 Xin Yuan
Xin Yuan Zhangying Wang
Zhangying Wang Jianzhong Huang
Jianzhong Huang Zhiyong Gao
Zhiyong Gao- State Key Laboratory of Hybrid Rice, Key Laboratory for Research and Utilization of Heterosis in Indica Rice of Ministry of Agriculture, College of Life Sciences, Wuhan University, Wuhan, China
RPM1 is a plant immune receptor that specially recognizes pathogen-released effectors to activate effector-triggered immunity (ETI) in Arabidopsis thaliana. RPM1 triggers ETI and hypersensitive response (HR) for disease resistance. Previous reports indicated that Phospholipase D (PLD) positively regulated RPM1-mediated HR. However, single, double, and triple pld knock-out mutants of 12 members of the PLD family in A. thaliana did not show suppressed RPM1-mediated HR, indicating the functional redundancy among PLD members. In this study, we revealed that PLD could negatively regulate the function of RPM1. We found that RPM1 interacted with PLDδ, but did not interact with PLDβ1, PLDβ2, and PLDγ3. Overexpression of PLDδ conducted to a reduction of protein level and corresponding activity of RPM1. We found that abscisic acid (ABA) reduced the protein level of RPM1, and the ABA-induced RPM1 reduction required PLD activity and PLD-derived phosphatidic acid (PA). Our study shows that PLD plays both negative and positive roles regulating the protein level and activity of RPM1 during stress responses in plants. PLD proteins are regulating points to integrate the abiotic and biotic responses of plants.
Introduction
Plants develop innate immunity systems to confront pathogenic invasions (Jones and Dangl, 2006). Plants have immunity receptors to recognize pathogen-associated molecular patterns (PAMPs), and stimulate PAMP triggered immunity (PTI) (Couto and Zipfel, 2016). Pathogens evolve effectors to suppress plant PTI, and plants evolve corresponding receptors to recognize effectors, and stimulate effector triggered immunity (ETI) (Cui et al., 2015). Most of the ETI receptors are characterized with a nucleotide-binding domain (NB) and a leucine-rich-repeat domain (LRR), and these NLRs stimulate strong and rapid defense responses, named hypersensitive responses (HR), and results in cell deaths at the infection sites (Baggs et al., 2017).
RPM1 is an NLR receptor in Arabidopsis thaliana (Grant et al., 1995). It does not directly bind its corresponding effector AvrRpm1 or AvrB, but is activated by perceiving the effector-induced phosphorylation of the guarded protein RIN4 (Mackey et al., 2002; Chung et al., 2011; Liu et al., 2011). RPM1 is a plasma-membrane associated protein, and it does not have any transmembrane domain (Boyes et al., 1998; Gao et al., 2011). RIN4, a plasma-membrane localized protein, can stabilize RPM1, and only trace amounts of RPM1 are detected in the rin4 knockout plant. However, the trace amounts of RPM1 are still plasma-membrane localized (Gao et al., 2011). The activation of RPM1 leads to downstream signal transductions, including the activation of phospholipase C (PLC), the influx of extracelluar Ca2+, the activation of phospholipase D (PLD), and the accumulation of reactive oxygen species (ROS) (Andersson et al., 2006). The calcium-channel blocker LaCl3 and inhibition the activity of PLC proteins and PLD proteins are able to suppress RPM1-induced HR (Grant et al., 2000; Andersson et al., 2006).
The PLD proteins are a family of enzymes that hydrolyze membrane phospholipids, such as phosphoatidyl choline (PC) and phosphoatidylethanolamine (PE), to produce phosphatidic acid (PA) and a free-head alcohol. PLD catalyzes the reaction of transferring the phosphoatidyl group to primary alcohols to form phosphatidylalcohols instead of PA (Ella et al., 1997). Thus, n-butanol can be used to suppress PLD-derived PA. The A. thaliana genome contains 12 PLD genes which are grouped into α, β, γ, δ, ε, and ζ six types. Each PLD has different properties in activity regulation and/or lipid preferences (Li et al., 2009). PLDs play essential roles in responses to various abiotic and biotic stresses (Bargmann and Munnik, 2006). PLDα regulates plant response on drought and salt stresses (Sang et al., 2001; Hong et al., 2008). PLDα1 mediates abscisic acid (ABA) signaling to control the stomata closure. ABA activates PLDα1 and results in the production of PA. PLDα1-derived PA binds to ABI1, a negative regulator of ABA signaling, and the binding suppresses the negative effects of ABI1 and results in the stomatal closure. Meanwhile, PLDα1 and PA interact with the Gα subunit of heterotrimeric G protein to mediate ABA inhibition of stomatal opening (Mishra et al., 2006). PLDβ1 mediates negative defense to bacterial and fungal pathogens (Zhao et al., 2013). PLDγ1 is involved in Aluminum tolerance (Zhao et al., 2011). Plasma-membrane associated PLDδ binds to microtubules and negatively regulates thermotolerance by means of microtubule disorganization (Zhang et al., 2017). PLDδ also involves in the cell wall based defense against non-host powdery mildew fungi (Pinosa et al., 2013). PLDε regulates root growth responding to Nitrogen availability (Hong et al., 2009). PLDζ involves in phosphate deficiency and salt stresses (Li et al., 2006; Ben Othman et al., 2017).
RPM1-mediated HR can be suppressed with n-butanol, an antagonist for PLD-derived PA, suggesting PLD and PLD-derived PA are required for HR response (Andersson et al., 2006). Based on this result, it is reasonable to hypothesize that some pld-KO mutants should display suppressed HR response. The single, double, and triple pld-KO mutants of the 12 PLD members were assayed for RPM1-mediated HR, and none of the mutants showed obviously deficient HR. The results were explained with the redundancy among PLD members (Johansson et al., 2014). In this study, we revealed that PLD can negatively regulate the function of RPM1. We found that RPM1 interacted with PLDδ, but did not interact with PLDβ1, PLDβ2, and PLDγ3. Overexpression of PLDδ obviously conducted a reduction of protein level and corresponding activity of RPM1. We found that abscisic acid (ABA) reduced the protein level of RPM1, and the ABA-induced RPM1 reduction required PLD activity and PLD-derived phosphatidic acid (PA). Thus, the PLD activity is not only a consequence of RPM1 signal transduction, but also a negative regulator for the function of RPM1.
Materials and Methods
Plant Growth and Mutants
Arabidopsis thaliana and Nicotiana benthamiana plants were grown in pots with autoclaved vermiculite and watered with Hoagland solution. The growth condition is at 24°C under a 16 h light/8 h dark cycle. A. thaliana plants for HR and disease resistance assays were grown under 8 h light/16 h dark cycle condition. All the lines of A. thaliana are in Columbia-0 background. The pldδ mutant 12B (SALK_023247C) were obtained from ABRC. The mutant line rpm1-3 and the transgenic line pRPM1::RPM1-Myc/rpm1-3 (AT5) were gifts from Dr. Jeff. Dangl (University of North Carolina at Chapel Hill, Chapel Hill, NC, United States).
Vector Construction
The gateway system was used to construct vectors. For transient expression, The CDSs of AtPLD genes were cloned into the expression vector pEarleyGate 101 containing the constitutive high-expression CaMV 35S promoter, and YFP-HA tag (Karimi et al., 2007). HA tag was used for protein detection. RPM1-Myc was cloned into the pGWB2 vector to obtain the 35S::RPM1-Myc expression construct (Nakagawa et al., 2007). RPM1(D505V)-Myc was cloned into the pMDC7 vector under the control of the estradiol-inducible promoter to obtain the Est:: RPM1(D505V)-Myc expression construct (Karimi et al., 2007). For the bimolecular fluorescence complementation (BiFC) assay, we modified the pEarleyGate 101 vector into vectors that could express the two complementary parts of YFP, the N terminus (nYFP) and the C terminus (cYFP) (Walter et al., 2004; Citovsky et al., 2006). The expression constructs of 35S:: RPM1 -nYFP-HA and 35S:: PLDδ-cYFP-HA were made respectively.
Transient Expression in the Leaves of N. benthamiana
The expression constructs were electro-transformed into Agrobacterium tumefaciens GV3101. Agrobacteria were cultured overnight at 28°C with suitable antibiotics. Overnight-grown Agrobacteria were centrifuged and re-suspended in the induction buffer (10 mM MES pH 5.6, 10 mM MgCl2, 150 μM acetosyringone). Samples were infiltrated into the leaves of 5-week-old N. benthamiana with a 1-mL needless syringe at desired OD600 nm values. Agrobacteria containing the P19 plasmid was always co-infiltrated with the samples at an OD600 nm of 0.2. The transiently expressed proteins were extracted and analyzed at 48 h after infiltration (Pitino et al., 2016).
