 Zuzana Jendželovská
Zuzana Jendželovská Rastislav Jendželovský
Rastislav Jendželovský Barbora Kuchárová
Barbora Kuchárová Peter Fedoročko
Peter Fedoročko
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Plant Sci. , 06 May 2016
Sec. Plant Metabolism and Chemodiversity
Volume 7 - 2016 | https://doi.org/10.3389/fpls.2016.00560
This article is part of the Research Topic Molecular and biotechnological advancements in Hypericum species View all 13 articles
Hypericin (4,5,7,4′,5′,7′-hexahydroxy-2,2′-dimethylnaphtodianthrone) is a naturally occurring chromophore found in some species of the genus Hypericum, especially Hypericum perforatum L. (St. John's wort), and in some basidiomycetes (Dermocybe spp.) or endophytic fungi (Thielavia subthermophila). In recent decades, hypericin has been intensively studied for its broad pharmacological spectrum. Among its antidepressant and light-dependent antiviral actions, hypericin is a powerful natural photosensitizer that is applicable in the photodynamic therapy (PDT) of various oncological diseases. As the accumulation of hypericin is significantly higher in neoplastic tissue than in normal tissue, it can be used in photodynamic diagnosis (PDD) as an effective fluorescence marker for tumor detection and visualization. In addition, light-activated hypericin acts as a strong pro-oxidant agent with antineoplastic and antiangiogenic properties, since it effectively induces the apoptosis, necrosis or autophagy of cancer cells. Moreover, a strong affinity of hypericin for necrotic tissue was discovered. Thus, hypericin and its radiolabeled derivatives have been recently investigated as potential biomarkers for the non-invasive targeting of tissue necrosis in numerous disorders, including solid tumors. On the other hand, several light-independent actions of hypericin have also been described, even though its effects in the dark have not been studied as intensively as those of photoactivated hypericin. Various experimental studies have revealed no cytotoxicity of hypericin in the dark; however, it can serve as a potential antimetastatic and antiangiogenic agent. On the contrary, hypericin can induce the expression of some ABC transporters, which are often associated with the multidrug resistance (MDR) of cancer cells. Moreover, the hypericin-mediated attenuation of the cytotoxicity of some chemotherapeutics was revealed. Therefore, hypericin might represent another St. John's wort metabolite that is potentially responsible for negative herb–drug interactions. The main aim of this review is to summarize the benefits of photoactivated and non-activated hypericin, mainly in preclinical and clinical applications, and to uncover the “dark side” of this secondary metabolite, focusing on MDR mechanisms.
Hypericin (4,5,7,4′,5′,7′-hexahydroxy-2,2′-dimethylnaphtodiant hrone) is a naturally occurring compound synthesized by some species of the genus Hypericum. Hypericin was first isolated from Hypericum perforatum L. (Brockmann et al., 1939), commonly known as St. John's wort, which is one of the best characterized and most important representatives of this genus, because of its broad pharmacological activity (antidepressant, antimicrobial, anticancer, anti-inflammatory, wound healing, etc.) (reviewed in Kasper et al., 2010; Wölfle et al., 2014). Hypericin and its derivatives are accumulated in special morphological structures, so called dark nodules, occurring in the aerial parts of hypericin-producing Hypericum species. The newest data on interspecific variation in localization of hypericins and spatial chemo-profiling of hypericin in some Hypericum species were published recently (Kusari et al., 2015; Kucharikova et al., 2016).
In addition to St. John's wort, this secondary metabolite was found in several other Hypericum species (Kitanov, 2001; Ayan et al., 2004) and in some basidiomycetes (Dermocybe spp.) (Dewick, 2002; Garnica et al., 2003) or endophytic fungi growing in Hypericum perforatum (Thielavia subthermophila) (Kusari et al., 2008, 2009). As hypericin is a bioactive compound that is applicable in several medicinal approaches, its content has been evaluated in in vitro grown Hypericum perforatum and in its transgenic clones (Čellárová et al., 1997; Košuth et al., 2003; Koperdáková et al., 2009), or in Hypericum cultures exposed to various biotechnological applications that focused on their preservation or stimulation of secondary metabolite production (Urbanová et al., 2006; Bruňáková et al., 2015; reviewed in: Čellárová, 2011).
Hypericin is well-known as a potent natural photosensitizing agent with great potential in anticancer photodynamic therapy (PDT) and photodynamic diagnosis (PDD). Besides its antineoplastic action, light-dependent in vitro fungicidal (Rezusta et al., 2012; Paz-Cristobal et al., 2014) and bactericidal effects (Kashef et al., 2013; García et al., 2015) have also been reported. In addition, light-activated hypericin is considered to be an effective antiviral agent (Hudson et al., 1993; Prince et al., 2000). However, some clinical studies have revealed that high doses of hypericin can induce phototoxic skin reactions without showing any detectable antiviral or antiretroviral activity in patients with viral infections (Gulick et al., 1999; Jacobson et al., 2001). The controversy concerning the virucidal effect of hypericin was summarized in detail by Kubin et al. (2005).
However, the potential use of this secondary metabolite in medicine might be broader than currently thought. Although hypericin has been extensively studied mainly because of its photodynamic and photocytotoxic properties, it also possesses various positive or negative biological activities without being activated by light.
Hypericin possesses several properties that make it a powerful fluorescent photosensitizer that is suitable for PDT and PDD—attractive applications for the treatment and detection of tumors. It possesses minimal or no toxicity in the dark (Thomas and Pardini, 1992; Vandenbogaerde et al., 1997; Miadokova et al., 2010; Jendželovská et al., 2014; Feruszová et al., 2016), accumulates preferentially in neoplastic tissues (Kamuhabwa et al., 2002; Noell et al., 2011) and generates reactive oxygen species (ROS) in the presence of light (at wavelengths around 600 nm) and oxygen (Diwu and Lown, 1993). Thus, hypericin represents a potent natural alternative to chemically synthesized photosensitizers.
PDT represents a non-invasive therapeutic approach that is beneficial in the treatment of various cancerous (reviewed in Agostinis et al., 2011) and even non-cancerous lesions and disorders (reviewed in Kim et al., 2015). In general, it is based on the combined action of a photosensitizer, light and molecular oxygen. PDT involves the administration of a non-toxic photosensitizer that preferentially accumulates in the target tissue, followed by its local illumination with harmless visible light of an appropriate wavelength, to activate and excite the photosensitizer. These photoreactions lead to the oxygen-dependent generation of cytotoxic ROS, resulting in cell death and tissue destruction. However, PDT is a multifactorial process and the degree of cellular photodamage depends on many factors, including cell permeability, the subcellular localization of the photosensitizer, the quantity of molecular oxygen, the light dose, the types of generated ROS and the attributes of cancer cells.
The exact mechanisms of cellular hypericin uptake are still unclear and require further investigation, but the results indicate that hypericin might be transported into or through cells via temperature-dependent diffusion (Thomas and Pardini, 1992; Sattler et al., 1997), partitioning, pinocytosis or endocytosis (Siboni et al., 2002). Concerning its subcellular redistribution, the co-labeling of cancer cells with hypericin and fluorescent dyes specific for cell organelles revealed that hypericin accumulates in the membranes of the endoplasmic reticulum, the Golgi apparatus, lysosomes and mitochondria (Agostinis et al., 2002; Ali and Olivo, 2002; Galanou et al., 2008; Mikeš et al., 2011). However, the cellular uptake and subcellular localization of hypericin might be affected by its lipophilicity, incubation concentrations and/or interaction with serum lipoproteins (Crnolatac et al., 2005; Galanou et al., 2008; Kascakova et al., 2008). In brief, upon light-activation, hypericin is efficient primarily in the generation of singlet oxygen (1O2; type II mechanism) and superoxide anion (; type I mechanism) (Thomas et al., 1992; Diwu and Lown, 1993), which can ultimately lead to necrosis (Du et al., 2003b; Mikeš et al., 2007, 2009), apoptosis (Ali and Olivo, 2002; Mikeš et al., 2009), autophagy-associated cell death (Buytaert et al., 2006; Rubio et al., 2012) or even to immunogenic cell death (ICD) (Garg et al., 2012a). As type II ICD inducer (Garg et al., 2015a), HY-PDT represents a promising form of active immunotherapy (Galluzzi et al., 2014) owing to spatiotemporally defined emission of damage-associated molecular patterns (DAMPs) (Garg et al., 2012b, 2015b, 2016; Zheng et al., 2016).
The photocytotoxicity of hypericin is strongly oxygen-dependent, as no such effects are present in hypoxic conditions (Thomas and Pardini, 1992; Delaey et al., 2000). Nevertheless, the final response of hypericin-mediated PDT (HY-PDT) might also be affected by the ability of cells to overcome oxidative stress through the activity of various cytoprotective mechanisms, including cellular redox systems (Mikeš et al., 2011; Mikešová et al., 2013). Furthermore, the light-dependent inhibitory effect of hypericin against various enzymes engaged in the regulation of cell survival and proliferation (Ser/Thr kinases, tyrosine kinases, etc.) has been reported (reviewed in Kubin et al., 2005). These activities might also contribute to the cytotoxic and antiproliferative effects of HY-PDT. The exact mechanisms of action and the cellular aspects of HY-PDT have been outlined and summarized in several reviews (Agostinis et al., 2002; Theodossiou et al., 2009; Mikeš et al., 2013; Garg and Agostinis, 2014).
Many in vitro studies have demonstrated the cytotoxicity of photoactivated hypericin in various cancer cell types (Xie et al., 2001; Head et al., 2006; Sacková et al., 2006; Mikeš et al., 2007; Koval et al., 2010; Mikešová et al., 2013; Kleemann et al., 2014). Moreover, recent in vivo, preclinical and clinical studies have indicated that HY-PDT might be an effective and relevant approach in the treatment of some skin tumors, carcinomas and sarcomas. In general, the depth of tumor destruction after PDT commonly ranges from a few mm to 1 cm, due to limited photosensitizer and light penetration through the tissues. Thus, PDT is effective mostly against superficial lesions and small tumors.
To our knowledge, three clinical trials of HY-PDT applied to various skin tumors have been published to date (Table 1). In the first study, Alecu et al. (1998) tested the intralesional injection of hypericin with subsequent photoactivation with visible light in the treatment of basal cell carcinoma (BCC) (eleven patients) and squamous cell carcinoma (SCC) (eight patients) and found that HY-PDT was effective in the treatment of both skin disorders. A reduction in tumor size and the generation of a new epithelium at the surface of lesions following HY-PDT were observed. Moreover, as no necrosis or cell loss was evident in the surrounding healthy tissues and no side effects were observed, with the exception of mild erythema in five cases (two patients with SCC, three patients with BCC), HY-PDT-mediated tumor targeting was selective. The treatment resulted in a complete clinical response in one SCC patient and two BCC patients, but in the remaining patients, only a partial clinical response was observed. Thus, the efficacy of HY-PDT appeared to be dependent on the initial lesion size, the total dose of hypericin, or the frequency and duration of the therapy (Alecu et al., 1998). Several years later, the potential use of HY-PDT in the treatment of non-melanoma skin cancers was explored (Kacerovská et al., 2008). A complete clinical response was observed in 50% of patients with actinic keratosis (AK) 3 months after HY-PDT, and in 22% of patients with superficial BCC and 40% of patients with Bowen's disease (BD) 6 months after HY-PDT. However, in the case of AK, the percentage reduced to 29% 6 months after HY-PDT and only partial remission was observed in patients with nodular BCC. On the other hand, complete histological remission was evident in 80% of patients with BD (Kacerovská et al., 2008). Only the partial response rate and suboptimal success of HY-PDT could be caused by the limited penetration of the skin by hypericin and by its low concentration in the final extraction product. In the third clinical trial, Rook et al. (2010) tested HY-PDT as a potentially well-tolerated and effective therapeutic modality for the treatment of lymphocyte-mediated skin disorders: malignant mycosis fungoides (MF; the most common type of cutaneous T-cell lymphoma) and non-cancerous autoimmune psoriasis. The results were promising for both diseases. In the case of MF, HY-PDT led to an improvement in the treated lesions (a size reduction by at least 50%) in the majority of patients, whereas the placebo was ineffective. Moreover, hypericin was well tolerated by the patients, with only mild to moderate phototoxic skin reactions occurring after exposure to visible light. No serious adverse effects or events were observed (Rook et al., 2010). However, the authors themselves recommended a phase III study with a greater number of patients. All these clinical data indicate that topically applied hypericin, combined with its photoactivation, might be a promising and safe alternative for the treatment of some cancerous and non-cancerous skin disorders. However, as the effectiveness of HY-PDT depends on the hypericin concentration, its total dose, its rate of tissue penetration, the frequency and duration of the therapy, or on the grade of malignancy, more clinical trials are necessary to define the optimal conditions for the whole procedure.
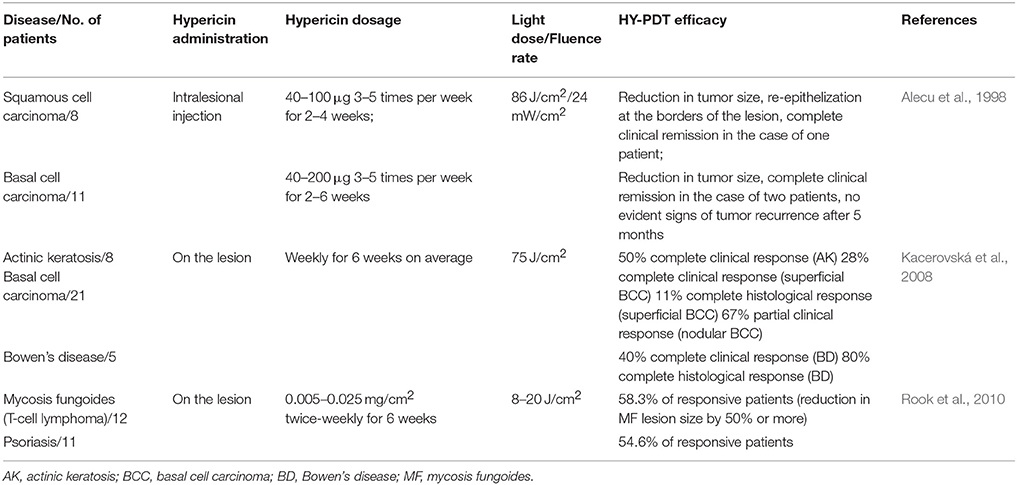
Table 1. Clinical studies to test HY-PDT efficacy.
