 Nazareth Torres
Nazareth Torres M. Carmen Antolín
M. Carmen Antolín Nieves Goicoechea
Nieves Goicoechea- Unidad Asociada al CSIC (EEAD, Zaragoza, ICVV, Logroño), Grupo de Fisiología del Estrés en Plantas (Departamento de Biología Ambiental), Facultades de Ciencias y Farmacia y Nutrición, Universidad de Navarra, Pamplona, Spain
Climate change and their resulting impacts are becoming a concern for winegrowers due to the high socioeconomic relevance of the winemaking sector worldwide. In fact, the projected climate change is expected to have detrimental impacts on the yield of grapevines, as well as on the quality and properties of grapes and wine. It is well known that arbuscular mycorrhizal fungi (AMF) can improve the nutritional quality of edible parts of crops and play essential roles in the maintenance of host plant fitness under stressed environments, including grapevines. The future scenarios of climate change may also modify the diversity and the growth of AMF in soils as well as the functionality of the mycorrhizal symbiosis. In this review, we summarize recent research progress on the effects of climate change on grapevine metabolism, paying special attention to the secondary compounds involved in the organoleptic properties of grapes and wines and to the levels of the phytohormones implied in the control of berry development and fruit ripening. In this context, the potential role of AMF for maintaining fruit quality in future climate change scenarios is discussed.
General Introduction
Grapevine is an important perennial crop worldwide, consumed as fresh or dried (raisins) fruit or produced to make mainly wine, but also grape juice and vinegars. Grapes can also be used for elaborating sweet spreads (marmalade, jelly, butter, and jam). The production of grapevine in Europe achieved of 23.4 million tons (Eurostat Statistics Division, 2016), which represents the 39% of the world production. The moderate consumption of wine (Georgiev et al., 2014; Artero et al., 2015) and the inclusion of grapes and grape products in the diet (Vislocky and Fernandez, 2010) may report beneficial effects for human health because it can positively influence risk factors associated with cardiovascular health, cancer, neurodegenerative disease, and age-related cognitive decline. However, projected climate change is expected to have detrimental impacts on the grapes and wine quality and properties because this crop is highly dependent upon weather conditions during the growing season. The impact of elevated temperatures is one of the environmental factors that most influence both primary and secondary metabolisms and, consequently, the quality of grape berries (Torres et al., 2017).
Mutualistic plant–microbe interactions offer a novel approach to enhance agricultural productivity while reducing environmental costs (Hamilton et al., 2016). Numerous studies have demonstrated that (1) climate change may affect all types of beneficial plant-microorganism interactions and (2) plant-associated microorganisms are an important factor modulating the response of plants to climate change (Compant et al., 2010). Among these beneficial microorganisms, it is worth noting the arbuscular mycorrhizal fungi (AMF), because they can establish mutualistic association with most crops (Smith and Read, 2008) and have an increasingly important role in vineyard production systems (Trouvelot et al., 2015). Although some aspects concerning the effects of global climate change on grapevine cultivation were recently reviewed (Alonso et al., 2016b; Costa et al., 2016; Mosedale et al., 2016; Schultz, 2016; van Leeuwen and Darriet, 2016) so far, the mycorrhizal-mediated responses to grapevine under changing environments have not been updated. In this review, we summarize recent research progress dealing with the effects of climate change on grapevine metabolism and we discuss if the benefits of mycorrhizal symbiosis on berry quality could be maintained under the predicted environmental conditions in future climate change scenarios.
Climate Change Scenarios for Viticulture
The increasing interest on the effect of climate change on viticulture is not surprising due to the high socioeconomic relevance of the winemaking sector worldwide. Furthermore, climate change and their resulting impacts are becoming a concern for winegrowers (Fraga et al., 2013; Neethling et al., 2017). For most wine-production regions of the world, long-term climate records have shown rising temperatures (Webb et al., 2013; Barnuud et al., 2014; Fraga et al., 2016) together with shifting patterns in rainfall and extreme weather events (Andrade et al., 2012; Intergovernmental Panel on Climate Change, [IPCC], 2014). Warming temperatures have been linked to anthropogenic climate change and are likely to continue. Thus, in most scenarios without additional mitigation efforts, atmospheric concentrations of CO2 could reach about 1000 ppm by 2100 and, thus, air temperature is likely to exceed 4°C above pre-industrial levels (Intergovernmental Panel on Climate Change, [IPCC], 2014). This rise of temperature is expected to exert detrimental impacts on grapevine physiology and, consequently, on berry quality (Mosedale et al., 2016), in addition to increase the risk of pests and diseases, especially in areas with warm winters (Caffarra et al., 2012). Heat waves during the growing season enhance the loss of water from the soil thus impairing vine productivity (Schultz, 2016). Moreover, the absence of precipitation, considered as a major limiting factor for plant growth, is frequently accompanied by increased UV-B radiation levels (Bandurska et al., 2013). This new scenario can entail future wine production in areas that are presently too cold for wine cultivation, while the actual grape growing regions may become unsuitable for premium wine production (Hannah et al., 2013; Roy et al., 2017). The establishment of vineyards at higher altitudes (colder areas) and the selection of exposures that lead to a lower interception of solar radiation would be some appropriated options facing these changes (Hannah et al., 2013; Palliotti et al., 2014). On the other hand, in south Mediterranean Europe, climate may limit grapevine yield and berry quality because most of the berry growth and ripening period occurs under conditions of high air temperature and soil water deficit. For this reason, irrigation is expanded fast in this region to mitigate environmental stress and to guarantee stable grape yield and quality (Costa et al., 2016; Resco et al., 2016). Indeed, future strategies to optimize the environmental performance of the viticulture in the Mediterranean region will have to be focused on an adequate selection of rootstocks and phenotypes resistant to drought and heat stress, although currently winegrowers identify them as the last resort strategies (Neethling et al., 2017). Unfortunately, in Mediterranean countries (e.g., Spain) these strategies have been carried out in a non-climate-smart way, this causing that almost a third of the 1 million hectares of grapevines in the Spanish territory will face a different climate to which they were planned (Resco et al., 2016).
Effects of Changing Environments on the Arbuscular Mycorrhizal Fungi
The plant growth-promoting microorganisms could play vital roles in the maintenance of plant fitness and soil health under stressed environments (Vimal et al., 2017). Climate change related factors may lead to increased C allocation to the root zone, which will potentially alter the composition of root exudates, the C/N ratio or the nutrient availability, with the consequent influence on the composition, abundance and/or activity of plant-associated microbial communities. The data collected by Compant et al. (2010) in their review demonstrated that warming, elevated CO2 and drought affect plant-beneficial microorganisms in many ways, the effects being dependent on the climate change factor studied, plant species, ecosystem type, soil type and microbial genotype.
Mohan et al. (2014) summarized the results obtained in research studies dealing with the effect of rising temperatures on mycorrhizal communities. While in 17% of such studies mycorrhizal abundance decreased, in 20% of them no significant change was observed and the 63% of the works concluded that the abundance of mycorrhizas can increase under elevated temperature. In subsequent studies, Augé et al. (2015) pointed out that AMF promotion was 10% higher when air temperatures were kept at or below 27°C than those that exceeded 27°C. Moreover, Wilson et al. (2016) concluded that the direct effect of increasing 3°C the temperature decreases AMF colonization, and this appeared to be regionally consistent across the Mediterranean climate gradient. On the other hand, although the growth of external hyphae and the diversity of AMF species can increase at high temperatures (Hawkes et al., 2008; Zhang et al., 2016), the mycorrhizal activity generally decreases (Mohan et al., 2014). Furthermore, in a warmer world, the presumed enhanced growth of AMF hyphae is unlikely to balance the carbon losses to the atmosphere from the AMF respiration (Hawkes et al., 2008).
Elevated CO2 can promote mycorrhizal colonization of plant roots as a consequence of the enhanced carbon allocation to roots (Zhu et al., 2016; Asha et al., 2017), which may result in an increased mineral uptake from soil but not necessarily correlated with nutrient transfer to the host plant (Smith and Read, 2008). Regarding the community composition of AMF, elevated CO2 increased the ratio of Glomeraceae to Gigasporaceae but this effect may be masked by the natural changes through time (Cotton et al., 2015) and also by the dependency of different fungi on water availability and precipitation (Veresoglou et al., 2016). The application of molecular tools revealed that, at present, Glomeraceae dominate the composition of the AMF communities in vineyards in Oregon (United States) (Schreiner and Mihara, 2009), Piedmont (Italy) (Balestrini et al., 2010) and Burgundy (France) (Bouffaud et al., 2016), being the AMF diversity relatively low, which contrasts with the high diversity of AMF in the rhizosphere of European wild grapevine V. vinifera L. ssp. sylvestris (Gmelin) Hegi found by Ocete et al. (2015). Projected droughts within the climate change are expected to cause AMF sporulation and reduction of the AMF activity (Guadarrama et al., 2014). Agricultural practices (high fertilizer inputs, tillage, weed control practices, and pest management practices, among others) may amplify the effect of environmental factors on the AMF communities present in vineyard soils (Trouvelot et al., 2015; Vukicevich et al., 2018). In semi-arid to arid regions, such as Mediterranean areas, soils of vineyards are periodically subjected to tillage or left totally bared in order to keep the soil free of weeds and grassland plant species, which influences on the development and reproduction of AMF. Oehl and Koch (2018) noted that AMF diversity decreased in Central European vineyards subjected to this cultivation management, which can in last instance affect yield and wine quality parameters.
A better understanding of the effect of climatic variability on the synchrony of plants and soil microorganisms which play a key role in the cycle of nutrients and disease cycles is necessary (Pritchard, 2011) and further research on the mechanisms involved in plant-microorganism interactions is required for developing new strategies to manage sustainable agriculture under stressful weather conditions (Vimal et al., 2017).
Metabolism in Grapevine Berries Under Challenging Environments
Primary and secondary metabolites in grapevine berries are directly involved in the organoleptic properties of grapes and wines (Conde et al., 2007). Climate change is particularly important for berry quality. In fact, drought and light intensity are just some environmental factors that dramatically affect phenolic metabolism and berry chemical composition.
