 Aurélie Charrier†
Aurélie Charrier† Emilie Vergne
Emilie Vergne Nicolas Dousset
Nicolas Dousset Aurélien Petiteau
Aurélien Petiteau Elisabeth Chevreau
Elisabeth Chevreau- IRHS, INRA, Agrocampus-Ouest, Université d’Angers, SFR 4207 QuaSaV, Beaucouzé, France
Targeted genome engineering has emerged as an alternative to classical plant breeding and transgenic methods to improve crop plants. Among other methods (zinc finger nucleases or TAL effector nucleases) the CRISPR-Cas system proved to be the most effective, convenient and least expensive method. In this study, we optimized the conditions of application of this system on apple and explored its feasibility on pear. As a proof of concept, we chose to knock-out the Phytoene Desaturase (PDS) and Terminal Flower 1 (TFL1) genes. To improve the edition efficiency, two different single guide RNAs (gRNAs) were associated to the Cas9 nuclease for each target gene. These gRNAs were placed under the control of the U3 and U6 apple promoters. Characteristic albino phenotype was obtained for 85% of the apple transgenic lines targeted in MdPDS gene. Early flowering was observed in 93% of the apple transgenic lines targeted in MdTFL1.1 gene and 9% of the pear transgenic lines targeted in PcTFL1.1. Sequencing of the target zones in apple and pear CRISPR-PDS and CRISPR-TFL1.1 transgenic lines showed that the two gRNAs induced mutations but at variable frequencies. In most cases, Cas9 nuclease cut the DNA in the twenty targeted base pairs near the protospacer adjacent motif and insertions were more frequent than deletions or substitutions. The most frequent edition profile of PDS as well as TFL1.1 genes was chimeric biallelic. Analysis of a sample of potential off-target sequences of the CRISPR-TFL1.1 construct indicated the absence of edition in cases of three mismatches. In addition, transient transformation with the CRISPR-PDS construct produced two T-DNA free edited apple lines. Our overall results indicate that, despite the frequent occurrence of chimerism, the CRISPR-Cas 9 system is a powerful and precise method to induce targeted mutagenesis in the first generation of apple and pear transgenic lines.
Introduction
Apple (Malus x domestica Bork.) is one of the major fruit crops produced in the world with a production over 89 million tons in 2016. The world pear production in 2016 reached 27 millions tons, including both European pears (Pyrus communis L.) and Asian pears (P. sp.) (FAOSTAT1). Conventional breeding of both species is limited by their long reproductive cycle and their high degree of heterozygosity. In addition, most fruit trees are produced by clonal propagation, traditional cultivars are still dominant and the speed of introduction of new hybrid varieties on the market is slow. In this context, genetic engineering appears as a powerful tool to accelerate the improvement of existing apple and pear elite cultivars. The sequencing of the apple (Velasco et al., 2010; Daccord et al., 2017) and pear (Wu et al., 2013; Chagné et al., 2014) genomes has opened the way to the development of many genomic resources, which also increases the need for accurate tools of gene function analysis in these species. Apple and pear are amenable to genetic transformation since 1989 (James et al., 1989) and 1996 (Mourgues et al., 1996), respectively. Numerous studies have improved genetic engineering tools for apple as well as pear and the number of clonal genotypes amenable to genetic transformation is now about 20 in Pyrus and 50 in Malus (Malnoy et al., 2008a,b).
Genome editing technologies have tremendously advanced during the last years and they now offer a mean for rational and precise modification of DNA sequences in many plant species. The Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR) nuclease Cas9 efficiently breaks the double strand of DNA at a predefined target site and non-homologous end-joining (NHEJ) permits the recovery of point mutations causing gene knock-out (Belhaj et al., 2013). This targeted mutagenesis technology is rapidly progressing in fruit trees and a number of successful gene knock-outs have been reported in Citrus (Jia and Wang, 2014; Jia et al., 2016, 2017; Peng et al., 2017), grape (Ren et al., 2016; Nakajima et al., 2017; Wang X. et al., 2018), kiwifruit (Wang Z. et al., 2018) and cacao (Fister et al., 2018). In apple, a first report indicated successful knock-out of the phytoene desaturase (PDS) gene in the rootstock ‘JM2’ (Nishitani et al., 2016). In this case, the Streptococcus pyogenes Cas9 was placed under the control of the CaMV35S promoter and several gRNAs of various lengths (18 or 20 bp) were placed under the control of the AtU6-1 promoter and tested separately. A rate of edition of 31.8% was obtained with clear or partial albino phenotypes. It is thus necessary to improve the methodology of genome editing in apple to reach higher efficiencies and to describe more precisely the complexity of the edition profiles. The only other report of apple gene editing concerns the efficient delivery of CRISPR-Cas9 ribonucleoproteins targeting the apple MLO-7 gene into apple protoplasts (Malnoy et al., 2016). However, no stably edited plants were regenerated from the edited protoplasts. Therefore, the production of T-DNA free edited apple lines is still a challenge. To our knowledge, no report of gene editing via CRISPR-Cas9 has been published so far on pear. The first proof of concept of pear genome editing is still to come.
In the present work our aims were: (i) to obtain high frequencies of gene knock-out of several apple easily scorable genes; (ii) to describe precisely the type of editions in T0 transformants; and (iii) to extend the technology to pear. For this purpose, we chose two target genes. The PDS gene disruption results in albino and dwarf phenotypes by impairing chlorophyll, carotenoid and gibberellin biosynthesis (Qin et al., 2007). The MdPDS gene is encoded by a single copy gene in the apple genome. The TFL1 gene is a floral repressor and its silencing leads to accelerate flowering (Shannon and Meeks-Wagner, 1991). Two TFL1 homologous genes are present in the apple genome and expressed in vegetative tissues (Mimida et al., 2009). MdTFL1 genes have been silenced through antisense (Kotoda et al., 2006), virus-induced gene silencing (Sasaki et al., 2011) or siRNA (Flachowsky et al., 2012; Weigl et al., 2015). In all cases, early flowering phenotypes were observed. Likewise in pear, expression of an RNAi cassette containing a sequence of the apple TFL1 led to the inhibition of both PcTFL-1 and Pc-TFL-2 and early flowering in the transgenic line (Freiman et al., 2012). We successfully edited MdPDS and MdTFL1.1 genes at high frequencies (>85%) in apple and demonstrated that chimeric biallelic patterns of edition are predominant. The PcTFL1.1 gene was also edited in pear at a lower frequency.
Materials and Methods
Biological Material
Bacterial strains included One Shot® TOP10 Chemically Competent Escherichia coli (Thermo Fisher Scientific) for cloning purposes, and Agrobacterium tumefaciens EHA105 (Hood et al., 1993) carrying both the binary vector of interest and the ternary plasmid pBBR1MCS-5 with a constitutive virG gene (van der Fits et al., 2000) for plant transformation.
The experiments were performed on two genotypes: the apple ‘Gala’ and the pear ‘Conference’. In vitro proliferating shoot cultures of the apple ‘Gala’ were micropropagated on Murashige and Skoog (1962) medium supplemented with 0.5 mg/l 6-benzyladenine (BA) and 0.1 mg/l 3-indolebutyric acid (IBA). Cultures of the pear ‘Conference’ were micropropagated as described by Leblay et al. (1991) on a derivative of Lepoivre’s medium supplemented with 0.5 mg/l 6-BA and 0.1 mg/l IBA. All cultures were grown at 22–24° C with a 16:8 h light:dark photoperiod (cool white fluorescent tubes, 40–60 μmol m-2 s-1) and transferred to fresh medium every 4 weeks.
Construction of Vectors
Binary vectors CRISPR-PDS and CRISPR-TFL1.1 used in this study were derived from pDE-CAS9 vector (Fauser et al., 2014) (Figure 1). Each construct contained two gRNAs with a different promoter (MdU3 or MdU6) and targeted MdPDS (MD04G0021400) or MdTFL1.1 (AB052994). First the bar resistance cassette in pDE-CAS9 vector was replaced by the nptII resistance cassette from pKGW vector (Karimi et al., 2002) by restriction/ligation at HindIII sites to create pDE-CAS9Kr vector. Each gRNA cassette marked out by attB gateway sites was synthesized independently by Integrated DNA Technology, Inc. (San Jose, CA, United States) and then cloned in pDONR207 vector by BP cloning (Gateway system; Thermo Fisher Scientific, MA, United States; Hartley et al., 2000). Then the ‘U6gRNA2’ cassette was placed after the ‘U3gRNA1’ cassette by restriction/ligation at XhoI/PstI sites in the donor vector and SalI/PstI sites in the destination vector, to create pDONR207-U3gRNA1-U6gRNA2 vector. Gateway LR cloning between pDONR2017-U3gRNA1-U6gRNA2 vector and pDE-CAS9Kr vector was then performed to create CRISPR-PDS or CRISPR-TFL1.1. Sequences are given in Supplementary File S1. Primers used to verify the cloning at each step (primers 7 and 8: BP cloning and gRNAs addition, primers 9 and 10: LR cloning) are indicated in Table 1. MdU3 or MdU6 promoters driving gRNAs were, respectively, found upstream MD10G1073100 and MD07G1138500 genes2 by BLAST with AtU3 and AtU6 sequences (respectively, found upstream X52629 and X52528; Marshallsay et al., 1990). Sequences are given in Supplementary File S1. Target sequences in gRNA were chosen with CRISPOR software3 (Haeussler et al., 2016) using the Malus domestica INRA GGDH13 Version 1.1 genome. Sequences are given in Supplementary File S1.