Protein Extraction and Co-immunoprecipitation (Co-IP) Assay
For protein expression, three leaf disks (7 mm diameter) were ground with 110 μL of the protein extraction buffer (20 mM Tris-HCl PH 8.0, 5 mM EDTA, 10 mM DTT, 1% SDS). The supernatants were collected with centrifugation at 8,000 × g for 3 min and mixed with fourfold protein loading buffer. Actin was used as an internal protein loading control. For the fractioning experiments, leaf tissues were ground in a mortar with the extraction buffer (20 mM Tris-HCl pH 8.0, 150 mM NaCl, 20% glycerol, 1 mM DTT, 10 mM EDTA, 1 mM PMSF, protease inhibitor cocktail). Samples were centrifuged at 6,000 × g for 10 min at 4°C to remove the debris. The supernatant containing total proteins (T) was separated into the cytosolic fraction (S) and the microsomal fraction (M) with centrifugation at 20,000 × g for 60 min at 4°C. The plasma-membrane localized H+-ATPase was used as the microsomal marker. The big subunit of Rubisco was stained with Ponceau S as the cytosolic marker.
For Co-IP experiment, because PLDδ and RPM1 are membrane associated proteins, the microsomal fractions were used for Co-IP. The protein content of RPM1-Myc in the microsomal fractions were determined with Western blots, and the microsomal fractions were re-suspended with the suspension buffer (40 mM Tris-HCl PH 7.5, 150 mM NaCl, 1 mM DTT, 5 mM EDTA, protease inhibitor cocktail, 1% Triton X-100) to equalize RPM1-Myc protein levels between the control and test samples. The samples were rotated at 4°C for 1 h and centrifuged at 20,000 × g for 1 h to remove the insoluble proteins. 30 μL of the anti-GFP conjugated agarose beads (MBL, #D153-8) were added into 1 mL of the supernatants and incubated at 4°C for 3 h. Beads were washed three times with the suspension buffer. Immunoprecipitates were eluted with 50 μL of the elution buffer (2% SDS) at 37°C for 5 min.
The primary antibodies used for Western blots were the anti-Myc (GeneScript, #A00704), the anti-HA (Roche, #11867423001), the anti-H+-ATPase (Agrisera, #AS07260-100), the anti-β-Actin (Abbkine, #A01050-2), and the anti-T7 (Novagen, #69522).
Confocal Microscopy
The nYFP-tagged proteins and the cYFP-tagged proteins were co-expressed in the leaves of N. benthamiana. The fluorescent images were observed using confocal microscopy according to the BiFC protocol (Walter et al., 2004). YFP was excited at 488 nm and fluorescent emissions were observed at 518–540 nm.
Disease Resistance and Ion-Leakage Assay
Pseudomonas syringae pv. tomato strain DC3000(avrB) was grown on King’s B medium with antibiotics at 28°C for 36 h. For spray inoculation, leaves from 5-week-old plants were sprayed with Pto DC3000(avrB) in 10 mM MgCl2 with 0.025% silwet L-77 at an OD600 of 0.05. Inoculated plants were covered with a plastic dome for 2 days. All experiments were repeated three times. For ion-leakage assay, five leaf disks (8 mm diameter) of A. thaliana or N. benthamiana were floated in a clean wild-mouth tube with 6 ml of distilled water. The conductivity of the samples was measured at the indicated time points with a conductivity meter (Hubert et al., 2009).
Results
RPM1 Interacts With PLDδ in planta
Phospholipase D mediates the signal transduction of RPM1 (Andersson et al., 2006). We cloned and transiently expressed all the members of AtPLD family in the leaves of N. benthamiana to determine their interactions with RPM1. Of the 12 PLD members, PLDδ, PLDα2, PLDβ1, PLDβ2, and PLDγ3 had detectable protein expression, and were used for Co-IP assay. We co-expressed 35S::PLD-YFP-HA and 35S::RPM1-Myc in the leaves of N. benthamiana, and immunoprecipitated PLD-YFP-HA with the anti-GFP antibody to determine the Co-IP of RPM1-Myc. Co-expression of 35S::YFP-HA and 35S::RPM1-Myc was used as the negative control. The Co-IP assay showed that PLDα2 and PLDδ interacted with RPM1, while PLDβ1, PLDβ2, and PLDγ3 did not (Table 1). PLDδ was used for subsequent experiments due to stronger interaction with RPM1 (Figure 1A). We validated the interaction with the BiFC assay. The Yellow fluorescence protein (YFP) was split into two complementary parts, the N terminus (nYFP) and the C terminus (cYFP) (Walter et al., 2004; Citovsky et al., 2006). The expression constructs 35S:: RPM-nYFP-HA and 35S:: PLDδ -cYFP-HA were co-expressed in the leaves of N. benthamiana. The results showed clear yellow fluorescence in the samples co-expressing RPM1-nYFP-HA and PLDδ-cYFP-HA, but no fluorescence was detected in the negative controls (Figure 1B). We transformed 35S::PLDδ-YFP-HA into the pRPM1::RPM1-Myc/rpm1-3 plant (AT5) to determine the interaction of RPM1 and PLDδ in A. thaliana. The Co-IP of RPM1-Myc with PLDδ-YFP-HA in the transgenic plants indicated the interaction of RPM1 and PLDδ in A. thaliana (Figure 1C).
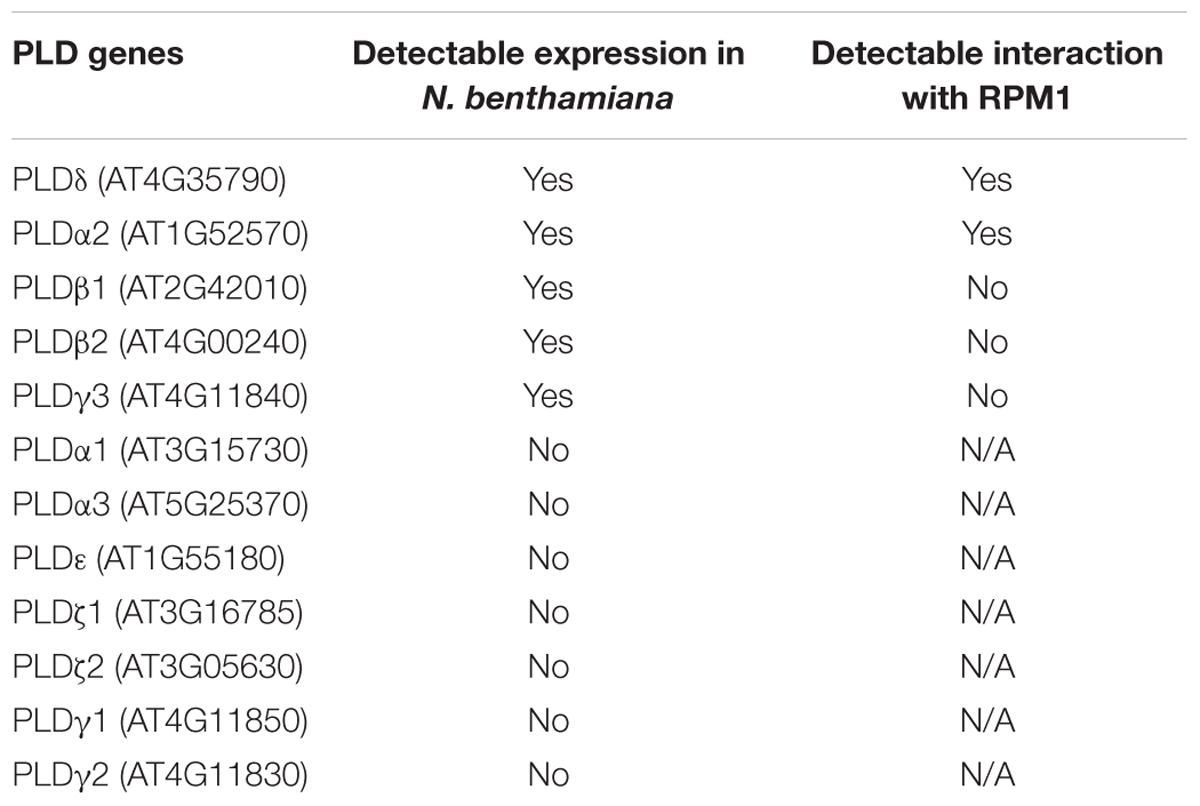
Table 1. Phospholipase D (PLD) genes and their protein expressions in N. benthamiana.