Warming temperatures hasten sugar accumulation and delay color development due to reduction of anthocyanin content (Table 1). This decoupling was explained by a relative shift in onset rather than rate of accumulation of these berry components leading to the elaboration of wines with higher alcohol contents (Sadras and Moran, 2012). Moreover, it has been reported that elevated temperatures resulted in higher proportion of acylated anthocyanins (De Rosas et al., 2017). These authors showed that pigment modifications under high temperature are regulated at transcriptional level by MYBA1 transcription factor, and by the UDP glucose: flavonoid-3-O-glucosyltransferase and anthocyanin acyltransferase genes. Berry acidity is another important quality trait dependent on the ratio of the concentration between free organic acids (mainly, malic and tartaric acids) and their potassium salt forms. Organic acid metabolism, and especially malic acid concentration, is highly responsive to warm temperatures during fruit ripening. High temperatures are known to induce the degradation of malic acid (Sweetman et al., 2014). Tartaric acid has been thought to be more stable than malic acid; however, several discrepancies were found when the impact of temperature on tartaric acid was studied (Table 1). Recent findings of Cholet et al. (2016) showed two groups of expression profiles for the genes involved in the biosynthetic pathway of tartaric acid: those upstream of ascorbic acid, belonging to the Smirnoff-Wheeler pathway and those downstream of ascorbic acid. This study proposed that both groups of genes might be modulated by different environmental factors, which could aid to explain the above mentioned discrepancies on tartaric acid content.
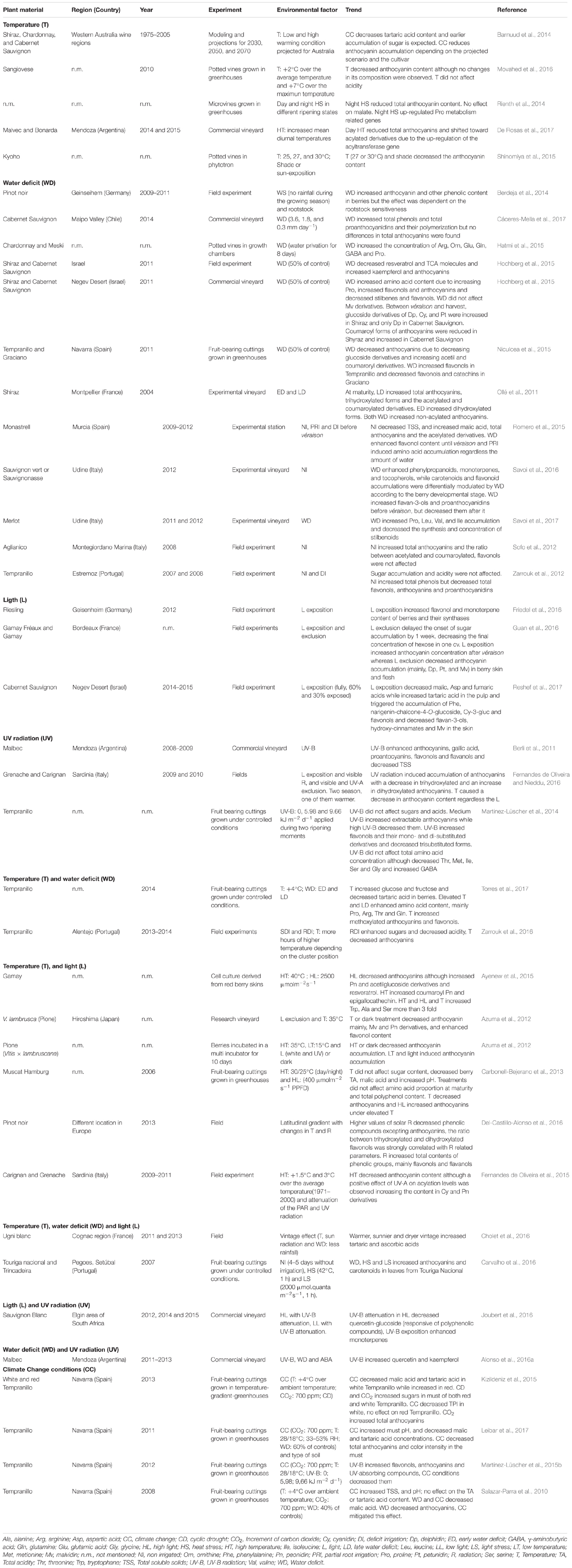
TABLE 1. Berry metabolism response to environmental factors.
Several studies have pointed out the role of water deficit on berry quality enhancing total phenolics, particularly, anthocyanins (Table 1). Moreover, it has been reported that water deficit changed anthocyanin composition, as well as the composition and the accumulation of flavonols or proanthocyanidins. In general, water deficit enhanced phenylpropanoids, monoterpenes, and tocopherols, while carotenoids and flavonoid accumulations were differentially modulated by water stress according to the berry developmental stage (Savoi et al., 2016), grapevine variety (Niculcea et al., 2015; Kizildeniz et al., 2015) and/or deficit irrigation program applied (Table 1). Recent findings showed that the effects of deficit irrigation on berry composition were attenuated at high temperature and that both factors (temperature and deficit irrigation) contributed to modify metabolite profiles of amino acids, anthocyanins and flavonols in Tempranillo variety (Torres et al., 2017). Thus, the combination of elevated temperature and deficit irrigation resulted in high amino acid content mainly due to the accumulation of arginine, proline, threonine, and glutamine. The high arginine and proline contents could be related to a transcriptional regulation of ornithine decarboxylase during water deficit (Berdeja et al., 2015), which could be exacerbated under warm temperatures. Torres et al. (2017) also showed that both temperature and irrigation modified anthocyanin profiles by increasing 3-acetyl-glucosides derivatives due to increased methoxylated forms (petunidin and malvidin). Berry skin flavonols were dominated by myricetin-3-O-glucoside but the changes in flavonol profiles were more pronounced at elevated temperatures when plants were subjected to the deficit irrigation. These changes on secondary metabolite profiles could be explained by the regulation at the transcriptional level of phenylpropanoid pathway genes that takes place during water deficit (Castellarin et al., 2007; Deluc et al., 2009) and under elevated temperatures (De Rosas et al., 2017).
Exposure to visible and/or UV radiation is a key factor in the synthesis of phenols and their accumulation in berries. On this subject, several studies have reported increasing trends in flavonols, anthocyanins, flavanols, monoterpenes, and decreases in hydroxy-cinnamates and flavan-3-ols in response to visible, UV-A and/or UV-B radiations (see Table 1 for further details). However, under climate change scenarios (elevated CO2 and temperature) the stimulation of UV-absorbing compound synthesis was reduced (Martínez-Lüscher et al., 2015b). Likewise, the combination of elevated CO2, elevated temperature and drought significantly reduced the phenolic content in the same variety, but no effect was observed when the environmental factors were applied individually (Kizildeniz et al., 2015). These results highlight the importance of approach the combined effects of different environmental factors on berry composition.
Grape and wine quality is extremely dependent on the fruit ripening process. Sensory and nutritional characteristics are crucial aspects for wine market, which are developed during berry ripening under a complex hormonal control. Grape berry development involves a complex series of changes, which can be divided into three major phases. Initial berry growth (Phase I) occurs along a sigmoid growth curve due to cell division and subsequent cell expansion. In this phase, the accumulation of organic acids, proanthocyanidins, and hydroxycinnamic acids starts to peak levels. In Phase II (lag phase), cell expansion ceases and sugars begin to accumulate. The beginning of Phase III is marked by the onset on ripening (véraison), in which berries undergo a second period of sigmoid growth due to mesocarp cell expansion and accumulate anthocyanin pigments. The accumulation of volatile compounds for aroma and sugars takes place in this phase, as well as the decline in organic acid content and the berry softening.
Several hormones participate in the control of grape berry development and ripening, such as auxin (IAA), ethylene, abscisic acid (ABA), gibberellins (GAs), cytokinins (CKs), and brassinosteroids (BRs) (Böttcher and Davies, 2012). To date, ABA has been the hormone most widely studied in berries in relation to environmental stress factors (Table 2). Some studies have reported that berry ABA content diminished under warm temperatures. Likewise, Shinomiya et al. (2015) reported that temperatures exceeding 27°C during the ripening season lead to insufficient berry coloration as a result of low levels of ABA and anthocyanin biosynthetic gene expression levels. Deficit irrigation during growing season also modifies the pattern of hormone accumulation in berries (Table 2). In this regard, Niculcea et al. (2013) showed that sustained deficit irrigation (SDI) caused a decrease in ABA and salicylic acid (SA) at véraison that affected the amount of anthocyanins at harvest. However, these modifications in berry hormonal patterns also depend on timing of deficit irrigation program applied. Thus, Niculcea et al. (2014) reported that both pre- and post-véraison water-deficit modified evolution of ABA, IAA, SA, and JA in berries, which was related to changes in berry size, increases in phenolic substances and accumulation of amines in Tempranillo and Graciano varieties.
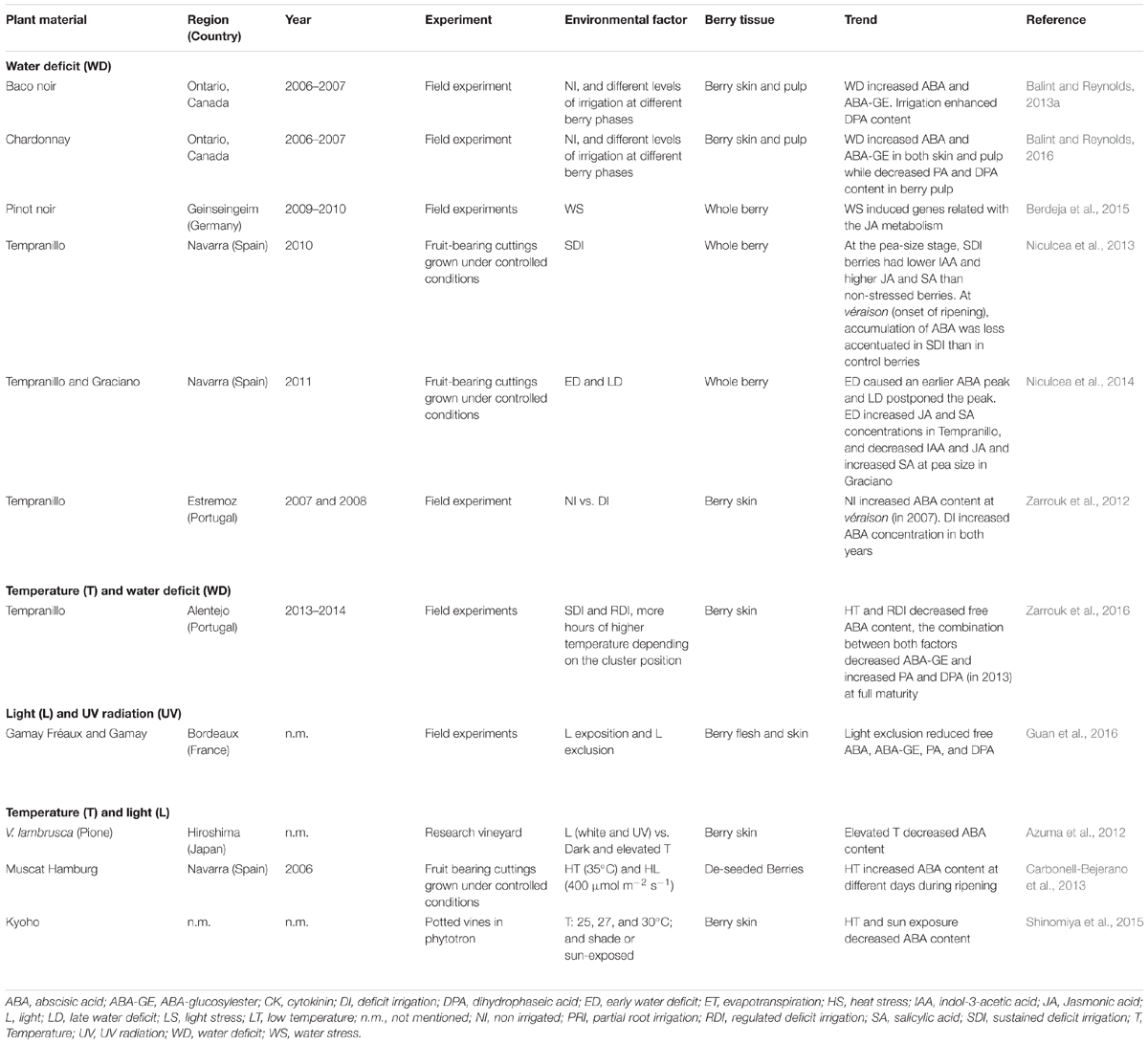
TABLE 2. Berry hormones in response to environmental factors.