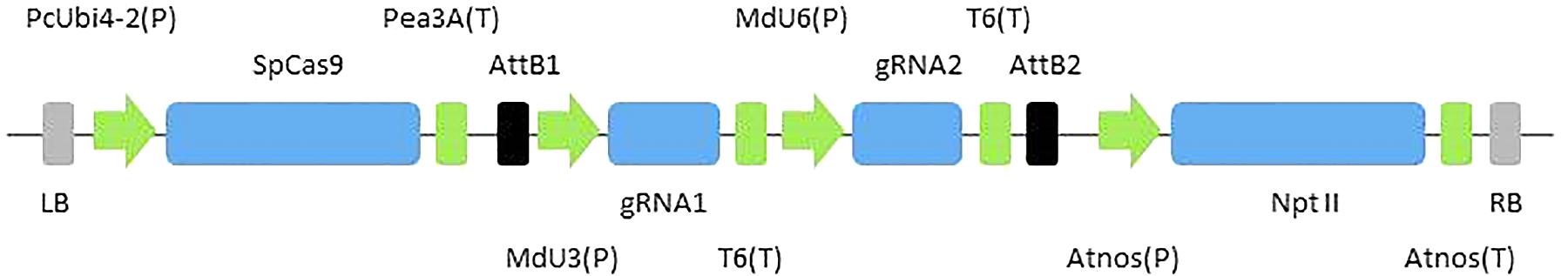
Figure 1. CRISPR-Cas expression construct. The Cas9 gene from Streptococcus pyogenes is driven by PcUbi4-2 promoter (P) from parsley (Petroselinum crispum) and transcription is terminated by the Pea3a terminator (T) from pea (Pisum sativum). gRNA1 and 2 are, respectively, driven by MdU3 and MdU6 promoters from Malus domestica and transcription is terminated by a polyT terminator. Transformants are selected with a nptII gene controlled by nos promoter and terminator from Agrobacterium tumefaciens. AttB1 and 2: sites resulting from the Gateway® LR recombination. LB and RB: T-DNA borders.
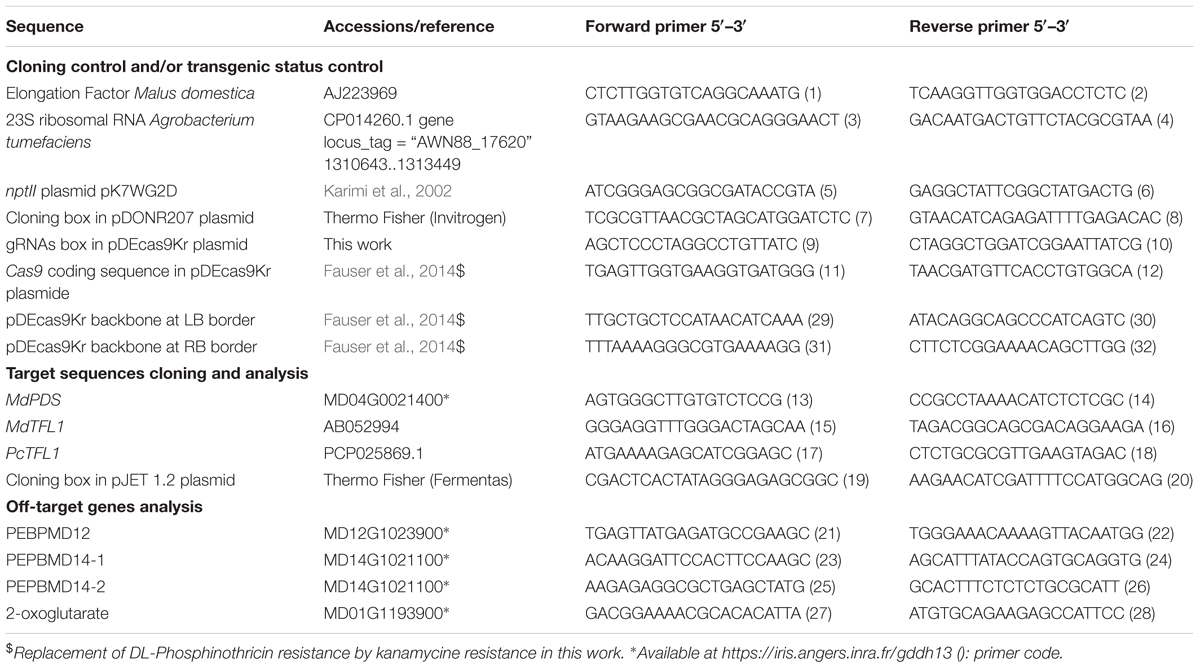
Table 1. Primers used in this work.
To perform a first proof of concept experiment of pear genome edition, we chose to use the same construct as for apple. However, in one of the two CRISPR-PDS gRNAs designed on the apple genome sequence, the PAM was lost because of a mismatch in the corresponding targets in PcPDS and the other gRNA presented also one mismatch with the corresponding target sequence in PcPDS, so the construct could not be used on pear. On the contrary, the two CRIPSR-TFL1.1 gRNAs presented one mismatch in position 18 after the PAM for gRNA 1 and no mismatch for gRNA2 in the corresponding targets in PcTFL1.1. Therefore the CRISPR-TFL1.1 was chosen to edit the pear genome.
Plant Transformation
For apple stable transformation, the youngest leaves of 4-week-old microshoots were vacuum-infiltrated in a suspension of A. tumefaciens at 108 bacteria/ml containing Silwet L-77® (Lehle Seeds, United States) at 0.002%, under -0.9 bar during 1 min. The leaves were then wounded transversely with a scalpel, and placed on apple regeneration medium consisting of MS medium containing 2–5 mg/l thidiazuron (TDZ), 0.5 mg/l IBA and 100 μM acetosyringone, solidified with PhytagelTM (SIGMA, United States) at 3 g/l, in the dark at 22–24°C.
For pear stable transformation, the youngest leaves of 4-week-old microshoots were vacuum-infiltrated in a suspension of A. tumefaciens at 107 bacteria/ml, under -0.9 bar during 1 min. The leaves were then wounded transversely with a scalpel, and placed on pear regeneration medium (Mourgues et al., 1996) containing 2 mg/l TDZ, 0.5 mg/l naphthalene acetic acid (NAA) and 100 μM acetosyringone, solidified with PhytagelTM (SIGMA, United States) at 3 g/l, in the dark at 22–24°C. For apple and pear, at the end of the co-culture, the leaves were placed on their respective regeneration medium containing 300 mg/l cefotaxime, 150 mg/l timentin and 100 mg/l kanamycin. The explants were kept in the dark and transferred to fresh medium every month for 6 months. Appearance of adventitious buds was monitored for a period of 6 months. All regenerated buds were micropropagated on the same medium as their mother plants, with the addition of 300 mg/l cefotaxime, 150 mg/l timentin, and 100 mg/l kanamycin.
For apple transient transformation, the protocol was modified as follows: the inoculum was a mix of the strain with a CRISPR construct and a strain carrying the gene coding the p19 protein of tomato bushy stunt virus as a suppresser of gene silencing (Voinnet et al., 2003), respectively, at 5 × 108 bacteria/ml and 2.5 × 108 bacteria/ml, supplemented with 0.002% Silwet L-77®. After agroinfiltration, the leaves were placed on regeneration medium containing 300 mg/l cefotaxime, 150 mg/l timentin, and 100 mg/l kanamycin for 1 week. Leaves were then transferred to the same medium without kanamycin. The explants were kept in the dark and transferred to fresh medium without kanamycin every month for 3 months. All regenerated buds were micropropagated on the same medium as their mother plants, with the addition of 300 mg/l cefotaxime, 150 mg/l timentin, without kanamycin.
