 Steffi Fritsche
Steffi Fritsche Amy L. Klocko
Amy L. Klocko Agnieszka Boron1
Agnieszka Boron1 Amy M. Brunner
Amy M. Brunner Glenn Thorlby
Glenn Thorlby
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Plant Sci. , 15 November 2018
Sec. Plant Biotechnology
Volume 9 - 2018 | https://doi.org/10.3389/fpls.2018.01671
This article is part of the Research Topic Forest Genomics and Biotechnology View all 20 articles
A considerable body of research exists concerning the development of technologies to engineer sterility in forest trees. The primary driver for this work has been to mitigate concerns arising from gene flow from commercial plantings of genetically engineered (GE) trees to non-GE plantations, or to wild or feral relatives. More recently, there has been interest in the use of sterility technologies as a means to mitigate the global environmental and socio-economic damage caused by the escape of non-native invasive tree species from planted forests. The current sophisticated understanding of the molecular processes underpinning sexual reproduction in angiosperms has facilitated the successful demonstration of a number of control strategies in hardwood tree species, particularly in the model hardwood tree Poplar. Despite gymnosperm softwood trees, such as pines, making up the majority of the global planted forest estate, only pollen sterility, via cell ablation, has been demonstrated in softwoods. Progress has been limited by the lack of an endogenous model system, long timescales required for testing, and key differences between softwood reproductive pathways and those of well characterized angiosperm model systems. The availability of comprehensive genome and transcriptome resources has allowed unprecedented insights into the reproductive processes of both hardwood and softwood tree species. This increased fundamental knowledge together with the implementation of new breeding technologies, such as gene editing, which potentially face a less oppressive regulatory regime, is making the implementation of engineered sterility into commercial forestry a realistic possibility.
Increasing global population coupled with transition to a sustainable bio-based economy is predicted to lead to growing pressure on forests to deliver wood-based products, energy, food, and ecosystem services whilst maintaining their role as major reservoirs of biodiversity. To accommodate this growing demand, it is estimated that the amount of wood we take from forests and plantations each year may need to triple by 2050 (WWF, 2015). Planted forests, which in 2015 made up 7% of forest lands, provide a means to sustainably increase production of forest products and reduce pressure on natural forests (FAO, 2015). Alongside improved silviculture, land management and other technological advances, biotech-based technologies offer tools to enhance the sustainability and productivity of planted forests (Al-Ahmad, 2018). Sterile trees have the ability to impact a number of obstacles to increasing productivity from planted forests.
Genetic engineering (GE) is able to provide solutions for many of the challenges forestry faces to sustainably increase forest production. Improved wood quantity and quality, processability, biotic, and abiotic stress tolerance and herbicide tolerance (Harfouche et al., 2011; Porth and El-Kassaby, 2014; Etchells et al., 2015; Ault et al., 2016; Zhou et al., 2017) are amongst the traits successfully demonstrated. The recent approval by the Brazilian Regulator for GE Eucalyptus that are able to grow 15–20% faster than the best existing clonal lines (Nature Biotechnology News, 2015) seems likely to lead to first large-scale commercial planting of trees.
There remain well documented regulatory and social challenges associated with commercial planting of GE trees (Porth and El-Kassaby, 2014; Strauss et al., 2016). Gene flow from transgenic trees remains a major concern, particularly as forest trees are virtually undomesticated and pollen is able to disseminate over great distances (DiFazio et al., 2004). Seeds also have the potential to spread, either locally or over distances, depending on the species. Transgene containment through the production of trees that are unable to produce fertile reproductive propagules has the ability to mitigate these concerns and prevent, or severely reduce, the flow of genes via sexual reproduction.
Increasing attention is being paid to the ecological, economic, and cultural damage caused by invasive tree species that have “escaped” by seed dispersal from planted forests (Breton et al., 2008; Nuñez et al., 2017). Globally, Pinus species are recognized as among the most widespread and influential of all invasive plants (Richardson and Rejmánek, 2004). These escapes, or wildings, are particularly a problem in the Southern Hemisphere where a large percentage of tree plantations are composed of exotic species (Franzese and Raffaele, 2017). South Africa, New Zealand, and Australia, who were early adopters of exotic conifer plantations, have been joined more recently by several South American nations in facing wilding challenges (Simberloff et al., 2010). For example, in New Zealand several exotic conifer species have become established and now occupy ∼1.8 million ha, and are expanding by about 6% per annum (Froude, 2011). Economic and ecological damage resulting from these wildings is challenging the license to operate, with commercially advantageous, but wilding-prone species such as Douglas-fir (Pseudotsuga menziesii). The ability to generate trees that are unable to reproduce would allow control programs to focus on the existing populations and give forest owners freedom to operate for new plantings.
The ability to either prevent reproduction or limit the development of reproductive propagules is predicted to boost growth and increase wood production in forest trees by redirecting energy and nutrients to increased vegetative growth (Strauss et al., 1995; Luis and José, 2014). Conclusive evidence for such a reproductive cost is lacking but is supported by evidence that in conifers cone production may utilize a significant proportion of the trees energy and assimilates (Cremer, 1992; Sala et al., 2012; Kramer et al., 2014). Unsurprisingly, in conifers, the long-lived female cones are more energy demanding than the generally more transient male cones (Obeso, 2002). These observations suggest that engineered sterility, particularly female sterility is likely to have a positive impact on vegetative growth and wood production. Long-term growth comparisons between sterile and reproductive trees would provide direct evidence for this and allow quantification of growth differences.
Pollen from many trees cause allergenic reactions and symptoms correlate with exposure (Buters et al., 2012). Planted forests can provide a major source of seasonal allergens (D’amato et al., 2007). For example, allergy to sugi (Cryptomeria japonica) pollen is reported to effect 26.5% of the Japanese population (Taniguchi, 2018). The ability to prevent or limit pollen production from planted forests would provide relief to allergy suffers and mitigate potential social license to operate challenges.
Both angiosperm (hardwood) and gymnosperm/conifer (softwood) trees are used as plantation species. Although they share broad similarities in their reproductive processes, there are distinct differences between them.
Perhaps no other plant development process has been studied more than flowering. For Arabidopsis, the in planta functions of a large number of flowering genes as well as their regulatory network context are known and studies in plants such as rice and petunia have revealed broad functional conservation (Pajoro et al., 2014). These include genes that regulate the transition to flowering, floral organ identity as well as pollen and ovule development. Although advances in sequencing have enabled the identification of flowering gene homologs in diverse angiosperm trees, there are few cases where in planta functions have been characterized in trees (Brunner et al., 2017; Klocko et al., 2018). This is due to the long non-flowering period that can last years to decades and that for most species, genetic transformation is a formidable hurdle. Trees also differ from herbaceous plants in the prolonged period between the floral transition and anthesis. In tropical species such as Eucalyptus, this occurs in one season, but temperate species exhibit indirect flowering, with flower development initiated in 1 year and completed the following year (Vining et al., 2015; Brunner et al., 2017). Thus, multi-year field trials that require monitoring of large trees and collecting flowers from the upper portion of the tree crown are typically required to demonstrate sterility or delay of flowering.
