
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Plant Sci., 14 June 2016
Sec. Plant Development and EvoDevo
Volume 7 - 2016 | https://doi.org/10.3389/fpls.2016.00788
 Analía Espinoza1
Analía Espinoza1 Rodrigo Contreras2,3
Rodrigo Contreras2,3 Gustavo E. Zúñiga2,3
Gustavo E. Zúñiga2,3 Raúl Herrera4
Raúl Herrera4 María Alejandra Moya-León4
María Alejandra Moya-León4 Lorena Norambuena1
Lorena Norambuena1 Michael Handford1*
Michael Handford1*White Chilean strawberries (Fragaria chiloensis) are non-climacteric fruits, with an exotic color and aroma. In order to discover genes involved in the development of these fruits, we identified a fragment of a gene encoding a late embryogenesis abundant domain protein, FcLDP1, that was expressed in early stages of fruit development, particularly in receptacles. Hormones play key roles in regulating the development of non-climacteric fruits. We show that the brassinosteroid content of the white strawberry varies during development. Additionally, FcLDP1 as well as the closest ortholog in the woodland strawberry, F. vesca (FvLDP1) possess multiple brassinosteroid, as well as abscisic acid (ABA) response motifs in the promoter region, consistent with the response of transiently expressed FcLDP1 promoter-GFP fusions to these hormones, and the rise in FcLDP1 transcript levels in white strawberry fruits treated with brassinosteroids or ABA. These findings suggest that both hormones regulate FcLDP1 expression during the development of white strawberries.
The octoploid white Chilean strawberry, Fragaria chiloensis produces exotic fruits with excellent taste, aroma, and color, and it is a parental species of the commercial red strawberry, F.× ananassa (Folta et al., 2005). The genus Fragaria develops aggregate accessory fruits with hundreds of achenes attached to a fleshy receptacle. Receptacles are composed of about 90% water and 10% soluble solids, substances which are imported from distal parts of the plant. As storage sinks, fruits are thus highly dependent on the xylem and phloem which together make up the vascular system. For this reason, the development of the vascular system is essential for fruit development, determining the texture and integrity of the mature fruit (Aharoni et al., 2002).
During the development of fruits, plant hormones guide multiple changes in physiological and biochemical processes (McAtee et al., 2013). In climacteric fruit, such as apples (Malus domestica) and tomatoes (Solanum lycopersicum), ethylene is the key hormone that regulates the ripening process (Cherian et al., 2014). However, the control of ripening in non-climacteric fruits, such as F. chiloensis (Aharoni et al., 2002) is less well understood. Final fruit size is determined by rapid initial cell division in early stages of development, and then by cell expansion, as water and other nutrients accumulate inside the cells (Rosli et al., 2004). During the early stages of development of both climacteric and non-climacteric fruits, changes in the levels of auxin, gibberellins, ABA, cytokinins, and brassinosteroids have been reported (Mounet et al., 2009; Symons et al., 2012). It has been observed that auxin gradients are essential for growth, and several genes, such as expansin, endo-xyloglucan, and pectate lyase families have been shown to be regulated by either auxins, gibberellins, or both (Catalá et al., 2000; Chen et al., 2002). In red strawberry (F. × ananassa), ABA levels rise gradually throughout fruit development and in the ripening process (Symons et al., 2012), consistent with the observation that down-regulation of an ABA receptor, FaPYR1, delays red strawberry fruit ripening (Chai et al., 2011). There are greater cytokinin levels after pollination, which favor the proliferation of ovary tissues, while brassinosteroids have been described as participating in elongation and cell division, vascular differentiation, flowering, and pollen development (Li, 2010). Nevertheless, to date the presence and potential effects of brassinosteroids have not been studied during the early stages of development of F. chiloensis.
Brassinosteroids and brassinosteroid signaling have been studied in Arabidopsis thaliana, based on the identification and analysis of mutants. The kinase receptor BRASSINOSTEROIDS INSENSITIVE1 (BRI1) activates transcription on recruitment of the co-receptor kinase BRI1-ASSOCIATED RECEPTOR KINASE1 (BAK1; Li et al., 2002). The Brassinazole-Resistant1 proteins (BZR1), bri1-EMS-Suppressor1 (BES1), and interacting BES1 MYC-like 1 (BIM) are transcription factors that regulate the brassinosteroid response (He et al., 2002; Yin et al., 2005; Jiang et al., 2015). In non-climacteric cucumbers (Cucumis sativus), brassinosteroids play a role in fruit set and development, as exogenous applications trigger development of unfertilized ovaries, and blocking perception of brassinosteroids with brassinazole prevents the development of parthenocarpic fruit (Fu et al., 2008). Also, in red strawberries the BRI1 transcript level is greater during the first stage of development, and brassinazole application prevents coloration (Chai et al., 2013).
Considering the role of brassinosteroids in non-climacteric fruit development, the aim of this study was to search for genes in white strawberry that could be regulated by this hormone. To do so, we used results from a suppression subtractive hybridization (SSH) library that contains candidate genes that are differentially expressed during development and ripening of F. chiloensis fruits (Pimentel et al., 2010). We selected a gene encoding a late embryogenesis abundant (LEA) domain protein, FcLDP1, as it was the most highly represented gene in the SSH library in the mid-stages of fruit development. FcLDP1 possesses brassinosteroid response elements (REs) in its promoter, and responds to exogenous applications of this hormone, as well as ABA, both when heterologously expressed in tobacco (Nicotiana tabacum) and also in white strawberry fruits.
Fruit of F. chiloensis cultivated in the same commercial field were obtained from Purén, Araucania region, Chile (latitude 38°.04′8.6′′S; longitude 73°.14′2.96′′W) in December 2014. Four developmental stages were considered, as reported in Figueroa et al. (2008), based on the size and color of the receptacle and achenes: C1, small fruits with a green receptacle and green achenes; C2, large fruits with a green receptacle and red achenes; C3, turning stage with a white receptacle and red achenes; and C4, ripe fruits with a pink receptacle and red achenes. Fruits in the laboratory were treated with various hormones and immediately frozen at -80°C until analysis.
Seeds of tobacco were germinated and grown under standard greenhouse conditions (16 h light/8 h dark, room temperature, 60–70% relative humidity) and watered with Murashige and Skoog nutrient solution (Phytotechnology Laboratories, USA). Leaves of 6-week-old plants were used to perform transient expression assays.
Four strawberry fruits for each replicate with three biological replicates for each time point were treated with the following hormonal compounds: 1 mM ABA (Sigma), 1 mM 1-NAA (auxin; Sigma), 10 μM 24-epibrassinolide (brassinosteroid; Sigma), 2 g/L Ethrel (ethylene, Bayer CropScience) or 100 μM methyl jasmonate (Sigma). All compounds were dissolved in ethanol, and the corresponding buffer-only controls were also performed. Fruits at the C2 developmental stage were placed in a plastic box and immersed in citrate buffer containing each hormone, and then collected at different time points post-treatment. All samples were frozen in liquid nitrogen and stored at -80°C.
DNA and RNA samples were isolated from each developmental stage, using the CTAB method with modifications (Chang et al., 1993; Porebski et al., 1997). RNA samples were treated with RNase-free DNase I (Thermo Fisher Scientific) to remove contaminant genomic DNA. The integrity and concentration of isolated DNA and RNA were measured in a NanoVue Plus Spectrophotometer (Nanodrop Technologies, GE Healthcare, UK). First-strand cDNAs from each developmental stage were synthesized from total RNA (2 μg) using the ImProm-IITM Reverse Transcription System (Promega), according to the manufacturer’s instructions.
A fragment of FcLDP1 was obtained from an SSH library constructed from F. chiloensis fruits at different developmental stages (Pimentel et al., 2010). To obtain the full-length cDNA sequence of FcLDP1, primers were designed from the orthologous sequence of Fragaria vesca (NC_020496.1, LG6:14021042-14026562; equivalent to gene15289, which we named FvLDP1)1. The cDNA sequence was amplified by PCR using Fv-LDPf and Fv-LDPr from a mixture of F. chiloensis fruits at different development stages and cloned in pCR8-TOPO (Invitrogen). Subsequently, the promoter of FcLDP1 was amplified and cloned in pCR8 from genomic DNA, using primers designed from the orthologous sequence in F. vesca (forward primer: Fv-pLDPf) and complementary to the cloned FcLDP1 cDNA sequence (reverse primer: Fc-pLDPr). All primer sequences are shown in Supplementary Table S1.