Presence of Transgenes
Presence of transgenes and absence of contaminating agrobacteria were monitored by PCR. Genomic DNA of apple and pear leaves was extracted as described in Fulton et al. (1995). The primers used for the detection of (i) the gRNAs cassette (9 and 10), (ii) the Cas9 coding sequence (11 and 12), (iii) A. tumefaciens presence (3 and 4), (iv) nptII gene (5 and 6) and (v) elongation factor 1α (EF1α) coding gene as a marker of plant DNA suitability for PCR (1 and 2) are listed in Table 1. Amplifications were performed using GoTaq® Flexi DNA Polymerase (Promega, Madison, WI, United States) according to the manufacturer’s recommendations. The PCR reaction conditions were identical for the five genes: 95°C for 5 min, followed by 35 cycles at 95°C for 30 s, 58°C for 45 s, 72°C for 1 min, with a final extension at 72°C for 5 min. The PCR products were separated on a 1.5% agarose gel. Absence of plasmid backbone was also monitored in putative T-DNA free edited lines. Primers pairs used on either sides of the T-DNA (left: 29 and 30; right: 31 and 32) are listed in Table 1. The PCR reaction conditions were as above but the hybridization at 53°C.
Detection of Mutations
For mutation analysis of each target region, primer pairs were designed based on apple MdPDS (primers 13 and 14) and MdTFL1.1 (primers 15 and 16) genomic sequences from the apple ‘Golden Delicious’ genome or on the PcTFl1.1 (primers 17 and 18) genomic sequence from Pyrus communis (PCP025869.1). These primers amplified a DNA fragment of approximately 600–750 bp for MdPDS gene and approximately 2 kb for TFL1.1 (Md and Pc) genes surrounding each target. The wild type apple MdPDS and MdTFL1.1 gene fragments were amplified by PCR using genomic DNA from ‘Gala’. In the same way, the wild type pear PcTFL1.1 gene fragments were amplified by PCR using genomic DNA from ‘Conference.’ All primer pairs were described in Table 1. The PCR reaction conditions for MdPDS gene were as described in part 2.4. The PCR reactions for the TFL1.1 gene in apple and pear were performed with the following conditions: 95°C for 5 min, followed by 40 cycles at 95°C for 30 s, 58.5°C for 45 s, 72°C for 3 min, with a final extension at 72°C for 5 min. A touch-down PCR program (Marchand et al., 2003) (initial annealing temperature of 59°C, decreasing by 0.5°C per cycle down to 56°C) was used on a few recalcitrant pear and apple TFL1.1 samples using Q5® High-Fidelity DNA polymerase (New England Biolabs, MA, United States) following by gel purification to isolate the right amplicon.
Blunt and ligation reactions were performed on amplification products using the Sticky-End cloning protocol from CloneJET PCR Cloning Kit (Thermo Scientific, Waltham, MA, United States) according to the manufacturer’s recommendations. Bacterial transformation was performed in One ShotTM TOP10 Chemically Competent E. coli and spread on LB plates with 50 μg/ml ampicillin. MdPDS and TFL1.1 insertion colonies were checked by PCR (primers 19 and 20) using GoTaq® Flexi DNA Polymerase with pJET 1.2 primers (Table 1) using conditions described in part 2.4. The bacteria containing putative edited sequences were directly sent for sequencing. All sequencing results were compared with the reference sequence of the wild type apple MdPDS (MD04G0021400) or TFL1.1 (AB052994) genes and pear (PCP025869.1) TFL1.1 gene by alignment in MultAlin software (Corpet, 1988).
Putative off-target genes for gRNAs in our constructions were obtained thanks to CRISPOR software4 (Haeussler et al., 2016). For mutation analyses in putative off-target genes, strategy was as before: genes primers pairs (21 and 22, 23 and 24, 25 and 26, 27 and 28; Table 1) were designed based on genomic sequences from the apple ‘Golden Delicious’ genome (Daccord et al., 2017). They amplified a DNA fragment between 201 and 475 bp surrounding each putative target. PCR conditions were as described in part 2.4 and subsequent cloning and sequencing were as described just above.
Results
Production of Apple and Pear Transgenic Lines
In total, five stable transformation experiments were performed and resulted in variable rates of transformation (Table 2). For the CRISPR-PDS construct, 57 ‘Gala’ kanamycin resistant lines were produced in a single experiment whereas three experiments were needed to produce 30 ‘Gala’ kanamycin resistant lines with the CRISPR-TFLl.1 construct. For pear, a very high rate of transformation was achieved (23%) and 100 ‘Conference’ kanamycin resistant lines were produced with the CRIPSR-TFL1.1 construct. Only 54 lines were further studied.
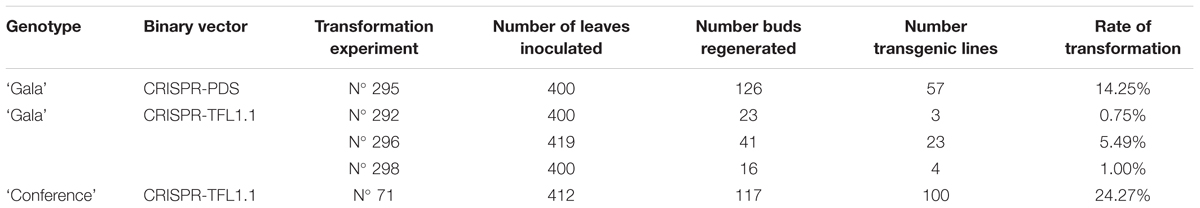
Table 2. Production of transgenic lines.
The ‘Gala’ kanamycin resistant-lines with the CRISPR-PDS construct which survived after 1 year of micropropagation were analyzed by PCR and EF1α gene was used as a marker of plant DNA suitability for PCR. Plasmid DNA extracted from A. tumefaciens strain containing the CRISPR-PDS construct and genomic DNAs extracted from a non-transgenic ‘Gala’ were used as controls. Amplification with UF/B1R primers showed that all the tested lines were free from A. tumefaciens contamination. Amplification with nptII primers confirmed that the 41 tested lines were true transgenic lines carrying the selectable marker gene. Amplification with primers for the gRNAs presence and for the Cas9 coding sequence showed that 39 of the 41 lines had integrated the full CRISPR-PDS construct. Two additional albino lines did not integrate the gRNAs box and the Cas9 coding gene. For the transgenic lines with the CRISPR-TFL1.1 construct, we decided to study only a sample of five early flowering lines per species. All of them had integrated the full CRISPR-TFL1.1 construct and were free from A. tumefaciens.
Phenotypic Analysis of Transgenic Lines
The expected phenotype of knock-out mutants of the PDS gene is dwarf and albinos. After the first subculture of the transgenic buds expressing the CRISPR-PDS construct on micropropagation medium, their original phenotype was recorded. At this early stage, 15 lines (26%) appeared pure white and 6 lines (10%) pure green. All the other lines (64%) showed various mixtures of white, green and variegated leaf phenotypes indicating a high level of chimerism (Figures 2A–C). All the transgenic lines were micropropagated over a period of 1 year and white sectors were selected at each subculture (Figure 2D). This led to the progressive disappearance of variegated phenotypes (only one line) and a majority of pure white (48 lines) or green (9 lines) remained. A rate of 84% of edition (48 out of 57 lines) was finally recorded.

Figure 2. Transgenic lines phenotypes. Phenotypes of ‘Gala’ transgenic lines containing the CRISPR-PDS construct: (A) pure green, (B) variegated, (C) pure white, (D) variegated line undergoing chimera dissociation through adventitious bud formation. Phenotypes of ‘Gala’ transgenic lines containing the CRISPR-TFL1 construct: (E) flower bud formation in apical position, (F) opened flower with all floral organs present, (G) loss of vegetative growth after 3 months of continuous flowering. Phenotype of ‘Conference’ transgenic line containing the CRISPR-TFL1construct: (H) opened flower with all floral organs present.
Flower buds appeared on terminal position on ‘Gala’ and ‘Conference’ transgenic lines expressing the CRISPR-TFL1.1 construct and the opened flowers showed the presence of all floral organs, often in irregular numbers (Figure 2). Flowering of ‘Gala’ transgenic lines occurred between one and 6 months after the beginning of their micropropagation (Figures 2E–H). Similarly to the albino phenotype, the flowering phenotype appeared progressively, first on one then on several shoots of each transgenic line. When the majority of the apical meristems of a single line turned floral, its micropropagation became very difficult due to the cessation of vegetative growth (Figure 2G). Most ‘Gala’ transgenic lines flowered after 3 months, and after 6 months 27 out of 30 lines had flowered, indicating a rate of 90% of edition. On the contrary, flowering of ‘Conference’ transgenic lines appeared between 4 and 12 months after the start of their micropropagation (Figure 3), and was limited to 5 lines out of 54, indicating a rate of edition of 9%.
