 Guang-hui Zhang
Guang-hui Zhang Ni-hao Jiang
Ni-hao Jiang Jun-wen Chen
Jun-wen Chen
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Plant Sci. , 16 August 2016
Sec. Plant Metabolism and Chemodiversity
Volume 7 - 2016 | https://doi.org/10.3389/fpls.2016.01209
This article is part of the Research Topic The world inside and outside phenylpropanoids View all 15 articles
Background: The medicinal herb, Pinellia ternata, is purported to be an anti-emetic with analgesic and sedative effects. Alkaloids are the main biologically active compounds in P. ternata, especially ephedrine that is a phenylpropylamino alkaloid specifically produced by Ephedra and Catha edulis. However, how ephedrine is synthesized in plants is uncertain. Only the phenylalanine ammonia lyase (PAL) and relevant genes in this pathway have been characterized. Genomic information of P. ternata is also unavailable.
Results: We analyzed the transcriptome of the tuber of P. ternata with the Illumina HiSeq™ 2000 sequencing platform. 66,813,052 high-quality reads were generated, and these reads were assembled de novo into 89,068 unigenes. Most known genes involved in benzoic acid biosynthesis were identified in the unigene dataset of P. ternata, and the expression patterns of some ephedrine biosynthesis-related genes were analyzed by reverse transcription quantitative real-time PCR (RT-qPCR). Also, 14,468 simple sequence repeats (SSRs) were identified from 12,000 unigenes. Twenty primer pairs for SSRs were randomly selected for the validation of their amplification effect.
Conclusion: RNA-seq data was used for the first time to provide a comprehensive gene information on P. ternata at the transcriptional level. These data will advance molecular genetics in this valuable medicinal plant.
Pinellia ternata (Thunb.) Berit. is an anti-emetic with analgesic and sedative effects, and has been applied as an antitussive and expectorant (He et al., 2005; Wang and Zhou, 2007; Zhang et al., 2007). The dried tuber of this herb, called “banxia” in Chinese, is in the top 10 most commonly used traditional Chinese medicines. The alkaloids isolated from the tubers of P. ternata are said to have anticancer properties (Wang and Zhou, 2007). P. ternata is widely distributed in China and other Asian countries. Due to overexploitation and lack of large-scale plantings, P. ternata sources are becoming increasingly scarce.
Secondary metabolites of P. ternata have been identified: alkaloids (main active ingredient; i.e., ephedrine), organic acids, volatile oils, sterols, and amino acids (Oshio et al., 1978; Masao, 1997; Ge and Wu, 2009). Ephedrine accumulates primarily in mature tubers and is of great interest to researchers (Wu et al., 1996; Xu et al., 2007). Ephedrine and other phenylpropylamino alkaloids, such as, (1S, 2S)-pseudoephedrine, (S)-cathinone, (1R, 2S)-norephedrine, and (1S, 2S)-pseudonorephedrine are particularly produced by plants in the genus Ephedra and by Catha edulis. The US Food and Drug Administration has banned ephedra-containing supplements. In addition, dietary supplements that contain ephedrine are illegal in the United States for its serious side effects. Despite these limitations, ephedrine still is listed on the WHO Model List of Essential Medicines, and has been used to prevent low blood pressure, asthma, narcolepsy, and obesity. This motivates efforts to increase the production of ephedrine in planta. Further, to obtain purified extracts of ephedrine from plant, it is paramount to understand the structurally-related metabolites on the ephedrine pathway. This knowledge will provide information on potentially unforeseen byproducts and on how intermediates on the pathway can be chemically separated by chromatographic techniques. Therefore, a complete understanding of the biosynthesis of phenylpropylamino alkaloids still needs to be fully elucidated (Groves et al., 2015).
Phenylpropylamino alkaloid biosynthesis begins with L-phenylalanine (Phe) (Hagel et al., 2011) which is deaminated by phenylalanine ammonia lyase (PAL) (Soerensen and Spenser, 1994). Recent studies suggest that Phe-derived benzoic acid is an intermediate in the formation of phenylpropylamino alkaloids (Krizevski et al., 2010), although the involvement of benzoyl-CoA or benzaldehyde cannot be ruled out. There are at least two possible pathways of Phe side-chain shortening in plant benzoic acid biosynthesis: β-oxidative and non-β-oxidative routes (Boatright et al., 2004). A proposed biosynthesis pathway for benzoic acid and ephedrine synthesis is depicted as in Figure 1 (Facchini, 2001; Long et al., 2009; Krizevski et al., 2012a,b).
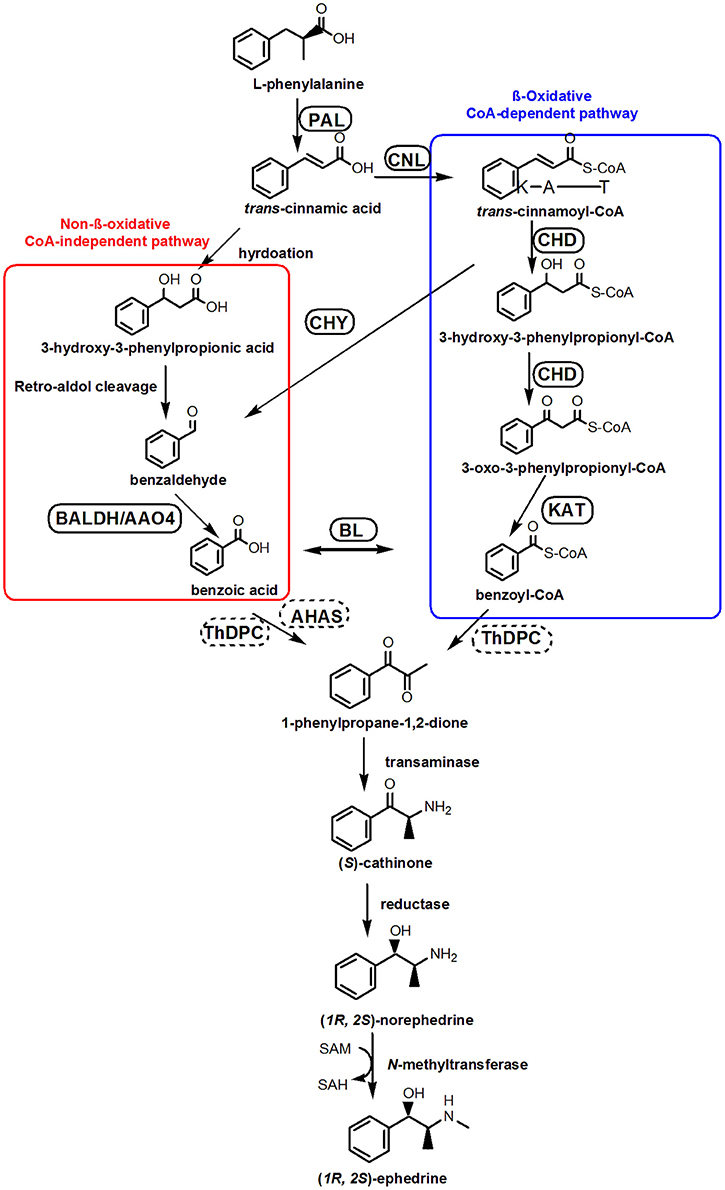
Figure 1. Proposed pathways for the biosynthesis of ephedrine in P. ternata. A CoA-independent, non-β-oxidative pathway of L-phenylalanine side-chain shortening is shown in red boxes, whereas a CoA-dependent, β-oxidative route is shown in blue boxes. Abbreviations: PAL, phenylalanine ammonia lyase; CNL, cinnamate:CoA ligase; CHD, cinnamoyl-CoA hydratase-dehydrogenase; CHY, 3-hydroxyisobutyryl-CoA hydrolase; AO4, aldehyde oxidases 4; KAT, 3-ketoacyl-CoA thiolase; BALDH, benzaldehyde dehydrogenase; BL, benzoate-CoA ligase; ThPDC, ThDP-dependent pyruvate decarboxylase; AHAS, acetolactate synthase.
In the β-oxidative pathway, the first step is cinnamoyl-CoA formation, catalyzed by cinnamate: CoA ligase (CNL) (Gaid et al., 2012; Klempien et al., 2012), which was also called as acyl activating enzyme (AAE) (Colquhoun et al., 2012). Subsequently, a bifunctional peroxisomal enzyme (cinnamoyl-CoA hydratase-dehydrogenase, CHD) converting cinnamoyl-CoA to 3-oxo-3-phenylpropanoyl-CoA, and 3-ketoacyl-CoA thiolase (KAT) catalyzing the formation of benzoyl-CoA (Figure 1; Van et al., 2009). In the non-β-oxidative pathway, two distinct dehydrogenase catalyzing the formation of benzoic acid from benzaldehyde have been characterized, and their genes, benzaldehyde dehydrogenase (BALDH) gene from Antirrhinum majus (Long et al., 2009) and aldehyde oxidases 4 (AO4) gene from Arabidopsis thaliana (Ibdah et al., 2009) have been cloned. After the formation of benzoic acid, the synthesis of phenylpropylamino alkaloid is initiated by condensation of pyruvic acid and benzoic acid to form 1-phenylpropane-1,2-dione (Figure 1). The enzyme that catalyzes this reaction has not been identified, but a ThDP-dependent pyruvate decarboxylase (ThPDC) or an acetolactate synthase (AHAS) have been suggested (Müller et al., 2009).
