 Marie-Laure Pilet-Nayel1,2*
Marie-Laure Pilet-Nayel1,2* Benoît Moury3
Benoît Moury3 Valérie Caffier4
Valérie Caffier4 Josselin Montarry1
Josselin Montarry1 Marie-Claire Kerlan1
Marie-Claire Kerlan1 Sylvain Fournet1
Sylvain Fournet1 Charles-Eric Durel4
Charles-Eric Durel4 Régine Delourme1
Régine Delourme1- 1Institute for Genetics, Environment and Plant Protection (INRA), UMR 1349, Leu Rheu, France
- 2PISOM, UMT INRA-Terres Inovia, Le Rheu, France
- 3Pathologie Végétale (INRA), Montfavet, France
- 4Research Institute of Horticulture and Seeds (INRA), UMR 1345, Beaucouzé, France
Quantitative resistance has gained interest in plant breeding for pathogen control in low-input cropping systems. Although quantitative resistance frequently has only a partial effect and is difficult to select, it is considered more durable than major resistance (R) genes. With the exponential development of molecular markers over the past 20 years, resistance QTL have been more accurately detected and better integrated into breeding strategies for resistant varieties with increased potential for durability. This review summarizes current knowledge on the genetic inheritance, molecular basis, and durability of quantitative resistance. Based on this knowledge, we discuss how strategies that combine major R genes and QTL in crops can maintain the effectiveness of plant resistance to pathogens. Combining resistance QTL with complementary modes of action appears to be an interesting strategy for breeding effective and potentially durable resistance. Combining quantitative resistance with major R genes has proven to be a valuable approach for extending the effectiveness of major genes. In the plant genomics era, improved tools and methods are becoming available to better integrate quantitative resistance into breeding strategies. Nevertheless, optimal combinations of resistance loci will still have to be identified to preserve resistance effectiveness over time for durable crop protection.
Introduction
Plant pathogens are major limiting factors in crop production and this has led to the extensive use of chemicals to control them. Plant genetic resistance is a promising key alternative to control crop diseases and pests. However, pathogens frequently adapt to and overcome genetic resistance especially when it is determined by major genes. Thus, quantitative resistance has gained interest in recent years to address the major challenge of genetic resistance durability.
Quantitative resistance can refer to an incomplete or partial level of resistance phenotype. It can also refer to a continuous distribution between resistant and susceptible phenotypes in a progeny, most often resulting from the segregation of alleles with variable effects at several loci. In contrast, qualitative resistance refers to either a complete or high level of resistance, or to a bimodal segregation of phenotypes in a progeny, classifying individuals into two distinct categories: resistant and susceptible. Accordingly, Niks et al. (2015) distinguished qualitative or quantitative resistance, depending on whether it refers to its effect on the phenotype or mode of inheritance.
Quantitative Disease Resistance: Inheritance, Function, Durability
Several genes usually control quantitative resistance and are associated with genomic regions or QTL (quantitative trait loci) which contribute, each with variable effect, to the phenotype of resistance to a pathogen. Quantitative resistance often confers a partial level of resistance to the plant. It does not block but only reduces pathogen multiplication, plant colonization, and/or symptom severity. However, a combination of resistance QTL can lead to total resistance in some cases, especially when QTL have strong effects (Niks et al., 2015). For example, three QTL, rx1, rx2, and rx3 were found to confer a high level of resistance of tomato to Xanthomonas campestris (Stall et al., 2009).
Over the past 20 years, since the development of molecular markers, many resistance QTL detection experiments have been conducted in all major crop species (Kover and Caicedo, 2001; Wilfert and Schmid-Hempel, 2008). The genetic architecture of quantitative resistance has often been associated with a small number of detected QTL, some with major effects and others with minor effects. In recent years, the development of next generation sequencing (NGS) technologies, coupled with the use of accession panels and multi-parental populations, has accelerated the discovery of a higher number of QTL controlling quantitative resistance. Markers closely linked to QTL, especially to QTL with minor effects, and marker haplotypes describing allelic diversity at QTL were identified (Yu and Buckler, 2006; Huang and Han, 2014; Desgroux et al., 2016). Recently, Bartoli and Roux (2017) reviewed 35 genome-wide association (GWA) studies of plant resistance to pathogens. Closely linked markers to resistance QTL were reported in 34 plant–pathogen systems and candidate genes validated in Arabidopsis thaliana/Pseudomonas syringae, Ralstonia solanacearum, Xanthomonas campestris, and Oryza sativa/Magnaporthe oryzae systems.
The effects of resistance QTL are often additive. However, QTL with epistatic effects were identified (Lefebvre and Palloix, 1996; Manzanares-Dauleux et al., 2000; Calenge et al., 2005). Resistance QTL can sometimes only be detected under certain environmental conditions (soil, climate, pathogen population), or in specific genetic backgrounds or cross types. Thus, stable QTL are highly sought after for their applicability in breeding (Calenge and Durel, 2006; Ballini et al., 2008; Danan et al., 2011; Hamon et al., 2011; Goudemand et al., 2013). When several isolates are used for QTL analysis in a given mapping population, resistance QTL can be detected as either isolate-specific (i.e., detected with only one isolate) or non-specific (i.e., broad-spectrum). In several systems, quantitative resistance was found to result from the combination of broad-spectrum QTL and QTL specific to one or several isolates (Caranta et al., 1997a; Calenge et al., 2004; Rocherieux et al., 2004). Resistance QTL were also shown to be specific or non-specific to a pathogen species, and some QTL can confer resistance to multiple pathogens (Ellis et al., 2014; Wiesner-Hanks and Nelson, 2016).
While the molecular mechanisms underlying major R genes have been described extensively (Michelmore et al., 2013), those underlying resistance QTL are much less well-known. In their review, Poland et al. (2009) proposed several possible functions for the genes underlying resistance QTL. These included host plant development or morphology, basal defense, detoxification, transduction of defense signals, or partially altered major R genes. Since then, to our knowledge, only 15 genes with partial effects underlying quantitative resistance QTL have been cloned in plants (Table 1) (Niks et al., 2015; French et al., 2016). Indeed, the causal genes encode proteins with roles in pathogen recognition (NB-LRR, wall-associated kinases), signal transduction (transporters), defense (lectins, prolines), and host metabolism (transferases). Some genes are altered forms of NB-LRR genes (e.g., Pi35) or loss-of-function susceptibility alleles (e.g., Pi21). Some are expressed during a specific developmental stage (e.g., Lr34) or in a specific environment (e.g., Yr36), or confer resistance when overexpressed (e.g., Rhg1). Positional candidate genes located in resistance QTL genomic regions were also identified in recent studies, using genomics and GWA mapping approaches (Corwin and Kliebenstein, 2017). These genes were often found to encode downstream regulatory or defense response proteins, including transcription factors influencing the response to defense hormones, defensins, pathogenesis-related proteins and secondary metabolite enzymes. Genes, proteins, and metabolites regulated by quantitative resistance in response to pathogen infection were also identified using transcriptomics and metabolomics approaches (Kushalappa et al., 2016). Resistance-related proteins and metabolites involved in antimicrobial, toxin-degrading and cell-wall enforcing activities were identified in several plant species. In potato, quantitative resistance to late blight was mainly associated with cell wall thickening due to deposition of hydroxycinnamic acid amides, flavonoids, and alkaloids (Yogendra et al., 2015). In Arabidopsis thaliana, a major phytoalexin, camalexin, was induced in response to clubroot infection and associated with reduced pathogen development in heterogeneous inbred family lines carrying a major resistance QTL (Lemarie et al., 2015).