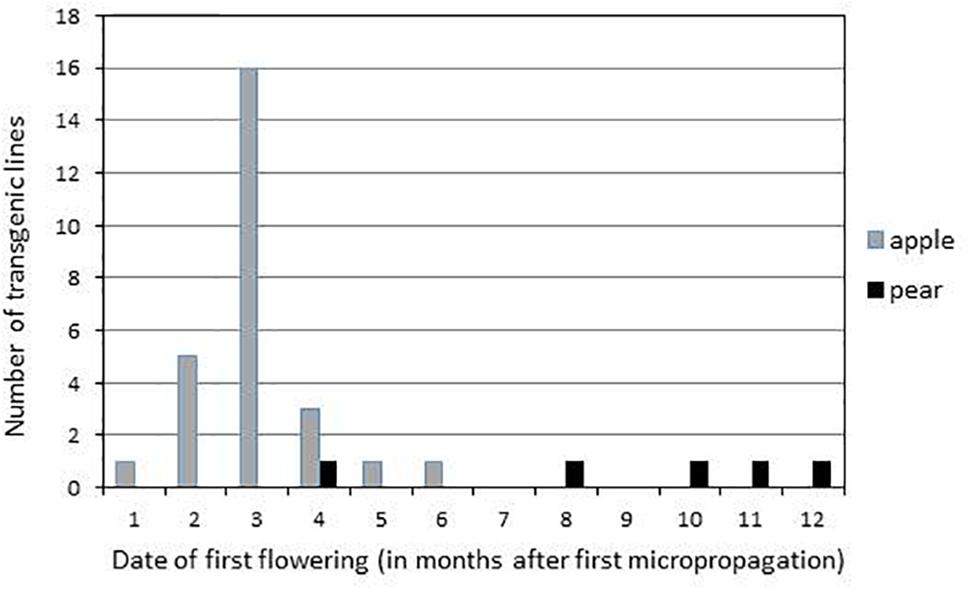
Figure 3. Date of first flowering of transgenic lines expressing the CRISPR-TFL1.1 construct.
Characterization of Targeted Mutation of MdPDS and TFL1.1 Genes
In total, 41 transgenic lines (subsequently named ‘lines’) with the CRISPR-PDS construct were analyzed including 37 albino and 4 green lines. For the majority of these lines, four bacterial clones (subsequently named ‘clones’) per transgenic line were sequenced, each containing an allele of the target sequence putatively edited. For two lines, only three clones could be obtained. In addition, for three lines 8 clones were sequenced in order to further explore the variability of edition profiles. A summary of the edition profiles is given in Table 3. For the MdPDS gene, a majority of edited lines were chimeric (87.8%). Among these chimeric lines, all albino lines presented at least one biallelic cell line and 8 transgenic lines presented at least one heterozygous cell line. Three of the four green lines were non-mutated and the last green line presented an edition on one clone out of four only. Among the 174 bacterial clones sequenced, both targets were simultaneously mutated in 51.1% of cases, target 1 was more often mutated alone (30.5%) than target 2 (3.5%). Small indels located in the target sequence were the most frequent outcome with 78.2% for target 1 and 50% for target 2. A great variability of types of indels was observed (Figure 4): 24 types of indels for target 1 and 17 for target 2. For target 1, 48.3% of the clones were represented by three specific indels: addition of a thymine at the 5th nucleotide after the PAM, addition of an adenine at the 4th nucleotide after the PAM and deletion of a thymine at the 4th nucleotide after the PAM. For target 2, 28.2% of the clones presented an addition of a cytosine at the 5th nucleotide after the PAM. Substitution indels were located more randomly in target sequence and frequently combined with deletions or additions. Larger deletions ranging from 11 to 29 bp were less frequent. Two particular cases involving reshuffling of large sequence fragments are detailed in Figure 5, the first case (A) resulted from a large substitution by a double copy of an upstream sequence, the second case (B) resulted from an inversion of the sequence between the two targets. Figure 6 presents the frequencies of the three types of indels according to their position at the target site. The majority of deletions (96.7% for target 1, 84.6% for target 2) were located only in targets. All additions were located on the 4th and the 5th nucleotides after PAM for both targets.
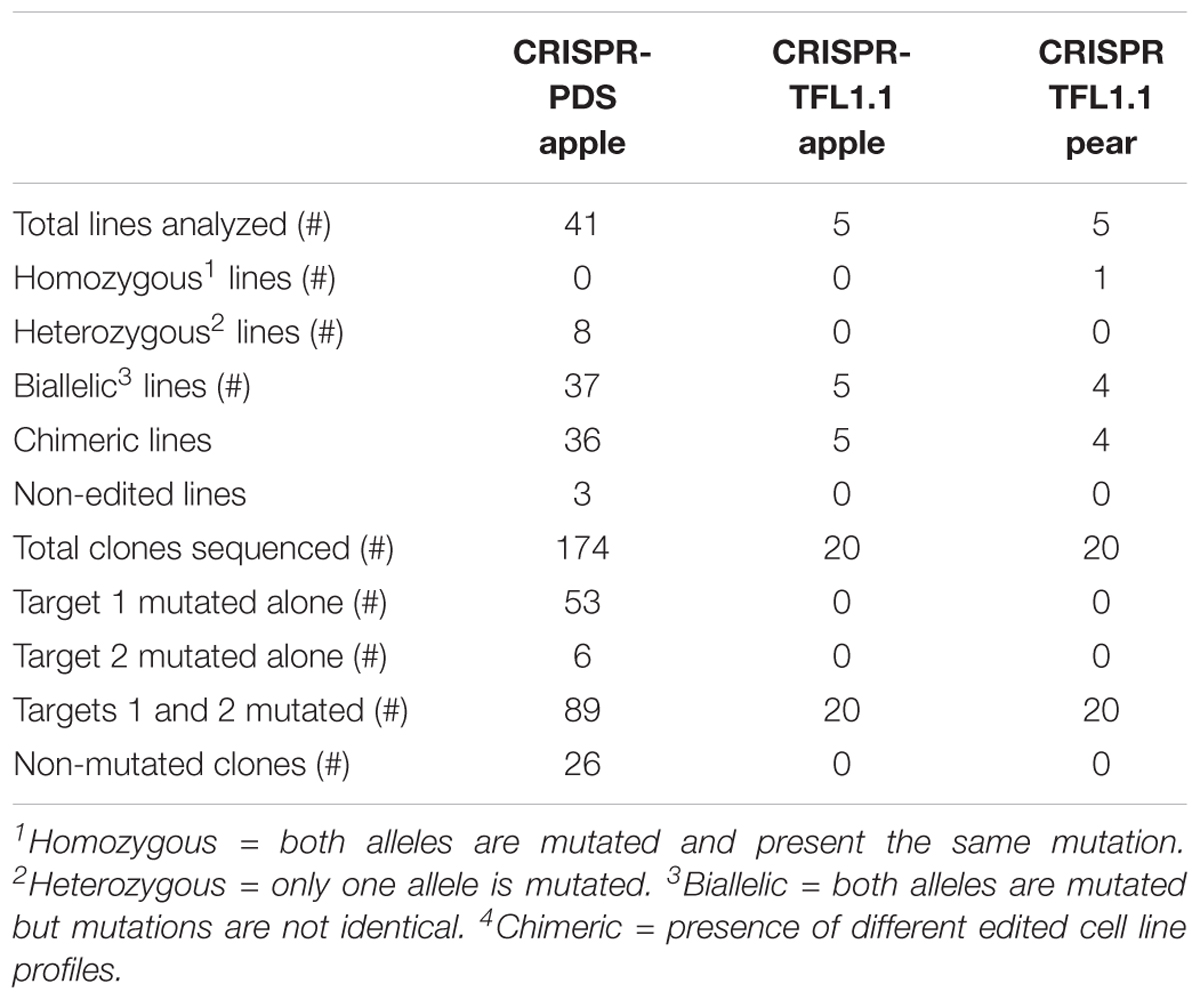
Table 3. Summary of edition profiles of transgenic lines.