Impact of AMF on the Quality of Crops and Fruits Undergoing Changing Environments
The application of mycorrhizal inocula has emerged as a reliable technique to enhance the agricultural productivity and nutritional value of edible vegetables whereas reducing environmental costs (Berruti et al., 2016; Goicoechea and Antolín, 2017). This is the case of strawberry fruits, whose levels of phenolic compounds and minerals increased when plants were inoculated with the arbuscular mycorrhizal fungus Glomus intraradices. Some color parameters of strawberry fruits were also affected by AMF (Castellanos-Morales et al., 2010). Similarly, Hart et al. (2015) found that mycorrhizal inoculation enhanced the concentrations of several minerals (N, P, Cu), carotenoids, and some flavor compounds, as well as the antioxidant capacity in tomato fruits. This beneficial effect of AMF on the quality of tomatoes was corroborated by Bona et al. (2017) in a field study performed in a real industrial tomato farm. In another study carried out under nature conditions, Zeng et al. (2014) found that Glomus versiforme improved the quality of citrus fruits by increasing the ratio of sugar to acid, and the amounts of vitamin C, flavonoids and minerals. Another woody plant beneficed by mycorrhization in field is Libidibia ferrea, a tree with medicinal properties whose bark accumulated higher amount of flavonoids and tannins when associated with AMF (dos Santos et al., 2017). Mycorrhizal fungi also improved the antioxidant potential of leaves from sweet basil, an aromatic plant widely used for medicinal and cooking purposes (Hristozkova et al., 2017). In lettuce, mycorrhizal symbiosis induced the accumulation of carotenoids, total soluble phenolics, anthocyanins, chlorophylls, tocopherol and some mineral nutrients in leaves (Baslam et al., 2011, 2013a), which makes the application of AMF a feasible tool for improving the nutritional quality of this horticultural crop. This improvement of the quality in lettuces associated with AMF was significant enough to allow extending cultivation of this crop to seasons in which non-mycorrhizal lettuces suffer relevant decreases in their levels of proteins, carotenoids and flavonoids (Baslam et al., 2013b).
However, different factors involved in the expected climate change can modulate or even change the effects of AMF on the metabolism and physiology of their host plants. In fact, drought, salinity, global warming and rising CO2 in the atmosphere affect plant growth and yield and constitute a threat to sustainable agriculture and global food security. In studies focused on the role of mycorrhizal symbiosis when plants are undergoing salt stress, some authors have found that AMF increase the plant salt-tolerance and improve fruit yield and quality. This is the case of cucumber cultivated under saline conditions: fruits produced by mycorrhizal plants had higher amounts of soluble proteins, sugars and vitamin C and lower levels of nitrate than those from non-mycorrhizal plants (Han et al., 2012). Likewise, Huang et al. (2013) measured higher contents of N, P, K, and Ca in tomato fruits of hybrid cultivars associated with AMF than in those of non-inoculated ones. Beneficial effects of mycorrhizal symbiosis on yield and fruit quality of crops under salty conditions, however, can vary depending on plant cultivars and fungal strains, as demonstrated by results of Huang et al. (2013) in tomato and those obtained by Sinclair et al. (2014) working with strawberry. Water deficit is one of the most important factors affecting crop survival, growth, and productivity. Most times the beneficial effect of AMF on the host plant development is more evident when water supply is restricted than under plentiful water availability. For example, the beneficial effect of AMF on the growth and quality of chile ancho pepper fruits was especially clear when plants were undergoing drought conditions: fruits of mycorrhizal plants subjected to water deficit showed similar color intensity and chlorophyll content and higher amount of carotenoids than those of non-mycorrhizal plants cultivated at optimal irrigation regime (Mena-Violante et al., 2006). In lettuce, while a moderate water deficit prolonged in time reinforced the capacity of AMF for increasing the levels of antioxidant compounds in leaves (Baslam and Goicoechea, 2012), elevated CO2 in the atmosphere impaired this beneficial effect probably due to the use of photoassimilates for enhancing growth of the host plant and spreading mycorrhizal colonization in detriment to the secondary metabolism (Baslam et al., 2012). Similarly, Goicoechea et al. (2016) observed a general depletion of contents of micro- and macro-nutrients and gliadins in grains of durum wheat cultivated under elevated CO2 in the air, regardless of mycorrhizal inoculation and water regime applied to plants, what contrasted with the higher accumulation of copper, iron, manganese, zinc and gliadins –wheat-seed storage proteins responsible together with glutenins for dough elasticity and extensibility that determine processing qualities in the production of end products- in grains of durum wheat inoculated with AMF and grown under water deficit at ambient CO2. Notwithstanding the above, elevated CO2 not always nullify the beneficial effect of mycorrhizal colonization on crop quality. For example, in alfalfa, the combination of AMF and elevated atmospheric CO2 improved forage quality by increasing the amount of hemicellulose and decreasing that of lignin in leaves (Baslam et al., 2014). Moreover, the positive effect of the synergism between AMF and elevated atmospheric CO2 may be reinforced by the simultaneous application of some cultural practices, such as the supply of humic substances to the soil. In this sense, Bettoni et al. (2014) concluded that the triple interaction between humic substances application, mycorrhizal inoculation and elevated CO2 enhanced the accumulation of soluble sugars, proteins and proline in leaves of onion seedlings in a greater extent than the application of those factors separately, which increased the quality of onion shoots as source organs for posterior growth and quality of bulbs. These same authors found that the application of humic substances, AMF inoculum and elevated CO2 in the air had an additive effect of increasing the content of soluble sugars, proteins, and phenolics in onion bulbs, thus reinforcing their energetic and antioxidant properties. This triple interaction also enhanced the ratio between soluble solids and total titratable acidity, which may favor the perception of sweetness and make onion more pleasant for consumption (Bettoni et al., 2017).
In addition to the induction of plant defenses, many times through the activation of pathways belonging to the antioxidant and secondary metabolism, Thirkell et al. (2017) suggested that one of the strongest benefits of mycorrhizal symbiosis for crop plants is related to the improved soil properties mediated by AMF. It is known that fungal hyphae increase the mineralisation of soil organic matter (SOM) (Paterson et al., 2016). Moreover, mycorrhizal fungi can enhance the fixation of atmospheric CO2 by their host plants, and then induce the transport of photoassimilates from the aerial part to the roots by exerting a sink effect. A portion of the carbon present in the fungal biomass will remain in the soil as a part of the SOM after the AMF senescence (Treseder, 2016). These benefits exerted by AMF will become especially relevant in the context of the projected loss of soil organic carbon caused by the global warming, which in last instance will decrease agricultural productivity (Wiesmeier et al., 2016). Erosion and low organic matter stocks are common problems affecting soils of vineyards in Mediterranean areas as a consequence of the concurrence of environmental factors and some management practices such as the abovementioned continuous tillage (García-Díaz et al., 2018).
Potentiality of AMF to Mitigate the Negative Impacts of Climate Change on Grapevine Berry Quality
Most studies dealing with the role that mycorrhizal association may play in avoiding or reducing the deleterious effects of the weather conditions predicted for the future have been focused on the secondary metabolism and, mainly, on the phenolic compounds. There is, however, scarce information on the influence of mycorrhizal symbiosis on the levels of phytohormones implied in the berry development and ripening, especially when grapevines are undergoing climatic change scenarios.
Several research works have demonstrated that the association of grapevine with AMF favors the synthesis of plant secondary metabolites -resveratrol, flavonols and anthocyanins, among others-, which are determinant not only for increasing plant tolerance to environmental stresses but also for enhancing berry quality (Table 3). In a recent study, Torres et al. (2016) reported that inoculation of grapevines with AMF might play a relevant role in a future climate-change scenario to maintain or even improve berry quality by improving parameters related to the phenolic maturity, such as anthocyanin content, and by enhancing antioxidant activity. This study also highlighted the different abilities of distinct clones from the same variety of grapevine –Tempranillo- to respond to elevated temperature and AMF inoculation. These intravarietal Tempranillo differences were also observed when studied the effects of elevated air temperature, water deficit and mycorrhizal inoculation, separately or in combination, on fruit quality (Torres et al., 2018). However, in all Tempranillo clones used in this study the loss of anthocyanins in berries from non-mycorrhizal plants grown under elevated temperature and subjected to water deficit from the veraison to the maturity of berries did not occur in the plants inoculated with AMF.