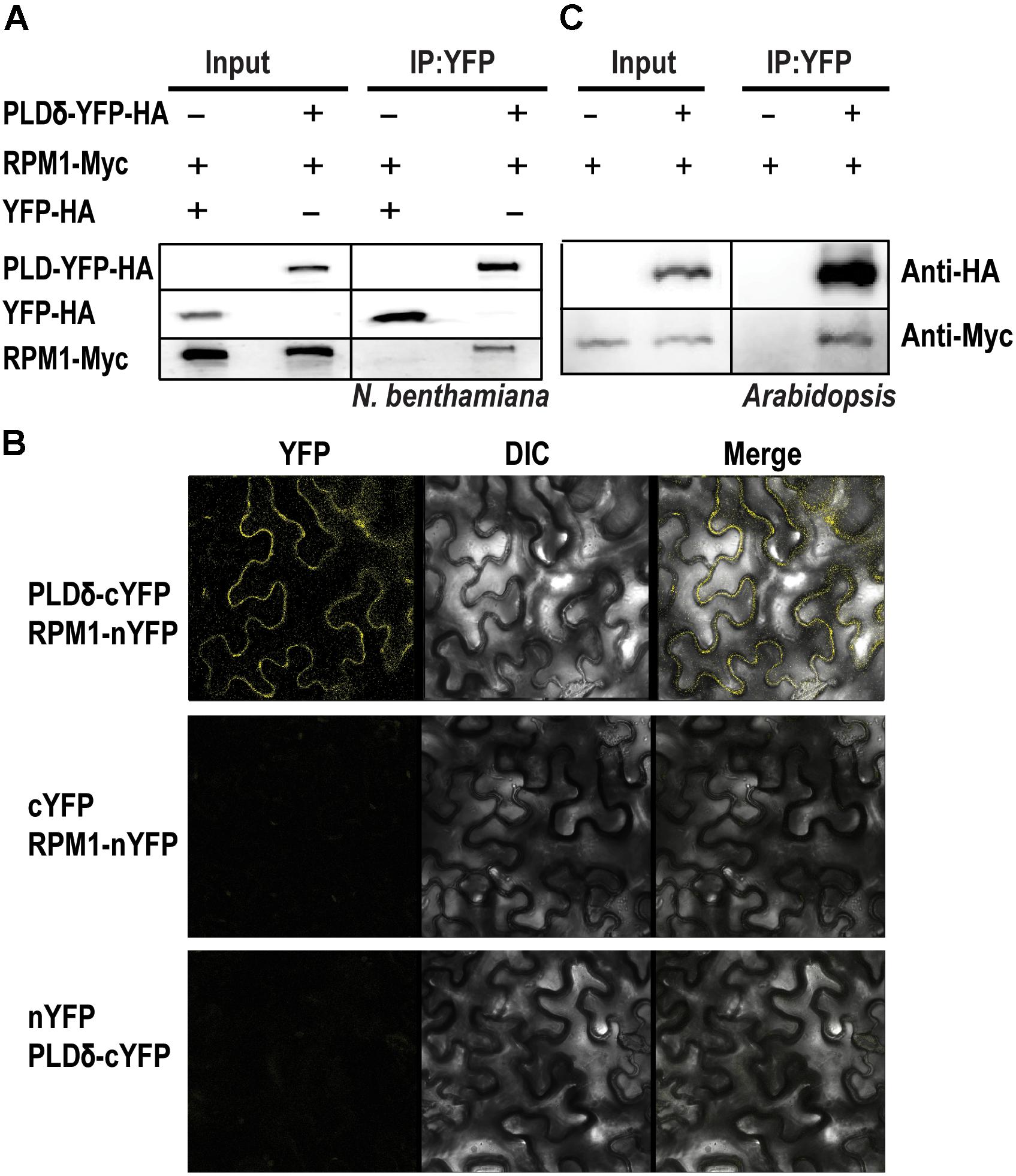
Figure 1. PLDδ interacts with RPM1. (A) The interaction of PLDδ and RPM1 in Nicotiana benthamiana. 35S::PLDδ-YFP-HA and 35S::RPM1-Myc were co-expressed in the leaves of N. benthamiana. Total membrane proteins were extracted from samples collected at 2 days after infiltration. PLDδ-YFP-HA was immunoprecipitated with agarose beads conjugated with anti-GFP antibody, and RPM1-Myc was detected for Co-IP. Co-expression of 35S::YFP-HA and 35S::RPM1-Myc was used as the negative control. (B) Representative confocal images of BiFC. BiFC analysis was performed in N. benthamiana. PLDδ was fused with the C-terminal portion of YFP (cYFP), and RPM1 was fused to the N-terminal portion of YFP (nYFP). Different plasmids were co-expressed in N. benthamiana. Images were pictured at 2.5 days after infiltration. All the experiments were repeated three times with similar results. (C) The interaction of PLDδ and RPM1 in Arabidopsis thaliana. Co-IP assay was performed with stable transgenic plants containing 35S::PLDδ-YFP-HA and pRPM1::RPM1-Myc. The transgenic plants containing pRPM1::RPM1-Myc was used as the negative control.
PLDδ Suppresses RPM1(D505V)-Induced Cell Death in N. benthamiana
Since PLDδ interacted with RPM1, we tested whether PLDδ affected the function of RPM1. We firstly determined whether PLDδ could affect RPM1-induced HR. RPM1(D505V), which mimics the active state of RPM1, is sufficient to induce the cell death (Gao et al., 2011). The estradiol inducible Est::RPM1(D505V)-Myc construct was co-expressed with 35S::PLDδ-YFP-HA or 35S::YFP-HA in the leaves of N. benthamiana to compare the occurrences of the cell death induced by RPM1(D505V). The cell death could be stained with trypan blue or be quantified with ion-leakage assay. Results showed that PLDδ-YFP-HA could obviously suppress RPM1(D505V) induced cell death (Figures 2A,B). We hypothesized that PLDδ either suppressed the activation of RPM1, or reduced the protein level of RPM1 to affect its function.
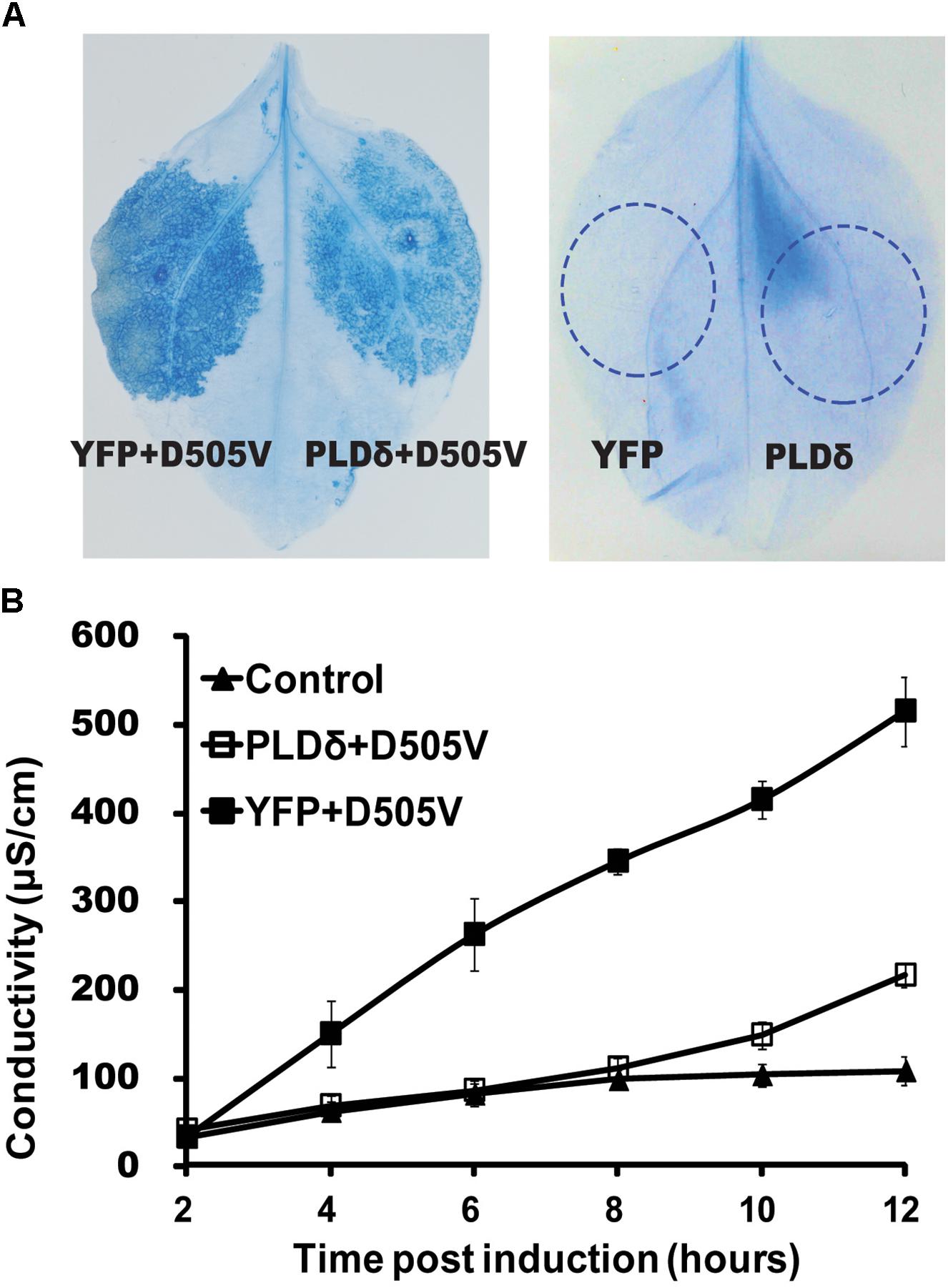
Figure 2. PLDδ suppresses RPM1(D505V) induced cell death in N. benthamiana. (A) Trypan blue staining of the infiltrated leaves of N. benthamiana. 35S::PLDδ-YFP-HA or 35S::YFP-HA were co-expressed with Est::RPM1(D505V)-Myc. RPM1(D505V) was induced with 20 μM estradiol at 2 days after infiltration and the leaves were stained at 6 h after induction. Leaves transiently expressed 35S::PLDδ-YFP-HA or 35S::YFP-HA were stained at 2 days after infiltration. (B) Ion-leakage assay. Leaf disks were collected from estradiol induced leaves and floated in distilled water. The conductivity of the water was measured at indicated time points. Each assay was performed with three duplicates. The error bars represent 2X standard deviation (SD). Leaves transiently expressed 35S::YFP-HA were used as control.
