 Oleg I. Yakhin
Oleg I. Yakhin Aleksandr A. Lubyanov
Aleksandr A. Lubyanov Ildus A. Yakhin
Ildus A. Yakhin Patrick H. Brown
Patrick H. Brown- 1Institute of Biochemistry and Genetics, Ufa Scientific Center, Russian Academy of Sciences, Ufa, Russia
- 2R&D Company Eco Priroda, Ulkundy, Russia
- 3Department of Plant Sciences, University of California, Davis, Davis, CA, USA
This review presents a comprehensive and systematic study of the field of plant biostimulants and considers the fundamental and innovative principles underlying this technology. The elucidation of the biological basis of biostimulant function is a prerequisite for the development of science-based biostimulant industry and sound regulations governing these compounds. The task of defining the biological basis of biostimulants as a class of compounds, however, is made more complex by the diverse sources of biostimulants present in the market, which include bacteria, fungi, seaweeds, higher plants, animals and humate-containing raw materials, and the wide diversity of industrial processes utilized in their preparation. To distinguish biostimulants from the existing legislative product categories we propose the following definition of a biostimulant as “a formulated product of biological origin that improves plant productivity as a consequence of the novel or emergent properties of the complex of constituents, and not as a sole consequence of the presence of known essential plant nutrients, plant growth regulators, or plant protective compounds.” The definition provided here is important as it emphasizes the principle that biological function can be positively modulated through application of molecules, or mixtures of molecules, for which an explicit mode of action has not been defined. Given the difficulty in determining a “mode of action” for a biostimulant, and recognizing the need for the market in biostimulants to attain legitimacy, we suggest that the focus of biostimulant research and validation should be upon proof of efficacy and safety and the determination of a broad mechanism of action, without a requirement for the determination of a specific mode of action. While there is a clear commercial imperative to rationalize biostimulants as a discrete class of products, there is also a compelling biological case for the science-based development of, and experimentation with biostimulants in the expectation that this may lead to the identification of novel biological molecules and phenomenon, pathways and processes, that would not have been discovered if the category of biostimulants did not exist, or was not considered legitimate.
Introduction
The regulation of plant growth and the development and alleviation of the negative effects of environmental stresses during ontogenesis, are important factors determining the productivity of cultivated plants. While it is well recognized that biotic and abiotic stress prevents essentially all crop systems from achieving their yield potential, current understanding of the mechanisms involved, and the strategies to mitigate these effects are limited. Abiotic stresses may be prevented by optimizing plant growth conditions and through provision of water and nutrients and plant growth regulators (PGRs—auxins, cytokinins, gibberellins, strigolactones, brassinosteroids). In addition to these traditional approaches, biostimulants are increasingly being integrated into production systems with the goal of modifying physiological processes in plants to optimize productivity. Plant biostimulants based on natural materials have received considerable attention by both the scientific community and commercial enterprises especially in the last two and a half decades (Crouch and van Staden, 1993a; Herve, 1994; Zhang and Schmidt, 1999; Maini, 2006; Khan et al., 2009; Apone et al., 2010; Craigie, 2011; Sharma et al., 2014; Brown and Saa, 2015; Du Jardin, 2015; Yakhin et al., 2016a). Biostimulants offer a potentially novel approach for the regulation/modification of physiological processes in plants to stimulate growth, to mitigate stress-induced limitations, and to increase yield. In the following review, we do not attempt to discern if the effects of biostimulants on plant productivity is a direct response of plants or soils to the biostimulant application or an indirect response of the biostimulant on the soil and plant microbiome with subsequent effects on plant productivity. Ultimately discerning if biostimulant effects are direct or microbially mediated will be critical to the development of this technology. The general goals of the current review are to provide a comprehensive analysis of the current situation in the field of biostimulants and to develop a science-based theoretical foundation for the conceptualization, classification, and practical application of these materials. A focus of this review is to understand and define the appropriate place of biostimulants among other agricultural products such as plant protection compounds and fertilizers, and to consider the unique attributes of complex, multi-component biostimulants. The structure of the review is based on the consideration of biostimulants in terms of their action on different regulatory and functional systems of plants (signaling, metabolism, uptake, and transport mechanisms, etc.) using both conceptual and methodological approaches. The overarching objective of the work is to highlight innovative concepts and to establish a scientific framework for future development of biostimulant science.
General Concepts and Methodology
To understand the development of biostimulant science, several seminal publications warrant discussion. To our knowledge, the first discussion of “biogenic stimulant” theory can be attributed to Prof. V.P. Filatov and was started in 1933 in the USSR (Filatov, 1944, 1951a,b; Gordon, 1947; Sukhoverkhov, 1967). Filatov proposed that biological materials derived from various organisms, including plants, that have been exposed to stressors could affect metabolic and energetic processes in humans, animals, and plants (Table 1). Blagoveshchensky (1945, 1955, 1956) further developed these ideas with specific reference to their application for plants, considering biogenic stimulants as “organic acids with stimulating effects due to their dibasic properties which can enhance the enzymatic activity in plants.” Filatov's concept (1951b), was, however, not limited to these compounds alone (Filatov, 1951b). Herve's (1994) pioneering review provides the first real conceptual approach to biostimulants. Herve suggests the development of novel “bio-rational products” should proceed on the basis of a systemic approach founded in chemical synthesis, biochemistry, and biotechnology as applied to real plant physiological, agricultural, and ecological constraints. He suggests these products should function at low doses, be ecologically benign and have reproducible benefits in agricultural plant cultivation. Zhang and Schmidt (1999) emphasized the need for comprehensive and empirical analysis of these products with particular emphasis on hormonal and antioxidant systems as the basis for many important benefits of biostimulants. They discuss the concept of biostimulants as “pre-stress conditioners,” their effects being manifested in improved photosynthetic efficiency, reduction of spread and intensity of some diseases and in better yields. Basak (2008) initiated the systematic discussion on biostimulants and created the conceptual preconditions for the formation of present biostimulant science while Du Jardin (2012, 2015) provided the first in-depth analysis of plant biostimulant science with an emphasis on biostimulant systematization and categorization on the basis of biochemical and physiological function and mode of action and origin. Du Jardin's (2015) analysis and categorization was influential in informing the development of subsequent legislation and regulation in the European Union.

Table 1. Terminology in the biostimulant field: Evolution and diversity of concepts*.
The study and development of biostimulants has been approached utilizing a wide range of methodological approaches including chemical and non-chemical characterization of composition (Crouch and van Staden, 1993b; Yakhin et al., 2005; Parrado et al., 2008; Sharma et al., 2012a,b; Ertani et al., 2013a,b; Aremu et al., 2015a,b), plant growth and yield studies (Khan et al., 2009; Kunicki et al., 2010; Parađiković et al., 2011; Zodape et al., 2011; Yakhin et al., 2012, 2016b; Chbani et al., 2013; Kurepin et al., 2014; Colla et al., 2015; Saa et al., 2015; Tandon and Dubey, 2015; Tian et al., 2015), application of the so-called -omics strategies with variations, including microarray and physiological analysis (Jannin et al., 2012, 2013), transcriptome (Wilson et al., 2015; Goñi et al., 2016), genomic (Santaniello et al., 2013), phenomic and molecular (Petrozza et al., 2014), proteomic (Martínez-Esteso et al., 2016), chemical and metabolomic (Ertani et al., 2014). Ultimately, the integrative synthesis of results from multiple methodologies, particularly when integrated with the most relevant—omic technology, “agronomics,” will be required if the science and legitimacy of plant biostimulants is to advance.
Several significant scientific meetings in the field of biostimulants have been held over the past ten years and have contributed greatly to our understanding of conceptual and methodological development of the biostimulant theory: “Biostimolanti in agrocoltura” (Italy, 2006), “Biostimulators in Modern Agriculture” (Poland, 2008), “Biostimulants and Plant Growth” (Belgium, 2014), among others. Of particular significance were the first (France, 2012) and the second (Italy, 2015) World Congresses on the “Use of Biostimulants in Agriculture” which were valuable in highlighting the development of novel concepts and methodology as applied to biostimulants. While many of the following papers are not published in a peer-reviewed format, they do represent important advances in this field. Dumas et al. (2012), for example, proposed a multi-part approach to study biostimulants based on large-scale genomic approaches and high-throughput screening tests with genetically-modified reporter plants. Others suggested that biostimulant mode of action can be best determined using molecular microarray analysis to identify gene changes in transcript levels (Gates et al., 2012). This approach has the potential to reveal biostimulant activated signaling pathways involved in the stimulation of plant response. Microarray analysis is not, however, adequate and must be supplemented with carefully conducted field testing or high throughput plant phenotyping (Summerer et al., 2013). The complexity of known biostimulant response, the dependency of crop environment and the diversity of biostimulant products demands the application of novel statistical approaches not commonly used in agronomic research (Sleighter et al., 2015). The principle espoused by Sleighter et al. (2015) is based on the identification of a subset of molecular markers that represent the active ingredients in complex biostimulants and then to correlate these markers with observations of plant response. Chemical genomics that utilizes small molecules to perturb target protein function is a useful strategy for biostimulant discovery as it overcomes constraints imposed by traditional molecular approaches that often fail due to gene redundancy and loss-of-function lethality. Botta et al. (2015) proposed probing the function of biostimulants using an enantiomeric analysis of active compounds in the biostimulant coupled with a proteomic profiling approach. In contrast, Conan et al. (2015) proposed identification of the bioactive compounds responsible for the plant growth response by means of a metabolomic profiling of biostimulant products and analysis of their physiological effects through transcriptomic and metabolomic strategies. Such methodology allows the determination of metabolite pathways affected by biostimulants as well as providing insight into gene regulation. To integrate the diversity of methodologies available Santaniello et al. (2015) emphasizes the need to use bioinformatics strategies to analyse similarities and differences in procedures of ingredient extraction and biostimulant formulation in terms of molecular plant responses. This integrative concept can be used to derive new technologies and novel biostimulant products through the identification of new target genes, enzymes and metabolites.
While the development of robust, multi-faceted approach to the analysis of biostimulant composition and function will greatly aid in the development of this field, all advances must ultimately be interpreted in the context of plant response. The complexity of plant response to the environment is daunting and was elegantly highlighted by Krouk (2015) who demonstrated that root response to nitrogen in the environment is mediated by combinations of signaling molecules and nitrogen sources in a manner that cannot be predicted by exposure to single compounds provided individually (Krouk, 2016; Krouk et al., 2009, 2010, 2011). Inevitably, as our understanding of the molecular networks that control plant growth improves our ability to predict plant response to biostimulants under specific environmental conditions, will improve. Only through a combination of methodologies will progress in biostimulant research be possible.
Terminology and Definitions
The development of plant biostimulant science, as well as the principles governing its legislation in the context of the existing legal frameworks of plant protection products and fertilizers, requires the development of a clear definition of term “biostimulant.” Currently, the term “biostimulant” is poorly defined and includes many products that have variously been described as biogenic stimulants, metabolic enhancers, plant strengtheners, positive plant growth regulators, elicitors, allelopathic preparation, plant conditioners, phytostimulators, biofertilisers, or biofertiliser/biostimulant (Table 1). One area of significant challenge is evoked in the question “are biostimulants PGRs?” Historically, biostimulants have been considered as a subgroup of growth regulators (Herve, 1994), as plant growth regulators (Huang, 2007), and as subgroup of bioregulators (Basak, 2008). “From a legal point of view, biostimulants can contain traces of natural plant hormones, but their biological action should not be ascribed to them, otherwise they should be registered as plant growth regulators” (Bulgari et al., 2015). Likewise, biostimulants cannot by definition be pesticides or fertilizers (Russo and Berlyn, 1991; Karnok, 2000; Hamza and Suggars, 2001; Banks and Percival, 2012; Du Jardin, 2012; Torre et al., 2013, 2016).
A concise and biologically meaningful definition of biostimulants has eluded researchers and regulators for many years. Table 1 presents a chronological evolution of concept of the term biostimulant. While several of biostimulant definitions presented are useful in their breadth, many of them have significant limitations and are overly generic, while several do not exclude possible effects of nutrients contained within any putative biostimulant product. In practice, biostimulants may deliberately include nutrients for regulatory approval as fertilizers and on occasions the included nutrients or hormones may be responsible for the perceived agronomic benefit. Given the state of public mistrust of many “biostimulant” products, it is necessary to provide a definition of biostimulants that explicitly denies the use of this term for products that do not have biological efficacy or have efficacy only by virtue of the inclusion of known plant hormones or nutrients.
While the adoption of a definition of biostimulants for regulatory purposes is important, any definition of biostimulant should also be based on scientific principles. Several concepts have been proposed to define plant biostimulants. Basak (2008), proposed that biostimulants could be classified depending on the mode of action and the origin of the active ingredient while Bulgari et al. (2015), proposed that “biostimulants should be classified on the basis of their action in the plants or, on the physiological plant responses rather than on their composition.” Du Jardin (2015), however, has emphasized the importance of the final impact on plant productivity when he suggests that “any definition of biostimulants should focus on the agricultural functions of biostimulants, not on the nature of their constituents nor on their modes of actions.” The term “plant productivity” is used here to describe any improvement in plant yield or quality or increased efficiency of production. These concepts reflect important differences in approaches to providing a definition of biostimulants as a discrete category of agricultural products. Thus, biostimulants could be defined by their demonstrated mode of action and origin, or solely by their demonstrated beneficial impact on plant productivity. The challenges in developing a definition are also complicated by the multi-component and largely undefined composition of many biostimulant products and the possibility that the activity of a biostimulant may not be explained by the presence of any individual constituent, but is a result of the interaction of many constituents in the product.
On this basis two approaches to the definition of complex biostimulants emerge. The first is based on the possibility that the biostimulant contains within it, previously unrecognized molecules that are the sole and discrete cause of the observed improvement in plant productivity. This concept emphasizes both the need for clear demonstration of plant productivity benefits and the unknown nature of the mode of action. Thus, a biostimulant could be defined as “a formulated product that improves plant productivity by a mechanism of action that is not the sole consequence of the presence of known essential plant nutrients, plant hormones, plant growth regulators or plant protective compounds.” By this definition, once the primary biological mechanism of biostimulant function has been identified it should henceforth, be subject to classification on the basis of that functional component.
The majority of biostimulants in use today are complex mixtures of chemicals derived from a biological process or extraction of biological materials. The complexity of these mixtures is often considered to be essential to the performance of the biostimulant, and biostimulants may have properties of the whole, that cannot be fully elucidated by knowing the characteristics of the separate components or their combinations. This theory of complexity or “emergence” was described by Mayr (1982), who argued that in many biological systems “the properties of the whole cannot be fully elucidated by knowing the characteristics of the separate components or their combinations.” “The term emergence describes the onset of novel properties that arise when a certain level of structural complexity is formed from components of lower complexity. In the last few decades, emergence has been discussed in a number of different research fields, such as cybernetics, theory of complexity, artificial intelligence, non-linear dynamics, information theory, and social systems organization” (Luisi, 2002). “Emergence” and “emergent properties” are thus closely related with the notion of the “systems biology” (Luisi, 2002; Johnson, 2006; Korosov, 2012; Lüttge, 2012; Bertolli et al., 2014). Emergence was described by Johnson (2006) as “unexpected behaviors that stem from interaction between the components of an application and their environment,” “there is, however, considerable disagreement about the nature of ‘emergent properties.’ Some include almost any unexpected properties exhibited by a complex system. Others refer to emergent properties when an application exhibits behaviors that cannot be identified through functional decomposition. In other words, the system is more than the sum of its component parts” (Johnson, 2006).
Thus, a biostimulant could also be defined as “a formulated product of biological origin that improves plant productivity as a consequence of the emergent properties of its constituents.”
To our knowledge, however, there have been no clear demonstrations that any biostimulant exhibits truly emergent properties. This is not however a unique challenge and all “biological systems are extremely complex and have emergent properties that cannot explained, or even predicted, by studying their individual parts” (Van Regenmortel, 2004). Emergent properties have been demonstrated in the networks of biological signaling pathways (Bhalla and Iyengar, 1999); in system-level study of traditional Chinese medicine (Chen et al., 2014), and in microbial communities (Wintermute and Silver, 2010; Chiu et al., 2014). To adequately explain the biological complexity present in plants and their interactions with the environment, Lüttge (2012) and Bertolli et al. (2014) emphasize that classic reductionist biology/chemistry is indeed insufficient.
While the two theoretical definitions provided in this section share a requirement that the mode of action is unknown, they differ in the core assumption that biostimulant function is a consequence of the discrete components in the biostimulant or as a consequence of the “emergent” properties of the biostimulant as a whole. Each of these definitions is also incomplete in that it is certainly possible that a biostimulant may contain several molecules that act synergistically while not being truly “emergent,” and it is indeed possible and indeed likely, that even if a biostimulant is demonstrated to have emergent properties, that not all components of that biostimulant are required for that property to be expressed.
We propose, therefore, a definition of a biostimulant that integrates these two concepts. Thus, a biostimulant is defined here as:
“a formulated product of biological origin that improves plant productivity as a consequence of the novel, or emergent properties of the complex of constituents, and not as a sole consequence of the presence of known essential plant nutrients, plant growth regulators, or plant protective compounds.”
Consistent with this definition, the ultimate identification of a novel molecule within a biostimulant that is found to be wholly responsible for the biological function of that biostimulant, would necessitate the classification of the biostimulant according to the discovered function.
Classification
A review of the history of biostimulants and related products provides insight into the diversity of these products and the development of this field of study. The evolution of biostimulant classifications as described by various authors is presented in the Table 2. To the best of our knowledge, one of the first attempts to categorize biostimulants was provided by Filatov (1951b) when 4 groupings of biogenic stimulants were suggested. Karnok (2000) compiled a list of 59 materials presenting in 15 biostimulants; Ikrina and Kolbin (2004) systematized patent literature and specified 9 categories of natural raw materials used to derive biostimulants; Basak (2008) suggested that biostimulants could be grouped on the basis of single or multicomponent formulations and classified on the origin of the active ingredient, and the mode of action of the active ingredient. Du Jardin (2012) developed a scientific rationale of classification considering 8 categories of biostimulants and subsequently reduced this list to 7 categories (Du Jardin, 2015). Du Jardin (2012) was explicit in his exclusion of microorganisms from his categorization primarily to avoid conflict with existing categorization of microorganisms as biopesticides and sources of plant hormones. Later Bulgari et al. (2015) proposed a biostimulant classification on the basis of their mode of action rather than on their composition.