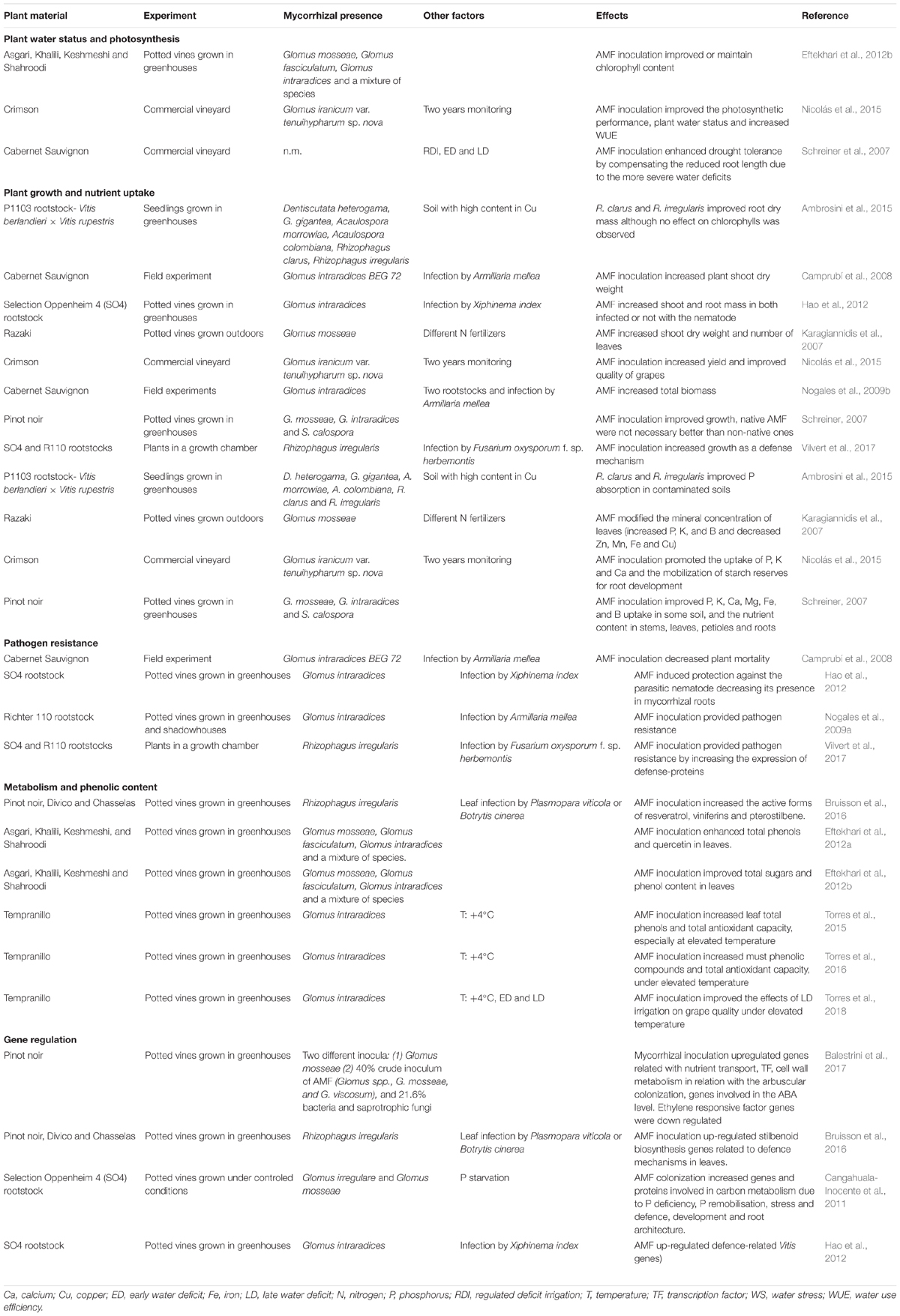
TABLE 3. Benefits of AMF for grapevines.
Summarizing the findings of diverse studies, the mechanisms that may be implied in the higher concentrations of secondary metabolites in tissues of plants associated with AMF are the improved photosynthesis and mineral nutrition of host plants, the activation of pathways belonging to the secondary metabolism, the production of signaling molecules, hormonal modifications and/or higher expression of genes involved in the secondary metabolism (dos Santos et al., 2017). In the case of grapevines, several authors have concluded that the positive effects due to AMF colonization are mediated by the up-regulation of some genes (Table 3). For example, Balestrini et al. (2017) showed that the expression of genes belonging to categories such as nutrient transport, transcription factors, and cell wall-related genes was significantly altered by AMF colonization. Moreover, the presence of AMF in roots of three grapevine varieties stimulated the transcription of the genes that codify the enzymes phenylalanine ammonia-lyase, stilbene synthase, and a resveratrol O-methyltransferase, involved in the response of grapevines to the attack of Plasmopara viticola and Botrytis cinerea (Bruisson et al., 2016).
Conclusion and Perspectives
In the last decades, a growing concern about the potential consequences of climate change on viticulture and the detrimental impact on grape and wine quality has addressed several researches. Nevertheless, few studies have highlighted the role of AMF symbiosis in this scenario, in spite of the known benefits that mycorrhizas provide to host plants. The general utilization of fertilizers and/or phytohormones may damage or unbalance soil ecosystem of viticultural areas, so that their application needs to be reduced. Thus, AMF has been presented as natural biofertilizers that can be the alternative to chemical fertilization without the concomitant loss of crop quality (Berruti et al., 2016) and there is evidence that co-adaptation of the partners to a new environment may maximize benefits and minimize costs of the symbiosis (Johnson et al., 2013). On the other hand, exogenous phytohormone application (especially, ABA) to the vine has been used as a tool to improve the quality of the grapes (Balint and Reynolds, 2013b; Alonso et al., 2016a). Taking into account that under abiotic stress ABA concentration was enhanced by AMF (Wang et al., 2017), the symbiotic association can offer an alternative to phytohormone supply to improve grape quality. However, there is a need for more studies that deepen into the influence of AMF in the levels of the ABA when grapevines undergo challenging environments. Moreover, the fact that the responses of grapevines to the inoculation with AMF and/or to the environmental conditions may vary according to the plant variety or clone (Torres et al., 2015, 2016, 2018) indicates that it may be profitable to identify the AMF inoculants most suitable for a given cultivar in a given environment.
Winegrowers are aware of reconsidering their viticultural practices in order to better manage climate-related risks and produce quality wines (Neethling et al., 2017). Given the global warming impact on berry quality traits, it is useful to reconsider the potential application of some new or traditional management techniques able to regulate sugar accumulation and/or to delay or balance berry ripening (Palliotti et al., 2014). Most wine-producing regions are subjected to seasonal drought but, based on the global climate models an increase in aridity is predicted in the future. Hence, an optimized irrigation schedule would still be one of the most desirable tools to improve crop productivity and fruit quality (Costa et al., 2016). Under low rainfall conditions, warm temperatures and high light intensity, spontaneous vegetation used as groundcover appears as an effective strategy to revert soil degradation in Mediterranean vineyards (García-Díaz et al., 2018) at the same time that it may benefit taste or quality of wines (Trigo-Córdoba et al., 2015). Although the impact of water deficit on berry ripening and quality has been extensively investigated during the last decades, the suitability of actual irrigation programs should be reviewed in the future climate conditions. Consequently, several researches have begun to address the combined effects of elevated temperature, UV-B radiation or high CO2 with water deficit on grapevine quality (Bonada et al., 2015; Kizildeniz et al., 2015; Martínez-Lüscher et al., 2015a; Torres et al., 2016; Zarrouk et al., 2016). However, more research is needed to elucidate the potential effects of AMF symbiosis on the ability of grapevines to cope with water deficit in interaction with other environmental factors as well as to identify the mycorrhizal inoculants most appropriate for a given variety, cultivar or accession cultivated under a real and specific environmental scenario. In addition, the long-term site history and the previous management practices employed must be considered before introducing the AMF inocula in order to obtain benefits and ensure future food security (Thirkell et al., 2017).
The asexual propagation of the grapevine varieties allows the appearance and accumulation of somatic mutations, which are the basis for the clonal selection, which leads to differences in vigor, berry and cluster weight, yield production, resistance to plagues and diseases or oenological potential (Fernandes et al., 2015). One of the adaptive agronomic strategies to use in modern viticulture under the on-going climate change conditions is the selection of the best adapted rootstock and clones. Thus, the clonal selection could be oriented toward late-ripening clones to avoid alterations caused by high temperatures on fruit quality (van Leeuwen and Darriet, 2016), or to obtain clones with better balance between yield, acidity and alcoholic degree (Gonçalves et al., 2016). Recent findings provide evidence for this clonal diversity, which resulted in different abilities to respond to AMF inoculation (Torres et al., 2016). Therefore, the use of AMF for improving the fruit quality of grapevines needs to be included in an integrated management program of clonal selection.
Author Contributions
NT carried out the bibliographic search and wrote the first draft of the manuscript. MCA and NG analyzed the related papers and supervised the manuscript. All authors read and approved the submitted version.
Funding
NT was the recipient of a FPU grant of the Spanish Ministry of Education, Culture and Sport.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The reviewer EB and handling Editor declared their shared affiliation.