PLDδ Reduces the Protein Level of RPM1(D505V) in N. benthamiana
We tested whether PLDδ could affect the protein level of RPM1(D505V) in the leaves of N. benthamiana. RPM1(D505V) is unstable due to the cell death induced by the protein, and it is hard to accumulate detectable protein level. Lanthanum chloride (LaCl3) can inhibit RPM-mediated HR, and prevent the degradation of the activated RPM1 or RPM1(D505V) (Grant et al., 2000; El Kasmi et al., 2017). We co-expressed Est::RPM1(D505V)-Myc and 35S::PLDδ-YFP-HA or 35S::YFP-HA in the leaves of N. benthamiana, and induced the expression of RPM1(D505V) with estradiol at 2 days after infiltration. The leaves were treated with 4 mM of LaCl3 at the same time of the induction. The protein levels of RPM1(D50V) were detected at 3, 5, and 8 h after induction. Our results showed that PLDδ-YFP-HA obviously reduced the protein level of RPM1(D505V) compared with the negative control YFP-HA (Figure 3A).
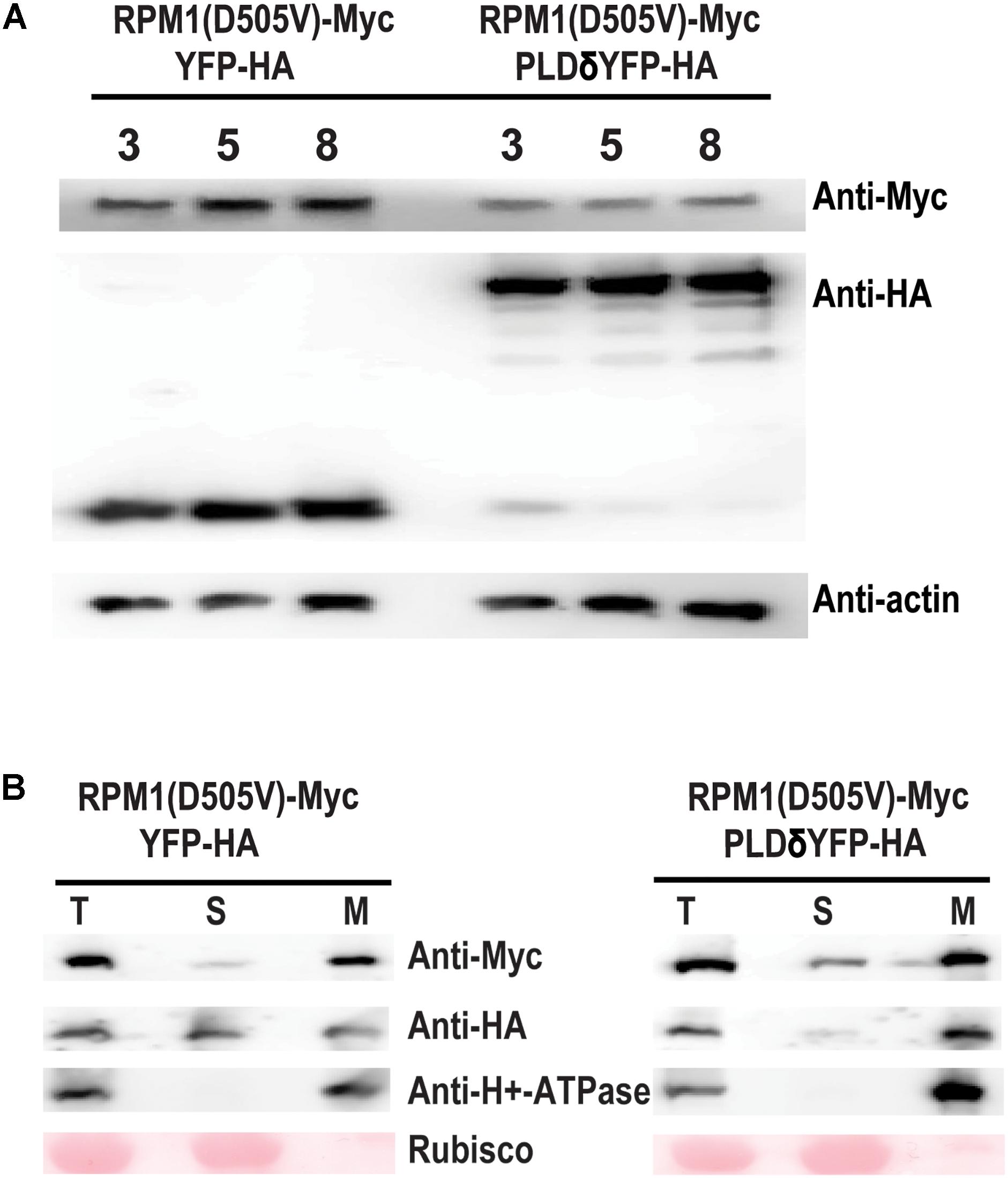
Figure 3. PLDδ reduces the protein level of RPM1(D505V) in N. benthamiana. (A) Protein level of RPM1(D505V). Est::RPM1(D505V)-Myc and 35S::PLDδ-YFP-HA were co-expressed in the leaves of N. benthamiana. RPM1(D505V) was induced with estradiol at 2 days after infiltration and the protein levels of RPM1(D505V)-Myc were detected at 3, 5, and 8 h after induction. 4 mM of LaCl3 was infiltrated into the leaves at 2 h before induction to block RPM1(D505V) induced cell death. Co-expression of YFP-HA and RPM1(D505V) was used as a control. Actin was used as a marker for protein equal loading. (B) PLDδ affects the membrane association of RPM1(D505V) in N. benthamiana. Samples were collected at 5 h after induction. The protein extraction solution (T) was separated into the cytosolic fraction (S) and the microsomal fraction (M) using centrifugation. The distribution of RPM1(D505V)-Myc in the two fractions were detected. The large subunit of Rubisco was used as a cytosolic protein marker. The plasma-membrane protein H+-ATPase was used as a microsomal protein marker. The RPM1(D505V)-Myc images were adjusted to the similar density to compare RPM1(D505V) levels distributed in the S fraction. The original western blots are available in Supplementary Materials.
RPM1(D505V) is a plasma-membrane associated protein. We collected the samples at 5 h after induction, and separated total proteins into the soluble and the microsomal membrane fractions using ultracentrifugation. Certain amount of RPM1(D505V) participated in the soluble fraction of the sample co-expressed with PLDδ, while less amount of RPM1(D505V) participated in the soluble fraction of the sample co-expressed with the negative control YFP-HA (Figure 3B).
PLDδ Activity Is Required to Negatively Regulate the Function of RPM1(D505V) in N. benthamiana
PLDδ is an oleate and phosphatidylinositol 4,5-bisphosphate (PIP2) activated enzyme. PLDδ(R410D) and PLDδ(R622P) are two activity-deficient mutants due to loss oleate binding or PIP2 stimulated activity respectively (Wang and Wang, 2001). We made PLDδ(R410D/R622P) double mutant to determine whether PLD activity was required to affect the function of RPM1(D505V). Contrast to PLDδ, the activity deficient mutant did not affect the autoactivity and the protein level of RPM1(D505V) (Figures 4A,B).