All cDNA and promoter clones were sequenced and analyzed by BLAST in order to assess their similarity and identity to orthologous sequences1,2. The transcriptional start site, TATA-box, CAAT-box, and putative cis-acting elements were identified using the Plant Pan Database3. The phylogenetic tree was constructed using http://phylogeny.lirmm.fr/phylo_cgi/simple_phylogeny.cgi.
The transcript levels of FcLDP1 in different stages of fruit development, and after hormone treatments were determined by RT-qPCR, based on the protocol described in Aguayo et al. (2013). Specific primers were designed with the Primer 3 software4 (Supplementary Table S1). Analysis by RT-qPCR was carried out using 10 μL SensiMix SYBR Hi-ROX kit (Bioline), with 5 μM of each primer, in a Stratagene MX3000P (Agilent Technologies) with the following parameters: 95°C for 10 min, then 40 cycles of 95°C for 15 s, 60°C for 15 s, and 72°C for 20 s. FvGAPDH (Pimentel et al., 2010) and FcRIB314 (Amil-Ruiz et al., 2013) were used as constitutively expressed control genes. The specificity of the amplification products was confirmed by registration of the melting curve of the PCR products, a heat dissociation protocol (from 55 to 95°C) and visualization in 3% agarose gels. The Ct values obtained (threshold cycle value) were analyzed by averaging the three biological replicas and two technical replicas of each sample (Livak and Schmittgen, 2001).
The promoter of FcLDP1 cloned in pCR8 was recombined into pKWSF7.0 (Karimi et al., 2002). Agrobacterium tumefaciens strain EHA105 was transformed by heat shock and a liquid culture was grown for 24 h at 28°C. Cells where then pelleted (4500 × g for 10 min) and resuspended in 10 mM MgSO4, 10 mM MES, and pH adjusted to 5.6, to reach a final OD600 nm of 0.5. Young leaves of 6-week-old tobacco plants were manually infiltrated with each bacterial suspension through the abaxial surface using a needleless plastic syringe. Transient GFP expression was evaluated by a Zeiss LSM510 Confocal, and images were processed using ImageJ, 3 days post-infiltration (Utz and Handford, 2015).
The extraction and measurement of brassinosteroid concentrations were performed by the protocol described by Pan et al. (2010). Twenty microliters of white strawberry fruit extract was resolved in a reverse-phase C18 Gemini LC-MS 1200s-6410 triplequad Agilent HPLC-ESI-MS/MS system (Palo Alto, CA, USA). Commercial epibrassinolide (Sigma) was used as a positive control.
The statistical significance of the differences of the mean data between treatment groups was calculated by analysis of variance and by Student’s t-test. Results were considered statistically significant when p ≤ 0.05 (Tukey test). All analyses were performed with GraphPad Prism 5.0 (GraphPad, San Diego, CA, USA).
The development of F. chiloensis fruits has been divided into four stages according to the size and color of achenes and receptacles (Figueroa et al., 2008). Specifically, these are small green fruits with green achenes (C1); large green fruits with red achenes (C2); white fruits with red achenes (C3), and pink fruits with red achenes (C4). An SSH library from cDNA of F. chiloensis collected at the four developmental stages was generated in order to better understand the processes underlying the development and ripening of the Chilean white strawberry (Pimentel et al., 2010), and the results analyzed in detail (Handford et al., 2014). Considering the changes in color, firmness and size in white strawberry fruits, and elevated brassinosteroid levels before the ripening of non-climacteric fruits, we focussed on those transcripts that were more abundant in C3 compared to C2 fruit stages. Altogether, 54 ESTs representing 11 contigs were differentially expressed between these two stages (Pimentel et al., 2010). We selected by far the most highly represented contig, SSHC-051 (represented by 25 of the 54 ESTs) for future analysis. Consistent with the premise that it may respond to brassinosteroids, the corresponding orthologous gene in the most closely related species to F. chiloensis, the woodland strawberry F. vesca (FvLDP1, Shulaev et al., 2011) possesses six elements that respond to brassinosteroids in its promoter. Due to the absence of a genome sequence for F. chiloensis, for cloning SSHC-051 we designed primers using the coding sequence of NC_020496.1. We determined that the coding region of the gene in F. chiloensis was 693 bp in length (Genbank accession number: KX246933), and that the corresponding genomic DNA lacked introns (Supplementary Figure S1). The predicted protein sequence was 52.2 and 41.1% identical to AtNDR1/Hairpin-induced protein (HIN1)-like 10 and AtSYP24 of A. thaliana, respectively. AtNDR1/HIN1-like 10 plays a crucial role in pathogen-induced plant responses to biotic stress (Zheng et al., 2004), whereas syntaxins of the SYP2 family are responsible for protein trafficking between pre-vacuolar compartments (PVC) and vacuoles (Sanderfoot et al., 2000; Shirakawa et al., 2010). Also, and like AtNDR1/HIN1-like 10 and AtSYP24, the sequence encoded by SSHC-051 possesses a LEA 14 (IPR004864) domain (Supplementary Figure S2), which is believed to be involved in abiotic stress tolerance and is characterized by accumulation in the late stages of embryogenesis in Arabidopsis (Hundertmark and Hincha, 2008) and in climacteric species such as peach (Prunus persica; Bonghi et al., 2011). However, in non-climacteric species like F. chiloensis, their role is more poorly understood. Considering the presence of LEA 14, we named the protein encoded by SSHC-051 as FcLDP1. Orthologs of FcLDP1 are present in the genomes of a wide diversity of plant species, including F. vesca (FvLDP1), N. tabacum, and the African oil palm (Elaeis guineensis; Supplementary Figure S3). With the aim of determining the expression profile of FcLDP1 throughout fruit development, RT-qPCR analysis was performed using fruits from the 2014 season. The transcripts of this gene were more abundant in whole fruits during the C1 stage (Figure 1A). In achenes, the transcript levels of FcLDP1 were higher in C1 and C3 stages (Figure 1B), indicating that the encoded protein may play a role in different stages of fruit and embryo development. Interestingly, FvLDP1 is also expressed during F. vesca fruit development. Highest levels of FvLDP1 expression are found in tissues from the achenes compared to receptacles in early stages of development (named as gene15289 in Kang et al., 2013), and while transcript levels of FcLDP1 between achenes and receptacles cannot be compared, greatest accumulation of FcLDP1 is found in achenes at the C1 stage, roughly equivalent to those F. vesca fruits examined by Kang et al. (2013). After dissecting F. vesca achenes, the maximum levels of expression are present in ovary wall tissues, which subsequently develop into the pericarp in mature fruits (Kang et al., 2013).