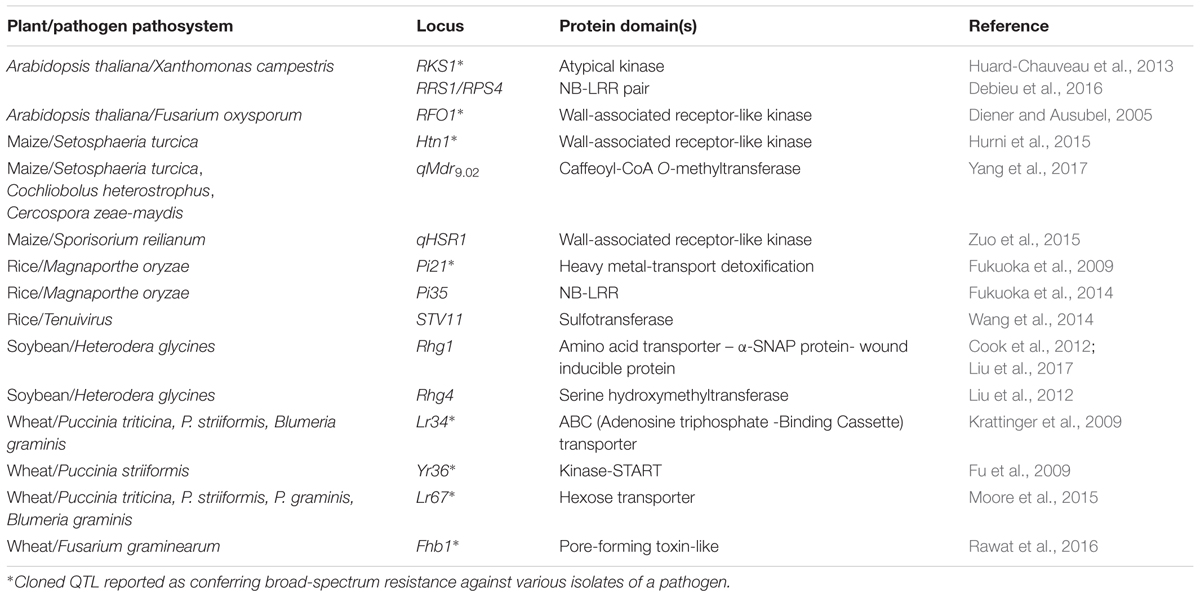
TABLE 1. Cloned genes with partial effects contributing to quantitative resistance in plants.
Quantitative resistance is generally more durable than qualitative resistance (Parlevliet, 2002). However, cases of major genes conferring durable resistance are known. For example, the Mlo gene has been used to control powdery mildew in varieties of spring barley in Europe since 1979 (Acevedo-Garcia et al., 2014). In addition, several genes conferring resistance to viruses have also shown durability (i.e., pvr22, Pvr4 in pepper, Ry in potato) (Garcia-Arenal and McDonald, 2003). However, in general, virulent isolates rapidly overcome major R genes (Brown, 2015). The increased durability of quantitative resistance could be due to (i) the partial resistance effect exerting a low selection pressure on the pathogen population, (ii) a combination of contradictory selection pressures on pathogen evolution, (iii) a low probability of multiple pathogen mutations needed to overcome multiple QTL, (iv) a combination of different resistance-associated mechanisms, which together are more difficult to overcome, and (v) a combination of resistance mechanisms acting successively at different times in the pathogen life cycle or throughout plant development (Palloix et al., 2009; Mundt, 2014). Nevertheless, experimental evolutionary studies carried out by successively passaging a pathogen on plants carrying quantitative resistance, suggested that fungi and viruses can still erode away this type of resistance (Kolmer and Leonard, 1986; Lehman and Shaner, 1997; Montarry et al., 2012). Indeed, the breakdown or erosion of quantitative resistance or resistance QTL by pathogen isolates was also observed under natural conditions. Studies of these systems have provided insight into the pathogen adaptation processes involved (Cowger and Mundt, 2002; Peressotti et al., 2010; Caffier et al., 2014, 2016; Delmas et al., 2016).
Quantitative Disease Resistance: Integration into Pyramiding Strategies for Durable Resistance
Different approaches have been proposed for adequately deploying major R genes and resistance QTL, with the aim of increasing the durability of crop resistance to pathogens. The sustainable management of available genetic resistance factors includes (i) the use of multi-line varieties or varietal mixtures carrying different R genes or QTL (Sapoukhina et al., 2013), (ii) the rotation in space or time of various R genes (Papaix et al., 2011) and (iii) the combination (i.e., pyramiding) of R genes or QTL in the same genotype (Mundt, 2014; Brown, 2015). In spite of a relative lack of data, comparative experimental and retrospective studies of these three approaches suggested that R genes may be more durable when deployed in pyramids than in varietal mixtures or rotations (Djian-Caporalino et al., 2014; Bourguet et al., 2016). However, a recent modeling approach found that rotations in space (i.e., mosaics of fields with either resistant or susceptible cultivars) were at least as efficient and durable as pyramids (Djidjou-Demasse et al., 2017).
Breeding schemes for pyramiding major R genes have been extensively trialed (Collard and Mackill, 2008; Ordon and Kühne, 2014) and resulted in the development of resistant varieties that are widely cultivated (Ellis et al., 2014). Strategies involving combinations of major genes, each conferring resistance to various specific isolates, were suggested to increase resistance durability (Feechan et al., 2015). However, the appearance of multi-virulent populations in different situations could compromise the effectiveness of resistance gene pyramids (i) when virulence mutations are not independent and generate limited fitness costs for the pathogen, (ii) when one or several virulence factors pre-exist in pathogen populations or (iii) when the modes of action of specific resistances are redundant (Brown, 2015). As the independent use of resistance genes in breeding programs is often unchecked, it is likely that pathogens gradually overcome resistance genes in deployed pyramids.
Breeding schemes for pyramiding resistance QTL were also applied to increase resistance levels in cultivated varieties, for example in barley, wheat, bean, and pepper (St. Clair, 2010). There are few reports of studies aiming to integrate disease resistance QTL in breeding strategies, in contrast to major R genes that have been widely used in plant breeding. Most focused on QTL with major effects, and very few on QTL with minor effects. Different schemes of marker-assisted-selection (MAS) have been developed to exploit and combine resistance QTL in plants, including early selection on haplotypes in F2 populations, marker-assisted back-crossing (MAB) and marker-assisted recurrent selection (MARS) (St. Clair, 2010). The successful MAS published to date were obtained essentially using the MAB method (Hospital, 2009). In several studies, this method was used to create near-isogenic lines (NILs) and validate the effects of previously detected resistance QTL (Lavaud et al., 2015).
The durability of QTL pyramids was rarely evaluated. Nevertheless, QTL combinations are expected to increase durability for different reasons.
Pyramiding resistance QTL showing a varying spectrum of action on pathogen strains may generate contradictory selection pressures on pathogen evolution. Le Van et al. (2013) showed that scab resistance QTL differentially selected for Venturia inaequalis strains co-inoculated in a mixture. Broad-spectrum QTL did not exert differential selection pressures between strains, whereas specific QTL decreased the frequency of some strains. Consequently, pyramiding QTL with different specificities or broad spectrum QTL may be expected to increase durability. Recently, the combination of two resistance QTL, which showed additive broad-spectrum effects in NILs, was suggested as a promising strategy for breeding durable resistance in rice against rice blast (Chaipanya et al., 2017).