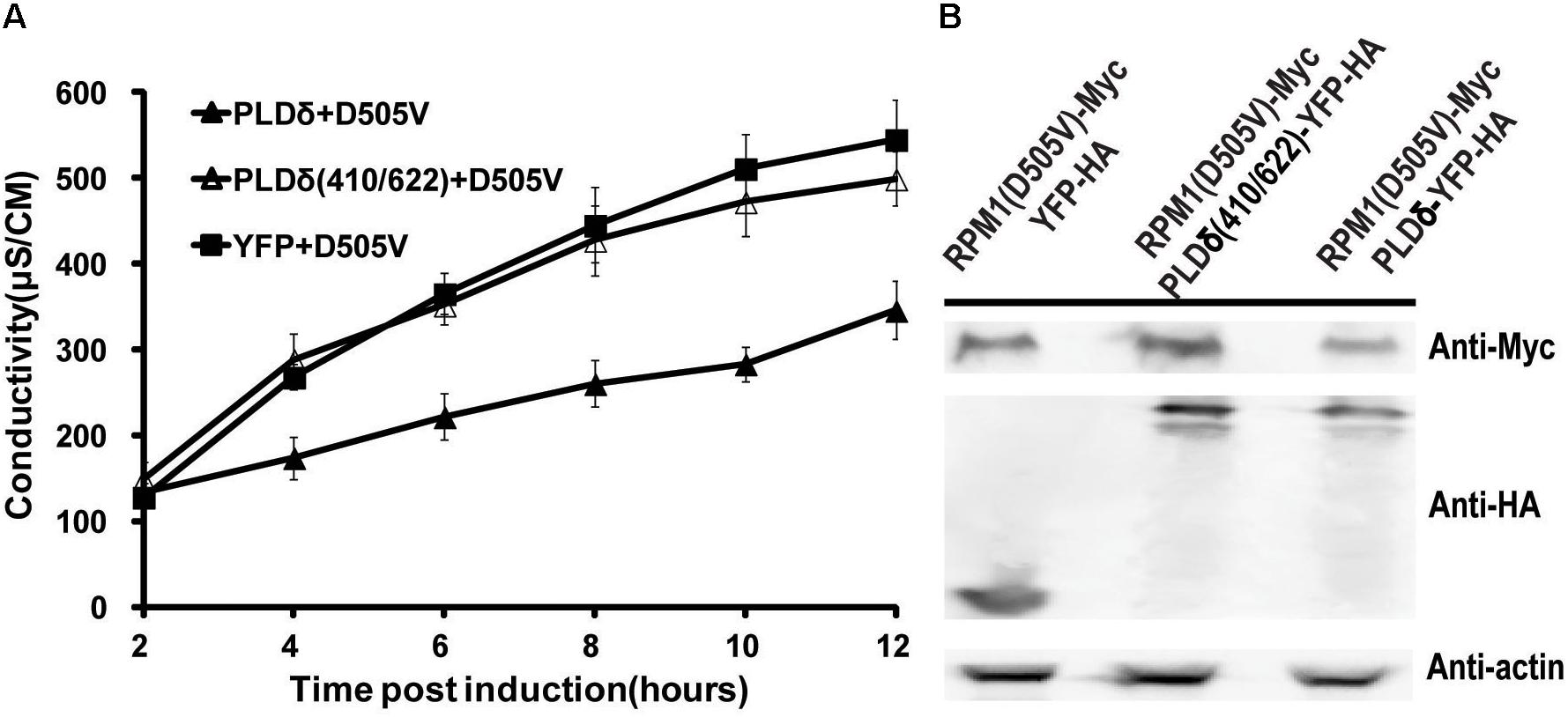
Figure 4. PLDδ activity is required to negatively regulate the function and the protein level of RPM1(D505V) in N. benthamiana. (A) PLDδ(R410D/R622P) is deficient to suppress the autoactivity of RPM1(D505V). Est::RPM1(D505V)-Myc was co-expressed with 35S::PLDδ-YFP-HA, 35S::PLDδ(R410D/R622P)-YFP-HA, or 35S::YFP-HA in the leaves of N. benthamiana. RPM1(D505V) was induced with estradiol at 2 days after infiltration. The autoactivity of RPM1(D505V) in the samples was measured with ion-leakage. (B) PLDδ(R410D/R622P) is deficient to reduce the protein level of RPM1(D505V). Samples were collected at 5 h after induction. The protein levels of RPM1(D505V)-Myc, PLDδ(R410D/R622P)-YFP-HA, and PLDδ-YFP-HA were detected. Actin was used as a marker for protein equal loading.
Overexpression of PLDδ Negatively Affects the Function of RPM1 in A. thaliana
We transformed 35S::PLDδ-YFP-HA into AT5 plant to determine the effects of PLDδ on RPM1 in A. thaliana. Three independent transgenic lines PLD-8, PLD-9, and PLD-21 were used to assay the protein level of RPM1. The results showed that the protein levels of RPM1 were lower in the PLDδ transgenic plants (Figure 5A). According to our assay, PLDγ3 did not interact with RPM1 in N. benthamiana. We transformed 35S::PLDγ3-YFP-HA into AT5 plant to determine the protein levels of RPM1 in three independent PLDγ3-OE plants. The results showed that the protein levels of RPM1 in the transgenic plants were the same as that in AT5 (Figure 5B). The results suggested that the interaction of PLD with RPM1 was necessary to reduce the protein level of RPM1.
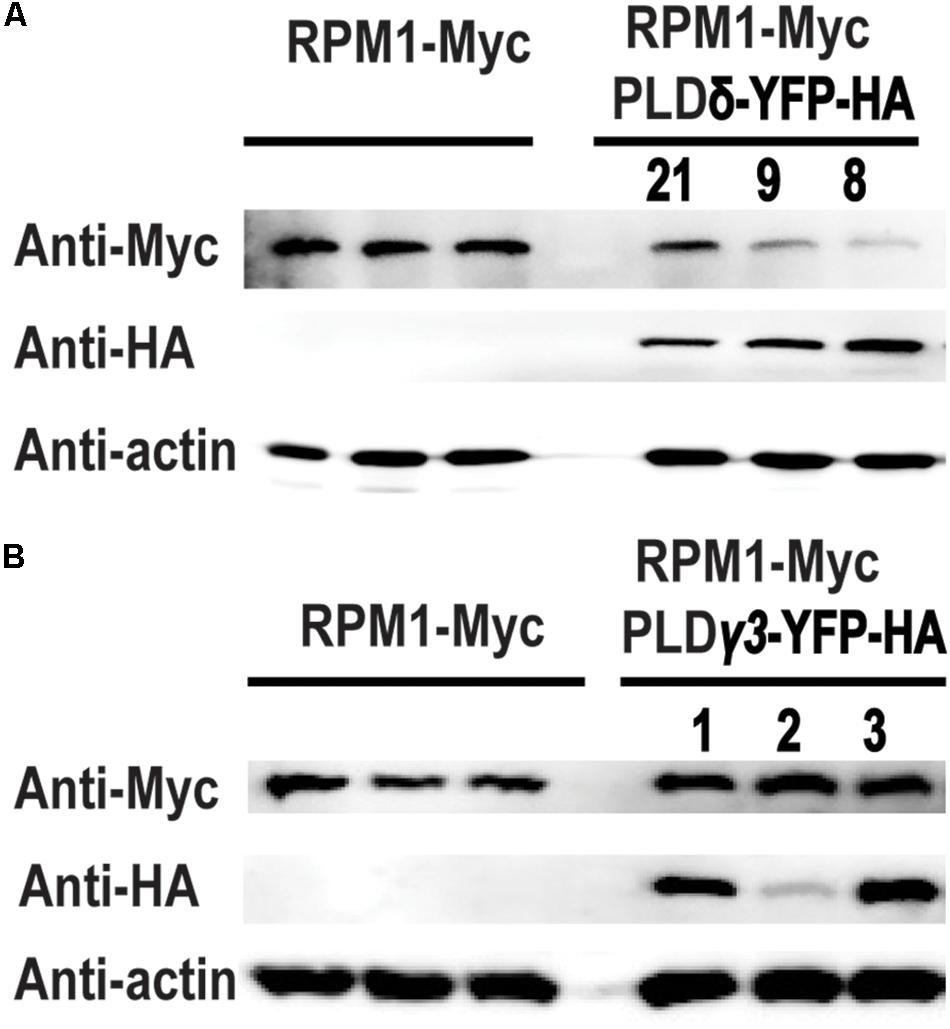
Figure 5. Overexpression of PLDδ affects the protein level of RPM1 in A. thaliana. (A) The protein level of RPM1 in the PLDδ-OE plants. Protein samples were collected from 5-week-old plants. Protein levels of RPM1-Myc in three independent PLDδ OE transgenic plants (PLD8, PLD9, and PLD21) and their background plant pRPM1::RPM1-Myc/rpm1 were detected. (B) The protein level of RPM1 in the PLDγ3-OE plants. The original western blots are available in Supplementary Materials.
Since the protein level of RPM1 reduced in the PLDδ-OE plants, we reasoned that the function of RPM1 should be deficient in the PLDδ-OE plants. We compared the functions of RPM1 in PLD-8 and PLD-9 with those in AT5 plant. The loss-of-function mutant rpm1-3 was used as a negative control, and the pldδ-KO mutant (SALK_023247C) was included for the assays. RPM1 can be activated by its corresponding effector AvrB, and thus restrict the growth of the avirulent pathogen Pto DC3000(avrB). We spray-inoculated the leaves of the plants with Pto DC3000(avrB), and counted bacteria numbers at 0 and 3 days after inoculation. The result indicated that PLD-8 and PLD-9 plants displayed obviously less disease resistance than AT5 (Figures 6A,B). The pldδ-KO mutant displayed the same bacteria growth restriction as AT5, which was consistent with previous report. We quantified the RPM1-mediated HR by measuring the ion-leakage of the plants. The ion-leakage data showed that the PLD-8 and PLD-9 had weaker HR than AT5 (Figure 6C). Based on above results, we concluded that overexpressing of PLDδ reduced the protein level of RPM1, and thus negatively affected the function of RPM1.