Selection of candidate genes for genetic containment in trees based on homology to Arabidopsis flowering genes and gene expression might be straightforward, but such conservation does not necessarily translate to the predicted or desired phenotype. Flowering time genes are attractive targets because prevention of flowering is easier to monitor (e.g., no need to demonstrate flowers are sterile) and to prevent resource allocation to reproduction. However, accumulating evidence supports that tree homologs of various flowering time and floral meristem identity genes have roles in both vegetative and reproductive phenology (Bohlenius et al., 2006; Bielenberg et al., 2008; Hoenicka et al., 2008; Mohamed et al., 2010; Hsu et al., 2011; Azeez et al., 2014; Tylewicz et al., 2015; Parmentier-Line and Coleman, 2016). Targeting such genes for manipulation can thus result in undesired vegetative effects, such as delayed bud flush, in addition to predicted effects on flowering or no effect on flowering (Hoenicka et al., 2012). However, promising results have also been achieved, such as the delayed flowering without growth reduction demonstrated by overexpressing the poplar ortholog of the floral repressor SHORT VEGETATIVE PHASE (SVP) (Klocko et al., 2018). Manipulation of floral organ identity genes might be less likely to have vegetative effects as these genes may show stronger conservation of reproductive-only function. For example, considerable evidence supports that the AGAMOUS (AG) subgroup of MADS-box genes have reproductive functions not only in angiosperms but also in gymnosperms (Dreni and Kater, 2014). However, even in these cases, results can differ from expectation. For example, downregulation of the conserved floral meristem identity gene LEAFY (LFY) in a male poplar genotype induced bisexual and female flowers (Klocko et al., 2018). Despite the challenges, the knowledge gained from gene function and sterility studies in trees along with more detailed and extensive genome-wide expression studies in different angiosperm trees will enable more accurate gene selection for manipulation of only-reproductive traits.
Unlike angiosperms, where there is extensive knowledge of the molecular factors involved in the reproduction process, relatively little is known regarding gymnosperms. A number of putative genes have been identified through comparative analyses of orthologous angiosperm genes, tissue-specific expression analysis or genome sequencing. However, several key floral genes including FD, SQUAMOSA- (SQUA-) or SEPALLATA-like (SEP-like) seem to be absent (Becker, 2003; Abe et al., 2005; Zahn et al., 2005; Melzer et al., 2010; Karlgren et al., 2011; Jaeger et al., 2013). Initial research was able to detect orthologs to only the B- and C-genes involved in the control of meristem formation and organ identity in the developing cones (Tandre et al., 1995, 1998; Mouradov et al., 1998; Rutledge et al., 1998; Fukui et al., 2001; Sundström and Engström, 2002; Gramzow et al., 2014; Katahata et al., 2014; Uddenberg et al., 2015). More recently, ABCE model prototype transcription factors, genes that define the developmental flower organ model (ABC(DE)) in angiosperms, have been confirmed in gymnosperms (Chen et al., 2017). Conifer-specific genes such as the DEFICIENS-AGAMOUS-LIKE (DAL) and NEEDLY have also been identified but functional knowledge is limited due to the lack of angiosperm orthologs (Carlsbecker et al., 2003, 2004; Rudall et al., 2011). It is not generally possible to predict which of the many vegetative meristems will undergo the reproductive bud transition before changes are initiated making research on reproductive initiation a bold venture (Williams, 2009).
The biggest bottleneck for conifer reproduction research is the inability to carry out functional characterization in an endogenous system which prevents the definitive elucidation of gene function. Testing of gene function in angiosperm model systems has produced inconclusive results. Whilst some such studies have confirmed the function of putative orthologs, others failed to find flowering related differences, found multiple phenotypic alteration or were unable to complement mutants, highlighting the need for a reliable conifer testing system (Rutledge et al., 1998; Tandre et al., 1998; Shindo et al., 2001; Sundström and Engström, 2002; Carlsbecker et al., 2003, 2004; Nilsson et al., 2007; Shiokawa et al., 2008; Klintenas et al., 2012; Katahata et al., 2014; Liu et al., 2018). As discussed above for angiosperm trees, it might be challenging to identify flowering time gene homologs in conifers that do not have roles in vegetative development that make their manipulation for reproductive sterility problematic. For example, the gymnosperm FLOWERING LOCUS T-like subfamily has been suggested to have roles in both vegetative and reproductive phenology (Klintenas et al., 2012; Karlgren et al., 2013; Nystedt et al., 2013; Liu et al., 2016). However, the lack of characterization in an endogenous system means that the function of the sub-family members remains unresolved.
The increasingly sophisticated understanding of the molecular processes underpinning sexual reproduction described above has facilitated the successful demonstration of a number of sterility strategies in plants. Chief amongst these are strategies using ablation of reproductive cells or structures and the inactivation or suppression of genes essential for normal reproductive processes. Here, we highlight only a selection of sterility approaches and refer readers to Brunner et al. (2007) and Hoenicka et al. (2016b) for additional examples.
The use of cell or tissue-specific promoters to direct the expression of cytotoxic genes (Palmiter et al., 1987) to reproductive tissues has been widely used to investigate and modify reproductive development in plants (Goldman et al., 1994; Beals and Goldberg, 1997). Numerous examples exist of using this technology in plants to generate male and female sterility (Mariani et al., 1990; Goldman et al., 1994; De Block et al., 1997). Complete (dual male and female) sterility has also been achieved using either independent male- and female-specific promoters or a single promoter targeting both tissues simultaneously (Liu and Liu, 2008; Huang et al., 2016). Male sterility using this cell ablation strategy has been demonstrated in both hardwood and softwood trees via expression of the BARNASE gene from Bacillus amyloliquefaciens under the control of reproductive tissue-specific promoters (Table 1). A key requirement for a cell ablation strategy is a promoter that tightly directs expression of the cytotoxin to the desired reproductive tissue to prevent pleiotropic effects on non-reproductive tissues. The conservation of expression of some floral genes has facilitated the use of a number of well characterized promoters across species (Strauss et al., 1995). Indeed, an anther-specific promoter derived from Pinus radiata has been used to express the BARNASE gene in both a softwood (pine) and hardwood (Eucalyptus) tree to deliver male sterility (Zhang et al., 2012). We are unware of dual male/female BARNASE-mediated sterility being demonstrated in trees without negative pleiotropic effects (Lemmetyinen et al., 2004) but this should be possible if a suitable promoter is used.