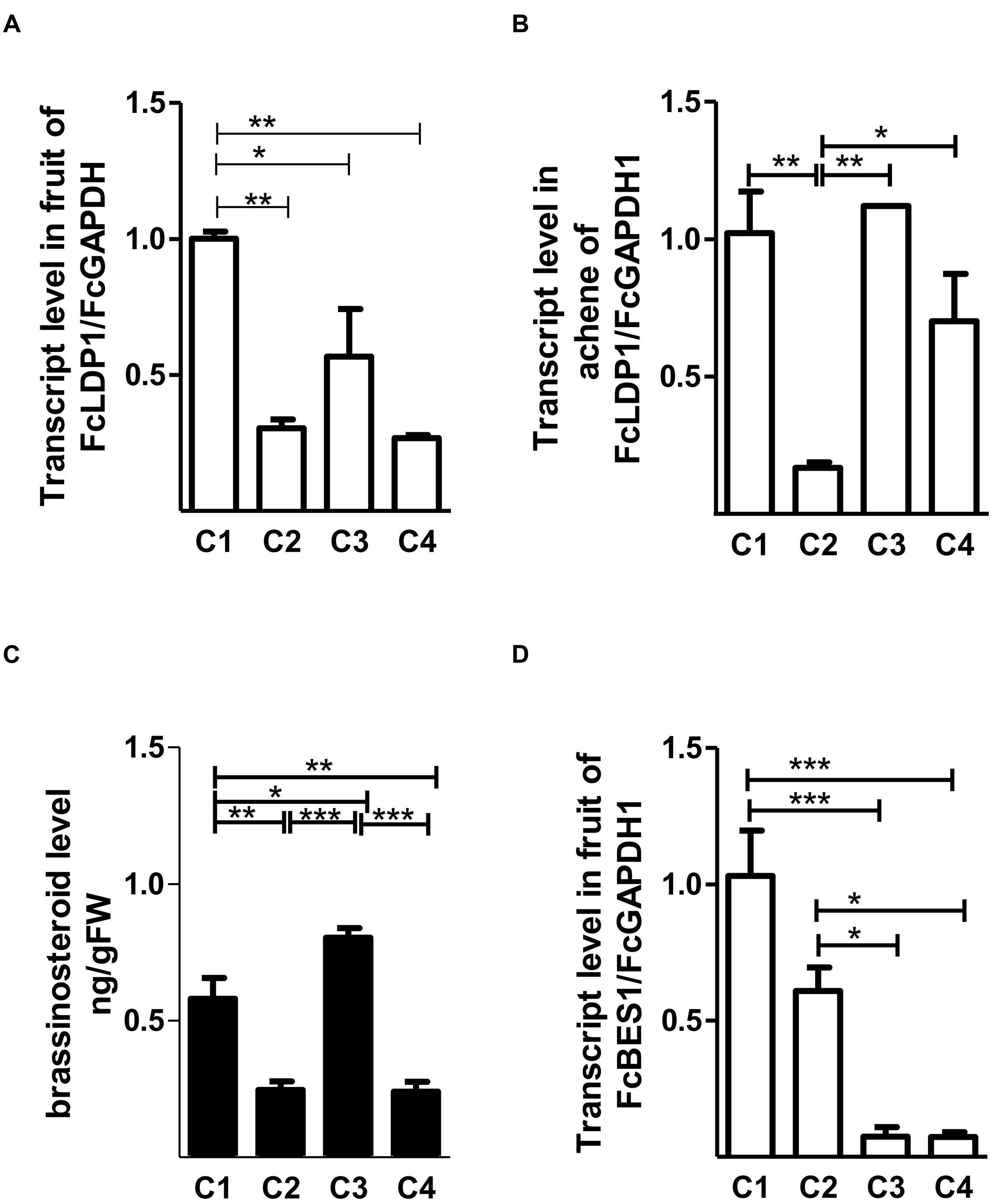
FIGURE 1. FcLDP1 transcript and brassinosteroid levels in Fragaria chiloensis. The relative transcript levels of FcLDP1 at different stages of white strawberry development in whole fruits (A) and achenes (B). Endogenous levels (ng/g-1 FW) of epibrassinolide during fruit development (C). The relative expression levels of the brassinosteroid marker gene FcBES1 during fruit development (D). The error bars show standard errors, and asterisks indicate statistical differences with one-way ANOVA and Tukey tests (*p < 0.05; **p < 0.01; ***p < 0.001).
Brassinosteroids are one of the hormones that are more abundant in the early stages of fruit development. This hormone belongs to a steroidal group of plant hormones, essential for normal plant development (Kauschmann et al., 1996). Exogenous brassinosteroid application promotes ripening in climacteric and non-climacteric fruits (Vidya Vardhini and Rao, 2002; Symons et al., 2006). Changes in levels of brassinosteroids can modulate maturation in conjunction with auxin, ABA, and other hormones (Symons et al., 2012). In white strawberry fruit, brassinosteroid levels peak at the C1 and C3 stages (Figure 1C). High levels in C1 may be related to the known role of brassinosteroids in promoting cell division, thus affecting the final size of the fruit, as shown when cucumbers are treated with exogenous applications of 24-epibrassinolide (Fu et al., 2008). On the other hand, the increase during the C3 stage may favor maturation, which is supported by the fact that ripening of F. × ananassa is accelerated when immature large green fruits (C2-equivalent) are exposed to brassinosteroids (Chai et al., 2013). Blocking the perception of brassinosteroids by using brassinazole generates a delay in maturation in F. × ananassa (Chai et al., 2013). Consistent with the high transcript levels of FcLDP1 during the C1 stage of fruit development of F. chiloensis, a greater accumulation of the transcript levels of the brassinosteroid transcriptional stimulator FcBES1 was found in C1 (Figure 1D). Furthermore, in whole fruits, levels of FcBES1 decreased progressively toward the C4 stage (Figure 1D), as did FcLDP1 levels (Figure 1A). Nevertheless, it is possible that other transcription factors regulate brassinosteroid responses in the C3 stage when a high content of brassinosteroid was detected and FcBES1 transcript levels were low. Considering the biological processes regulated by the BES1 transcription factor (Sun et al., 2010), it is possible that FcLDP1 is involved during the early stages of fruit development, when cell division rates are high.
With the purpose of determining whether brassinosteroids control FcLDP1 expression, the FcLDP1 promoter region was obtained using a forward primer designed against the equivalent orthologous region approximately 2000 bp upstream of the transcription initiation sequence in F. vesca (FvLDP1), and a reverse primer in the 5′ coding sequence of FcLDP1. Consequently, a 1924 bp putative promoter sequence of FcLDP1 was amplified and cloned. We verified that the promoter and cDNA sequences were indeed contiguous, by amplifying F. chiloensis genomic DNA. A fragment of the expected size (approximately 2600 bp; Supplementary Figure S4) was obtained. In silico analysis of the promoter sequence revealed the presence of a TATA box -111 bp upstream of the predicted start site of transcription (Genbank accession number: KX246932). Furthermore, multiple sequences associated with regulatory hormone binding sites, as well as light REs were present (Figure 2) which may be involved in controlling reproductive development. Of the brassinosteroid responsive transcription factors, BZR1 binds to brassinosteroid response elements (BRRE, CGTGYG; He et al., 2005) while BIM/BES1 binds to E-boxes (CANNTG), driving expression of target genes (Yin et al., 2005). In silico analysis of the putative promoter sequence of FcLDP1 shows that 1 BRRE (He et al., 2002) and 10 E-boxes (He et al., 2005) are present, suggesting that FcLDP1 expression is controlled by this hormone. Furthermore, we found putative REs to auxin, gibberellin, ABA, and salicylic acid (Table 1; Gubler and Jacobsen, 1992; Itzhaki et al., 1994; Shen and Ho, 1995; Shah and Klessig, 1996; Guilfoyle et al., 1998). Moreover, putative binding sites for transcription factors described in floral development [SEP4 AGL3 (Liu et al., 2013); AG and ATH-1 (Li et al., 2012; Ó’Maoiléidigh et al., 2013)], synthesis of lignin [LIM1, (Kawaoka et al., 2000; Kawaoka and Ebinuma, 2001)], flavonoid synthesis (Corn P site, Zhang et al., 2000) and a bellringer transcription factor site associated with vascular differentiation (Etchells et al., 2012) are also contained within the FcLDP1 promoter. Using data from the re-annotation of the F. vesca genome (Darwish et al., 2015), we analyzed the equivalent putative promoter region of the closest ortholog to FcLDP1, FvLDP1 (2.0 kb; Table 1). While REs to most hormones are present in both species, fewer elements that respond to brassinosteroids (2 BRRE and 4 E-boxes in FvLDP1; 1 BRRE and 10 E-boxes in FcLDP1) and fewer ABA response elements (1 ABRE in FvLDP1 and 3 ABRE in FcLDP1) are present in the woodland strawberry compared to the white Chilean strawberry (Table 1). These differences may indicate subtle variations in the hormone responsiveness that modulate the expression of LDP1 during reproductive strawberry development.