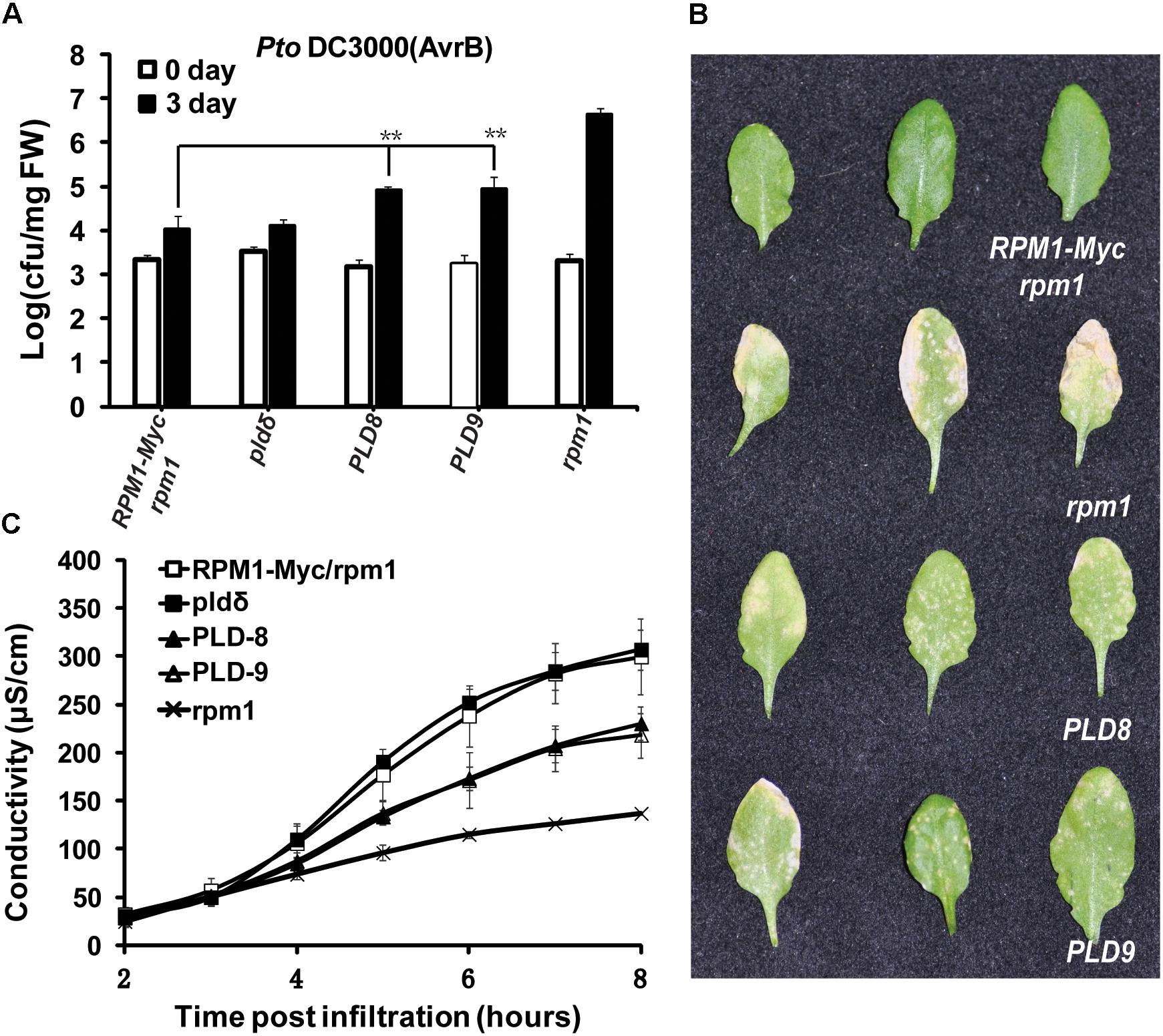
Figure 6. Overexpression of PLDδ suppresses RPM1-mediated disease resistance in A. thaliana. (A) Bacteria growth assay. Pto DC3000(avrB) was spray-inoculated to plant leaves at an OD600 of 0.05. Bacteria numbers were measured at 0 and 3 days after inoculation. The assay was performed with four duplicates. Error bars represent 2X SD. The significant difference (P < 0.01) is marked as ∗∗. PLD8 and PLD9 are two independent PLDδ-OE transgenic plants. (B) Symptoms of the inoculated leaves. Pictures were taken at 5 days after inoculation. (C) Ion-leakage assay. DC3000(avrB) was infiltrated into plant leaves at an OD600 of 0.05. Leaf disks were collected at 1 h after infiltration.
ABA-Induced PLD Activity Reduces the Protein Level of RPM1
Because PLD is enzyme, we want to determine whether PLD activity is required to affect the protein level of RPM1. ABA was able to stimulate the activity of PLD (Jacob et al., 1999), so we treated the leaves of AT5 with or without 100 μM of ABA, and compared the protein levels of RPM1 between the samples. The protein levels of RPM1 assayed at 3 h after treatments, and the results showed that ABA treatment obviously reduced the protein level of RPM1 (Figure 7A). Since PLD activity leads to the synthesis of PA, we treated the plants with n-butanol to block the synthesis of PA and thus the PLD enzymatic function. When the plants were treated with both n-butanol and ABA at the same time, ABA-induced RPM1 reduction was abolished (Figure 7B). The results indicated that PLD activity and its derived PA were required to mediate the ABA-induced RPM1 reduction.
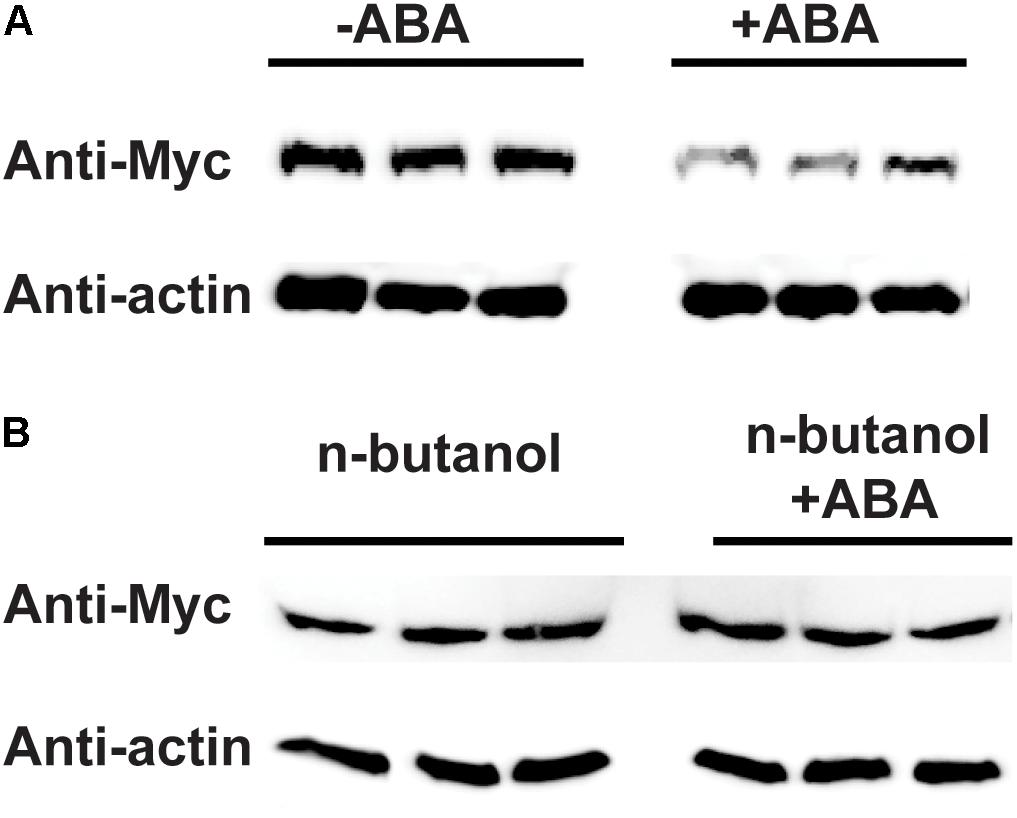
Figure 7. Abscisic acid (ABA)-induced PLD activity negatively regulates the protein level of RPM1. (A) ABA treatment reduces the protein level of RPM1. The leaves of pRPM1::RPM1-Myc/rpm1plants were incubated with or without 100 μM of ABA. Protein samples were collected at 3 h after ABA treatment. Three duplicates were detected. (B) n-butanol suppresses ABA-induced RPM1reduction. Plant leaves were incubated with 100 μM of ABA and 1% n-butanol. The leaves were incubated with 1% n-butanol were used as the negative control. The protein levels of RPM1-Myc were compared between the two treatments. Three duplicates were detected. The original western blots are available in Supplementary Materials.