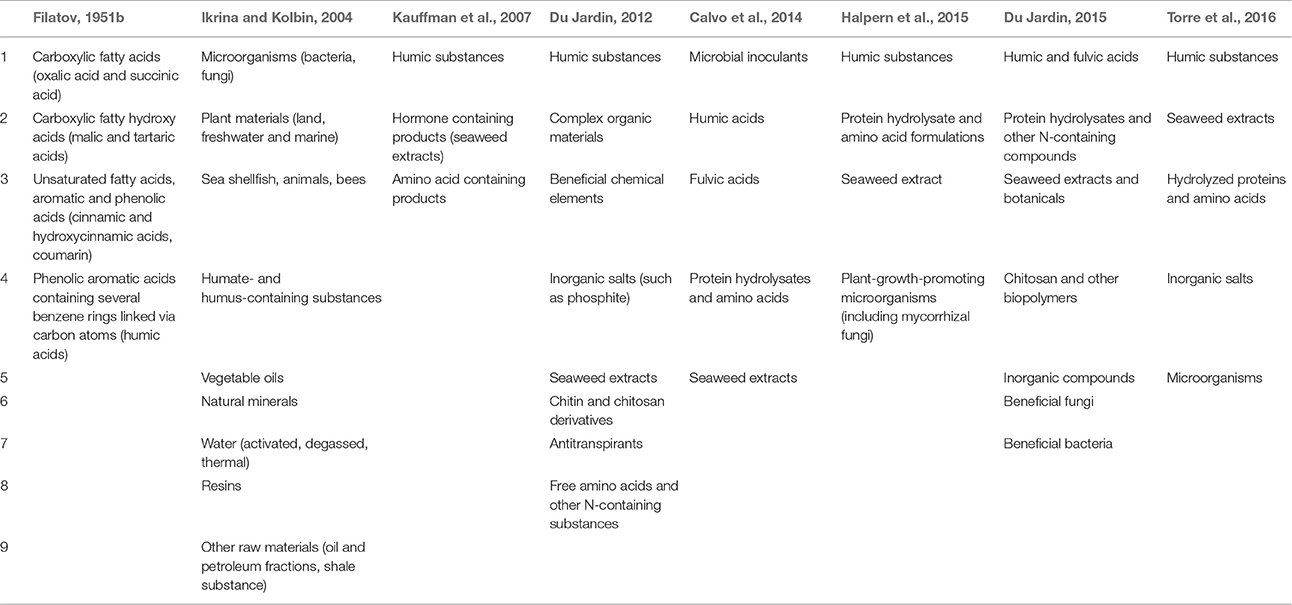
Table 2. Proposed Biostimulant Categories.
Many biostimulant products have been classified into completely divergent groups and categories of function, use, and type of activity (Tables 3, 4). For example, humate-based products are often described as soil health amendments while plant growth promoting rhizobacteria (PGPRs) could be categorized as biofertilizers, phytostimulators, and biopesticides (Martínez-Viveros et al., 2010; Bhattacharyya and Jha, 2012). Du Jardin (2015) has proposed that biofertilisers are a subcategory of biostimulants. Seaweed extracts have been considered as biofertilizers (Zodape, 2001) and microorganisms have also been described as biofertilizers (Vessey, 2003; Fuentes-Ramirez and Caballero-Mellado, 2006; Roy et al., 2006; Malusá et al., 2012; Bhardwaj et al., 2014; Malusá and Vassilev, 2014). Some inorganic elements or small molecules that are not known to be essential may also be classified as biostimulants if evidence of plant growth promotion is available (Michalski, 2008; Kleiber and Markiewicz, 2013; Radkowski and Radkowska, 2013). Thao and Yamakawa (2009), for example, consider phosphites to be biostimulants since plant response to phosphites frequently cannot be explained as a consequence of the known anti-fungal function of these molecules. While the categorization of biostimulants by their origin does not, a priori, provide information on their mode of action this categorization may still be a useful tool to aid in the process of discovery and facilitate comparison between similar products.
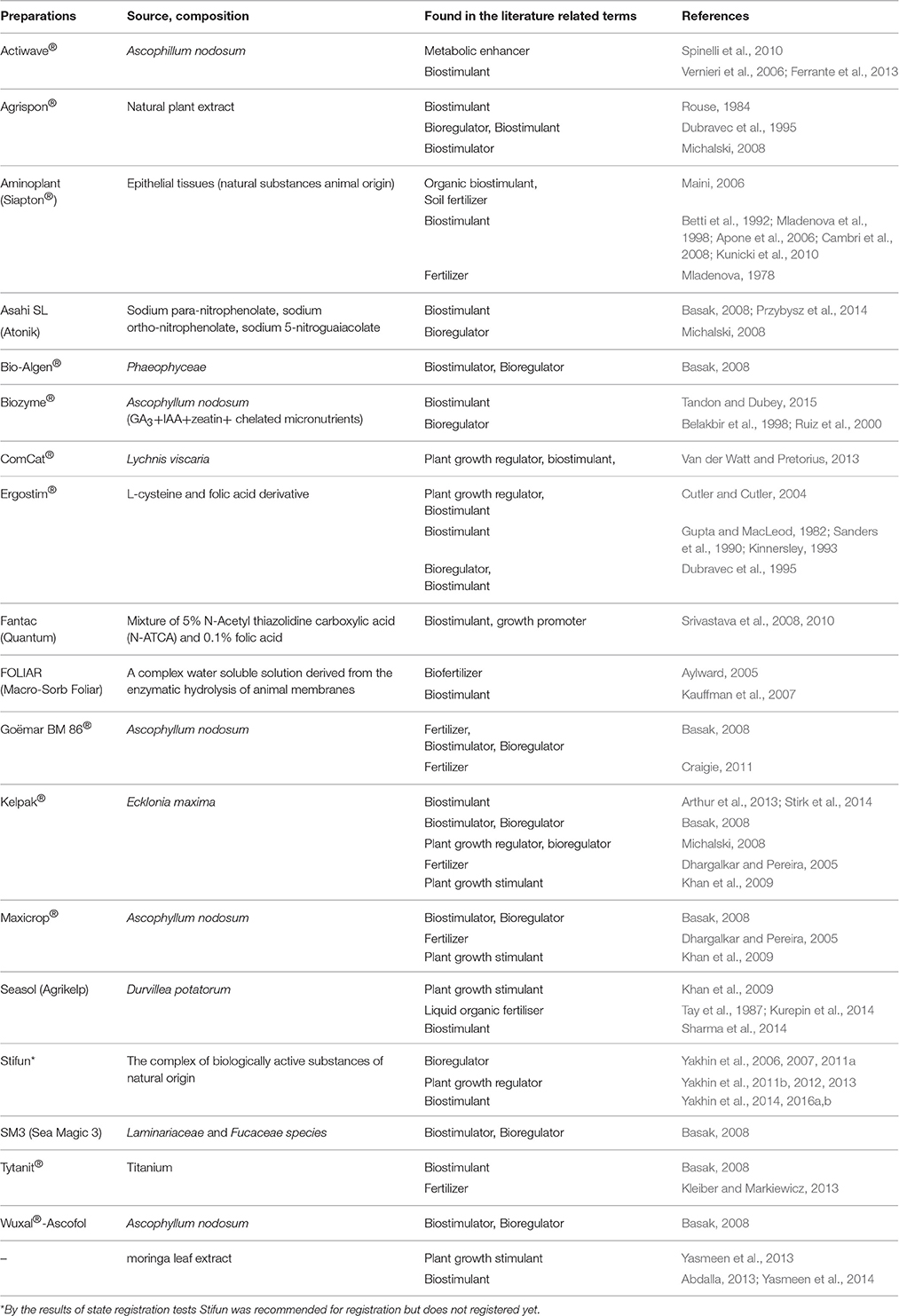
Table 3. Examples of different terminology used for commercial biostimulants.

Table 4. Biostimulants: sources, production, compositions, and activities.
Legislation and Legal Framework
Registration of products used in agriculture is crucial to ensure their practical, safe and legitimate application. In the absence of a sound definition of biostimulants as a discrete group of products (Basak, 2008), the registration procedure and subsequent classification regime is untenable and this inevitably creates a barrier to trade and development. Various countries, states, and administrative regions have developed different categories for registration of potential biostimulants including terminology such as plant conditioners, “other fertilizers,” supplements, soil improvers, plant strengtheners, fitofortificants, etc. (Basak, 2008; Torre et al., 2013; Traon et al., 2014). In many jurisdictions regulatory practices require an itemized description and identification of substances in all commercial product classifications while in others the registration of non-fully identified substances is allowed if those products are considered of complex composition. There is even a proposal for complex biostimulants to not specify the chemical name (IUPAC) and note as “None” with the definition that “this product is a complex mixture of chemical substances” (Traon et al., 2014). If we accept the concept that a biostimulant is a product of clear benefit but unknown mode of action, then it can only be regulated by its safety and proof of efficacy. For example, in pharmacology it has been suggested that “the demand to demonstrate the mode of action of each single component in a phytopharmaceutical may not be obligatory any more” (Ulrich-Merzenich et al., 2009).
The complex multicomponent nature of many biostimulants clearly complicates discovery of their modes/mechanisms of action, production, registration and use. What is clearly needed however, is a regulatory mechanism to ensure that the products are “generally recognized as safe,” have “a positive benefit on crop productivity” and are discrete from exisiting categories of products. The task of identifying function and agronomic utility can then be pursued independently and will be driven by the marketplace imperative for product quality and consistency. Coordinating national legislation within this framework will become critical for the optimization of biostimulants and trade between different countries. The possible place of biostimulants in the regulatory system of pesticides and agrochemicals is illustrated in Figure 1.
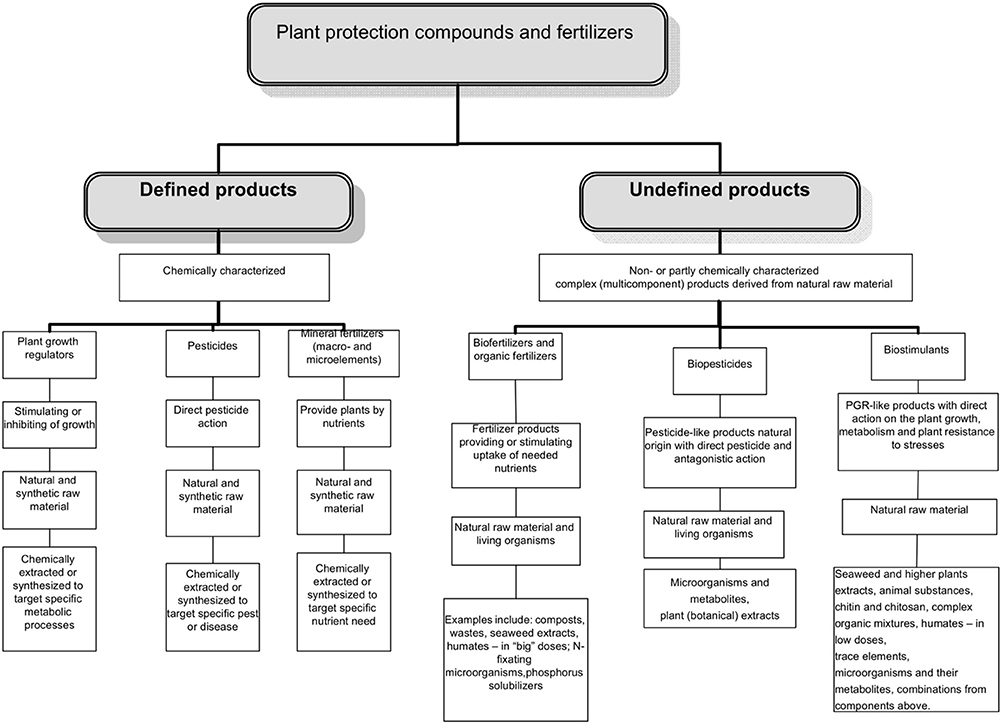
Figure 1. The distribution of various categories of products among the plant protection products and fertilizers.
Primary Sources of Raw Materials
We have conducted an exhaustive analysis of the literature and categorized the majority of the reported biostimulants by origin (Table 4). Microorganisms are widely used for the production of biostimulants and may be derived from bacteria, yeasts, and fungi. These preparations may include living and/or non-living microorganisms and their metabolites. The concept of microorganism-based preparations as biostimulants is described by Xavier and Boyetchko (2002), Sofo et al. (2014), Colla et al. (2015), Matyjaszczyk (2015), and Ravensberg (2015). Different species of algae, mostly seaweeds, are also commonly used for producing biostimulants. Seaweed-based preparations as biostimulants are described in reviews by Crouch and van Staden (1993a), Khan et al. (2009), Craigie (2011), Sharma et al. (2014); and experimental papers by Goatley and Schmidt (1991), Jannin et al. (2013), Billard et al. (2014), Aremu et al. (2015b). Raw materials for biostimulants are also commonly based on higher plant parts including seeds, leaves, and roots and exudates from families Amaryllidaceae, Brassicacae, Ericaceae, Fabaceae, Fagaceae, Moringaceae, Plantaginaceae, Poaceae, Rosaceae, Solanaceae, Theaceae, Vitaceae, among others (Naumov et al., 1993; Yakhin et al., 1998, 2011a, 2012, 2014; Pretorius, 2007, 2013; Parrado et al., 2008; Apone et al., 2010; Ertani et al., 2011a, 2013a, 2014; Colla et al., 2014; Yasmeen et al., 2014; Lucini et al., 2015; Ugolini et al., 2015). Biostimulants may also be based on protein hydrolysates and amino acids of animal origin including wastes and by-products (Mladenova et al., 1998; Maini, 2006; Kolomazník et al., 2012; Ertani et al., 2013b; Rodríguez-Morgado et al., 2014), and insect derived chitin and chitosan derivatives (Sharp, 2013). Humate-based raw materials are widely used to derive biostimulants and have been reviewed by Sanders et al. (1990), Kelting et al. (1998), Ertani et al. (2011b), and Jannin et al. (2012). A final category of biostimulants includes those derived from extracts of food waste or industrial waste streams, composts and compost extracts, manures, vermicompost, aquaculture residues and waste streams, and sewage treatments among others. Because of the diversity of source materials and extraction technologies, the mode of action of these products is not easily determined.
Technologies of Production
The technologies used in the production and preparation of biostimulants are highly diverse and include cultivation, extraction, fermentation, processing and purification, hydrolysis, and high-pressure cell rupture treatment (Table 4). In some instances, a biostimulant product may also contain mixes of components derived from different sources and production methods. Frequently the rationale for utilizing extracts rather than raw biomass is a consequence of the need for a standardized manufacturing process to produce a uniform commercial product (Michalak and Chojnacka, 2014). For many products, the production processes are driven by process and marketing demands and are not the result of a targeted strategy to optimize the biological efficacy of the commercial product. While the ultimate composition and possible function of commercial biostimulant products may be partially determined by both the source of raw material and the process by which it is prepared (Traon et al., 2014), there may be manufacturing processes and product treatments utilized that result in compounds that are not present in the initial (primary raw) material. An example of this is the multitude of commercial seaweed extracts, often derived from the same species, that are rarely equivalent (Craigie, 2011). Commercial biostimulant manufactured from similar sources are usually marketed as equivalent products, but may differ considerably in composition and thereby in efficiency (Lötze and Hoffman, 2016). Many manufacturers do not reveal the technology of biostimulant production since that is a commercial secret (Traon et al., 2014).
Bioactive Components and Methods of Quality Control
A diversity of substances contained in raw materials is used for the production of biostimulants. Whereas, primary metabolites are contained in most preparations de facto, the presence of secondary metabolites is more specific and depends to a large extent on the raw material used (species, tissue, growing conditions). Primary metabolites include amino acids, sugars, nucleotides, and lipids (Aharoni and Galili, 2011). Secondary metabolites are formed from different primary metabolic pathways, including glycolysis, the tricarboxylic acid cycle (TCA), aliphatic amino acids (AA), the pentose-phosphate and shikimate pathways which are primarily the source of aromatic AA and phenolic compounds (PC), terpenoids/isoprenoids, nitrogen-containing compounds (alkaloids), sulfur-containing compounds (glucosinolates); (Aharoni and Galili, 2011). Frequently, biostimulants are shown to have a multicomponent composition and may include plant hormones or hormone-like substances, amino acids, betaines, peptides, proteins, sugars (carbohydrates, oligo-, and polysaccharides), aminopolysaccharides, lipids, vitamins, nucleotides or nucleosides, humic substances, beneficial elements, phenolic compounds, furostanol glycosides, sterols, etc. (Table 4). While many articles have attempted to describe the composition of complex biostimulants, these descriptions are frequently incomplete since the vast majority of biological molecules that would be present in crude extracts of complex origin, have not yet been characterized and the mere presence of a specific compound does not a priori demonstrate that compound is functional. The composition of most biologically derived biostimulant feed stock will also vary with the season of production, species, physiological state of the source organism and growth conditions. Indeed, there is an implication in the marketing of many biostimulants that stress conditions experienced by the plant or microbe utilized to produce the biostimulant, results in the production of stress metabolites and amino acids with consequent beneficial effects on plant response. In the absence of knowledge of the functional component of a biostimulant, changes in composition of a biostimulant over time and between batches and commercial sources cannot be interpreted. In the most rigorously prepared biostimulants from leading companies, high-throughput analytical methods have been employed to ensure consistent product quality (Sharma et al., 2012b). Methods such as chromatography, mass spectrometry, NMR spectroscopy, elemental analysis, ELISA, spectrophotometry, etc. are typically used for this purpose (Table 4). The complexity of this challenge is illustrated in the analysis of a four-year algae composition sequence using a profile or fingerprint technique employing NMR (Craigie et al., 2009).
Function and Effects on Whole Plants
Biostimulants have been used at all stages of agricultural production including as seed treatments, as foliar sprays during growth and on harvested products. The mode/mechanisms action of “biostimulants” is equally diverse and may include the activation of nitrogen metabolism or phosphorus release from soils, generic stimulation of soil microbial activity or stimulation of root growth and enhanced plant establishment. Various biostimulants have been reported to stimulate plant growth by increasing plant metabolism, stimulating germination, enhancing photosynthesis, and increasing the absorption of nutrients from the soil thereby increasing plant productivity (Table 4). Biostimulants may also mitigate the negative effects of abiotic stress factors on plants and marked effects of biostimulants on the control of drought, heat, salinity, chilling, frost, oxidative, mechanical, and chemical stress, have been observed (Table 4). Alleviation of abiotic stress is perhaps the most frequently cited benefit of biostimulant formulations. The following text describes the primary modes/mechanisms of action that have been demonstrated or claimed for biostimulants in the primary scientific literature.
Modes of Action/Mechanisms of Action
Understanding the modes of action of an agricultural chemical has been a fundamental requirement for effective marketing and frequently a regulatory requirement for manufactured products used in agriculture. Mode of action is used here to mean “a specific effect on a discrete biochemical or regulatory process,” thus the “mode of action” of Glyphosate is to inhibit the activity of the enzyme enolpyruvylshikimate-3-phosphate synthase (EPSPS). Biostimulants frequently do not meet this standard of specificity and indeed there are few biostimulant products for which a specific biochemical target site and known mode of action has been identified. For a small subset of biostimulants, however, a demonstrated impact on general biochemical or molecular pathways or physiological processes, termed here as a “mechanism of action,” has been identified even though the explicit “mode of action” may not be known. An example of a “mechanism of action” would be a stimulation of photosynthesis or the down regulation of a plant stress signaling pathway without an understanding of the explicit biochemical or molecular “mode of action.”
For many biostimulant products, however, neither a specified mode of action, nor a known mechanism of action, has been identified. The presence of some spurious products in the marketplace compromises the market for all players resulting in the assumption by many, that biostimulants as a whole, are “snake oils” (Basak, 2008), a pejorative term implying the product is of no value. Multicomponent biostimulants are particularly difficult to reconcile since they may have constituents for which the mode of action is known and components of no known functional benefit. Furthermore, multicomponent biostimulants will frequently contain measureable but biologically irrelevant concentrations of known essential elements, amino acids, and plant hormones etc., for which the mode of action is known but the concentrations are irrelevant when used at recommended rates. Thus, for many of the multicomponent biostimulant in the marketplace today, we propose that a demonstration of a clear “mechanism of action” is a more rationale and attainable regulatory goal than requiring an unequivocal demonstration of the “mode of action.”