RNA-sequencing (RNA-seq) is a particularly effective technology for gene discovery, especially in non-model species for which reference genome sequences are not available. As mentioned above, the biosynthesis of ephedrine and other phenylpropylamino alkaloids is still largely unknown. Recently, candidate genes potentially involved in phenylpropylamino alkaloids biosynthesis in C. edulis and E. sinica were revealed using Illumina next-generation sequencing (NGS) (Groves et al., 2015). Here, we characterized the transcriptome of the tuber of P. ternata and identified candidate genes that encode enzymes in the ephedrine biosynthetic pathway. Based on the transcriptome sequences, SSR markers were predicted in P. ternata to facilitate molecular genetics in this valuable medicinal plant.
No specific permits were required for the described field studies. No specific permissions were required for these locations and activities. The location is not privately-owned or protected in any way and the field studies did not involve endangered or protected species.
P. ternata was cultivated in experimental fields at Purui Bio Pharmaceutical Co., Ltd., located in Shizong County, Yunnan province, southwest of China (24° 46′ 40″N, 104° 5′ 34″E, alt. 1886 m). P. ternata tubers ~1.5–2.0 cm in diameter were harvested from 1-year-old plants (Data Sheet 1: Figure S1). Tubers were collected and immediately frozen in liquid nitrogen and stored at −80°C until use.
Total RNA was extracted from the mature tubers using the Trizol Kit (Promega, USA) according to the manufacturer's instructions, and poly (A) mRNA was purified from 20 μg of total RNA using Oligo (dT) magnetic beads. Subsequently, mRNA was fragmented into smaller pieces (200–700 bp), which were used for first-strand cDNA synthesis with reverse transcriptase and random hexamer-primer. Subsequently, second-strand cDNA was synthesized using buffer, dNTPs, RNaseH, and DNA polymerase I. The short double-stranded cDNA fragments were purified with QiaQuick PCR extraction kit and resolved with EB buffer. These cDNA fragments underwent an end-repair process and poly(A) was added and then ligated with the Illumina paired-end sequencing adaptors. Ligation products were purified with magnetic beads and separated by agarose gel electrophoresis. A range of cDNA fragments (200 ± 25 bp) were excised from the gel and selected for PCR amplification as templates. The cDNA library was constructed with a fragment-length range of 200 bp (±25 bp). Finally, the cDNA libraries were sequenced on a paired-end flow cell using an Illumina HiSeq™ 2000 at Genedenovo Bio-Tech Co., Ltd (Guangzhou, China). The dataset of high-quality reads was deposited in a NCBI database under accession number SRX484200.
The image data output from the sequencing machine was transformed by base calling into sequence data (raw data/reads). Raw reads are transformed into clean reads by removing reads with sequencing adaptors; removing reads with frequency of unknown nucleotides above 5%; and removing low-quality reads (containing more than 50% bases with Q ≤ 20) using a custom Perl script. Transcripts de novo assembly was carried out using two short read assembly programs: Trinity (Grabherr et al., 2011) and Bridger (Chang et al., 2015). Clean reads were de novo assembled with Trinity with the fixed default k-mer size of 25. Trinity initially combines reads with certain length of overlap to form longer fragments without N (using “N” to represent unknown sequences), or contigs. Then, contigs are processed with sequence clustering software TIGR Gene Indices clustering tools (TGICL) (Pertea et al., 2003) to form longer sequences without N and these sequences are defined as unigenes.
Finally, all assembled unigenes were searched using BLASTX against protein databases, such as non-redundant (NR) protein database (http://www.ncbi.nlm.nih.gov/), Swiss-Prot database (http://www.expasy.ch/sprot), the Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway database (http://www.genome.jp/kegg) (Kanehisa et al., 2006), and the Cluster of Orthologous Groups (COG) (Tatusov et al., 1997) database (http://www.ncbi.nlm.nih.gov/COG/), with an E-value cutoff of 1e-5. The best match from the four databases was used to decide unigene sequence direction. If database information was conflicting, a priority order of NR, SwissProt, KEGG, and COG was followed when deciding unigene sequence direction. When a unigene would not align to any database, unigene orientation was predicted using ESTScan (Iseli et al., 1999). We also plotted the ratio of assembled unigene length to Oryza sativa ortholog length against coverage depth for assessing the extent of transcript coverage provided by unigenes and to evaluate how coverage depth affected unigenes assembly. Firstly, we compared the sequences of all unigenes by BLASTX against the Nr database and found O. sativa is one of the top-hit species, which is also monocot as same as P. ternata. The Oryza sativa orthologs, coverage, and their CDS regions were also obtained in NCBI.
In functional annotation, all assembled unigenes were searched against the NR, Swiss-Prot, KEGG, and COG databases using BLASTN (E < 10−5) to predict possible functional classifications and molecular pathways. All unigenes were annotated by BLASTX (E < 10−5) against the Arabidopsis TAIR10 peptide database (Swarbreck et al., 2008). Moreover, the conserved domains and families of the assembled unigenes encoding proteins were searched against the Pfam database (version 26.0) (http://pfam.xfam.org/) using the Pfam_Scan program (Finn et al., 2014). To obtain the final functional annotation of the unigenes, the best annotation was chosen based on the BLASTX scores (Camacho et al., 2009). If a unigene did not have annotations in any of the above-mentioned databases, Pfam annotation was assigned to the unigene. Based on the results from NR database annotation, the Blast2GO program (Conesa et al., 2005) was used to obtain GO unigene annotations. Then, WEGO software (Ye et al., 2006) was used to perform GO functional classification for all unigenes to view gene function distribution.
The unigenes coding sequence (CDS) was predicted by Blastx and ESTscan. The unigene sequences were first aligned with the protein databases using BLASTX (E < 10−5) in the following order: NR, SwissProt, KEGG, and COG. Unigenes aligned to a higher priority database were not aligned to lower priority databases. The best alignment results were used to determine the unigenes sequence directions. Unigene oorientation and CDS with no hits in Blast were predicted using ESTScan (Iseli et al., 1999). To identify the transcription factors, all unigenes were searched against PlnTFDB database (Pérez-Rodríguez et al., 2010) using iTAK analysis tool (http://bioinfo.bti.cornell.edu/cgi-bin/itak/index.cgi) (Dai et al., 2013).
Total RNA from tubers and leaves of P. ternata were extracted individually using Trizol Kit (Promega, USA) following the manufacturer's protocol. Subsequently, RNA was treated with 4 × gDNA wiperMix at 42°C for 2 min to remove DNA. The purified RNA (1 ug) was reverse transcribed to cDNA using HiScript QRT SuperMix for qPCR (Vazyme, Nanjing, China). The qPCR reactions were performed in a 20 μl volume composed of 2 μl of cDNA, 0.4 μl of each primer, and 10 μl 2 × SYBR Green Master mix (TaKaRa) in Roche LightCycler 2.0 system (Roche Applied Science, Branford, CT). PCR amplification was performed under the following conditions: 3 min at 94°C, followed by 45 cycles of 94°C for 20 s, 55°C for 20 s, and 72°C for 20 s. Three technical replications were performed for all quantitative PCRs. The glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was chosen as reference gene control for normalization. The relative changes in gene expression levels were calculated using the 2−ΔΔCt method (Livak and Schmittgen, 2001). The log2 value of 2−ΔΔCt was used for representing the relative expressions of each gene. All primers used for the reverse transcription quantitative real-time PCR (RT-qPCR) assay are listed in Data Sheet 1: Table S1.
Simple sequence repeats (SSRs) were identified using the MIcroSAtellite Identification Tool (http://pgrc.ipk-gatersleben.de/misa/misa.html). Parameters were designed for identifying di-, tri-, tetra-, penta- and hexa-nucleotide motifs with a minimum of 6, 5, 4, 4, and 4 repeats, respectively (Zeng et al., 2010). A maximum distance of 100 nucleotides was allowed between two SSRs. Primer3 (http://primer3.ut.ee/) was employed to design PCR primers flanking each unique SSR region that was identified.
A total of 20 primer pairs (Data Sheet 1: Table S2) were randomly selected to evaluate their amplification effect. The method was performed as our previously described (Jiang et al., 2014), but with the following modification: The PCR reactions were performed at 94°C for 5 min, and followed by 30 cycles of 1 min at 94°C, 50 s at Tm (annealing temperature), 90 s at 72°C and a final step at 72°C for 10 min.