Discussion
We revealed that PLD could negatively regulate the function of RPM1 (Figure 6), but this did not conflict with previous finding that PLD positively regulated RPM1-mediated HR. As an immune receptor, RPM1 has pro-signaling inactive state and active state. RPM1 is activated by its cognate effectors. We detected the protein levels of RPM1 in PLDδ-OE plants and ABA-treated plants without the inoculation of pathogenic bacteria. The results reflect the protein levels of the inactive RPM1. Since the gene expression of RPM1 is not induced by pathogens, the protein level of the inactive pro-signaling RPM1 determines the output of the RPM1-initiated signaling (Adams-Phillips et al., 2008). PLD negatively regulates the function of RPM1 by reducing the protein level of the pro-signaling inactive RPM1. Previous report indicated that PLD played a positive role on the function of RPM1 (Andersson et al., 2006). PLD activity and its derived PA are downstream components in the RPM1 signal transduction pathway, and their positive roles are effective after the activation of RPM1. PLD activity induced by RPM1 activation may also negatively regulate the protein level and function of RPM1, but active RPM1 is quickly degraded accompanying with the HR response (Boyes et al., 1998), indicating the potential negative regulation of active RPM1 by PLD activity is not necessary.
The exact mechanism on how PLD reduces the protein level of RPM1 is not clear. Overexpression of PLDδ but not PLDγ3, which did not interact with RPM1, reduced the protein level of RPM1 in A. thaliana, suggesting the PLD-RPM1 interaction is required (Figure 5). Our study indicates that PLD activity is also required to reduce RPM1 level. We found that the protein level of RPM1 obviously reduced at 3 hours after ABA treatment, and PLD activity is required for ABA-induced RPM1 reduction in Arabidopsis (Figure 7). In addition, PLDδ(R410D/R622P), an activity-deficient mutant, could not reduce the protein level of RPM1(D505V) in N. benthamiana further demonstrated the necessity of PLD activity to reduce the protein level of RPM1 (Figure 4).
Transiently expressed PLD resulted in the partial distribution of RPM1(D505V) to the soluble fraction of plant cells, suggesting that PLD could interfere the plasma-membrane location of RPM1 (Figure 3B). RPM1 does not have transmembrane domains. The isoelectric point of RPM1 is 8.51, and it should be positively charged at physiological conditions. The potential interaction of the positively charged RPM1 with the negatively charged phospholipids may facilitate the membrane location of RPM1. PLD can hydrolyze PC and PE to PA which still is negative charged. It seemed that PLD activity would not affect RPM1 location. However, PA can be further dephosphated by phosphatidic acid phosphohydrolase (PAP) to diacylglycerol (DAG) (Becker and Hannun, 2005). Therefore, PLD activity should result in the net increase of DAG and decrease of PC and PE. Because DAG does not contain negatively charged phosphate group, RPM1 is released from the membrane and degraded in the cytosol of the cells. It would be interested to determine the binding of RPM1 with phospholipids such as PE and PC, and compare the lipid components between PLDδ OE plants and the wild type or plants treated with and without ABA treatment (Wang and Wang, 2001; Andersson et al., 2006). PLDδ reduced the protein level of RPM1(D505V), a mimic mutant of the active RPM1, in N. benthamiana, but PLDδ did not obviously reduce the protein level of RPM1 in N. benthamiana. The different effects of PLDδ on RPM1(D505V) and RPM1 reflect the conclusion that PLDδ activity is required to reduce the protein level of RPM1. RPM1-Myc itself is not active in N. benthamiana. Overexpression of PLDδ does not definitely lead to PLD activity, especially the transiently expressed PLDδ only accumulated two days. RPM1(D505V) is autoactive, and can stimulate PLD activity. Therefore, PLDδ obviously reduces the protein level of RPM1(D505V) in N. benthamiana. The reason that overexpression of PLDδ reduced the protein level of RPM1 in Arabidopsis (Figure 5A) is possibly due to the long-term effects of PLDδ during the time its activity can be stimulated by environmental and physiological factors (Li et al., 2009).
PLD mediates the cross-talking between ABA signaling and RPM1-mediated disease resistance. We found that ABA treatment could reduce the protein level of RPM1 in A. thaliana. ABA is a phytohormone responding to the drought stress, while RPM1 is an immune receptor for disease resistance. ABA signaling activates PLD, and PLD activity regulates the protein level of RPM1 and its function. Therefore, PLD could be an integrating point to balance the plant responses to the complex environmental stimuli. Other abiotic stresses such as cold, heat, and ROS can also affect PLD activity (Li et al., 2009; Guo et al., 2012; Zhang et al., 2017). It has been reported that temperature affects RPM1 function (Wang et al., 2009). PLD activity may mediate this correlation. Although we clearly determined that PLDδ could negatively regulate the protein level of RPM1, we haven’t tested all the members of the PLD family yet. Since each PLD member has specific and overlapping functions, further research about the effects of PLD members with RPM1 and other immune receptors will help to understand how PLD proteins integrate environmental and physiological responses with disease resistances.
Author Contributions
XY and ZG designed the experiments, analyzed the data, and wrote the manuscript. XY, ZW, JH, and HX conducted the experiments.
Funding
This study was supported by the National Key Research and Development Program of China (Grant No. 2016YFD0100600) and the National Natural Science Foundation of China (Grant No. 31270315) to ZG.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Dr. Jeff Dangl (University of North Carolina) for providing A. thaliana seeds, Dr. Yingtang Lu (Wuhan University) for providing pathogen bacteria. We also thank Dr. Yunkuan Liang (Wuhan University) for critical reading.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2018.01991/full#supplementary-material
References
Adams-Phillips, L., Wan, J., Tan, X., Dunning, F. M., Meyers, B. C., Michelmore, R. W., et al. (2008). Discovery of ADP-ribosylation and other plant defense pathway elements through expression profiling of four different Arabidopsis–Pseudomonas R-avr interactions. Mol. Plant Microbe Interact. 21, 646–657. doi: 10.1094/mpmi-21-5-0646
Andersson, M. X., Kourtchenko, O., Dangl, J. L., Mackey, D., and Ellerstrom, M. (2006). Phospholipase-dependent signalling during the AvrRpm1- and AvrRpt2-induced disease resistance responses in Arabidopsis thaliana. Plant J. 47, 947–959. doi: 10.1111/j.1365-313X.2006.02844.x
Baggs, E., Dagdas, G., and Krasileva, K. V. (2017). NLR diversity, helpers and integrated domains: making sense of the NLR IDentity. Curr. Opin. Plant Biol. 38, 59–67. doi: 10.1016/j.pbi.2017.04.012
Bargmann, B. O., and Munnik, T. (2006). The role of phospholipase D in plant stress responses. Curr. Opin. Plant Biol. 9, 515–522. doi: 10.1016/j.pbi.2006.07.011
Becker, K. P., and Hannun, Y. A. (2005). Protein kinase C and phospholipase D: intimate interactions in intracellular signaling. Cell. Mol. Life Sci. 62, 1448–1461. doi: 10.1007/s00018-005-4531-7
Ben Othman, A., Ellouzi, H., Planchais, S., De Vos, D., Faiyue, B., Carol, P., et al. (2017). Phospholipases Dzeta1 and Dzeta2 have distinct roles in growth and antioxidant systems in Arabidopsis thaliana responding to salt stress. Planta 246, 721–735. doi: 10.1007/s00425-017-2728-2
Boyes, D. C., Nam, J., and Dangl, J. L. (1998). The Arabidopsis thaliana RPM1 disease resistance gene product is a peripheral plasma membrane protein that is degraded coincident with the hypersensitive response. Proc. Natl. Acad. Sci. U.S.A. 95, 15849–15854. doi: 10.1073/pnas.95.26.15849