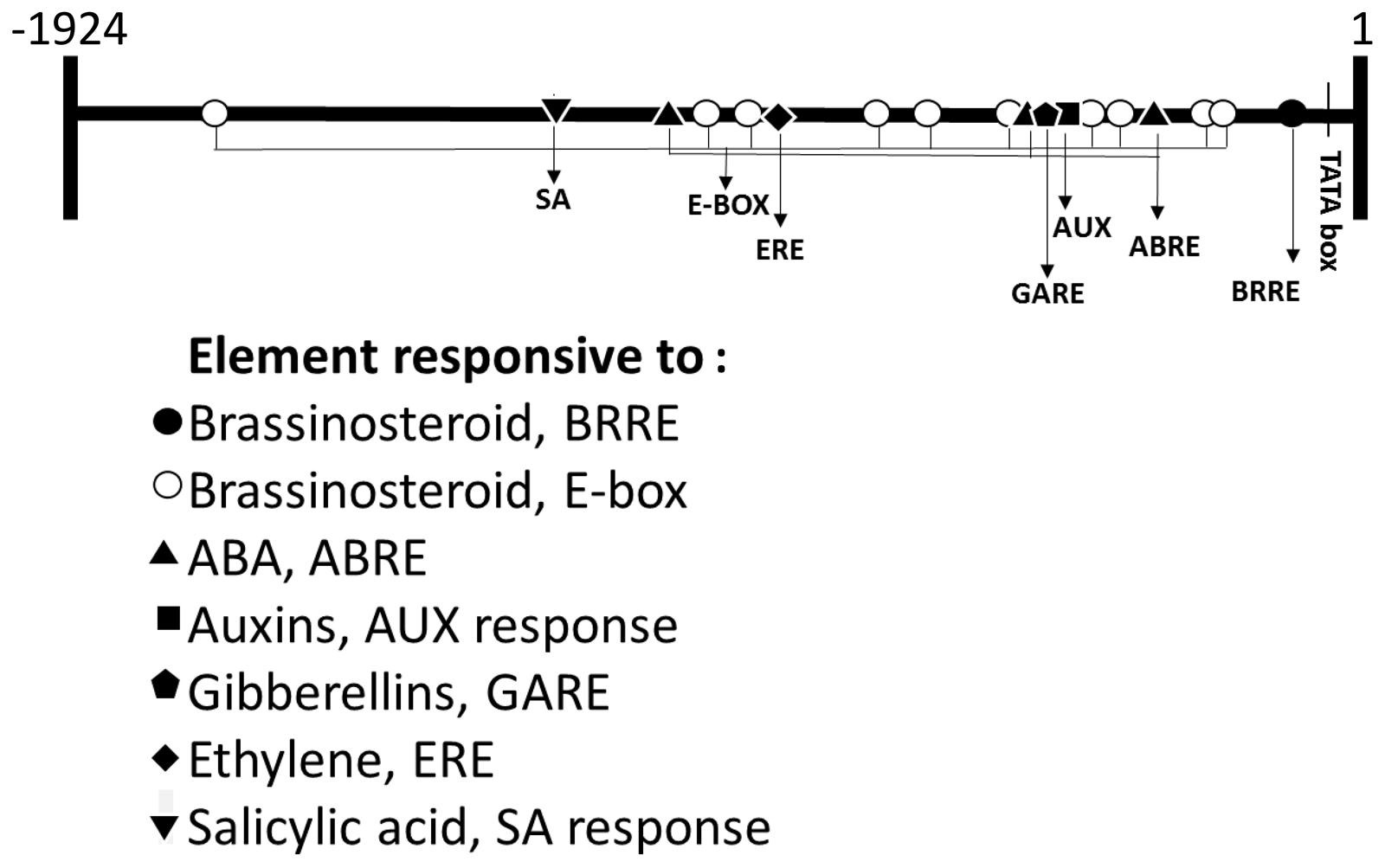
FIGURE 2. In silico analysis of the promoter of FcLDP1. The promoter and gene sequence of FcLDP1 were amplified, cloned, and the prediction of transcriptional start site, TATA-box, and hormonal response elements (REs) was performed using the Plant Pan Database (plantpan2.itps.ncku.edu.tw).
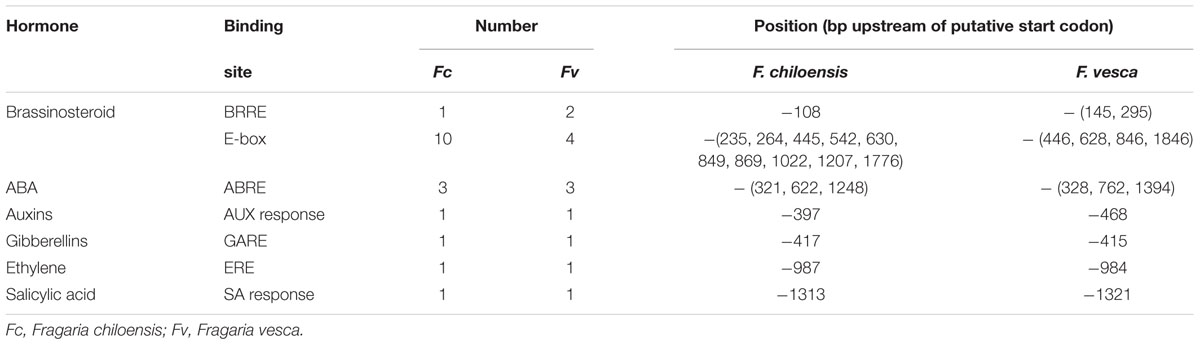
TABLE 1. Predicted hormone response elements in the FcLDP1 and FvLDP1 promoters.
In order to assess the role that hormones believed to act during non-climacteric fruit development have on FcLDP1 expression, two approaches were followed; the effect of exogenous hormone applications on promoter activity in tobacco, and on FcLDP1 transcript levels in white strawberries. First, to evaluate the regulatory capacity of the promoter, the isolated sequence was fused to the GFP coding sequence and then transiently expressed in tobacco leaves (Yang et al., 2000; Liu et al., 2011). After 3 days, the untreated infiltrated leaves were analyzed by confocal microscopy; no fluorescence was observed, indicating that no basal promoter activity is detectable. Subsequently, infiltrated leaves were sprayed with 24-epibrassinolide and observed over time. This hormone generated a marked increase in fluorescence (Figures 3A,B). This result is consistent with the highest levels of FcLDP1 transcripts in C1 fruits which possess the greatest levels of this hormone (Figures 1A,C), and suggests that the REs involved in brassinosteroid responsiveness present in the promoter are indeed functional. Specifically, it is known that BES1 induces transcription of brassinosteroid-responsive genes via E-box motifs (Yin et al., 2005). The presence of E-box motifs within 1800 nucleotides upstream of the start codon in the putative promoter region of FcLDP1 (Figure 2), together with the similar expression profiles of FcBES1 and FcLDP1 (Figures 1A,D) is consistent with FcLDP1 being a brassinosteroid responsive gene, possibly mediated by the BES1 transcription factor.
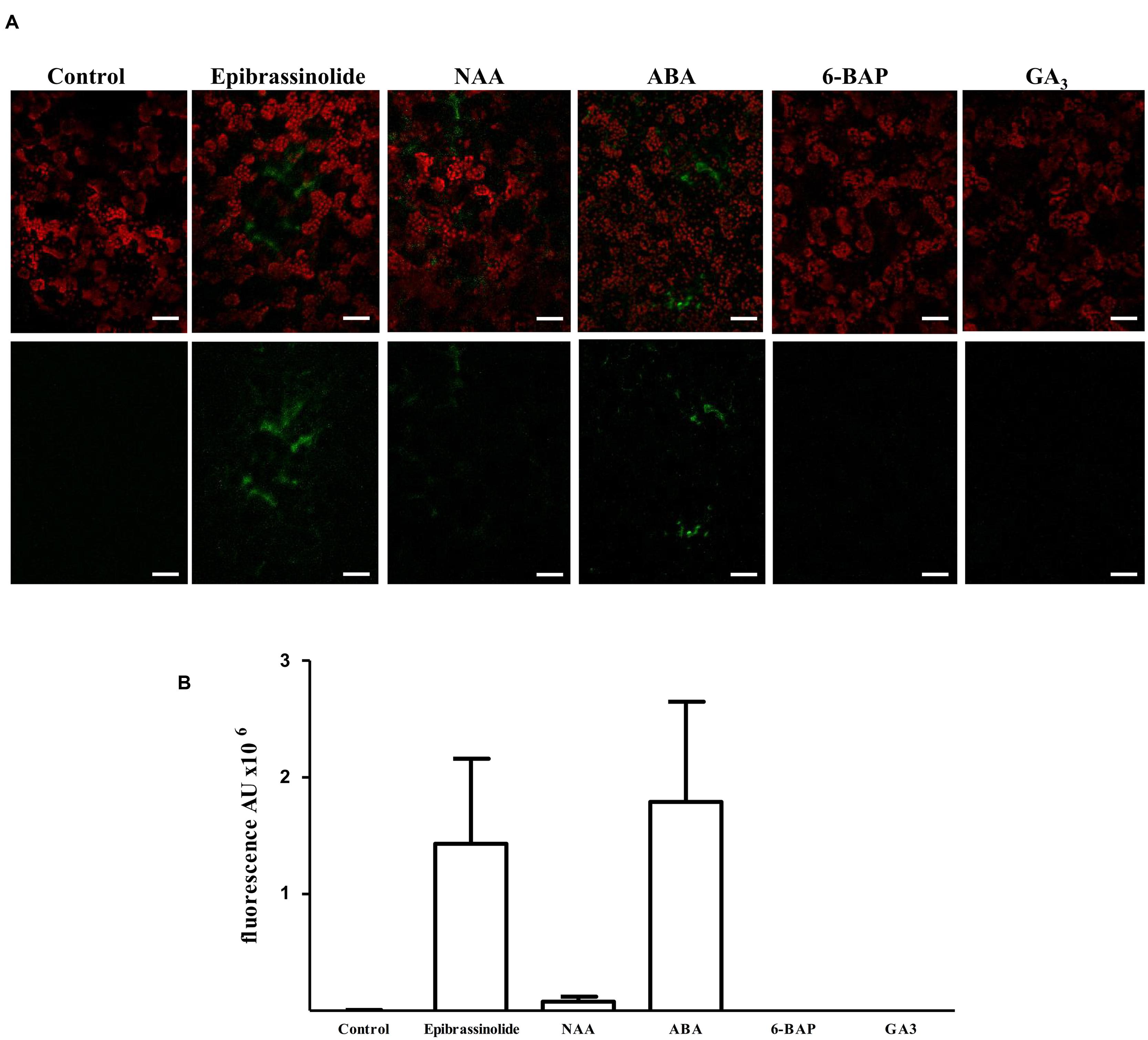
FIGURE 3. Response of the promoter of FcLDP1 to hormonal treatments. The 1.9 kb promoter of FcLDP1 was cloned upstream of GFP in pKWSF7.0, and transiently transformed using Agrobacterium tumefaciens into tobacco. Three days post-infiltration, hormones were applied for 60 min, and GFP fluorescence monitored by confocal microscopy. (A) Brassinosteroid (10 μM epibrassinolide), auxin (10 μM NAA), ABA (100 μM), cytokinin (10 μM 6-BAP), and gibberellins (1 μM GA3) were applied. Scale bar: 50 μm. (B) The fluorescence intensities of five representative images were quantified using ImageJ software in arbitrary units (AU). Error bars represent standard errors.