Many further studies to test HY-PDT efficacy have been performed using mouse or rat animal models (Table 2). Several in vivo studies indicate that HY-PDT might be a promising approach in the treatment of bladder carcinomas. Kamuhabwa et al. (2002) reported selective hypericin uptake in bladder tumors and subsequently, even HY-PDT-mediated tumor damage was observed without the destruction of normal tissue (Kamuhabwa et al., 2003). In both studies, female Fisher rats with an orthotopic superficial transitional cell carcinoma (TCC) were used as an experimental model and hypericin was administered directly into the bladder via the catheter. The instilled hypericin accumulated selectively in the bladder urothelial tumors and the normal urothelium (in a ratio of 12:1), but no hypericin was detected in normal bladder submucosa and muscle layers, which is an important factor to avoid underlying tissue damage. In addition, no hypericin was detected in plasma; thus, systemic side-effects should not appear (Kamuhabwa et al., 2002).
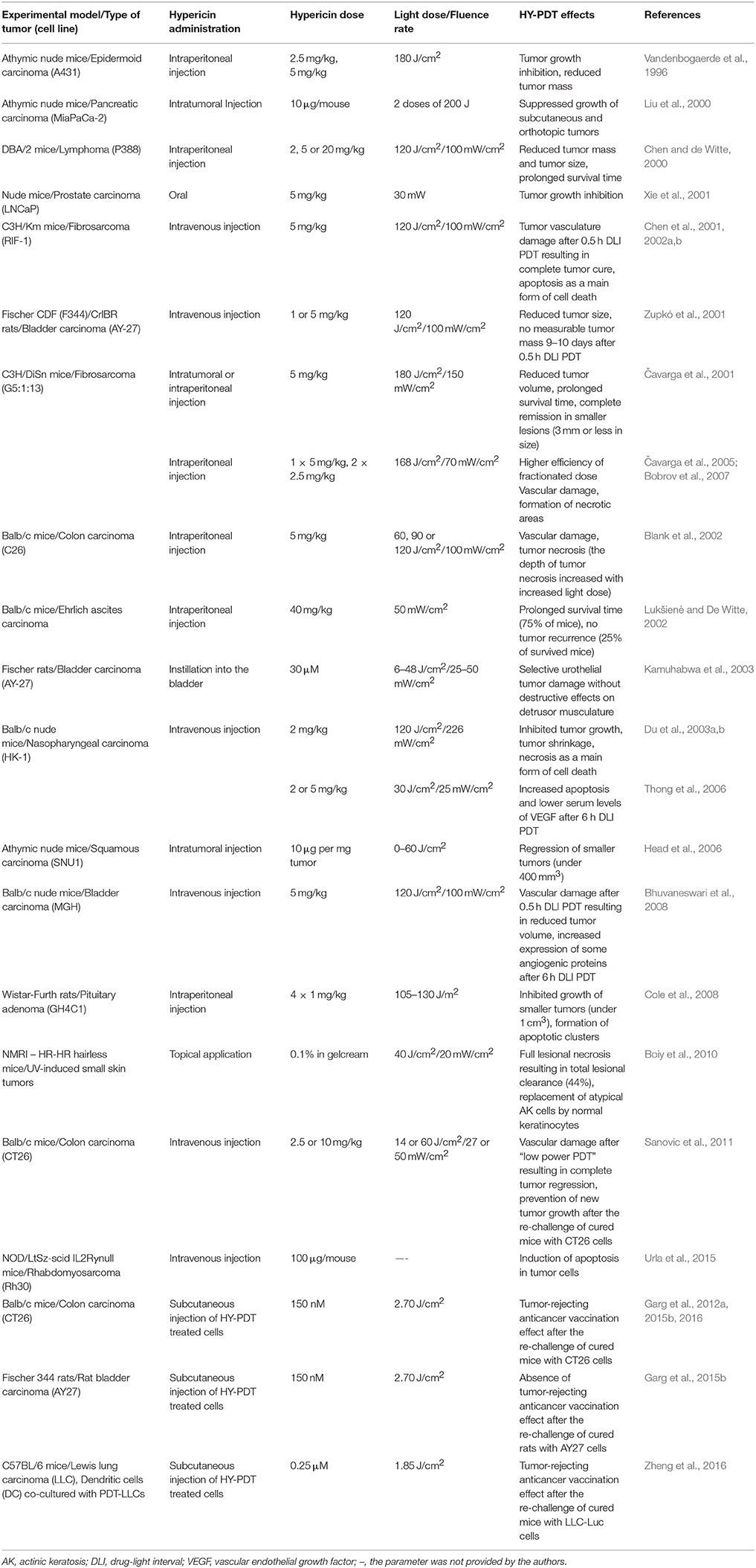
Table 2. Preclinical in vivo studies to test HY-PDT effects and conditions.
Furthermore, photoactivated hypericin resulted in selective urothelial tumor damage, with tumor cells shrinking and detaching from the bladder wall, indicating that HY-PDT might be beneficial in the treatment of superficial carcinomas and premalignant changes in the bladder. The HY-PDT that was performed under suitable light conditions had no significant effects on the other bladder layers; nevertheless, 2–5% of tumor cells survived and were responsible for tumor regrowth (Kamuhabwa et al., 2003). However, following the results of in vitro study based on TCC-derived spheroids, the same authors suggested that hyperoxygenation could overcome this problem and might enhance the efficacy of HY-PDT (Huygens et al., 2005). In addition to the orthotopic tumor model, Liu et al. (2000) also used a xenograft model in their experiments. Human MiaPaCa-2 pancreatic adenocarcinoma cells were injected subcutaneously and orthotopically into the pancreatic bed of nude, athymic mice. To allow hypericin photoactivation in orthotopic pancreatic tumor nodules, mice underwent a laparotomy that was necessary for the positioning of the optical fiber. A significant decrease in growth of subcutaneous shoulder tumors (91.2% ± 2.3%) and even pancreatic tumor nodules (42.2% ± 8.1%) was observed 4 weeks after HY-PDT, indicating that intratumor hypericin and laser therapy might also be beneficial in the treatment of unresectable pancreatic cancer (Liu et al., 2000).
However, instead of more clinically relevant orthotopic tumor models, more in vivo studies have been performed to test the efficacy, conditions or responses of HY-PDT after the treatment, only in the murine or rat xenograft or allograft models of subcutaneous carcinomas or sarcomas. Various positive effects of HY-PDT involving the inhibition of tumor growth, a prolonged survival time of the treated animals, tumor necrosis, apoptosis or damage to the tumor vasculature were observed in mice bearing human epidermoid carcinoma (Vandenbogaerde et al., 1996), human prostate adenocarcinoma cells (Xie et al., 2001), human nasopharyngeal carcinoma cells (Du et al., 2003a,b; Thong et al., 2006), human squamous carcinoma cells (Head et al., 2006), human bladder carcinoma cells (Bhuvaneswari et al., 2008), human rhabdomyosarcoma cells (Urla et al., 2015), murine lymphoma cells (Chen and de Witte, 2000), murine colon adenocarcinoma cells (Blank et al., 2002; Sanovic et al., 2011), murine fibrosarcoma cells (Čavarga et al., 2001, 2005; Chen et al., 2001, 2002a,b; Bobrov et al., 2007) or murine Ehrlich ascites carcinoma cells (Lukšienė and De Witte, 2002) and in rats bearing rat bladder transitional bladder carcinoma (Zupkó et al., 2001) or rat pituitary adenoma cells (Cole et al., 2008) (Table 2). In addition, Blank et al. (2002) demonstrated the dependence of HY-PDT efficacy on the irradiation conditions (light dose and wavelength). Tumor necrosis was much more pronounced at 590 nm than at 550 nm and even increased when the light dose was raised from 60 to 120 J/cm2; however, the maximum depth of tumor necrosis was 9.9 ± 0.8 mm at 590 nm (Blank et al., 2002). Considering the relationship between HY-PDT efficacy and tumor volume, similar results were obtained in other studies. Head et al. (2006) and Cole et al. (2008) observed a regression or reduction in tumor size only in tumors smaller than 0.4 or 1 cm3, respectively, whereas larger tumors showed only a partial response followed by their regrowth (Head et al., 2006), or did not respond to the treatment (Cole et al., 2008). Thus, light penetration into the tissue appeared to be a limiting factor. However, Cole et al. (2008) also concluded that HY-PDT can be effective in the elimination of small solid tumor residues. Čavarga et al. (2001) also determined complete remission only in smaller tumors (3 mm or less in height), but the main aim of their study was to compare the impact of intraperitoneal and intratumoral hypericin injection on the effectiveness of HY-PDT. Both schedules of hypericin administration significantly reduced tumor volume and increased the survival rate of animals. However, considering the complete response, a higher HY-PDT efficacy was observed for hypericin that was administered intraperitoneally (44.4%) compared to intratumorally (33.3%) (Čavarga et al., 2001). Moreover, it was later demonstrated that a better therapeutic response was obtained after fractionated hypericin administration (two 2.5 mg/kg doses; 6 and 1 h before irradiation) than after a single hypericin dose (5 mg/kg; 1 or 6 h before irradiation) (Čavarga et al., 2005).
Furthermore, Chen et al. (2001) reported the correlation between hypericin biodistribution, HY-PDT efficacy and various administration–irradiation time intervals (drug–light intervals—DLIs). It was found that shortly (0.5 h) after the intravenous administration of hypericin (5 mg/kg), the photosensitizer was located preferentially within tumor blood vessels. At 6 h after injection, maximum intratumoral hypericin content was evident and only poor fluorescence was detected in the tumor vasculature. Despite high tumor hypericin levels, no tumor cure was observed after longer DLI HY-PDT (6 h) treatment. However, the efficacy of PDT was maximal (100% tumor cure) when irradiation was performed at 0.5 h after hypericin administration (short DLI), indicating strong HY-PDT-induced damage to the tumor vasculature (Chen et al., 2001). Similar results were obtained by Zupkó et al. (2001). In TCC tumors, the outcome of HY-PDT was highly dependent on DLI (0.5, 6, or 24 h). The strongest effect, which resulted in no tumor regrowth in some rats, was evident after 0.5 h DLI PDT. At the same time, the highest hypericin concentration was detected in the plasma, indicating that PDT-mediated tumor vascular damage was responsible for its antineoplastic action (Zupkó et al., 2001). The antivascular and strong antitumoral effects of short DLI HY-PDT were also subsequently confirmed in the murine fibrosarcoma model (Chen et al., 2002a,b). Damage to tumor vessels was also detected following long DLI HY-PDT treatments, but many viable tumor cells were present, especially at the tumor periphery, indicating that this PDT modality only induced partial vascular collapse (Chen et al., 2002a). In agreement with these results, Bobrov et al. (2007) also observed primary vascular damage after HY-PDT with either a single dose (5 mg/kg; 1 or 6 h before irradiation) or fractionated hypericin administration (two 2.5 mg/kg doses; 6 and 1 h before irradiation), which subsequently progressed to tumor tissue necrosis. The preference of HY-PDT-mediated vasculature or cellular damage appears to be dependent on the distribution and accumulation of the photosensitizer; thus, greater vascular destruction is expected after short DLI PDT and more direct killing of tumor cells is expected after long DLI PDT. However, because the damage to tumor vessels can have devastating consequences for whole tumor mass, the targeting of the tumor vasculature via short DLI PDT might be more effective in the treatment of solid tumors than a long DLI PDT. Moreover, apoptosis was the main form of cell death responsible for tumor eradication following short DLI HY-PDT (Chen et al., 2002b). In contrast, Thong et al. (2006) observed significantly more apoptosis after long DLI PDT (6 h) compared to short DLI PDT (1 h). However, different irradiation conditions and tumor models were used in both studies: whereas Chen et al. (2002b) photoactivated hypericin with a light dose of 120 J/cm2 delivered at a fluence rate of 100 mW/cm2, a lower fluence rate HY-PDT (light dose of 30 J/cm2, fluence rate of 25 mW/cm2) was applied by Thong et al. (2006). The results indicate that long DLI PDT might also be effective in the induction of programmed cell death, but only under low fluence rate conditions. Moreover, lower serum levels of vascular endothelial growth factor (VEGF) were detected after HY-PDT using a long DLI and a low fluence rate, which can reduce the risk of new tumor vasculature formation (Thong et al., 2006).
As PDT-mediated tissue damage can lead to various cellular and molecular responses, Bhuvaneswari et al. (2008) examined the potential anti-angiogenic vs. angiogenic properties of short (0.5 h) and long (6 h) DLI HY-PDT. Both HY-PDT scenarios led to a reduction in tumor volume, but the effect was much more pronounced for a short DLI. These findings agree with the above-mentioned results (Chen et al., 2001, 2002a,b) and suggest the destruction of the tumor vasculature after short DLI HY-PDT. However, in addition to its antitumor activities, cellular-targeted long DLI HY-PDT induced the expression of some angiogenic proteins in tumor tissue, including VEGF, tumor necrosis growth factor-α (TNF-α) and interferon-α (INF-α), which potentially lead to the formation of new vessels (Bhuvaneswari et al., 2008). In subsequent studies, the efficacy of HY-PDT was enhanced using monoclonal antibodies against VEGF and the epidermal growth factor receptor (EGFR) (Bhuvaneswari et al., 2010, 2011). For short DLI HY-PDT, similar results were obtained by Sanovic et al. (2011) in mice bearing CT26 colon carcinoma cells. Complete tumor regression was observed after “low-power HY-PDT” (a hypericin dose of 2.5 mg/kg, a short DLI of 0.5 h, a light dose of 14 J/cm2 and a fluence rate of 27 mW/cm2), as no visible or palpable tumors were detected for at least 60 days after the treatment. In contrast, all mice exposed to “high-power HY-PDT” (a hypericin dose of 10 mg/kg, a short DLI of 1 h, a light dose of 60 J/cm2 and a fluence rate of 50 mW/cm2) died 2 days after the treatment as a consequence of internal bleeding. Thus, both HY-PDT modalities with short DLIs appeared to preferentially target the vessels, but a higher hypericin concentration and light dose produced a much stronger response. These results suggest that “low-power HY-PDT” is strong enough to completely eliminate tumors by damaging their vasculature. Moreover, the re-challenge of cured mice with tumorigenic CT26 cells did not result in new tumor growth, indicating the HY-PDT-mediated induction of the antitumor immune response (Sanovic et al., 2011). HY-PDT-mediated induction of anticancer immunity was also observed in the studies utilizing different in vivo experimental models. Immunization of BALB/c mice with “dying or dead” colon carcinoma CT26 cells prevented the tumor growth at the rechallenge site treated with live CT26 tumor cells. Approximately 70–85% of the mice immunized with HY-PDT treated CT26 cells efficiently rejected the formation of CT26-derived tumors at challenge site (Garg et al., 2012a, 2015b, 2016). Activation of adaptive immune system was also detected in immunocompetent C57BL/6 mice immunized with HY-PDT treated Lewis lung carcinoma (LLC) cells and LLC cells co-cultured with dendritic cells (Zheng et al., 2016). The results of these three independent experimental groups suggest great potential of HY-PDT in development of anticancer vaccines.