To obtain an overview of the transcriptome of P. ternata, a cDNA library was generated from total RNA of mature tubers, and pair-end sequenced using the Illumina HiSeq™ 2000 sequencing platform. After the removal of adaptor, sequences, ambiguous reads, and low-quality reads (Q20 < 20), a total of 66,813,052 clean reads (total length of 6,681,305,200; 6.7 Gb) nucleotides were obtained. The Q20 (sequencing error rate < 1%) and GC percentages were 95.21 and 53.04%, respectively (Table 1).
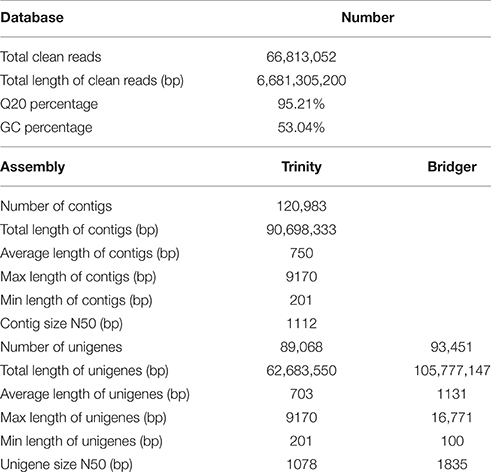
Table 1. Summary of Illumina Paired-end sequencing and assembly for P. ternata.
Because no reference genome exists for P. ternata, the reads were assembled de novo. Using the Trinity assembling program, clean reads were assembled into 120,983 contigs ranging from 201 to 9170 bp and with a mean length of 750 bp and an N50 length of 1112 bp. Among these contigs, 61,268 (50.64%) were longer than 500 bp, and 15,781 (13.04%) were longer than 1000 bp. Using paired-end joining and gap-filling methods, the contigs were further assembled into 89,068 unigenes with an average length of 703 bp and an N50 length of 1078 bp (Table 1). Among all unigenes, 19,898 (22.34%) were longer than 1000 bp, and 48,673 (54.64%) were less than 500 bp. In this study, we obtained a total of 51,642 CDSs and 9902 CDSs (19.17%) were longer than 1000 bp and 22,499 CDSs (43.57%) exceeded 500 bp. The length distributions of contigs, unigenes and CDSs are depicted in Figure 2 and the sequences of all unigenes assembled by Trinity are shown in Data Sheet 2.
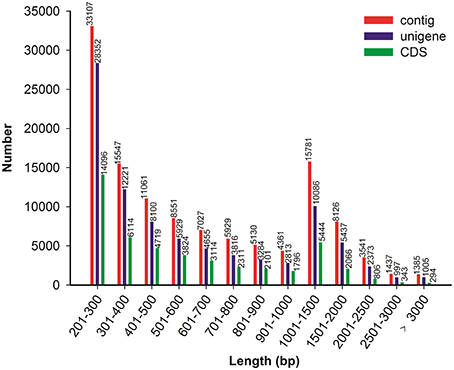
Figure 2. Overview of the P. ternata transcripts assembly and the length distribution of the CDS.
Moreover, a new de novo transcriptome assembler, Bridger (version: r2014-12-01), was also used for assembly in our study (Chang et al., 2015). Compared with Trinity, more unigenes (93,451) with longer average length (1131 bp) and N50 length (1835 bp) were obtained by Bridger (Table 1). The sequences of all unigenes assembled by Bridger are shown in Data Sheet 3. Bridger can assemble more full-length reference transcripts, and reducing false positive transcripts in comparison with the state-of-the-art assemblers (Chang et al., 2015). Our results also indicated that Bridger is better than Trinity in transcriptome assembly in non-model plant, and the unigenes with longer average length will be helpful in cloning the full-length of cDNA and functional characterization of the genes. Nevertheless, Trinity is also a good assembler for transcriptome assembly from RNA-seq data without a reference genome (Grabherr et al., 2011; Bankar et al., 2015); the assembled results of Trinity are used for further analysis in this study.
To evaluate the quality and coverage of the assembled unigenes, all usable sequencing reads were realigned to the unigenes using SOAPaligner (Li et al., 2008), allowing up to 2 base mismatches. The sequencing depth ranged from 0.025 to 52,968-fold, with an average of 31.96-fold. About 62.17% of the unigenes were realigned by more than 10 reads, 22.87% were supported by more than 100 reads and 5.04% were supported by more than 1000 reads (Data Sheet 1: Figure S2). To assess the extent of transcript coverage provided by unigenes and to evaluate how coverage depth affected unigenes assembly, we plotted the ratio of assembled unigene length to Oryza sativa ortholog length against coverage depth (Data Sheet 1: Figure S3A). Although many of deeply covered P. ternata unigenes failed to cover the complete coding regions of their O. sativa orthologs, our unigenes covered most O. sativa ortholog coding regions. In our study, 2206 unigenes had ratios greater than 1, and 18,913 unigenes had ratios less than 1. Of note, to certain extent, increased coverage depth can result in higher coverage of the coding regions. The percentage of O. sativa ortholog coding sequences covered by all P. ternata unigenes was also measured and 3195 of the orthologs were covered by more than 80% of the unigenes and 2285 of the orthologs were covered by 40–80% of the unigenes. Around 45% of the orthologs were covered by only 20% or less (Data Sheet 1: Figure S3B) indicating that additional sequencing is essential for a more comprehensive coverage of the transcriptome of P. ternata.
To provide putative annotations of the assembled unigenes, all of the unigenes was subjected to a BLASTN search against the public protein databases (NR, Swiss-Prot, KEGG, COG) with E < 10−5. The unigenes were also searched against the Arabidopsis TAIR10 peptide database using the BLASTX algorithm with E < 10−5. Using this approach, ~53.33% of unigenes (47,504) were annotated in the five public databases (Data Sheet 1: Table S3). Among them, 4248 unigenes had significant matches in all five databases, 9753 unigenes annotated uniquely in NR database, 182 unigenes annotated uniquely in Swiss-Prot database, 8 unigenes annotated uniquely in COG database, 46 unigenes annotated uniquely in KEGG database, and there were no unigenes annotated uniquely in TAIR10 database (Data Sheet 1: Figure S4). The annotations of unigenes in all five databases are shown in Data Sheet 4.
Previous studies indicate that the longer sequences were more likely to obtain BLAST matches in the protein databases (Wang et al., 2010; Li et al., 2012; Liu et al., 2013), and this assumption was validated by our data which indicated that more than 83% of the unigenes larger than 1000 bp in length had BLAST matches in NR and Swiss-Prot protein databases. In contrast, only ~30% of unigenes shorter than 500 bp did (Data Sheet 1: Figure S5). The E-value distribution of the top hits in the NR database revealed that 46.11% of the mapped sequences had significant homology (E < 1e−50), and 17.80% of the sequences with greater than 80% similarity were found (Data Sheet 1: Figures S6A,C). The E-value and similarity distributions of the top hits in the Swiss-Prot database had comparable patterns with 36.41 and 15% of the sequences possessing significant homology and similarity, respectively (Data Sheet 1: Figures S6B,D). Our results also indicated that 35.79% of the unigenes had significant homology with sequences of Vitis vinifera (9011, 19.07%), followed by Theobroma cacao (6824, 14.44%), O. sativa (6158, 13.03%), and Setaria italica (3124, 6.61%) (Data Sheet 1: Figure S7). This suggests that the genome of P. ternata is more closely related to V. vinifera than to other model plant genomes.
Based on the NR annotation, Gene Ontology (GO) classification was used to classify the functions of all unigenes. Based on sequence homology, 16,671 unigenes were assigned to one or more ontologies, including 35,061 unigenes at the cellular component, 29,454 unigenes at the biological process, and 17,857 sequences at the molecular function (Figure 3). Within the cellular component category, cell (10,728, 30.60%), cell parts (10,728, 30.60%), and organelles (8457, 24.12%) represented the majorities. Under the biological process category, metabolic (7950, 26.99%) and cellular processes (7362, 24.99%) and response to stimulus processes (2959, 10.05%) were the most highly represented groups. Under the molecular function category, catalytic activity (8235, 46.12%) and binding (7816, 43.77%) were the most highly represented GO terms (Data Sheet 1: Table S4).
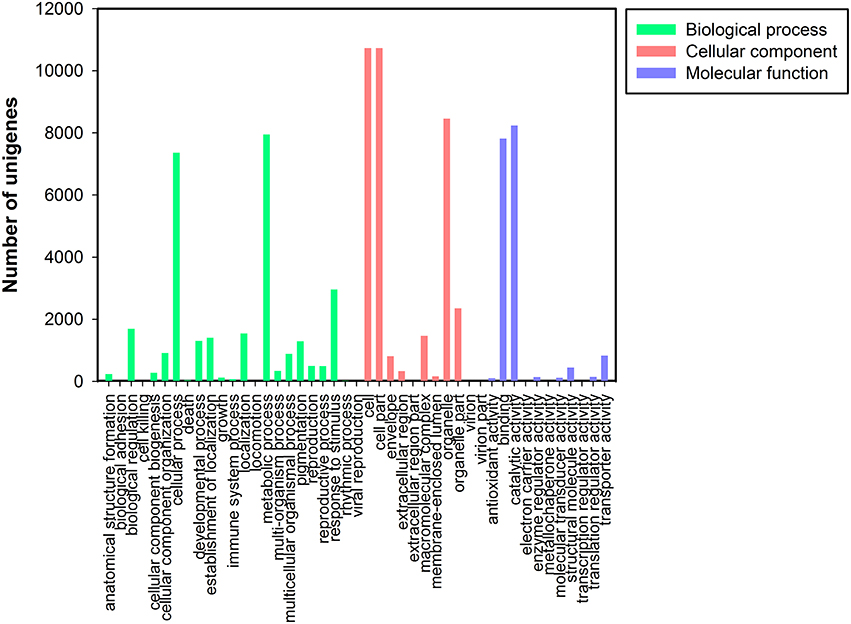
Figure 3. Gene Ontology classification of assembled unigenes. Total 16,671 unigenes were categorized into three main categories: biological process, cellular component, and molecular function.