Pyramiding resistance QTL associated with different resistance mechanisms may affect different pathogen life-history traits such as latency, infection efficiency, plant colonization, and pathogen multiplication (Table 2). Resistance QTL with diversified actions on the pathogen’s life-cycle were shown for yellow rust resistance in barley NILs (Richardson et al., 2006) and brown rust resistance in wheat (Azzimonti et al., 2014). QTL acting at different stages of plant development, corresponding to different stages of the pathogen infection cycle, were also identified in two wheat cultivars with durable resistance to yellow rust (Dedryver et al., 2009). In apple, the pyramiding of three resistance QTL to scab was suggested to result in more durable resistance since the QTL were shown to act at different stages of the fungal infection cycle, from as soon as it penetrated the plant to a later stage during subcutaneous growth and sporulation (Laloi et al., 2016). In pea NILs, combinations of resistance QTL individually acting on delaying symptom appearance and/or on slowing down root colonization by Aphanomyces euteiches had an increased action on the two pathogen life-history traits (Lavaud et al., 2016). In maize NILs, Chung et al. (2010) identified two QTL acting on different stages of the infectious cycle (penetration, colonization) of Setosphaeria turcica, one of which also conferred an accumulation of callose and phenolic compounds at the points of infection. In rice, NILs that pyramided four blast disease resistance QTL (Pi21, Pi34, qBR4-2, qBR12-1) underlying different putative functions, exhibited increased, broad-spectrum and stable levels of resistance in multiple environments (Fukuoka et al., 2015). In addition to these experimental data, theoretical studies highlighted that a combination of QTL will be more durable if the QTL affect distinct pathogen life-history traits, especially when the evolution of repressed traits is antagonistic (Bourget et al., 2015).
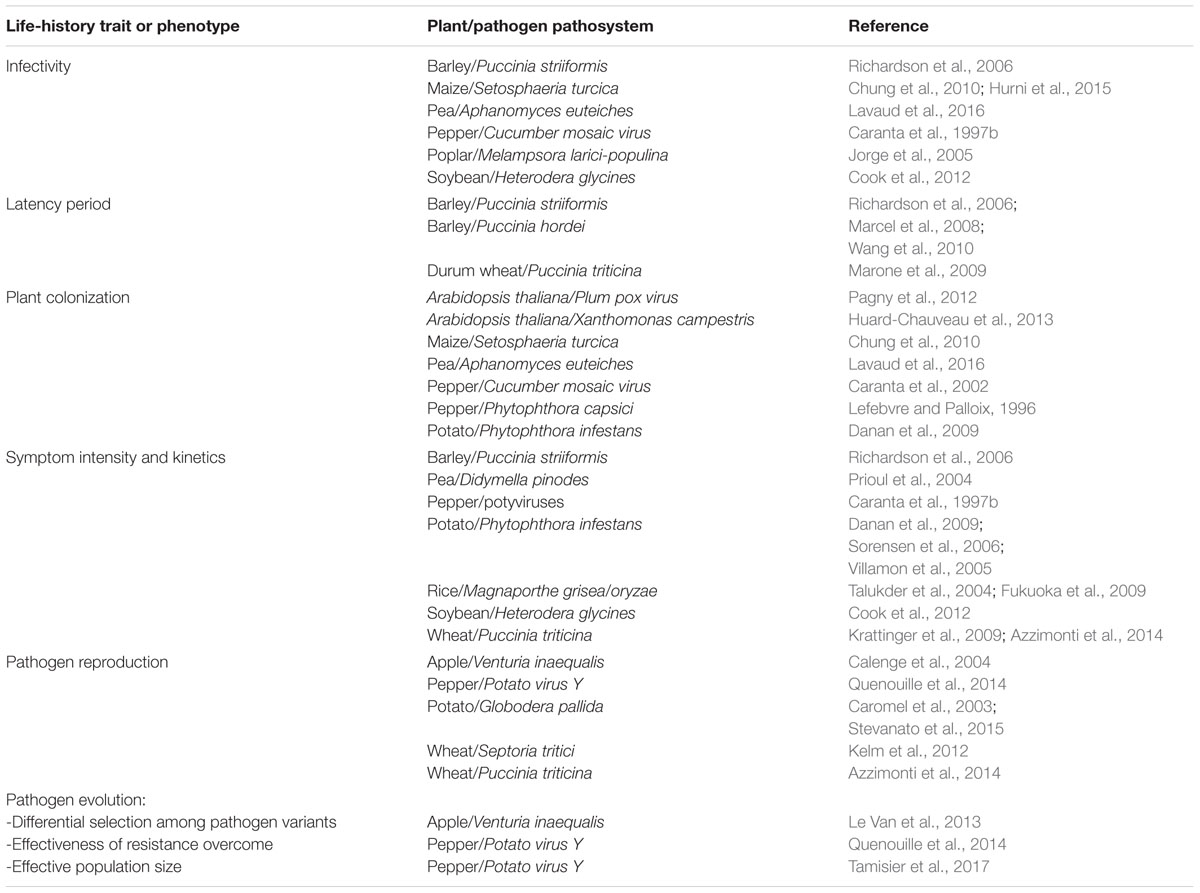
TABLE 2. Examples of life-history traits or phenotypes associated with resistance QTL in plants.
Breeding schemes for pyramiding resistance QTL and major genes were developed to increase the effectiveness of disease resistance by diversifying the putative resistance mechanisms combined (Baumgartner et al., 2015). They were also evaluated for their potential to preserve the effectiveness of resistance conferred by major genes, either by: (i) reducing the total pathogen population size, (ii) reducing the effective pathogen population size, i.e., the number of individuals that pass on their genes to the next generation, and consequently potentially increasing genetic drift, (iii) reducing the selection effects among pathogen variants or even (iv) exerting diversifying selection pressures on these variants (Quenouille et al., 2014).
In oilseed rape, major genes conferring resistance to Leptosphaeria maculans were shown to be rapidly overcome under field experimental conditions by recurrent selection of pathogen populations on the Rlm6 major gene (Brun et al., 2000) and under cultivation conditions for varieties carrying the Rlm1 or Rlm3 genes (Rouxel et al., 2003; Zhang et al., 2016). Eight years of recurrent selection were necessary to overcome the Rlm6 resistance introduced in a partially resistant genetic background, compared to only 3 years in a susceptible genetic background (Brun et al., 2010; Delourme et al., 2014).
In potato, resistance to the cyst nematode Globodera pallida conferred by a major-effect QTL (GpaVvrn) from Solanum vernei, which acts by masculinizing the nematode populations, was overcome after 8 years of recurrent selection in the laboratory. The speed at which the resistance was overcome depended on the genetic background into which GpaVvrn was introduced (Fournet et al., 2013), suggesting that low-effect QTL could enhance the durability of QTL GpaVvrn. In the case of resistances to Meloidogyne spp. root-knot nematodes, Barbary et al. (2014, 2016) also demonstrated that the effectiveness and durability of the major genes Me1 and Me3 depended on the genetic background into which they were introduced.
In pepper, the major pvr23 resistance allele to Potato virus Y (PVY) was rapidly overcome under controlled experimental conditions (Palloix et al., 2009; Quenouille et al., 2014). The combination of pvr23 with three partial resistance QTL significantly increased resistance durability under the same conditions. Durability QTL controlling the frequency of plants with pvr23-resistance breakdown were mapped onto the pepper genome (Quenouille et al., 2014). The evolutionary mechanisms underlying the protective effect of the resistance QTL on the major gene present in the genetic background appeared to be multiple (Quenouille et al., 2013). These included: (i) the reduced ability of PVY to multiply, (ii) the increased number of mutations required for the virus to become virulent and (iii) the slowed-down selection of virulent PVY variants.
Conclusion and Perspectives
The development of molecular markers has led to an increase in the use of disease resistance QTL in breeding. However, minor-effect QTL are still difficult to exploit and resistance can have physiological costs for the plant (Brown and Rant, 2013). The development of genomics over the last 10 years has opened up the prospects for genomic selection of quantitative resistance, which should lead to an improved consideration of minor-effect resistance QTL in breeding programs (Poland and Rutkoski, 2016). This progress has also made it possible to reduce confidence intervals and fine-map resistance QTL, so that unfavorable linkages, such as late maturity and partial resistance to Phytophthora infestans in potato (Muktar et al., 2015), can be broken. It has also allowed favorable pleiotropic effects of resistance loci to be identified, such as the QTL Lr34 which confers resistance to multiple diseases in cereals (Kolmer et al., 2008; Krattinger et al., 2016), or the major locus Xa4 which improves multiple agronomic traits by strengthening the cell wall (Hu et al., 2017). However, phenotyping methods still need to be improved, especially for evaluating resistance components which best predict field quantitative resistance. In this way, the actions of QTL on pathogen epidemics could be better described and their use in breeding optimized (Huang et al., 2009; Willocquet et al., 2017).
