fpls.2019.00165
Received: 30 September 2018; Accepted: 31 January 2019;
Published: 25 February 2019.
Edited by:
Dorothea Tholl, Virginia Tech, United StatesReviewed by:
Tariq Akhtar, University of Guelph, CanadaCopyright © 2019 Lakusta, Kwon, Kwon, Stonebloom, Scheller and Ro. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Dae-Kyun Ro, ZGFla3l1bi5yb0B1Y2FsZ2FyeS5jYQ==
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.