Figure 4. Mutations in the apple MdPDS gene induced by CRISPR/Cas9. (A) Results for target 1, (B) results for target 2. Alignment was done in comparison to the wild type (WT). In color blue, the targeted sequence. This synthesis excludes two particular cases described in Figure 5 but included in the total for the frequency calculation. In orange, the PAM sequence. Mut: type of mutation with “–” for deletion, “+” for addition, “/” for substitution. Rep, number of occurrences of each indel. Freq, (%) frequencies calculated in relation to the number of occurrences of each indel among the total number of bacterial clones sequenced (174).
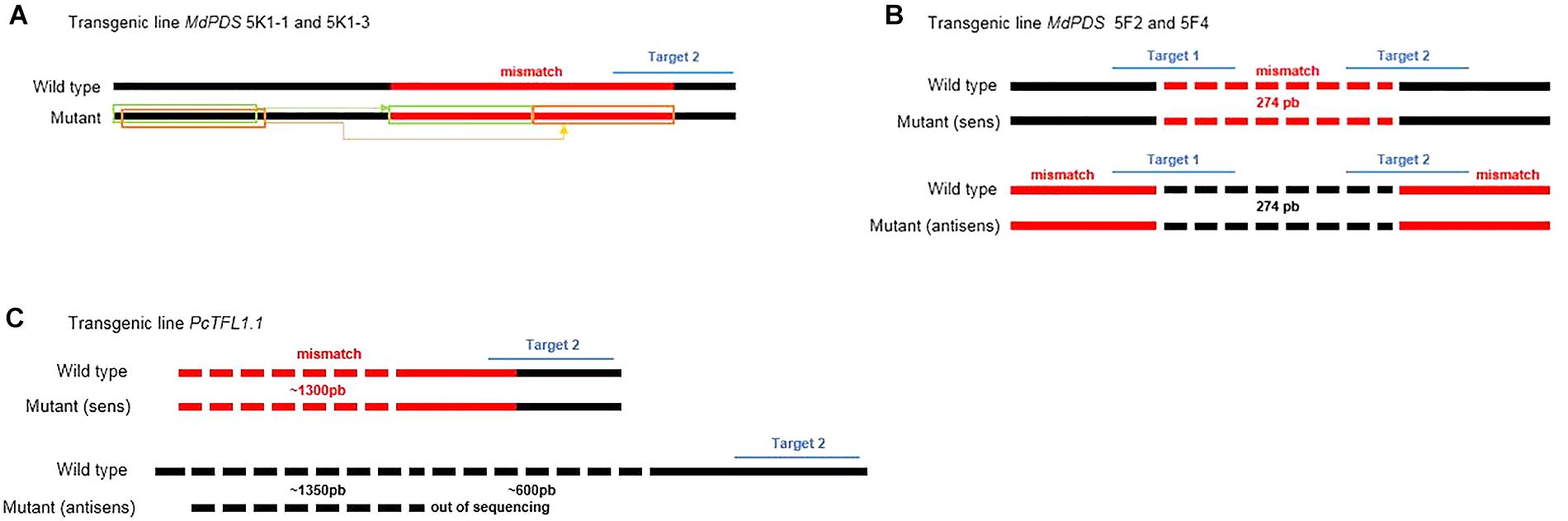
Figure 5. Schemes of particular edition profiles involving large scale mutations. (A) Mutation on target 2 of the MdPDS gene, observed on 2 bacterial clones from one edited line (MdPDS 5K1-1 and 5K1-3): a large mismatch (57 bp) concerns a part of target 2 resulting from a substitution of copies of two fragments upstream of the sequence. (B) Mutation concerning the two targets of the MdPDS gene, observed on 2 bacterial clones from one edited line (MdPDS 5F2 and 5F4): the large mismatch (274 bp) between the two targets results from the sequence reversal after Cas9 cutting and repair by the NHEJ system. (C) Mutation on target 2 of the PcTFL1.1 gene, observed on the 8 bacterial clones from the two edited lines PcTFL1.1 71AJ and 71CI: substitution of a reverse fragment from a locus upstream of the gene. Sequencing being limited in length, the mutation is not completely described and the inversion could concern a fragment longer than 1300 bp.

Figure 6. Frequencies of nucleotide editions of MdPDS targets according to indel positions. Frequencies were calculated in relation to the total number of edited MdPDS clones (174). Particular cases (alleles 5F2, 5F4 and 5K1-1, 5K1-3) are not presented. The distance between the end of the 1st target and the beginning of the 2nd is 272 bp.
A sample of five apple and five pear early flowering transgenic lines with the CRISPR-TFL1.1 construct were analyzed and four bacterial clones were sequenced per line. As for the CRISPR-PDS construct, results (Table 3) indicated a majority of chimeric biallelic profiles. However, one pear transgenic line showed an homozygous non-chimeric profile. The two targets were simultaneously edited in all apple and pear lines with the CRISPR-TFL1.1 construct, despite the presence of one mismatch between the gRNA1 and the target in pear. All edited lines for MdTFL1.1 and PcTFL1.1 analyzed are described in Figure 7. For both genes, a majority of alleles presented a deletion of one or more bases at the target sites. On target 1, major editions, including substitutions and additions were observed for 9 among 20 edited alleles. On target 2, 8 edited alleles presented a major reshuffling substitution schematized in Figure 5.

Figure 7. Mutations in the apple MdTFL1.1 (A) and pear PcTFL1.1 (B) genes induced by CRISPR/Cas9. Particular case (∗) is described in Figure 5. In orange, the PAM sequence. Mut: type of mutation with “–” for deletion, “+” for addition, “/” for substitution. Rep, number of repetitions of each indel. Freq, (%) frequencies calculated in relation to the number of repetition of each indel among the total number of clones sequenced (20 for both species).
Analysis of Potential Off-Target Mutations
Potential off-target sequences were identified using the CRISPOR software. We found no off-target genes for gRNA2 in CRISPR-PDS construct and two for gRNA1 with three mismatches each. In CRISPR-TFL1.1 construct, we found two putative off-target genes for each gRNA. gRNA1 could target MD12G1023900 and MD14G1021100 with, respectively, zero and three mismatches and gRNA2 could target MD14G1021100 and MD01G1193900 with three mismatches each (Table 4). Given the greater probability to affect off-target genes with CRISPR-TFL1.1 construct than with CRISPR-PDS construct, we analyzed only putative off-target genes of CRISPR-TFL1.1 construct.

Table 4. Analysis of the off-target gene with 0 mismatch for CRISPR-TFL1.1 gRNA1.
Two to four bacterial clones were sequenced for each off-target gene in five different transgenic lines (CRISPR-TFL1.1-1 to 5). We failed to design specific primers to amplify the putative target of gRNA2 in MD14G1021100. We found no mutations in putative off-target genes with three mismatches but, in each transgenic line, we found editions of MD12G1023900, targeted by gRNA1 without mismatch (Table 4).
Production of T-DNA Free Edited Lines
In order to evaluate the feasibility of production of T-DNA-free edited lines, a transient transformation with the CRISPR-PDS construct was performed on apple. The protocol was adapted to increase the frequency of transient expression by (i) a higher concentration of inoculum, (ii) co-inoculation with an A. tumefaciens strain carrying an inhibitor of silencing, (iii) kanamycin selection applied only during the period of T-DNA transient expression (1 week after the end of the co-culture). In total, 229 apple leaves were inoculated. A very high rate of regeneration was observed (71%). Among 747 regenerated buds, three albino buds were detected (0.4% mutation efficiency). PCR analysis indicated for two of them the absence of integration of Cas9 or nptII genes, gRNAs and left and right backbone sequences (Figure 8). Sequencing of four clones per edited line indicated that the two T-DNA free lines were homozygous and mutated only on target 1. One line showed a single deletion at the fourth base after the PAM and the other line presented a more complex mutation with three insertions and four substitutions. These two lines can be considered as T-DNA free edited lines.