Insight into the use of the terms “mode and mechanism” of action can be drawn from the pesticide science and pesticide development. In pesticide science, the “mechanism of action” describes the integral of all the biochemical events following application while the “mode of action” characterizes the main features of a bioactive molecule and its specific biochemical action leading to its effect in treated plants (Aliferis and Jabaji, 2011). In reference to plant bioregulators, Halmann (1990) suggests that ideally an understanding of the mode of action of plant bioregulators on the molecular level requires the identification of the receptor site for each regulator, as well as the elucidation of the subsequent reactions. In reality this standard is often not met in biopesticide or biostimulant products where the identification of the molecular targets of all bioavailable (and frequently uncharacterized) compounds within a given extract cannot be easily achieved. The identification of the target binding sites of the natural biomolecules has, however, proven to be helpful in the design of new insecticidal molecules with novel modes of action (Rattan, 2010).
At the present time, given the difficulty in determining a “mode of action” for a complex multicomponent product such as a biostimulant, and recognizing the need for the market in biostimulants to attain legitimacy, we suggest that the focus of biostimulant research and validation should be upon determining the mechanism of action, without a requirement for the determination of a mode of action. This is the standard of practice for many pharmacological products. With the development of advanced analytical equipment, bioinformatics, systems biology and other fundamentally new methodologies a more complete understanding of the mechanisms and even possible modes of action of these materials may be achieved in the future. While this proposal suggests that the development and marketing of a biostimulant may not require a demonstration of the mode of action, it is still in the interest of the manufacturers of these products to pursue an understanding of the mode of action so that the product can be improved and the use can be optimized for various environments and cropping systems.
The mechanisms of action of all but a few biostimulants remain largely unknown (Rayorath et al., 2008; Khan et al., 2009; Rathore et al., 2009). This is primarily due to the heterogeneous nature of raw materials used for production and the complex mixtures of components contained in biostimulant products which makes it almost impossible to identify exactly the component(s) responsible for biological activity and to determine the involved mode(s) of action (Parađiković et al., 2011). Therefore, focus should be upon the identification of the “mechanisms of action” of biostimulants as indicated by general positive impacts on plant productivity through enhancement in processes such as photosynthesis, senescence, modulation of phytohormones, uptake of nutrients and water, and activation of genes responsible for resistance to abiotic stresses and altered plant architecture and phenology (Khan et al., 2009; Sharma et al., 2012b). An example of this process is the advances in use of protein-based biostimulants for which recent studies have identified the target metabolic pathways and some of the mechanisms through which they exert their effects on plants (Nardi et al., 2016).
To further our understanding of modes/mechanisms of biostimulant action we have systematized the stages of biostimulants action on plants after their application: (1) penetration into tissues, translocation and transformation in plants, (2) gene expression, plant signaling and the regulation of hormonal status, (3) metabolic processes and integrated whole plant effects.
Penetration Into Tissues, Translocation, Transformation in Plants
The penetration of amino acids and peptide based biostimulants into plant tissues has been investigated using radiolabeled amino acids (Maini, 2006) and mathematical modeling (Kolomazník et al., 2012; Pecha et al., 2012). The components of a biostimulant preparation of animal origin, labeled with 14C proline and glycine, were shown to penetrate rapidly into treated leaves and where subsequently distributed to other leaves (Maini, 2006). The mathematical model based on the “mechanism of diffusion” allows the estimation of the time required for the absorption of a minimal amount of the active component of a biostimulant. Furthermore, it describes the process of its transport from the moment of penetration into the leaf until the arrival at more distant tissues (Kolomazník et al., 2012; Pecha et al., 2012). The penetration of protein hydrolysates into a plant tissue occurs via diffusion of protein molecules through membrane pores (Kolomazník et al., 2012) and is energy-dependent (Parrado et al., 2008). Biostimulants must have a good solubility in water or other suitable solvents. This is a precondition for most types of application and for sufficient penetration of active ingredients into internal structures of treated plants. Surfactants and other additives may be required to overcome solubility and uptake limitations including lipophilicity and molecular size of active components (Kolomazník et al., 2012; Pecha et al., 2012).
Gene Expression, Signaling, and Hormone Interactions
Ultimately a full understanding of the biological activity of complex biostimulant preparations will require a detailed understanding of the mechanism of action and effects on plant productivity and the identification of the biologically active molecules and their molecular mode of action (Henda and Bordenave-Juchereau, 2014). A wide array of molecular methods has been used to attempt to discern the active compounds found in biostimulants including microarrays, metabolomics, proteomic, and transcriptomics methods. These technologies have been applied to biostimulants to probe changes in gene expression following the application of biostimulants (Jannin et al., 2012, 2013; Santaniello et al., 2013). Further research on the effects of complex biostimulants and their components on the complete genome/transcriptome of plants will be required to understand the mechanisms of action involved in growth responses and stress mitigation (Khan et al., 2009). The search for the mode of action of biostimulants is complicated by the observation that many biostimulants have been shown to induce genes and benefit productivity only when plants are challenged by abiotic and biotic stress. Experimental methods must therefore be developed to produce relevant and reproducible stress conditions so that the application of any molecular tool to probe gene function produces results that are relevant to the purported effects on plant productivity.
The role of signaling molecules in plant response to environmental cues has been an area of active research in plant biology. The process of signal transmission involves the synthesis of signaling molecules (ligands), their translocation, their binding to receptors, the resulting cellular responses, and, finally, the degradation of the signaling molecules (Zhao et al., 2005; Wang and Irving, 2011). When the signaling molecule binds to its receptor, the initial cellular response is the activation of secondary messengers, or intracellular signaling mediators, which cause a further series of cellular responses. Among the substances that may act as secondary messengers are: lipids, sugars, ions, nucleotides, gases, Ca2+, cAMP, cGMP, cyclic ADP-ribose, small GTPase, 1,2-diacylglycerol, inositol-1,4,5-triphosphate, nitric oxide, phosphoinosides, and others (Zhao et al., 2005; Wang and Irving, 2011). Generally, a membrane-mediated action is typical for water-soluble compounds, while cytosol-mediated activity is primarily triggered by lipophilic compounds.
Whereas, enzymes interact with their substrates in a geometrical way (“lock and key”), signaling molecules are thought to have a topochemical affinity to their receptors. It is assumed that the interaction of such components at the receptor site is cooperative and quantized (Gafurov and Zefirov, 2007). The bioactive compounds in some biostimulants are also proposed to display signaling activity in plants or induce signaling pathways. Various amino acids (Forde and Lea, 2007; Arbona et al., 2013), and peptides (Ivanov, 2010) function as signaling molecules in the regulation of plant growth and development (Ertani et al., 2009; Mochida and Shinozaki, 2011). Peptide signaling is important in various aspects of plant development and growth regulation including meristem organization, leaf morphogenesis, and defense responses to biotic and abiotic stress (Schiavon et al., 2008). Specific signaling peptides contained in a plant-derived protein hydrolysate have been shown to affect plant growth and development, defense responses, callus growth, meristem organization, root growth, leaf-shape regulation, and nodule development (Matsubayashi and Sakagami, 2006; Colla et al., 2013). Protein hydrolysates from soybean and casein have been shown to act as elicitors to enhance grapevine immunity against Plasmopara viticola (Lachhab et al., 2014).
Proteins may also contain hidden peptide sites, “cryptides” or “crypteins” in their amino acid sequence, which may have their own biological activities, distinct from its precursor (Ivanov, 2010; Samir and Link, 2011). Evidence that cryptides can trigger plants defense reactions have recently been demonstrated (Yamaguchi and Huffaker, 2011) and there are reports of the isolation of cryptides by hydrolysis of proteins from marine organisms, including seaweeds, and cryptides may be present naturally in a variety of biological derived products (Henda and Bordenave-Juchereau, 2014; Hayes et al., 2015).
Many small molecular weight substances are known to participate in signaling cascades in vivo. Exogenous amino acids may affect biological processes by acting directly as signal molecules or by influencing hormone action via amino acid conjugation (Tegeder, 2012). It has been suggested that amino acid based biostimulants are readily absorbed and translocated by plant tissues and once absorbed, they have the capacity to function as compatible osmolytes, transport regulators, signaling molecules, modulators of stomatal opening, and may detoxify heavy metals among other benefits (Kauffman et al., 2007). Sugars (Smeekens, 2000; Eveland and Jackson, 2012) and fatty acids and plant lipids (Kachroo and Kachroo, 2009) are also known to act as signaling molecules and mitigators of stress response in plants (Okazaki and Saito, 2014). Animal based lipid soluble fractions, have also been observed to produce an auxin-like response (Kauffman et al., 2007), while sugars, sucrose, and its cleavage products (hexoses), are also known to act as signaling molecules through regulation of gene expression and by interaction with other hormone signals including auxins. In a sunflower meal hydrolyzate, amino acids, humic substances, microelements, and sugars present in the biostimulant appeared to coordinate, with auxin-like compounds in complex signaling cross-talk promoting plant growth, enhancing plant transplanting success and increasing final crop yield (Ugolini et al., 2015).
Hormones are of central importance for the regulation of metabolic processes and plant development in a complex system of interacting hormones and cofactors, the functions of which are closely intertwined and mutually dependent (Wang and Irving, 2011). Biostimulants developed from humic substances, complex organic materials, seaweeds, antitranspirants, free amino acids (Du Jardin, 2012), and crude extracts of lower (Rathore et al., 2009) and higher plants (Yakhin et al., 2012) have been frequently demonstrated to have an effect on plant hormonal status (Kurepin et al., 2014). While hormone-like compounds may be present in biostimulants, it is also possible that de novo synthesis of hormones may be induced by such preparations in treated plants (Jannin et al., 2012) and amino acids, glycosides, polysaccharides and organic acids are contained in many biostimulants and may act as precursors or activators of endogenous plant hormones (Parađiković et al., 2011). Hormones or hormone-like effects could therefore be responsible for the action of natural biostimulants derived from microorganisms, algae, higher plants, animal, and humate based raw material (Table 4).
Metabolic Effects
Information on currently available biostimulants gives some insight into the possible biochemical and molecular genetic effects of biostimulants derived from different natural raw materials (Table 4). Many published reports are available suggesting various biostimulants improve plant productivity through increased assimilation of N, C, and S (Jannin et al., 2012, 2013), improved photosynthesis, improved stress responses, altered senescence, and enhanced ion transport (Gajic, 1989; Khan et al., 2009; Parađiković et al., 2011). Biostimulants are also reported to increase free amino acids, protein, carbohydrates, phenolic compounds, pigment levels, and various enzymes (Table 4). The protective effect of many biostimulants against biotic and abiotic stresses has been associated with a reduction of stress-induced reactive oxygen species, activation of the antioxidant defense system of plants, or increased levels of phenolic compounds (Ertani et al., 2011a, 2013a).
While it is clear that many biologically derived biostimulants contain small molecular weight compounds that are involved in signaling events and may directly influence plant metabolic processes, it remains unclear how an exogenous soil or foliar application of an uncharacterized product can have predictable and beneficial responses in plants. It is well-known, for example, that application of exogenous plant hormones or compounds that disrupt hormone function (PGR's) can have markedly negative effects on plants and that optimization of PGR materials and their applicaitons requires precise information on dosage and timing. Application of biostimulants for which the dosage and efficacy of the functional compounds is unknown, cannot, therefore, be expected to result in predictable plant responses and identification of molecules with effects on plant metabolic processes is not, in of itself, a sufficient explanation for the function of a biostimulant. It is also uncertain why the application of a biostimulant with purported function as a PGR, signaling molecule or other discrete compound would be superior to, or more easily controlled, than a direct application of the purified product itself.
Toxicological and Ecological Aspects
Modern crop production requires a balance of high and consistent productivity with maximum safety for consumers, agricultural workers, and the environment (Rathore et al., 2009; Jannin et al., 2012; Pecha et al., 2012). While some biostimulants have been analyzed with regard to unwanted side effects including negative impact on the natural environment (Janas and Posmyk, 2013) most biostimulants have not been fully characterized but have been regarded as generally recognized as safe (GRAS in the US) on the basis of the biological origin of their constituents (Thomas et al., 2013). Generally, biostimulants are assumed to be biodegradable, non-toxic, non-polluting and non-hazardous to various organisms. While this may be a rational conclusion for many formulations derived from biological materials such as seaweed extracts and their components (Turan and Köse, 2004; Dhargalkar and Pereira, 2005; Rathore et al., 2009; Michalak and Chojnacka, 2014; Stadnik and de Freitas, 2014), higher plants (Onatsky et al., 2001; Abdalla, 2013; Yakhin et al., 2013), chitin and chitosan (Bautista-Baños et al., 2006; Cabrera et al., 2013) it is not clear that this is a valid assumption for microbial products or products that would not normally be present in agricultural fields.
Biostimulants have been utilized as bioremediants and have been shown to improve ATP levels and phosphatase and urease activity (Tejada et al., 2011a), and hence increase the rate of degradation of xenobiotics in the soil (Tejada et al., 2010, 2011b) and to enhance beneficial soil microbial communities under semi-arid climates (Tejada et al., 2011b). Biostimulants may also help reduce the amount of potentially risky agrochemicals (Kolomazník et al., 2012) including reducing the use of fertilizers and pesticides (Hamza and Suggars, 2001). Most compounds contained in biostimulants are natural constituents of terrestrial and aquatic ecosystems (Jannin et al., 2012) and metabolites of plant and microbial origin and as such most are generally regarded as safe, particularly at the low rates at which they are typically applied. Thus, it has been proposed that biostimulants can be positioned as eco-friendly products for sustainable agriculture (Mladenova et al., 1998; Ertani et al., 2011a; Ghannam et al., 2013; Vijayanand et al., 2014). In many countries, however, biostimulants are not subject to rigorous toxicological screening (Traon et al., 2014) and there remains the potential for the persistence of human pathogens in materials of animal origin and for the synthesis of novel compounds of unknown function or toxicology during the manufacturing process.
Economic Aspects
Even though there have been relatively few rigorous demonstrations of the benefit of biostimulants, and to a large extent the mode of action of these products remains uncertain, the industry for biostimulants is substantial and rapidly growing. Though many recent “market” studies show that the market for these products is growing at a remarkable rate, the validity of these analyses must be considered with care as they frequently do not provide an explicit definition of term “biostimulants.” The value of the European biostimulants market ranged from €200 to €400 million in 2011, €500 million in 2013 and may grow to more than €800 million in 2018 with annual growth potential in 10% and more (EBIC, 2011a, 2013; Traon et al., 2014). France, Italy, Spain are the leading EU countries in the production of biostimulants (Traon et al., 2014). In North America, the biostimulant market was valued at $0.27 billion in 20131, and is expected to grow at a growth rate of 12.4% annually, to reach $0.69 billion by 2018, the USA is the largest producer and consumer of biostimulants in the region (http://www.micromarketmonitor.com/). In 2014, the USA market was assessed at $313.0 million and is projected to reach $605.1 million by 20192, at a CAGR of 14.1% (http://news.agropages.com/). The biostimulants market in the Asia-Pacific was valued at $0.25 billion in 2013, and is expected to grow at a CAGR of 12.9% annually, to reach $0.47 billion by 2018 (Asia Biostimulants Market, 2015)3. China and India are key countries playing a significant role. The Southeast Asian & Australasian biostimulants market was valued at $233.8 million in 2015, and is projected to reach $451.8 million by 2021 (http://news.agropages.com/)4. The market in Latin America was valued at $0.16 billion in 20135, and is expected to grow at a CAGR of 14.4% annually, to reach $0.32 billion by 2018 (http://www.micromarketmonitor.com/). This market is mostly concentrated in Brazil and Argentina. The regional market shares of the global biostimulants market6 are: EU—41.7%, North America—21.5%, the Asia-Pacific region—20%, Latin America—12.9%. Globally, it biostimulants were valued at $1402.15 million in 2014 and are projected to have aCAGR of 12.5% reaching $2524.02 million by 20197, largely as a consequence of growing interest in organic products. Wu (2016) summised that “the global biostimulants market is projected to reach $2.91 billion by 2021, with a CAGR (compound annual growth rate) of 10.4% from 2016 to 2021. In terms of area of application, the biostimulants market is projected to reach 24.9 million hectares by 2021 and is projected to grow at a CAGR of 11.7% from 2016 to 2021” (Wu, 2016).”
Problems and Prospects
The biostimulant industry faces many problems and challenges. Until recently biostimulant products based on natural raw materials and particularly waste stream has mainly been developed based on observational and less commonly, empirical data. While many contemporary biostimulants have been shown to be effective in practice, very few biostimulants can claim to understand the mechanisms or modes of action (Khan et al., 2009). Furthermore, while biostimulants can be categorized by source of origin, this is frequently inadequate as very substantial differences can exist between products even within a common feed stock origin. The challenge to biostimulant science is further exacerbated since composition and content of active substances in the original plant raw material can be affected by many factors including the location and growing conditions, season, species, variety, organ, and the phase of growth (Naumov et al., 1993; Dragovoz et al., 2009; Sharma et al., 2012b). Similarly, the response of the target crop can be expected to vary across crops and environments. One solution to this problem is to derive the raw materials for the biostimulant under highly regulated conditions. This approach has been successfully implemented by leading seaweed producers and fermentation based products that have developed harvesting and manufacturing processes that ensure uniformity of product performance through time. The development of a product with uniformity of response is not, however, a guarantee that the product is optimized for biological efficacy.
To address these issues, developments in -omics approaches will be critical in accelerating the discovery of mode of action of bioactive compounds (Aliferis and Jabaji, 2011; Craigie, 2011; Jannin et al., 2012) and optimizing their use. Metabolomics, phenomics and agronomics represent the integration of gene expression, protein interactions, and other regulatory processes as they impact on plant productivity and thus are more appropriate tools for discovery in this field than mRNA, transcripts, or proteins analyzed in isolation (Arbona et al., 2013). Integrative, multidisciplinary approaches using tools from transcriptomics in conjunction with metabolomics and biochemical analysis are necessary to establish the mechanism of action and to identify the active components in the extracts (Lee et al., 2012). The difficulty in identifying modes of action and subsequent standardization of composition of multicomponent biostimulants based on natural raw materials will continue to hamper the use, certification and registration of biostimulants. The solution to this problem will require the collaborative efforts of specialists from different fields: chemists, biologists, plant physiologists, industrial manufacture, sales and distribution and those with expertise in practical agricultural production (Raldugin, 2004; Craigie, 2011; Jannin et al., 2012; Lee et al., 2012).
Products with a single active substance represent a simpler construct in which the physiological effects and mechanism of action can be more readily determined and hence certification and registration is simpler. The multicomponent composition of many preparations, however, are much more difficult to characterize (Bozhkov et al., 1996), though they may offer novel insight into biological synergy (Bulgari et al., 2015), multifunctionality and emergence which may be crucial to product efficacy (Gerhardson, 2002). In the absence of a functional rationale for every constituent in a multicomponent biostimulant, it is likely that there will be molecules present that may positively or negatively influence plant productivity. Currently, it is almost impossible using available chemical-synthetic, and genetic engineering approaches to reproduce the full suite of molecules and complexes of biologically active substances (Kershengolts et al., 2008) that are present in most biostimulants.