The conserved domains/families of the assembled unigenes encoding proteins were searched against the Pfam database (version 26.0) (Finn et al., 2014) using Pfam_Scan program. A total of 3601 protein domains were identified in 29,909 unigenes of P. ternata (Data Sheet 5). Among these protein domains/families, the pentatricopeptide repeat domain (PPR) was the most abundant domain type, found in 3204 unigenes. PPR-containing proteins are commonly found in plants and although their functions are still unclear, the PPR domain has been reported to exist in proteins involved in RNA editing (Fujii and Small, 2011; Shikanai and Okuda, 2011). Moreover, highly represented domains were the WD40 domain (1015 unigenes) and leucine-rich repeats (949 unigenes), which are primarily involved in protein-protein interactions (Kobe and Kajava, 2001). Then, a protein kinase domain (901 unigenes) was predicted in the derived transcriptomic sequences of P. ternata, which is involved in signal transduction pathways, development, cell division, and metabolism in higher organisms (Hanks and Quinn, 1991; Hanks and Hunter, 1995; Ahier et al., 2009). Other domains identified as being abundant included RNA recognition motifs (608 unigenes), protein tyrosine kinase (397 unigenes), reverse transcriptase (374 unigenes), mitochondrial carrier proteins (272 unigenes), and cytochrome P450s (271 unigenes). The 15 most abundant protein domains/families are represented in Data Sheet 1: Figure S8.
To further predict gene function and to evaluate the completeness of the transcriptome library of P. ternata, all unigenes were subjected to a search against the COG database for functional prediction and classification. In total, 50,534 unigenes were annotated and grouped into 25 COG classifications (Figure 4). Among the 25 COG categories, the largest cluster was for general function prediction (6282, 12.43%), and this was followed by translation, ribosomal structure and biogenesis(4726, 9.35%), transcription (4661, 9.22%), unknown functions (4347, 8.60%), replication, recombination and repair (4009, 7.93%), and cell cycle control, cell division, and chromosome partitioning (3537, 7.00%), posttranslational modification, protein turnover, chaperones (3285, 6.50%), and signal transduction mechanisms (2955, 5.85%), and finally cell wall/membrane/envelope biogenesis (2878, 5.70%).
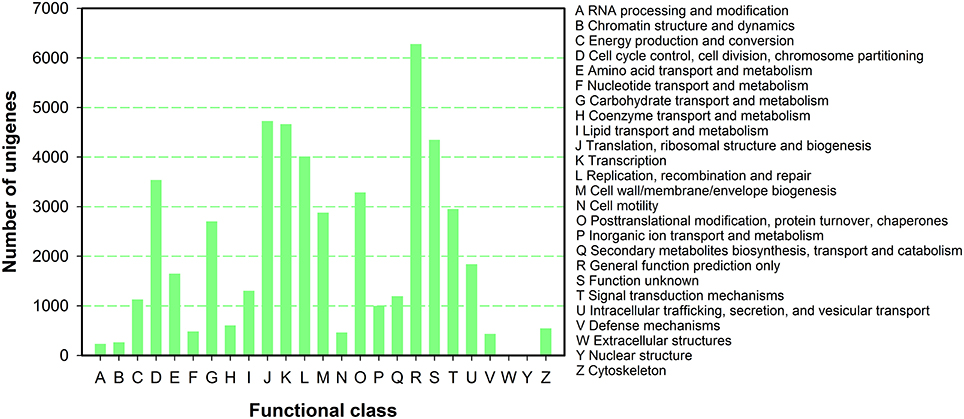
Figure 4. COG function classification of P. ternata unigenes.
To identify active biochemical pathways in P. ternata, unigenes were mapped to typical pathways in KEGG. Pathway-based analysis can help us understand the biological functions of genes. Based on a comparison against the KEGG database, a total of 13,899 unigenes (15.60%) were annotated in KEGG and 18,927 unigenes (21.25%) were assigned to 126 KEGG pathways (Data Sheet 1: Table S5). Among them, the metabolic pathway containing 3612 unigenes is the largest one, followed by biosynthesis of secondary metabolites (1673 unigenes), the ribosome (592 unigenes), RNA transport (590 unigenes), the spliceosome (441 unigenes), an mRNA surveillance pathway (394 unigenes), and plant-pathogen interaction (376 unigenes). The tubers grown in soil are more prone to pathogenic attack, so P. ternata needs an adequate supply of secondary metabolites to produce an adequate pathogen defense. Thus, 376 unigenes were assigned to the plant-pathogen interaction term as expected. In the metabolic pathway, the most represented subcategories were carbohydrate metabolism (1943 unigenes), followed by metabolism of amino acids (1107 unigenes), lipids (1065 unigenes), energy (900 unigenes), nucleotides (635 unigenes), cofactors and vitamins (516 unigenes), other amino acids (351 unigenes), terpenoids and polyketides (300 unigenes), and glycan biosynthesis and metabolism (298 unigenes), as well as biosynthesis of others secondary metabolites (291 unigenes) (Figure 5A). In the amino acid metabolism category, 14 subcategories comprised 1107 unigenes, the most represented categories were for metabolism of arginine and proline (156 unigenes), cysteine and methionine (151 unigenes), glycine, serine and threonine (125 unigenes), aspartate and glutamate (114 unigenes), and valine, leucine and isoleucine degradation (101 unigenes) (Figure 5B).
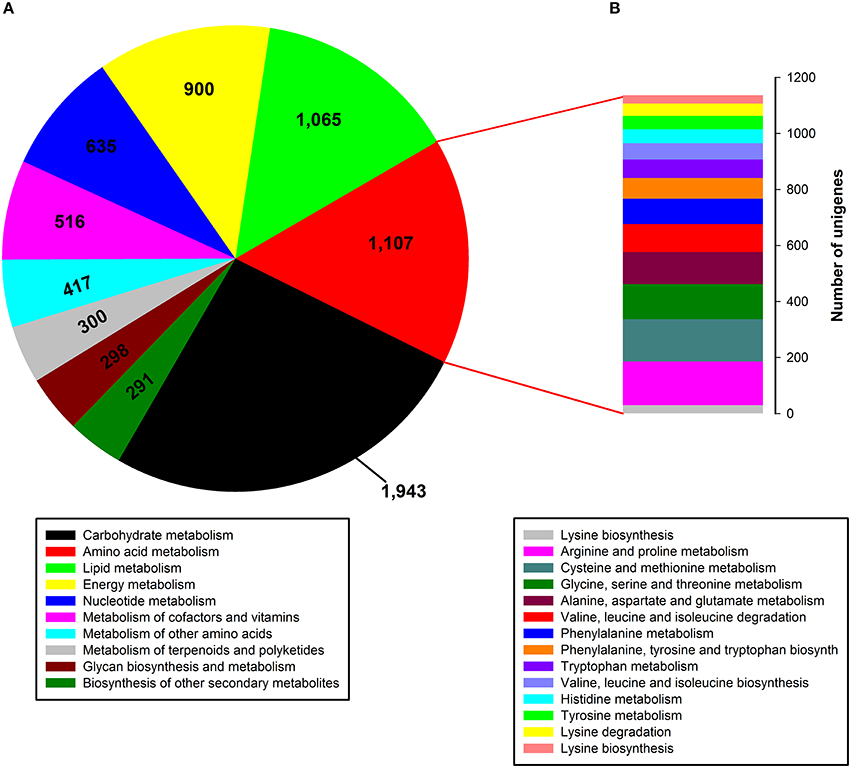
Figure 5. Pathway assignment based on KEGG. (A) Classification based on metabolism categories; (B) Classification based on amino acid metabolism categories.
Here, we focused on the discovery of genes involved in ephedrine and its precursor benzoic acid biosynthesis. Ephedrine biosynthesis in plants begins with L-phenylalanine, which is converted to trans-cinnamic acid by PAL. In E. sinica, at least four isoforms of PAL genes exist, and their expression in roots was higher than in aerial plant components (Okada et al., 2008). Similarly, in the transcriptome dataset of P. ternata, 6 unigenes annotated to PAL gene were identified (Table 2; Data Sheet 6).