Quantitative resistance appears to be more durable than qualitative resistance and has the potential to preserve major-gene effects. However, there is still a need to adequately choose resistance QTL to create optimal combinations and limit QTL erosion. The choice of QTL could be based on their spectrum of action on different strains, their effect on pathogen life traits and their underlying molecular mechanisms (Niks et al., 2015; Kushalappa et al., 2016). Recently, plant resistance genes targeted by conserved essential pathogen effectors were suggested to confer more durable resistance than those targeted by non-essential effectors (Dangl, 2013; Clarke et al., 2015). Furthermore, the development of genome editing approaches will open new prospects for creating novel specificities in resistance genes which can then be combined to preserve resistance effectiveness (Andolfo et al., 2016).
Breeding strategies for quantitative resistance still require a better understanding of the genetic, ecological and agronomic determinants of pathogen adaptation involved in QTL erosion. Further knowledge is needed on the evolutionary capacity of pathogen species, which would depend on their mutation rate, reproduction mode, dispersal ability and effective population size (McDonald and Linde, 2002). Five recent studies identified bacterial and fungal genetic determinants of pathogenicity using GWA mapping (Bartoli and Roux, 2017). A genome scan approach also identified pathogen genomic regions involved in adaptation to resistance QTL (Eoche-Bosy et al., 2017a,b). Finally, in order to reduce the risks of pathogen adaptation, resistance breeding schemes should be more thoroughly integrated with deployment strategies of resistant varieties and combination strategies of diverse disease control methods (Didelot et al., 2016).
Author Contributions
All the authors listed made substantial intellectual contribution to the work, wrote the manuscript together and approved it for publication.
Funding
This work was supported by the “PeaMUST” project (Pea MUlti-STress adaptation and biological regulations for yield improvement and stability; Investments for the Future call 2011 ANR-11-BTBR-0002) and the CTPS grant C07-01-pomme de terre, from the French Government. It also benefited from the INRA metaprogram SMaCH – Sustainable Management of Plant Health – especially the projects “ARAMIS” (Apple partial Resistance durability Assessed through MetabolIc pathways and pathogen adaptation to Selective pressures) and “Take Control” (Deployment strategies of plant quantitative resistance to take control of plant pathogen evolution). It was supported by the European Network of Excellence “ENDURE” (European Network for the Durable Exploitation of Crop Protection).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors thank colleagues from UMRs IGEPP, IRHS, and GAFL for their contributions to the works cited, with a special tribute to Alain Palloix for his decisive and visionary role in the development of INRA research on durability and durable management of resistances. The authors apologize for the necessary omission of bibliography of interest, due to manuscript length limitations.
References
Acevedo-Garcia, J., Kusch, S., and Panstruga, R. (2014). Magical mystery tour: MLO proteins in plant immunity and beyond. New Phytol. 204, 273–281. doi: 10.1111/nph.12889
Andolfo, G., Lovieno, P., Frusciante, L., and Ercolano, M. R. (2016). Genome-editing technologies for enhancing plant disease resistance. Front. Plant Sci. 7:1813. doi: 10.3389/fpls.2016.01813
Azzimonti, G., Marcel, T. C., Robert, O., Paillard, S., Lannou, C., and Goyeau, H. (2014). Diversity, specificity and impacts on field epidemics of QTLs involved in components of quantitative resistance in the wheat leaf rust pathosystem. Mol. Breed. 34, 549–567. doi: 10.1007/s11032-014-0057-8
Ballini, E., Morel, J. B., Droc, G., Price, A., Courtois, B., Notteghem, J. L., et al. (2008). A genome-wide meta-analysis of rice blast resistance genes and quantitative trait loci provides new insights into partial and complete resistance. Mol. Plant Microbe Interact. 21, 859–868. doi: 10.1094/mpmi-21-7-0859
Barbary, A., Djian-Caporalino, C., Marteu, N., Fazari, A., Caromel, B., Castagnone-Sereno, P., et al. (2016). Plant genetic background increasing the efficiency and durability of major resistance genes to root-knot nematodes can be resolved into a few resistance QTLs. Front. Plant Sci. 7:632. doi: 10.3389/fpls.2016.00632
Barbary, A., Palloix, A., Fazari, A., Marteu, N., Castagnone-Sereno, P., and Djian-Caporalino, C. (2014). The plant genetic background affects the efficiency of the pepper major nematode resistance genes Me1 and Me3. Theor. Appl. Genet. 127, 499–507. doi: 10.1007/s00122-013-2235-1
Bartoli, C., and Roux, F. (2017). Genome-wide association studies in plant pathosystems: toward an ecological genomics approach. Front. Plant Sci. 8:763. doi: 10.3389/fpls.2017.00763
Baumgartner, I. O., Patocchi, A., Frey, J. E., Peil, A., and Kellerhals, M. (2015). Breeding elite lines of apple carrying pyramided homozygous resistance genes against apple scab and resistance against powdery mildew and fire blight. Plant Mol. Biol. Rep. 33, 1573–1583. doi: 10.1007/s11105-015-0858-x
Bourget, R., Chaumont, L., Durel, C. E., and Sapoukhina, N. (2015). Sustainable deployment of QTLs conferring quantitative resistance to crops: first lessons from a stochastic model. New Phytol. 206, 1163–1171. doi: 10.1111/nph.13295
Bourguet, D., Delmotte, F., Franck, P., Guillemaud, T., Reboud, X., Vacher, C., et al. (2016). Combining selective pressures to enhance the durability of disease resistance genes. Front. Plant Sci. 7:1916. doi: 10.3389/fpls.2016.01916
Brown, J. K. M. (2015). “Durable resistance of crops to disease: a darwinian perspective,” in Annual Review of Phytopathology, ed. N. K. VanAlfen (Palo Alto, CA: Annual Reviews), 513–539.