References
Alonso, R., Berli, F. J., Fontana, A., Piccoli, P., and Bottini, R. (2016a). Malbec grape (Vitis vinifera L.) responses to the environment: Berry phenolics as influenced by solar UV-B, water deficit and sprayed abscisic acid. Plant Physiol. Biochem. 109, 84–90. doi: 10.1016/j.plaphy.2016.09.007
Alonso, R., Berli, F. J., Piccoli, P., and Bottini, R. (2016b). Ultraviolet-B radiation, water deficit and abscisic acid: a review of independent and interactive effects on grapevines. Theor. Exp. Plant Physiol. 28, 11–22. doi: 10.1007/s40626-016-0053-y
Ambrosini, V. G., Voges, J. G., Canton, L., da Rosa Couto, R., Ferreira, P. A. A., Comin, J. J., et al. (2015). Effect of arbuscular mycorrhizal fungi on young vines in copper-contaminated soil. Braz. J. Microbiol. 46, 1045–1052. doi: 10.1590/S1517-838246420140622
Andrade, C., Leite, S. M., and Santos, J. A. (2012). Temperature extremes in Europe: overview of their driving atmospheric patterns. Nat. Hazards Earth Syst. Sci. 12, 1671–1691. doi: 10.5194/nhess-12-1671-2012
Artero, A., Artero, A., Tarín, J. J., and Cano, A. (2015). The impact of moderate wine consumption on health. Maturitas 80, 3–13. doi: 10.1016/j.maturitas.2014.09.007
Asha, H. N., Nirmalnath, P. J., Sagarkar, M. A., and Venkatesh, H. (2017). Impact of elevated CO2 and/or temperature on the am fungal diversity in groundnut rhizosphere under open top chamber facility. Int. J. Curr. Microbiol. Appl. Sci. 6, 882–895. doi: 10.20546/ijcmas.2017.601.104
Augé, R. M., Toler, H. D., and Saxton, A. M. (2015). Arbuscular mycorrhizal symbiosis alters stomatal conductance of host plants more under drought than under amply watered conditions: a meta-analysis. Mycorrhiza 25, 13–24. doi: 10.1007/s00572-014-0585-4
Ayenew, B., Degu, A., Manela, N., Perl, A., Shamir, M. O., and Fait, A. (2015). Metabolite profiling and transcript analysis reveal specificities in the response of a berry derived cell culture to abiotic stresses. Front. Plant Sci. 6:728. doi: 10.3389/fpls.2015.00728
Azuma, A., Yakushiji, H., Koshita, Y., and Kobayashi, S. (2012). Flavonoid biosynthesis-related genes in grape skin are differentially regulated by temperature and light conditions. Planta 236, 1067–1080. doi: 10.1007/s00425-012-1650-x
Balestrini, R., Magurno, F., Walker, C., Lumini, E., and Bianciotto, V. (2010). Cohorts of arbuscular mycorrhizal fungi (AMF) in Vitis vinifera, a typical Mediterranean fruit crop. Environ. Microbiol. Rep. 2, 594–604. doi: 10.1111/j.1758-2229.2010.00160.x
Balestrini, R., Salvioli, A., Dal Molin, A., Novero, M., Gabelli, G., Paparelli, E., et al. (2017). Impact of an arbuscular mycorrhizal fungus versus a mixed microbial inoculum on the transcriptome reprogramming of grapevine roots. Mycorrhiza 27, 417–430. doi: 10.1007/s00572-016-0754-8
Balint, G., and Reynolds, A. G. (2013a). Impact of irrigation strategies on abscisic acid and its catabolites profiles in leaves and berries of Baco noir grapes. J. Plant Growth Regul. 32, 884–900. doi: 10.1007/s00344-013-9354-4
Balint, G., and Reynolds, A. G. (2013b). Impact of exogenous abscisic acid on vine physiology and grape composition of Cabernet Sauvignon. Am. J. Enol. Vitic. 64, 74–87. doi: 10.5344/ajev.2012.12075
Balint, G., and Reynolds, A. G. (2016). Abscisic acid and its catabolites in leaves and berries of Chardonnay are affected by water status. J. Plant Growth Regul. 35, 1126–1140. doi: 10.1007/s00344-016-9610-5
Bandurska, H., Niedziela, J., and Chadzinikolau, T. (2013). Separate and combined responses to water deficit and UV-B radiation. Plant Sci. 213, 98–105. doi: 10.1016/j.plantsci.2013.09.003
Barnuud, N. N., Zerihun, A., Mpelasoka, F., Gibberd, M., and Bates, B. (2014). Responses of grape berry anthocyanin and titratable acidity to the projected climate change across the Western Australian wine regions. Int. J. Biometeorol. 58, 1279–1293. doi: 10.1007/s00484-013-0724-1
Baslam, M., Antolín, M. C., Gogorcena, Y., Muñoz, F., and Goicoechea, N. (2014). Changes in alfalfa forage quality and stem carbohydrates induced by arbuscular mycorrhizal fungi and elevated atmospheric CO2. Ann. Appl. Biol. 164, 190–199. doi: 10.1111/aab.12092
Baslam, M., Esteban, R., García-Plazaola, J. I., and Goicoechea, N. (2013a). Effectiveness of arbuscular mycorrhizal fungi (AMF) for inducing the accumulation of major carotenoids, chlorophylls and tocopherol in green and red leaf lettuces. Appl. Microbiol. Biotechnol. 97, 3119–3128. doi: 10.1007/s00253-012-4526-x
Baslam, M., Garmendia, I., and Goicoechea, N. (2013b). The arbuscular mycorrhizal symbiosis can overcome reductions in yield and nutritional quality in greenhouse-lettuces cultivated at inappropriate growing seasons. Sci. Hortic. 164, 145–154. doi: 10.1016/j.scienta.2013.09.021
Baslam, M., Garmendia, I., and Goicoechea, N. (2011). Arbuscular mycorrhizal fungi (AMF) improved growth and nutritional quality of greenhouse grown lettuce. J. Agric. Food Chem. 59, 5504–5515. doi: 10.1021/jf200501c
Baslam, M., Garmendia, I., and Goicoechea, N. (2012). Elevated CO2 may impair the beneficial effect of arbuscular mycorrhizal fungi on the mineral and phytochemical quality of lettuce. Ann. Appl. Biol. 161, 180–191. doi: 10.1111/j.1744-7348.2012.00563.x
Baslam, M., and Goicoechea, N. (2012). Water deficit improved the capacity of arbuscular mycorrhizal fungi (AMF) for inducing the accumulation of antioxidant compounds in lettuce leaves. Mycorrhiza 22, 347–359. doi: 10.1007/s00572-011-0408-9
Berdeja, M., Hilbert, G., Dai, Z. W., Lafontaine, M., Stoll, M., Schultz, H. R., et al. (2014). Effect of water stress and rootstock genotype on Pinot Noir berry composition. Aust. J. Grape Wine Res. 20, 409–421. doi: 10.1111/ajgw.12091
Berdeja, M., Nicolas, P., Kappel, C., Dai, Z. W., Hilbert, G., Peccoux, A., et al. (2015). Water limitation and rootstock genotype interact to alter grape berry metabolism through transcriptome reprogramming. Hortic. Res. 2:15012. doi: 10.1038/hortres.2015.12
Berli, F. J., Fanzone, M., Piccoli, P., and Bottini, R. (2011). Solar UV-B and ABA are involved in phenol metabolism of Vitis vinifera L. increasing biosynthesis of berry skin polyphenols. J. Agric. Food Chem. 59, 4874–4884. doi: 10.1021/jf200040z
Berruti, A., Lumini, E., Balestrini, R., and Bianciotto, V. (2016). Arbuscular mycorrhizal fungi as natural biofertilizers: Let’s benefit from past successes. Front. Microbiol. 6:1559. doi: 10.3389/fmicb.2015.01559
Bettoni, M. M., Mogor, A. F., Pauletti, V., and Goicoechea, N. (2017). The interaction between mycorrhizal inoculation, humic acids supply and elevated athmospheric CO2 increses energetic and antioxidant properties and sweetness of yellow onion. Hortic. Environ. Biotechnol. 58, 432–440. doi: 10.1007/s13580-017-0122-4
Bettoni, M. M., Mogor,Á. F., Pauletti, V., and Goicoechea, N. (2014). Growth and metabolism of onion seedlings as affected by the application of humic substances, mycorrhizal inoculation and elevated CO2. Sci. Hortic. 180, 227–235. doi: 10.1016/j.scienta.2014.10.037
Bona, E., Cantamessa, S., Massa, N., Manassero, P., Marsano, F., Copetta, A., et al. (2017). Arbuscular mycorrhizal fungi and plant growth-promoting pseudomonads improve yield, quality and nutritional value of tomato: a field study. Mycorrhiza 27, 1–11. doi: 10.1007/s00572-016-0727-y
Bonada, M., Jeffery, D. W., Petrie, P. R., Moran, M. A., and Sadras, V. O. (2015). Impact of elevated temperature and water deficit on the chemical and sensory profiles of Barossa Shiraz grapes and wines. Aust. J. Grape Wine Res. 21, 240–253. doi: 10.1111/ajgw.12142
Böttcher, C., and Davies, C. (2012). “Hormonal control of grape berry development and ripening,” in The Biochemistry of the Grape Berry, eds H. Gerós, M. M. Chaves, and S. Delrot (Potomac, MD: Bentham Science Publishers), 194–217. doi: 10.2174/978160805360511201010194
Bouffaud, M.-L., Bernaud, E., Colombet, A., van Tuinen, D., Wipf, D., and Redecker, D. (2016). Regional-scale analysis of arbuscular mycorrhizal fungi: the case of Burgundy vineyards. J. Int. Sci. Vigne Vin. 50, 1–8. doi: 10.20870/oeno-one.2016.50.1.49
Bruisson, S., Maillot, P., Schellenbaum, P., Walter, B., Gindro, K., and Deglène-Benbrahim, L. (2016). Arbuscular mycorrhizal symbiosis stimulates key genes of the phenylpropanoid biosynthesis and stilbenoid production in grapevine leaves in response to downy mildew and grey mould infection. Phytochemistry 131, 92–99. doi: 10.1016/j.phytochem.2016.09.002
Cáceres-Mella, A., Talaverano, M. I., Villalobos-González, L., Ribalta-Pizarro, C., and Pastenes, C. (2017). Controlled water deficit during ripening affects proanthocyanidin synthesis, concentration and composition in Cabernet Sauvignon grape skins. Plant Physiol. Biochem. 117, 34–41. doi: 10.1016/j.plaphy.2017.05.015
Caffarra, A., Rinaldi, M., Eccel, E., Rossi, V., and Pertot, I. (2012). Modelling the impact of climate change on the interaction between grapevine and its pests and pathogens: European grapevine moth and powdery mildew. Agric. Ecosyst. Environ. 148, 89–101. doi: 10.1016/j.agee.2011.11.017
Camprubí, A., Estaún, V., Nogales, A., García-Figueres, F., Pitet, M., and Calvet, C. (2008). Response of the grapevine rootstock Richter 110 to inoculation with native and selected arbuscular mycorrhizal fungi and growth performance in a replant vineyard. Mycorrhiza 18, 211–216. doi: 10.1007/s00572-008-0168-3
Cangahuala-Inocente, G. C., da Silva, M. F., Johnson, J. M., Manga, A., van Tuinen, D., Henry, C., et al. (2011). Arbuscular mycorrhizal symbiosis elicits proteome responses opposite of P-starvation in SO4 grapevine rootstock upon root colonisation with two Glomus species. Mycorrhiza 21, 473–493. doi: 10.1007/s00572-010-0352-0