Given that hormones may interact and influence fruit development in different ways (Ozga and Reinecke, 2003), and taking into consideration the presence of other hormonal REs in the FcLDP1 promoter (Figure 2), we applied other hormones to the transiently transformed tobacco leaves and monitored GFP-fluorescence after 1 h. Application of NAA slightly induced transient GFP expression (Figures 3A,B). This hormone is essential for the development of F. × ananassa, where it is synthesized primarily in the achenes (Aharoni et al., 2002). During the later stages of development of F. × ananassa, several authors have reported a sustained increase in the accumulation of ABA (Chai et al., 2011; Jia et al., 2011; Symons et al., 2012). Our results show an induction of promoter activity after application of this hormone, which indicates that at least one of the three ABRE motifs in the promoter could be functional (Figures 3A,B). However, not all putative hormone REs were found to be functional. There was no induction of GFP fluorescence after gibberellin (GA3) or cytokinin (6-BAP) applications (Figures 3A,B). Gibberellins are present in higher concentrations in the early stages of development of F. × ananassa fruits, and they are involved in the final size of the fruit because REs to this hormone influence receptacle size (Csukasi et al., 2011). Although the promoter of FcLDP1 presents one GARE motif, this appears insufficient to trigger a response to this hormone, because a coupled CARE element, essential for gibberellin action, is absent (Sutoh and Yamauchi, 2003). On the other hand, cytokinins are mainly associated with vegetative tissue, and even though they have been proposed to influence non-climacteric sweet grape development (Böttcher et al., 2015), the absence of corresponding REs in the white strawberry promoter under study, may explain the lack of influence of this hormone over promoter activity. In addition, the distance between the REs and the transcription start site also alters the potential for a hormone to control gene expression (Shen and Ho, 1995).
Second, to determine whether the responsiveness of the FcLDP1 promoter to analyzed hormones is also reflected in the strawberry itself, the transcript abundance of FcLDP1 was quantified directly in stage C2 fruits. The C2 stage was chosen, as FcLDP1 expression and brassinosteroid levels had yet to peak (Figures 1A,C). At this stage, various metabolic changes occur and there is a 72% loss of firmness between stages C2 and C3 (Opazo et al., 2010). As fruit development is a complex process that involves changes in color, texture, flavor, and aroma, non-climacteric fruits such as F. × ananassa, present a program of development and maturation guided by different hormones, including auxin, gibberellin, cytokinins, brassinosteroids, ABA, and ethylene (Symons et al., 2012). These hormones were applied to white strawberry fruits as described in Section “Materials and Methods”, and the expression of known response genes was monitored to verify the effectiveness of each treatment (Figure 4). Brassinosteroid treatment at stage C2 leads to a significant increase in transcript levels of FcLDP1 (Figure 4A). In non-climacteric fruit, from flowering and fruit set, up to the first stages of fruit development, levels of brassinosteroids are high (Chai et al., 2013). Moreover, exogenous applications of this hormone in grape and F. × ananassa promote ripening, whereas treatment with brassinazole, a blocker of brassinosteroid perception, delays this process (Symons et al., 2006; Chai et al., 2013). Similarly, auxins have a higher concentration during the early stages of development favoring fruit growth by cell division and expansion in unripe fruits, yet toward maturation, auxin levels fall in F. × ananassa fruits (Aharoni et al., 2002; Fait et al., 2008; Pattison and Catalá, 2012). In this study, the application of NAA, a synthetic auxin leads to an increase in levels of FcLDP1 transcripts, with a peak at 60 min and decreasing toward 12 h after applying the treatment (Figure 4B). On the other hand, application of 1 mM ABA generated a decrease in transcript levels of FcLDP1 after 1 h post-treatment (Figure 4C). Ethylene is a key hormone in climacteric fruit ripening and although in non-climacteric fruit it is present in lower concentrations, F. × ananassa mutants in the perception of ethylene show temporal affects in phenylpropanoid metabolism, cell wall degradation, and aroma (Merchante et al., 2013). However, this phytohormone treatment did not produce changes in transcript levels of FcLDP1 in white strawberries (Figure 4D). Finally, methyl jasmonate applications resulted in a decrease in transcript levels after 6 and 12 h (Figure 4E). In F. × ananassa, it has been observed that applications of this hormone at the green C2 stage induce anthocyanin biosynthesis and degradation of chlorophyll associated with increased accumulation of ethylene (Pérez et al., 1997). In all cases after hormone applications to C2 fruits, the changes in FcLDP1 transcript levels occur within a short time frame, including a sixfold increase after just 30 min in the case of brassinosteroid treatment. While FcLDP1 transcript levels increase approximately twofold between stages C2 and C3 (Figure 1A), this transition takes approximately 1 week (Salvatierra et al., 2013). Therefore, the most-likely reason for raised FcLDP1 expression is due to the action of hormone-activated transcription factors on the accessible regions of the FcLDP1 promoter, and not to the progressive ripening of the fruits during the course of the hormone application experiments.