Brown, J. K. M., and Rant, J. C. (2013). Fitness costs and trade-offs of disease resistance and their consequences for breeding arable crops. Plant Pathol. 62, 83–95. doi: 10.1111/ppa.12163
Brun, H., Chevre, A. M., Fitt, B. D. L., Powers, S., Besnard, A. L., Ermel, M., et al. (2010). Quantitative resistance increases the durability of qualitative resistance to Leptosphaeria maculans in Brassica napus. New Phytol. 185, 285–299. doi: 10.1111/j.1469-8137.2009.03049.x
Brun, H., Levivier, S., Somda, I., Ruer, D., Renard, M., and Chevre, A. M. (2000). A field method for evaluating the potential durability of new resistance sources: application to the Leptosphaeria maculans-Brassica napus pathosystem. Phytopathology 90, 961–966. doi: 10.1094/phyto.2000.90.9.961
Caffier, V., Lasserre-Zuber, P., Giraud, M., Lascostes, M., Stievenard, R., Lemarquand, A., et al. (2014). Erosion of quantitative host resistance in the apple x Venturia inaequalis pathosystem. Infect. Genet. Evol. 27, 481–489. doi: 10.1016/j.meegid.2014.02.003
Caffier, V., Le Cam, B., Al Rifai, M., Bellanger, M. N., Comby, M., Denance, C., et al. (2016). Slow erosion of a quantitative apple resistance to Venturia inaequalis based on an isolate-specific quantitative trait Locus. Infect. Genet. Evol. 44, 541–548. doi: 10.1016/j.meegid.2016.07.016
Calenge, F., Drouet, D., Denance, C., Van de Weg, W. E., Brisset, M. N., Paulin, J. P., et al. (2005). Identification of a major QTL together with several minor additive or epistatic QTLs for resistance to fire blight in apple in two related progenies. Theor. Appl. Genet. 111, 128–135. doi: 10.1007/s00122-005-2002-z
Calenge, F., and Durel, C. E. (2006). Both stable and unstable QTLs for resistance to powdery mildew are detected in apple after four years of field assessments. Mol. Breed. 17, 329–339. doi: 10.1007/s11032-006-9004-7
Calenge, F., Faure, A., Goerre, M., Gebhardt, C., Van de Weg, W. E., Parisi, L., et al. (2004). Quantitative trait loci (QTL) analysis reveals both broad-spectrum and isolate-specific QTL for scab resistance in an apple progeny challenged with eight isolates of Venturia inaequalis. Phytopathology 94, 370–379. doi: 10.1094/phyto.2004.94.4.370
Caranta, C., Lefebvre, V., and Palloix, A. (1997a). Polygenic resistance of pepper to potyviruses consists of a combination of isolate-specific and broad-spectrum quantitative trait loci. Mol. Plant Microbe Interact. 10, 872–878. doi: 10.1094/mpmi.1997.10.7.872
Caranta, C., Palloix, A., Lefebvre, V., and Daubeze, A. M. (1997b). QTLs for a component of partial resistance to cucumber mosaic virus in pepper: restriction of virus installation in host-cells. Theor. Appl. Genet. 94, 431–438. doi: 10.1007/s001220050433
Caranta, C., Pflieger, S., Lefebvre, V., Daubeze, A. M., Thabuis, A., and Palloix, A. (2002). QTLs involved in the restriction of cucumber mosaic virus (CMV) long-distance movement in pepper. Theor. Appl. Genet. 104, 586–591. doi: 10.1007/s001220100753
Caromel, B., Mugniery, D., Lefebvre, V., Andrzejewski, S., Ellisseche, D., Kerlan, M. C., et al. (2003). Mapping QTLs for resistance against Globodera pallida (Stone) Pa2/3 in a diploid potato progeny originating from Solanum spegazzinii. Theor. Appl. Genet. 106, 1517–1523. doi: 10.1007/s00122-003-1211-6
Chaipanya, C., Telebanco-Yanoria, M. J., Quime, B., Longya, A., Korinsak, S., Korinsak, S., et al. (2017). Dissection of broad-spectrum resistance of the Thai rice variety Jao Hom Nin conferred by two resistance genes against rice blast. Rice 10, 18. doi: 10.1186/s12284-017-0159-0
Chung, C. L., Longfellow, J. M., Walsh, E. K., Kerdieh, Z., Van Esbroeck, G., Balint-Kurti, P., et al. (2010). Resistance loci affecting distinct stages of fungal pathogenesis: use of introgression lines for QTL mapping and characterization in the maize - Setosphaeria turcica pathosystem. BMC Plant Biol. 10:103. doi: 10.1186/1471-2229-10-103
Clarke, C. R., Studholme, D. J., Hayes, B., Runde, B., Weisberg, A., Cai, R. M., et al. (2015). Genome-enabled phylogeographic investigation of the quarantine pathogen Ralstonia solanacearum Race 3 Biovar 2 and screening for sources of resistance against its core effectors. Phytopathology 105, 597–607. doi: 10.1094/phyto-12-14-0373-r
Collard, B. C. Y., and Mackill, D. J. (2008). Marker-assisted selection: an approach for precision plant breeding in the twenty-first century. Philos. Trans. R. Soc. B Biol. Sci. 363, 557–572. doi: 10.1098/rstb.2007.2170
Cook, D. E., Lee, T. G., Guo, X. L., Melito, S., Wang, K., Bayless, A. M., et al. (2012). Copy number variation of multiple genes at Rhg1 mediates nematode resistance in soybean. Science 338, 1206–1209. doi: 10.1126/science.1228746
Corwin, J. A., and Kliebenstein, D. J. (2017). Quantitative resistance: more than just perception of a pathogen. Plant Cell 29, 655–665. doi: 10.1105/tpc.16.00915
Cowger, C., and Mundt, C. C. (2002). Aggressiveness of Mycosphaerella graminicola isolates from susceptible and partially resistant wheat cultivars. Phytopathology 92, 624–630. doi: 10.1094/phyto.2002.92.6.624
Danan, S., Chauvin, J. E., Caromel, B., Moal, J. D., Pelle, R., and Lefebvre, V. (2009). Major-effect QTLs for stem and foliage resistance to late blight in the wild potato relatives Solanum sparsipilum and S. spegazzinii are mapped to chromosome X. Theor. Appl. Genet. 119, 705–719. doi: 10.1007/s00122-009-1081-7
Danan, S., Veyrieras, J. B., and Lefebvre, V. (2011). Construction of a potato consensus map and QTL meta-analysis offer new insights into the genetic architecture of late blight resistance and plant maturity traits. BMC Plant Biol. 11:16. doi: 10.1186/1471-2229-11-16
Dangl, J. L. (2013). Pivoting the plant immune system from dissection to deployment. Science 341, 1175–1175. doi: 10.1126/science.1236011
Debieu, M., Huard-Chauveau, C., Genissel, A., Roux, F., and Roby, D. (2016). Quantitative disease resistance to the bacterial pathogen Xanthomonas campestris involves an Arabidopsis immune receptor pair and a gene of unknown function. Mol. Plant Pathol. 17, 510–520. doi: 10.1111/mpp.12298
Dedryver, F., Paillard, S., Mallard, S., Robert, O., Trottet, M., Negre, S., et al. (2009). Characterization of genetic components involved in durable resistance to stripe rust in the bread wheat ’Renan’. Phytopathology 99, 968–973. doi: 10.1094/phyto-99-8-0968
Delmas, C. E. L., Fabre, F., Jolivet, J., Mazet, I. D., Cervera, S. R., Deliere, L., et al. (2016). Adaptation of a plant pathogen to partial host resistance: selection for greater aggressiveness in grapevine downy mildew. Evol. Appl. 9, 709–725. doi: 10.1111/eva.12368
Delourme, R., Bousset, L., Ermel, M., Duffe, P., Besnard, A. L., Marquer, B., et al. (2014). Quantitative resistance affects the speed of frequency increase but not the diversity of the virulence alleles overcoming a major resistance gene to Leptosphaeria maculans in oilseed rape. Infect. Genet. Evol. 27, 490–499. doi: 10.1016/j.meegid.2013.12.019
Desgroux, A., L’Anthoene, V., Roux-Duparque, M., Riviere, J. P., Aubert, G., Tayeh, N., et al. (2016). Genome-wide association mapping of partial resistance to Aphanomyces euteiches in pea. BMC Genomics 17:124. doi: 10.1186/s12864-016-2429-4