Similar results as in the case of partial remission in patients with nodular BCC (Kacerovská et al., 2008) were obtained in an in vivo study using hairless mice with UV-induced non-melanoma skin tumors (AK, SCC) as an experimental model (Boiy et al., 2010). Photoactivated hypericin induced a total and partial response in 44 and 22% of lesions (diameter of 1–2 mm), respectively, with evident lesional necrosis and the replacement of atypical AK cells for normal keratinocytes. However, 33% of lesions were non-responsive to HY-PDT. Tumor penetration and the selectivity of topically applied hypericin was also limited by mouse skin, as the accumulation of hypericin was highest in the outermost epidermal layer and lower hypericin levels were detected in the rest of the epidermis and the dermis (Boiy et al., 2010).
Some preclinical and clinical results gave relatively disappointing results and in some cases, the efficacy of HY-PDT was not as high as expected. The above-mentioned studies contributed to a better understanding of HY-PDT-induced responses and highlighted the importance of optimizing the conditions, such as suitable hypericin administration, light dose, fluence rate, or time intervals and implicated HY-PDT as a promising anticancer therapeutic approach; however, more clinically relevant studies and trials are required for the implementation of HY-PDT into clinical practice.
In addition to their therapeutic abilities, most photosensitizers are also potent diagnostic agents. The main principle and relevance of PDD is to enhance the contrast between neoplastic and surrounding healthy tissue, which should contribute to the surgical clearance of the whole tumor mass or its small residues. Due to the fluorescent properties of hypericin and its specificity for neoplastic tissue, hypericin-mediated PDD (HY-PDD) is being tested for various clinical uses, including optical tumor imaging, and the targeting, monitoring or detection of tumor stages and grades. To date, fluorescence diagnosis using hypericin has been clinically tested in bladder, head and neck cancers or gliomas (Table 3).
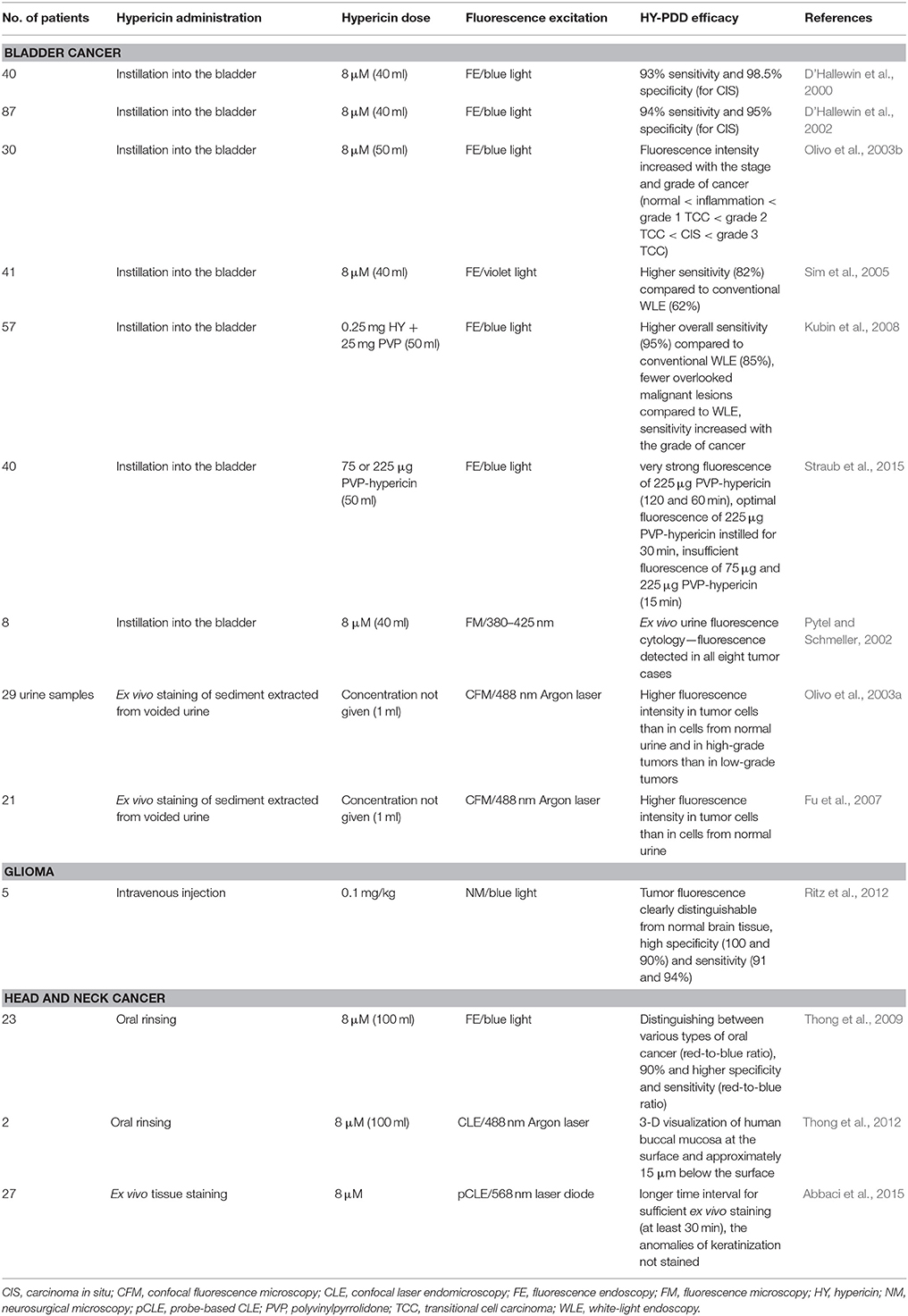
Table 3. Clinical studies to test HY-PDD efficacy, sensitivity and specificity.
In most published clinical studies, HY-PDD was applied to bladder tumors. As already mentioned in the HY-PDT section, Kamuhabwa et al. (2002) (Table 4) revealed selective hypericin uptake in rat bladder tumors using in situ laser-induced fluorescence (LIF) and fluorescence microscopy. The results suggest that hypericin is very beneficial in visualization and distinguishing the tumor mass from normal tissue using various fluorescent techniques. Thus, hypericin could be used not only in PDT, but also in the PDD of superficial bladder tumors.
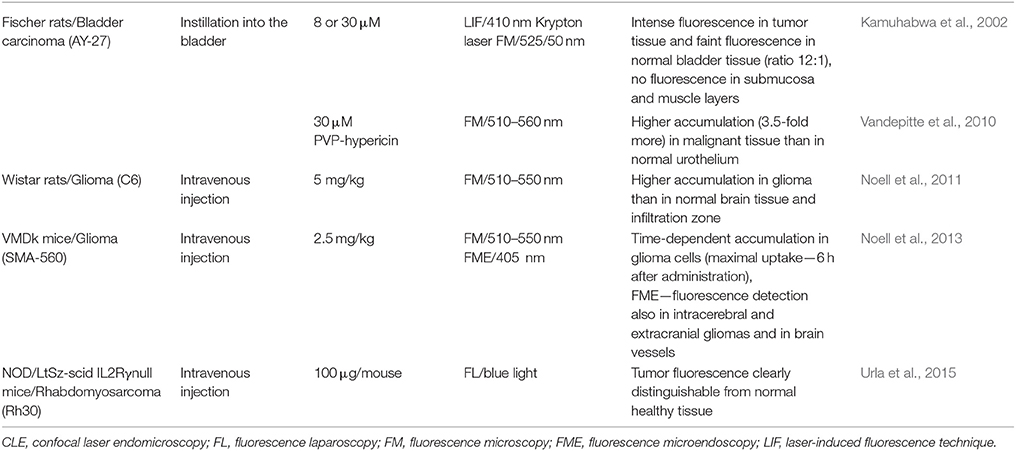
Table 4. Preliminary data for clinical HY-PDD applications and in vivo studies concerning hypericin accumulation.
At about the same time, the first clinical studies were performed by D'Hallewin et al. (2000, 2002), who examined the fluorescence-based detection of flat bladder carcinomas in situ (CIS) and in papillary non-invasive bladder tumors after the intravesical instillation of hypericin (at least 2 h). The fluorescence emission was induced using fluorescence endoscopy (FE) under blue-light illumination. Hypericin accumulated selectively in tumor cells and papillary and flat lesions showed red fluorescence, whereas no fluorescence was evident in the normal bladder tissue. Subsequently, biopsies were taken from fluorescent regions for microscopic analyses. The results of both clinical studies suggested that HY-PDD has a high sensitivity and specificity for the detection of bladder cancer (D'Hallewin et al., 2000, 2002). Moreover, the results obtained by confocal microscopy, white-light endoscopy (WLE) and histopathology revealed that the intensity of hypericin fluorescence increased with the stage and grade of bladder cancer (normal bladder tissue < inflammation in the bladder < grade 1 TCC < grade 2 TCC < CIS < grade 3 TCC) (Olivo et al., 2003b). Therefore, HY-PDD could be used as a diagnostic aid to the histopathology of bladder tumors.
Nowadays, bladder lesions are conventionally diagnosed under WLE followed by biopsies that are necessary for the histological examination of suspicious tissues. However, in previous studies, some lesions were barely visible or were absent in white light (D'Hallewin et al., 2000, 2002), thus, WLE appears not to be sensitive enough to reveal all CIS lesions and leads to a high risk of missing the tumor. Therefore, another clinical study compared the WLE and HY-PDD methods (Sim et al., 2005). Hypericin was instilled into the bladder (for 2 h) and immediately after WLE, FE (violet light) was used to induce fluorescence emission in the same bladder regions. Despite the comparable specificity of both approaches (91% for HY-PDD, 98% for WLE), HY-PDD was more sensitive (82%) than WLE (62%) (Sim et al., 2005), suggesting that hypericin might be very potent in the labeling and early detection of flat superficial bladder tumors. Similar results were obtained by Kubin et al. (2008) using polyvinylpyrrolidone (PVP) bound to hypericin as a new water-soluble formula for the improvement of hypericin-mediated bladder cancer detection and diagnosis. Hypericin-PVP was intravesically instilled 1–2 h prior to FE. The overall sensitivity of PDD with PVP-hypericin (95%) was significantly higher than WLE (85%). The maximum contrast in sensitivity was evident in the case of CIS (100% for PDD vs. 33% for WLE) and dysplasia (85% for PDD vs. 31% for WLE) (Kubin et al., 2008).
As the PVP–hypericin complex represents a potent water-soluble PDD agent without the necessity of binding to serum proteins, its biodistribution (Vandepitte et al., 2010) and optimal dosage and instillation time were evaluated (Straub et al., 2015) in tumor-bearing rats and in patients with bladder cancer, respectively. Vandepitte et al. (2010) (Table 4) demonstrated the uniform distribution of instilled PVP–hypericin in all cell layers of the malignant urothelium, whereas its penetration into the normal bladder epithelium was very limited. Straub et al. (2015) tested various combinations of PVP–hypericin dosage (75 and 225 μg) and instillation time (15, 30, 60, and 120 min) to identify the optimal PDD conditions. Even though the fluorescence of 225 μg PVP-hypericin instilled for 120 and 60 min was very strong, the shorter instillation time (30 min) for 225 μg PVP–hypericin was evaluated as optimal. A lower photosensitizer dose (75 μg) and 15 min with a dose of 225 μg were insufficient to detect the lesions (Straub et al., 2015). The authors established the most suitable dosage and instillation time of PDD with PVP–hypericin, but they suggest a larger phase IIB study should be performed to determine the sensitivity and specificity of these optimal conditions.
All the above-mentioned results indicate that HY-PDD is highly sensitive in the detection of early bladder cancer and could be routinely used as a diagnostic approach. Moreover, no photobleaching during FE and resection or side effects were detected (D'Hallewin et al., 2002; Olivo et al., 2003b; Sim et al., 2005; Kubin et al., 2008). Kamuhabwa et al. (2005) also conclude that either photosensitization or systemic side-effects should not be expected in patients after intravesical hypericin administration, as the hypericin concentration in plasma was below the detection limit (< 6 nM).
Another common method used for bladder cancer diagnosis is ex vivo urine cytology, which microscopically analyzes the exfoliated bladder cells from voided urine. This diagnostic technique is non-invasive and less time-consuming than taking biopsy specimens. However, its sensitivity to detect early-stage or low-grade cancer is relatively low. Thus, several teams have focused on a technique that combines HY-PDD and urine cytology (Pytel and Schmeller, 2002; Olivo et al., 2003a; Fu et al., 2007). In the first study conducted by Pytel and Schmeller (2002), voided urine was analyzed in eight patients following intravesically instilled hypericin (for at least 1 h). Even though the number of patients was quite low, hypericin fluorescence was detected in all cases of bladder cancer. On the contrary, Olivo et al. (2003a) and Fu et al. (2007) performed HY-PDD-mediated urine cytology without intravesical instillation of hypericin. In both studies, sediments extracted from patient urine samples were incubated with hypericin in the dark for 15 min and were subsequently analyzed using confocal fluorescence microscopy. The overall fluorescence intensity of the urothelial cells was significantly higher in urine from early-grade TCC than in normal samples, which enabled the differentiation between normal and early bladder cancer specimens (Olivo et al., 2003a). This finding was later confirmed through a diagnostic algorithm (Fu et al., 2007). Moreover, fluorescence was even higher in high-grade tumors than in low-grade tumors (Olivo et al., 2003a). The results indicate that ex vivo fluorescence cytology using hypericin might be a promising diagnostic method for the detection and identification of early and low-grade bladder cancer.
It is well-known that malignant gliomas are tumors with a very poor prognosis and their complete resection significantly improves and extends the survival of patients. Thus, in these cases, the enhancement of the contrast between tumor and surrounding healthy tissue would be very beneficial for surgeons.
In the first in vivo study, Noell et al. (2011) (Table 4) investigated the accumulation of hypericin in tumors arising from intracerebrally implanted C6 glioma cells, in the zones surrounding the tumors and in healthy brain tissue. Hypericin was injected intravenously and its uptake was maximal 24 h after injection. Considering tissue autofluorescence, the ratios of fluorescence intensities were as follows: tumor:infiltration zone:normal tissue = 19.8:2.5:1.0. Because hypericin accumulation was significantly higher in the tumor than in normal tissue, it could be effectively used as a fluorescence marker for glioma detection (Noell et al., 2011). According to these promising preliminary results, the hypericin-mediated visualization of tumor tissue during its surgical resection was examined in five patients with recurrent glioblastomas (Ritz et al., 2012). Hypericin was injected intravenously 6 h prior to the surgical procedure, which was performed using a neurosurgical microscope under switchable white- and blue-light modes. Malignant tumor tissue (red fluorescence) was clearly distinguishable from the healthy brain tissue (blue color) in all patients and the margins of the tumors showed weaker pink fluorescence. Moreover, specimens were taken for histological evaluation, which was carried out by two neuropathologists and showed 100 and 90% specificity and 91 and 94% sensitivity. The obtained results suggest that HY-PDD is well-tolerated and represents a method that is sufficiently sensitive and specific for the intraoperative visualization of malignant gliomas (Ritz et al., 2012).