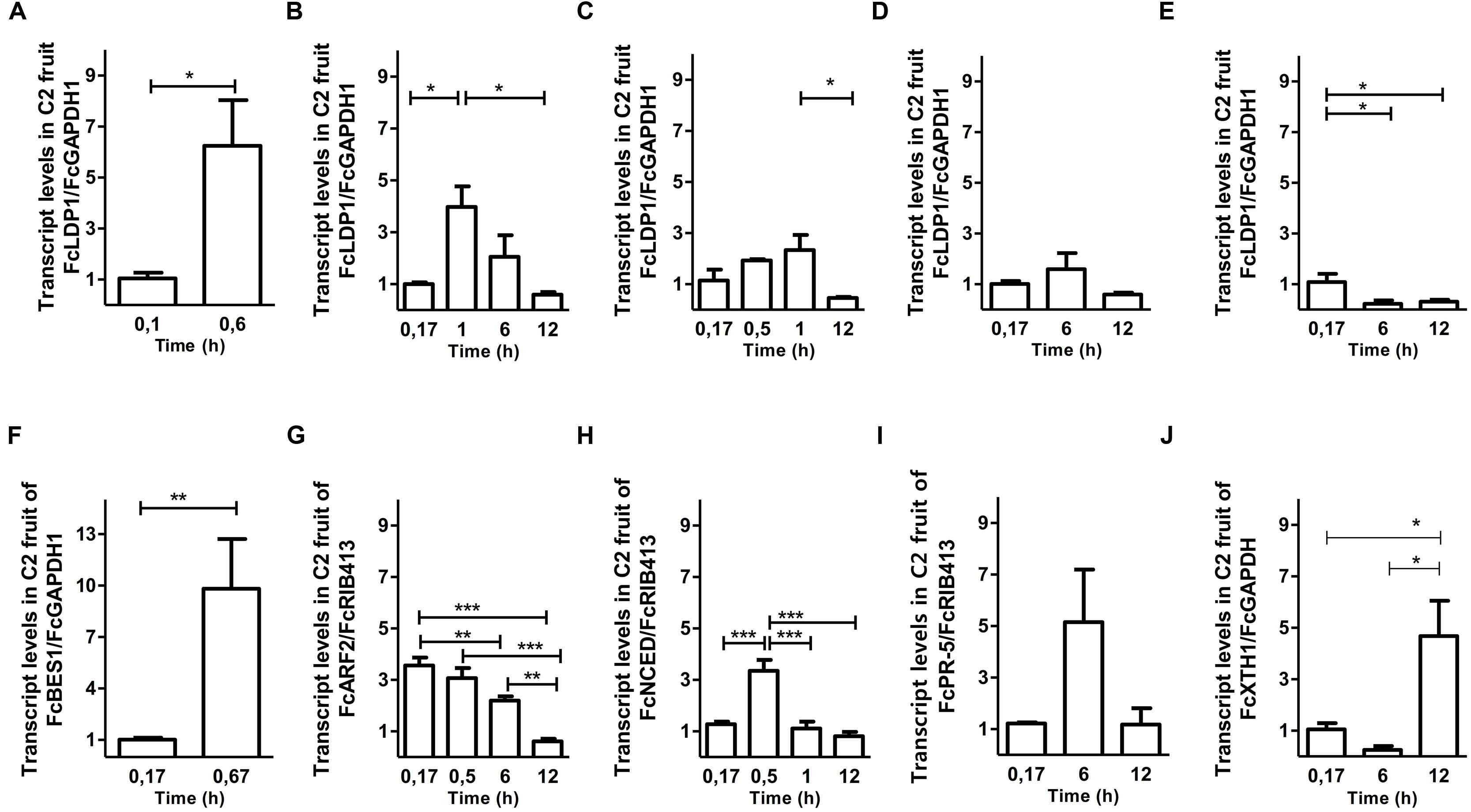
FIGURE 4. Effect of hormonal treatments on FcLDP1 transcript levels. Effect of 10 μM epibrassinolide (A), 1 mM ABA (B), 1 mM auxin (C), 2 g/L Ethrel (D), and 100 μM methyl jasmonate (E) on F. chiloensis C2-stage fruits by qRT-PCR. FcGAPDH and FcRIB413 transcript levels were used as a normalizer. FcBES1 (F), FcARP2 (G), FcNCED (H), FcXTH (I), and FcPR5 (J) were used as a positive control in their respective hormonal treatments. The error bars show standard errors and asterisks indicate statistical differences using a Student’s t-test (*p < 0.05; **p < 0.01; ***p < 0.001; A,F) or one-way ANOVA and Tukey tests (*p < 0.05; **p < 0.01; ***p < 0.001; B–E, G–J).
In F. × ananassa fruits, high levels of auxins, brassinosteroids, gibberellins, and methyl jasmonate are found in the initial stages of development (Aharoni et al., 2002). Variations in hormonal contents, particularly in ABA and ethylene, guide the subsequent development of the fruit, leading ultimately to fruits with a greater size, decreased acidity, raised sugar content, loss of chlorophyll, and reduced firmness. In F. chiloensis, we analyzed the content of brassinosteroids, which presents two peaks at the C1 and C3 stages. The presence of brassinosteroids induces the expression of a cascade of genes that trigger the development of fruits (Fu et al., 2008). Therefore, considering that FcLDP1 transcripts accumulate when there are greater brassinosteroid levels, and that it responds transcriptionally to this hormone, it is highly plausible that FcLDP1 plays a role in processes regulated by brassinosteroids during fruit development. For example, SYP2-family syntaxins including AtSYP24 are involved in vasculature development (Shirakawa et al., 2010), and FcLDP1 could play a similar role in the development of the vasculature connections within receptacles, or between achenes and receptacles. Alternatively, as a NDR1/HIN1-like10 protein, it could be involved in pathogen resistance in white strawberry fruits. However, as transcript levels and promoter activity also respond to synthetic auxin, methyl jasmonate and ABA, but not ethylene, it is possible that the function of this gene is also associated with cell division and expansion according to the timing and hormones that induce expression. To test these possibilities, functional characterization of FcLDP1 is required. Nevertheless, the substantial genetic redundancy within the SYP2 family (Shirakawa et al., 2010) and the present lack of characterized insertion lines in AtSYP24 (At1g32270) and AtNDR1/HIN1-like10 (At2g35980) pose challenges if the heterologous expression and complementation strategy were to be pursued. Moreover, the generation of F. chiloensis mutants, or over-expressing lines for the study of FcLDP1 (and other genes) is limited due to the current dearth of stable genetic transformation protocols for this specie, compounded by the extended life cycle and octoploidy of the white strawberry. The use of transient transformation, using for example an RNAi-based approach (Salvatierra et al., 2013) could therefore provide a potential alternative means for determining the effects in the shorter term of reducing FcLDP1 expression.
AE designed and performed experiments, and wrote the manuscript. RC performed experiments, GZ performed experiments. RH provided data, MM-L provided data, LN designed experiments and wrote the manuscript. MH designed experiments and wrote the manuscript.
This work was primarily funded by grant PBCT Anillo Ciencia y Tecnología ACT-1110, and partially supported by FONDECYT 1140527 to MH and 1120289 to LN.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The authors thank members of the CBMV for helpful discussions, especially Uri Aceituno, Francisca Aguayo, Milagros Bracamonte, Aliosha Figueroa, Vicente Oliva, and Sara Zapata.
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fpls.2016.00788
ABA, abscisic acid; FcLDP1, Fragaria chiloensis LEA domain protein 1; FvLDP1, Fragaria vesca LEA domain protein 1; NAA, 1-naphthaleneacetic acid.
Aguayo, M. F., Ampuero, D., Mandujano, P., Parada, R., Muñoz, R., Gallart, M., et al. (2013). Sorbitol dehydrogenase is a cytosolic protein required for sorbitol metabolism in Arabidopsis thaliana. Plant Sci. 205–206, 63–75. doi: 10.1016/j.plantsci.2013.01.012
Aharoni, A., Keizer, L. C. P., Van Den Broeck, H. C., Blanco-Portales, R., Muñoz-Blanco, J., Bois, G., et al. (2002). Novel Insight into vascular, stress, and auxin-dependent and -independent gene expression programs in strawberry, a non-climacteric fruit. Plant Physiol. 129, 1019–1031. doi: 10.1104/pp.003558
Amil-Ruiz, F., Garrido-Gala, J., Blanco-Portales, R., Folta, K. M., Muñoz-Blanco, J., and Caballero, J. L. (2013). Identification and validation of reference genes for transcript normalization in Strawberry (Fragaria × ananassa) Defense Responses. PLoS ONE 8:e70603. doi: 10.1371/journal.pone.0070603
Bonghi, C., Trainotti, L., Botton, A., Tadiello, A., Rasori, A., Ziliotto, F., et al. (2011). A microarray approach to identify genes involved in seed-pericarp cross-talk and development in peach. BMC Plant Biol. 11:107. doi: 10.1186/1471-2229-11-107
Böttcher, C., Burbidge, C. A., Boss, P. K., and Davies, C. (2015). Changes in transcription of cytokinin metabolism and signalling genes in grape (Vitis vinifera L.) berries are associated with the ripening-related increase in isopentenyladenine. BMC Plant Biol. 15:223. doi: 10.1186/s12870-015-0611-5
Catalá, C., Rose, J. K. C., and Bennett, A. B. (2000). Auxin-regulated genes encoding cell wall-modifying proteins are expressed during early tomato fruit growth. Plant Physiol. 122, 527–534. doi: 10.1104/pp.122.2.527
Chai, Y.-M., Jia, H.-F., Li, C.-L., Dong, Q.-H., and Shen, Y.-Y. (2011). FaPYR1 is involved in strawberry fruit ripening. J. Exp. Bot. 62, 5079–5089. doi: 10.1093/jxb/err207
Chai, Y.-M., Zhang, Q., Tian, L., Li, C.-L., Xing, Y., Qin, L., et al. (2013). Brassinosteroid is involved in strawberry fruit ripening. Plant Growth Regul. 69, 63–69. doi: 10.1007/s10725-012-9747-6
Chang, S., Puryear, J., and Cairney, J. (1993). A simple and efficient method for isolating RNA from pine trees. Plant Mol. Biol. Rep. 11, 113–116. doi: 10.1007/BF02670468
Chen, F., Nonogaki, H., and Bradford, K. J. (2002). A gibberellin-regulated xyloglucan endotransglycosylase gene is expressed in the endosperm cap during tomato seed germination. J. Exp. Bot. 53, 215–223. doi: 10.1093/jexbot/53.367.215
Cherian, S., Figueroa, C. R., and Nair, H. (2014). ‘Movers and shakers’ in the regulation of fruit ripening: a cross-dissection of climacteric versus non-climacteric fruit. J. Exp. Bot. 65, 4705–4722. doi: 10.1093/jxb/eru280