Carbonell-Bejerano, P., Santa María, E., Torres-Pérez, R., Royo, C., Lijavetzky, D., Bravo, G., et al. (2013). Thermotolerance responses in ripening berries of Vitis vinifera L. cv. Muscat Hamburg. Plant Cell Physiol. 54, 1200–1216. doi: 10.1093/pcp/pct071
Carvalho, L. C., Coito, J. L., Gonçalves, E. F., Chaves, M. M., and Amâncio, S. (2016). Differential physiological response of the grapevine varieties Touriga Nacional and Trincadeira to combined heat, drought and light stresses. Plant Biol. 18, 101–111. doi: 10.1111/plb.12410
Castellanos-Morales, V., Villegas, J., Wendelin, S., Vierheiling, H., Eder, R., and Cárdenas-Navarro, R. (2010). Root colonisation by the arbuscular mycorrhizal fungus Glomus intraradices alters the quality of strawberry fruits (Fragaria × ananassa Duch.) at different nitrogen levels. J. Sci. Food Agric. 90, 1774–1782. doi: 10.1002/jsfa.3998
Castellarin, S. D., Pfeiffer, A., Sivilotti, P., Degan, M., Peterlunger, E., and Di Gaspero, G. (2007). Transcriptional regulation of anthocyanin biosynthesis in ripening fruits of grapevine under seasonal water deficit. Plant Cell Environ. 30, 1381–1399. doi: 10.1111/j.1365-3040.2007.01716.x
Cholet, C., Claverol, S., Claisse, O., Rabot, A., Osowsky, A., Dumot, V., et al. (2016). Tartaric acid pathways in Vitis vinifera L. (cv. Ugni blanc): a comparative study of two vintages with contrasted climatic conditions. BMC Plant Biol. 16:144. doi: 10.1186/s12870-016-0833-1
Compant, S., van der Heijden, M. G. A., and Sessitsch, A. (2010). Climate change effects on beneficial plant-microorganism interactions. FEMS Microbiol. Ecol. 73, 197–214. doi: 10.1111/j.1574-6941.2010.00900.x
Conde, C., Silva, P., Fontes, N., Dias, A. C. P., Tavares, R. M., Sousa, M. J., et al. (2007). Biochemical changes throughout grape berry development and fruit and wine quality. Food 1, 1–22. doi: 10.1186/s12864-016-2660-z
Costa, J. M., Vaz, M., Escalona, J., Egipto, R., Lopes, C., Medrano, H., et al. (2016). Modern viticulture in southern Europe: vulnerabilities and strategies for adaptation to water scarcity. Agric. Water Manage. 164, 5–18. doi: 10.1016/j.agwat.2015.08.021
Cotton, T. E. A., Fitter, A. H., Miller, R. M., Dumbrell, A. J., and Helgason, T. (2015). Fungi in the future: Interannual variation and effects of atmospheric change on arbuscular mycorrhizal fungal communities. New Phytol. 205, 1598–1607. doi: 10.1111/nph.13224
De Rosas, I., Ponce, M. T., Malovini, E., Deis, L., Cavagnaro, B., and Cavagnaro, P. (2017). Loss of anthocyanins and modification of the anthocyanin profiles in grape berries of Malvec and Bonarda grown under high temperature conditions. Plant Sci. 258, 137–145. doi: 10.1016/j.plantsci.2017.01.015
Del-Castillo-Alonso, M. A., Castagna, A., Csepregi, K., Hideg, E., Jakab, G., Jansen, M. A. K., et al. (2016). Environmental factors correlated with the metabolite profile of Vitis vinifera cv. Pinot Noir berry skins along a European latitudinal gradient. J. Agric. Food Chem. 64, 8722–8734. doi: 10.1021/acs.jafc.6b03272
Deluc, L. G., Quilici, D. R., Decendit, A., Grimplet, J., Wheatley, D., Schlauch, A., et al. (2009). Water deficit alters differentially metabolic pathways affecting important flavour and quality traits in grape berries of Cabernet Sauvignon and Chardonnay. BMC Genomics 10:212. doi: 10.1186/1471-2164-10-212
dos Santos, E. L., Alves da Silva, F., and Barbosa da Silva, F. S. (2017). Arbuscular mycorrhizal fungi increase the phenolic compounds concentrations in the bark of the stem of Libidibia ferrea in field conditions. Open Microbiol. J. 11, 283–291. doi: 10.2174/1874285801711010283
Eftekhari, M., Alizadeh, M., and Ebrahimi, P. (2012a). Evaluation of the total phenolics and quercetin content of foliage in mycorrhizal grape (Vitis vinifera L.) varieties and effect of postharvest drying on quercetin yield. Ind. Crops Prod. 38, 160–165. doi: 10.1016/j.indcrop.2012.01.022
Eftekhari, M., Alizadeh, M., Mashayekhi, K., and Asghari, H. R. (2012b). In vitro propagation of four Iranian grape varieties: influence of genotype and pretreatment with arbuscular mycorrhiza. Vitis 51, 175–182.
Eurostat Statistics Division. (2016). Available at: http://ec.europa.eu/eurostat/statistics-explained/index.php/Agricultural_production_-_crops#Grapes [accessed August 21, 2017].
Fernandes, A. M., Melo-Pinto, P., Millan, B., Tardaguila, J., and Diago, M. P. (2015). Automatic discrimination of grapevine (Vitis vinifera L.) clones using leaf hyperspectral imaging and partial least squares. J. Agric. Sci. 153, 455–465. doi: 10.1017/S0021859614000252
Fernandes de Oliveira, A., Mercenaro, L., Del Caro, A., Pretti, L., and Nieddu, G. (2015). Distinctive anthocyanin accumulation responses to temperature and natural UV radiation of two field-grown Vitis vinifera L. cultivars. Molecules 20, 2061–2080. doi: 10.3390/molecules20022061
Fernandes de Oliveira, A., and Nieddu, G. (2016). Accumulation and partitioning of anthocyanins in two red grape cultivars under natural and reduced UV solar radiation. Aust. J. Grape Wine Res. 22, 96–104. doi: 10.1111/ajgw.12174
Fraga, H., Malheiro, A. C., Moutinho-Pereira, J., and Santos, J. A. (2013). Future scenarios for viticultural zoning in Europe: ensemble projections and uncertainties. Int. J. Biometeorol. 57, 909–925. doi: 10.1007/s00484-012-0617-8
Fraga, H., Santos, J. A., Moutinho-Pereira, J., Carlos, C., Silvestre, J., Eiras-Dias, J., et al. (2016). Climatic suitability of Portuguese grapevine varieties and climate change adaptation. J. Agric. Sci. 154, 795–811. doi: 10.1017/S0021859615000933
Friedel, M., Frotscher, J., Nitsch, M., Hofmann, M., Bogs, J., Stoll, M., et al. (2016). Light promotes expression of monoterpene and flavonol metabolic genes and enhances flavour of winegrape berries (Vitis vinifera L. cv. Riesling). Aust. J. Grape Wine Res. 22, 409–421. doi: 10.1111/ajgw.12229
García-Díaz, A., Marqués, M. J., Sastre, B., and Bienes, R. (2018). Labile and stable soil organic carbón and physical improvements using groundcovers in vineyards from central Spain. Sci. Total Environ. 621, 387–397. doi: 10.1016/j.scitotenv.2017.11.240
Georgiev, V., Ananga, A., and Tsolova, V. (2014). Recent advances and uses of grape flavonoids as nutraceuticals. Nutrients 6, 391–415. doi: 10.3390/nu6010391
Goicoechea, N., and Antolín, M. C. (2017). Increased nutritional value in food crops. Microb. Biotechnol. 10, 1004–1007. doi: 10.1111/1751-7915.12764
Goicoechea, N., Bettoni, M. M., Fuertes-Mendizábal, T., González-Murua, C., and Aranjuelo, I. (2016). Durum wheat quality traits affected by mycorrhizal inoculation, water availability and atmospheric CO2 concentration. Crop Past Sci. 67, 147–155. doi: 10.1071/CP15212
Gonçalves, E., Carrasquinho, I., Almeida, R., Pedroso, V., and Martins, A. (2016). Genetic correlations in grapevine and their effects on selection. Aust. J. Grape Wine Res. 22, 52–63. doi: 10.1111/ajgw.12164
Guadarrama, P., Castillo, S., Ramos-Zapata, J. A., Hernández-Cuevas, L. V., and Camargo-Ricalde, S. L. (2014). Arbuscular mycorrhizal fungal communities in changing environments: the effects of seasonality and anthropogenic disturbance in a seasonal dry forest. Pedobiologia 57, 87–95. doi: 10.1016/j.pedobi.2014.01.002
Guan, L., Dai, Z., Wu, B. H., Wu, J., Merlin, I., Hilbert, G., et al. (2016). Anthocyanin biosynthesis is differentially regulated by light in the skin and flesh of white-fleshed and teinturier grape berries. Planta 243, 23–41. doi: 10.1007/s00425-015-2391-4
Hamilton, C. E., Bever, J. D., Labbé, J., Yang, X., and Yin, H. (2016). Mitigating climate change through managing constructed-microbial communities in agriculture. Agric. Ecosyst. Environ. 216, 304–308. doi: 10.1016/j.agee.2015.10.006
Han, B., Guo, S. R., He, C. X., Yan, Y., and Yu, X. C. (2012). Effects of arbuscular mycorrhiza fungi (AMF) on the plant growth, fruit yield, and fruit quality of cucumber under salt stress. J. Appl. Ecol. 23, 154–158.
Hannah, L., Roehrdanz, P. R., Ikegami, M., Shepard, A. V., Shaw, M. R., Tabor, G., et al. (2013). Climate change, wine, and conservation. Proc. Natl. Acad. Sci. U.S.A. 110, 6907–6912. doi: 10.1073/pnas.1210127110
Hao, Z., Fayolle, L., van Tuinen, D., Chatagnier, O., Li, X., Gianinazzi, S., et al. (2012). Local and systemic mycorrhiza-induced protection against the ectoparasitic nematode Xiphinema index involves priming of defence gene responses in grapevine. J. Exp. Bot. 63, 3657–3672. doi: 10.1093/jxb/ers046
Hart, M., Ehret, D. L., Krumbein, A., Leung, C., Murch, S., Turi, C., et al. (2015). Inoculation with arbuscular mycorrhizal fungi improves the nutritional value of tomatoes. Mycorrhiza 25, 359–376. doi: 10.1007/s00572-014-0617-0
Hatmi, S., Gruau, C., Trotel-Aziz, P., Villaume, S., Rabenoelina, F., Baillieul, F., et al. (2015). Drought stress tolerance in grapevine involves activation of polyamine oxidation contributing to improved immune response and low susceptibility to Botrytis cinerea. J. Exp. Bot. 66, 775–787. doi: 10.1093/jxb/eru436
Hawkes, C. V., Hartley, I. P., Ineson, P., and Fitter, A. H. (2008). Soil temperature affects carbon allocation within arbuscular mycorrhizal networks and carbon transport from plant to fungus. Glob. Change Biol. 14, 1181–1190. doi: 10.1111/j.1365-2486.2007.01535.x
Hochberg, U., Batushansky, A., Degu, A., Rachmilevitch, S., and Fait, A. (2015). Metabolic and physiological responses of Shiraz and Cabernet Sauvignon (Vitis vinifera L.) to near optimal temperatures of 25 and 35°C. Int. J. Mol. Sci. 16, 24276–24294. doi: 10.3390/ijms161024276
Hristozkova, M., Gigova, L., Geneva, M., Stancheva, I., Vasileva, I., Sichanova, M., et al. (2017). Mycorrhizal fungi and microalgae modulate antioxidant capacity of basil plants. J. Plant Prot. Res. doi: 10.1515/jppr-2017-0057 [Epub ahead of print].