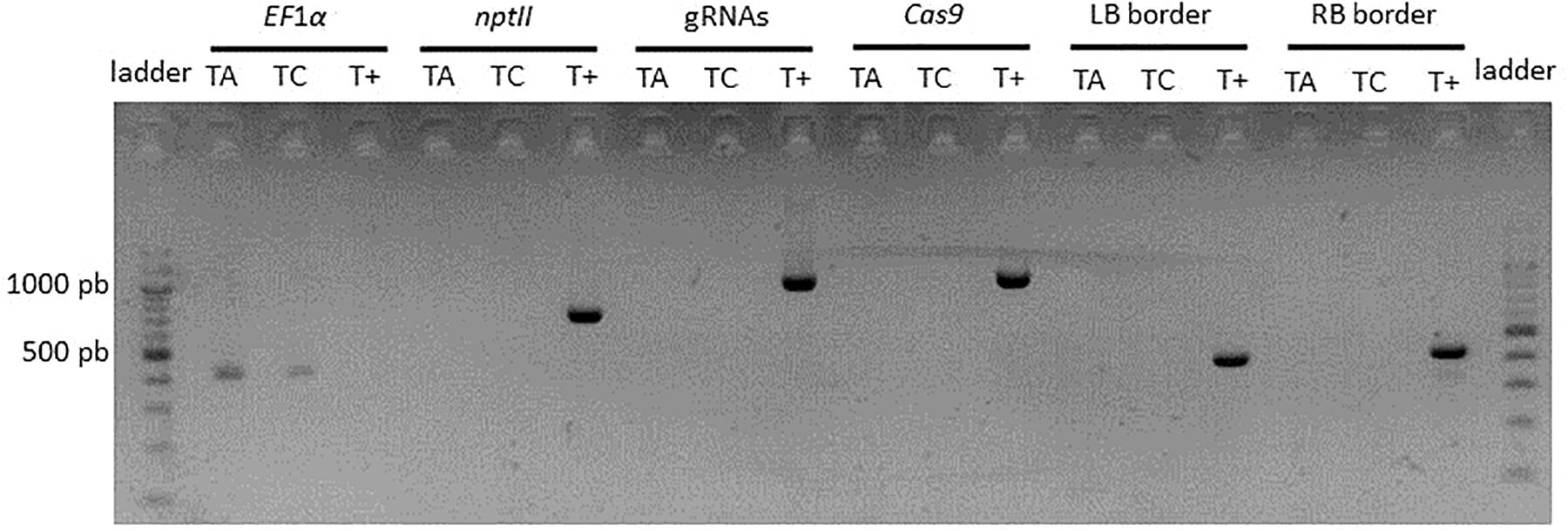
Figure 8. PCR analysis of T-DNA free edited lines. TA, T-DNA free edited line A. TC, T-DNA free edited line C. T+, binary vector CRISPR-TFL1.1 as a control.
Discussion
High Efficiencies of Editing Can Be Obtained in Apple by the Simultaneous Use of Two gRNAS Driven by Apple U3 and U6 Promoters
The present study demonstrates successful CRISPR-Cas9-mediated targeted edition of two different genes in apple, with very high rates of edition (84–90%) in the first generation after transformation (T0 plants). These ratios are higher than comparable ratios of other perennial crop: 51.7% of edited T0 plants in poplar (Fan et al., 2015), 30.6% in grape (Wang X. et al., 2018), 34.5% in orange (Peng et al., 2017). The only other report of CRISPR-Cas9 edition of the MdPDS gene in regenerated apple plants indicated a rate of edition of 31.8% in the rootstock ‘JM2’ (Nishitani et al., 2016). The difference of genotype could explain in part the difference of edition efficiency. But other differences in the construct design must be taken into account. First, Nishitani et al. (2016) used a Cas9 fused to GFBSD2 (GFP fused to the N-terminus of blasticidin S deaminase) under the control of the CaMV35S promoter. This fusion protein may be expressed less efficiently than the simple Cas9 [codon-optimized for Arabidopsis thaliana by Fauser et al. (2014)] placed under the control of the parsley ubiquitin promoter, in our study. Secondly, we used two gRNAs simultaneously and placed them under the control of the apple U3 and U6 promoters, whereas Nishitani et al. (2016) used several gRNAs separately under the control of the A. thaliana U6 promoter. This strategy probably increased the probability of edition of the target gene.
Pear TFL1.1 Gene Can Be Edited Using gRNAs Designed on the Apple Genome
Our study also reports for the first time the CRISPR-Cas9- mediated targeted mutation of a pear gene (PcTFL1.1) with a rate of edition of 9% of the observed T0 plants. To perform this first proof of concept experiment of pear genome edition, we chose to use the same construct as for apple edition of MdTFL1.1 gene. However, the CRIPSR-TFL1.1 gRNA1 designed on the apple genome sequence presented one mismatch in position 18 after the PAM with the corresponding target sequence in pear PcTFL1.1. The presence of this mismatch could explain the lower rate of mutated phenotype observed in pear. It is also possible, that in pear, the knockout of both TFL1 genes (PcTFL1.1 and PcTFL1.2) is necessary to release totally the floral repression. The only previous experiment to silence pear TFL1 used an RNAi construct based on MdTFL1.1 which silenced both pear TFL1 genes and led to an early flowering phenotype (Freiman et al., 2012). Yet, in our study, both gRNAs in the CRIPSR-TFL1.1 construct presented three mismatches with the corresponding target sequences in pear PcTFL1.2. Thus, in light of our results on off-target genes, we assume that PcTFL1.2 was not edited. On the contrary, targeting MdTFL1.1 alone and not other TFL1/CEN-like genes (MdTFL1a, MdCENa, and MdCENb; Mimida et al., 2009) appeared to be sufficient to get the early flowering phenotype. Indeed CRIPSR-TFL1.1 gRNA1 and gRNA2 showed three or more mismatches with the corresponding target sequences in those TFL1/CEN-like genes, which exclude their edition in our study.
Complex Edition Profiles and Chimerism Are Frequent in Apple and Pear Edited Lines
Phenotypic and molecular analyses of the PDS-edited apple lines produced in our study clearly indicated that the Cas9 protein is acting progressively during the whole period of stable transgenic bud regeneration (about 6 months). As already described by Nishitani et al. (2016), a majority of transgenic CRISPR-PDS lines initially appeared as a mixture of white, green and variegated sectors, indicating a high level of chimerism between edited and non-edited tissues in the same plant. Similarly, the flowering phenotype appeared very progressively on apple and pear CRISPR-TFL1.1 lines. The sequencing analyses were performed about 1 year after the regeneration of the transgenic buds, and a constant selection for the mutated phenotypes (albino or flowering) was applied during this period. Therefore most of the non-edited tissues were probably eliminated at the time of the molecular analyses. The sequencing of more than 214 bacterial clones revealed a minority of non-mutated alleles (12%). In the majority of lines (88%), more than two mutated allele sequences were detected, indicating chimerism between different edited cell-lines in the analyzed tissues. Up to six different alleles were observed in one of the lines for which eight clones were sequenced. Furthermore, bi-allelic mutants were the most frequent outcome and only one putatively mono-allelic homozygous line was observed among the 51 independent edited lines analyzed. Multiple mutated sequences and frequent biallelic mutations were also observed in grape (Nakajima et al., 2017; Wang X. et al., 2018), orange (Peng et al., 2017) or tomato (D’Ambrosio et al., 2018) T0 plants.
A clear difference of efficiency between the two gRNAs was observed in MdPDS-edited lines but not in MdTFL1.1 or PcTFL1.1-edited lines. Most of the edition profiles observed in our study were small additions or deletions at the targets sites, as a consequence of repair through NHEJ after gRNA-directed Cas9 cleavage. This is in agreement with the majority of reports which indicate that about half of CRISPR-Cas9-induced mutations are single-base insertions and the rest small deletions (Ma et al., 2016). It has been reported that multiple gRNAs targeting close targets in one gene can result in a large deletion in poplar (Fan et al., 2015). Similarly, paired-sgRNA vectors led to large fragment deletions in kiwifruit (Wang X. et al., 2018; Wang Z. et al., 2018). Despite of the use of two gRNAs, no deletions between the two targets were observed in our study. The only case of large mutation involving the two targets was the inversion of the fragment between the two targets observed in one edited line. In the previous report on apple using CRISPR-Cas9 to target MdPDS (Nishitani et al., 2016), only insertion or deletions (+1 to -8 bases) were obtained. This is probably due to the use of a single gRNA.
Occurrence of Off-Target Edition Depends on the Number of Mismatches in the gRNAs
It is well known that Cas9 can act on non-selective regions of genomic DNA known as “off-target” sites (Kadam et al., 2018). A high homology between the desired target and other zones in the genome increases the risk of off-target activity. Our results on apple confirmed that when complete homology exists between a gRNA and a non-target sequence, this off-target sequence was systematically edited. Our results on pear showed that in case of one mismatch distal from the PAM, the target sequence is also edited. On the contrary, potential off-target sequences containing three mismatches with the gRNA were never edited in apple.