Csukasi, F., Osorio, S., Gutierrez, J. R., Kitamura, J., Giavalisco, P., Nakajima, M., et al. (2011). Gibberellin biosynthesis and signalling during development of the strawberry receptacle. New Phytol. 191, 376–390. doi: 10.1111/j.1469-8137.2011.03700.x
Darwish, O., Shahan, R., Liu, Z., Slovin, J. P., and Alkharouf, N. W. (2015). Re-annotation of the woodland strawberry (Fragaria vesca) genome. BMC Genomics 16:29. doi: 10.1186/s12864-015-1221-1
Etchells, J. P., Moore, L., Jiang, W. Z., Prescott, H., Capper, R., Saunders, N., et al. (2012). A role for BELLRINGER in cell wall development is supported by loss-of-function phenotypes. BMC Plant Biol. 12:212. doi: 10.1186/1471-2229-12-212
Fait, A., Hanhineva, K., Beleggia, R., Dai, N., Rogachev, I., Nikiforova, V. J., et al. (2008). Reconfiguration of the achene and receptacle metabolic networks during strawberry fruit development. Plant Physiol. 148, 730–750. doi: 10.1104/pp.108.120691
Figueroa, C. R., Pimentel, P., Gaete-Eastman, C., Moya, M., Herrera, R., Moya-León, M. A., et al. (2008). Softening rate of the Chilean strawberry (Fragaria chiloensis) fruit reflects the expression of polygalacturonase and pectate lyase genes. Postharvest Biol. Technol. 49, 210–220. doi: 10.1016/j.postharvbio.2008.01.018
Folta, K., Staton, M., Stewart, P., Jung, S., Bies, D., Jesdurai, C., et al. (2005). Expressed sequence tags (ESTs) and simple sequence repeat (SSR) markers from octoploid strawberry (Fragaria × ananassa). BMC Plant Biol. 5:12. doi: 10.1186/1471-2229-5-12
Fu, F. Q., Mao, W. H., Shi, K., Zhou, Y. H., Asami, T., and Yu, J. Q. (2008). A role of brassinosteroids in early fruit development in cucumber. J. Exp. Bot. 59, 2299–2308. doi: 10.1093/jxb/ern093
González, G., Fuentes, L., Moya-León, M. A., Sandoval, C., and Herrera, R. (2013). Characterization of two PR genes from Fragaria chiloensis in response to Botrytis cinerea infection: a comparison with Fragaria × ananassa. Physiol. Mol. Plant Pathol. 82, 73–80. doi: 10.1016/j.pmpp.2013.02.001
Gubler, F., and Jacobsen, J. V. (1992). Gibberellin-responsive elements in the promoter of a barley high-pI alpha-amylase gene. Plant Cell 4, 1435–1441.
Guilfoyle, T., Hagen, G., Ulmasov, T., and Murfett, J. (1998). How does auxin turn on genes? Plant Physiol. 118, 341–347. doi: 10.1104/pp.118.2.341
Handford, M., Espinoza, A., Bracamonte, M., Figueroa, A., Zapata, S., Aceituno, U., et al. (2014). Identification and characterisation of key genes involved in fruit ripening of the Chilean strawberry. New Biotechnol. 31:S182. doi: 10.1016/j.nbt.2014.05.912
He, J.-X., Gendron, J. M., Sun, Y., Gampala, S. S. L., Gendron, N., Sun, C. Q., et al. (2005). BZR1 is a transcriptional repressor with dual roles in brassinosteroid homeostasis and growth responses. Science 307, 1634–1638. doi: 10.1126/science.1107580
He, J.-X., Gendron, J. M., Yang, Y., Li, J., and Wang, Z.-Y. (2002). The GSK3-like kinase BIN2 phosphorylates and destabilizes BZR1, a positive regulator of the brassinosteroid signaling pathway in Arabidopsis. Proc. Natl. Acad. Sci. U.S.A. 99, 10185–10190. doi: 10.1073/pnas.152342599
Hundertmark, M., and Hincha, D. (2008). LEA (late embryogenesis abundant) proteins and their encoding genes in Arabidopsis thaliana. BMC Genomics 9:118. doi: 10.1186/1471-2164-9-118
Itzhaki, H., Maxson, J. M., and Woodson, W. R. (1994). An ethylene-responsive enhancer element is involved in the senescence-related expression of the carnation glutathione-S-transferase (GST1) gene. Proc. Natl. Acad. Sci. U.S.A. 91, 8925–8929. doi: 10.1073/pnas.91.19.8925
Jia, H.-F., Chai, Y.-M., Li, C.-L., Lu, D., Luo, J.-J., Qin, L., et al. (2011). Abscisic acid plays an important role in the regulation of strawberry fruit ripening. Plant Physiol. 157, 188–199. doi: 10.1104/pp.111.177311
Jiang, J., Zhang, C., and Wang, X. (2015). A recently evolved isoform of the transcription factor BES1 promotes brassinosteroid signaling and development in Arabidopsis thaliana. Plant Cell 27, 361–374. doi: 10.1105/tpc.114.133678
Kang, C., Darwish, O., Geretz, A., Shahan, R., Alkharouf, N., and Liu, Z. (2013). Genome-scale transcriptomic insights into early-stage fruit development in woodland strawberry Fragaria vesca. Plant Cell 25, 1960–1978. doi: 10.1105/tpc.113.111732
Karimi, M., Inzé, D., and Depicker, A. (2002). GATEWAYTM vectors for Agrobacterium-mediated plant transformation. Trends Plant Sci. 7, 193–195. doi: 10.1016/S1360-1385(02)02251-3
Kauschmann, A., Jessop, A., Koncz, C., Szekeres, M., Willmitzer, L., and Altmann, T. (1996). Genetic evidence for an essential role of brassinosteroids in plant development. Plant J. 9, 701–713. doi: 10.1046/j.1365-313X.1996.9050701.x
Kawaoka, A., and Ebinuma, H. (2001). Transcriptional control of lignin biosynthesis by tobacco LIM protein. Phytochemistry 57, 1149–1157. doi: 10.1016/S0031-9422(01)00054-1
Kawaoka, A., Kaothien, P., Yoshida, K., Endo, S., Yamada, K., and Ebinuma, H. (2000). Functional analysis of tobacco LIM protein Ntlim1 involved in lignin biosynthesis. Plant J. 22, 289–301. doi: 10.1046/j.1365-313x.2000.00737.x
Li, J. (2010). Regulation of the nuclear activities of brassinosteroid signaling. Curr. Opin. Plant Biol. 13, 540–547. doi: 10.1016/j.pbi.2010.08.007
Li, J., Wen, J., Lease, K. A., Doke, J. T., Tax, F. E., and Walker, J. C. (2002). BAK1, an Arabidopsis LRR receptor-like protein kinase, interacts with BRI1 and modulates brassinosteroid signaling. Cell 110, 213–222. doi: 10.1016/S0092-8674(02)00812-7
Li, Y., Pi, L., Huang, H., and Xu, L. (2012). ATH1 and KNAT2 proteins act together in regulation of plant inflorescence architecture. J. Exp. Bot. 63, 1423–1433. doi: 10.1093/jxb/err376
Liu, C., Teo, Zhi Wei N., Bi, Y., Song, S., Xi, W., Yang, X., et al. (2013). A conserved genetic pathway determines inflorescence architecture in Arabidopsis and rice. Dev. Cell 24, 612–622. doi: 10.1016/j.devcel.2013.02.013
Liu, W., Mazarei, M., Rudis, M. R., Fethe, M. H., and Stewart, C. N. (2011). Rapid in vivo analysis of synthetic promoters for plant pathogen phytosensing. BMC Biotechnol. 11:108. doi: 10.1186/1472-6750-11-108
Livak, K. J., and Schmittgen, T. D. (2001). Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT Method. Methods 25, 402–408. doi: 10.1006/meth.2001.1262
McAtee, P., Karim, S., Schaffer, R., and David, K. (2013). A dynamic interplay between phytohormones is required for fruit development, maturation, and ripening. Front. Plant Sci. 4:79. doi: 10.3389/fpls.2013.00079
Merchante, C., Vallarino, J. G., Osorio, S., Aragüez, I., Villarreal, N., Ariza, M. T., et al. (2013). Ethylene is involved in strawberry fruit ripening in an organ-specific manner. J. Exp. Bot. 64, 4421–4439. doi: 10.1093/jxb/ert257
Mounet, F., Moing, A., Garcia, V., Petit, J., Maucourt, M., Deborde, C., et al. (2009). Gene and metabolite regulatory network analysis of early developing fruit tissues highlights new candidate genes for the control of tomato fruit composition and development. Plant Physiol. 149, 1505–1528. doi: 10.1104/pp.108.133967