Pro's and Con's of Biostimulants Science and Practice
Many have noted the state confusion in the field of biostimulants (Torre et al., 2013; Traon et al., 2014) and this has resulted in the opinion that much of the biostimulant market is not based on science or efficacy and that many products are little more than recycled waste products sold on the basis of pseudoscience and marketing. Indeed, research on several biostimulant products has shown them to be ineffective or to contain inactive, unstable or inconsistent properties with several showing negative effects compared when contrasted with well-designed controls (Csizinszky, 1984, 1986; Albregts et al., 1988; Di Marco and Osti, 2009; Vasconcelos et al., 2009; Banks and Percival, 2012; Cerdan et al., 2013; de Oliveira et al., 2013; Carvalho et al., 2014). For example, foliar and root application of a product containing amino acids from animal origin have been reported to cause severe plant-growth depression and negative effects on Fe nutrition while a second product containing amino acids from plant origin stimulated plant growth (Cerdan et al., 2013). In another report that tested several biostimulant products it was concluded that “none of the biostimulant products tested achieved a sufficient degree of pathogen control to warrant replacement of or supplementation with conventional synthetic fungicides” (Banks and Percival, 2012), and there have been demonstrated positive and negative impacts and overall questions of the economic feasibility of the use of humic substances for increasing crop yields (Rose et al., 2014). Since biological systems are inherently complex, and given that most biostimulant products have not been characterized and have received relatively little replicated and rigorous independent validation, it is perhaps not surprising that many products are ineffective or highly variable in response. Nevertheless, there are a significant number of rigorous independent reports of benefits from some biostimulant formulations and market growth data demonstrates that there is a good deal of support for these products within agricultural producer communities. That such market growth has occured, even in the absence of a known “mechanism of function” suggests that there are aspects of plant metabolism and productivity constraints that are not understood but are potentially important if we are to achieve the goal of increased global food production.
The market euphoria that is taking place in the biostimulant industry recognizes these unknowns and biostimulants are viewed by many innovators and investors as a mechanism to conduct broadscale, if unfocussed, discovery of novel biologically derived molecules. Much as the exploration of marine organisms, and plants and microbes from diverse ecosystems has led to the discovery of novel pharmaceuticals, so too the development of biostimulants from the broad range of source materials, holds significant promise of discovery. Recent years have seen rapid growth in the number of published studies, increased numbers of scientific conferences and development of legal framework and legislation. These trends will inevitably improve the image of this industry and the efficacy of products. Two significant problems still exist within the industry broadly: (1) preparations of products with highly complex multicomponent and incompletely identified composition make the identification of a primary mode of action extremely difficult and (2) the current classification and legislation/legal framework for regulation of biostimulants is based primarily on source material and not on biological mode of action. Hence there is insufficient capacity to differentiate products, and there is the potential for the successful demonstration of a single product within a biostimulant category, to falsely indicate the efficacy of the group as whole.
Several topical questions need consideration in the future:
1. Can living cultures of microorganisms, which have the ability to stimulate the growth of plants be referred to biostimulants?
2. Are non-essential elements that result in improved plant productivity, biostimulants?
3. How should biostimulants with a complex completely unidentified structure where all the components and modes/mechanisms involved have not been established be registered and regulated in national and international legislation?
4. What standard of proof of efficacy is appropriate that both stimulates development and discourages the sale of materials of no benefit?
5. On what principles, should the final classification of biostimulants be based and what categories should it contain?
Conclusions
Modern biostimulants are complex mixtures derived from raw materials of highly diverse origin utilizing highly diverse manufacturing processes and as such can be expected to have a broad spectrum of possible biological activity and safety. To distinguish biostimulants from the existing legislative product categories including essential nutrients, pesticides, or plant hormones a biostimulant should not solely function by virtue of the presence of elements or compounds of known function. We propose, therefore, a definition of a biostimulant as “a formulated product of biological origin that improves plant productivity as a consequence of the novel or emergent properties of the complex of constituents and not as a sole consequence of the presence of known essential plant nutrients, plant growth regulators, or plant protective compounds.” Consistent with this definition, the ultimate identification of a novel molecule within a biostimulant that is found to be wholly responsible for the biological function of that biostimulant, would necessitate the classification of the biostimulant according to the discovered function.
This novel definition is inspired by three observations: (1) that the development of the biostimulant industry will inevitably result in the discovery of novel biologically active molecules and that the identification and classification of these molecules will benefit biological discovery more greatly if these molecules are explicitly described than if they were merely labeled as “biostimulants,” (2) that there is a need for the nascent biostimulant industry to explicitly discourage the inclusion of nutrient elements and known biologically active molecules under the guise of a “biostimulant” and (3) that there is a need to recognize that classic reductionist biology/chemistry may indeed be insufficient to explain biological complexity (Luisi, 2002; Lüttge, 2012; Bertolli et al., 2014).
The definition provided here is important as it emphasizes the principle that biological function can be modulated through application of complex mixtures of molecules for which an explicit mode of action has not been defined. The definition also requires a demonstration of beneficial impacts of the biostimulant on plant productivity. Given the difficulty in determining a “mode of action” for a biostimulant, and recognizing the need for the market in biostimulants to attain legitimacy, we suggest that the focus of biostimulant research and validation should be upon determining the mechanism of action, without a requirement for the determination of a mode of action. This can be achieved through careful agronomic experimentation, molecular or biochemical demonstration of positive impact on biological processes or the use of advanced analytical equipment to identify functional constituents. Given the prerequisite multi-component and emergent characteristics of biostimulants, the discovery of the mode of action is likely to require application of new techniques in bioinformatics and systems biology. While the definition proposed here suggests that the development and marketing of a biostimulant does not require a demonstration of the mode of action, it is still in the interest of the commercial producers of these products to pursue an understanding of these products so that the product can be improved and optimized for use in various environments and cropping systems.
While there is a clear commercial imperative to rationalize biostimulants as a discrete class of products, there is also a compelling biological case for the science-based development of the biostimulant science that is grounded in the observation that the application of biological materials derived from various organisms, including plants, that have been exposed to stressors can affect metabolic and energetic processes in humans, animals, and plants (Filatov, 1951a,b). This hypothesis is based upon the premise that functional chemical communication occurs between individuals or organs that favorably modulate metabolic pathways and networks at different plant hierarchical levels. Inter and intra organism communication and consequent molecular and metabolic regulation are at the heart of the science of systems biology and the tools of systems biology will inevitably be critical to the realization of mode of action of many biostimulants. Continued investments by commercial entities in biostimulant research and product development will serve as a critical driver of discovery in this realm and will inevitably lead to the identification of novel biological phenomenon, pathways and processes that would not have been discovered if the category of biostimulants did not exist, or was not considered legitimate.
Author Contributions
All authors OY, AL, IY, PB, contributed equally to this review.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Russian Foundation for Basic Research (RFBR-Agidel No 02-04-97917), Russian Science Support Foundation, Foundation for Assistance to Small Innovative Enterprises in Science and Technology (“Start Program,” FASIE, Russia).
Footnotes
1. ^North America biostimulants market (2015). Available online at http://www.micromarketmonitor.com/ (Accessed February 27, 2015).
2. ^North America biostimulants market to reach $605.1 million by 2019 (2015). Available online at: http://news.agropages.com/ (accessed August 18, 2015).
3. ^Available online at: http://www.micromarketmonitor.com/ (Accessed February 27, 2015).
4. ^Southeast Asian and Australasian Biostimulants Market Trends and Forecasts to 2021 Available online at: http://news.agropages.com/ (Accessed August 15, 2016).
5. ^Latin America biostimulants market (2015). Available online at: http://www.micromarketmonitor.com/ (Accessed February 27, 2015).
6. ^Global biostimulants product market to reach $2241.0 million by 2018. Available online at: http://news.agropages.com/ (Accessed December 30, 2015).
7. ^Global biostimulants market to reach $2.52 bn by 2019. Available online at http://news.agropages.com/ (Accessed December 30, 2015).
References
Abbas, S. M. (2013). The influence of biostimulants on the growth and on the biochemical composition of Vicia faba CV. Giza 3 beans. Rom. Biotech. Lett. 18, 8061–8068. Available online at: http://www.rombio.eu/vol18nr2/1%20Salwa%20Mohamed.pdf
Abd El-Baky, H. H., Hussein, M. M., and El-Baroty, G. S. (2008). Algal extracts improve antioxidant defense abilities and salt tolerance of wheat plant irrigated with sea water. Afr. J. Biochem. Res. 7, 151–164. Available online at: http://www.academicjournals.org/journal/AJBR/article-abstract/82070DD10085
Abdalla, M. M. (2013). The potential of Moringa oleifera extract as a biostimulant in enhancing the growth, biochemical and hormonal contents in rocket (Eruca vesicaria subsp. sativa) plants. Int. J. Plant Physiol. Biochem. 5, 42–49. doi: 10.5897/IJPPB2012.026
Adani, F., Genevini, P., Zaccheo, P., and Zocchi, G. (1998). The effect of commercial humic acid on tomato plant growth and mineral nutrition. J. Plant. Nutr. 21, 561–575. doi: 10.1080/01904169809365424
Adholeya, A., Tiwari, P., and Singh, R. (2005). “Large-scale production of arbuscular mycorrhizal fungi on root organs and inoculation strategies,” in In vitro Culture of Mycorrhizas, eds S. Declerck, D. G. Strullu, and J. A. Fortin (Heidelberg: Springer), 315–338.
Aguado-Santacruz, G. A., Moreno-Gómez, B., Rascón-Cruz, Q., Aguirre-Mancilla, C., Espinosa-Solís, J. A., and González-Barriga, C. D. (2014). “Biofertilizers as complements to synthetic and organic fertilization,” in Components, Uses in Agriculture and Environmental Impacts, eds F. López-Valdes and F. Fernández-Luqueno (New York, NY: Nova Science Publishers Inc.), 155–180.
Aguirre, E., Leménager, D., Bacaicoa, E., Fuentes, M., Baigorri, R., Angel, M. Z., et al. (2009). The root application of a purified leonardite humic acid modifies the transcriptional regulation of the main physiological root responses to Fe deficiency in Fe-sufficient cucumber plants. Plant Physiol. Biochem. 47, 215–223. doi: 10.1016/j.plaphy.2009.06.004
Aharoni, A., and Galili, G. (2011). Metabolic engineering of the plant primary–secondary metabolism interface. Curr. Opin. Biotechnol. 22, 239–244. doi: 10.1016/j.copbio.2010.11.004
Aitken, J. B., and Senn, T. L. (1965). Seaweed products as a fertilizer and soil conditioner for horticultural crops. Bot. Mar. 8, 144–148. doi: 10.1515/botm.1965.8.1.144
Albregts, E. E., Howard, C. M., Chandler, C., and Mitchell, R. L. (1988). Effect of biostimulants on fruiting of strawberry. Proc. Fla. State Hort. Soc. 101, 370–372.
Aliferis, K. A., and Jabaji, S. (2011). Metabolomics – a robust bioanalytical approach for the discovery of the modes-of-action of pesticides: a review. Pestic. Biochem. Phys. 100, 105–117. doi: 10.1016/j.pestbp.2011.03.004
Apone, F., Arciello, S., Colucci, G., Filippini, L., and Portoso, D. (2006). Alle radici della biostimolazione: indagini scientifiche a supporto. Fertilitas Agrorum 1, 55–63. Available online at: http://fertilitasagrorum.ciec-italia.it/index_file/volumi.htm
Apone, F., Tito, A., Carola, A., Arciello, S., Tortora, A., Filippini, L., et al. (2010). A mixture of peptides and sugars derived from plant cell walls increases plant defense responses to stress and attenuates ageing-associated molecular changes in cultured skin cells. J. Biotechnol. 145, 367–376. doi: 10.1016/j.jbiotec.2009.11.021
Arbona, V., Manzi, M., Ollas, C., and Gómez-Cadenas, A. (2013). Metabolomics as a tool to investigate abiotic stress tolerance in plants. Int. J. Mol. Sci. 14, 4885–4911. doi: 10.3390/ijms14034885
Aremu, A. O., Masondo, N. A., Rengasamy, K. R. R., Amoo, S. O., Gruz, J., Bíba, O., et al. (2015b). Physiological role of phenolic biostimulants isolated from brown seaweed Ecklonia maxima on plant growth and development. Planta 246, 1313–1324. doi: 10.1007/s00425-015-2256-x
Aremu, A. O., Stirk, W. A., Kulkarni, M. G., Tarkowská, D., Turečková, V., Gruz, J., et al. (2015a). Evidence of phytohormones and phenolic acids variability in garden-waste-derived vermicompost leachate, a well-known plant growth stimulant. Plant Growth Regul. 75, 483–492. doi: 10.1007/s10725-014-0011-0
Arnao, M. B., and Hernández-Ruiz, J. (2014). Melatonin: plant growth regulator and/or biostimulator during stress? Trends Plant Sci. 19, 789–797. doi: 10.1016/j.tplants.2014.07.006
Arthur, G. D., Aremu, A. O., Moyo, M., Stirk, W. A., and van Staden, J. (2013). Growth-promoting effects of a seaweed concentrate at various pH and water hardness conditions. S. Afr. J. Sci. 109, 1–6. doi: 10.1590/sajs.2013/20120013
Aydin, A., Kant, C., and Turan, M. (2012). Humic acid application alleviate salinity stress of bean (Phaseolus vulgaris L.) plants decreasing membrane leakage. Afr. J. Agric. Res. 7, 1073–1086. doi: 10.5897/ajar10.274
Aylward, L. (2005). More superintendents are using biostimulants and trusting in them, companies say. Golfdom 54–58. doi: 10.5897/AJAR10.274
Baglieri, A., Cadili, V., Mozzetti Monterumici, C., Gennari, M., Tabasso, S., Montoneri, E., et al. (2014). Fertilization of bean plants with tomato plants hydrolysates. Effect on biomass production, chlorophyll content and N assimilation. Sci. Hortic. 176, 194–199. doi: 10.1016/j.scienta.2014.07.002
Banks, J., and Percival, G. C. (2012). Evaluation of biostimulants to control Guignardia leaf blotch (Guignardia aesculi) of horsechestnut and black spot (Diplocarpon rosae) of roses. Arboric. Urban Forest. 38, 258–261. Available online at: http://www.ncufc.org/uploads/biostimulantsanddisease.pdf
Bargiacchi, E., Miele, S., Romani, A., and Campo, M. (2013). Biostimulant activity of hydrolyzable tannins from sweet chestnut (Castanea sativa Mill.). Acta Hortic. 1009, 111–116. doi: 10.17660/ActaHortic.2013.1009.13
Basak, A. (2008). “Biostimulators – definitions, classification and legislation,” in Monographs Series: Biostimulators in Modern Agriculture. General Aspects, ed H. Gawrońska (Warsaw: Wieś Jutra), 7–17.
Bashan, Y. (1998). Inoculants of plant growth-promoting bacteria for use in agriculture. Biotechnol. Adv. 16, 729–770. doi: 10.1016/S0734-9750(98)00003-2
Bautista-Baños, S., Hernández-Lauzardo, A. N., Velázquez-del Valle, M. G., Hernández-López, M., Ait Barka, E., Bosquez-Molina, E., et al. (2006). Chitosan as a potential natural compound to control pre and postharvest diseases of horticultural commodities. Crop Prot. 25, 108–118. doi: 10.1016/j.cropro.2005.03.010
Beaudreau, D. G. Jr. (2013). Biostimulants in Agriculture: Their Current and Future Role in a Connected Agricultural Economy. Available online at: http://www.biostimulantcoalition.org/ (Accessed September 27, 2014).
Belakbir, A., Ruiz, J. M., and Romero, L. (1998). Yield and fruit quality of pepper (Capsicum annuum L.) in response to bioregulators. HortScience, 33, 85–87.
Berbara, R. L. L., and García, A. C. (2014). “Humic substances and plant defense metabolism,” in Physiological Mechanisms and Adaptation Strategies in Plants Under Changing Enviornoment, eds P. Ahmad and M. R. Wani (New York, NY: Springer Science+Business Media), 297–319.
Bertolli, S. C., Mazzafera, P., and Souza, G. M. (2014). Why is it so difficult to identify a single indicator of water stress in plants? a proposal for a multivariate analysis to assess emergent properties. Plant Biol. 16, 578–585. doi: 10.1111/plb.12088
Betti, L., Canova, A., Maini, P., Merendino, A., and Paolini, M. (1992). Effects of foliar application of an aminoacid-based biostimulant on the response of pepper seedling to PepMV infections. Adv. Hortic. Sci. 6, 97–103.
Bhalla, U. S., and Iyengar, R. (1999). Emergent properties of networks of biological signaling pathways. Science 283, 381–387. doi: 10.1126/science.283.5400.381
Bhardwaj, D., Ansari, M. W., Sahoo, R. K., and Tuteja, N. (2014). Biofertilizers function as key player in sustainable agriculture by improving soil fertility, plant tolerance and crop productivity. Microb. Cell Fact. 13:66. doi: 10.1186/1475-2859-13-66
Bhattacharyya, P. N., and Jha, D. K. (2012). Plant growth promoting rhizobacteria (PGPR): emergence in agriculture. World J. Microbiol. Biotechnol. 28, 1327–1350. doi: 10.1007/s11274-011-0979-9
Billard, V., Etienne, P., Jannin, L., Garnica, M., Cruz, F., Garcia-Mina, J.-M., et al. (2014). Two biostimulants derived from algae or humic acid induce similar responses in the mineral content and gene expression of winter oilseed rape (Brassica napus L.). J. Plant Growth Regul. 33, 305–316. doi: 10.1007/s00344-013-9372-2
Blagoveshchensky, A. V. (1945). Biochemical factors of natural selection in plants. J. Gen. Biol. 6, 217–234.
Blagoveshchensky, A. V. (1956). Biogenic stimulants and biochemical nature of their action. Bull. Main Bot. Garden. 25, 79–86.
Borriss, R. (2011). “Use of plant-associated Bacillus strains as biofertilizers and biocontrol agents in agriculture,” in Bacteria in Agrobiology: Plant Growth Responses, ed D. K. Maheshwari (Berlin: Springer), 41–76.
Botta, A., Sierras, N., Sánchez-Hernández, L., Martinez, M. J., and Bru, R. (2015). “Understanding the effect of an amino acids based biostimulant by an enantiomeric analysis of their active principles and a proteomic profiling approach,” in Abstracts Book for Oral and Poster Presentations of the 2st World Congress on the use of Biostimulants in Agriculture; 2015 Nov 16-19, eds P. Perata, P. Brown, R. A. Alvarez, and M. Ponchet (Florence: New Ag International), 40.
Bozhkov, A. I., Menzyanova, N. G., and Leontovich, V. P. (1996). Lipid-composition and antibacterial activity of root exudates secreted by wheat seedlings. Russ. J. Plant Physiol. 43, 795–799.