Huang, J.-C., Lai, W.-A., Singh, S., Hameed, A., and Young, C.-C. (2013). Response of mycorrhizal hybrid tomato cultivars under saline stress. J. Soil Sci. Plant Nutr. 13, 469–484. doi: 10.4067/S0718-95162013005000037
Intergovernmental Panel on Climate Change, [IPCC] (2014). in Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change, eds Core Writing Team, R. K. Pachauri, and L. A. Meyer (Geneva: IPCC). doi: 10.1017/CBO9781107415416
Johnson, N. C., Angelard, C., Sanders, I. R., and Kiers, E. T. (2013). Predicting community and ecosystem outcomes of mycorrhizal responses to global change. Ecol. Lett. 16, 140–153. doi: 10.1111/ele.12085
Joubert, C., Young, P. R., Eyeghe-Bickong, H. A., and Vivier, M. A. (2016). Field-grown grapevine berries use carotenoids and the associated xanthophyll cycles to acclimate to UV exposure differentially in high and low light (shade) conditions. Front. Plant Sci. 7:786. doi: 10.3389/FPLS.2016.00786
Karagiannidis, N., Nikolaou, N., Ipsilantis, I., and Zioziou, E. (2007). Effects of different N fertilizers on the activity of Glomus mosseae and on grapevine nutrition and berry composition. Mycorrhiza 18, 43–50. doi: 10.1007/s00572-007-0153-2
Kizildeniz, T., Mekni, I., Santesteban, H., Pascual, I., Morales, F., and Irigoyen, J. J. (2015). Effects of climate change including elevated CO2 concentration, temperature and water deficit on growth, water status, and yield quality of grapevine (Vitis vinifera L.) cultivars. Agric. Water. Manage. 159, 155–164. doi: 10.1016/j.agwat.2015.06.015
Leibar, U., Pascual, I., Morales, F., Aizpurua, A., and Unamunzaga, O. (2017). Grape yield and quality responses to simulated year 2100 expected climatic conditions under different soil textures. J. Sci. Food Agric. 97, 2633–2640. doi: 10.1002/jsfa.8086
Martínez-Lüscher, J., Morales, F., Delrot, S., Sánchez-Díaz, M., Gomès, E., Aguirreolea, J., et al. (2015a). Characterization of the adaptive response of grapevine (cv. Tempranillo) to UV-B radiation under water deficit conditions. Plant Sci. 232, 13–22. doi: 10.1016/j.plantsci.2014.12.013
Martínez-Lüscher, J., Morales, F., Sánchez-Díaz, M., Delrot, S., Aguirreolea, J., Gomès, E., et al. (2015b). Climate change conditions (elevated CO2 and temperature) and UV-B radiation affect grapevine (Vitis vinifera cv. Tempranillo) leaf carbon assimilation, altering fruit ripening rates. Plant Sci. 236, 168–176. doi: 10.1016/j.plantsci.2015.04.001
Martínez-Lüscher, J., Torres, N., Hilbert, G., Richard, T., Sánchez-Díaz, M., Delrot, S., et al. (2014). Ultraviolet-B radiation modifies the quantitative and qualitative profile of flavonoids and amino acids in grape berries. Phytochemistry 102, 106–114. doi: 10.1016/j.phytochem.2014.03.014
Mena-Violante, H. G., Ocampo-Jiménez, O., Dendooven, L., Martínez-Soto, G., González-Castañeda, J., Davies, F. T. Jr., et al. (2006). Arbuscular mycorrhizal fungi enhance fruit growth and quality of chile ancho (Capsicum annuum L. cv. San Luis) plants exposed to drought. Mycorrhiza 16, 261–267. doi: 10.1007/s00572-006-0043-z
Mohan, J. E., Cowden, C. C., Baas, P., Dawadi, A., Frankson, P. T., Helmick, K., et al. (2014). Mycorrhizal fungi mediation of terrestrial ecosystem responses to global change: Mini-review. Fungal Ecol. 10, 3–19. doi: 10.1016/j.funeco.2014.01.005
Mosedale, J. R., Abernethy, K. E., Smart, R. E., Wilson, R. J., and Maclean, I. M. D. (2016). Climate change impacts and adaptive strategies: lessons from the grapevine. Glob. Change Biol. 22, 3814–3828. doi: 10.1111/gcb.13406
Movahed, N., Pastore, C., Cellini, A., Allegro, G., Valentini, G., Zenoni, S., et al. (2016). The grapevine VviPrx31 peroxidase as a candidate gene involved in anthocyanin degradation in ripening berries under high temperature. J. Plant Res. 129, 513–526. doi: 10.1007/s10265-016-0786-3
Neethling, E., Petitjean, T., Quénol, H., and Barbeau, G. (2017). Assessing local climate vulnerability and winegrowers’ adaptive processes in the context of climate change. Mitig. Adapt. Strat. Global Change 22, 777–803. doi: 10.1007/s11027-015-9698-0
Nicolás, E., Maestre-Valero, J. F., Alarcón, J. J., Pedrero, F., Vicente-Sánchez, J., Bernabé, A., et al. (2015). Effectiveness and persistence of arbuscular mycorrhizal fungi on the physiology, nutrient uptake and yield of Crimson seedless grapevine. J. Agric. Sci. 153, 1084–1096. doi: 10.1017/S002185961400080X
Niculcea, M., López, J., Sánchez-Díaz, M., and Antolín, M. C. (2014). Involvement of berry hormonal content in the response to pre- and post-veraison water deficit in different grapevine (Vitis vinifera L.) cultivars. Aust. J. Grape Wine Res. 20, 281–291. doi: 10.1111/ajgw.12064
Niculcea, M., Martínez-Lapuente, L., Guadalupe, Z., Sánchez-Díaz, M., Ayestarán, B., and Antolín, M. C. (2015). Characterization of phenolic composition of Vitis vinifera L. “Tempranillo” and “Graciano” subjected to deficit irrigation during berry development. Vitis 54, 9–16.
Niculcea, M., Martínez-Lapuente, L., Guadalupe, Z., Sánchez-Díaz, M., Morales, F., Ayestarán, B., et al. (2013). Effects on water deficit irrigation on hormonal content and nitrogen compounds in developing berries of Vitis vinifera L. cv. Tempranillo. J. Plant Growth Regul. 32, 551–563. doi: 10.1007/s00344-013-9322-z
Nogales, A., Aguirreolea, J., Santa María, E., Camprubí, A., and Calvet, C. (2009a). In vitro interaction studies between Glomus intraradices and Armillaria mellea in vines. Span. J. Agric. Res. 8, 62–68. doi: 10.5424/sjar/201008S1-1223
Nogales, A., Luque, J., Estaún, V., Camprubí, A., Garcia-Figueres, F., and Calvet, C. (2009b). Differential growth of mycorrhizal field-inoculated grapevine rootstocks in two replant soils. Am. J. Enol. Vitic. 60, 484–489.
Ocete, R., Armendariz, I., Cantos, M., Álvarez, D., and Azcón, R. (2015). Ecological characterization of wild grapevine habitats focused on arbuscular mycorrhizal simbiosis. Vitis 54, 207–211.
Oehl, F., and Koch, B. (2018). Diversity of arbuscular mycorrhizal fungi in no-till and conventionally tilled vineyards. JABFQ 91, 56–60. doi: 10.5073/JABFQ.2018.091.008
Ollé, D., Guiraud, J. L., Souquet, J. M., Terrier, N., Ageorge, S. A., Cheynier, V., et al. (2011). Effect of pre- and post-veraison water deficit on proanthocyanidins and anthocyanin accumulation during Shiraz berry development. Aust. J. Grape Wine Res. 17, 90–100. doi: 10.1111/j.1755-0238.2010.00121.x
Palliotti, A., Tombesi, S., Silvestroni, O., Lanari, V., Gatti, M., and Poni, S. (2014). Changes in vineyard establishment and canopy management urged by earlier climate-related grape ripening: a review. Sci. Hortic. 178, 43–54. doi: 10.1016/j.scienta.2014.07.039
Paterson, E., Sim, A., Davidson, J., and Daniell, T. J. (2016). Arbuscular mycorrhizal hyphae promote priming of native soil organic matter mineralization. Plant Soil 408, 243–254. doi: 10.1107/s11104-016-2928-8
Pritchard, S. G. (2011). Soil organisms and global climate change. Plant Pathol. 60, 82–99. doi: 10.1111/j.1365-3059.2010.02405.x
Resco, P., Iglesias, A., Bardají, I., and Sotés, V. (2016). Exploring adaptation choices for grapevine regions in Spain. Reg. Environ. Change 16, 979–993. doi: 10.1007/s10113-015-0811-4
Reshef, N., Walbaum, N., Agam, N., and Fait, A. (2017). Sunlight modulates fruit metabolic profile and shapes the spatial pattern of compound accumulation within the grape cluster. Front. Plant Sci. 8:70. doi: 10.3389/fpls.2017.00070
Rienth, M., Torregrosa, L., Luchaire, N., Chatbanyong, R., Lecourieux, D., Kelly, M. T., et al. (2014). Day and night heat stress trigger different transcriptomic responses in green and ripening grapevine (Vitis vinifera) fruit. BMC Plant Biol. 14:108. doi: 10.1186/1471-2229-14-108
Romero, P., Muñoz, R. G., Fernández-Fernández, J. I., del Amor, F. M., Martínez-Cutillas, A., and García-García, J. (2015). Improvement of yield and grape and wine composition in field-grown Monastrell grapevines by partial root zone irrigation, in comparison with regulated deficit irrigation. Agric. Water Manage. 149, 55–73. doi: 10.1016/j.agwat.2014.10.018
Roy, P., Grenier, P., Barriault, E., Logan, T., Blondlot, A., Bourgeois, G., et al. (2017). Probabilistic climate change scenarios for viticultural potential in Québec. Clim. Change 143, 43–58. doi: 10.1007/s10584-017-1960-x
Sadras, V. O., and Moran, M. A. (2012). Elevated temperature decouples anthocyanins and sugars in berries of Shiraz and Cabernet Franc. Aust. J. Grape Wine Res. 18, 115–122. doi: 10.1111/j.1755-0238.2012.00180.x
Salazar-Parra, C., Aguirreolea, J., Sánchez-Díaz, M., Irigoyen, J. J., and Morales, F. (2010). Effects of climate change scenarios on Tempranillo grapevine (Vitis vinifera L.) ripening: Response to a combination of elevated CO2 and temperature, and moderate drought. Plant Soil 337, 179–191. doi: 10.1007/s11104-010-0514-z
Savoi, S., Wong, D. C. J., Arapitsas, P., Miculan, M., Bucchetti, B., Peterlunger, E., et al. (2016). Transcriptome and metabolite profiling reveals that prolonged drought modulates the phenylpropanoid and terpenoid pathway in white grapes (Vitis vinifera L.). BMC Plant Biol. 16:67. doi: 10.1186/s12870-016-0760-1
Savoi, S., Wong, D. C. J., Degu, A., Herrera, J. C., Bucchetti, B., Peterlunger, E., et al. (2017). Multi-omics and integrated network analyses reveal new insights into the systems relationships between metabolites, structural genes, and transcriptional regulators in developing grape berries (Vitis vinifera L.) exposed to water deficit. Front. Plant Sci. 8:1124. doi: 10.3389/fpls.2017.01124
Schreiner, R. P. (2007). Effects of native and nonnative arbuscular mycorrhizal fungi on growth and nutrient uptake of ‘Pinot noir’(Vitis vinifera L.) in two soils with contrasting levels of phosphorus. Appl. Soil Ecol. 36, 205–215. doi: 10.1007/s00572-007-0128-3
Schreiner, R. P., and Mihara, K. L. (2009). The diversity of arbuscular mycorrhizal fungi amplified from grapevine roots (Vitis vinifera L.) in Oregon vineyards is seasonally stable and influenced by soil and vine age. Mycologia 101, 599–611. doi: 10.3852/08-169
Schreiner, R. P., Tarara, J. M., and Smithyman, R. P. (2007). Deficit irrigation promotes arbuscular colonization of fine roots by mycorrhizal fungi in grapevines (Vitis vinifera L.) in an arid climate. Mycorrhiza 17, 551–562. doi: 10.1007/s00572-007-0128-3
Schultz, H. R. (2016). Global climate change, sustainability, and some challenges for grape and wine production. J. Wine Econom. 11, 181–200. doi: 10.1017/jwe.2015.31
Shinomiya, R., Fujishima, H., Muramoto, K., and Shiraishi, M. (2015). Impact of temperature and sunlight on the skin coloration of the “Kyoho” table grape. Sci. Hortic. 193, 77–83. doi: 10.1016/j.scienta.2015.06.042
Sinclair, G., Charest, C., Dalpé, Y., and Khanizadeh, S. (2014). Influence of colonization by arbuscular mycorrhizal fungi on three strawberry cultivars under salty conditions. Agric. Food Sci. 23, 146–158.