Didelot, F., Caffier, V., Orain, G., Lemarquand, A., and Parisi, L. (2016). Sustainable management of scab control through the integration of apple resistant cultivars in a low-fungicide input system. Agric. Ecosyst. Environ. 217, 41–48. doi: 10.1016/j.agee.2015.10.023
Diener, A. C., and Ausubel, F. M. (2005). Resistance to Fusarium oxysporum 1, a dominant Arabidopsis disease-resistance gene, is not race specific. Genetics 171, 305–321. doi: 10.1534/genetics.105.042218
Djian-Caporalino, C., Palloix, A., Fazari, A., Marteu, N., Barbary, A., Abad, P., et al. (2014). Pyramiding, alternating or mixing: comparative performances of deployment strategies of nematode resistance genes to promote plant resistance efficiency and durability. BMC Plant Biol. 14:53. doi: 10.1186/1471-2229-14-53
Djidjou-Demasse, R., Moury, B., and Fabre, F. (2017). Mosaics often outperform pyramids: insights from a model comparing strategies for the deployment of plant resistance genes against viruses in agricultural landscapes. New Phytol. 216, 239–253. doi: 10.1111/nph.14701
Ellis, J. G., Lagudah, E. S., Spielmeyer, W., and Dodds, P. N. (2014). The past, present and future of breeding rust resistant wheat. Front. Plant Sci. 5:641. doi: 10.3389/fpls.2014.00641
Eoche-Bosy, D., Gauthier, J., Juhel, A. S., Esquibet, M., Fournet, S., Grenier, E., et al. (2017a). Experimentally evolved populations of the potato cyst nematode Globodera pallida allow the targeting of genomic footprints of selection due to host adaptation. Plant Pathol. 66, 1022–1030. doi: 10.1111/ppa.12646
Eoche-Bosy, D., Gautier, M., Esquibet, M., Legeai, F., Bretaudeau, A., Bouchez, O., et al. (2017b). Genome scans on experimentally evolved populations reveal candidate regions for adaptation to plant resistance in the potato cyst nematode Globodera pallida. Mol. Ecol. 26, 4700–4711. doi: 10.1111/mec.14240
Feechan, A., Kocsis, M., Riaz, S., Zhang, W., Gadoury, D. M., Walker, M. A., et al. (2015). Strategies for RUN1 deployment using RUN2 and REN2 to manage grapevine powdery mildew informed by studies of race specificity. Phytopathology 105, 1104–1113. doi: 10.1094/phyto-09-14-0244-r
Fournet, S., Kerlan, M. C., Renault, L., Dantec, J. P., Rouaux, C., and Montarry, J. (2013). Selection of nematodes by resistant plants has implications for local adaptation and cross-virulence. Plant Pathol. 62, 184–193. doi: 10.1111/j.1365-3059.2012.02617.x
French, E., Kim, B. S., and Iyer-Pascuzzi, A. S. (2016). Mechanisms of quantitative disease resistance in plants. Semin. Dev. Biol. 56, 201–208. doi: 10.1016/j.semcdb.2016.05.015
Fu, D. L., Uauy, C., Distelfeld, A., Blechl, A., Epstein, L., Chen, X. M., et al. (2009). A Kinase-START gene confers temperature-dependent resistance to wheat stripe rust. Science 323, 1357–1360. doi: 10.1126/science.1166289
Fukuoka, S., Saka, N., Koga, H., Ono, K., Shimizu, T., Ebana, K., et al. (2009). Loss of function of a proline-containing protein confers durable disease resistance in rice. Science 325, 998–1001. doi: 10.1126/science.1175550
Fukuoka, S., Saka, N., Mizukami, Y., Koga, H., Yamanouchi, U., Yoshioka, Y., et al. (2015). Gene pyramiding enhances durable blast disease resistance in rice. Sci. Rep. 5:7773. doi: 10.1038/srep07773
Fukuoka, S., Yamamoto, S. I., Mizobuchi, R., Yamanouchi, U., Ono, K., Kitazawa, N., et al. (2014). Multiple functional polymorphisms in a single disease resistance gene in rice enhance durable resistance to blast. Sci. Rep. 4:4550. doi: 10.1038/srep04550
Garcia-Arenal, F., and McDonald, B. A. (2003). An analysis of the durability of resistance to plant viruses. Phytopathology 93, 941–952. doi: 10.1094/phyto.2003.93.8.941
Goudemand, E., Laurent, V., Duchalais, L., Ghaffary, S. M. T., Kema, G. H. J., Lonnet, P., et al. (2013). Association mapping and meta-analysis: two complementary approaches for the detection of reliable Septoria tritici blotch quantitative resistance in bread wheat (Triticum aestivum L.). Mol. Breed. 32, 563–584. doi: 10.1007/s11032-013-9890-4
Hamon, C., Baranger, A., Coyne, C. J., McGee, R. J., Le Goff, I., L’Anthoene, V., et al. (2011). New consistent QTL in pea associated with partial resistance to Aphanomyces euteiches in multiple French and American environments. Theor. Appl. Genet. 123, 261–281. doi: 10.1007/s00122-011-1582-z
Hospital, F. (2009). Challenges for effective marker-assisted selection in plants. Genetica 136, 303–310. doi: 10.1007/s10709-008-9307-1
Hu, K. M., Cao, J. B., Zhang, J., Xia, F., Ke, Y. G., Zhang, H. T., et al. (2017). Improvement of multiple agronomic traits by a disease resistance gene via cell wall reinforcement. Nat. Plants 3:17009. doi: 10.1038/nplants.2017.9
Huang, X. H., and Han, B. (2014). “Natural variations and genome-wide association studies in crop plants,” in Annual Review of Plant Biology, Vol. 65, ed. S. S. Merchant (Palo Alto, CA: Annual Reviews), 531–551.
Huang, Y. J., Pirie, E. J., Evans, N., Delourme, R., King, G. J., and Fitt, B. D. L. (2009). Quantitative resistance to symptomless growth of Leptosphaeria maculans (phoma stem canker) in Brassica napus (oilseed rape). Plant Pathol. 58, 314–323. doi: 10.1111/j.1365-3059.2008.01957.x
Huard-Chauveau, C., Perchepied, L., Debieu, M., Rivas, S., Kroj, T., Kars, I., et al. (2013). An atypical kinase under balancing selection confers broad-spectrum disease resistance in Arabidopsis. PLOS Genet. 9:e1003766. doi: 10.1371/journal.pgen.1003766
Hurni, S., Scheuermann, D., Krattinger, S. G., Kessel, B., Wicker, T., Herren, G., et al. (2015). The maize disease resistance gene Htn1 against northern corn leaf blight encodes a wall-associated receptor-like kinase. Proc. Natl. Acad. Sci. U.S.A. 112, 8780–8785. doi: 10.1073/pnas.1502522112
Jorge, V., Dowkiw, A., Faivre-Rampant, P., and Bastien, C. (2005). Genetic architecture of qualitative and quantitative Melampsora larici-populina leaf rust resistance in hybrid poplar: genetic mapping and QTL detection. New Phytol. 167, 113–127. doi: 10.1111/j.1469-8137.2005.01424.x
Kelm, C., Ghaffary, S. M. T., Bruelheide, H., Roder, M. S., Miersch, S., Weber, W. E., et al. (2012). The genetic architecture of seedling resistance to Septoria tritici blotch in the winter wheat doubled-haploid population Solitar x Mazurka. Mol. Breed. 29, 813–830. doi: 10.1007/s11032-011-9592-8
Kolmer, J. A., and Leonard, K. J. (1986). Genetic selection and adaptation of Cochliobolus heterostrophus to corn host with partial resistance. Phytopathology 76, 774–777. doi: 10.1094/Phyto-76-774
Kolmer, J. A., Singh, R. P., Garvin, D. F., Viccars, L., William, H. M., Huerta-Espino, J., et al. (2008). Analysis of the Lr34/Yr18 rust resistance region in wheat germplasm. Crop Sci. 48, 1841–1852. doi: 10.2135/cropsci2007.08.0474
Kover, P. X., and Caicedo, A. L. (2001). The genetic architecture of disease resistance in plants and the maintenance of recombination by parasites. Mol. Ecol. 10, 1–16. doi: 10.1046/j.1365-294X.2001.01124.x