Brice-o-Domínguez, D., Hernández-Carmona, G., Moyo, M., Stirk, W., and van Staden, J. (2014). Plant growth promoting activity of seaweed liquid extracts produced from Macrocystis pyrifera under different pH and temperature conditions. J. Appl. Phycol. 26, 2203–2210. doi: 10.1007/s10811-014-0237-2
Brown, P., and Saa, S. (2015). Biostimulants in agriculture. Front. Plant Sci. 6:671. doi: 10.3389/fpls.2015.00671
Bulgari, R., Cocetta, G., Trivellini, A., Vernieri, P., and Ferrante, A. (2015). Biostimulants and crop responses: a review. Biol. Agric. Hortic. 31, 1–17. doi: 10.1080/01448765.2014.964649
Cabrera, J. C., Wégria, G., Onderwater, R. C. A., Nápoles, M. C., Falcón-Rodríguez, A. B., Costales, D., et al. (2013). Practical use of oligosaccharins in agriculture. Acta Hortic. 1009, 195–212. doi: 10.17660/ActaHortic.2013.1009.24
Cacco, G., and Dell'Agnola, G. (1984). Plant growth regulator activity of soluble humic complexes. Can. J. Soil Sci. 64, 225–228. doi: 10.4141/cjss84-023
Calvo, P., Nelson, L., and Kloepper, J. W. (2014). Agricultural uses of plant biostimulants. Plant Soil 383, 3–41. doi: 10.1007/s11104-014-2131-8
Cambri, D., Filippini, L., Apone, F., Arciello, S., Colucci, G., and Portoso, D. (2008). “Effect of aminoplant® on expression of selected genes in Arabidopsis thaliana L. plants,” in Biostimulators in Modern Agriculture. General Aspects, ed H. Gawrońska Warsaw: Wieś Jutra), 77–82.
Canellas, L. P., and Olivares, F. L. (2014). Physiological responses to humic substances as plant growth promoter. Chem. Biol. Technol. Agric. 1:3. doi: 10.1186/2196-5641-1-3
Canellas, L. P., Olivares, F. L., Okorokova-Façanha, A. L., and Façanha, A. R. (2002). Humic acids isolated from earthworm compost enhance root elongation, lateral root emergence, and plasma membrane H+-ATPase activity in maize roots. Plant Physiol. 130, 1951–1957. doi: 10.1104/pp.007088
Canellas, L. P., Piccolo, A., Dobbss, L. B., Spaccini, R., Olivares, F. L., Zandonadi, D. B., et al. (2010). Chemical composition and bioactivity properties of size-fractions separated from a vermicompost humic acid. Chemosphere 78, 457–466. doi: 10.1016/j.chemosphere.2009.10.018
Carvalho, M. E. A., Castro, P. R. D. C. E., Gallo, L. A., and Ferraz, M. V. D. C. Jr. (2014). Seaweed extract provides development and production of wheat. Rev. Agrarian 7, 166–170. doi: 10.5281/zenodo.51607
Caulet, R. P., Gradinariu, G., Iurea, D., and Morariu, A. (2014). Influence of furostanol glycosides treatments on strawberry (Fragaria × ananassa Duch.) growth and photosynthetic characteristics under drought condition. Sci. Hortic. 169, 179–188. doi: 10.1016/j.scienta.2014.02.031
Cerdan, M., Sanchez-Sanchez, A., Jorda, J. D., Juarez, M., and Sanchez-Andreu, J. (2013). Effect of commercial amino acids on iron nutrition of tomato plants grown under lime-induced iron deficiency. J. Plant Nutr. Soil Sci. 176, 859–866. doi: 10.1002/jpln.201200525
Chambers, J. W. (2014). Tea Extracts and Uses in Promoting Plant Growth. U.S. Patent No. 20140113814 A1, 27. Available online at: http://www.freepatentsonline.com/y2014/0113814.html
Chambolle, C. (2005). Biostimulants: humus substances. PHM Rev. Hortic. 468, 21–23. Available online at: https://www.cabdirect.org/cabdirect/abstract/20053088318
Chbani, A., Mawlawi, H., and Zaouk, L. (2013). Evaluation of brown seaweed (Padina pavonica) as biostimulant of plant growth and development. Afr. J. Agric. Res. 13, 1155–1165. doi: 10.5897/AJAR12.1346
Chen, T., Gu, J., Zhang, X., Ma, Y., Cao, L., Wang, Z., et al. (2014). System-level study on synergism and antagonism of active ingredients in traditional chinese medicine by using molecular imprinting technology. Sci. Rep. 4:7159. doi: 10.1038/srep07159
Chen, Y., Clapp, C. E., and Magen, H. (2004). Mechanisms of plant growth stimulation by humic substances: the role of organo iron complexes. Soil Sci. Plant Nutr. 50, 1089–1095. doi: 10.1080/00380768.2004.10408579
Chiu, H. C., Levy, R., and Borenstein, E. (2014). Emergent biosynthetic capacity in simple microbial communities. PLoS Comput. Biol. 10:e1003695. doi: 10.1371/journal.pcbi.1003695
Chojnacka, K., Michalak, I., Dmytryk, A., Wilk, R., and Gorecki, H. (2015). “Innovative natural plant growth biostimulants,” in Fertilizer Technology: II Biofertilizer, eds S. Sinha, K. K. Pant, S. Bajpai, and J. N. Govil (Houston, TX: Studium Press LLC), 451–489.
Christofoletti, C. A., Escher, J. P., Correia, J. E., Marinho, J. F. U., and Fontanetti, C. S. (2013). Sugarcane vinasse: environmental implications of its use. Waste Manage. 33, 2752–2761. doi: 10.1016/j.wasman.2013.09.005
Ciavatta, C., and Cavani, L. (2006). Problematiche per l'inserimento dei biostimolanti nella legislazione dei fertilizzanti. Fertilitas Agrorum 1, 11–15. Available online at: http://fertilitasagrorum.ciec-italia.it/index_file/volumi.htm
Ciesiołka, D., Gulewicz, P., Martinez-Villaluenga, C., Pilarski, R., Bednarczyk, M., and Gulewicz, K. (2005). Products and biopreparations from alkaloid-rich lupin in animal nutrition and ecological agriculture. Folia Biol. 53, 59–66. doi: 10.3409/173491605775789443
Colla, G., Rouphael, Y., Canaguier, R., Svecova, E., and Cardarelli, M. (2014). Biostimulant action of a plant-derived protein hydrolysate produced through enzymatic hydrolysis. Front. Plant Sci. 5:448. doi: 10.3389/fpls.2014.00448
Colla, G., Rouphael, Y., Di Mattia, E., El-Nakhel, C., and Cardarelli, M. (2015). Co-inoculation of Glomus intraradices and Trichoderma atroviride acts as a biostimulant to promote growth, yield and nutrient uptake of vegetable crops. J. Sci. Food Agric. 95, 1706–1715. doi: 10.1002/jsfa.6875
Colla, G., Svecova, E., Rouphael, Y., Cardarelli, M., Reynaud, H., Canaguier, R., et al. (2013). Effectiveness of a plant-derived protein Hydrolysate to improve crop performances under different growing conditions. Acta Hortic. 1009, 175–180. doi: 10.17660/ActaHortic.2013.1009.21
Conan, C., Guiboileau, A., Joubert, J.-M., and Potin, P. (2015). “Investigations of seaweed filtrate as Biostimulant,” in Abstracts Book for Oral and Poster Presentations of the 2st World Congress on the use of Biostimulants in Agriculture; 2015 Nov 16-19, eds P. Perata, P. Brown, R. A. Alvarez, and M. Ponchet (Florence: New Ag International), 75.
Cordeiro, F. C., Santa-Catarina, C., Silveira, V., and Souza, S. R. D. (2011). Humic acid effect on catalase activity and the generation of reactive oxygen species in corn (Zea mays). Biosci. Biotechnol. Biochem. 75, 70–74. doi: 10.1271/bbb.100553
Corte, L., Dell'Abate, M. T., Magini, A., Migliore, M., Felici, B., Roscini, L., et al. (2014). Assessment of safety and efficiency of nitrogen organic fertilizers from animal-based protein hydrolysates-a laboratory multidisciplinary approach. J. Sci. Food Agr. 94, 235–245. doi: 10.1002/jsfa.6239
Craigie, J. S. (2011). Seaweed extract stimuli in plant science and agriculture. J. Appl. Phycol. 23, 371–393. doi: 10.1007/s10811-010-9560-4
Craigie, J. S., MacKinnon, S. L., and Walter, J. A. (2009). Liquid seaweed extracts identified using 1H NMR profiles. J. Appl. Phycol. 20, 665–671. doi: 10.1007/s10811-007-9232-1
Crouch, I. J., and van Staden, J. (1993a). Commercial seaweed products as biostimulants in horticulture. J. Home Consum. Hort. 1, 19–76.
Crouch, I. J., and van Staden, J. (1993b). Evidence for the presence of plant growth regulators in commercial seaweed products. Plant Growth Reg. 13, 21–29.
Crouch, I. J., Smith, M. T., van Staden, J., Lewis, M. J., and Hoad, G. V. (1992). Identification of auxins in a commercial seaweed concentrate. J. Plant. Physiol. 139, 590–594. doi: 10.1016/S0176-1617(11)80375-5
Csizinszky, A. A. (1984). Response of tomatoes to seaweed based nutrient sprays. Proc. Fla. State Hort. Sco. 97, 151–157.
Csizinszky, A. A. (1986). Response of tomatoes to foliar biostimulant sprays. Proc. Fla. State Hort. Soc. 99, 353–358.
Cutler, H. G., and Cutler, S. J. (2004). “Growth regulators, plant,” in Kirk-Othmer Encyclopedia of Chemical Technology. John Wiley and Sons Inc. doi: 10.1002/0471238961.1612011403212012.a01.pub2
Daniels, R. S (2013). Corn Steep Liquor as A Biostimulant Composition. U.S. Patent No. 8568758 B2, 12. Available online at: http://www.freepatentsonline.com/y2012/0028801.html
de Fretes, C. E., Sembiring, L., and Purwestri, Y. A. (2013). Characterization of Streptomyces spp. producing indole-3-acetic acid as biostimulant agent. Indones. J. Biotech. 18, 83–91. doi: 10.22146/ijbiotech.7872
De Lucia, B., and Vecchietti, L. (2012). Type of bio-stimulant and application method effects on stem quality and root system growth in L.A. Lily. Europ. J. Hort. Sci. 77, 10–15. Available online at: http://www.jstor.org/stable/24126519
de Oliveira, F. D. A. D., Medeiros, J. F. D., Oliveira, M. K. T. D., Souza, A. A. T., Ferreira, J. A., and Souza, M. S. (2013). Interaction between water salinity and biostimulant in the cowpea plants. Rev. Brasil. de Eng. Agríc. Ambien. 17, 465–471. doi: 10.1590/S1415-43662013000500001
Dhargalkar, V. K., and Pereira, N. (2005). Seaweed: promising plant of the millennium. Sci. Cult. 71, 60–66. Available online at: http://drs.nio.org/drs/handle/2264/489
Di Marco, S., and Osti, F. (2009). Effect of biostimulant sprays on Phaeomoniella chlamydospora and esca proper infected vines under greenhouse and field conditions. Phytopathol. Mediterr. 48, 47–58. doi: 10.14601/Phytopathol_Mediterr-2874
Dixon, G. R., and Walsh, U. F. (2004). “Suppressing Pythium ultimum induced damping-off in cabbage seedlings by biostimulation with proprietary liquid seaweed extracts managing soil-borne pathogens: a sound rhizosphere to improve productivity in intensive horticultural systems,” in Proceedings of the XXVI Inter. Horticultural Congr (Toronto, ON).
Doak, S. O., Schmidt, R. E., and Ervin, E. H. (2005). Metabolic enhancer impact on creeping bentgrass leaf sodium and physiology under salinity. Int. Turfgr. Soc. Res. J. 10, 845–849. Available online at: http://turfsociety.com/itsjournal.html
Dobbss, L. B., Canellas, L. P., Olivares, F. L., Aguiar, N. O., Peres, L. E. P., Azevedo, M., et al. (2010). Bioactivity of chemically transformed humic matter from vermicompost on plant root growth. J. Agric. Food Chem. 58, 3681–3688. doi: 10.1021/jf904385c
Dragovoz, I. V., Yavorskaya, V. K., Antoniuk, V. P., and Kurchii, B. A. (2009). Hormonal substances produced by microorganism association from ginseng roots. Physiol. Biochem. Cultivated plants 41, 393–399.
Dubravec, K., Dubravec, I., and Manitasevic, J. (1995). The effect of the bioregulators Agrispon® and Ergostim on the vegetative and reproductive growth of apples. J. Sustain. Agr. 5, 73–83. doi: 10.1300/J064v05n01_06
Du Jardin, P. (2012). The Science of Plant Biostimulants - A Bibliographic Analysis, Ad hoc Study Report. Brussels: European Commission. Available online at: http://hdl.handle.net/2268/169257 (Accessed April 25, 2013).
Du Jardin, P. (2015). Plant biostimulants: definition, concept, main categories and regulation. Sci. Hortic. 196, 3–14. doi: 10.1016/j.scienta.2015.09.021
Dumas, B., Vergnes, S., Attia, F., and Noël, D. (2012). “Characterization of a new phyto-stimulating preparation: mode of action and evaluation of agronomic performance,” in Abstracts Book for Oral and Poster Presentations of the 1st World Congress on the use of Biostimulants in Agriculture; 2012 Nov 26-29, eds P. Perata, P. Brown, and M. Ponchet (Strasbourg: New Ag International), 86.
EBIC (2011a). Economic Overview of the Biostimulants Sector in Europe. Available online at: http://www.biostimulants.eu/ (Accessed October 23, 2012).
EBIC (2011b). Available online at: http://www.biostimulants.eu/2011/10/biostimulants-definition-agreed/ (Accessed July 29, 2014)
EBIC (2012). Available online at: http://www.biostimulants.eu/ (Accessed September 27, 2014).
EBIC (2013). Economic Overview of the Biostimulants Sector in Europe. Available online at: http://www.biostimulants.eu/ (Accessed September 23, 2014).
Elliott, M. L., and Prevatte, M. (1996). Response of ‘Tifdwarf’ Bermudagrass to Seaweed-derived Biostimulants. HortTechnology 6, 261–263.
Ertani, A., Cavani, L., Pizzeghello, D., Brandellero, E., Altissimo, A., Ciavatta, C., et al. (2009). Biostimulant activity of two protein hydrolysates in the growth and nitrogen metabolism of maize seedlings. J. Plant Nutr. Soil Sci. 172, 237–244. doi: 10.1002/jpln.200800174
Ertani, A., Francioso, O., Tugnoli, V., Righi, V., and Nardi, S. (2011b). Effect of commercial lignosulfonate-humate on Zea mays L. metabolism. J. Agric. Food Chem. 59, 11940–11948. doi: 10.1021/jf202473e
Ertani, A., Pizzeghello, D., Altissimo, A., and Nardi, S. (2013b). Use of meat hydrolysate derived from tanning residues as plant biostimulant for hydroponically grown maize. J. Plant Nutr. Soil Sci. 176, 287–295. doi: 10.1002/jpln.201200020
Ertani, A., Pizzeghello, D., Baglieri, A., Cadili, V., Tambone, F., Gennari, M., et al. (2013c). Humic-like substances from agro-industrial residues affect growth and nitrogen assimilation in maize (Zea mays L.) plantlets. J. Geochem. Explor. 129, 103–111. doi: 10.1016/j.gexplo.2012.10.001
Ertani, A., Pizzeghello, D., Francioso, O., Sambo, P., Sanchez-Cortes, S., and Nardi, S. (2014). Capsicum chinensis L. growth and nutraceutical properties are enhanced by biostimulants in a long-term period: chemical and metabolomic approaches. Front. Plant Sci. 5:375. doi: 10.3389/fpls.2014.00375
Ertani, A., Schiavon, M., Altissimo, A., Franceschi, C., and Nardi, S. (2011a). Phenol-containing organic substances stimulate phenylpropanoid metabolism in Zea mays. J. Plant Nutr. Soil Sci. 174, 496–503. doi: 10.1002/jpln.201000075
Ertani, A., Schiavon, M., Muscolo, A., and Nardi, S. (2013a). Alfalfa plant-derived biostimulant stimulate short-term growth of salt stressed Zea mays L. plants. Plant Soil 364, 145–158. doi: 10.1007/s11104-012-1335-z
Eveland, A. L., and Jackson, D. P. (2012). Sugars, signalling, and plant development. J. Exp. Bot. 63, 3367–3377. doi: 10.1093/jxb/err379
Featonby-Smith, B. C., and van Staden, J. (1983). The effect of seaweed concentrate on the growth of tomato plants in nematode-infested soil. Sci. Hortic. 20, 137–146. doi: 10.1016/0304-4238(83)90134-6
Ferrante, A., Trivellini, A., Vernieri, P., and Piaggesi, A. (2013). Application of Actiwave® for improving the rooting of Camellia cuttings. Acta Hortic. 1009, 213–218. doi: 10.17660/ActaHortic.2013.1009.25
Filatov, V. P. (1951a). Tissue treatment. (Doctrine on biogenic stimulators). I. Background, methods and the clinical tissue treatment. Priroda 11, 39–46.
Filatov, V. P. (1951b). Tissue treatment. (Doctrine on biogenic stimulators). II. Hypothesis of tissue therapy, or the doctrine on biogenic stimulators. Priroda 12, 20–28.
Finnie, J. F., and van Staden, J. (1985). Effect of seaweed concentrate and applied hormones on in vitro cultured tomato roots. J. Plant Physiol. 120, 215–222. doi: 10.1016/S0176-1617(85)80108-5
Fleming, C. C., Turner, S. J., and Hunt, M. (2006). Management of root knot nematodes in turfgrass using mustard formulations and biostimulants. Commun. Agric. Appl. Biol. Sci. 71, 653–658.
Forde, B. G., and Lea, P. J. (2007). Glutamate in plants: metabolism, regulation, and signalling. J. Exp. Bot. 58, 2339–2358. doi: 10.1093/jxb/erm121
Fuentes-Ramirez, L. E., and Caballero-Mellado, J. (2006). “Bacterial biofertilizers,” in PGPR: Biocontrol and Biofertilization, ed Z. A. Siddiqui (Dordrecht: Springer), 143–172.
Gafurov, R. G., and Zefirov, N. S. (2007). A role of the molecular structure of phytoregulators in chemical signal perception by receptors of plant hormonal systems. Mosc. Univ. Chem. Bull. 62, 52–56. doi: 10.3103/S0027131407010129
Gajic, B. R. (1989). Bioregulator Based on Plant Raw Materials and Method for Producing Same. U.S. Patent No. 4872900. Available online at: http://www.freepatentsonline.com/4872900.html
Gandarillas Infante, M. J. (2012). Biostimulant Composition for Growth and Production of Capsicum chinense L. that Comprises a Carbohydrate Solution Originating from Chitosan and a Partial Hydrolysate of Saccharomyces Cerevisiae and Method for the Use of said Composition. Patent CL: WO2012045189A2, 16. Available online at: http://www.freepatentsonline.com/WO2012045189A2.html
García, A. C., Izquierdo, F. G., and Berbara, R. L. L. (2014). “Effects of humic materials on plant metabolism and agricultural productivity,” in Emerging Technologies and Management of Crop Stress Tolerance, ed P. Ahmad (Amsterdam; Boston, MA; Heidelberg; London; New York, NY; Oxford; Paris; San Diego, CA; San Francisco, CA; Singapore; Sydney, NSW; Tokyo: Elsevier Inc.), 449–466.