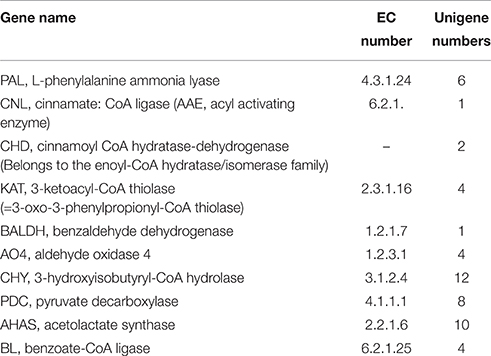
Table 2. Transcripts involved in ephedrine and benzoic acid biosynthesis in P. ternata.
Biosynthesis of benzoic acid from L-phenylalanine requires shortening of the side chain by two carbons, which can occur via the β-oxidative or non-β-oxidative pathways. The transcripts encoding enzymes in both of β-oxidative and non-β-oxidative pathways are found in the transcriptome dataset of P. ternata. One unique sequence was identified as with 74% similarity to Hypericum calycinum CNL, and four were annotated to the KAT gene (Table 2; Data Sheet 6). No unigene annotated to CHD gene was found, but 2 unigenes belong to the enoyl-CoA hydratase/isomerase family and showed high similarity (>75%) to petunia CHD gene (Qualley et al., 2012), suggesting that those might be the CHD candidate gene of P. ternata (Figure 6).

Figure 6. Alignment of amino acid sequences of the putative P. ternata CHDs with Petunia hybrid CHD (AFS41246.1). Identical amino acid residues are shaded in dark blue. Light blue shade indicates 50% or more identity among all the aligned sequences.
In this study, 1 unigene was identified as having 66% similarity to the BALDH gene of A. thaliana, and 5 unigenes were annotated to the AO4 gene. Interestingly, 12 unigenes were annotated to CHY genes, which encoded 3-hydroxyisobutyryl-CoA hydrolase for catalyzing the conversion of cinnamoyl-CoA to benzaldehyde (Ibdah and Pichersky, 2009), suggested that benzaldehyde might come from cinnamoyl-CoA in P. ternata. Benzoate-CoA ligase (BL) catalyzes the formation of benzoyl-CoA from benzoate and CoA (Kliebenstein et al., 2007; Ibdah et al., 2009). In P. ternata transcriptome database, no unigene was annotated to BL, but we found 4 ungenes (unigene0092667, 0089644, 0080607, and 0044033) have close relationship to Arabidopsis thaliana BL (Genbank no. NP_176763.1) with the identities of 45–51%, respectively. The similar result was also found in C. edulis and E. sinica (Groves et al., 2015) (Table 2; Data Sheet 6). To the best of our knowledge, these putative BALDH, AO4, CHY and BL genes are first reported in P. ternata. Genes in the β-oxidative and non-oxidative pathways were identified, and we propose that benzoic acid biosynthesis in P. ternata likely occurs via both of these pathways.
ThPDC or AHAS might catalyze the condensation of pyruvic acid and benzoic acid to form 1-phenylpropane-1,2-dione (Müller et al., 2009), in the transcriptome of P. ternata, we identified 8 unigenes that may encode the PDC gene, and 10 unigenes were annotated as candidate AHAS genes Table 2; Data Sheet 6). After the formation of 1-phenylpropane-1,2-dione intermediate, there are three enzymes involved in ephedrine biosynthesis, viz. transaminases, reductases and N-methyltransferases (Figure 1). Up to now, no such enzymes and relevant genes in phenylpropylamino alkaloids biosynthesis were functionally characterized; only some candidates were predicted based on phylogenetic analysis (Groves et al., 2015). In C. edulis and E. sinica transcriptomes, the transaminases candidates closely related to aromatic amino acid transaminases or prokaryotic-type amino transferases (Groves et al., 2015). Unlikely, in P. ternata transcriptome database, 14 unigenes were annotated to transaminase, among them, 8 unigenes were annotated to alanine-glyoxylate transaminase, 4 unigenes were annotated to alanine transaminase, and 2 were annotated to ornithine-oxo-acid transaminase (Data Sheet 7). This result suggested the complexity of transaminase in phenylpropylamino alkaloids-producing plants. Moreover, 265 unigenes annotated to reductase (Data Sheet 8) and 211 unigenes annotated to N-methyltransferase (Data Sheet 9) were also found in P. ternata transcriptome database. Those results indicated that identifying the transaminase, reductase and N-methyltransferase in phenylpropylamino alkaloids biosynthesis should be very difficult and further research is needed.
The expression level of ephedrine related genes in the leaves and tubers of P. ternata were analyzed by RT-qPCR. The results showed that these genes exhibited different expression level in the leaves and tubers (Figure 7). All these genes were more highly expressed in the leaves than in tubers. Due to lack of detailed information concerning the downstream steps of ephedrine biosynthesis, the genes selected for RT-qPCR analysis were all located at the upstream of ephedrine biosynthesis. Therefore, our results were not unexpected, and it indicated that leaves may be the main organ for synthesizing the precursors of ephedrine. It is noteworthy that the transcript abundance of the AHAS gene was higher in the tubers as compared to that of PDC gene. Obviously, AHAS appears to have greater activity in the tubers. Although no more evidence is presently available, we suggested that AHAS was the major enzyme responsible for the biosynthesis of 1-phenylpropane-1,2-dione in P. ternata. We believe that these data will be helpful to further understand the mechanism of ephedrine biosynthesis.
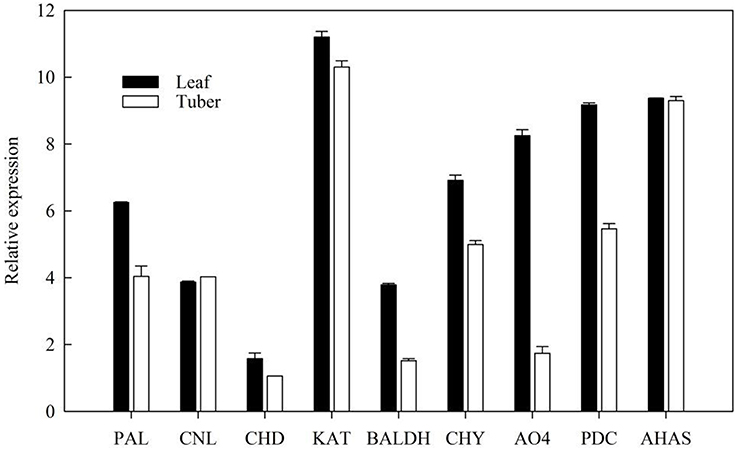
Figure 7. Validation of candidate P. ternata unigenes involved in ephedrine biosynthesis by RT-qPCR. Bars represent the mean (± SD) of three experiments.
Transcription factors (TFs) play a crucial role in regulating the secondary metabolism, because they can regulate the expression of related genes at the transcriptional level to control the flux of secondary metabolites. The identification of such transcription factors will help us gain a better understanding of gene regulatory networks. Based on the BLASTX search against the Plant Transcription Factor Database database (PlnTFDB) (Pérez-Rodríguez et al., 2010), 1218 unigenes were annotated in 848 independent coding sequences of plant transcription factors (identity >80%) that belong to 68 known transcription factor families (Table 3 and Data Sheet 10). Among them, 79, 68, 67, 64, 57, and 53 unigenes were annotated to the AP2-EREBP, bHLH, HB, MYB, C3H, and WRKY families, respectively.
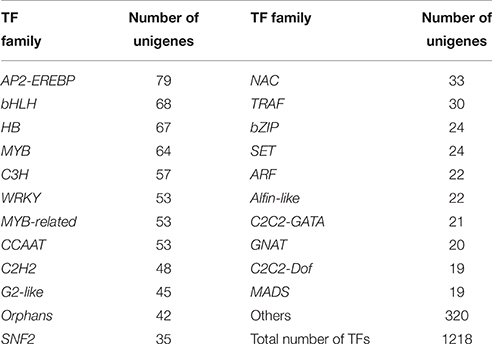
Table 3. Statistics for putative transcription factors in P. ternata unigenes.
Some TFs are essential for alkaloid biosynthesis in plants. AP2-EREBP is a large super family of TFs, and it can be distinguished by containing one or two AP2/ERF DNA binding domain (Riechmann and Meyerowitz, 1998). In Catharanthus roseus, the AP2/ERF-domain transcription factor ORCA2 and ORCA3 in turn regulate a subset of alkaloid biosynthesis genes (van der Fits and Memelink, 2000; Zhang et al., 2011; Guo et al., 2013). In our study, 79 unigenes encoding AP2-EREBP transcription factors were found. Meanwhile, the CrMYC2 transcription factor, a member of the bHLH family from C. roseus, may play a functional role in the regulation of expression of ORCA genes (Zhang et al., 2011). Besides, isoquinoline alkaloid biosynthesis in Coptis japonica is regulated by a unique bHLH-type transcription factor, CjbHLH1 (Yamada et al., 2011). In present study, a total of 68 unigenes encoding bHLH TFs were identified. The strictosidine synthase (STR) contributes to the forming of strictosidine, the central intermediate leading to all monoterpenoid indole alkaloids. A CrBPF1 transcription factor is similar to the MYB-like factor BPF1, and it appears to enhance elicitor-mediated STR1 gene expression (Facchini and De Luca, 2008). In our database, 64 unigenes were annotated as MYB transcription factors. In C. roseus, CrGBF1, and CrGBF2 are members of the bZIP TFs and participate in the regulation of expression of STR gene (Sibéril et al., 2001). A total of 24 unigenes coded for bZIP TFs have been identified. Transcription factors identified here may facilitate the research on the regulation of alkaloid biosynthesis in P. ternata.