Krattinger, S. G., Lagudah, E. S., Spielmeyer, W., Singh, R. P., Huerta-Espino, J., McFadden, H., et al. (2009). A putative ABC transporter confers durable resistance to multiple fungal pathogens in wheat. Science 323, 1360–1363. doi: 10.1126/science.1166453
Krattinger, S. G., Sucher, J., Selter, L. L., Chauhan, H., Zhou, B., Tang, M. Z., et al. (2016). The wheat durable, multipathogen resistance gene Lr34 confers partial blast resistance in rice. Plant Biotechnol. J. 14, 1261–1268. doi: 10.1111/pbi.12491
Kushalappa, A. C., Yogendra, K. N., and Karre, S. (2016). Plant innate immune response: qualitative and quantitative resistance. Crit. Rev. Plant Sci. 35, 38–55. doi: 10.1080/07352689.2016.1148980
Laloi, G., Vergne, E., Durel, C.-E., Le Cam, B., and Caffier, V. (2016). Efficiency of pyramiding of three quantitative resistance loci to apple scab. Plant Pathol. 66, 412–422. doi: 10.1111/ppa.12581
Lavaud, C., Baviere, M., Le Roy, G., Herve, M. R., Moussart, A., Delourme, R., et al. (2016). Single and multiple resistance QTL delay symptom appearance and slow down root colonization by Aphanomyces euteiches in pea near isogenic lines. BMC Plant Biol. 16:166. doi: 10.1186/s12870-016-0822-4
Lavaud, C., Lesne, A., Piriou, C., Le Roy, G., Boutet, G., Moussart, A., et al. (2015). Validation of QTL for resistance to Aphanomyces euteiches in different pea genetic backgrounds using near-isogenic lines. Theor. Appl. Genet. 128, 2273–2288. doi: 10.1007/s00122-015-2583-0
Le Van, A., Caffier, V., Lasserre-Zuber, P., Chauveau, A., Brunel, D., Le Cam, B., et al. (2013). Differential selection pressures exerted by host resistance quantitative trait loci on a pathogen population: a case study in an apple Venturia inaequalis pathosystem. New Phytol. 197, 899–908. doi: 10.1111/nph.12086
Lefebvre, V., and Palloix, A. (1996). Both epistatic and additive effects of QTLs are involved in polygenic induced resistance to disease: a case study, the interaction pepper - Phytophthora capsici Leonian. Theor. Appl. Genet. 93, 503–511. doi: 10.1007/bf00417941
Lehman, J. S., and Shaner, G. (1997). Selection of populations of Puccinia recondita f. sp. tritici for shortened latent period on a partially resistant wheat cultivar. Phytopathology 87, 170–176. doi: 10.1094/phyto.1997.87.2.170
Lemarie, S., Robert-Seilaniantz, A., Lariagon, C., Lemoine, J., Marnet, N., Levrel, A., et al. (2015). Camalexin contributes to the partial resistance of Arabidopsis thaliana to the biotrophic soilborne protist Plasmodiophora brassicae. Front. Plant Sci. 6:539. doi: 10.3389/fpls.2015.00539
Liu, S. M., Kandoth, P. K., Lakhssassi, N., Kang, J. W., Colantonio, V., Heinz, R., et al. (2017). The soybean GmSNAP18 gene underlies two types of resistance to soybean cyst nematode. Nat. Commun. 8:14822. doi: 10.1038/ncomms14822
Liu, S. M., Kandoth, P. K., Warren, S. D., Yeckel, G., Heinz, R., Alden, J., et al. (2012). A soybean cyst nematode resistance gene points to a new mechanism of plant resistance to pathogens. Nature 492, 256–260. doi: 10.1038/nature11651
Manzanares-Dauleux, M. J., Delourme, R., Baron, F., and Thomas, G. (2000). Mapping of one major gene and of QTLs involved in resistance to clubroot in Brassica napus. Theor. Appl. Genet. 101, 885–891. doi: 10.1007/s001220051557
Marcel, T. C., Gorguet, B., Ta, M. T., Kohutova, Z., Vels, A., and Niks, R. E. (2008). Isolate specificity of quantitative trait loci for partial resistance of barley to Puccinia hordei confirmed in mapping populations and near-isogenic lines. New Phytol. 177, 743–755. doi: 10.1111/j.1469-8137.2007.02298.x
Marone, D., Del Olmo, A. I., Laido, G., Sillero, J. C., Emeran, A. A., Russo, M. A., et al. (2009). Genetic analysis of durable resistance against leaf rust in durum wheat. Mol. Breed. 24, 25–39. doi: 10.1007/s11032-009-9268-9
McDonald, B. A., and Linde, C. (2002). Pathogen population genetics, evolutionary potential, and durable resistance. Annu. Rev. Phytopathol. 40, 349–379. doi: 10.1146/annurev.phyto.40.120501.101443
Michelmore, R. W., Christopoulou, M., and Caldwell, K. S. (2013). “Impacts of resistance gene genetics, function, and evolution on a durable future,” in Annual Review of Phytopathology, Vol. 51, ed. N. K. VanAlfen (Palo Alto, CA: Annual Reviews), 291–319.
Montarry, J., Cartier, E., Jacquemond, M., Palloix, A., and Moury, B. (2012). Virus adaptation to quantitative plant resistance: erosion or breakdown? J. Evol. Biol. 25, 2242–2252. doi: 10.1111/j.1420-9101.2012.02600.x
Moore, J. W., Herrera-Foessel, S., Lan, C. X., Schnippenkoetter, W., Ayliffe, M., Huerta-Espino, J., et al. (2015). A recently evolved hexose transporter variant confers resistance to multiple pathogens in wheat. Nat. Genet. 47, 1494–1498. doi: 10.1038/ng.3439
Muktar, M. S., Lubeck, J., Strahwald, J., and Gebhardt, C. (2015). Selection and validation of potato candidate genes for maturity corrected resistance to Phytophthora infestans based on differential expression combined with SNP association and linkage mapping. Front. Genet. 6:294. doi: 10.3389/fgene.2015.00294
Mundt, C. C. (2014). Durable resistance: a key to sustainable management of pathogens and pests. Infect. Genet. Evol. 27, 446–455. doi: 10.1016/j.meegid.2014.01.011
Niks, R. E., Qi, X. Q., and Marcel, T. C. (2015). “Quantitative resistance to biotrophic filamentous plant pathogens: concepts, misconceptions and mechanisms,” in Annual Review of Phytopathology, ed. N. K. VanAlfen (Palo Alto, CA: Annual Reviews), 445–470.
Ordon, F., and Kühne, T. (2014). “Response to viral pathogens,” in Biotechnological Approaches to Barley Improvement: Biotechnology in Agriculture and Forestry, Vol. 69, Chapter. 10, eds J. Kumlehn and N. Stein (Berlin: Springer-Verlag), 181–196. doi: 10.1007/978-3-662-44406-1_10es
Pagny, G., Paulstephenraj, P. S., Poque, S., Sicard, O., Cosson, P., Eyquard, J. P., et al. (2012). Family-based linkage and association mapping reveals novel genes affecting Plum pox virus infection in Arabidopsis thaliana. New Phytol. 196, 873–886. doi: 10.1111/j.1469-8137.2012.04289.x
Palloix, A., Ayme, V., and Moury, B. (2009). Durability of plant major resistance genes to pathogens depends on the genetic background, experimental evidence and consequences for breeding strategies. New Phytol. 183, 190–199. doi: 10.1111/j.1469-8137.2009.02827.x
Papaix, J., Goyeau, H., Du Cheyron, P., Monod, H., and Lannou, C. (2011). Influence of cultivated landscape composition on variety resistance: an assessment based on wheat leaf rust epidemics. New Phytol. 191, 1095–1107. doi: 10.1111/j.1469-8137.2011.03764.x
Parlevliet, J. E. (2002). Durability of resistance against fungal, bacterial and viral pathogens; present situation. Euphytica 124, 147–156. doi: 10.1023/a:1015601731446
Peressotti, E., Wiedemann-Merdinoglu, S., Delmotte, F., Bellin, D., Di Gaspero, G., Testolin, R., et al. (2010). Breakdown of resistance to grapevine downy mildew upon limited deployment of a resistant variety. BMC Plant Biol. 10:147. doi: 10.1186/1471-2229-10-147
Poland, J., and Rutkoski, J. (2016). “Advances and challenges in genomic selection for disease resistance,” in Annual Review of Phytopathology, eds J. E. Leach and S. Lindow (Palo Alto, CA: Annual Reviews), 79–98.