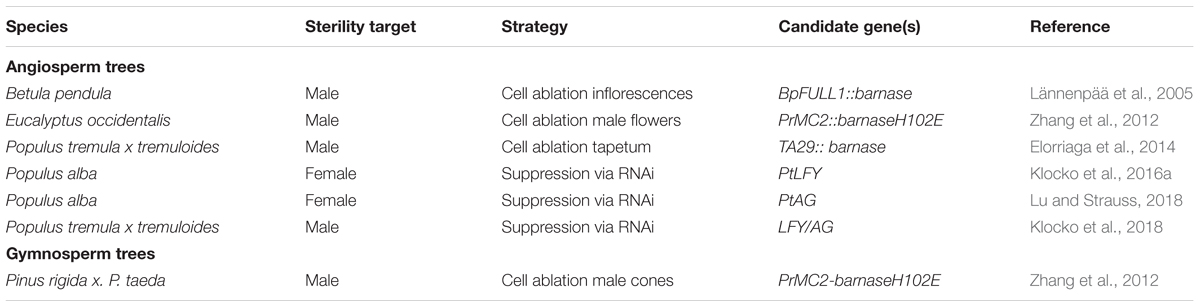
TABLE 1. Examples of Engineered Sterility in Forest Trees.
RNA interference (RNAi) is a well proven homology-dependent gene silencing technology that involves double-stranded RNA directed against a target gene or its promoter region (Mansoor et al., 2006). Numerous demonstrations of engineered sterility through the suppression of genes essential for normal reproduction are available in angiosperm species (Wang et al., 2012). RNAi silencing has been used in angiosperm trees to engineer sterility with constructs targeting LFY and AG successfully producing sterile trees (Table 1). The production of male and female sterile plants via the use of chimeric repressors targeting transcription factors involved in flower development has also been demonstrated (Mitsuda et al., 2006; Katahata et al., 2014). In conifers the use of gene suppression methods to prevent reproduction has not been demonstrated even though such methods have been widely used to investigate wood quality traits (Wagner et al., 2005, 2009; Souza et al., 2007; Trontin et al., 2007). Attempts have been reported of expressing conifer flowering-associated genes in an endogenous system (Karlgren et al., 2013) but these studies have not directly sought to address sterility.
This lack of success in conifers reflects both a lack of fundamental knowledge regarding conifer reproduction and the inherent difficulties in working with conifers including the long timescale required for testing. For example, attempts to investigate the effects of over-expressing the Arabidopsis LFY gene in P. radiata were not informative as neither modified or control plants initiated reproduction during the 8 years that the trees were grown (NZ-EPA, 2008; Lottmann et al., 2010).
The social, legal and ecological impacts of sterile trees is still controversial (Williams, 2005; Kazana et al., 2015; Strauss et al., 2017). Although sterility provides mitigation for some of the social and ecological objections to the deployment of both GE trees and species with the potential to become invasive, this may be challenged if the sterility technology is itself GE.
The recent development of a number of new breeding technologies, including gene editing, that are already seeing widespread application in crop species (Nekrasov et al., 2017; Waltz, 2018) have great potential in forest trees. Site-directed mutagenesis would allow the inactivation of genes that are essential for normal reproductive processes and the generation of sterile trees. Gene editing-mediated mutagenesis would be particularly advantageous in forest trees where, to date, mutagenesis breeding has played an extremely limited role. The permanent inactivation of a gene would provide assurance of enduring containment and reduce concerns associated with the stability of long-term transgene expression associated with silencing of over-expression technologies (Li et al., 2008). Site directed mutagenesis via CRISPR-cas9 has been demonstrated in a number of tree species including Poplar (Fan et al., 2015) where mutagenesis of genes involved in flowering (Elorriaga et al., 2018) has also been shown. To date, gene editing has not been published in conifer species. However, the existence of a small number of natural spontaneous sterile conifer mutants (Orr-Ewing, 1977; Wilson and Owens, 2003; Rudall et al., 2011) suggest that a targeted-mutagenesis strategy would be successful if suitable targets can be identified.
Although the regulatory landscape regarding gene editing technologies remains complex, it is likely that in many jurisdictions versions of the technology that do not include foreign DNA in the final organisms will not be regulated as GMOs (Waltz, 2016; Davison and Ammann, 2017; Ishii and Araki, 2017). This would provide a more straightforward and less costly route to commercial release than is currently the case for products of GE technology (Waltz, 2018). This regulatory approach would hold particular promise in applications where sterility is a standalone trait, such as for the control of invasive tree species, rather than providing a means of containment for other (GE) traits. This strategy would require DNA-free editing technologies as outcrossing of transgenes would not be possible with sterile trees.
The second major challenge has been the inability to carry out timely prototyping of sterility constructs in commercially important species. To facilitate testing in conifers it is desirable to develop a system analogous to the Poplar model system (Jansson and Douglas, 2007; Douglas, 2017) which has allowed relatively rapid prototyping of sterility constructs (Klocko et al., 2018). Although effective transformation systems exist for a number of commercially important conifers including P. radiata, P. taeda, and Picea abies, these species have long pre-reproductive juvenile growth periods that limits their use as sterility-testing platforms (Tang and Newton, 2003; Uddenberg et al., 2015). Some conifer species are able to reproduce at a much younger age (Righter, 1939; Pharis et al., 1987; Uddenberg et al., 2013) or can be induced to undergo early reproduction. Such precocious reproduction has been demonstrated in both hardwoods and softwoods by grafting onto older rootstock (Simak, 1978; Zhang et al., 2012), and by the application of external stimuli such as hormone treatments (Pharis et al., 1965; Ross and Pharis, 1985; Meilan, 1997). Stable introduction of FT transgenes induced precocious fertile flowers in Eucalyptus (Klocko et al., 2016b) and in Populus when combined with a low temperature treatment (Hoenicka et al., 2016a). In fruit trees, viral vectors that express floral promoters or silence repressors induced early flowering (Velázquez et al., 2016; Yamagishi et al., 2016). Although these offer potential routes to earlier testing of sterility strategies, developing the required tissue culture and transformation capabilities for a new tree species remains a significant barrier.