SSRs are useful molecular genetic markers because of their relative abundance, and they have been widely applied for molecular-assisted selection (MAS) in plant breeding programs. Potential SSRs were detected in all of the 89,068 assembled unigenes using MISA software. A total of 14,468 SSRs were identified in 12,000 unigenes (Data Sheet 1: Table S6). Of all SSR-containing unigenes, 2053 sequences contained more than one SSR and 824 SSRs were present in compound form. SSRs derived from all unigenes are shown in Data Sheet 11. On average, we found 1 SSR per 19 Kb, which is similar to the frequency in cotton (1 SSR per 20 KB) (Cardle et al., 2000). Among the SSRs, the di-nucleotide repeat motifs were the most abundant types. SSRs with five tandem repeats were the most common (Data Sheet 1: Table S7). The most common type of di-nucleotide was AG/CT which accounted for 41.54% of the repeats, followed by AT/AT (5.04%) and AC/GT (4.29%). Among the tri-nucleotide repeats, both of CCG/CGG was the most frequent motifs (11.84%) (Data Sheet 1: Figure S9). Using Primer3, a total of 13,644 pairs of primers (Data Sheet 12) was designed and these SSRs could serve the foundation for future molecular breeding and genetic applications in this herb. To evaluate the amplification effect of these primer pairs, 20 primer pairs were randomly selected for the test. A total of 13 (65%) primer pairs successfully amplified the clear and repeatable bands, and 7 (35%) pair primers failed to generate PCR products. Among the 13 successful primer pairs, 10 primer pairs produced PCR amplicons at the expected size. However, 3 primer pairs generated PCR products longer than expected, and this may be due to the presence of introns. We believed that these unigene-derived SSR markers identified in our research will facilitate molecular genetics and molecular breeding in this valuable medicinal plant.
In this study we performed RNA sequencing and report a de novo assembly of an important medicinal herb in China, Pinellia ternata, which particularly accumulates ephedrine in the tubers. In total, 89,068 unigenes were obtained with an average length of 703 bp and an N50 length of 1078 bp and we obtained a total of 51,642 CDSs and 9902 CDSs (19.17%) were longer than 1000 bp and 22,499 CDSs (43.57%) exceeded 500 bp.
NGS does not require a reference genome to gain the useful transcriptomic information, making this technology particularly useful for non-model organisms that often lack genomic sequence data (Strickler et al., 2012). In this study, the assembly results indicated that the length distribution pattern and mean contig and unigene lengths were similar to those in the previous studies of Illumina-transcriptome (Hao et al., 2011; Huang et al., 2012; Shu et al., 2013), suggesting that transcriptome sequencing data from P. ternata were effectively assembled. To our knowledge, this is the first report of large-scale transcriptome sequencing and analysis in P. ternata and these data offer abundant genomic information for P. ternata. In all of 89,068 unigenes, ~53.33% of unigenes (47,504) were annotated in the public databases. Furthermore, about 46.67% of unigenes (41,564) could not be matched to known genes, indicating that there is limited information about the genomes or transcriptomes of P. ternata and its related species.
In functional classification by KEGG, we found 91 and 71 unigenes were assigned to phenylalanine metabolism and phenylalanine, tyrosine and tryptophan biosynthesis, respectively (Figure 5B). The phenylalanine, tyrosine and tryptophan biosynthesis pathway is involved in L-phenylalanine synthesis, as a precursor common phenylpropylamino alkaloid biosynthesis. The phenylalanine metabolism pathway is involved in making 1-phenylpropane-1, 2-dione, as a putative precursor in the formation of phenylpropylamino alkaloids in plants (Krizevski et al., 2007). KEGG annotation data offer a valuable resource for investigating specific processes, functions, and pathways that will guide P. ternata research.
It is believed that benzoic acid is the intermediate in the formation of phenylpropylamino alkaloids (Krizevski et al., 2010), so the genes in benzoic acid biosynthesis might also be involved in the biosynthesis of phenylpropylamino alkaloids. In P. ternata transcriptome database, the transcripts encoding enzymes involved in benzoic acid biosynthesis were found in this study, included both in β-oxidative or non-β-oxidative pathways, and two genes (CHY and BL) connecting the two pathways. But the main pathway and the functions of those genes in phenylpropylamino alkaloids biosynthesis are still far from elucidated. RT-qPCR has shown that the relative expressions of the genes involved in benzoic acid biosynthesis are highly expressed in the leaves, indicating that leaves may be the main organ for synthesizing the precursors of ephedrine. We also predicted that AHAS might be the major enzyme involved in 1-phenylpropane-1,2-dione biosynthesis in P. ternata.
For the first time, this study provides the comprehensive data regarding the transcriptome of P. ternata and establishes P. ternata as an ideal herb for investigating ephedrine biosynthesis. We identified several candidate genes involved in ephedrine biosynthesis and these data can facilitate the study of molecular mechanisms of ephedrine synthesis as well as the engineering of microorganisms for de novo production of similar active ingredients. Our data can also be useful for molecular genetic research or genetic engineering as it represents the most abundant genetic resource for P. ternata, and will serve as a foundation for other functional genomics of P. ternata or closely related species.
Phenylpropylamino alkaloids are only produced in a few plant species. There is a lack of attention to their biosynthesis and only some candidate genes were predicted (Groves et al., 2015). This study also provided candidate genes in ephedrine biosynthesis in P. ternata. But the functions of those genes should be characterized in the future. Furthermore, the relationships between gene expression profiles and natural abundance of phenylpropylamino alkaloids also needs to be evaluated; this will help us understand the concentrations of pathway intermediates under steady-state flux and the accumulation phenylpropylamino alkaloid end-products.
GZ and NJ designed the experiment, prepared samples for RNA-seq, and analyzed the data. WS and CM helped in data interpretation and manuscript preparation. SY and JC analyzed the data and performed RT-qPCR analysis, and GZ and JC prepared the manuscript. All authors read and approved the final manuscript.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
This work was funded by the Research and Demonstration Project of Standardized Cultivation Technology of Pinellia ternata of Yunnan Geo-herbalism (No. 2012CG008) and the project of young and middle-aged talent of Yunnan province (Grant No. 2014HB011). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fpls.2016.01209
Data Sheet 1. Supplementary figures and tables in this study.
Data Sheet 2. The unigene sequence assembled by Trinity (Fasta).
Data Sheet 3. The unigene sequence assembled by Bridger (Fasta).
Data Sheet 4. Annotation of P. ternata unigene.
Data Sheet 5. List of Pfam domain families.
Data Sheet 6. The main identified phenylpropylamino alkaloid biosynthetic genes from P. ternata unigenes.
Data Sheet 7. Transaminase discovery.
Data Sheet 8. Reductase discovery.
Data Sheet 9. N-methyltransferase discovery.
Data Sheet 10. Putative transcription factors encoding unigenes in P. ternata.
Data Sheet 11. The information of SSR derived from all unigene.
Data Sheet 12. The sequences information of SSR primers.
Ahier, A., Rondard, P., Gouignard, N., and Khayath, N. (2009). A new family of receptor tyrosine kinases with a Venus Flytrap binding domain in insects and other invertebrates activated by amino acids. PLoS ONE 4:e5651. doi: 10.1371/journal.pone.0005651
Bankar, K. G., Todur, V. N., Shukla, R. N., and Vasudevan, M. (2015). Ameliorated de novo transcriptome assembly using Illumina paired end sequence data with Trinity Assembler. Genom. Data 5, 352–359. doi: 10.1016/j.gdata.2015.07.012
Boatright, J., Negre, F., Chen, X., Kish, C. M., Wood, B., Peel, G., et al. (2004). Understanding in vivo benzenoid metabolism in petunia petal tissue. Plant Physiol. 135, 1993–2011. doi: 10.1104/pp.104.045468
Camacho, C., Coulouris, G., Avagyan, V., Ma, N., Papadopoulos, J., Bealer, K., et al. (2009). BLAST+: architecture and applications. BMC Bioinformatics 10:421. doi: 10.1186/1471-2105-10-421
Cardle, L., Ramsay, L., Milbourne, D., Macaulay, M., Marshall, D., and Waugh, R. (2000). Computational and experimental characterization of physically clustered simple sequence repeats in plants. Genetics 156, 847–854.