Furthermore, time-dependent hypericin uptake was investigated and observed in a subcutaneous glioma mouse model using microendoscopy, an approach that is not designed for microsurgical tumor resection, but is very useful for applications such as the visualization of different tissue compartments, the identification of vessels or the detection of optimal regions for biopsy. To verify the potential to detect intracerebral gliomas using microendoscopy, tumor cells were also implanted into the brain. After craniotomy, hypericin fluorescence was detected in intracerebral and extracranial gliomas and also in the vessels located in the cortical surface of the contralateral hemisphere (Noell et al., 2013) (Table 4).
In addition to the above-mentioned studies, HY-PDD was also examined in mice bearing rhabdomyosarcoma (Urla et al., 2015) and in patients with various types of head and neck cancer (Thong et al., 2009, 2012; Abbaci et al., 2015) (Tables 3, 4).
Similarly to bladder cancer, lesions in the oral cavity are conventionally diagnosed using WLE and histopathology. However, the results obtained by Thong et al. (2009) demonstrate the great potential of hypericin for the diagnosis of various oral cancer types (hyperplasia, cellular pleomorphic adenoma of the palate, dysplasia, SCC). After oral rinsing with hypericin solution (over 30 min), FE was performed and the captured images were analyzed using several parameters. Firstly, the selective uptake of hypericin in tumor tissue was confirmed. Moreover, an increase in the red-to-blue fluorescence intensity ratio was evaluated from normal tissue (0.3) to hyperplasia (1.0) to SCC (2.0), which makes this parameter suitable for distinguishing between these tissue types with high specificity and sensitivity (over 90%) (Thong et al., 2009). Subsequently, the endomicroscopy imaging technique was improved by the same research team (Thong et al., 2012). Preliminary study already suggested that confocal laser endomicroscopy (CLE) might be a potent approach for the surface and subsurface imaging of oral cavity tissues using various fluorescent dyes in both animal and human models (Thong et al., 2007). Recently, a computing system was interfaced to CLE, which enables the 3-D fluorescence visualization of the oral cavity in real-time (Thong et al., 2012). This system could be integrated into current techniques for oral cancer diagnosis and might ultimately lead to better clinical outcomes.
The fluorescent properties of hypericin and four additional fluorescent dyes were also tested for their ability to characterize normal and cancerous head and neck tissue; however, the staining procedure was performed ex vivo on fresh samples obtained from head and neck surgeries (glossectomy, pharyngolaryngectomy, laryngectomy, etc.). Hypericin accumulated in the cytoplasm of normal and tumor cells, but was the only fluorescent dye that did not stain the anomalies of keratinization. Thus, the authors conclude that hypericin might not be a suitable photosensitizer for use in such head and neck specimens (Abbaci et al., 2015).
However, more promising in vivo results were obtained for intra-operative HY-PDD of rhabdomyosarcoma. In the preclinical study conducted by Urla et al. (2015), mice were injected intraperitoneally with human alveolar rhabdomyosarcoma cells and 3 weeks later, hypericin was administered intravenously. After 24 h, conventional and fluorescence laparoscopy were performed and the tumors were surgically resected using hypericin-mediated red fluorescence as guidance. Tumor specimens were processed for histological analyses. Conventional laparoscopy revealed 24 tumors (ranging in size from 1.6 to 13.5 mm) and 28 tumors were detected only by fluorescence laparoscopy (0.5–11 mm). The results indicate that intraoperative HY-PDD is more sensitive than conventional laparoscopy and can clearly distinguish rhabdomyosarcoma from healthy tissue (Urla et al., 2015). Moreover, the authors inform about clinical trial that will be initiated in children with advanced-stage rhabdomyosarcoma.
Although hypericin has been extensively studied mainly because of its photodynamic and photocytotoxic properties, it also possesses various positive and negative biological activities without light-activation.
As some in vitro studies revealed cytotoxic or growth-inhibitory effects of non-activated hypericin (Blank et al., 2001, 2003; Berlanda et al., 2010; Besic Gyenge et al., 2012) and antiproliferative (Blank et al., 2001) and antimetastatic (Blank et al., 2004) activities in vivo and antiangiogenic actions in vitro (Martínez-Poveda et al., 2005) have been described, hypericin might also show antitumor potential in the absence of light. Moreover, one clinical study has demonstrated antiglioma activity of hypericin in dark conditions (Couldwell et al., 2011). However, the potential use of this natural compound in medicine might be broader. Regarding its avidity to necrotic tissues, the radiolabeled hypericin derivative ([123I]iodohypericin) can be used for radio-imaging of numerous necrosis-related pathologies (acute myocardial infarction, liver infarction) (Ni et al., 2006; Fonge et al., 2008), including solid tumors (Van de Putte et al., 2012).
On the other hand, hypericin can be associated with a decrease in the chemosensitivity of cancer cells because of its ability to induce the expression of some ATP-binding cassette (ABC) transporters, which are well-known multidrug resistance (MDR) components (Jendželovský et al., 2009; Jendželovská et al., 2014; Kuchárová et al., 2015). In addition, the hypericin-mediated attenuation of the cytotoxicity of some chemotherapeutic agents was demonstrated (Jendželovská et al., 2014).
Hypericin-mediated photocytotoxic effects have always been a relevant and attractive issue for researchers in the field of oncology; however, the abilities of hypericin in the absence of light-activation have not been studied as intensively. Although several studies indicate that hypericin might possess some anticancer activities even in dark conditions (Table 5).
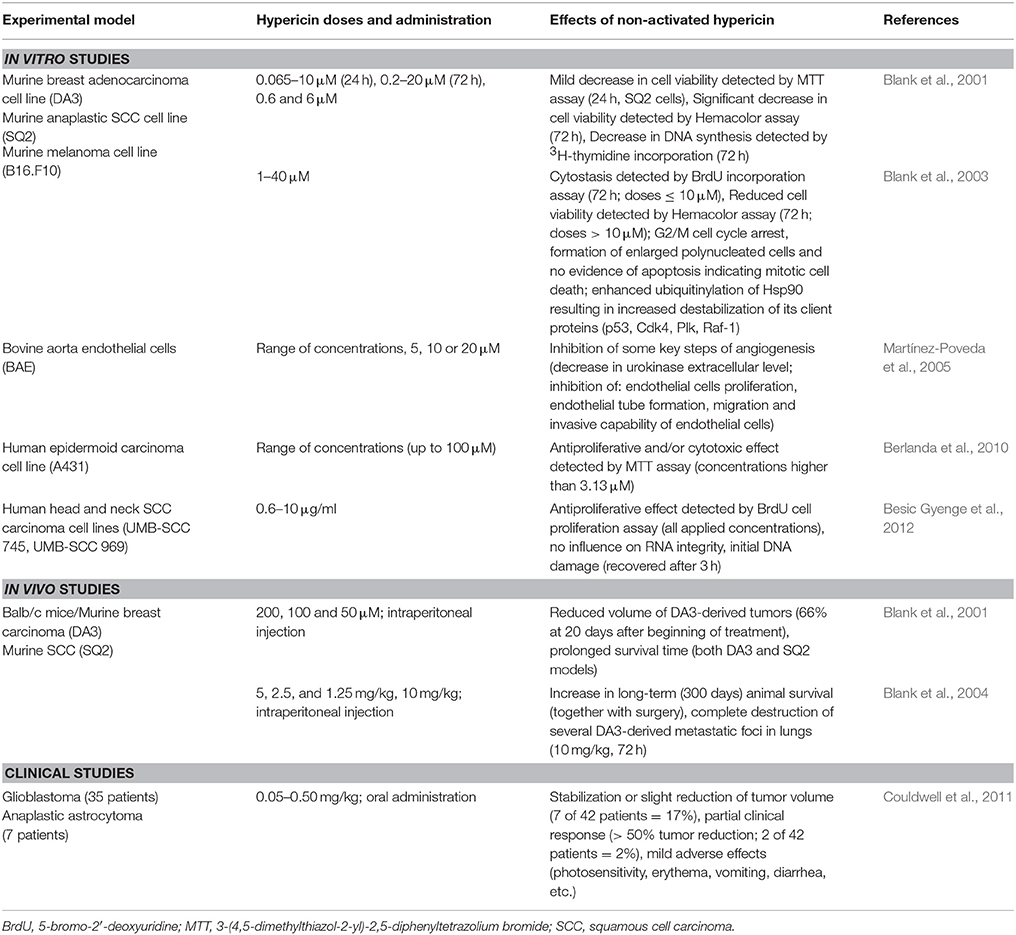
Table 5. Anticancer effects of hypericin in dark conditions.
It has been found that non-activated hypericin possesses no cytotoxicity toward various cancer cell lines at concentrations sufficient for its photocytotoxic action (Thomas and Pardini, 1992; Hadjur et al., 1996; Vandenbogaerde et al., 1997). However, some in vitro studies have shown that hypericin can act as a cytotoxic or antiproliferative agent even in the dark (Blank et al., 2001, 2003; Berlanda et al., 2010; Besic Gyenge et al., 2012). The presence or absence of these effects often strongly depends on the hypericin concentration (higher concentrations are required than for HY-PDT-mediated toxicity), treatment conditions, the applied experimental methods, as well as on the type, origin and sensitivity of cancer cells. Moreover, most in vitro studies have tested the cytotoxicity of non-activated hypericin following its single dose or during relatively short time intervals; however, the situation after multiple fractionated hypericin dosages or during a long-term investigation might be completely different.
Firstly, Blank et al. (2001) demonstrated the cytotoxic and antiproliferative effect of non-activated hypericin both in vitro and in vivo. Hypericin significantly decreased the viability of highly metastatic murine breast adenocarcinoma (DA3HI) and SCC (SQ2) cells. Moreover, the anticancer potential of hypericin was observed even in vivo in DA3HI- and SQ2-derived tumors. Even though hypericin slightly accelerated death in mice with DA3HI-derived tumor development that was very rapid, intraperitoneal hypericin administration (6 declining doses) in other animals led to the inhibition of tumor growth, which was accompanied by prolonged survival time. Moreover, the hypericin-mediated improvement of survival was also evident in mice with high-grade SCC tumors (Blank et al., 2001). Subsequently, Blank et al. (2004) examined the antimetastatic potential of non-activated hypericin and evaluated its influence on long-term survival (up to 300 days). Again, mice with breast adenocarcinoma or SCC tumors, which develop metastases predominantly in the lungs, were used as experimental models. To evaluate the impact of hypericin only on metastases, primary tumors were surgically excised at a stage when micrometastases already existed. Hypericin was administered intraperitoneally in multiple declining dosages (up to 6 doses of hypericin at 5-day intervals). In both cases of tumor origin, hypericin therapy together with the resection of primary tumors resulted in a significant increase in long-term animal survival compared to the untreated control or to those animals that received surgery alone. Moreover, the complete destruction of several, but not all lung metastases, was evident 72 h after hypericin treatment. The results indicate that a single hypericin dose was insufficient to prolong the survival of animals, but fractionated hypericin doses could prevent animal death when administered shortly after the resection of the primary tumor (Blank et al., 2004). Similarly, multiple hypericin doses were applied in a previous study to obtain positive anticancer therapeutic results (Blank et al., 2001).
Considering the potential of hypericin to destroy metastases (Blank et al., 2004), the results are consistent with previous in vitro findings, where a decrease in the viability of DA-3HI and SQ2 cells was also evident 72 h after treatment (Blank et al., 2001). However, no hypericin-mediated induction of apoptosis in the dark was observed. The inhibition of DNA synthesis indicated that the anticancer action of non-activated hypericin in these cell lines was more cytostatic than cytotoxic (Blank et al., 2001). Nevertheless, another in vitro experiment conducted by the same research group established mitotic cell death as the mechanism of hypericin-mediated cytotoxicity. Hypericin was responsible for the enhanced ubiquitinylation of the Hsp90 chaperone, resulting in the destabilization of its client proteins engaged in the regulation of cell proliferation, including p53, Cdk4, Plk, and Raf-1. Ultimately, cytostasis and a decrease in cell viability with no apoptosis were observed. Mitotic cell death is generally characterized by cell-cycle arrest in the G2/M phase, increased cell volume and multinucleation. All these phenotypes were evident in DA3 and SQ2 cells and even in B16.F10 melanoma cells after hypericin treatment (Blank et al., 2003). Thus, mitotic cell death might participate in the tumoricidal and antimetastatic functions of hypericin in the absence of light.
Martínez-Poveda et al. (2005) described another mechanism that might be implicated in the anticancer effects of hypericin in the dark. The results of several in vitro assays indicated that hypericin can inhibit several key steps of angiogenesis, including the proliferation, migration and invasion of endothelial cells, extracellular matrix-degrading urokinase or tubular formation on Matrigel (Martínez-Poveda et al., 2005). All these effects might be beneficial in the prevention of tumor neovascularization.
To our knowledge, one clinical trial has investigated the impact of hypericin on recurrent malignant gliomas (anaplastic astrocytoma and glioblastoma) and has monitored the tolerance of the patients to this treatment. Hypericin was administered orally at gradually increasing dosages ranging from 0.05 to 0.5 mg/kg (once each morning for up to 3 months) and was well-tolerated (mean maximum tolerated daily dose of 0.40 ± 0.098 mg/kg), although some mild skin or gastrointestinal side effects were observed. More importantly, hypericin stabilized or slightly reduced tumor volume (in seven out of 42 patients). In addition, a partial response (>50% reduction of tumor volume) was observed in two patients (Couldwell et al., 2011). These results indicate that synthetic orally administered hypericin can be moderately effective as an adjuvant therapy in cases of malignant glioma; however, further clinical studies are required.
In addition to its preferential accumulation in tumors compared to in normal healthy tissue, hypericin also has a specific strong affinity toward necrotic tissue (Van de Putte et al., 2008a,b,c). The mechanisms of this phenomenon have not yet been fully elucidated; although some hypotheses already exist that consider the binding of hypericin to specific constituents in the necrotic space. Several radiolabeled hypericin derivatives, particularly [123I]iodohypericin (Fonge et al., 2007) and [131I]iodohypericin (Li et al., 2011), possess similar necrosis avidity; thus, hypericin can be used as a potent necrosis-avid contrast agent (NACA) for the non-invasive detection and imaging of various necrosis-related pathologies and diseases or to assess tissue viability and therapeutic responses. Moreover, iodohypericins as NACAs might also be effective in relatively new approaches that combine both tumor diagnosis and therapy, so-called “theranostic” modalities (Table 6).