Chung, E. H., da Cunha, L., Wu, A. J., Gao, Z., Cherkis, K., Afzal, A. J., et al. (2011). Specific threonine phosphorylation of a host target by two unrelated type III effectors activates a host innate immune receptor in plants. Cell Host Microbe 9, 125–136. doi: 10.1016/j.chom.2011.01.009
Citovsky, V., Lee, L. Y., Vyas, S., Glick, E., Chen, M. H., Vainstein, A., et al. (2006). Subcellular localization of interacting proteins by bimolecular fluorescence complementation in planta. J. Mol. Biol. 362, 1120–1131. doi: 10.1016/j.jmb.2006.08.017
Couto, D., and Zipfel, C. (2016). Regulation of pattern recognition receptor signalling in plants. Nat. Rev. Immunol. 16, 537–552. doi: 10.1038/nri.2016.77
Cui, H., Tsuda, K., and Parker, J. E. (2015). Effector-triggered immunity: from pathogen perception to robust defense. Annu. Rev. Plant Biol. 66, 487–511. doi: 10.1146/annurev-arplant-050213-040012
El Kasmi, F., Chung, E. H., Anderson, R. G., Li, J., Wan, L., Eitas, T. K., et al. (2017). Signaling from the plasma-membrane localized plant immune receptor RPM1 requires self-association of the full-length protein. Proc. Natl. Acad. Sci. U.S.A. 114, E7385–E7394. doi: 10.1073/pnas.1708288114
Ella, K. M., Meier, K. E., Kumar, A., Zhang, Y., and Meier, G. P. (1997). Utilization of alcohols by plant and mammalian phospholipase D. Biochem. Mol. Biol. Int. 41, 715–724. doi: 10.1080/15216549700201761
Gao, Z., Chung, E. H., Eitas, T. K., and Dangl, J. L. (2011). Plant intracellular innate immune receptor Resistance to Pseudomonas syringae pv. maculicola 1 (RPM1) is activated at, and functions on, the plasma membrane. Proc. Natl. Acad. Sci. U.S.A. 108, 7619–7624. doi: 10.1073/pnas.1104410108
Grant, M., Brown, I., Adams, S., Knight, M., Ainslie, A., and Mansfield, J. (2000). The RPM1 plant disease resistance gene facilitates a rapid and sustained increase in cytosolic calcium that is necessary for the oxidative burst and hypersensitive cell death. Plant J. 23, 441–450. doi: 10.1046/j.1365-313x.2000.00804.x
Grant, M. R., Godiard, L., Straube, E., Ashfield, T., Lewald, J., Sattler, A., et al. (1995). Structure of the Arabidopsis RPM1 gene enabling dual specificity disease resistance. Science 269, 843–846. doi: 10.1126/science.7638602
Guo, L., Devaiah, S. P., Narasimhan, R., Pan, X., Zhang, Y., Zhang, W., et al. (2012). Cytosolic glyceraldehyde-3-phosphate dehydrogenases interact with phospholipase Dδ to transduce hydrogen peroxide signals in the Arabidopsis response to stress. Plant Cell 24, 2200–2212. doi: 10.1105/tpc.111.094946
Hong, Y., Devaiah, S. P., Bahn, S. C., Thamasandra, B. N., Li, M., Welti, R., et al. (2009). Phospholipase D epsilon and phosphatidic acid enhance Arabidopsis nitrogen signaling and growth. Plant J. 58, 376–387. doi: 10.1111/j.1365-313X.2009.03788.x
Hong, Y., Pan, X., Welti, R., and Wang, X. (2008). Phospholipase Dalpha3 is involved in the hyperosmotic response in Arabidopsis. Plant Cell 20, 803–816. doi: 10.1105/tpc.107.056390
Hubert, D. A., He, Y., McNulty, B. C., Tornero, P., and Dangl, J. L. (2009). Specific Arabidopsis HSP90.2 alleles recapitulate RAR1 cochaperone function in plant NB-LRR disease resistance protein regulation. Proc. Natl. Acad. Sci. U.S.A. 106, 9556–9563. doi: 10.1073/pnas.0904877106
Jacob, T., Ritchie, S., Assmann, S. M., and Gilroy, S. (1999). Abscisic acid signal transduction in guard cells is mediated by phospholipase D activity. Proc. Natl. Acad. Sci. U.S.A. 96, 12192–12197. doi: 10.1073/pnas.96.21.12192
Johansson, O. N., Fahlberg, P., Karimi, E., Nilsson, A. K., Ellerstrom, M., and Andersson, M. X. (2014). Redundancy among phospholipase D isoforms in resistance triggered by recognition of the Pseudomonas syringae effector AvrRpm1 in Arabidopsis thaliana. Front. Plant Sci. 5:639. doi: 10.3389/fpls.2014.00639
Jones, J. D., and Dangl, J. L. (2006). The plant immune system. Nature 444, 323–329. doi: 10.1038/nature05286
Karimi, M., Depicker, A., and Hilson, P. (2007). Recombinational cloning with plant gateway vectors. Plant Physiol. 145, 1144–1154. doi: 10.1104/pp.107.106989
Li, M., Hong, Y., and Wang, X. (2009). Phospholipase D- and phosphatidic acid-mediated signaling in plants. Biochim. Biophys. Acta 1791, 927–935. doi: 10.1016/j.bbalip.2009.02.017
Li, M., Qin, C., Welti, R., and Wang, X. (2006). Double knockouts of phospholipases Dzeta1 and Dzeta2 in Arabidopsis affect root elongation during phosphate-limited growth but do not affect root hair patterning. Plant Physiol. 140, 761–770. doi: 10.1104/pp.105.070995
Liu, J., Elmore, J. M., Lin, Z. J., and Coaker, G. (2011). A receptor-like cytoplasmic kinase phosphorylates the host target RIN4, leading to the activation of a plant innate immune receptor. Cell Host Microbe 9, 137–146. doi: 10.1016/j.chom.2011.01.010
Mackey, D., Holt, B. F. III, Wiig, A., and Dangl, J. L. (2002). RIN4 interacts with Pseudomonas syringae type III effector molecules and is required for RPM1-mediated resistance in Arabidopsis. Cell 108, 743–754. doi: 10.1016/S0092-8674(02)00661-X
Mishra, G., Zhang, W., Deng, F., Zhao, J., and Wang, X. (2006). A bifurcating pathway directs abscisic acid effects on stomatal closure and opening in Arabidopsis. Science 312, 264–266. doi: 10.1126/science.1123769
Nakagawa, T., Kurose, T., Hino, T., Tanaka, K., Kawamukai, M., Niwa, Y., et al. (2007). Development of series of gateway binary vectors, pGWBs, for realizing efficient construction of fusion genes for plant transformation. J. Biosci. Bioeng. 104, 34–41. doi: 10.1263/jbb.104.34
Pinosa, F., Buhot, N., Kwaaitaal, M., Fahlberg, P., Thordal-Christensen, H., Ellerstrom, M., et al. (2013). Arabidopsis phospholipase ddelta is involved in basal defense and nonhost resistance to powdery mildew fungi. Plant Physiol. 163, 896–906. doi: 10.1104/pp.113.223503
Pitino, M., Armstrong, C. M., Cano, L. M., and Duan, Y. (2016). Transient expression of Candidatus liberibacter asiaticus effector induces cell death in Nicotiana benthamiana. Front. Plant Sci. 7:982. doi: 10.3389/fpls.2016.00982
Sang, Y., Zheng, S., Li, W., Huang, B., and Wang, X. (2001). Regulation of plant water loss by manipulating the expression of phospholipase Dalpha. Plant J. 28, 135–144. doi: 10.1046/j.1365-313X.2001.01138.x
Walter, M., Chaban, C., Schutze, K., Batistic, O., Weckermann, K., Nake, C., et al. (2004). Visualization of protein interactions in living plant cells using bimolecular fluorescence complementation. Plant J. 40, 428–438. doi: 10.1111/j.1365-313X.2004.02219.x
Wang, C., and Wang, X. (2001). A novel phospholipase D of Arabidopsis that is activated by oleic acid and associated with the plasma membrane. Plant Physiol. 127, 1102–1112. doi: 10.1104/pp.010444
Wang, Y., Bao, Z., Zhu, Y., and Hua, J. (2009). Analysis of temperature modulation of plant defense against biotrophic microbes. Mol. Plant Microbe Interact. 22, 498–506. doi: 10.1094/mpmi-22-5-0498
Zhang, Q., Song, P., Qu, Y., Wang, P., Jia, Q., Guo, L., et al. (2017). Phospholipase D delta negatively regulates plant thermotolerance by destabilizing cortical microtubules in Arabidopsis. Plant Cell Environ. 40, 2220–2235. doi: 10.1111/pce.13023
Zhao, J., Devaiah, S. P., Wang, C., Li, M., Welti, R., and Wang, X. (2013). Arabidopsis phospholipase Dbeta1 modulates defense responses to bacterial and fungal pathogens. New Phytol. 199, 228–240. doi: 10.1111/nph.12256
Keywords: phospholipase D, RPM1, ABA, HR, plant immunity
Citation: Yuan X, Wang Z, Huang J, Xuan H and Gao Z (2019) Phospholipidase Dδ Negatively Regulates the Function of Resistance to Pseudomonas syringae pv. Maculicola 1 (RPM1). Front. Plant Sci. 9:1991. doi: 10.3389/fpls.2018.01991
Received: 27 August 2018; Accepted: 20 December 2018;
Published: 18 January 2019.
Edited by:
Jens Staal, Ghent University, BelgiumReviewed by:
Volkan Cevik, University of Bath, United KingdomMarta Berrocal-Lobo, Polytechnic University of Madrid, Spain
Copyright © 2019 Yuan, Wang, Huang, Xuan and Gao. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Zhiyong Gao, enlnYW9Ad2h1LmVkdS5jbg==