García, A. C., Santos, L. A., Izquierdo, F. G., Sperandio, M. V. L., Castro, R. N., and Berbara, R. L. L. (2012). Vermicompost humic acids as an ecological pathway to protect rice plant against oxidative stress. Ecol. Eng. 47, 203–208. doi: 10.1016/j.ecoleng.2012.06.011
Gates, D. E., Parker, C., Boston, C. L., Allen, A., Pabon, A., Nelson, M. L., et al. (2012). “Effects of biostimulants containing fermentation metabolites and nutrients on plant response to abiotic stress,” in Abstracts Book for Oral and Poster Presentations of the 1st World Congress on the use of Biostimulants in Agriculture; 2012 Nov 26-29, eds P. Perata, P. Brown, and M. Ponchet (Strasbourg: New Ag International), 96.
Gerhardson, B. (2002). Biological substitutes for pesticides. Trends Biotechnol. 20, 338–343. doi: 10.1016/S0167-7799(02)02021-8
Ghannam, A., Abbas, A., Alek, H., Al-Waari, Z., and Al-Ktaifani, M. (2013). Enhancement of local plant immunity against tobacco mosaic virus infection after treatment with sulphated-carrageenan from red alga (Hypnea musciformis). Physiol. Mol. Plant. 84, 19–27. doi: 10.1016/j.pmpp.2013.07.001
Giannattasio, M., Vendramin, E., Fornasier, F., Alberghini, S., Zanardo, M., Stellin, F., et al. (2013). Microbiological features and bioactivity of a fermented manure product (Preparation 500) used in biodynamic agriculture. J. Microbiol. Biotechnol. 23, 644–651. doi: 10.4014/jmb.1212.12004
Goatley, J. M., and Schmidt, R. E. (1991). Biostimulator enhancement of Kentucky bluegrass sod. HortScience 26, 254–255.
Goñi, O., Fort, A., Quille, P., Mckeown, P. C., Spillane, C., and O'Connell, S. (2016). Comparative transcriptome analysis of two Ascophyllum nodosum extract biostimulants: same seaweed but different. J. Agr. Food Chem. 64, 2980–2989. doi: 10.1021/acs.jafc.6b00621
González, A., Castro, J., Vera, J., and Moenne, A. (2013). Seaweed oligosaccharides stimulate plant growth by enhancing carbon and nitrogen assimilation, basal metabolism, and cell division. J. Plant Growth. Regul. 32, 443–448. doi: 10.1007/s00344-012-9309-1
González, A., Contreras, R. A., Zúiga, G., and Moenne, A. (2014). Oligo-carrageenan kappa-induced reducing redox status and activation of TRR/TRX system increase the level of indole-3-acetic acid, gibberellin A3 and trans-zeatin in Eucalyptus globulus trees. Molecules 19, 12690–12698. doi: 10.3390/molecules190812690
Gordon, D. M. (1947). The treatment of retinitis pigmentosa with special reference to the Filatov method. Am. J. Ophthalmol. 30, 565–580. doi: 10.1016/0002-9394(47)92310-6
Gupta, U. C., and MacLeod, J. A. (1982). Effect of sea crop 16 and ergostim on crop yields and plant composition. Can. J. Soil Sci. 62, 527–532. doi: 10.4141/cjss82-057
Halmann, M. (1990). Synthetic plant growth regulators. Adv. Agron. 43, 47–105. doi: 10.1016/S0065-2113(08)60476-9
Halpern, M., Bar-Tal, A., Ofek, M., Minz, D., Muller, T., and Yermiyahu, U. (2015). The use of biostimulants for enhancing nutrient uptake. Adv. Agron. 130, 141–174. doi: 10.1016/bs.agron.2014.10.001
Hammad, S. A. R., and Ali, O. A. M. (2014). Physiological and biochemical studies on drought tolerance of wheat plants by application of amino acids and yeast extract. Ann. Agr. Sci. 59, 133–145. doi: 10.1016/j.aoas.2014.06.018
Hamza, B., and Suggars, A. (2001). Biostimulants: myths and realities. TurfGrass Trends 8, 6–10. Available online at: http://archive.lib.msu.edu/tic/tgtre/2001.html#aug; http://archive.lib.msu.edu/tic/tgtre/article/2001aug6.pdf
Hanafy, M. S., Saadawy, F. M., Milad, S. M. N., and Ali, R. M. (2012). Effect of some natural extracts on growth and chemical constituents of Schefflera arboricola plants. J. Hort. Sci. Ornamen. Plants 4, 26–33. Available online at: http://idosi.org/jhsop/jhsop4(1)12.htm; http://idosi.org/jhsop/4(1)12/4.pdf
Hayes, M., García-García, M., Fitzgerald, C., and Lafarga, T. (2015). “Seaweed and milk derived bioactive peptides and small molecules in functional foods and cosmeceuticals,” in Biotechnology of Bioactive Compounds: Sources and applications, eds V. K. Gupta and M. G. Tuohy (Chichester: John Wiley and Sons, Ltd.), 669–691.
Henda, Y. B., and Bordenave-Juchereau, S. (2014). “Using marine cryptides against metabolic syndrome,” in Bioactive Compounds from Marine Foods: Plant and Animal Sources, eds B. Hernández-Ledesma and M. Herrero (Chichester: John Wiley & Sons, Ltd.), 95–112.
Herbreteau, F., Coiffard, L. J. M., Derrien, A., and De Roeck-Holtzhauer, Y. (1997). The fatty acid composition of five species of macroalgae. Bot. Mar. 40, 25–27.
Hernandez, O. L., Calderín, A., Huelva, R., Martínez-Balmori, D., Guridi, F., Aguiar, N. O., et al. (2015). Humic substances from vermicompost enhance urban lettuce production. Agron. Sustain. Dev. 35, 225–232. doi: 10.1007/s13593-014-0221-x
Hernandez-Herrera, R. M., Santacruz-Ruvalcaba, F., Ruiz-Lopez, M. A., Norrie, J., and Hernandez-Carmona, G. (2014). Effect of liquid seaweed extracts on growth of tomato seedlings (Solanum lycopersicum L.). J. Appl. Phycol. 26, 619–628. doi: 10.1007/s10811-013-0078-4
Herve, J. J. (1994). Biostimulants, a new concept for the future; prospects offered by the chemistry of synthesis and biotechnology. Comptes Rendus Acad. Agric. Fr. 80, 91–102.
Hirsch, R., Hartung, W., and Gimmler, H. (1989). Abscisic acid content of algae under stress. Bot. Acta 102, 326–334. doi: 10.1111/j.1438-8677.1989.tb00113.x
IJdo, M., Cranenbrouck, S., and Declerck, S. (2010). Methods for large-scale production of AM fungi: past, present, and future. Mycorrhiza 21, 1–16. doi: 10.1007/s00572-010-0337-z
Ikrina, M. A., and Kolbin, A. M. (2004). Regulators of Plant Growth and Development, Vol. 1, Stimulants. Moscow: Chimia.
Ivanov, V. T. (2010). Peptides as universal biological regulators. Her. Russ. Acad. Sci. 80, 419–429. doi: 10.1134/S1019331610050011
Janas, K. M., and Posmyk, M. M. (2013). Melatonin, an underestimated natural substance with great potential for agricultural application. Acta Physiol. Plant 35, 3285–3292. doi: 10.1007/s11738-013-1372-0
Jannin, L., Arkoun, M., Etienne, P., Laîné, P., Goux, D., Garnica, M., et al. (2013). Brassica napus growth is promoted by Ascophyllum nodosum (L.) Le Jol. seaweed extract: microarray analysis and physiological characterization of N, C, and S metabolisms. J. Plant Growth Regul. 32, 31–52. doi: 10.1007/s00344-012-9273-9
Jannin, L., Arkoun, M., Ourry, A., Laîné, P., Goux, D., Garnica, M., et al. (2012). Microarray analysis of humic acid effects on Brassica napus growth: involvement of N, C and S metabolisms. Plant Soil. 359, 297–319. doi: 10.1007/s11104-012-1191-x
Jenkins, T. A (2014). Bio-Stimulant for Improved Plant Growth and Development. U.S. Patent No. 20140162877A1, 7. Available online at: http://www.freepatentsonline.com/y2014/0162877.html
Johnson, C. W. (2006). What are emergent properties and how do they affect the engineering of complex systems? Reliability Eng. Syst. Safety 91, 1475–1481. doi: 10.1016/j.ress.2006.01.008
Kachroo, A., and Kachroo, P. (2009). Fatty acid–derived signals in plant defense. Ann. Rev. Phytopathol. 47, 153–176. doi: 10.1146/annurev-phyto-080508-081820
Kauffman, G. L., Kneivel, D. P., and Watschke, T. L. (2007). Effects of a biostimulant on the heat tolerance associated with photosynthetic capacity, membrane thermostability, and polyphenol production of perennial ryegrass. Crop Sci. 47, 261–267. doi: 10.2135/cropsci2006.03.0171
Kelting, M., Harris, J. R., Fanelli, J., and Appleton, B. (1998). Humate-based biostimulants affect early post-transplant root growth and sapflow of red maple. HortScience 33, 342–344.
Kershengolts, B. M., Zhuravskaya, A. N., Filippova, G. V., An'shakova, V. V., Shein, A. A., Khlebny, E. S., et al. (2008). Innovational nano-biotechnologies in decision of problems in northern regions of Russia. Bull. Yakut. State Univ. 5, 20–31. Available online at: http://cyberleninka.ru/article/n/innovatsionnye-nanobiotehnologii-v-reshenii-aktualnyh-problem-severnyh-regionov-rossii
Khan, W., Hiltz, D., Critchley, A. T., and Prithiviraj, B. (2011). Bioassay to detect Ascophyllum nodosum extract-induced cytokinin-like activity in Arabidopsis thaliana. J. Appl. Phycol. 23, 409–414. doi: 10.1007/s10811-010-9583-x
Khan, W., Prithiviraj, B., and Smith, D. L. (2003). Chitosan and chitin oligomers increase phenylalanine ammonia-lyase and tyrosine ammonia-lyase activities in soybean leaves. J Plant Physiol. 160, 859–863. doi: 10.1078/0176-1617-00905
Khan, W., Rayirath, U. P., Subramanian, S., Jithesh, M. N., Rayorath, P., Hodges, D. M., et al. (2009). Seaweed extracts as biostimulants of plant growth and development. J. Plant Growth Regul. 28, 386–399. doi: 10.1007/s00344-009-9103-x
Kinnersley, A. M. (1993). The role of phytochelates in plant growth and productivity. Plant Growth Regul. 12, 207–218. doi: 10.1007/BF00027200
Kleiber, T., and Markiewicz, B. (2013). Application of “Tytanit” in greenhouse tomato growing. Acta Sci. Pol. Hortorum Cultus 12, 117–126. Available online at: http://www.acta.media.pl/pl/action/getfull.php?id=3436
Kolomazník, K., Pecha, J., Friebrová, V., Janáčová, D., and Vašek, V. (2012). Diffusion of biostimulators into plant tissues. Heat Mass Transfer. 48, 1505–1512. doi: 10.1007/s00231-012-0998-6
Korosov, A. V. (2012). An emergent principle in ecology. Princ. Èkol. 1, 48–66. doi: 10.15393/j1.art.2012.1481
Krouk, G. (2015). “How plants respond to a combination of signals: are biostimulant effects triggered by a cocktail of molecules?” in Abstracts Book for Oral and Poster Presentations of the 2nd World Congress on the Use of Biostimulants in Agriculture; 2015 Nov 16–19, eds P. Perata, P. Brown, R. A. Alvarez, and M. Ponchet (Florence: New Ag International), 28.
Krouk, G. (2016). Hormones and nitrate: a two-way connection. Plant Mol. Biol. 91, 599–606. doi: 10.1007/s11103-016-0463-x
Krouk, G., Crawford, N. M., Coruzzi, G. M., and Tsay, Y. F. (2010). Nitrate signaling: adaptation to fluctuating environments. Curr. Opin. Plant Biol. 13, 265–272. doi: 10.1016/j.pbi.2009.12.003
Krouk, G., Ruffel, S., Gutiérrez, R. A., Gojon, A., Crawford, N. M., Coruzzi, G. M., et al. (2011). A framework integrating plant growth with hormones and nutrients. Trends Plant Sci. 16, 178–182. doi: 10.1016/j.tplants.2011.02.004
Krouk, G., Tranchina, D., Lejay, L., Cruikshank, A. A., Shasha, D., Coruzzi, G. M., et al. (2009). A systems approach uncovers restrictions for signal interactions regulating genome-wide responses to nutritional cues in Arabidopsis. PLoS Comput Biol. 5:e1000326. doi: 10.1371/journal.pcbi.1000326
Kumar, D., and Shivay, Y. S. (2008). Definitional Glossary of Agricultural Terms. V. I. IK. New Delhi: International Publishing House Pvt Ltd.
Kunicki, E., Grabowska, A., Sękara, A., and Wojciechowska, R. (2010). The effect of cultivar type, time of cultivation, and biostimulant treatment on the yield of spinach (Spinacia oleracea L.). Folia Hortic. 22, 9–13. doi: 10.2478/fhort-2013-0153
Kurepin, L. V., Zaman, M., and Pharis, R. P. (2014). Phytohormonal basis for the plant growth promoting action of naturally occurring biostimulators. J. Sci. Food Agric. 94, 1715–1722. doi: 10.1002/jsfa.6545
Lachhab, N., Sanzani, S. M., Adrian, M., Chiltz, A., Balacey, S., Boselli, M., et al. (2014). Soybean and casein hydrolysates induce grapevine immune responses and resistance against Plasmopara viticola. Front. Plant Sci. 5:716. doi: 10.3389/fpls.2014.00716
Leal, D., Matsuhiro, B., Rossi, M., and Caruso, F. (2008). FT-IR spectra of alginic acid block fractions in three species of brown seaweeds. Carbohydr. Res. 343, 308–316. doi: 10.1016/j.carres.2007.10.016
Lee, S., Yoon, J. Y., Jung, H. I., Lee, D. J., Shin, D. Y., Hyun, K. H., et al. (2012). Ameliorative effects of squash (Cucurbita moschata Duchesne ex Poiret) leaf extracts on oxidative stress. Plant Growth Regul. 67, 9–17. doi: 10.1007/s10725-011-9655-1
Linser, A., Cazzara, L., and Barbieri, G. (2006). Plant growth promoting rhizobacteria: a new opportunity for a sustainable agriculture. Fertilitas Agrorum. 1, 65–75. Available online at: http://fertilitasagrorum.ciec-italia.it/index_file/volumi.htm
Lisiecka, J., Knaflewski, M., Spizewski, T., Fraszczak, B., Kaluzewicz, A., and Krzesinski, W. (2011). The effect of animal protein hydrolysate on quantity and quality of strawberry daughter plants cv. ‘Elsanta’. Acta Sci. Pol.-Hortoru. 10, 31–40. Available online at: http://www.acta.media.pl/pl/action/getfull.php?id=2706
Lötze, E., and Hoffman, E. W. (2016). Nutrient composition and content of various biological active compounds of three South African-based commercial seaweed biostimulants. J. Appl. Phycol. 28, 1379–1386. doi: 10.1007/s10811-015-0644-z
Lovatt, C. J (2015). Use of a Natural Metabolite to Increase Crop Production. U.S. Patent Application No. 14/880,120. Available online at: http://www.freepatentsonline.com/y2016/0088842.html
Lucini, L., Rouphael, Y., Cardarelli, M., Canaguier, R., Kumar, P., and Colla, G. (2015). The effect of a plant-derived biostimulant on metabolic profiling and crop performance of lettuce grown under saline conditions. Sci. Hortic. 182, 124–133. doi: 10.1016/j.scienta.2014.11.022
Luisi, P. L. (2002). Emergence in chemistry: Chemistry as the embodiment of emergence. Found. Chem. 4, 183–200. doi: 10.1023/A:1020672005348
Lüttge, U. (2012). Modularity and emergence: biology's challenge in understanding life. Plant Biol. 14, 865–871. doi: 10.1111/j.1438-8677.2012.00659.x
Maini, P. (2006). The experience of the first biostimulant, based on aminoacids and peptides: a short retrospective review on the laboratory researches and the practical results. Fertilitas Agrorum. 1, 29–43. Available online at: http://fertilitasagrorum.ciec-italia.it/index_file/volumi.htm
Malusá, E., and Vassilev, N. A. (2014). Contribution to set a legal framework for biofertilisers. Appl. Microbiol. Biotechnol. 98, 6599–6607. doi: 10.1007/s00253-014-5828-y
Malusá, E., Sas-Paszt, L., and Ciesielska, J. (2012). Technologies for beneficial microorganisms inocula used as biofertilizers. Sci. World J. 98, 6599–6607. doi: 10.1100/2012/491206
Martínez-Esteso, M. J., Vilella-Antón, M. T., Sellés-Marchart, S., Martínez-Márquez, A., Botta-Català, A., Piñol-Dastis, R., et al. (2016). A DIGE proteomic analysis of wheat flag leaf treated with TERRA-SORB® foliar, a free amino acid high content biostimulant. J. Integr. Omics 6, 9–17. doi: 10.5584/jiomics.v6i1.188
Martínez-Viveros, O., Jorquera, M. A., Crowley, D. E., Gajardo, G., and Mora, M. L. (2010). Mechanisms and practical considerations involved in plant growth promotion by rhizobacteria. J. Soil Sci. Plant Nutr. 10, 293–319. doi: 10.4067/S0718-95162010000100006
Matsubayashi, Y., and Sakagami, Y. (2006). Peptide hormones in plants. Annu. Rev. Plant Biol. 57, 649–674. doi: 10.1146/annurev.arplant.56.032604.144204
Matyjaszczyk, E. (2015). Products containing microorganisms as a tool in integrated pest management and the rules of their market placement in the European Union. Pest Manag. Sci. 71, 1201–1206. doi: 10.1002/ps.3986
Mayr, E. (1982). The Growth of Biological Thought: Diversity, Evolution, and Inheritance. Cambridge, MA; London: Harvard University Press.
McCarty, L. B. (2001). Best Golf Course Management Practices. Upper Saddle River, NJ: Prentice-Hall.
Mercier, L., Lafitte, C., Borderies, G., Briand, X., Esquerré-Tugayé, M. T., and Fournier, J. (2001). The algal polysaccharide carrageenans can act as an elicitor of plant defence. New Phytol. 149, 43–51. doi: 10.1046/j.1469-8137.2001.00011.x
Michalak, I., and Chojnacka, K. (2014). Algal extracts: Technology and advances. Eng Life Sci. 14, 581–591. doi: 10.1002/elsc.201400139
Michalski, T. (2008). “Possibilities of maize production increase using non-conventional technologies,” in Monographs series: Biostimulators in Modern Agriculture. General Aspects, ed H. Gawrońska (Warsaw: Wieś Jutra), 30–53.