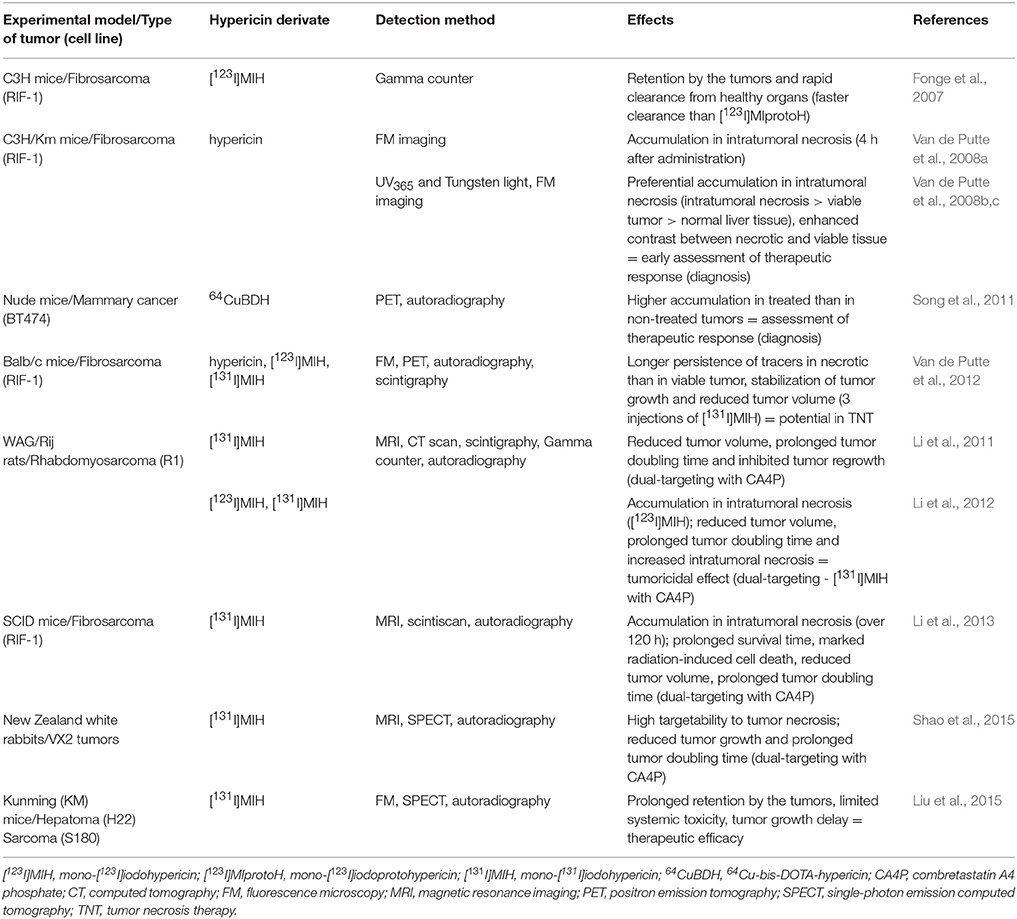
Table 6. Preclinical in vivo studies to test hypericin as a necrosis-avid agent.
In a few initial in vivo studies, hypericin was investigated as a potential indicator of therapeutic responses, following various necrosis-inducing anticancer treatments. Mice bearing intrahepatic fibrosarcoma tumors were used as an experimental model and hypericin was injected intravenously 1 h before, or 24 h after intratumoral ethanol injection (Van de Putte et al., 2008b) or radiofrequency ablation (Van de Putte et al., 2008c), which both induced tumor necrosis. Fluoromacroscopic and fluoromicroscopic examinations confirmed that hypericin accumulated preferentially in necrotic tissue. In the cases of necrosis, mean fluorescence densities were about 4.5- and 5-fold higher than in viable tumor tissue and 14- and 12-fold higher than in normal liver tissue. These results demonstrate the ability of hypericin to enhance the imaging contrast between necrotic and viable tissues and ultimately, its potential role in the early assessment of the therapeutic response (Van de Putte et al., 2008b,c). At about the same time, tumor uptake of radiolabeled hypericin (mono-[123I]iodohypericin) and protohypericin (mono-[123I]iodoprotohypericin) derivatives were tested and compared, because of the applicability of tomographic imaging techniques instead of fluorescence-based techniques. Radioactivity was measured using a gamma counter. Both radiolabeled compounds were retained by the tumors, but mono-[123I]iodohypericin appeared to be a more suitable tumor diagnostic agent, due to its faster clearance from healthy organs (Fonge et al., 2007). Another radiolabeled derivate, 64Cu-bis-DOTA-hypericin, was also applicable in the early determination of the therapeutic response as its accumulation was significantly lower in non-treated tumors than in those treated by photothermal ablation therapy, inducing necrosis (Song et al., 2011).
As necrotic tissue represents 30–80% of the solid tumor mass and is rarely present in normal healthy tissue and organs, it is a suitable target not only for cancer diagnosis, but also for anticancer therapy. The strong avidity of iodohypericins for necrotic tissue makes these compounds very potent in the imaging of tumor necrosis. Furthermore, as hypericin can persist in necrotic tumor areas much longer (up to 72 h) than in viable tumor tissue (up to 24 h) (Van de Putte et al., 2012), its radiolabeled derivatives could be also used for so-called tumor necrosis therapy (TNT). This therapy is based on the destruction of adjacent viable tumor cells by the deposition and accumulation of radiation energy. In other words, attached radioactive iodine “bombards” the neighboring living tumor cells with radiation. Each successive treatment kills more tumor cells, thus, increasing the necrotic region, which allows higher efficacy with each treatment. As three injections of [131I]iodohypericin reduced the volume of RIF-1-derived tumors, this radiolabeled derivate might have potential in TNT (Van de Putte et al., 2012). In addition, similar results, especially the intense retention of [131I]iodohypericin in necrotic tumor tissue (over 168 h) and inhibited tumor growth after a single dose, were obtained by Liu et al. (2015) in mice bearing hepatomas or sarcomas.
Moreover, the approach of a necrosis-based anticancer treatment has been expanded into a dual-targeting theranostic strategy by administering the vascular-disrupting agent prior to the hypericin iododerivate. Firstly, a vascular-disrupting agent, such as combretastatin A4 phosphate (CA4P), targets the tumor microenvironment and subsequently, iodine radioactivity kills residual cancer cells. Several preclinical studies have demonstrated that dual-targeting using CA4P with [131I]iodohypericin was much more effective than single treatments. A reduced tumor volume, a prolonged tumor doubling time, an increase in radiation-induced cell death and intratumoral necrosis or prolonged survival time were reported in rats bearing liver rhabdomyosarcomas (Li et al., 2011, 2012), in mice bearing fibrosarcomas (Li et al., 2013) and in rabbits with liver and muscular VX2 tumors (Shao et al., 2015). Thus, the dual-targeting theranostic approach appears to be well tolerated and can enhance the therapeutic response, encouraging further development for other preclinical and even clinical applications.
All the above-mentioned preclinical and clinical results suggest that hypericin offers great potential in tumor diagnosis as well as in anticancer therapy. However, this secondary metabolite might also cause some other effects that would not be beneficial for therapeutic outcomes. It is well-known that the efficacy of commonly used anticancer treatment modalities is often limited by intrinsic or acquired MDR—a multifactorial phenomenon of the increased tolerance of cancer cells to various tumoricidal agents. A number of cellular mechanisms can contribute to MDR (reviewed in Stavrovskaya, 2000; Zahreddine and Borden, 2013), including the increased elimination of anticancer drugs by tumor cells, which is mostly linked to the elevated expression and/or activity of several ABC transporters. It has been shown that non-activated hypericin can modulate some of these efflux pumps. In vitro experiments conducted by our research group (Jendželovský et al., 2009; Jendželovská et al., 2014; Kuchárová et al., 2015) have revealed an increased expression of multidrug resistance-associated protein 1 (MRP1) and breast cancer resistance protein (BCRP) in colorectal HT-29 cells or in ovarian A2780 and A2780cis cells following hypericin treatment in the dark. In A2780 and A2780cis cells, 0.5 μM hypericin elevated MRP1 protein levels already 6 h after the treatment (Jendželovská et al., 2014). For HT-29 cells, an even lower hypericin concentration (0.1 μM) was sufficient to increase MRP1 and BCRP expression (16 h after hypericin addition) (Jendželovský et al., 2009; Kuchárová et al., 2015). Therefore, because many chemotherapeutic agents and photosensitizers are substrates of the above-mentioned transporters (reviewed in Nies et al., 2007; Robey et al., 2007), the hypericin-mediated stimulation of efflux systems might lead to a decrease in the efficacy of these therapeutic approaches when they are applied at the same time or shortly following hypericin treatment.
Wada et al. (2002) evaluated the impact of hypericin on the action of several anticancer drugs, using the cervical HeLa cell line and its resistant subline Hvr100-6 that overexpress another MDR-related ABC transporter, P-glycoprotein (P-gp), but no effect was observed. Several studies suggest that hypericin can neither modulate P-gp expression nor its activity (Wada et al., 2002; Tian et al., 2005; Jendželovský et al., 2009), which explains its poor ability to influence the cytotoxicity and transport of P-gp substrates, such as paclitaxel, daunorubicin, doxorubicin or vinblastin (Wada et al., 2002). On the other hand, 24 h pre-treatment with hypericin resulted in the attenuation of mitoxantrone cytotoxicity in HL-60 cells and cisplatin cytotoxicity in sensitive A2780 and resistant A2780cis cells (Jendželovská et al., 2014). However, this effect was probably not caused by modulation of the analyzed ABC transporters. However, the results suggest that hypericin in the dark might have a negative impact on the onset or progress of cell death induced by some anticancer agents, possibly by affecting some other mechanisms. Further studies are required, to elucidate the specific mechanisms responsible for the above-mentioned changes and in vivo studies will be necessary to verify the impact of non-activated hypericin on the outcome of chemotherapy.
Moreover, some MDR mechanisms, including ABC transporters, are also involved in drug pharmacokinetics. The modulation of these mechanisms can affect the absorption, distribution or clearance and ultimately, the action of the administered xenobiotics, resulting in negative drug interactions. Several clinical trials have demonstrated the interactions between some chemotherapeutic agents and St. John's wort extract, which is often taken by oncological patients as an antidepressant. In the first study, the enhanced metabolism of irinotecan and consequently, decreased plasma levels of its active metabolite (SN-38) were evaluated in cancer patients following St. John's wort treatment (Mathijssen et al., 2002). Furthermore, Frye et al. (2004) and Smith et al. (2004) examined the effect of St. John's wort extract on the pharmacokinetics of imatinib mesylate in healthy adult volunteers. In both studies, imatinib was administered before and after the treatment with St. John's wort (300 mg three times daily for 2 weeks) and its clearance and half-life was significantly increased or decreased, respectively, by the herb extract. Similar results were obtained by Goey et al. (2014) using a similar experimental design. Besides the enhanced clearance and decreased plasma concentrations of docetaxel in cancer patients, St. John's wort lowered the incidence of docetaxel-mediated toxicities. Thus, due to the risk of potential undertreatment, combining anticancer therapeutic approaches with St. John's wort extracts should not be recommended to oncological patients.
The probable reason for these effects is the induction of the metabolic enzyme (CYP3A4) and/or the P-gp transporter by hyperforin (Komoroski et al., 2004; Tian et al., 2005), another St. John's wort metabolite. However, considering the potential of hypericin to induce the expression of some ABC efflux pumps, this secondary metabolite might also contribute to negative drug interactions with St. John's wort. Therefore, a much broader spectrum of antineoplastic drugs might exist, including various chemotherapeutic agents or photosensitizers, whose action might be altered due to the presence of hypericin.
In this review, we have summarized the medicinal applications of light-activated and non-activated hypericin in the field of oncology. We have preferentially highlighted the “primary beneficial side” of this secondary metabolite concerning its anticancer potential, but have also outlined its “second non-beneficial side,” which was revealed by preliminary in vitro studies.
The vast majority of the results summarized here suggest that hypericin in light and dark conditions might be a very potent agent in cancer treatment and diagnosis. Besides the well-known and intensively investigated HY-PDT and HY-PDD, some new and promising approaches using hypericin as NACA are becoming the focus of various research groups. Moreover, some tested modalities, including dual-targeting, might even improve the clinical outcome of cancer treatments. However, in the dark, hypericin might be responsible for the limited efficacy of conventionally applied chemotherapy or even PDT, due to its ability to induce the expression of some ABC transporters. Moreover, there is a suspicion that non-activated hypericin possesses much broader biological activity. Thus, the chronic usage of St. John's wort extracts as an antidepressant by oncological patients undergoing anticancer treatment should be avoided.
ZJ designed the concept and issue of the review, studied the literature, contributed to all chapters and summarized the bulk of the text, revised the text after it was completed and also following English revision and approved the final version. RJ discussed the concept of the review with first author, studied the literature and contributed to the chapters about HY-PDT and non-activated hypericin, revised the text and approved the final version. BK studied the literature and contributed to the chapters about HY-PDT and non-activated hypericin, revised the text and approved the final version. PF discussed the concept of the review with first author, revised the text and approved the final version.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
This work was supported by the Slovak Research and Development Agency under contract No. APVV-14-0154 and the Scientific Grant Agency of the Ministry of Education of the Slovak Republic under contract No. VEGA 1/0147/15.
1O2, singlet oxygen; 64Cu-bis-DOTA-hypericin, 64Cu-Labeled bis-1, 4, 7, 10-tetraazacyclododecane-N, N′,N,N′-tetraacetic acid conjugated hypericin; ABC, ATP-binding cassette; AK, actinic keratosis; BCC, basal cell carcinoma; BCRP, breast cancer resistance protein; BD, Bowen's disease; CA4P, combretastatin A4 phosphate; Cdk4, cyclin-dependent kinase 4; CIS, carcinoma in situ; CLE, confocal laser endomicroscopy; CYP3A4, cytochrome P450 3A4; DAMPs, damage-associated molecular patterns; DLI, drug–light interval; EGFR, epidermal growth factor receptor; FE, fluorescence endoscopy; Hsp90, heat shock protein 90; HY-PDD, hypericin-mediated photodynamic diagnosis; HY-PDT, hypericin-mediated photodynamic therapy; ICD, immunogenic cell death; INF-α, interferon-α; LIF, laser-induced fluorescence; MDR, multidrug resistance; MF, mycosis fungoides; MRP1, multidrug resistance-associated protein 1; NACA, necrosis-avid contrast agent; , superoxide anion; p53, phosphoprotein p53, tumor suppressor p53, tumor protein p53; PDD, photodynamic diagnosis; PDT, photodynamic therapy; P-gp, P-glycoprotein; Plk, Polo-like kinase; PVP, polyvinylpyrrolidone; Raf-1, serine/threonine kinase, Raf-1 proto-oncogene; ROS, reactive oxygen species; SCC, squamous cell carcinoma; Ser, serine; SN-38, 7-Ethyl-10-hydroxy-camptothecin; TCC, transitional cell carcinoma; Thr, threonine; TNF-α, tumor necrosis growth factor-α; TNT, tumor necrosis therapy; VEGF, vascular endothelial growth factor; WLE, white-light endoscopy.