Ó’Maoiléidigh, D. S., Wuest, S. E., Rae, L., Raganelli, A., Ryan, P. T., Kwaśniewska, K., et al. (2013). Control of reproductive floral organ identity specification in Arabidopsis by the C function regulator AGAMOUS. Plant Cell 25, 2482–2503. doi: 10.1105/tpc.113.113209
Opazo, M. C., Figueroa, C. R., Henríquez, J., Herrera, R., Bruno, C., Valenzuela, P. D. T., et al. (2010). Characterization of two divergent cDNAs encoding xyloglucan endotransglycosylase/hydrolase (XTH) expressed in Fragaria chiloensis fruit. Plant Sci. 179, 479–488. doi: 10.1016/j.plantsci.2010.07.018
Ozga, J., and Reinecke, D. (2003). Hormonal interactions in fruit development. J. Plant Growth Regul. 22, 73–81. doi: 10.1007/s00344-003-0024-9
Pan, X., Welti, R., and Wang, X. (2010). Quantitative analysis of major plant hormones in crude plant extracts by high-performance liquid chromatography-mass spectrometry. Nat. Protoc. 5, 986–992. doi: 10.1038/nprot.2010.37
Pattison, R. J., and Catalá, C. (2012). Evaluating auxin distribution in tomato (Solanum lycopersicum) through an analysis of the PIN and AUX/LAX gene families. Plant J. 70, 585–598. doi: 10.1111/j.1365-313X.2011.04895.x
Pérez, A. G., Sanz, C., Olías, R., and Olías, J. M. (1997). Effect of methyl jasmonate on in vitro strawberry ripening. J. Agric. Food Chem. 45, 3733–3737. doi: 10.1021/jf9703563
Pimentel, P., Salvatierra, A., Moya-León, M.A. and Raúl, H. (2010). Isolation of genes differentially expressed during development and ripening of Fragaria chiloensis fruit by suppression subtractive hybridization. J. Plant Physiol. 167, 1179–1187. doi: 10.1016/j.jplph.2010.03.006
Porebski, S., Bailey, L. G., and Baum, B. (1997). Modification of a CTAB DNA extraction protocol for plants containing high polysaccharide and polyphenol components. Plant Mol. Biol. Rep. 15, 8–15. doi: 10.1007/BF02772108
Rosli, H. G., Civello, P. M., and Martínez, G. A. (2004). Changes in cell wall composition of three Fragaria x ananassa cultivars with different softening rate during ripening. Plant Physiol. Biochem. 42, 823–831. doi: 10.1016/j.plaphy.2004.10.002
Salvatierra, A., Pimentel, P., Moya-León, M. A., and Herrera, R. (2013). Increased accumulation of anthocyanins in Fragaria chiloensis fruits by transient suppression of FcMYB1 gene. Phytochemistry 90, 25–36. doi: 10.1016/j.phytochem.2013.02.016
Sanderfoot, A. A., Assaad, F. F., and Raikhel, N. V. (2000). The Arabidopsis genome. an abundance of soluble N-ethylmaleimide-sensitive factor adaptor protein receptors. Plant Physiol. 124, 1558–1569. doi: 10.1104/pp.124.4.1558
Shah, J., and Klessig, D. F. (1996). Identification of a salicylic acid-responsive element in the promoter of the tobacco pathogenesis-related β-1,3-glucanase gene, PR-2d. Plant J. 10, 1089–1101. doi: 10.1046/j.1365-313X.1996.10061089.x
Shen, Q., and Ho, T. H. (1995). Functional dissection of an abscisic acid (ABA)-inducible gene reveals two independent ABA-responsive complexes each containing a G-box and a novel cis-acting element. Plant Cell 7, 295–307. doi: 10.1105/tpc.7.3.295
Shirakawa, M., Ueda, H., Shimada, T., Koumoto, Y., Shimada, T. L., Kondo, M., et al. (2010). Arabidopsis Qa-SNARE SYP2 proteins localized to different subcellular regions function redundantly in vacuolar protein sorting and plant development. Plant J. 64, 924–935. doi: 10.1111/j.1365-313X.2010.04394.x
Shulaev, V., Sargent, D. J., Crowhurst, R. N., Mockler, T. C., Folkerts, O., Delcher, A. L., et al. (2011). The genome of woodland strawberry (Fragaria vesca). Nat. Genet. 43, 109–116. doi: 10.1038/ng.740
Sun, Y., Fan, X.-Y., Cao, D.-M., Tang, W., He, K., Zhu, J.-Y., et al. (2010). Integration of brassinosteroid signal transduction with the transcription network for plant growth regulation in Arabidopsis. Dev. Cell 19, 765–777. doi: 10.1016/j.devcel.2010.10.010
Sutoh, K., and Yamauchi, D. (2003). Two cis-acting elements necessary and sufficient for gibberellin-upregulated proteinase expression in rice seeds. Plant J. 34, 635–645. doi: 10.1046/j.1365-313X.2003.01753.x
Symons, G. M., Chua, Y.-J., Ross, J. J., Quittenden, L. J., Davies, N. W., and Reid, J. B. (2012). Hormonal changes during non-climacteric ripening in strawberry. J. Exp. Bot. 63, 4741–4750. doi: 10.1093/jxb/ers147
Symons, G. M., Davies, C., Shavrukov, Y., Dry, I. B., Reid, J. B., and Thomas, M. R. (2006). Grapes on Steroids. Brassinosteroids are involved in grape berry ripening. Plant Physiol. 140, 150–158. doi: 10.1104/pp.105.070706
Utz, D., and Handford, M. (2015). VvGONST-A and VvGONST-B are Golgi-localised GDP-sugar transporters in grapevine (Vitis vinifera L.). Plant Sci. 231 191–197. doi: 10.1016/j.plantsci.2014.11.009
Vidya Vardhini, B., and Rao, S. S. R. (2002). Acceleration of ripening of tomato pericarp discs by brassinosteroids. Phytochemistry 61, 843–847. doi: 10.1016/S0031-9422(02)00223-6
Yang, Y., Li, R., and Qi, M. (2000) In vivo analysis of plant promoters and transcription factors by agroinfiltration of tobacco leaves. Plant J. 22, 543–551. doi: 10.1046/j.1365-313x.2000.00760.x
Yin, Y., Vafeados, D., Tao, Y., Yoshida, S., Asami, T., and Chory, J. (2005). A new class of transcription factors mediates brassinosteroid-regulated gene expression in Arabidopsis. Cell 120, 249–259. doi: 10.1016/j.cell.2004.11.044
Zhang, P., Chopra, S., and Peterson, T. (2000). A segmental gene duplication generated differentially expressed myb-homologous genes in maize. Plant Cell 12, 2311–2322. doi: 10.1105/tpc.12.12.2311
Zheng, M. S., Takahashi, H., Miyazaki, A., Hamamoto, H., Shah, J., Yamaguchi, I. A., et al. (2004). Up-regulation of Arabidopsis thaliana NHL10 in the hypersensitive response to cucumber mosaic virus infection and in senescing leaves is controlled by signalling pathways that differ in salicylate involvement. Planta 218, 740–750. doi: 10.1007/s00425-003-1169-2
Keywords: abscisic acid, brassinosteroids, Fragaria chiloensis, fruit development, LEA domain protein, non-climacteric
Citation: Espinoza A, Contreras R, Zúñiga GE, Herrera R, Moya-León MA, Norambuena L and Handford M (2016) FcLDP1, a Gene Encoding a Late Embryogenesis Abundant (LEA) Domain Protein, Responds to Brassinosteroids and Abscisic Acid during the Development of Fruits in Fragaria chiloensis. Front. Plant Sci. 7:788. doi: 10.3389/fpls.2016.00788
Received: 03 February 2016; Accepted: 22 May 2016;
Published: 14 June 2016.
Edited by:
Stefan De Folter, Centro de Investigación y de Estudios Avanzados del Instituto Politécnico Nacional, MexicoReviewed by:
David Smyth, Monash University, AustraliaCopyright © 2016 Espinoza, Contreras, Zúñiga, Herrera, Moya-León, Norambuena and Handford. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Michael Handford, bWhhbmRmb3JAdWNoaWxlLmNs
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.