Transient Expression Permits the Production of T-DNA Free Apple Edited Lines at Low Frequency
Using a modified protocol in order to favor transient expression of Cas9, we have been able to produce two lines mutated for MdPDS without integration of T-DNA elements or backbone sequences. To our knowledge, this is the first case of production of T-DNA free edited plants in a woody fruit species. The fact that these two lines present a single homozygous mutation at the target 1 site indicates a very early activity of Cas9 at the beginning of the regeneration process. However, the overall efficiency of this transient system remains very low (0.4% of edited lines). This is lower than the mutation frequencies obtained after agroinfiltration of CRISPR constructs targeting PDS gene in Nicotiana benthamiana leaves (Li et al., 2013; Nekrasov et al., 2013; Chen et al., 2018). The only other attempt to produce apple T-DNA free edited lines was through direct delivery of CRISPR-Cas9 ribonucleoproteins to apple protoplasts (Malnoy et al., 2016). Efficient edition of two target genes (MLO-7 and DIPM-1) was observed in the protoplasts after PEG treatment, but no plants were regenerated.
Conclusion
Our overall results indicate that CRISPR-Cas9-mediated knockout of targeted genes is very efficient in apple and possible in pear. Careful design of gRNAs without mismatch with the target gene should increase the frequency of pear edition. The high proportion of biallelic mutants obtained in our study is a good indication that CRISPR-Cas9-mediated knock-out is an efficient strategy for loss of function experiments in apple or pear. However, occurrence of chimerism in T0 plants is of particular concern for the genome editing of these trees which cannot be crossed to segregate the chimera without losing the genotype of the edited variety. Strategies to separate chimeras before analysis of the edited lines should be developed. Adventitious regeneration already proved to be efficient for chimeral release in naturally variegated cultivars of pear (Abu-Quaoud et al., 1990). Finally, our study demonstrates that the production of T-DNA free edited apple lines is feasible. Various strategies such as biolistics or viral delivery could be tested to increase the efficiency of recovery of these lines. In conclusion, the CRISPR-Cas9 system presented here now allows the establishment of a robust and precise genomic edition platform in apple and pear.
Author Contributions
AC and EV performed part of the experiments, analyzed the data, and revised the manuscript. ND and AR performed part of the experiments and revised the manuscript. AP performed part of the experiments, analyzed the data, and drafted the manuscript. EC designed the study and drafted the manuscript. All authors read and approved the final manuscript.
Funding
Thisproject has received funding from the French Government managed by the Research National Agency (ANR) under the Investment for the Future program (GENIUS project ANR11-BTBR-0001).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors sincerely thank F. Nogué (INRA Versailles) for his assistance in vector design and cloning strategy. The authors also thank M. Mazier and B. Danilo (INRA Avignon) for providing the plasmid pDE-CAS9Kr. The authors gratefully acknowledge the technical platform ANAN (SFR 4207 QUASAV) and L. Ogé (IRHS-GDO) for his helpful advice on gel purification.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2019.00040/full#supplementary-material
Footnotes
References
Abu-Quaoud, H., Skirvin, R. M., and Chevreau, E. (1990). In vitro separation of chimeral pears into their component genotypes. Euphytica 48, 189–196. doi: 10.1007/BF00037199
Belhaj, K., Chaparro-Garcia, A., Kamoun, S., and Nekrasov, V. (2013). Plant genome editing made easy: targeted mutagenesis in model and crop plants using the CRISPR/Cas system. Plant Methods 9:39. doi: 10.1186/1746-4811-9-39
Chagné, D., Crowhurst, R. N., Pindo, M., Thrimawithana, A., Deng, C., Ireland, H., et al. (2014). The draft genome sequence of European pear (Pyrus communis L. ‘Bartlett’). PLoS One 9:e92644. doi: 10.1371/journal.pone.0092644
Chen, L., Li, W., Katin-Grazzini, L., Ding, J., Li, Y., Gu, T., et al. (2018). A method for the production and expedient screening of CRISPR/Cas9-mediated non-transgenic mutant plants. Hortic. Res. 5:13. doi: 10.1038/s41438-018-0023-4
Corpet, F. (1988). Multiple sequence alignment with hierarchical clustering. Nucl. Acids Res. 16, 10881–10890. doi: 10.1093/nar/16.22.10881
Daccord, N., Celton, J. M., Linsmith, G., Becker, C., Choisne, N., Schijlen, E., et al. (2017). High-quality de novo assembly of the apple genome and methylome dynamics of early fruit development. Nat. Genet. 49, 1099–1106. doi: 10.1038/ng.3886
D’Ambrosio, C., Stigliani, A. L., and Giorio, G. (2018). CRISPR/Cas9 editing of carotenoid genes in tomato. Transgenic Res. 27, 367–378. doi: 10.1007/s11248-018-0079-9
Fan, D., Liu, T., Li, C., Jiao, B., Li, S., Hou, Y., et al. (2015). Efficient CRISPR/Cas9-mediated targeted mutagenesis in Populus in the first generation. Sci. Rep. 5:12217. doi: 10.1038/srep12217
Fauser, F., Schiml, S., and Puchta, H. (2014). Both CRISPR/Cas-based nucleases and nickases can be used efficiently for genome engineering in Arabidopsis thaliana. Plant J. 79, 348–359. doi: 10.1111/tpj.12554
Fister, A., Landherr, L., Maximova, S., and Guiltinan, M. J. (2018). Transient expression of CRISPR/Cas9 machinery targeting TcNPR3 enhances defense response in Theobroma cacao. Front. Plant Sci. 9:268. doi: 10.3389/fpls.2018.00268
Flachowsky, H., Szankowski, I., Waidmann, S., Peil, A., Tränker, C., and Hanke, M. V. (2012). The MdTFL1 gene of apple (Malus x domestica Borkh.) reduces vegetative growth and generation time. Tree Physiol. 32, 1288–1301. doi: 10.1093/treephys/tps/080
Freiman, A., Shlizerman, L., Golubowicz, S., Yabloviz, Z., Korchinsky, R., Cohen, Y., et al. (2012). Development of a transgenic early flowering pear (Pyrus communis L.) genotype by RNAi silencing of PcTFL1-1 and PcTFL1-2. Planta 235, 1239–1251. doi: 10.1007/s00425-011-1571-0
Fulton, T. M., Chunzoongse, J., and Tanksley, S. D. (1995). Microprep protocol for extraction of DNA from tomato and other herbaceous plants. Plant Mol. Biol. Rep. 13, 207–209. doi: 10.1007/BF02670897
Haeussler, M., Schönig, K., Eckert, H., Eschstruth, A., Mianné, J., Renaud, J. B., et al. (2016). Evaluation of off-target and on-target scoring algorithms and integration into the guide RNA selection tool CRISPOR. Genome Biol. 17:148. doi: 10.1186/s13059-016-1012-2
Hartley, J. L., Temple, G. F., and Brasch, M. A. (2000). DNA cloning using in vitro site-specific recombination. Genome Res. 10, 1788–1795. doi: 10.1101/gr.143000
Hood, E. E., Gelvin, S. B., Melchers, L. S., and Hoekema, A. (1993). New Agrobacterium helper plasmids for gene transfer to plants. Transgenic Res. 2, 208–218. doi: 10.1007/BF01977351
James, D. J., Passey, A. J., Barbara, D. J., and Bevan, M. (1989). Genetic transformation of apple (Malus pumila Mill) using a disarmed Ti-binary vector. Plant Cell Rep. 7, 658–661.
Jia, H., Orbovic, V., Jones, J. B., and Wang, N. (2016). Modification of the PthA4 effector binding elements in Type I CsLOB1 promoter using Cas9/sgRNA to produce transgenic Duncan grapefruit alleviating XccΔpthA4:dCsLOB1.3 infection. Plant Biotechnol. J. 14, 1291–1301. doi: 10.1111/pbi.12495
Jia, H., Zhang, Y., Orbovic, V., Zu, J., White, F., Bones, J., et al. (2017). Genome editing of the disease susceptibility gene CsLOB1 in citrus confers resistance to citrus canker. Plant Biotechnol. J. 15, 817–823. doi: 10.1111/pbj.12677
Jia, H. G., and Wang, N. (2014). Targeted genome editing of sweet orange using Cas9/sgRNA. PLoS One 9:e93806. doi: 10.1371/journal.pone.0093806
Kadam, U. S., Shelake, R. M., Chavhan, R. L., and Suprasanna, P. (2018). Concerns regarding ‘off-target’ activity of genome editing endonucleases. Plant Pysiol. Biochem. 131, 22–30. doi: 10.1016/j.plaphy.2018.03.027
Karimi, M., Inzé, D., and Depicker, A. (2002). Gateway vectors for Agrobacterium-mediated plant transformation. Trends Plant Sci. 7, 193–195. doi: 10.1016/S1360-1385(02)02251-3
Kotoda, N., Iwanami, H., Takahashi, S., and Abe, K. (2006). Antisense expression of MdTFL1, a TFL1-like gene, reduces the juvenile phase in apple. J. Am. Soc. Hort. Sci. 131, 74–81.