Abbaci, M., Casiraghi, O., Temam, S., Ferchiou, M., Bosq, J., Dartigues, P., et al. (2015). Red and far-red fluorescent dyes for the characterization of head and neck cancer at the cellular level. J. Oral Pathol. Med. 44, 831–841. doi: 10.1111/jop.12316
Agostinis, P., Berg, K., Cengel, K. A., Foster, T. H., Girotti, A. W., Gollnick, S. O., et al. (2011). Photodynamic therapy of cancer: an update. CA Cancer J. Clin. 61, 250–281. doi: 10.3322/caac.20114
Agostinis, P., Vantieghem, A., Merlevede, W., and de Witte, P. A. (2002). Hypericin in cancer treatment: more light on the way. Int. J. Biochem. Cell Biol. 34, 221–241. doi: 10.1016/S1357-2725(01)00126-1
Alecu, M., Ursaciuc, C., Hãlãlãu, F., Coman, G., Merlevede, W., Waelkens, E., et al. (1998). Photodynamic treatment of basal cell carcinoma and squamous cell carcinoma with hypericin. Anticancer Res. 18, 4651–4654.
Ali, S. M., and Olivo, M. (2002). Bio-distribution and subcellular localization of Hypericin and its role in PDT induced apoptosis in cancer cells. Int. J. Oncol. 21, 531–540. doi: 10.3892/ijo.21.3.531
Ayan, A. K., Cirak, C., Kevseroglu, K., and Ozen, T. (2004). Hypericin in some Hypericum species from Turkey. Asian J. Plant Sci. 3, 200–202. doi: 10.3923/ajps.2004.200.202
Berlanda, J., Kiesslich, T., Engelhardt, V., Krammer, B., and Plaetzer, K. (2010). Comparative in vitro study on the characteristics of different photosensitizers employed in PDT. J. Photochem. Photobiol. B. Biol. 100, 173–180. doi: 10.1016/j.jphotobiol.2010.06.004
Besic Gyenge, E., Forny, P., Lüscher, D., Laass, A., Walt, H., and Maake, C. (2012). Effects of hypericin and a chlorin based photosensitizer alone or in combination in squamous cell carcinoma cells in the dark. Photodiagnosis Photodyn. Ther. 9, 321–331. doi: 10.1016/j.pdpdt.2012.03.006
Bhuvaneswari, R., Gan, Y. Y., Lucky, S. S., Chin, W. W., Ali, S. M., Soo, K. C., et al. (2008). Molecular profiling of angiogenesis in hypericin mediated photodynamic therapy. Mol. Cancer. 7:56. doi: 10.1186/1476-4598-7-56
Bhuvaneswari, R., Thong, P. S., Gan, Y. Y., Soo, K. C., and Olivo, M. (2010). Evaluation of hypericin-mediated photodynamic therapy in combination with angiogenesis inhibitor bevacizumab using in vivo fluorescence confocal endomicroscopy. J. Biomed. Opt. 15:011114. doi: 10.1117/1.3281671
Bhuvaneswari, R., Yuen, G. Y., Chee, S. K., and Olivo, M. (2011). Antiangiogenesis agents avastin and erbitux enhance the efficacy of photodynamic therapy in a murine bladder tumor model. Lasers Surg. Med. 43, 651–662. doi: 10.1002/lsm.21109
Blank, M., Kostenich, G., Lavie, G., Kimel, S., Keisari, Y., and Orenstein, A. (2002). Wavelength-dependent properties of photodynamic therapy using hypericin in vitro and in an animal model. Photochem. Photobiol. 76, 335–340. doi: 10.1562/0031-8655(2002)0760335WDPOPT2.0.CO2
Blank, M., Lavie, G., Mandel, M., Hazan, S., Orenstein, A., Meruelo, D., et al. (2004). Antimetastatic activity of the photodynamic agent hypericin in the dark. Int. J. Cancer. 111, 596–603. doi: 10.1002/ijc.20285
Blank, M., Mandel, M., Hazan, S., Keisari, Y., and Lavie, G. (2001). Anti-cancer activities of hypericin in the dark. Photochem. Photobiol. 74, 120–125. doi: 10.1562/0031-8655(2001)0740120ACAOHI2.0.CO2
Blank, M., Mandel, M., Keisari, Y., Meruelo, D., and Lavie, G. (2003). Enhanced ubiquitinylation of heat shock protein 90 as a potential mechanism for mitotic cell death in cancer cells induced with hypericin. Cancer Res. 63, 8241–8247.
Bobrov, N., Cavarga, I., Longauer, F., Rybárová, S., Fedorocko, P., Brezáni, P., et al. (2007). Histomorphological changes in murine fibrosarcoma after hypericin-based photodynamic therapy. Phytomedicine 14, 172–178. doi: 10.1016/j.phymed.2006.09.017
Boiy, A., Roelandts, R., and de Witte, P. A. (2010). Photodynamic therapy using topically applied hypericin: comparative effect with methyl-aminolevulinic acid on UV induced skin tumours. J. Photochem. Photobiol. B. Biol. 102, 123–131. doi: 10.1016/j.jphotobiol.2010.09.012
Brockmann, H., Haschad, M. N., Maier, K., and Pohl, F. (1939). Über das Hypericin, den photodynamisch wirksamen Farbstoff aus Hypericum perforatum. Naturwissenschaften 27, 550. doi: 10.1007/BF01495453
Bruňáková, K., Petijová, L., Zámečník, J., Turečková, V., and Čellárová, E. (2015). The role of ABA in the freezing injury avoidance in two Hypericum species differing in frost tolerance and potential to synthesize hypericins. Plant Cell Tissue Organ Cult. 122, 45–56. doi: 10.1007/s11240-015-0748-9
Buytaert, E., Callewaert, G., Hendrickx, N., Scorrano, L., Hartmann, D., Missiaen, L., et al. (2006). Role of endoplasmic reticulum depletion and multidomain proapoptotic BAX and BAK proteins in shaping cell death after hypericin-mediated photodynamic therapy. FASEB J. 20, 756–758. doi: 10.1096/fj.05-4305fje
Čavarga, I., Brezáni, P., Cekanová-Figurová, M., Solár, P., Fedorocko, P., and Miskovský, P. (2001). Photodynamic therapy of murine fibrosarcoma with topical and systemic administration of hypericin. Phytomedicine 8, 325–330. doi: 10.1078/0944-7113-00057
Čavarga, I., Brezáni, P., Fedorocko, P., Miskovský, P., Bobrov, N., Longauer, F., et al. (2005). Photoinduced antitumour effect of hypericin can be enhanced by fractionated dosing. Phytomedicine 12, 680–683. doi: 10.1016/j.phymed.2004.02.011
Čellárová, E. (2011). Effect of exogenous morphogenetic signals on differentiation in vitro and secondary metabolite formation in the genus Hypericum in Medicinal and Aromatic Plant Science and Biotechnology 5 (Special Issue 1), eds M. S. Odabas and C. Çırak (Ikenobe: Global Science Books), 62–69.
Čellárová, E., Brutovská, R., Daxnerová, Z., Bruňáková, K., and Weigel, R. C. (1997). Correlation between hypericin content and the ploidy of somaclones of Hypericum perforatum L. Acta Biotechnol. 17, 83–90. doi: 10.1002/abio.370170111
Chen, B., and de Witte, P. A. (2000). Photodynamic therapy efficacy and tissue distribution of hypericin in a mouse P388 lymphoma tumor model. Cancer Lett. 150, 111–117. doi: 10.1016/S0304-3835(99)00381-X
Chen, B., Roskams, T., and de Witte, P. A. (2002a). Antivascular tumor eradication by hypericin-mediated photodynamic therapy. Photochem. Photobiol. 76, 509–513. doi: 10.1562/0031-8655(2002)0760509ATEBHM2.0.CO2
Chen, B., Roskams, T., Xu, Y., Agostinis, P., and de Witte, P. A. (2002b). Photodynamic therapy with hypericin induces vascular damage and apoptosis in the RIF-1 mouse tumor model. Int. J. Cancer. 98, 284–290. doi: 10.1002/ijc.10175
Chen, B., Xu, Y., Roskams, T., Delaey, E., Agostinis, P., Vandenheede, J. R., et al. (2001). Efficacy of antitumoral photodynamic therapy with hypericin: relationship between biodistribution and photodynamic effects in the RIF-1 mouse tumor model. Int. J. Cancer. 93, 275–282. doi: 10.1002/ijc.1324
Cole, C. D., Liu, J. K., Sheng, X., Chin, S. S., Schmidt, M. H., Weiss, M. H., et al. (2008). Hypericin-mediated photodynamic therapy of pituitary tumors: preclinical study in a GH4C1 rat tumor model. J. Neurooncol. 87, 255–261. doi: 10.1007/s11060-007-9514-0
Couldwell, W. T., Surnock, A. A., Tobia, A. J., Cabana, B. E., Stillerman, C. B., Forsyth, P. A., et al. (2011). A phase 1/2 study of orally administered synthetic hypericin for treatment of recurrent malignant gliomas. Cancer 117, 4905–4915. doi: 10.1002/cncr.26123
Crnolatac, I., Huygens, A., van Aerschot, A., Busson, R., Rozenski, J., and de Witte, P. A. (2005). Synthesis, in vitro cellular uptake and photo-induced antiproliferative effects of lipophilic hypericin acid derivatives. Bioorg. Med. Chem. 13, 6347–6353. doi: 10.1016/j.bmc.2005.09.003
Delaey, E., Vandenbogaerde, A., Merlevede, W., and de Witte, P. (2000). Photocytotoxicity of hypericin in normoxic and hypoxic conditions. J. Photochem. Photobiol. B. Biol. 56, 19–24. doi: 10.1016/S1011-1344(00)00051-8
Dewick, P. M. (2002). Medicinal Natural Products: A Biosynthetic Approach, 2nd Edn. Chichester: John Wiley & Sons Ltd.
D'Hallewin, M. A., De Witte, P. A., Waelkens, E., Merlevede, W., and Baert, L. (2000). Fluorescence detection of flat bladder carcinoma in situ after intravesical instillation of hypericin. J. Urol. 164, 349–351. doi: 10.1016/S0022-5347(05)67357-0
D'Hallewin, M. A., Kamuhabwa, A. R., Roskams, T., De Witte, P. A., and Baert, L. (2002). Hypericin-based fluorescence diagnosis of bladder carcinoma. BJU Int. 89, 760–763. doi: 10.1046/j.1464-410X.2002.02690.x
Diwu, Z., and Lown, J. W. (1993). Photosensitization with anticancer agents. 17. EPR studies of photodynamic action of hypericin: formation of semiquinone radical and activated oxygen species on illumination. Free Radic. Biol. Med. 14, 209–215. doi: 10.1016/0891-5849(93)90012-J
Du, H. Y., Bay, B. H., and Olivo, M. (2003a). Biodistribution and photodynamic therapy with hypericin in a human NPC murine tumor model. Int. J. Oncol. 22, 1019–1024. doi: 10.3892/ijo.22.5.1019
Du, H. Y., Olivo, M., Tan, B. K., and Bay, B. H. (2003b). Hypericin-mediated photodynamic therapy induces lipid peroxidation and necrosis in nasopharyngeal cancer. Int. J. Oncol. 23, 1401–1405. doi: 10.3892/ijo.23.5.1401
Feruszová, J., Imreová, P., Bodnárová, K., Ševčovičová, A., Kyzek, S., Chalupa, I., et al. (2016). Photoactivated hypericin is not genotoxic. Gen. Physiol. Biophys. 35, 223–230. doi: 10.4149/gpb_2015045
Fonge, H., Van de Putte, M., Huyghe, D., Bormans, G., Ni, Y., de Witte, P., et al. (2007). Evaluation of tumor affinity of mono-[(123)I]iodohypericin and mono-[(123)I]iodoprotohypericin in a mouse model with a RIF-1 tumor. Contrast Media Mol. Imaging. 2, 113–119. doi: 10.1002/cmmi.136
Fonge, H., Vunckx, K., Wang, H., Feng, Y., Mortelmans, L., Nuyts, J., et al. (2008). Non-invasive detection and quantification of acute myocardial infarction in rabbits using mono-[123I]iodohypericin microSPECT. Eur. Heart J. 29, 260–269. doi: 10.1093/eurheartj/ehm588
Frye, R. F., Fitzgerald, S. M., Lagattuta, T. F., Hruska, M. W., and Egorin, M. J. (2004). Effect of St John's wort on imatinib mesylate pharmacokinetics. Clin. Pharmacol. Ther. 76, 323–329. doi: 10.1016/j.clpt.2004.06.007
Fu, C. Y., Ng, B. K., Razul, S. G., Chin, W. W., Tan, P. H., Lau, W. K., et al. (2007). Fluorescence detection of bladder cancer using urine cytology. Int. J. Oncol. 31, 525–530. doi: 10.3892/ijo.31.3.525
Galanou, M. C., Theodossiou, T. A., Tsiourvas, D., Sideratou, Z., and Paleos, C. M. (2008). Interactive transport, subcellular relocation and enhanced phototoxicity of hypericin encapsulated in guanidinylated liposomes via molecular recognition. Photochem. Photobiol. 84, 1073–1083. doi: 10.1111/j.1751-1097.2008.00392.x
Galluzzi, L., Vacchelli, E., Bravo-San Pedro, J. M., Buqué, A., Senovilla, L., Baracco, E. E., et al. (2014). Classification of current anticancer immunotherapies. Oncotarget 5, 12472–12508. doi: 10.18632/oncotarget.2998
García, I., Ballesta, S., Gilaberte, Y., Rezusta, A., and Pascual, Á. (2015). Antimicrobial photodynamic activity of hypericin against methicillin-susceptible and resistant Staphylococcus aureus biofilms. Future Microbiol. 10, 347–356. doi: 10.2217/fmb.14.114
Garg, A. D., and Agostinis, P. (2014). ER stress, autophagy and immunogenic cell death in photodynamic therapy-induced anti-cancer immune responses. Photochem. Photobiol. Sci. 13, 474–487. doi: 10.1039/c3pp50333j
Garg, A. D., Elsen, S., Krysko, D. V., Vandenabeele, P., de Witte, P., and Agostinis, P. (2015b). Resistance to anticancer vaccination effect is controlled by a cancer cell-autonomous phenotype that disrupts immunogenic phagocytic removal. Oncotarget 6, 26841–26860. doi: 10.18632/oncotarget.4754
Garg, A. D., Galluzzi, L., Apetoh, L., Baert, T., Birge, R. B., Bravo-San Pedro, J. M., et al. (2015a). Molecular and translational classifications of DAMPs in immunogenic cell death. Front. Immunol. 6:588. doi: 10.3389/fimmu.2015.00588
Garg, A. D., Krysko, D. V., Vandenabeele, P., and Agostinis, P. (2012b). Hypericin-based photodynamic therapy induces surface exposure of damage-associated molecular patterns like HSP70 and calreticulin. Cancer Immunol. Immunother. 61, 215–221. doi: 10.1007/s00262-011-1184-2
Garg, A. D., Krysko, D. V., Vandenabeele, P., and Agostinis, P. (2016). Extracellular ATP and P2X7 receptor exert context-specific immunogenic effects after immunogenic cancer cell death. Cell Death Dis. 7, e2097. doi: 10.1038/cddis.2015.411