Migliore, M., Felici, B., Benedetti, A., and Florio, A. (2013). Proposal of bioassays as a tool for screening biostimulant properties of protein hydrolysates from animal waste materials. Acta Hortic. 1009, 235–240. doi: 10.17660/ActaHortic.2013.1009.28
Mikiciuk, M., and Dobromilska, R. (2014). Assessment of yield and physiological indices of small-sized tomato cv. ‘Bianka F1’ under the influence of biostimulators of marine algae origin. Acta Sci. Pol.-Hortoru. 13, 31–41. Available online at: http://www.acta.media.pl/pl/action/getfull.php?id=3851
Miller, C. O., Skoog, F., Von Saltza, M. H., and Strong, F. M. (1955). Kinetin, a cell division factor from deoxyribonucleic acid. J. Am. Chem. Soc. 77, 1392–1392. doi: 10.1021/ja01610a105
Mladenova, Y. I. (1978). Effect of L-glutamic acid and Siapton leaf organic fertilizer on oxidized nicotinamide adenine dinucleotide dependent glutamate dehydrogenase of different maize genotypes. J. Agric. Food. Chem. 26, 1274–1276. doi: 10.1021/jf60220a034
Mladenova, Y. I., Maini, P., Mallegni, C., Goltsev, V., Vladova, R., Vinarova, K., et al. (1998). Siapton – an amino-acid-based biostimulant reducing osmostress metabolic changes in maize. Agro Food Ind. Hi-Tech. 9, 18–22.
Mochida, K., and Shinozaki, K. (2011). Advances in omics and bioinformatics tools for systems analyses of plant functions. Plant Cell Physiol. 52, 2017–2038. doi: 10.1093/pcp/pcr153
Mora, V., Bacaicoa, E., Zamarreño, A.-M., Aguirre, E., Garnica, M., Fuentes, M., et al. (2010). Action of humic acid on promotion of cucumber shoot growth involves nitrate-related changes associated with the root-to-shoot distribution of cytokinins, polyamines and mineral nutrients. J. Plant Physiol. 167, 633–642. doi: 10.1016/j.jplph.2009.11.018
Murch, S. J., and Saxena, P. K. (2002). Melatonin: a potential regulator of plant growth and development? In vitro Cell. Dev. Biol. Plant 38, 531–536. doi: 10.1079/IVP2002333
Nabil, S., and Cosson, J. (1996). Seasonal variations in sterol composition of Delesseria sanguinea (Ceramiales, Rhodophyta). Hydrobiologia 326, 511–514. doi: 10.1007/BF00047854
Nardi, S., Ertani, A., Concheri, G., and Pizzeghello, D. (2006). Metodi di determinazione dell'attivita biostimolante. Fertilitas Agrorum. 1, 47–53. Available online at: http://fertilitasagrorum.ciec-italia.it/index_file/volumi.htm
Nardi, S., Muscolo, A., Vaccaro, S., Baiano, S., Spaccini, R., and Piccolo, A. (2007). Relationship between molecular characteristics of soil humic fractions and glycolytic pathway and Krebs cycle in maize seedlings. Soil Biol. Biochem. 39, 3138–3146. doi: 10.1016/j.soilbio.2007.07.006
Nardi, S., Pizzeghello, D., Schiavon, M., and Ertani, A. (2016). Plant biostimulants: physiological responses induced by protein hydrolyzed-based products and humic substances in plant metabolism. Sci. Agric. 73, 18–23. doi: 10.1590/0103-9016-2015-0006
Nardi, S., Tosoni, M., Pizzeghello, D., Provenzano, M. R., Cilenti, A., Sturaro, A., et al. (2005). Chemical characteristics and biological activity of organic substances extracted from soils by root exudates. Soil Sci. Soc. Am. J. 69, 2012–2019. doi: 10.2136/sssaj2004.0401
Naumov, G. F., Bozhkov, A. I., Leontovich, V. P., Sklyar, A. I., and Belous, A. M. (1993). Polyfunctionality of allelopathic substance allelostim. Doklady Akademii nauk Ukrainy 11, 166–169.
Nelson, W. R., and van Staden, J. (1985). 1-Aminocyclopropane-1-carboxylic acid in seaweed concentrate. Bot. Mar. 28, 415–418.
Okazaki, Y., and Saito, K. (2014). Roles of lipids as signaling molecules and mitigators during stress response in plants. Plant J. 79, 584–596. doi: 10.1111/tpj.12556
Omar, H., Abdullatif, B., Al-Kazan, M., and El-Gendy, A. (2015). Various applications of seaweed improves growth and biochemical constituents of Zea mays L. and Helianthus annuus L. J. Plant Nutr. 38, 28–40. doi: 10.1080/01904167.2014.911893
Onatsky, N. M., Yakhin, I. A., Rybalkin, S. P., Mikhina, L. V., Yakhin, O. I., and Ibatullina, R. B. (2001). Toxicological assessment of the preparation Stifun and substantiation of its MAC in the occupational air. Toxicol. Rev. 5, 20–24. Available online at: http://en.toxreview.ru/
Painter, T. J. (1983). “Algal polysaccharides,” in The Polysaccharides, ed G. O. Aspinall (New York, NY: Academic Press), 195–285.
Parađiković, N., Vinković, T., Vinković Vrček, I., and Tkalec, M. (2013). Natural biostimulants reduce the incidence of BER in sweet yellow pepper plants (Capsicum annuum L.). Agric. Food Sci. 22, 307–317. Available online at: http://ojs.tsv.fi/index.php/AFS/article/view/7354
Parađiković, N., Vinković, T., Vinković Vrček, I., Žuntar, I., Bojić, M., and Medić-Šarić, M. (2011). Effect of natural biostimulants on yield and nutritional quality: an example of sweet yellow pepper (Capsicum annuum L.) plants. J. Sci. Food Agric. 91, 2146–2152. doi: 10.1002/jsfa.4431
Pardo-García, A. I., Martínez-Gil, A. M., Cadahía, E., Pardo, F., Alonso, G. L., and Salinas, M. R. (2014). Oak extract application to grapevines as a plant biostimulant to increase wine polyphenols. Food Res. Int. 55, 150–160. doi: 10.1016/j.foodres.2013.11.004
Parrado, J., Bautista, J., Romero, E. J., García-Martínez, A. M., Friaza, V., and Tejada, M. (2008). Production of a carob enzymatic extract: potential use as a biofertilizer. Bioresour. Technol. 99, 2312–2318. doi: 10.1016/j.biortech.2007.05.029
Parrado, J., Escudero-Gilete, M. L., Friaza, V., García-Martínez, A., González-Miret, M. L., Bautista, J. D., et al. (2007). Enzymatic vegetable extract with bio-active components: influence of fertiliser on the colour and anthocyanins of red grapes. J. Sci. Food. Agric. 87, 2310–2318. doi: 10.1002/jsfa.2989
Pecha, J., Fürst, T., Kolomazník, K., Friebrová, V., and Svoboda, P. (2012). Protein biostimulant foliar uptake modeling: the impact of climatic conditions. AICHE J. 58, 2010–2019. doi: 10.1002/aic.12739
Petrozza, A., Santaniello, A., Summerer, S., Di Tommaso, G., Di Tommaso, D., Paparelli, E., et al. (2014). Physiological responses to Megafol® treatments in tomato plants under drought stress: a phenomic and molecular approach. Sci. Hortic. 174, 185–192. doi: 10.1016/j.scienta.2014.05.023
Pizzeghello, D., Francioso, O., Ertani, A., Muscolo, A., and Nardi, S. (2013). Isopentenyladenosine and cytokinin-like activity of different humic substances. J. Geochem. Explor. 129, 70–75. doi: 10.1016/j.gexplo.2012.10.007
Pretorius, J. C (2007). Seed Suspensions from “Lupinus albus”, Isolated Compounds Thereof and Use as Biological Plant Strengthening Agent. Patent No. WO2007090438 A1, 59. Available online at: http://www.freepatentsonline.com/WO2007090438A1.html
Pretorius, J. C (2013). Extracts and Compounds from “Agapanthus africanus” and Their Use as Biological Plant Protecting Agents. Patent No. WO2007003286 A2. Available online at: http://www.freepatentsonline.com/WO2007003286A2.html
Przybysz, A., Gawrońska, H., and Gajc-Wolska, J. (2014). Biological mode of action of a nitrophenolates-based biostimulant: case study. Front. Plant Sci. 5:713c. doi: 10.3389/fpls.2014.00713
Radkowski, A., and Radkowska, I. (2013). Effect of foliar application of growth biostimulant on quality and nutritive value of meadow sward. Ecol. Chem. Eng. A. 20, 1205–1211. doi: 10.2428/ecea.2013.20(10)110
Raldugin, V. A. (2004). Triterpenoids of fir and highly efficient plant growth regulator based on them. Russ. Chem. J. 48, 84–88.
Rathore, S. S., Chaudhary, D. R., Boricha, G. N., Ghosh, A., Bhatt, B. P., Zodape, S. T., et al. (2009). Effect of seaweed extract on the growth, yield and nutrient uptake of soybean (Glycine max) under rainfed conditions. South Afr. J. Bot. 75, 351–355. doi: 10.1016/j.sajb.2008.10.009
Rattan, R. S. (2010). Mechanism of action of insecticidal secondary metabolites of plant origin. Crop Prot. 29, 913–920. doi: 10.1016/j.cropro.2010.05.008
Ravensberg, W. J. (2015). “Commercialisation of microbes: present situation and future prospects,” in Principles of Plant-Microbe Interactions, ed B. Lugtenberg (Cham; Heidelberg; New York, NY; Dordrecht; London: Springer International Publishing), 309–317.
Rayorath, P., Khan, W., Palanisamy, R., MacKinnon, S. L., Stefanova, R., Hankins, S. D., et al. (2008). Extracts of the brown seaweed Ascophyllum nodosum induce gibberellic acid (GA3)-independent amylase activity in barley. J. Plant Growth Regul. 27, 370–379. doi: 10.1007/s00344-008-9063-6
Rengasamy, K. R. R., Kulkarni, M. G., Stirk, W. A., and Van Staden, J. (2015a). Eckol improves growth, enzyme activities, and secondary metabolite content in maize (Zea mays cv. Border King). J. Plant Growth Regul. 34, 410–416. doi: 10.1007/s00344-015-9479-8
Rengasamy, K. R. R., Kulkarni, M. G., Stirk, W. A., and Van Staden, J. (2015b). Eckol - a new plant growth stimulant from the brown seaweed Ecklonia maxima. J. Appl. Phycol. 27, 581–587. doi: 10.1007/s10811-014-0337-z
Rivera, C. M., Salerno, A., Sequi, P., Rea, E., and Trinchera, A. (2010). Exploring biostimulant effect of a brassicacea plant extract: use of maize seedling development as reference bioassay. Acta Hortic. 884, 737–744. doi: 10.17660/ActaHortic.2010.884.100
Rodríguez-Morgado, B., Gómez, I., Parrado, J., and Tejada, M. (2014). Behaviour of oxyfluorfen in soils amended with edaphic biostimulants/biofertilizers obtained from sewage sludge and chicken feathers. Effects on soil biological properties. Environ. Sci. Pollut. R. 21, 11027–11035. doi: 10.1007/s11356-014-3040-3
Rose, M. T., Patti, A. F., Little, K. R., Brown, A. L., Jackson, W. R., and Cavagnaro, T. R. (2014). A meta-analysis and review of plant-growth response to humic substances: practical implications for agriculture. Adv. Agron. 124, 37–89. doi: 10.1016/B978-0-12-800138-7.00002-4
Rouse, R. E. (1984). Evaluation of two commercially available biostimulants on citrus. J. Rio Grande Valley Hortic. Soc. 37, 107–112.
Roy, R. N., Finck, A., Blair, G. J., and Tandon, H. L. S. (2006). Plant Nutrition for Food Security-A Guide for Integrated Nutrient Management. FAO Fertilizer and Plant Nutrition Bulletin 16, Rome: FAO.
Ruiz, J. M., Castilla, N., and Romero, L. (2000). Nitrogen metabolism in pepper plants applied with different bioregulators. J. Agric. Food Chem. 48, 2925–2929. doi: 10.1021/jf990394h
Russo, R. O., and Berlyn, G. P. (1991). The use of organic biostimulants to help low-input sustainable agriculture. J. Sustain. Agric. 1, 19–42. doi: 10.1300/J064v01n02_04
Saa, S., Olivos-Del Rio, A., Castro, S., and Brown, P. H. (2015). Foliar application of microbial and plant based biostimulants increases growth and potassium uptake in almond (Prunus dulcis Mill. D. A. Webb). Front. Plant Sci. 6:87. doi: 10.3389/fpls.2015.00087
Samir, P., and Link, A. J. (2011). Analyzing the cryptome: uncovering secret sequences. AAPS J. 13, 152–158. doi: 10.1208/s12248-011-9252-2
Sánchez-Gómez, R., Zalacain, A., Alonso, G. L., and Salinas, M. R. (2014). Vine-shoot waste aqueous extracts for re-use in agriculture obtained by different extraction techniques: phenolic, volatile, and mineral compounds. J. Agric. Food Chem. 62, 10861–10872. doi: 10.1021/jf503929v
Sanders, D. C., Ricotta, J. A., and Hodges, L. (1990). Improvement of carrot stands with plant biostimulants and fluid drilling. HortScience 25, 181–183.
Sanderson, K. J., and Jameson, P. E. (1986). The cytokinins in a liquid seaweed extract: could they be the active ingredients? Acta Hortic. 179, 113–116.
Santaniello, A., Di Tommaso, D., Povero, D., Piaggesi, A., and Perata, P. (2015). “Comparative analysis of the effects induced by three different Ascophyllum nodosum extracts: Algea® Fert BASE, SOLID and SOLID G,” in Abstracts Book for Oral and Poster Presentations of the 2st World Congress on the use of Biostimulants in Agriculture; 2015 Nov 16-19, eds P. Perata, P. Brown, R. A. Alvarez, and M. Ponchet (Florence: New Ag International), 100.
Santaniello, A., Giorgi, F. M., Di Tommaso, D., Di Tommaso, G., Piaggesi, A., and Perata, P. (2013). Genomic approaches to unveil the physiological pathways activated in Arabidopsis treated with plant-derived raw extracts. Acta Hortic. 1009, 161–174. doi: 10.17660/ActaHortic.2013.1009.20
Satish, L., Rameshkumar, R., Rathinapriya, P., Pandian, S., Rency, A. S., Sunitha, T., et al. (2015). Effect of seaweed liquid extracts and plant growth regulators on in vitro mass propagation of brinjal (Solanum melongena L.) through hypocotyl and leaf disc explants. J. Appl. Phycol. 27, 993–1002. doi: 10.1007/s10811-014-0375-6
Schiavon, M., Ertani, A., and Nardi, S. (2008). Effects of an alfalfa protein hydrolysate on the gene expression and activity of enzymes of the tricarboxylic acid (TCA) Cycle and nitrogen metabolism in Zea mays L. J. Agric. Food Chem. 56, 11800–11808. doi: 10.1021/jf802362g
Schiavon, M., Pizzeghello, D., Muscolo, A., Vaccaro, S., Francioso, O., and Nardi, S. (2010). High molecular size humic substances enhance phenylpropanoid metabolism in maize (Zea mays L.). J. Chem. Ecol. 36, 662–669. doi: 10.1007/s10886-010-9790-6
Schmidt, R. E., Ervin, E. H., and Zhang, X. (2003). Questions and answers about biostimulants. Golf Course Manage. 71, 91–94.
Sharma, H. S. S., Fleming, C., Selby, C., Rao, J. R., and Martin, T. (2014). Plant biostimulants: a review on the processing of macroalgae and use of extracts for crop management to reduce abiotic and biotic stresses. J. Appl. Phycol. 26, 465–490. doi: 10.1007/s10811-013-0101-9
Sharma, K., Bruns, C., Butz, A. F., and Finckh, M. R. (2012). Effects of fertilizers and plant strengtheners on the susceptibility of tomatoes to single and mixed isolates of Phytophthora infestans. Eur J. Plant Pathol. 133, 739–751. doi: 10.1007/s10658-012-9954-z
Sharma, S. H. S., Lyons, G., McRoberts, C., McCall, D., Carmichael, E., Andrews, F., et al. (2012a). Biostimulant activity of brown seaweed species from Strangford Lough: compositional analyses of polysaccharides and bioassay of extracts using mung bean (Vigna mungo L.) and pak choi (Brassica rapa chinensis L.). J. Appl. Phycol. 24, 1081–1091. doi: 10.1007/s10811-011-9737-5
Sharma, S. H. S., Lyons, G., McRoberts, C., McCall, D., Carmichael, E., Andrews, F., et al. (2012b). Brown seaweed species from Strangford Lough: compositional analyses of seaweed species and biostimulant formulations by rapid instrumental methods. J. Appl. Phycol. 24, 1141–1157. doi: 10.1007/s10811-011-9744-6
Sharp, R. G. (2013). A Review of the applications of chitin and its derivatives in agriculture to modify plant-microbial interactions and improve crop yields. Agronomy 3, 757–793. doi: 10.3390/agronomy3040757
Sleighter, R. L., Caricasole, P., Richards, K. M., Hanson, T., and Hatcher, P. G. (2015). “Molecular level characterization of humic substances and correlation with plant growth stimulation,” in Abstracts Book for Oral and Poster Presentations of the 2st World Congress on the use of Biostimulants in Agriculture 2015 Nov 16-19, eds P. Perata, P. Brown, R. A. Alvarez, and M. Ponchet (Florence: New Ag International), 10.
Smeekens, S. (2000). Sugar-induced signal transduction in plants. Annu. Rev. Plant Phys. 51, 49–81. doi: 10.1146/annurev.arplant.51.1.49
Smoleń, S. (2012). “Foliar nutrition: current state of knowledge and opportunities,” in advances in citrus nutrition, ed A. K. Srivastava (Dordrecht; Heidelberg; New York, NY; London: Springer Science+Business Media BV), 41–58.
Sofo, A., Nuzzaci, M., Vitti, A., Tataranni, G., and Scopa, A. (2014). “Control of biotic and abiotic stresses in cultivated plants by the use of biostimulant microorganisms,” in Improvement of Crops in the Era of Climatic Changes, eds P. Ahmad, M. R. Wani, M. M. Azooz, and L. S. Phan Tran (New York, NY: Springer Science +Business Media), 107–117.
Spaepen, S. (2015). “Plant hormones produced by microbes,” in Principles of plant-microbe interactions, ed B. Lugtenberg (Cham; Heidelberg; New York, NY; Dordrecht; London: Springer International Publishing), 247–256.
Spinelli, F., Fiori, G., Noferini, M., Sprocatti, M., and Costa, G. (2010). A novel type of seaweed extract as a natural alternative to the use of iron chelates in strawberry production. Sci. Hortic. 125, 263–269. doi: 10.1016/j.scienta.2010.03.011
Srivastava, A., Bhatia, G., Pant, R., and Srivastava, P. C. (2010). Bioefficacy and residue studies of Fantac (biostimulant) in rice crop under sub-tropical conditions. J. Environ. Prot. 1, 261–263. doi: 10.4236/jep.2010.13031
Srivastava, A., Bhatia, G., and Srivastava, P. C. (2008). Persistence behavior of Fantac biostimulant in Chili and soil under subtropical conditions. B. Environ. Contam. Tox. 80, 403–406. doi: 10.1007/s00128-008-9390-0
Stadnik, M. J., and de Freitas, M. B. (2014). Algal polysaccharides as source of plant resistance inducers. Trop. Plant Pathol. 39, 111–118. doi: 10.1590/S1982-56762014000200001
Stirk, W. A., and Staden, J. (1996). Comparison of cytokinin- and auxin-like activity in some commercially used seaweed extracts. J. Appl. Phycol. 8, 503–508. doi: 10.1007/BF02186328
Stirk, W. A., and Van Staden, J. (2006). “Seaweed products as biostimulatns in agriculture,” in World Seaweed Resources, eds A. T. Critchley, M. Ohno, and D. B. Largo (Amsterdam: ETI Information Services Ltd.), 1–32.