Chang, Z., Li, G., Liu, J., Zhang, Y., Ashby, C., Liu, D., et al. (2015). Bridger: a new framework for de novo transcriptome assembly using RNA-seq data. Genome Biol. 16, 30. doi: 10.1186/s13059-015-0596-2
Colquhoun, T. A., Marciniak, D. M., Wedde, A. E., Kim, J. Y., Schwieterman, M. L., Levin, L. A., et al. (2012). A peroxisomally localized acyl-activating enzyme is required for volatile benzenoid formation in a Petunia x hybrida cv. ‘Mitchell Diploid’ flower. J. Exp. Bot. 63, 4821–4833. doi: 10.1093/jxb/ers153
Conesa, A., Gotz, S., Garcia-Gomez, J. M., Terol, J., and Talon, M. (2005). Blast2GO: a universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 21, 3674–3676. doi: 10.1093/bioinformatics/bti610
Dai, X., Sinharoy, S., Udvardi, M., and Zhao, P. X. (2013). PlantTFcat: an online plant transcription factor and transcriptional regulator categorization and analysis tool. BMC Bioinformatics 14:321. doi: 10.1186/1471-2105-14-321
Facchini, P. J. (2001). Alkaloid biosynthesis in plants: biochemistry, cell biology, molecular regulation, and metabolic engineering applications. Annu. Rev. Plant Physiol. Plant Mol. Biol. 52, 29–66. doi: 10.1146/annurev.arplant.52.1.29
Facchini, P. J., and De Luca, V. (2008). Opium poppy and Madagascar periwinkle: model non-model systems to investigate alkaloid biosynthesis in plants. Plant J. 54, 763–784. doi: 10.1111/j.1365-313X.2008.03438.x
Finn, R. D., Bateman, A., Clements, J., Coggill, P., Eberhardt, R. Y., Eddy, S. R., et al. (2014). Pfam: the protein families database. Nucleic Acids Res. 42, D222–D230. doi: 10.1093/nar/gkt1223
Fujii, S., and Small, I. (2011). The evolution of RNA editing and pentatricopeptide repeat genes. New Phytol. 191, 37–47. doi: 10.1111/j.1469-8137.2011.03746.x
Gaid, M. M., Sircar, D., Müller, A., Beuerle, T., Liu, B., Ernst, L., et al. (2012). Cinnamate:CoA ligase initiates the biosynthesis of a benzoate-derived xanthone phytoalexin in Hypericum calycinum cell cultures. Plant Physiol. 160, 1267–1280. doi: 10.1104/pp.112.204180
Ge, X. Y., and Wu, H. (2009). Phytochemical properties and quality evaluation methods of Pinelliae ternata. China Pharm. 18, 3–5. doi: 10.3969/j.issn.1006-4931.2009.09.002
Grabherr, M. G., Haas, B. J., Yassour, M., Levin, J. Z., Thompson, D. A., Amit, I., et al. (2011). Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 29, 644–652. doi: 10.1038/nbt.1883
Groves, R. A., Hagel, J. M., Zhang, Y., Kilpatrick, K., Levy, A., Marsolais, F., et al. (2015). Transcriptome profiling of khat (Catha edulis) and Ephedra sinica reveals gene candidates potentially involved in amphetamine-type alkaloid biosynthesis. PLoS ONE 10:e0119701. doi: 10.1371/journal.pone.0119701
Guo, X., Li, Y., Li, C., Luo, H., Wang, L., Qian, J., et al. (2013). Analysis of the Dendrobium officinale transcriptome reveals putative alkaloid biosynthetic genes and genetic markers. Gene 527, 131–138. doi: 10.1016/j.gene.2013.05.073
Hagel, J. M., Krizevski, R., Kilpatrick, K., Sitrit, Y., Marsolais, F., Lewinsohn, E., et al. (2011). Expressed sequence tag analysis of khat (Catha edulis) provides a putative molecular biochemical basis for the biosynthesis of phenylpropylamino alkaloids. Genet. Mol. Biol. 34, 640–646. doi: 10.1590/S1415-47572011000400017
Hanks, S. K., and Hunter, T. (1995). Protein kinases 6. the eukaryotic protein kinase superfamily: kinase (catalytic) domain structure and classification. FASEB J. 9, 576–596.
Hanks, S. K., and Quinn, A. M. (1991). Protein kinase catalytic domain sequence database: identification of conserved features of primary structure and classification of family members. Methods Enzymol. 200, 38–62. doi: 10.1016/0076-6879(91)00126-H
Hao, D. C., Ge, G. B., Xiao, P. G., Zhang, Y. Y., and Yang, L. (2011). The first insight into the tissue specific Taxus transcriptome via Illumina second generation sequencing. PLoS ONE 6:e21220. doi: 10.1371/journal.pone.0021220
He, P., Li, S., Wang, S. J., Yang, Y. C., and Shi, J. G. (2005). Study on chemical constituents in rhizome of Pinellia ternata. China J. Chin. Mater. Med. 20, 671–674.
Huang, L. L., Yang, X., Sun, P., Tong, W., and Hu, S. Q. (2012). The first Illumina-based de novo transcriptome sequencing and analysis of safflower flowers. PLoS ONE 7:e38653. doi: 10.1371/journal.pone.0038653
Ibdah, M., Chen, Y. T., Wilkerson, C. G., and Pichersky, E. (2009). An aldehyde oxidase in developing seeds of Arabidopsis converts benzaldehyde to benzoic Acid. Plant Physiol. 150, 416–423. doi: 10.1104/pp.109.135848
Ibdah, M., and Pichersky, E. (2009). Arabidopsis Chy1 null mutants are deficient in benzoic acid-containing glucosinolates in the seeds. Plant Biol. 11, 574–581. doi: 10.1111/j.1438-8677.2008.00160.x
Iseli, C., Jongeneel, C. V., and Bucher, P. (1999). ESTScan: a program for detecting, evaluating, and reconstructing potential coding regions in EST sequences. Proc. Int. Conf. Intell. Syst. Mol. Biol. 99, 138–148.
Jiang, N. H., Zhang, G. H., Zhang, J. J., Shu, L. P., Zhang, W., Long, G. Q., et al. (2014). Analysis of the transcriptome of Erigeron breviscapus uncovers putative scutellarin and chlorogenic acids biosynthetic genes and genetic markers. PLoS ONE 9:e100357. doi: 10.1371/journal.pone.0100357
Kanehisa, M., Goto, S., Hattori, M., Aoki-Kinoshita, K. F., Itoh, M., Kawashima, S., et al. (2006). From genomics to chemical genomics: new developments in KEGG. Nucleic Acids Res. 34, D354–D357. doi: 10.1093/nar/gkj102
Klempien, A., Kaminaga, Y., Qualley, A., Nagegowda, D. A., Widhalm, J. R., Orlova, I., et al. (2012). Contribution of CoA ligases to benzenoid biosynthesis in petunia flowers. Plant Cell 24, 2015–2030. doi: 10.1105/tpc.112.097519
Kliebenstein, D. J., D'Auria, J. C., Behere, A. S., Kim, J. H., Gunderson, K. L., Breen, J. N., et al. (2007). Characterization of seed-specific benzoyloxyglucosinolate mutations in Arabidopsis thaliana. Plant J. 51, 1062–1076. doi: 10.1111/j.1365-313X.2007.03205.x
Kobe, B., and Kajava, A. V. (2001). The leucine-rich repeat as a protein recognition motif. Curr. Opin. Struct. Biol. 11, 725–732. doi: 10.1016/s0959-440x(01)00266-4
Krizevski, R., Bar, E., Dudai, N., Levy, A., Lewinsohn, E., Sitrit, Y., et al. (2012a). Naturally occurring norephedrine oxazolidine derivatives in khat (Catha edulis). Planta Med. 78, 838–842. doi: 10.1055/s-0031-1298430
Krizevski, R., Bar, E., Shalit, O. R., Levy, A., Hagel, J. M., Kilpatrick, K., et al. (2012b). Benzaldehyde is a precursor of phenylpropylamino alkaloids as revealed by targeted metabolic profiling and comparative biochemical analyses in Ephedra spp. Phytochemistry 81, 71–79. doi: 10.1016/j.phytochem.2012.05.018
Krizevski, R., Bar, E., Shalit, O., Sitrit, Y., Ben-Shabat, S., and Lewinsohn, E. (2010). Composition and stereochemistry of ephedrine alkaloids accumulation in Ephedra sinica Stapf. Phytochemistry 71, 895–903. doi: 10.1016/j.phytochem.2010.03.019
Krizevski, R., Dudai, N., Bar, E., and Lewinsohn, E. (2007). Developmental patterns of phenylpropylamino alkaloids accumulation in khat (Catha edulis, Forsk.). J. Ethnopharmacol. 114, 432–438. doi: 10.1016/j.jep.2007.08.042
Li, D. J., Deng, Z., Qin, B., Liu, X. H., and Men, Z. H. (2012). De novo assembly and characterization of bark transcriptome using Illumina sequencing and development of EST-SSR markers in rubber tree (Hevea brasiliensis Muell. Arg.). BMC Genomics 13:192. doi: 10.1186/1471-2164-13-192
Li, R., Li, Y., Kristiansen, K., and Wang, J. (2008). SOAP: short oligonucleotide Alignment program. Bioinformatics 24, 713–714. doi: 10.1093/bioinformatics/btn025
Liu, C., Ma, N., Wang, P. Y., Fu, N., and Shen, H. L. (2013). Transcriptome sequencing and de novo analysis of a cytoplasmic male sterile line and its near-isogenic restorer line in Chili pepper (Capsicum annuum L.). PLoS ONE 8:e65209. doi: 10.1371/journal.pone.0065209
Livak, K. J., and Schmittgen, T. D. (2001). Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 4, 402–408. doi: 10.1006/meth.2001.1262
Long, M. C., Nagegowda, D. A., Kaminaga, Y., Ho, K. K., Kish, C. M., Schnepp, J., et al. (2009). Involvement of snapdragon benzaldehyde dehydrogenase in benzoic acid biosynthesis. Plant J. 59, 256–265. doi: 10.1111/j.1365-313X.2009.03864
Masao, M. (1997). Research for active principles of Pinelliae tuber and new preparation of crude drug. J. Tradit. Med. 14, 81.