The increasing availability of genome and transcriptome resources for forest trees is providing new insights into reproductive processes. This is reducing the reliance on non-tree model systems and providing novel species-specific knowledge of reproductive processes and candidate genes for modification. The development of gene-editing-based targeted mutagenesis is likely to be the most attractive route to engineered sterility as it offers precise and predictable modifications combined with assurance of phenotypic stability. The lack of global consensus on the regulation of gene editing technology remains a barrier to research investment and commercialization and complicates the public debate that must go hand-in-hand with progress toward implementation.
All authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
This work was supported by Scion’s Strategic Science Investment Funding (SSIF) from the Science and Innovation Group, Ministry of Innovation, Business and Science.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Abe, M., Kobayashi, Y., Yamamoto, S., Daimon, Y., Yamaguchi, A., Ikeda, Y., et al. (2005). FD, a bZIP protein mediating signals from the floral pathway integrator FT at the shoot apex. Science 309, 1052–1056. doi: 10.1126/science.1115983
Al-Ahmad, H. (2018). Biotechnology for bioenergy dedicated trees: meeting future energy demands. Z. Naturforsch. 73, 15–32. doi: 10.1515/znc-2016-0185
Ault, K., Viswanath, V., Jayawickrama, J., Ma, C., Eaton, J., Meilan, R., et al. (2016). Improved growth and weed control of glyphosate-tolerant poplars. New For. 47, 653–667. doi: 10.1007/s11056-016-9536-6
Azeez, A., Miskolczi, P., Tylewicz, S., and Bhalerao, R. P. (2014). A tree ortholog of APETALA1 mediates photoperiodic control of seasonal growth. Curr. Biol. 24, 717–724. doi: 10.1016/j.cub.2014.02.037
Beals, T. P., and Goldberg, R. B. (1997). A novel cell ablation strategy blocks tobacco anther dehiscence. Plant Cell 9, 1527–1545. doi: 10.1105/tpc.9.9.1527
Becker, A. (2003). The major clades of MADS-box genes and their role in the development and evolution of flowering plants. Mol. Phylogenet. Evol. 29, 464–489. doi: 10.1016/s1055-7903(03)00207-0
Bielenberg, D. G., Wang, Y., Li, Z. G., Zhebentyayeva, T., Fan, S. H., Reighard, G. L., et al. (2008). Sequencing and annotation of the evergrowing locus in peach [Prunus persica (L.) Batsch] reveals a cluster of six MADS-box transcription factors as candidate genes for regulation of terminal bud formation. Tree Genet. Genomes 4, 495–507. doi: 10.1007/s11295-007-0126-9
Bohlenius, H., Huang, T., Charbonnel-Campaa, L., Brunner, A. M., Jansson, S., Strauss, S. H., et al. (2006). CO/FT regulatory module controls timing of flowering and seasonal growth cessation in trees. Science 312, 1040–1043. doi: 10.1126/science.1126038
Breton, C., Guerin, J., Ducatillion, C., Medail, F., Kull, C. A., and Berville, A. (2008). Taming the wild and ‘wilding’the tame: tree breeding and dispersal in Australia and the Mediterranean. Plant Sci. 175, 197–205. doi: 10.1016/j.plantsci.2008.05.013
Brunner, A. M., Li, J., DiFazio, S. P., Shevchenko, O., Montgomery, B. E., Mohamed, R., et al. (2007). Genetic containment of forest plantations. Tree Genet. Genomes 3, 75–100. doi: 10.1007/s11295-006-0067-8
Brunner, A. M., Varkonyi-Gasic, E., and Jones, R. C. (2017). “Phase change and phenology in trees,” in Comparative and Evolutionary Genomics of Angiosperm Trees, eds A. Groover and Q. Cronk (Cham: Springer), 227–274. doi: 10.1007/7397_2016_30
Buters, J. T. M., Thibaudon, M., Smith, M., Kennedy, R., Rantio-Lehtimäki, A., Albertini, R., et al. (2012). Release of Bet v 1 from birch pollen from 5 European countries. Results from the HIALINE study. Atmos. Environ. 55, 496–505. doi: 10.1016/j.atmosenv.2012.01.054
Carlsbecker, A., Sundström, J., Tandre, K., Englund, M., Kvarnheden, A., Johanson, U., et al. (2003). The DAL10 gene from norway spruce (Picea abies) belongs to a potentially gymnosperm-specific subclass of MADS-box genes and is specifically active in seed cones and pollen cones. Evol. Dev. 5, 551–561. doi: 10.1046/j.1525-142X.2003.03060.x
Carlsbecker, A., Tandre, K., Johanson, U., Englund, M., and Engstrom, P. (2004). The MADS-box gene DAL1 is a potential mediator of the juvenile-to-adult transition in Norway spruce (Picea abies). Plant J. 40, 546–557. doi: 10.1111/j.1365-313X.2004.02226.x
Chen, F., Zhang, X., Liu, X., and Zhang, L. (2017). Evolutionary analysis of MIKC(c)-type MADS-box genes in gymnosperms and angiosperms. Front. Plant Sci. 8:895. doi: 10.3389/fpls.2017.00895
Cremer, K. (1992). Relations between reproductive growth and vegetative growth of Pinus radiata. For. Ecol. Manage. 52, 179–199. doi: 10.1016/0378-1127(92)90501-Y
D’amato, G., Cecchi, L., Bonini, S., Nunes, C., Annesi-Maesano, I., Behrendt, H., et al. (2007). Allergenic pollen and pollen allergy in Europe. Allergy 62, 976–990. doi: 10.1111/j.1398-9995.2007.01393.x
Davison, J., and Ammann, K. (2017). New GMO regulations for old: determining a new future for EU crop biotechnology. GM Crops Food 8, 13–34. doi: 10.1080/21645698.2017.1289305
De Block, M., Debrouwer, D., and Moens, T. (1997). The development of a nuclear male sterility system in wheat. Expression of the barnase gene under the control of tapetum specific promoters. Theor. Appl. Genet. 95, 125–131. doi: 10.1007/s001220050540
DiFazio, S. P., Slavov, G. T., Burczyk, J., Leonardi, S., and Strauss, S. H. (2004). “Gene flow from tree plantations and implications for transgenic risk assessment,” in Plantation Forest Biotechnology for the 21st Century, eds C. Walter and M. Carson (Trivandrum: Research Signpost), 405–422.