Smith, S. E., and Read, D. (2008). “The symbionts forming arbuscular mycorrhizas,” in Mycorrhizal Symbiosis, 3rd Edn, eds S. E. Smith and D. Read (London: Academic Press), 13–41. doi: 10.1016/B978-012370526-6.50003-9
Sofo, A., Nuzzo, V., Tataranni, G., Manfra, M., De Nisco, M., and Scopa, A. (2012). Berry morphology and composition in irrigated and non-irrigated grapevine (Vitis vinifera L.). J. Plant Physiol. 169, 1023–1031. doi: 10.1016/j.jplph.2012.03.007
Sweetman, C., Sadras, V. O., Hancock, R. D., Soole, K. L., and Ford, C. M. (2014). Metabolic effects of elevated temperature on organic acid degradation in ripening Vitis vinifera fruit. J. Exp. Bot. 65, 5975–5988. doi: 10.1093/jxb/eru343
Thirkell, T. J., Charters, M. D., Elliott, A. J., Sait, S. M., and Field, K. J. (2017). Are mycorrhizal fungi our sustainable saviours? Considerations for achieving food security. J. Ecol. 105, 921–929. doi: 10.1111/1365-2745.12788
Torres, N., Goicoechea, N., and Antolín, M. C. (2015). Antioxidant properties of leaves from different accessions of grapevine (Vitis vinifera L.) cv. Tempranillo after applying biotic and/or environmental modulator factors. Ind. Crops Prod. 76, 77–85. doi: 10.1016/j.indcrop.2015.03.093
Torres, N., Goicoechea, N., and Antolín, M. C. (2018). Influence of irrigation strategy and mycorrhizal inoculation on fruit quality in different clones of Tempranillo grown under elevated temperatures. Agric. Water Manag. 202, 285–298. doi: 10.1016/j.agwat.2017.12.004
Torres, N., Goicoechea, N., Morales, F., and Antolín, M. C. (2016). Berry quality and antioxidant properties in Vitis vinifera cv. Tempranillo as affected by clonal variability, mycorrhizal inoculation and temperature. Crop Past. Sci. 67, 961–977. doi: 10.1071/CP16038
Torres, N., Hilbert, G., Luquin, J., Goicoechea, N., and Antolín, M. C. (2017). Flavonoid and amino acid profiling on Vitis vinifera L. cv Tempranillo subjected to deficit irrigation under elevated temperatures. J. Food Comp. Anal. 62, 51–62. doi: 10.1016/j.jfca.2017.05.001
Treseder, K. K. (2016). Model behavior of arbuscular mycorrhizal fungi: predicting soil carbon dynamics under climate change. Botany 94, 417–423. doi: 10.1139/cjb-2015-0245
Trigo-Córdoba, E., Bouzas-Cid, Y., Orriols-Fernández, I., Díaz-Losada, E., and Mirás-Avalos, J. (2015). Influence of cover crop treatments on the performance of a vineyard in a humid región. Span. J. Agric. Res. 13:e0907. doi: 10.5424/sjar/2015134-8265
Trouvelot, S., Bonneau, L., Redecker, D., van Tuinen, D., Adrian, M., and Wipf, D. (2015). Arbuscular mycorrhiza symbiosis in viticulture: a review. Agron. Sustain. Dev. 35, 1449–1467. doi: 10.1007/s13593-015-0329-7
van Leeuwen, C., and Darriet, P. (2016). The impact of climate change on viticulture and wine quality. J. Wine Econom. 11, 150–167. doi: 10.1017/jwe.2015.21
Veresoglou, S. D., Anderson, I. C., de Sousa, N. M. F., Hempel, S., and Rillig, M. C. (2016). Resilience of fungal communities to elevated CO2. Microb. Ecol. 72, 493–495. doi: 10.1007/s00248-016-0795-8
Vilvert, E., Dalla Costa, M., Cangahuala-Inocente, G. C., and Lovato, P. E. (2017). Root proteomic analysis of grapevine rootstocks inoculated with Rhizophagus irregularis and Fusarium oxysporum f. sp. herbemontis. Rev. Bras. Cienc. Solo 41, 1–14. doi: 10.1590/18069657rbcs20160134
Vimal, S. R., Singh, J. S., Arora, N. K., and Singh, S. (2017). Soil-Plant-Microbe interactions in stressed agriculture management: a review. Pedosphere 27, 177–192. doi: 10.1016/S1002-0160(17)60309-6
Vislocky, L. M., and Fernandez, M. L. (2010). Biomedical effects of grape products. Nutr. Rev. 68, 656–670. doi: 10.1111/j.1753-4887.2010.00335.x
Vukicevich, E., Lowery, D. T., Úrbez-Torres, J. R., Bowen, P., and Hart, M. (2018). Groundcover management changes grapevine root fungal communities and plant-soil feedback. Plant Soil 424, 419–433. doi: 10.1007/s11104-017-3532-2
Wang, W.-X., Zhang, F., Chen, Z.-L., Liu, J., Gou, C., He, J.-D., et al. (2017). Responses of phytohormones and gas exchange to mycorrhizal colonisation in trifoliate orange subjected to drought stress. Arch. Agron. Soil Sci. 63, 14–23. doi: 10.1080/03650340.2016.1175556
Webb, L. B., Watterson, I., Bhend, J., Whetton, P. H., and Barlow, E. W. R. (2013). Global climate analogues for winegrowing regions in future periods: projections of temperature and precipitation. Aust. J. Grape Wine Res. 19, 331–341. doi: 10.1111/ajgw.12045
Wiesmeier, M., Poeplau, C., Sierra, C. A., Maier, H., Frühauf, C., Hübner, R., et al. (2016). Projected loss of soil organic carbon in temperate agricultural soils in the 21st century: effects of climate change and carbon input trends. Sci. Rep. 6:32525. doi: 10.1038/srep32525
Wilson, H., Johnson, B. R., Bohannan, B., Pfeifer-Meister, L., Mueller, R., and Bridgham, S. D. (2016). Experimental warming decreases arbuscular mycorrhizal fungal colonization in prairie plants along a Mediterranean climate gradient. Peer J. 4:e2083. doi: 10.7717/peerj.2083
Zarrouk, O., Brunetti, C., Egipto, R., Pinheiro, C., Genebra, T., Gori, A., et al. (2016). Grape ripening is regulated by deficit irrigation/elevated temperatures according to cluster position in the canopy. Front. Plant Sci. 7:1640. doi: 10.3389/fpls.2016.01640
Zarrouk, O., Francisco, R., Pinto-Marijuan, M., Brossa, R., Santos, R. R., Pinheiro, C., et al. (2012). Impact of irrigation regime on berry development and flavonoids composition in Aragonez (Syn. Tempranillo) grapevine. Agric. Water Manage. 114, 18–29. doi: 10.1016/j.agwat.2012.06.018
Zeng, L., JianFu, L., JianFu, L., and MingYuan, W. (2014). Effects of arbuscular mycorrhizal (AM) fungi on citrus quality under nature conditions. Southwest China J. Agric. Sci. 27, 2101–2105.
Zhang, T., Yang, X., Guo, R., and Guo, J. (2016). Response of AM fungi spore population to elevated temperature and nitrogen addition and their influence on the plant community composition and productivity. Sci. Rep. 6:24749. doi: 10.1038/srep24749
Keywords: berry metabolism, climate change, fruit quality, mycorrhizal fungi, Vitis vinifera
Citation: Torres N, Antolín MC and Goicoechea N (2018) Arbuscular Mycorrhizal Symbiosis as a Promising Resource for Improving Berry Quality in Grapevines Under Changing Environments. Front. Plant Sci. 9:897. doi: 10.3389/fpls.2018.00897
Received: 07 February 2018; Accepted: 07 June 2018;
Published: 29 June 2018.
Edited by:
Guido Lingua, Università degli Studi del Piemonte Orientale, ItalyReviewed by:
Sergio Saia, Consiglio per la Ricerca in Agricoltura e l’Analisi dell’Economia Agraria (CREA), ItalyElisa Bona, Università degli Studi del Piemonte Orientale, Italy
Copyright © 2018 Torres, Antolín and Goicoechea. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Nieves Goicoechea, bmllZ29pQHVuYXYuZXM=