Leblay, C., Chevreau, E., and Raboin, L. M. (1991). Adventitious shoot regeneration from in vitro leaves of several pear cultivars (Pyrus communis L.). Plant Cell Rep. 25, 99–105.
Li, J. F., Norville, J. E., Aach, J., McCormack, M., Zhang, D., Bush, J., et al. (2013). Multiplex and homologous recombination-mediated genome editing in Arabidopsis and Nicotiana benthamiana using guide RNA and Cas9. Nat. Biotechnol. 31, 688–691.
Ma, X., Zhu, Q., Chen, Y., and Liu, Y. G. (2016). CRISPR/Cas9 platforms for genome editing in plants: developments and applications. Mol. Plant 9, 961–974. doi: 10.1016/j.molp.2016.04.009
Malnoy, M., Viola, R., Jung, M. H., Koo, O. J., Kim, S., Kim, J. S., et al. (2016). DNA-free genetically edited grapevine and apple protoplast using CRISPR/Cas9 ribonucleoproteins. Front. Plant Sci. 7:194. doi: 10.03389/fpls.2016.0194
Malnoy, M. A., Chevreau, E., Bell, R. L., Dolgov, S. V., and Aldwinckle, H. C. (2008a). “Pear,” in Compendium of Transgenic Crop Plants: Transgenic Temperate Fruits and Nuts, eds C. Kole and T. C. Hall (Hoboken, NJ: Blackwell Publishing Ltd.), 53–77. doi: 10.1002/9781405181099.k0402
Malnoy, M. A., Korban, S., Boresjza-Wysocka, E., and Aldwinckle, H. C. (2008b). “Apple,” in Compendium of Transgenic Crop Plants: Transgenic Temperate Fruits and Nuts, eds C. Kole and T. C. Hall (Hoboken, NJ: Blackwell Publishing Ltd.), 1–51. doi: 10.1002/9781405181099.k0401
Marchand, S., Hajdari, P., Hackman, P., Udd, B., and Richard, I. (2003). Touch-down method for high-performance sequencing of polymerase chain reaction products. Anal. Biochem. 315, 270–272. doi: 10.1016/S0003-2697(02)00613-9
Marshallsay, C., Kiss, T., and Filipowicz, W. (1990). Amplification of plant U3 and U6 snRNA gene sequences using primers specific for an upstream promoter element and conserved intragenic regions. Nucleic Acids Res. 18, 3459–3466. doi: 10.1093/nar/18.12.3459
Mimida, N., Kotoda, N., Ueada, T., Igarashi, M., Hatsuyama, Y., Iwanami, H., et al. (2009). Four TFL1/CEN-like genes on distinct linkage groups show different expression patterns to regulate vegetative and reproductive development in apple (Malus x domestica Borkh.). Plant Cell Physiol. 50, 394–412. doi: 10.1093/pcp/pcp001
Mourgues, F., Chevreau, E., Lambert, C., and de Bondt, A. (1996). Efficient Agrobacterium-mediated transformation and recovery of transgenic plants from pear (Pyrus communis L.). Plant Cell Rep. 16, 245–249. doi: 10.1007/BF01890877
Murashige, T., and Skoog, F. (1962). A revised medium for rapid growth and bioassays with tobacco tissue culture. Physiol. Plant. 15, 473–497. doi: 10.1111/j.1399-3054.1962.tb08052.x
Nakajima, I., Ban, Y., Azuma, A., Onoue, N., Moriguchi, T., Yamamoto, T., et al. (2017). CRIPSR/Cas9-mediated targeted mutagenesis in grape. PLoS One 12:e0177966. doi: 10.1371/journal.pone.0177966
Nekrasov, V., Staskawicz, B., Weigel, D., Jones, J. D. G., and Kamoun, S. (2013). Targeted mutagenesis in the model plant Nicotiana benthamiana using Cas9 RNA-guided endonucleases. Nat. Biotechnol. 31, 691–693. doi: 10.1007/s11103-014-0263-0
Nishitani, C., Hirai, N., Komori, S., Wada, M., Okada, K., Osakabe, K., et al. (2016). Efficient genome editing in apple using a CRISPR/Cas9 system. Sci. Rep. 6:31481. doi: 10.1038/srep31481
Peng, A., Chen, S., Lei, T., Xu, L., He, Y., Wu, L., et al. (2017). Engineering canker-resistant plants through CRISPR/Cas9-targeted editing of the susceptibility gene CsLOB1 promoter in citrus. Plant Biotechnol. J. 15, 1509–1519. doi: 10.1111/pbi.12733
Qin, G., Gu, H., Peng, Y., Deng, X. W., Chen, Z., and Qu, L. J. (2007). Disruption of phytoene desaturase gene results in albino and dwarf phenotypes in Arabidopsis by impairing chlorophyll, carotenoid and gibberellin biosynthesis. Cell Res. 17, 471–482. doi: 10.1038/cr.2007.40
Ren, C., Liu, X. J., Zhang, Z., Wang, Y., Duan, W., Li, S. H., et al. (2016). CRISPR/Cas9-mediated efficient targeted mutagenesis in Chardonnay (Vitis vinifera L.). Sci. Rep. 6:32289. doi: 10.1038/srep32289
Sasaki, S., Yamagishi, N., and Yoshikawa, N. (2011). Efficient virus-induced gene silencing in apple, pear and Japanese pear using Apple latent spherical virus vectors. Plant Methods 7:15. doi: 10.1186/1746-4811-7-15
Shannon, S., and Meeks-Wagner, D. R. (1991). A mutation in the Arabidopsis TFL1 gene affects inflorescence meristem development. Plant Cell 3, 877–892. doi: 10.1105/tpc.3.9.877
van der Fits, L., Deakin, E. A., Hoge, J. H., and Memelink, J. (2000). The ternary transformation system: constitutive virG on a compatible plasmid dramatically increases Agrobacterium-mediated plant transformation. Plant Mol. Biol. 43, 495–502. doi: 10.1023/A:1006440221718
Velasco, R., Zharkikh, A., Affourtit, J., Dhingra, A., Cestaro, A., Kalyanaraman, A., et al. (2010). The genome of the domesticated apple (Malus × domestica Borkh.). Nat. Genet. 42, 833–839. doi: 10.1038/ng.654
Voinnet, O., Rivas, S., Mestre, P., and Baulcombe, D. (2003). An enhanced transient expression system in plants based on suppression of gene silencing by the p19 protein of tomato bushy stunt virus. Plant J. 33, 949–956. doi: 10.1046/j.1365-313X.2003.01676.x
Wang, X., Tu, M., Wang, D., Liu, J., Li, Y., Li, Z., et al. (2018). CRIPSR/Cas9-mediated efficient targeted mutagenesis in grape in the first generation. Plant Biotechnol. J. 16, 844–855. doi: 10.1111/pbi.12832
Wang, Z., Wang, S., Li, D., Zhang, Q., Li, L., Zhong, C., et al. (2018). Optimized paired-sgRNA/Cas9 cloning and expression cassette triggers high-efficiency multiplex genome editing in kiwifruit. Plant Biotechnol. J. 16, 1424–1433. doi: 10.1111/pbi.12884
Weigl, K., Flachowsky, H., Peil, A., and Hanke, M. V. (2015). Heat mediated silencing of MdTFL1 genes in apple. Plant Cell Tis. Org. Cult. 123, 511–521. doi: 10.1007/s11240-015-0855-7
Keywords: apple, pear, gene editing, knock-out, CRISPR, PDS, TFL1
Citation: Charrier A, Vergne E, Dousset N, Richer A, Petiteau A and Chevreau E (2019) Efficient Targeted Mutagenesis in Apple and First Time Edition of Pear Using the CRISPR-Cas9 System. Front. Plant Sci. 10:40. doi: 10.3389/fpls.2019.00040
Received: 22 October 2018; Accepted: 11 January 2019;
Published: 06 February 2019.
Edited by:
Vladimir Orbovic, University of Florida, United StatesReviewed by:
Vinay Kumar, Central University of Punjab, IndiaKaijun Zhao, Institute of Crop Sciences (CAAS), China
Copyright © 2019 Charrier, Vergne, Dousset, Richer, Petiteau and Chevreau. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Elisabeth Chevreau, ZWxpc2FiZXRoLmNoZXZyZWF1QGlucmEuZnI=
†These authors have contributed equally to this work