Douglas, C. J. (2017). “Populus as a model tree,” in Comparative and Evolutionary Genomics of Angiosperm Trees, eds A. Groover and Q. Cronk (Berlin: Springer International publishing), 61–84. doi: 10.1007/7397_2016_12
Dreni, L., and Kater, M. M. (2014). MADS reloaded: evolution of the AGAMOUS subfamily genes. New Phytol. 201, 717–732. doi: 10.1111/nph.12555
Elorriaga, E., Klocko, A. L., Ma, C., and Strauss, S. H. (2018). Variation in mutation spectra among CRISPR/Cas9 mutagenized poplars. Front. Plant Sci. 9:594. doi: 10.3389/fpls.2018.00594
Elorriaga, E., Meilan, R., Ma, C., Skinner, J. S., Etherington, E., Brunner, A., et al. (2014). A tapetal ablation transgene induces stable male sterility and slows field growth in Populus. Tree Genet. Genomes 10, 1583–1593. doi: 10.1007/s11295-014-0781-6
Etchells, J. P., Mishra, L. S., Kumar, M., Campbell, L., and Turner, S. R. (2015). Wood formation in trees is increased by manipulating PXY-regulated cell division. Curr. Biol. 25, 1050–1055. doi: 10.1016/j.cub.2015.02.023
Fan, D., Liu, T., Li, C., Jiao, B., Li, S., Hou, Y., et al. (2015). Efficient CRISPR/Cas9-mediated targeted mutagenesis in Populus in the first generation. Sci. Rep. 5:12217. doi: 10.1038/srep12217
FAO (2015). Global Forest Resources Assessement 2015: How are the Worlds Forests Changing? Rome: Food and Agricultural Organization of the United Nations. Available at: http://www.fao.org/3/a-i4793e.pdf [accessed May 2018 2015].
Franzese, J., and Raffaele, E. (2017). Fire as a driver of pine invasions in the Southern Hemisphere: a review. Biol. Invasions 19, 2237–2246. doi: 10.1007/s10530-017-1435-z
Froude, V. A. (2011). Wilding Conifers in New Zealand: Beyond the Status Report. Bay of Islands: Report prepared for the Ministry of Agriculture and Forestry, Pacific Eco-Logic, 44.
Fukui, M., Futamura, N., Mukai, Y., Wang, Y., Nagao, A., and Shinohara, K. (2001). Ancestral MADS box genes in sugi, Cryptomeria japonica D. Don (Taxodiaceae), homologous to the B function genes in angiosperms. Plant Cell Physiol. 42, 566–575. doi: 10.1093/pcp/pce069
Goldman, M., Goldberg, R., and Mariani, C. (1994). Female sterile tobacco plants are produced by stigma-specific cell ablation. EMBO J. 13, 2976–2984. doi: 10.1002/j.1460-2075.1994.tb06596.x
Gramzow, L., Weilandt, L., and Theissen, G. (2014). MADS goes genomic in conifers: towards determining the ancestral set of MADS-box genes in seed plants. Ann. Bot. 114, 1407–1429. doi: 10.1093/aob/mcu066
Harfouche, A., Meilan, R., and Altman, A. (2011). Tree genetic engineering and applications to sustainable forestry and biomass production. Trends Biotechnol. 29, 9–17. doi: 10.1016/j.tibtech.2010.09.003
Hoenicka, H., Lautner, S., Klingberg, A., Koch, G., El-Sherif, F., Lehnhardt, D., et al. (2012). Influence of over-expression of the FLOWERING PROMOTING FACTOR 1 gene (FPF1) from Arabidopsis on wood formation in hybrid poplar (Populus tremula L. × P. tremuloides Michx.). Planta 235, 359–373. doi: 10.1007/s00425-011-1507-8
Hoenicka, H., Lehnhardt, D., Briones, V., Nilsson, O., and Fladung, M. (2016a). Low temperatures are required to induce the development of fertile flowers in transgenic male and female early flowering poplar (Populus tremula L.). Tree Physiol. 36, 667–677. doi: 10.1093/treephys/tpw015
Hoenicka, H., Lehnhardt, D., Nunna, S., Reinhardt, R., Jeltsch, A., Briones, V., et al. (2016b). Level of tissue differentiation influences the activation of a heat-inducible flower-specific system for genetic containment in poplar (Populus tremula L.). Plant Cell Rep. 35, 369–384. doi: 10.1007/s00299-015-1890-x
Hoenicka, H., Nowitzki, O., Hanelt, D., and Fladung, M. (2008). Heterologous overexpression of the birch FRUITFULL-like MADS-box gene BpMADS4 prevents normal senescence and winter dormancy in Populus tremula L. Planta 227, 1001–1011. doi: 10.1007/s00425-007-0674-0
Hsu, C.-Y., Adams, J. P., Kim, H., No, K., Ma, C., Strauss, S. H., et al. (2011). FLOWERING LOCUS T duplication coordinates reproductive and vegetative growth in perennial poplar. Proc. Natl. Acad. Sci. U.S.A. 108, 10756–10761. doi: 10.1073/pnas.1104713108
Huang, J., Smith, A. R., Zhang, T., and Zhao, D. (2016). Creating completely both male and female sterile plants by specifically ablating microspore and megaspore mother cells. Front. Plant Sci. 7:30. doi: 10.3389/fpls.2016.00030
Ishii, T., and Araki, M. (2017). A future scenario of the global regulatory landscape regarding genome-edited crops. GM Crops Food 8, 44–56. doi: 10.1080/21645698.2016.1261787
Jaeger, K. E., Pullen, N., Lamzin, S., Morris, R. J., and Wigge, P. A. (2013). Interlocking feedback loops govern the dynamic behavior of the floral transition in Arabidopsis. Plant Cell 25, 820–833. doi: 10.1105/tpc.113.109355
Jansson, S., and Douglas, C. J. (2007). Populus: a model system for plant biology. Annu. Rev. Plant Biol. 58, 435–458. doi: 10.1146/annurev.arplant.58.032806.103956
Karlgren, A., Gyllenstrand, N., Clapham, D., and Lagercrantz, U. (2013). FLOWERING LOCUS T/TERMINAL FLOWER1-like genes affect growth rhythm and bud set in Norway spruce. Plant Physiol. 163, 792–803. doi: 10.1104/pp.113.224139
Karlgren, A., Gyllenstrand, N., Källman, T., Sundström, J. F., Moore, D., Lascoux, M., et al. (2011). Evolution of the PEBP gene family in plants: functional diversification in seed plant evolution. Plant Physiol. 156, 1967–1977. doi: 10.1104/pp.111.176206
Katahata, S.-I., Futamura, N., Igasaki, T., and Shinohara, K. (2014). Functional analysis of SOC1-like and AGL6-like MADS-box genes of the gymnosperm Cryptomeria japonica. Tree Genet. Genomes 10, 317–327. doi: 10.1007/s11295-013-0686-9
Kazana, V., Tsourgiannis, L., Iakovoglou, V., Stamatiou, C., Alexandrov, A., Araújo, S., et al. (2015). Public attitudes towards the use of transgenic forest trees: a cross-country pilot survey. iForest-Biogeosci. For. 9:344.