Poland, J. A., Balint-Kurti, P. J., Wisser, R. J., Pratt, R. C., and Nelson, R. J. (2009). Shades of gray: the world of quantitative disease resistance. Trends Plant Sci. 14, 21–29. doi: 10.1016/j.tplants.2008.10.006
Prioul, S., Frankewitz, A., Deniot, G., Morin, G., and Baranger, A. (2004). Mapping of quantitative trait loci for partial resistance to Mycosphaerella pinodes in pea (Pisum sativum L.), at the seedling and adult plant stages. Theor. Appl. Genet. 108, 1322–1334. doi: 10.1007/s00122-003-1543-2
Quenouille, J., Montarry, J., Palloix, A., and Moury, B. (2013). Farther, slower, stronger: how the plant genetic background protects a major resistance gene from breakdown. Mol. Plant Pathol. 14, 109–118. doi: 10.1111/j.1364-3703.2012.00834.x
Quenouille, J., Paulhiac, E., Moury, B., and Palloix, A. (2014). Quantitative trait loci from the host genetic background modulate the durability of a resistance gene: a rational basis for sustainable resistance breeding in plants. Heredity 112, 579–587. doi: 10.1038/hdy.2013.138
Rawat, N., Pumphrey, M. O., Liu, S. X., Zhang, X. F., Tiwari, V. K., Ando, K., et al. (2016). Wheat Fhb1 encodes a chimeric lectin with agglutinin domains and a pore-forming toxin-like domain conferring resistance to Fusarium head blight. Nat. Genet. 48, 1576–1580. doi: 10.1038/ng.3706
Richardson, K. L., Vales, M. I., Kling, J. G., Mundt, C. C., and Hayes, P. M. (2006). Pyramiding and dissecting disease resistance QTL to barley stripe rust. Theor. Appl. Genet. 113, 485–495. doi: 10.1007/s00122-006-0314-2
Rocherieux, J., Glory, P., Giboulot, A., Boury, S., Barbeyron, G., Thomas, G., et al. (2004). Isolate-specific and broad-spectrum QTLs are involved in the control of clubroot in Brassica oleracea. Theor. Appl. Genet. 108, 1555–1563. doi: 10.1007/s00122-003-1580-x
Rouxel, T., Penaud, A., Pinochet, X., Brun, H., Gout, L., Delourme, R., et al. (2003). A 10-year survey of populations of Leptosphaeria maculans in France indicates a rapid adaptation towards the Rlm1 resistance gene of oilseed rape. Eur. J. Plant Pathol. 109, 871–881. doi: 10.1023/a:1026189225466
Sapoukhina, N., Paillard, S., Dedryver, F., and de Vallavieille-Pope, C. (2013). Quantitative plant resistance in cultivar mixtures: wheat yellow rust as a modeling case study. New Phytol. 200, 888–897. doi: 10.1111/nph.12413
Sorensen, K. K., Madsen, M. H., Kirk, H. G., Madsen, D. K., and Torp, A. M. (2006). Linkage and quantitative trait locus mapping of foliage late blight resistance in the wild species Solanum vernei. Plant Breed. 125, 268–276. doi: 10.1111/j.1439-0523.2006.01219.x
St. Clair, D. A. (2010). “Quantitative disease resistance and quantitative resistance loci in breeding,” in Annual Review of Phytopathology, eds N. K. VanAlfen, G. Bruening, and J. E. Leach (Palo Alto, CA: Annual Reviews), 247–268.
Stall, R. E., Jones, J. B., and Minsavage, G. V. (2009). Durability of resistance in tomato and pepper to xanthomonads causing bacterial spot. Annu. Rev. Phytopathol. 47, 265–284. doi: 10.1146/annurev-phyto-080508-081752
Stevanato, P., Trebbi, D., Panella, L., Richardson, K., Broccanello, C., Pakish, L., et al. (2015). Identification and validation of a SNP marker linked to the gene HsBvm-1 for nematode resistance in sugar beet. Plant Mol. Biol. Rep. 33, 474–479. doi: 10.1007/s11105-014-0763-8
Talukder, Z. I., Tharreau, D., and Price, A. H. (2004). Quantitative trait loci analysis suggests that partial resistance to rice blast is mostly determined by race-specific interactions. New Phytol. 162, 197–209. doi: 10.1111/j.1469-8137.2004.01010.x
Tamisier, L., Rousseau, E., Barraillé, S., Nemouchi, G., Szadkowski, M., Mailleret, L., et al. (2017). Quantitative trait loci in pepper control the effective population size of two RNA viruses at inoculation. J. Gen. Virol. 98, 1923–1931. doi: 10.1099/jgv.0.000835
Villamon, F. G., Spooner, D. M., Orrillo, M., Mihovilovich, E., Perez, W., and Bonierbale, M. (2005). Late blight resistance linkages in a novel cross of the wild potato species Solanum paucissectum (series Piurana). Theor. Appl. Genet. 111, 1201–1214. doi: 10.1007/s00122-005-0053-9
Wang, L. J., Wang, Y. J., Wang, Z., Marcel, T. C., Niks, R. E., and Qi, X. Q. (2010). The phenotypic expression of QTLs for partial resistance to barley leaf rust during plant development. Theor. Appl. Genet. 121, 857–864. doi: 10.1007/s00122-010-1355-0
Wang, Q., Liu, Y. Q., He, J., Zheng, X. M., Hu, J. L., Liu, Y. L., et al. (2014). STV11 encodes a sulphotransferase and confers durable resistance to rice stripe virus. Nat. Commun. 5, 4768. doi: 10.1038/ncomms5768
Wiesner-Hanks, T., and Nelson, R. (2016). “Multiple disease resistance in plants,” in Annual Review of Phytopathology, eds J. E. Leach and S. Lindow (Palo Alto, CA: Annual Reviews), 229–252.
Wilfert, L., and Schmid-Hempel, P. (2008). The genetic architecture of susceptibility to parasites. BMC Evol. Biol. 8:187. doi: 10.1186/1471-2148-8-187
Willocquet, L., Savary, S., and Yuen, J. (2017). Multiscale phenotyping and decision strategies in breeding for resistance. Trends Plant Sci. 22, 420–432. doi: 10.1016/j.tplants.2017.01.009
Yang, Q., He, Y. J., Kabahuma, M., Chaya, T., Kelly, A., Borrego, E., et al. (2017). A gene encoding maize caffeoyl-CoA O-methyltransferase confers quantitative resistance to multiple pathogens. Nat. Genet. 49, 1364–1372. doi: 10.1038/ng.3919
Yogendra, K. N., Kushalappa, A. C., Sarmiento, F., Rodriguez, E., and Mosquera, T. (2015). Metabolomics deciphers quantitative resistance mechanisms in diploid potato clones against late blight. Funct. Plant Biol. 42, 284–298. doi: 10.1071/fp14177
Yu, J. M., and Buckler, E. S. (2006). Genetic association mapping and genome organization of maize. Curr. Opin. Biotechnol. 17, 155–160. doi: 10.1016/j.copbio.2006.02.003
Zhang, X. H., Peng, G., Kutcher, H. R., Balesdent, M. H., Delourme, R., and Fernando, W. G. D. (2016). Breakdown of Rlm3 resistance in the Brassica napus-Leptosphaeria maculans pathosystem in western Canada. Eur. J. Plant Pathol. 145, 659–674. doi: 10.1007/s10658-015-0819-0
Keywords: quantitative trait loci, major R genes, durability, resistance mechanisms, marker-assisted-selection
Citation: Pilet-Nayel M-L, Moury B, Caffier V, Montarry J, Kerlan M-C, Fournet S, Durel C-E and Delourme R (2017) Quantitative Resistance to Plant Pathogens in Pyramiding Strategies for Durable Crop Protection. Front. Plant Sci. 8:1838. doi: 10.3389/fpls.2017.01838
Received: 14 July 2017; Accepted: 10 October 2017;
Published: 27 October 2017.
Edited by:
Thomas Miedaner, University of Hohenheim, GermanyReviewed by:
Wolfgang Friedt, Justus Liebig University Giessen, GermanyHarsh Raman, New South Wales Department of Primary Industries, Australia
Copyright © 2017 Pilet-Nayel, Moury, Caffier, Montarry, Kerlan, Fournet, Durel and Delourme. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Marie-Laure Pilet-Nayel, bWFyaWUtbGF1cmUucGlsZXQtbmF5ZWxAaW5yYS5mcg==