Stirk, W. A., Novák, O., Strnad, M., and Van Staden, J. (2003). Cytokinins in macroalgae. Plant Growth Regul. 41, 13–24. doi: 10.1023/A:1027376507197
Stirk, W. A., Tarkowská, D., Turečová, V., Strnad, M., and van Staden, J. (2014). Abscisic acid, gibberellins and brassinosteroids in Kelpak®, a commercial seaweed extract made from Ecklonia maxima. J. Appl. Phycol. 26, 561–567. doi: 10.1007/s10811-013-0062-z
Sukhoverkhov, F. M. (1967). “The effect of cobalt, vitamins, tissue preparations and antibiotics on carp production,” in Proceedings of the FAO world Symposium on Warm-Water Pond Fish Culture (Rome), 44, 400–407.
Summerer, S., Petrozza, A., and Cellini, F. (2013). High throughput plant phenotyping: a new and objective method to detect and analyse the biostimulant properties of different products. Acta Hortic. 1009, 143–148. doi: 10.17660/ActaHortic.2013.1009.17
Tachibana, S., Summer, P., Ewing, J., Miwa, T., and Kitazawa, D (2012). Method of Producing Plant Biostimulant. U.S. Patent No. 20120129695 A1 p. 14. Available online at: http://www.freepatentsonline.com/y2012/0129695.html
Tagliavini, S., and Kubiskin, C. (2006). Effetti della biostimolazione in ortofrutticoltura: alcune esperienze a confronto. Fertilitas Agrorum 1, 23–28. Available online at: http://fertilitasagrorum.ciec-italia.it/index_file/volumi.htm
Tandon, S., and Dubey, A. (2015). Effects of Biozyme (Ascophyllum nodosum) biostimulant on growth and development of soybean [Glycine max (L.) Merill]. Commun. Soil Sci. Plant Anal. 46, 845–858. doi: 10.1080/00103624.2015.1011749
Tay, S. A. B., Palni, L. M. S., and MacLeod, J. K. (1987). Identification of cytokinin glucosides in seaweed extract. J. Plant Growth Regul. 5, 133–138. doi: 10.1007/BF02087181
Tegeder, M. (2012). Transporters for amino acids in plant cells: some functions and many unknowns. Curr. Opin. Plant. Biol. 15, 315–321. doi: 10.1016/j.pbi.2012.02.001
Tejada, M., Benitez, C., and Parrado, J. (2011a). Application of biostimulants in benzo(a)pyrene polluted soils: short-time effects on soil biochemical properties. Appl. Soil Ecol. 50, 21–26. doi: 10.1016/j.apsoil.2011.08.002
Tejada, M., Benitez, C., Gymeza, I., and Parrado, J. (2011b). Use of biostimulants on soil restoration: effects on soil biochemical properties and microbial community. Appl. Soil Ecol. 49, 11–17. doi: 10.1016/j.apsoil.2011.07.009
Tejada, M., García-Martínez, A. M., Gómez, I., and Parrado, J. (2010). Application of MCPA herbicide on soils amended with biostimulants: short-time effects on soil biological properties. Chemosphere 80, 1088–1094. doi: 10.1016/j.chemosphere.2010.04.074
Tejada, M., Rodríguez-Morgado, B., Gómez, I., and Parrado, J. (2014). Degradation of chlorpyrifos using different biostimulants/biofertilizers: effects on soil biochemical properties and microbial community. Appl. Soil Ecol. 84, 158–165. doi: 10.1016/j.apsoil.2014.07.007
Thao, H. T. B., and Yamakawa, T. (2009). Phosphite (phosphorous acid): fungicide, fertilizer or bio-stimulator? Soil Sci. Plant. Nutr. 55, 228–234. doi: 10.1111/j.1747-0765.2009.00365.x
Thomas, M., Chauhan, D., Patel, J., and Panchal, T. (2013). Analysis of biostimulants made by fermentation of Sargassum tenerimum seaweed. Int. J. Curr. Trop. Res. 2, 405–407.
Tian, S. K., Lu, L. L., Xie, R. H., Zhang, M. Z., Jernstedt, J. A., Hou, D. D., et al. (2015). Supplemental macronutrients and microbial fermentation products improve the uptake and transport of foliar applied zinc in sunflower (Helianthus annuus L.) plants. Studies utilizing micro X-ray florescence. Front. Plant Sci. 6:87. doi: 10.3389/fpls.2014.00808
Torre, L. A., Battaglia, V., and Caradonia, F. (2013). Legal aspects of the use of plant strengtheners (biostimulants) in Europe. Bulg. J. Agric. Sci. 19, 1183–1189. Available online at: http://www.agrojournal.org/19/06-02.pdf; http://www.agrojournal.org/19/06-02.html
Torre, L. A., Battaglia, V., and Caradonia, F. (2016). An overview of the current plant biostimulant legislations in different European Member States. J. Sci. Food Agric. 96, 727–734. doi: 10.1002/jsfa.7358
Traon, D., Amat, L., Zotz, F., and du Jardin, P. (2014). A Legal Framework for Plant Biostimulants and Agronomic Fertiliser Additives in the EU. Report to the European Commission, DG Enterprise & Industry, Arcadia International, 115.
Trevisan, S., Francioso, O., Quaggiotti, S., and Nardi, S. (2010). Humic substances biological activity at the plant-soil interface from environmental aspects to molecular factors. Plant Signal. Behav. 5, 635–643. doi: 10.4161/psb.5.6.11211
Turan, M., and Köse, C. (2004). Seaweed extracts improve copper uptake of grapevine. Acta Agric. Scand. B 54, 213–220. doi: 10.1080/09064710410030311
Ugolini, L., Cinti, S., Righetti, L., Stefan, A., Matteo, R., D'Avino, L., et al. (2015). Production of an enzymatic protein hydrolyzate from defatted sunflower seed meal for potential application as a plant biostimulant. Ind. Crop Prod. 75, 15–23. doi: 10.1016/j.indcrop.2014.11.026
Ulrich-Merzenich, G., Panek, D., Zeitler, H., Wagner, H., and Vetter, H. (2009). New perspectives for synergy research with the “omic”-technologies. Phytomedicine 16, 495–508. doi: 10.1016/j.phymed.2009.04.001
Valepyn, E., Cabrera, J. C., Richel, A., and Paquot, M. (2014). Water soluble exo-polysaccharide from Syncephalastrum racemosum, a strong inducer of plant defence reactions. Carbohyd Polym. 101, 941–946. doi: 10.1016/j.carbpol.2013.10.018
Van der Watt, E., and Pretorius, J. C. (2011). In vitro and in vivo bio-stimulatory properties of a Lupinus albus L. seed suspension. Crop Pasture Sci. 62, 189–197. doi: 10.1071/CP10391
Van der Watt, E., and Pretorius, J. C. (2013). A triglyceride from Lupinus albus L. seed with biostimulatory properties. Afr. J. Biotechnol. 35, 5431–5443. doi: 10.5897/AJB12.2851
Van Regenmortel, M. H. (2004). Reductionism and complexity in molecular biology. EMBO Rep. 5, 1016–1020. doi: 10.1038/sj.embor.7400284
Vasconcelos, A. C. F., Zhang, X., Ervin, E. H., and Kiehl, J. C. (2009). Enzymatic antioxidant responses to biostimulants in maize and soybean subjected to drought. Sci. Agric. 66, 395–402. doi: 10.1590/S0103-90162009000300015
Vaskova, H., Kolomaznik, K., and Vasek, V. (2013). Hydrolysis process of collagen protein from tannery waste materials for production of biostimulator and its mathematical model. Int. J. Math. Mod. Methods Appl. Sci. 7, 568–575. Available online at: http://naun.org/cms.action?id=5358
Vera, J., Castro, J., Gonzalez, A., and Moenne, A. (2011). Seaweed polysaccharides and derived oligosaccharides stimulate defense responses and protection against pathogens in plants. Mar. Drugs 9, 2514–2525. doi: 10.3390/md9122514
Verkleij, F. N. (1992). Seaweed extracts in agriculture and horticulture: a review. Biol. Agric. Hortic. 8, 309–324. doi: 10.1080/01448765.1992.9754608
Vernieri, P., Borghesi, E., Ferrante, A., and Magnani, G. (2005). Application of biostimulants in floating system for improving rocket quality. J. Food Agric. Environ. 3, 86–88. Available online at: http://world-food.net/application-of-biostimulants-in-floating-system-for-improving-rocket-quality/
Vernieri, P., Borghesi, E., Tognoni, F., Serra, G., Ferrante, A., and Piagessi, A. (2006). Use of biostimulants for reducing nutrient solution concentration in floating system. Acta Hortic. 718, 477–484. doi: 10.17660/ActaHortic.2006.718.55
Veselá, M., and Friedrich, J. (2009). Amino acid and soluble protein cocktail from waste keratin hydrolysed by a fungal keratinase of Paecilomyces marquandii. Biotechnol. Bioprocess Eng. 14, 84–90. doi: 10.1007/s12257-008-0083-7
Vessey, J. K. (2003). Plant growth promoting rhizobacteria as biofertilizers. Plant Soil. 255, 571–586. doi: 10.1023/A:1026037216893
Vijayanand, N., Ramya, S. S., and Rathinavel, S. (2014). Potential of liquid extracts of Sargassum wightii on growth, biochemical and yield parameters of cluster bean plant. Asian Pac. J. Reprod. 3, 150–155. doi: 10.1016/S2305-0500(14)60019-1
Vinoth, S., Gurusaravanan, P., and Jayabalan, N. (2014). Optimization of somatic embryogenesis protocol in Lycopersicon esculentum L. using plant growth regulators and seaweed extracts. J. Appl. Phycol. 26, 1527–1537. doi: 10.1007/s10811-013-0151-z
Viriji, M. G. (2007). A Plant Derived Biostimulant Formulation and Method for Preparation Thereof. Patent No. WO2007052282 A1. Available online at: http://www.freepatentsonline.com/WO2007052282A1.html
Vyas, S., Guha, S., Bhattacharya, M., and Rao, I. U. (2009). Rapid regeneration of plants of Dendrobium lituiflorum Lindl. (Orchidaceae) by using banana extract. Sci Hortic. 121, 32–37. doi: 10.1016/j.scienta.2009.01.012
Wang, Y. H., and Irving, H. R. (2011). Developing a model of plant hormone interactions. Plant Signal. Behav. 6, 494–500. doi: 10.4161/psb.6.4.14558
Weissabach, H., King, W., Sjoerdsma, A., and Udenfriend, S. (1959). Formation of indole-3-acetic acid and tryptamine in animals: a method for estimation of indole-3-acetic acid in tissue. J. Biol. Chem. 234, 81–86.
Wilson, H. T., Xu, K., and Taylor, A. G. (2015). Transcriptome analysis of gelatin seed treatment as a biostimulant on cucumber plant growth. ScientificWorldJournal 2015:391234. doi: 10.1155/2015/391234
Wintermute, E. H., and Silver, P. A. (2010). Emergent cooperation in microbial metabolism. Mol. Syst. Biol. 6:407. doi: 10.1038/msb.2010.66
Wu, A. (2016). Hot Bio-Stimulants Gain Traction in Latin American market. Available online at: http://news.agropages.com/News/NewsDetail---19117.htm (Accessed August 25, 2016).
Wu, Y., Jenkins, T., Blunden, G., von Mende, N., and Hankins, S. D. (1998). Suppression of fecundity of the root-knot nematode, Meloidogyne javanica, in monoxenic cultures of Arabidopsis thaliana treated with an alkaline extract of Ascophyllum nodosum. J. Appl. Phycol. 10, 91–94. doi: 10.1023/A:1008067420092
Xavier, I. J., and Boyetchko, S. M. (2002). “Arbuscular mycorrhizal fungi as biostimulants and bioprotectants of crops,” in Applied Mycology and Biotechnology. Agriculture and Food Production, Vol. 2, eds G. G. Khachatourians and D. K. Arora (Amsterdam: Elsevier), 311–330.
Yakhin, I. A., Ibragimov, R. I., Yakhin, O. I., Isaev, R. F., and Vakhitov, V. A. (1998). The induced effect of biopreparation stifun on the accumulation of trypsin inhibitors in potato tubers during storage. Russ. Agric. Sci. 4, 12–13.
Yakhin, O. I., Lubyanov, A. A., and Yakhin, I. A. (2012). Changes in cytokinins, auxin, and abscisic acid contents in wheat seedlings treated with the growth regulator stifun. Russ. J. Plant Physiol. 59, 398–405. doi: 10.1134/S1021443712030193
Yakhin, O. I., Lubyanov, A. A., Yakhin, I. A., and Vakhitov, V. A. (2007). Protective role of bioregulator stifun under the negative effect of cadmium. Russ. Agric. Sci. 33, 233–235. doi: 10.3103/S1068367407040064
Yakhin, O. I., Lubyanov, A. A., Yakhin, I. A., Vakhitov, V. A., Ibragimov, R. I., Yumaguzhin, M. S., et al. (2011a). Metabolic changes in wheat (Triticum aestivum L.) plants under action of bioregulator stifun. Appl. Biochem. Micro. 47, 621–626. doi: 10.1134/S0003683811060123
Yakhin, O. I., Lubyanov, A. A., and Yakhin, I. A. (2014). Modern concepts on biostimulators. Agrokhimiya 7, 85–90. Available online at: http://elibrary.ru/item.asp?id=21779483
Yakhin, O. I., Lubyanov, A. A., and Yakhin, I. A. (2016a). Biostimulants in agrotechnologies: problems, solutions, outlook. Agrochemical Her. 1, 15–21. Available online at: http://www.agrochemv.ru/en/nomer/2016/1; http://elibrary.ru/item.asp?id=25940647
Yakhin, O. I., Lubyanov, A. A., and Yakhin, I. A. (2016b). Physiological activity and efficiency of application of biostimulants. Agrokhimya 6, 72–94. Available online at: http://elibrary.ru/item.asp?id=26470157
Yakhin, O. I., Lubyanov, A. A., Yakhin, I. A., Gareeva, G. B., Markelova, E. M., Kabirov, R. R., et al. (2013). Ecological evaluation of plant growth regulator stifun with the use of a multi-component test system. Agrokhimiya 3, 65–71. Available online at: http://elibrary.ru/item.asp?id=18962629
Yakhin, O. I., Lubyanov, A. A., Yakhin, I. A., Postrigan, B. N., Chemeris, A. V., Vakhitov, V. A., et al. (2011b). Effect of the plant growth regulator stifun on the accumulation of cadmium in grain crops. Agrokhimiya 5, 76–83. Available online at: http://elibrary.ru/item.asp?id=16356500
Yakhin, O. I., Yakhin, I. A., Spirikhin, L. V., and Khalilov, L. M. (2005). “Development of methods for standardization of multicomponent bioregulators based on vegetable raw materials,” in Mass Spectrometry and Its Application Problems. II Congress RSMS. Russian Conference with International Participation (Moscow), MBS-19.
Yakhin, O. I., Yakhin, I. A., Vakhitov, V. A., and Lubyanov, A. A. (2006). The mechanism of action of the natural bioregulator stifun. Dokl. Biochem Biophys. 411, 327–330. doi: 10.1134/S1607672906060019
Yamaguchi, Y., and Huffaker, A. (2011). Endogenous peptide elicitors in higher plants. Curr. Opin. Plant Biol. 14, 351–357. doi: 10.1016/j.pbi.2011.05.001
Yasmeen, A., Basra, S. M. A., Farooq, M., Rehman, H., Hussain, N., and Athar, H. R. (2013). Exogenous application of moringa leaf extract modulates the antioxidant enzyme system to improve wheat performance under saline conditions. Plant Growth Regul. 69, 225–233. doi: 10.1007/s10725-012-9764-5
Yasmeen, A., Nouman, W., Basra, S. M. A., Wahid, A., Rehman, H., Hussain, N., et al. (2014). Morphological and physiological response of tomato (Solanum lycopersicum L.) to natural and synthetic cytokinin sources: a comparative study. Acta Physiol. Plant. 36, 3147–3155. doi: 10.1007/s11738-014-1662-1
Zandonadi, D. B., Canellas, L. P., and Façanha, A. R. (2007). Indolacetic and humic acids induce lateral root development through a concerted plasmalemma and tonoplast H+ pumps activation. Planta 225, 1583–1595. doi: 10.1007/s00425-006-0454-2
Zandonadi, D. B., Santos, M. P., Dobbss, L. B., Olivares, F. L., Canellas, L. P., Binzel, M. L., et al. (2010). Nitric oxide mediates humic acids-induced root development and plasma membrane H+-ATPase activation. Planta 231, 1025–1036. doi: 10.1007/s00425-010-1106-0
Zhang, X., and Schmidt, R. E. (2000). Hormone containing products impact on antioxidant status of tall fescue and creeping bent grass subjected to drought. Crop Sci. 40, 1344–1349. doi: 10.2135/cropsci2000.4051344x
Zhao, J., Davis, L. C., and Verpoorte, R. (2005). Elicitor signal transduction leading to production of plant secondary metabolites. Biotechnol. Adv. 23, 283–333. doi: 10.1016/j.biotechadv.2005.01.003
Ziosi, V., Zandoli, R., Di Nardo, A., Biondi, S., Antognoni, F., and Calandriello, F. (2013). Biological activity of different botanical extracts as evaluated by means of an array of in vitro and in vivo bioassays. Acta Hortic. 1009, 69–66. doi: 10.17660/ActaHortic.2013.1009.5
Zodape, S. T. (2001). Seaweeds as a biofertilizer. J. Sci. Ind. Res. India. 60, 378–382. Available online at: http://nopr.niscair.res.in/handle/123456789/26485
Zodape, S. T., Gupta, A., Bhandari, S. C., Rawat, U. S., Cahudhary, D. R., Eswara, K., et al. (2011). Foliar application of seaweed sap as biostimulant for enhancement of yield and yield quality of tomato (Lycopersicon esculentum Mill.). J. Sci. Ind. Res. India. 70, 215–219. Available online at: http://nopr.niscair.res.in/handle/123456789/11089
Keywords: biostimulants, mode of action, definition, classification, regulation, concepts, methodology, emergent properties
Citation: Yakhin OI, Lubyanov AA, Yakhin IA and Brown PH (2017) Biostimulants in Plant Science: A Global Perspective. Front. Plant Sci. 7:2049. doi: 10.3389/fpls.2016.02049
Received: 27 October 2016; Accepted: 21 December 2016;
Published: 26 January 2017.
Edited by:
Stefania De Pascale, University of Naples Federico II, ItalyReviewed by:
Yuksel Tuzel, Ege University Faculty of Agriculture, TurkeyPatrick Du Jardin, Gembloux Agro-Bio Tech - University of Liège, Belgium
Copyright © 2017 Yakhin, Lubyanov, Yakhin and Brown. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Oleg I. Yakhin, eWFraGluQGFucmIucnU=