Klintenas, M., Pin, P. A., Benlloch, R., Ingvarsson, P. K., and Nilsson, O. (2012). Analysis of conifer FLOWERING LOCUS T/TERMINAL FLOWER1-like genes provides evidence for dramatic biochemical evolution in the angiosperm FT lineage. New Phytol. 196, 1260–1273. doi: 10.1111/j.1469-8137.2012.04332.x
Klocko, A. L., Brunner, A. M., Huang, J., Meilan, R., Lu, H., Ma, C., et al. (2016a). Containment of transgenic trees by suppression of LEAFY. Nat. Biotechnol. 34:918. doi: 10.1038/nbt.3636
Klocko, A. L., Ma, C., Robertson, S., Esfandiari, E., Nilsson, O., and Strauss, S. H. (2016b). FT overexpression induces precocious flowering and normal reproductive development in Eucalyptus. Plant Biotechnol. J. 14, 808–819. doi: 10.1111/pbi.12431
Klocko, A. L., Lu, H., Magnuson, A., Brunner, A. M., Ma, C., and Strauss, S. H. (2018). Phenotypic expression and stability in a large-scale field study of genetically engineered poplars containing sexual containment transgenes. Front. Bioeng. Biotechnol. 6:100. doi: 10.3389/fbioe.2018.00100
Kramer, R. D., Sillett, S. C., and Carroll, A. L. (2014). Structural development of redwood branches and its effects on wood growth. Tree Physiol. 34, 314–330. doi: 10.1093/treephys/tpu011
Lännenpää, M., Hassinen, M., Ranki, A., Hölttä-Vuori, M., Lemmetyinen, J., Keinonen, K., et al. (2005). Prevention of flower development in birch and other plants using a BpFULL1:: BARNASE construct. Plant Cell Rep. 24, 69–78. doi: 10.1007/s00299-004-0903-y
Lemmetyinen, J., Keinonen, K., and Sopanen, T. (2004). Prevention of the flowering of a tree, silver birch. Mol. Breed. 13, 243–249. doi: 10.1023/b:Molb.0000022525.96200.53
Li, J., Brunner, A. M., Shevchenko, O., Meilan, R., Ma, C., Skinner, J. S., et al. (2008). Efficient and stable transgene suppression via RNAi in field-grown poplars. Transgen. Res. 17, 679–694. doi: 10.1007/s11248-007-9148-1
Liu, Y. Y., Yang, K. Z., Wei, X. X., and Wang, X. Q. (2016). Revisiting the phosphatidylethanolamine-binding protein (PEBP) gene family reveals cryptic FLOWERING LOCUS T gene homologs in gymnosperms and sheds new light on functional evolution. New Phytol. 212, 730–744. doi: 10.1111/nph.14066
Liu, Z., and Liu, Z. (2008). The second intron of AGAMOUS drives carpel-and stamen-specific expression sufficient to induce complete sterility in Arabidopsis. Plant Cell Rep. 27, 855–863. doi: 10.1007/s00299-008-0511-3
Liu, Z. -X., Xiong, H. -Y., Li, L. -Y., and Fei, Y. -J. (2018). Functional conservation of an AGAMOUS orthologous gene controlling reproductive organ development in the gymnosperm species Taxus chinensis var. mairei. J. Plant Biol. 61, 50–59. doi: 10.1007/s12374-017-0154-4
Lottmann, J., O’Callaghan, M., Baird, D., and Walter, C. (2010). Bacterial and fungal communities in the rhizosphere of field-grown genetically modified pine trees (Pinus radiata D.). Environ. Biosaf. Res. 9, 25–40. doi: 10.1051/ebr/2010007
Lu, H., and Strauss, S. H. (2018). “Cross-suppression of AG and AG-LIKE 11 genes gives sterility in field grown poplar,” in Proceedings of PAG XXVI, San Diego.
Luis, S. D. B., and José, C. (2014). Costs of female reproduction in a conifer tree: a whole-tree level assessment. J. Ecol. 102, 1310–1317. doi: 10.1111/1365-2745.12283
Mansoor, S., Amin, I., Hussain, M., Zafar, Y., and Briddon, R. W. (2006). Engineering novel traits in plants through RNA interference. Trends Plant Sci. 11, 559–565. doi: 10.1016/j.tplants.2006.09.010
Mariani, C., De Beuckeleer, M., Truettner, J., Leemans, J., and Goldberg, R. B. (1990). Induction of male sterility in plants by a chimaeric ribonuclease gene. Nature 347:737. doi: 10.1038/347737a0
Meilan, R. (1997). Floral induction in woody angiosperms. New For. 14, 179–202. doi: 10.1023/A:1006560603966
Melzer, R., Wang, Y. Q., and Theissen, G. (2010). The naked and the dead: the ABCs of gymnosperm reproduction and the origin of the angiosperm flower. Semin. Cell Dev. Biol. 21, 118–128. doi: 10.1016/j.semcdb.2009.11.015
Mitsuda, N., Hiratsu, K., Todaka, D., Nakashima, K., Yamaguchi-Shinozaki, K., and Ohme-Takagi, M. (2006). Efficient production of male and female sterile plants by expression of a chimeric repressor in Arabidopsis and rice. Plant Biotechnol. J. 4, 325–332. doi: 10.1111/j.1467-7652.2006.00184.x
Mohamed, R., Wang, C. T., Ma, C., Shevchenko, O., Dye, S. J., Puzey, J. R., et al. (2010). Populus CEN/TFL1 regulates first onset of flowering, axillary meristem identity and dormancy release in Populus. Plant J. 62, 674–688. doi: 10.1111/j.1365-313X.2010.04185.x
Mouradov, A., Glassick, T. V., Hamdorf, B. A., Murphy, L. C., Marla, S. S., Yang, Y., et al. (1998). Family of MADS-Box genes expressed early in male and female reproductive structures of monterey pine. Plant Physiol. 117, 55–62. doi: 10.1104/pp.117.1.55
Nature Biotechnology News (2015). Brazil approves transgenic eucalyptus. Nat. Biotechnol. 33:577. doi: 10.1038/nbt0615-577c
Nekrasov, V., Wang, C., Win, J., Lanz, C., Weigel, D., and Kamoun, S. (2017). Rapid generation of a transgene-free powdery mildew resistant tomato by genome deletion. Sci. Rep. 7:482. doi: 10.1038/s41598-017-00578-x
Nilsson, L., Carlsbecker, A., Sundas-Larsson, A., and Vahala, T. (2007). APETALA2 like genes from Picea abies show functional similarities to their Arabidopsis homologues. Planta 225, 589–602. doi: 10.1007/s00425-006-03741
Nuñez, M. A., Chiuffo, M. C., Torres, A., Paul, T., Dimarco, R. D., Raal, P., et al. (2017). Ecology and management of invasive Pinaceae around the world: progress and challenges. Biol. Invasions 19, 3099–3120. doi: 10.1007/s10530-017-1483-4
Nystedt, B., Street, N. R., Wetterbom, A., Zuccolo, A., Lin, Y. C., Scofield, D. G., et al. (2013). The Norway spruce genome sequence and conifer genome evolution. Nature 497, 579–584. doi: 10.1038/nature12211
NZ-EPA (2008). Annual Report for Field Test GMF99001. Available at: https://www.epa.govt.nz/assets/FileAPI/hsno-ar/GMF99001/GMF99001-2008-GMF99001-5-annual-report.pdf [accessed 14/08/2018].