Garg, A. D., Krysko, D. V., Verfaillie, T., Kaczmarek, A., Ferreira, G. B., Marysael, T., et al. (2012a). A novel pathway combining calreticulin exposure and ATP secretion in immunogenic cancer cell death. EMBO J. 31, 1062–1079. doi: 10.1038/emboj.2011.497
Garnica, S., Weiss, M., and Oberwinkler, F. (2003). Morphological and molecular phylogenetic studies in South American Cortinarius species. Mycol. Res. 107, 1143–1156. doi: 10.1017/S0953756203008414
Goey, A. K., Meijerman, I., Rosing, H., Marchetti, S., Mergui-Roelvink, M., Keessen, M., et al. (2014). The effect of St John's wort on the pharmacokinetics of docetaxel. Clin. Pharmacokinet. 53, 103–110. doi: 10.1007/s40262-013-0102-5
Gulick, R. M., McAuliffe, V., Holden-Wiltse, J., Crumpacker, C., Liebes, L., Stein, D. S., et al. (1999). Phase I studies of hypericin, the active compound in St. John's Wort, as an antiretroviral agent in HIV-infected adults. AIDS Clinical Trials Group Protocols 150 and 258. Ann Intern Med. 130, 510–514. doi: 10.7326/0003-4819-130-6-199903160-00015
Hadjur, C., Richard, M. J., Parat, M. O., Jardon, P., and Favier, A. (1996). Photodynamic effects of hypericin on lipid peroxidation and antioxidant status in melanoma cells. Photochem. Photobiol. 64, 375–381. doi: 10.1111/j.1751-1097.1996.tb02474.x
Head, C. S., Luu, Q., Sercarz, J., and Saxton, R. (2006). Photodynamic therapy and tumor imaging of hypericin-treated squamous cell carcinoma. World J. Surg. Oncol. 4:87. doi: 10.1186/1477-7819-4-87
Hudson, J. B., Harris, L., and Towers, G. H. (1993). The importance of light in the anti-HIV effect of hypericin. Antiviral Res. 20, 173–178. doi: 10.1016/0166-3542(93)90006-5
Huygens, A., Kamuhabwa, A. R., Van Laethem, A., Roskams, T., Van Cleynenbreugel, B., Van Poppel, H., et al. (2005). Enhancing the photodynamic effect of hypericin in tumour spheroids by fractionated light delivery in combination with hyperoxygenation. Int. J. Oncol. 26, 1691–1697. doi: 10.3892/ijo.26.6.1691
Jacobson, J. M., Feinman, L., Liebes, L., Ostrow, N., Koslowski, V., Tobia, A., et al. (2001). Pharmacokinetics, safety, and antiviral effects of hypericin, a derivative of St. John's wort plant, in patients with chronic hepatitis C virus infection. Antimicrob. Agents Chemother. 45, 517–524. doi: 10.1128/AAC.45.2.517-524.2001
Jendželovská, Z., Jendželovský, R., Hil'ovská, L., Koval', J., Mikeš, J., and Fedoročko, P. (2014). Single pre-treatment with hypericin, a St. John's wort secondary metabolite, attenuates cisplatin- and mitoxantrone-induced cell death in A2780, A2780cis and HL-60 cells. Toxicol. In vitro 28, 1259–1273. doi: 10.1016/j.tiv.2014.06.011
Jendželovský, R., Mikes, J., Koval,', J., Soucek, K., Procházková, J., Kello, M., et al. (2009). Drug efflux transporters, MRP1 and BCRP, affect the outcome of hypericin-mediated photodynamic therapy in HT-29 adenocarcinoma cells. Photochem. Photobiol. Sci. 8, 1716–1723. doi: 10.1039/b9pp00086k
Kacerovská, D., Pizinger, K., Majer, F., and Smíd, F. (2008). Photodynamic therapy of nonmelanoma skin cancer with topical Hypericum perforatum extract–a pilot study. Photochem. Photobiol. 84, 779–785. doi: 10.1111/j.1751-1097.2007.00260.x
Kamuhabwa, A. A., Cosserat-Gerardin, I., Didelon, J., Notter, D., Guillemin, F., Roskams, T., et al. (2002). Biodistribution of hypericin in orthotopic transitional cell carcinoma bladder tumors: implication for whole bladder wall photodynamic therapy. Int. J. Cancer. 97, 253–260. doi: 10.1002/ijc.1594
Kamuhabwa, A. A., Di Mavungu, J. D., Baert, L., D'Hallewin, M. A., Hoogmartens, J., and de Witte, P. A. (2005). Determination of hypericin in human plasma by high-performance liquid chromatography after intravesical administration in patients with transitional cell carcinoma of the bladder. Eur. J. Pharm. Biopharm. 59, 469–474. doi: 10.1016/j.ejpb.2004.09.013
Kamuhabwa, A. A., Roskams, T., D'Hallewin, M. A., Baert, L., Van Poppel, H., and de Witte, P. A. (2003). Whole bladder wall photodynamic therapy of transitional cell carcinoma rat bladder tumors using intravesically administered hypericin. Int. J. Cancer. 107, 460–467. doi: 10.1002/ijc.11396
Kascakova, S., Nadova, Z., Mateasik, A., Mikes, J., Huntosova, V., Refregiers, M., et al. (2008). High level of low-density lipoprotein receptors enhance hypericin uptake by U-87 MG cells in the presence of LDL. Photochem. Photobiol. 84, 120–127. doi: 10.1111/j.1751-1097.2007.00207.x
Kashef, N., Borghei, Y. S., and Djavid, G. E. (2013). Photodynamic effect of hypericin on the microorganisms and primary human fibroblasts. Photodiagnosis Photodyn. Ther. 10, 150–155. doi: 10.1016/j.pdpdt.2012.11.007
Kasper, S., Caraci, F., Forti, B., Drago, F., and Aguglia, E. (2010). Efficacy and tolerability of Hypericum extract for the treatment of mild to moderate depression. Eur. Neuropsychopharmacol. 20, 747–765. doi: 10.1016/j.euroneuro.2010.07.005
Kim, M., Jung, H. Y., and Park, H. J. (2015). Topical PDT in the treatment of benign skin diseases: principles and new applications. Int. J. Mol. Sci. 16, 23259–23278. doi: 10.3390/ijms161023259
Kitanov, G. M. (2001). Hypericin and pseudohypericin in some Hypericum species. Biochem. Syst. Ecol. 29, 171–178. doi: 10.1016/S0305-1978(00)00032-6
Kleemann, B., Loos, B., Scriba, T. J., Lang, D., and Davids, L. M. (2014). St John's Wort (Hypericum perforatum L.) photomedicine: hypericin-photodynamic therapy induces metastatic melanoma cell death. PLoS ONE 9:e103762. doi: 10.1371/journal.pone.0103762
Komoroski, B. J., Zhang, S., Cai, H., Hutzler, J. M., Frye, R., Tracy, T. S., et al. (2004). Induction and inhibition of cytochromes P450 by the St. John's wort constituent hyperforin in human hepatocyte cultures. Drug Metab. Dispos. 32, 512–518. doi: 10.1124/dmd.32.5.512
Koperdáková, J., Komarovská, H., Košuth, J., Giovannini, A., and Čellárová, E. (2009). Characterization of hairy root-phenotype in transgenic Hypericum perforatum L. clones. Acta Physiol. Plant. 31, 351–358. doi: 10.1007/s11738-008-0241-8
Košuth, J., Koperdáková, J., Tolonen, A., Hohtola, A., and Cellárová, E. (2003). The content of hypericins and phloroglucinols in Hypericum perforatum L. seedlings at early stage of development. Plant Sci. 165, 515–521. doi: 10.1016/S0168-9452(03)00210-3
Koval, J., Mikes, J., Jendzelovský, R., Kello, M., Solár, P., and Fedorocko, P. (2010). Degradation of HER2 receptor through hypericin-mediated photodynamic therapy. Photochem. Photobiol. 86, 200–205. doi: 10.1111/j.1751-1097.2009.00639.x
Kubin, A., Meissner, P., Wierrani, F., Burner, U., Bodenteich, A., Pytel, A., et al. (2008). Fluorescence diagnosis of bladder cancer with new water soluble hypericin bound to polyvinylpyrrolidone: PVP-hypericin. Photochem. Photobiol. 84, 1560–1563. doi: 10.1111/j.1751-1097.2008.00384.x
Kubin, A., Wierrani, F., Burner, U., Alth, G., and Grünberger, W. (2005). Hypericin–the facts about a controversial agent. Curr. Pharm. Des. 11, 233–253. doi: 10.2174/1381612053382287
Kucharikova, A., Kimakova, K., Janfelt, C., and Čellarova, E. (2016). Interspecific variation in localization of hypericins and phloroglucinols in the genus Hypericum as revealed by Desorption Electrospray Ionization Mass Spectrometry imaging. Physiol Plant. 157, 2–12. doi: 10.1111/ppl.12422
Kuchárová, B., Mikeš, J., Jendželovský, R., Vargová, J., Mikešová, L., Jendželovská, Z., et al. (2015). Potentiation of hypericin-mediated photodynamic therapy cytotoxicity by MK-886: focus on ABC transporters, GDF-15 and redox status. Photodiagnosis Photodyn. Ther. 12, 490–503. doi: 10.1016/j.pdpdt.2015.04.008
Kusari, S., Lamshöft, M., Zühlke, S., and Spiteller, M. (2008). An endophytic fungus from Hypericum perforatum that produces hypericin. J. Nat. Prod. 71, 159–162. doi: 10.1021/np070669k
Kusari, S., Sezgin, S., Nigutová, K., Čellárová, E., and Spiteller, M. (2015). Spatial chemo-profiling of hypericin and related phytochemicals in Hypericum species using MALDI-HRMS imaging. Anal. Bioanal. Chem. 407, 4779–4791. doi: 10.1007/s00216-015-8682-6
Kusari, S., Zühlke, S., Košuth, J., Čellárová, E., and Spiteller, M. (2009). Light-independent metabolomics of endophytic Thielavia subthermophila provides insight into microbial hypericin biosynthesis. J. Nat. Prod. 72, 1825–1835. doi: 10.1021/np9002977
Li, J., Cona, M. M., Chen, F., Feng, Y., Zhou, L., Yu, J., et al. (2012). Exploring theranostic potentials of radioiodinated hypericin in rodent necrosis models. Theranostics 2, 1010–1019. doi: 10.7150/thno.4924
Li, J., Cona, M. M., Chen, F., Feng, Y., Zhou, L., Zhang, G., et al. (2013). Sequential systemic administrations of combretastatin A4 Phosphate and radioiodinated hypericin exert synergistic targeted theranostic effects with prolonged survival on SCID mice carrying bifocal tumor xenografts. Theranostics 3, 127–137. doi: 10.7150/thno.5790
Li, J., Sun, Z., Zhang, J., Shao, H., Cona, M. M., Wang, H., et al. (2011). A dual-targeting anticancer approach: soil and seed principle. Radiology 260, 799–807. doi: 10.1148/radiol.11102120
Liu, C. D., Kwan, D., Saxton, R. E., and McFadden, D. W. (2000). Hypericin and photodynamic therapy decreases human pancreatic cancer in vitro and in vivo. J. Surg. Res. 93, 137–143. doi: 10.1006/jsre.2000.5949
Liu, W., Zhang, D., Feng, Y., Li, Y., Huang, D., Jiang, C., et al. (2015). Biodistribution and anti-tumor efficacy of intratumorally injected necrosis-avid theranostic agent radioiodinated hypericin in rodent tumor models. J. Drug Target. 23, 371–379. doi: 10.3109/1061186X.2014.1000337
Lukšienė, Ž., and De Witte, P. (2002). Hypericin-based photodynamic therapy: I. Comparative antitumor activity uptake studies in Ehrlich ascite tumor. Acta Med Litu. 9, 195–199.
Martínez-Poveda, B., Quesada, A. R., and Medina, M. A. (2005). Hypericin in the dark inhibits key steps of angiogenesis in vitro. Eur. J. Pharmacol. 516, 97–103. doi: 10.1016/j.ejphar.2005.03.047
Mathijssen, R. H., Verweij, J., de Bruijn, P., Loos, W. J., and Sparreboom, A. (2002). Effects of St. John's wort on irinotecan metabolism. J. Natl. Cancer Inst. 94, 1247–1249. doi: 10.1093/jnci/94.16.1247
Miadokova, E., Chalupa, I., Vlckova, V., Sevcovicova, A., Nadova, S., Kopaskova, M., et al. (2010). Genotoxicity and antigenotoxicity evaluation of non-photoactivated hypericin. Phytother. Res. 24, 90–95. doi: 10.1002/ptr.2901
Mikeš, J., Hýžd'alová, M., Kočí, L., Jendželovský, R., Koval', J., Vaculová, A., et al. (2011). Lower sensitivity of FHC fetal colon epithelial cells to photodynamic therapy compared to HT-29 colon adenocarcinoma cells despite higher intracellular accumulation of hypericin. Photochem. Photobiol. Sci. 10, 626–632. doi: 10.1039/c0pp00359j
Mikeš, J., Jendželovský, R., and Fedoročko, P. (2013). Cellular aspects of photodynamic therapy with hypericin in Photodynamic Therapy: New Research, ed M. L. T. Elsaie (New York, NY: Nova Science Publishers), 111–147.
Mikeš, J., Kleban, J., Sacková, V., Horváth, V., Jamborová, E., Vaculová, A., et al. (2007). Necrosis predominates in the cell death of human colon adenocarcinoma HT-29 cells treated under variable conditions of photodynamic therapy with hypericin. Photochem. Photobiol. Sci. 6, 758–766. doi: 10.1039/B700350A
Mikeš, J., Koval', J., Jendzelovský, R., Sacková, V., Uhrinová, I., Kello, M., et al. (2009). The role of p53 in the efficiency of photodynamic therapy with hypericin and subsequent long-term survival of colon cancer cells. Photochem. Photobiol. Sci. 8, 1558–1567. doi: 10.1039/b9pp00021f
Mikešová, L., Mikeš, J., Koval', J., Gyurászová, K., Culka, L., Vargová, J., et al. (2013). Conjunction of glutathione level, NAD(P)H/FAD redox status and hypericin content as a potential factor affecting colon cancer cell resistance to photodynamic therapy with hypericin. Photodiagnosis Photodyn. Ther. 10, 470–483. doi: 10.1016/j.pdpdt.2013.04.003
Ni, Y., Huyghe, D., Verbeke, K., de Witte, P. A., Nuyts, J., Mortelmans, L., et al. (2006). First preclinical evaluation of mono-[123I]iodohypericin as a necrosis-avid tracer agent. Eur. J. Nucl. Med. Mol. Imaging 33, 595–601. doi: 10.1007/s00259-005-0013-2
Nies, A. T., Rius, M., and Keppler, D. (2007). Multidrug resistance proteins of the ABCC subfamily in Drug Transporters: Molecular Characterization and Role in Drug Disposition, eds G. You and M. E. Morris (New Jersey, NJ: John Wiley & Sons, Inc.), 223–262.