Müller, M., Gocke, D., and Pohl, M. (2009). Thiamin diphosphate in biological chemistry: exploitation of diverse thiamin diphosphate-dependent enzymes for asymmetric chemoenzymatic synthesis. FEBS J. 276, 2894–2904. doi: 10.1111/j.1742-4658.2009.07017.x
Okada, T., Mikage, M., and Sekita, S. (2008). Molecular characterization of the phenylalanine ammonia-lyase from Ephedra sinica. Biol. Pharm. Bull. 31, 2194–2199. doi: 10.1248/bpb.31.2194
Oshio, H., Tsukui, M., and Matsuoka, T. (1978). Isolation of L-ephedrine from “Pinelliae tuber”. Chem. Pharm. Bull. 26, 2096–2097. doi: 10.1248/cpb.26.2096
Pérez-Rodríguez, P., Riaño-Pachón, D. M., Corrêa, L. G., Rensing, S. A., Kersten, B., and Mueller-Roeber, B. (2010). PlnTFDB: updated content and new features of the plant transcription factor database. Nucleic Acids Res. 38, D822–D827. doi: 10.1093/nar/gkp805
Pertea, G., Huang, X., Liang, F., Antonescu, V., Sultana, R., Karamycheva, S., et al. (2003). TIGR Gene Indices clustering tools (TGICL): a software system for fast clustering of large EST datasets. Bioinformatics 19, 651–652. doi: 10.1093/bioinformatics/btg034
Qualley, A. V., Widhalm, J. R., Adebesin, F., Kish, C. M., and Dudareva, N. (2012). Completionof the core β-oxidative pathway of benzoic acid biosynthesis in plants. Proc. Natl. Acad. Sci. U.S.A. 109, 16383–16388. doi: 10.1073/pnas.1211001109
Riechmann, J. L., and Meyerowitz, E. M. (1998). The AP2/EREBP family of plant transcription factors. Biol. Chem. 379, 633–646.
Shikanai, T., and Okuda, K. (2011). In vitro RNA-binding assay for studying trans-factors for RNA editing in chloroplasts. Methods Mol. Biol. 774, 199–208. doi: 10.1007/978-1-61779-234-2_13
Shu, S., Chen, B., Zhou, M., Zhao, X., Xia, H., and Wang, M. (2013). De novo sequencing and transcriptome analysis of Wolfiporia cocosto reveal genes related to biosynthesis of triterpenoids. PLoS ONE 8:e71350. doi: 10.1371/journal.pone.0071350
Sibéril, Y., Benhamron, S., Memelink, J., Giglioli-Guivarc'h, N., Thiersault, M., Boisson, B., et al. (2001). Catharanthus roseus G-box binding factors 1 and 2 act as repressors of strictosidine synthase gene expression in cell cultures. Plant Mol. Biol. 45, 477–488. doi: 10.1023/A:1010650906695
Soerensen, G. G., and Spenser, I. D. (1994). Biosynthetic route to the Ephedra alkaloids. J. Am. Chem. Soc. 116, 6195–6200. doi: 10.1021/ja00093a019
Strickler, S. R., Bombarely, A., and Mueller, L. A. (2012). Designing a transcriptome next-generation sequencing project for a nonmodel plant species. Am. J. Bot. 99, 257–266. doi: 10.3732/ajb.1100292
Swarbreck, D., Wilks, C., Lamesch, P., Berardini, T. Z., Garcia-Hernandez, M., Foerster, H., et al. (2008). The Arabidopsis Information Resource (TAIR): gene structure and function annotation. Nucleic Acids Res. 36, D1009–D1014. doi: 10.1093/nar/gkm965
Tatusov, R. L., Koonin, E. V., and Lipman, D. J. (1997). A genomic perspective on protein families. Science 5338, 631–637. doi: 10.1126/science.278.5338.631
van der Fits, L., and Memelink, J. (2000). ORCA3, a jasmonate-responsive transcriptional regulator of plant primary and secondary metabolism. Science 289, 295–297. doi: 10.1126/science.289.5477.295
Van, M. A., Schauvinhold, I., Pichersky, E., Haring, M. A., and Schuurink, R. C. (2009). A plant thiolase involved in benzoic acid biosynthesis and volatile benzenoid production. Plant J. 60, 292–302. doi: 10.1111/j.1365-313X.2009.03953.x
Wang, G. M., and Zhou, R. (2007). Progress in pharmacological studies of Pinelliae tuber. Guiding. J. TCM. 13, 97–99. doi: 10.3969/j.issn.1672-951X.2007.02.047
Wang, Z. Y., Fang, B. P., Chen, J. Y., Zhang, X. J., Luo, Z. X., Huang, L. F., et al. (2010). De novo assembly and characterization of root transcriptome using Illumina paired-end sequencing and development of cSSR markers in sweet potato (Ipomoea batatas). BMC Genomics 11:726. doi: 10.1186/1471-2164-11-726
Wu, H., Tan, X. H., Cai, B. C., and Ye, D. J. (1996). Effect of ginger-processing on L-ephedrine contents in rhizome pinelliae. China J. Chin. Mater. Med. 2, 157–159.
Xu, W. F., Zhang, B. G., Li, M., and Liu, G. H. (2007). Determination of ephedrine in rhizoma Pinelliae, rhizoma Typhonii flagelliformis and their processed products by HPLC. Lishizhen Med. Mate. Med. Res. 18, 884–885. doi: 10.3969/j.issn.1008-0805.2007.04.061
Yamada, Y., Kokabu, Y., Chaki, K., Yoshimoto, T., Ohgaki, M., Yoshida, S., et al. (2011). Isoquinoline alkaloid biosynthesis is regulated by a unique bHLH-type transcription factor in Coptis japonica. Plant Cell Physiol. 52, 1131–1141. doi: 10.1093/pcp/pcr062
Ye, J., Fang, L., Zheng, H., Zhang, Y., Chen, J., Zhang, Z., et al. (2006). WEGO: a web tool for plotting GO annotations. Nucleic Acids Res. 4, W293–W297. doi: 10.1093/nar/gkl031
Zeng, S., Xiao, G., Guo, J., Fei, Z., Xu, Y., Roe, B. A., et al. (2010). Development of a EST dataset and characterization of EST-SSRs in a traditional Chinese medicinal plant, Epimedium sagittatum (Sieb. Et Zucc.) Maxim. BMC Genomics 11:94. doi: 10.1186/1471-2164-11-94
Zhang, H., Hedhili, S., Montiel, G., Zhang, Y., Chatel, G., Pré, M., et al. (2011). The basic helix-loop-helix transcription factor CrMYC2 controls the jasmonate-responsive expression of the ORCA genes that regulate alkaloid biosynthesis in Catharanthus roseus. Plant J. 67, 61–71. doi: 10.1111/j.1365-313X.2011.04575.x
Keywords: Pinellia ternata, transcriptome, phenylpropylamino alkaloids, ephedrine, De novo sequencing
Citation: Zhang GH, Jiang NH, Song WL, Ma CH, Yang SC and Chen JW (2016) De novo Sequencing and Transcriptome Analysis of Pinellia ternata Identify the Candidate Genes Involved in the Biosynthesis of Benzoic Acid and Ephedrine. Front. Plant Sci. 7:1209. doi: 10.3389/fpls.2016.01209
Received: 25 March 2016; Accepted: 29 July 2016;
Published: 16 August 2016.
Edited by:
Jacqueline Grima-Pettenati, Centre National de la Recherche Scientifique, FranceReviewed by:
Wanchai De-Eknamkul, Chulalongkorn University, ThailandCopyright © 2016 Zhang, Jiang, Song, Ma, Yang and Chen. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sheng-chao Yang, c2hlbmdjaGFveWFuZ0AxNjMuY29t
Jun-wen Chen, Y2p3MzE0MTJAaG90bWFpbC5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.