Obeso, J. R. (2002). The costs of reproduction in plants. New Phytol. 155, 321–348. doi: 10.1046/j.1469-8137.2002.00477.x
Pajoro, A., Biewers, S., Dougali, E., Leal Valentim, F., Mendes, M. A., Porri, A., et al. (2014). The (r)evolution of gene regulatory networks controlling Arabidopsis plant reproduction: a two-decade history. J. Exp. Bot. 65, 4731–4745. doi: 10.1093/jxb/eru233
Palmiter, R. D., Behringer, R. R., Quaife, C. J., Maxwell, F., Maxwell, I. H., and Brinster, R. L. (1987). Cell lineage ablation in transgenic mice by cell-specific expression of a toxin gene. Cell 50, 435–443. doi: 10.1016/0092-8674(87)90497-1
Parmentier-Line, C. M., and Coleman, G. D. (2016). Constitutive expression of the Poplar FD-like basic leucine zipper transcription factor alters growth and bud development. Plant Biotechnol. J. 14, 260–270. doi: 10.1111/pbi.12380
Pharis, R. P., Ruddat, M. D., Phillips, C. C., and Heftmann, E. (1965). Precocious flowering of Arizona cypress with gibberellin. Can. J. Bot. 43, 923–927. doi: 10.1139/b65-102
Pharis, R. P., Webber, J. E., and Ross, S. D. (1987). The promotion of flowering in forest trees by gibberellin A47 and cultural treatments: a review of the possible mechanisms. For. Ecol. Manage. 19, 65–84. doi: 10.1016/0378-1127(87)90012-0
Porth, I., and El-Kassaby, Y. A. (2014). Current status of the development of genetically modified (GM) forest trees world-wide: a comparison with the development of other GM plants in agriculture. CAB Rev. 9:12. doi: 10.1079/PAVSNNR20149008
Richardson, D. M., and Rejmánek, M. (2004). Conifers as invasive aliens: a global survey and predictive framework. Divers. Distribut. 10, 321–331. doi: 10.1111/j.1366-9516.2004.00096.x
Ross, S., and Pharis, R. (1985). Promotion of Flowering in Tree Crops: Different Mechanisms and Techniques, with Special Reference to Conifers. Dorset: Institute of Terrestrial Ecology.
Rudall, P. J., Hilton, J., Vergara-Silva, F., and Bateman, R. M. (2011). Recurrent abnormalities in conifer cones and the evolutionary origins of flower-like structures. Trends Plant Sci. 16, 151–159. doi: 10.1016/j.tplants.2010.11.002
Rutledge, R., Regan, S., Nicolas, O., Fobert, P., Côté, C., Bosnich, W., et al. (1998). Characterization of an AGAMOUS homologue from the conifer black spruce (Picea mariana) that produces floral homeotic conversions when expressed in Arabidopsis. Plant J. 15, 625–634. doi: 10.1046/j.1365-313x.1998.00250.x
Sala, A., Hopping, K., McIntire, E. J., Delzon, S., and Crone, E. E. (2012). Masting in whitebark pine (Pinus albicaulis) depletes stored nutrients. New Phytol. 196, 189–199. doi: 10.1111/j.1469-8137.2012.04257.x
Shindo, S., Sakakibara, K., Sano, R., Ueda, K., and Hasebe, M. (2001). Characterization of a FLORICAULA/LEAFY homologue of Gnetum parvifolium and its implications for the evolution of reproductive organs in seed plants. Int. J. Plant Sci. 162, 1199–1209. doi: 10.1086/323417
Shiokawa, T., Yamada, S., Futamura, N., Osanai, K., Murasugi, D., Shinohara, K., et al. (2008). Isolation and functional analysis of the CjNdly gene, a homolog in Cryptomeria japonica of FLORICAULA/LEAFY genes. Tree Physiol. 28, 21–28. doi: 10.1093/treephys/28.1.21
Simak, M. (1978). “Precocius flower induction in Pinus sylvestris by grafting,” Proceedings of the Symposium on Flowering and Seed Development in Trees, Starkville, 63–76.
Simberloff, D., Nunez, M. A., Ledgard, N. J., Pauchard, A., Richardson, D. M., Sarasola, M., et al. (2010). Spread and impact of introduced conifers in South America: lessons from other southern hemisphere regions. Austral Ecol. 35, 489–504. doi: 10.1111/j.1442-9993.2009.02058.x
Souza, A. J. D., Mendes, B. M. J., and Mourão Filho, F. D. A. A. (2007). Gene silencing: concepts, applications, and perspectives in woody plants. Sci. Agric. 64, 645–656. doi: 10.1590/S0103-90162007000600014
Strauss, S. H., Jones, K. N., Lu, H., Petit, J. D., Klocko, A. L., Betts, M. G., et al. (2017). Reproductive modification in forest plantations: impacts on biodiversity and society. New Phytol. 213, 1000–1021. doi: 10.1111/nph.14374
Strauss, S. H., Ma, C., Ault, K., and Klocko, A. L. (2016). “Lessons from two decades of field trials with genetically modified trees in the USA: biology and regulatory compliance,” in Biosafety of Forest Transgenic Trees, eds C. Vettori, F. Gallardo, H. Häggman, V. Kazana, F. Migliacci, G. Pilate, et al. (Dordrecht: Springer), 101–124.