Noell, S., Feigl, G. C., Serifi, D., Mayer, D., Naumann, U., Göbel, W., et al. (2013). Microendoscopy for hypericin fluorescence tumor diagnosis in a subcutaneous glioma mouse model. Photodiagnosis Photodyn. Ther. 10, 552–560. doi: 10.1016/j.pdpdt.2013.06.001
Noell, S., Mayer, D., Strauss, W. S., Tatagiba, M. S., and Ritz, R. (2011). Selective enrichment of hypericin in malignant glioma: pioneering in vivo results. Int. J. Oncol. 38, 1343–1348. doi: 10.3892/ijo.2011.968
Olivo, M., Lau, W., Manivasager, V., Bhuvaneswari, R., Wei, Z., Soo, K. C., et al. (2003a). Novel photodynamic diagnosis of bladder cancer: ex vivo fluorescence cytology using hypericin. Int. J. Oncol. 23, 1501–1504. doi: 10.3892/ijo.23.6.1501
Olivo, M., Lau, W., Manivasager, V., Tan, P. H., Soo, K. C., and Cheng, C. (2003b). Macro-microscopic fluorescence of human bladder cancer using hypericin fluorescence cystoscopy and laser confocal microscopy. Int. J. Oncol. 23, 983–990. doi: 10.3892/ijo.23.4.983
Paz-Cristobal, M. P., Gilaberte, Y., Alejandre, C., Pardo, J., Revillo, M. J., and Rezusta, A. (2014). In vitro fungicidal photodynamic effect of hypericin on Trichophyton spp. Mycopathologia 178, 221–225. doi: 10.1007/s11046-014-9797-6
Prince, A. M., Pascual, D., Meruelo, D., Liebes, L., Mazur, Y., Dubovi, E., et al. (2000). Strategies for evaluation of enveloped virus inactivation in red cell concentrates using hypericin. Photochem. Photobiol. 71, 188–195. doi: 10.1562/0031-8655(2000)0710188SFEOEV2.0.CO2
Pytel, A., and Schmeller, N. (2002). New aspect of photodynamic diagnosis of bladder tumors: fluorescence cytology. Urology 59, 216–219. doi: 10.1016/S0090-4295(01)01528-X
Rezusta, A., López-Chicón, P., Paz-Cristobal, M. P., Alemany-Ribes, M., Royo-Díez, D., Agut, M., et al. (2012). In vitro fungicidal photodynamic effect of hypericin on Candida species. Photochem. Photobiol. 88, 613–619. doi: 10.1111/j.1751-1097.2011.01053.x
Ritz, R., Daniels, R., Noell, S., Feigl, G. C., Schmidt, V., Bornemann, A., et al. (2012). Hypericin for visualization of high grade gliomas: first clinical experience. Eur. J. Surg. Oncol. 38, 352–360. doi: 10.1016/j.ejso.2011.12.021
Robey, R. W., Polgar, O., Deeken, J., To, K. K. W., and Bates, S. (2007). Breast cancer resistance protein, in Drug Transporters: Molecular Characterization and Role in Drug Disposition, eds G. You and M. E. Morris (New Jersey, NJ: John Wiley & Sons, Inc.), 319–358. doi: 10.1002/9780470140505.ch12
Rook, A. H., Wood, G. S., Duvic, M., Vonderheid, E. C., Tobia, A., and Cabana, B. (2010). A phase II placebo-controlled study of photodynamic therapy with topical hypericin and visible light irradiation in the treatment of cutaneous T-cell lymphoma and psoriasis. J. Am. Acad. Dermatol. 63, 984–990. doi: 10.1016/j.jaad.2010.02.039
Rubio, N., Coupienne, I., Di Valentin, E., Heirman, I., Grooten, J., Piette, J., et al. (2012). Spatiotemporal autophagic degradation of oxidatively damaged organelles after photodynamic stress is amplified by mitochondrial reactive oxygen species. Autophagy 8, 1312–1324. doi: 10.4161/auto.20763
Sacková, V., Fedorocko, P., Szilárdiová, B., Mikes, J., and Kleban, J. (2006). Hypericin-induced photocytotoxicity is connected with G2/M arrest in HT-29 and S-phase arrest in U937 cells. Photochem. Photobiol. 82, 1285–1291. doi: 10.1562/2006-02-22-RA-806
Sanovic, R., Verwanger, T., Hartl, A., and Krammer, B. (2011). Low dose hypericin-PDT induces complete tumor regression in BALB/c mice bearing CT26 colon carcinoma. Photodiagnosis Photodyn. Ther. 8, 291–296. doi: 10.1016/j.pdpdt.2011.04.003
Sattler, S., Schaefer, U., Schneider, W., Hoelzl, J., and Lehr, C. M. (1997). Binding, uptake, and transport of hypericin by Caco-2 cell monolayers. J. Pharm. Sci. 86, 1120–1126. doi: 10.1021/js970004a
Shao, H., Zhang, J., Sun, Z., Chen, F., Dai, X., Li, Y., et al. (2015). Necrosis targeted radiotherapy with iodine-131-labeled hypericin to improve anticancer efficacy of vascular disrupting treatment in rabbit VX2 tumor models. Oncotarget 6, 14247–14259. doi: 10.18632/oncotarget.3679
Siboni, G., Weitman, H., Freeman, D., Mazur, Y., Malik, Z., and Ehrenberg, B. (2002). The correlation between hydrophilicity of hypericins and helianthrone: internalization mechanisms, subcellular distribution and photodynamic action in colon carcinoma cells. Photochem. Photobiol. Sci. 1, 483–491. doi: 10.1039/b202884k
Sim, H. G., Lau, W. K., Olivo, M., Tan, P. H., and Cheng, C. W. (2005). Is photodynamic diagnosis using hypericin better than white-light cystoscopy for detecting superficial bladder carcinoma? BJU Int. 95, 1215–1218. doi: 10.1111/j.1464-410X.2005.05508.x
Smith, P., Bullock, J. M., Booker, B. M., Haas, C. E., Berenson, C. S., and Jusko, W. J. (2004). The influence of St. John's wort on the pharmacokinetics and protein binding of imatinib mesylate. Pharmacotherapy 24, 1508–1514. doi: 10.1592/phco.24.16.1508.50958
Song, S., Xiong, C., Zhou, M., Lu, W., Huang, Q., Ku, G., et al. (2011). Small-animal PET of tumor damage induced by photothermal ablation with 64Cu-bis-DOTA-hypericin. J. Nucl. Med. 52, 792–799. doi: 10.2967/jnumed.110.086116
Stavrovskaya, A. A. (2000). Cellular mechanisms of multidrug resistance of tumor cells. Biochemistry Mosc. 65, 95–106.
Straub, M., Russ, D., Horn, T., Gschwend, J. E., and Abrahamsberg, C. (2015). A phase IIA dose-finding study of PVP-hypericin fluorescence cystoscopy for detection of nonmuscle-invasive bladder cancer. J. Endourol. 29, 216–222. doi: 10.1089/end.2014.0282
Theodossiou, T. A., Hothersall, J. S., De Witte, P. A., Pantos, A., and Agostinis, P. (2009). The multifaceted photocytotoxic profile of hypericin. Mol. Pharm. 6, 1775–1789. doi: 10.1021/mp900166q
Thomas, C., MacGill, R. S., Miller, G. C., and Pardini, R. S. (1992). Photoactivation of hypericin generates singlet oxygen in mitochondria and inhibits succinoxidase. Photochem. Photobiol. 55, 47–53. doi: 10.1111/j.1751-1097.1992.tb04208.x
Thomas, C., and Pardini, R. S. (1992). Oxygen dependence of hypericin-induced phototoxicity to EMT6 mouse mammary carcinoma cells. Photochem. Photobiol. 55, 831–837. doi: 10.1111/j.1751-1097.1992.tb08531.x
Thong, P. S., Kho, K. W., Zheng, W., Harris, M., Soo, K. C., and Olivo, M. (2007). Development of a laser confocal endomicroscope for in vivo fluorescence imaging. J. Mech. Med. Biol. 7, 11–18. doi: 10.1142/S0219519407002108
Thong, P. S., Olivo, M., Chin, W. W., Bhuvaneswari, R., Mancer, K., and Soo, K. C. (2009). Clinical application of fluorescence endoscopic imaging using hypericin for the diagnosis of human oral cavity lesions. Br. J. Cancer. 101, 1580–1584. doi: 10.1038/sj.bjc.6605357
Thong, P. S., Tandjung, S. S., Movania, M. M., Chiew, W. M., Olivo, M., Bhuvaneswari, R., et al. (2012). Toward real-time virtual biopsy of oral lesions using confocal laser endomicroscopy interfaced with embedded computing. J. Biomed. Opt. 17:056009. doi: 10.1117/1.JBO.17.5.056009
Thong, P. S., Watt, F., Ren, M. Q., Tan, P. H., Soo, K. C., and Olivo, M. (2006). Hypericin-photodynamic therapy (PDT) using an alternative treatment regime suitable for multi-fraction PDT. J. Photochem. Photobiol. B. Biol. 82, 1–8. doi: 10.1016/j.jphotobiol.2005.08.002
Tian, R., Koyabu, N., Morimoto, S., Shoyama, Y., Ohtani, H., and Sawada, Y. (2005). Functional induction and de-induction of P-glycoprotein by St. John's wort and its ingredients in a human colon adenocarcinoma cell line. Drug Metab. Dispos. 33, 547–554. doi: 10.1124/dmd.104.002485
Urbanová, M., Košuth, J., and Čellárová, E. (2006). Genetic and biochemical analysis of Hypericum perforatum L. plants regenerated after cryopreservation. Plant Cell Rep. 25, 140–147. doi: 10.1007/s00299-005-0050-0
Urla, C., Armeanu-Ebinger, S., Fuchs, J., and Seitz, G. (2015). Successful in vivo tumor visualization using fluorescence laparoscopy in a mouse model of disseminated alveolar rhabdomyosarcoma. Surg. Endosc. 29, 1105–1114. doi: 10.1007/s00464-014-3770-9
Vandenbogaerde, A. L., Cuveele, J. F., Proot, P., Himpens, B. E., Merlevede, W. J., and de Witte, P. A. (1997). Differential cytotoxic effects induced after photosensitization by hypericin. J. Photochem. Photobiol. B. Biol. 38, 136–142. doi: 10.1016/S1011-1344(96)07446-5
Vandenbogaerde, A. L., Geboes, K. R., Cuveele, J. F., Agostinis, P. M., Merlevede, W. J., and De Witte, P. A. (1996). Antitumour activity of photosensitized hypericin on A431 cell xenografts. Anticancer Res. 16, 1619–1625.
Vandepitte, J., Van Cleynenbreugel, B., Hettinger, K., Van Poppel, H., and de Witte, P. A. (2010). Biodistribution of PVP-hypericin and hexaminolevulinate-induced PpIX in normal and orthotopic tumor-bearing rat urinary bladder. Cancer Chemother. Pharmacol. 67, 775–781. doi: 10.1007/s00280-010-1375-0
Van de Putte, M., Marysael, T., Fonge, H., Roskams, T., Cona, M. M., Li, J., et al. (2012). Radiolabeled iodohypericin as tumor necrosis avid tracer: diagnostic and therapeutic potential. Int. J. Cancer. 131, E129–E137. doi: 10.1002/ijc.26492
Van de Putte, M., Ni, Y., and De Witte, P. A. (2008a). Exploration of the mechanism underlying the tumor necrosis avidity of hypericin. Oncol. Rep. 19, 921–926. doi: 10.3892/or.19.4.921
Van de Putte, M., Wang, H., Chen, F., de Witte, P. A., and Ni, Y. (2008b). Hypericin as a marker for determination of tissue viability after intratumoral ethanol injection in a murine liver tumor model. Acad. Radiol. 15, 107–113. doi: 10.1016/j.acra.2007.08.008
Van de Putte, M., Wang, H., Chen, F., De Witte, P. A., and Ni, Y. (2008c). Hypericin as a marker for determination of tissue viability after radiofrequency ablation in a murine liver tumor model. Oncol. Rep. 19, 927–932. doi: 10.3892/or.19.4.927
Wada, A., Sakaeda, T., Takara, K., Hirai, M., Kimura, T., Ohmoto, N., et al. (2002). Effects of St John's wort and hypericin on cytotoxicity of anticancer drugs. Drug Metab. Pharmacokinet. 17, 467–474. doi: 10.2133/dmpk.17.467
Wölfle, U., Seelinger, G., and Schempp, C. M. (2014). Topical application of St. John's wort (Hypericum perforatum). Planta Med. 80, 109–120. doi: 10.1055/s-0033-1351019
Xie, X., Hudson, J. B., and Guns, E. S. (2001). Tumor-specific and photodependent cytotoxicity of hypericin in the human LNCaP prostate tumor model. Photochem. Photobiol. 74, 221–225. doi: 10.1562/0031-8655(2001)0740221TSAPCO2.0.CO2
Zahreddine, H., and Borden, K. L. (2013). Mechanisms and insights into drug resistance in cancer. Front. Pharmacol. 4:28. doi: 10.3389/fphar.2013.00028
Zheng, Y., Yin, G., Le, V., Zhang, A., Chen, S., Liang, X., et al. (2016). Photodynamic-therapy activates immune response by disrupting immunity homeostasis of tumor cells, which generates vaccine for cancer therapy. Int. J. Biol. Sci. 12, 120–132. doi: 10.7150/ijbs.12852
Keywords: hypericin, St. John's wort, anticancer activities, photodynamic therapy, photodynamic diagnosis, drug resistance
Citation: Jendželovská Z, Jendželovský R, Kuchárová B and Fedoročko P (2016) Hypericin in the Light and in the Dark: Two Sides of the Same Coin. Front. Plant Sci. 7:560. doi: 10.3389/fpls.2016.00560
Received: 12 February 2016; Accepted: 11 April 2016;
Published: 06 May 2016.
Edited by:
Gregory Franklin, Polish Academy of Sciences, PolandReviewed by:
Jiřina Hofmanová, Academy of Sciences of the Czech Republic, Czech RepublicCopyright © 2016 Jendželovská, Jendželovský, Kuchárová and Fedoročko. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Peter Fedoročko, cGV0ZXIuZmVkb3JvY2tvQHVwanMuc2s=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.