Strauss, S. H., Rottmann, W. H., Brunner, A. M., and Sheppard, L. A. (1995). Genetic engineering of reproductive sterility in forest trees. Mol. Breed. 1, 5–26. doi: 10.1007/BF01682086
Sundström, J., and Engström, P. (2002). Conifer reproductive development involves B-type MADS-box genes with distinct and different activities in male organ primordia. Plant J. 31, 161–169. doi: 10.1046/j.1365-313X.2002.01343.x
Tandre, K., Albert, V. A., Sundås, A., and Engström, P. (1995). Conifer homologues to genes that control floral development in angiosperms. Plant Mol. Biol. 27, 69–78. doi: 10.1007/BF00019179
Tandre, K., Svenson, M., Svensson, M. E., and Engström, P. (1998). Conservation of gene structure and activity in the regulation of reproductive organ development of conifers and angiosperms. Plant J. 15, 615–623. doi: 10.1046/j.1365-313x.1998.00236.x
Tang, W., and Newton, R. J. (2003). Genetic transformation of conifers and its application in forest biotechnology. Plant Cell Rep. 22, 1–15. doi: 10.1007/s00299-003-0670-1
Taniguchi, T. (2018). “Biotechnological research in Cryptomeria japonica,” in Biofuels: Greenhouse Gas Mitigation and Global Warming: Next Generation Biofuels and Role of Biotechnology, eds A. Kumar, S. Ogita and Y.-Y. Yau (New Delhi: Springer), 357–374. doi: 10.1007/978-81-322-3763-1_20
Trontin, J.-F., Walter, C., Klimaszewska, K., Park, Y.-S., and Lelu-Walter, M.-A. (2007). Recent progress in genetic transformation of four Pinus spp. Trans. Plant J. 1, 314–329.
Tylewicz, S., Tsuji, H., Miskolczi, P., Petterle, A., Azeez, A., Jonsson, K., et al. (2015). Dual role of tree florigen activation complex component FD in photoperiodic growth control and adaptive response pathways. Proc. Natl. Acad. Sci. U.S.A. 112, 3140–3145. doi: 10.1073/pnas.1423440112
Uddenberg, D., Akhter, S., Ramachandran, P., Sundström, J. F., and Carlsbecker, A. (2015). Sequenced genomes and rapidly emerging technologies pave the way for conifer evolutionary developmental biology. Front. Plant Sci. 6:970. doi: 10.3389/fpls.2015.00970
Uddenberg, D., Reimegard, J., Clapham, D., Almqvist, C., von Arnold, S., Emanuelsson, O., et al. (2013). Early cone setting in Picea abies acrocona is associated with increased transcriptional activity of a MADS box transcription factor. Plant Physiol. 161, 813–823. doi: 10.1104/pp.112.207746
Velázquez, K., Agüero, J., Vives, M. C., Aleza, P., Pina, J. A., Moreno, P., et al. (2016). Precocious flowering of juvenile citrus induced by a viral vector based on Citrus leaf blotch virus: a new tool for genetics and breeding. Plant Biotechnol. J. 14, 1976–1985. doi: 10.1111/pbi.12555
Vining, K. J., Romanel, E., Jones, R. C., Klocko, A., Alves-Ferreira, M., Hefer, C. A., et al. (2015). The floral transcriptome of Eucalyptus grandis. New Phytol. 206, 1406–1422. doi: 10.1111/nph.13077
Wagner, A., Donaldson, L., Kim, H., Phillips, L., Flint, H., Steward, D., et al. (2009). Suppression of 4-coumarate-CoA ligase in the coniferous gymnosperm Pinus radiata. Plant Physiol. 149, 370–383. doi: 10.1104/pp.108.125765
Wagner, A., Phillips, L., Narayan, R. D., Moody, J. M., and Geddes, B. (2005). Gene silencing studies in the gymnosperm species Pinus radiata. Plant Cell Rep. 24, 95–102. doi: 10.1007/s00299-004-0911-y
Waltz, E. (2016). CRISPR-Edited Crops Free to Enter Market, Skip Regulation. London: Nature Publishing Group.
Waltz, E. (2018). With a free pass. CRISPR-edited plants reach market in record time. Nat. Biotechnol. 36:6. doi: 10.1038/nbt0118-6b
Wang, X., Singer, S. D., and Liu, Z. (2012). Silencing of meiosis-critical genes for engineering male sterility in plants. Plant Cell Rep. 31, 747–756. doi: 10.1007/s00299-011-1193-9
Williams, C. G. (2005). Framing the issues on transgenic forests. Nat. Biotechnol. 23:530. doi: 10.1038/nbt0505-530
Williams, C. G. (2009). Conifer Reproductive Biology. Berlin: Springer Science & Business Media. doi: 10.1093/aob/mcq081
Wilson, V. R., and Owens, J. N. (2003). Histology of sterile male and female cones in Pinus monticola (western white pine). Sex. Plant Reproduct. 15, 301–310.
WWF (2015). WWF Living Forests Report: Chapter 5: Saving Forests at Risk. Gland: WWF – World Wide Fund for Nature. Available at: http://awsassets.panda.org/downloads/living_forests_report_chapter_5_1.pdf [accessed May 2018].
Yamagishi, N., Li, C., and Yoshikawa, N. (2016). Promotion of flowering by apple latent spherical virus vector and virus elimination at high temperature allow accelerated breeding of apple and pear. Front. Plant Sci. 7:171. doi: 10.3389/fpls.2016.00171
Zahn, L. M., Kong, H., Leebens-Mack, J. H., Kim, S., Soltis, P. S., Landherr, L. L., et al. (2005). The evolution of the SEPALLATA subfamily of MADS-box genes: a preangiosperm origin with multiple duplications throughout angiosperm history. Genetics 169, 2209–2223. doi: 10.1534/genetics.104.037770
Zhang, C., Norris-Caneda, K. H., Rottmann, W. H., Gulledge, J. E., Chang, S., Kwan, B. Y.-H., et al. (2012). Control of pollen-mediated gene flow in transgenic trees. Plant Physiol. 159, 1319–1334. doi: 10.1104/pp.112.197228
Keywords: sterility, reproduction, forest trees, gene editing, genetic engineering, containment
Citation: Fritsche S, Klocko AL, Boron A, Brunner AM and Thorlby G (2018) Strategies for Engineering Reproductive Sterility in Plantation Forests. Front. Plant Sci. 9:1671. doi: 10.3389/fpls.2018.01671
Received: 12 August 2018; Accepted: 26 October 2018;
Published: 15 November 2018.
Edited by:
Steven Henry Strauss, Oregon State University, United StatesReviewed by:
Ove Nilsson, Umeå Plant Science Centre, SwedenCopyright © 2018 Fritsche, Klocko, Boron, Brunner and Thorlby. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Glenn Thorlby, Z2xlbm4udGhvcmxieUBzY2lvbnJlc2VhcmNoLmNvbQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.