 Mansoor Karimi Jashni1,2Rahim Mehrabi1,3Jérôme Collemare1,4Carl H. Mesarich1,5Pierre J. G. M. de Wit1*
Mansoor Karimi Jashni1,2Rahim Mehrabi1,3Jérôme Collemare1,4Carl H. Mesarich1,5Pierre J. G. M. de Wit1*- 1Laboratory of Phytopathology, Wageningen University and Research Centre, Wageningen, Netherlands
- 2Department of Plant Pathology, Tarbiat Modares University, Tehran, Iran
- 3Cereal Research Department, Seed and Plant Improvement Institute, Karaj, Iran
- 4UMR1345, IRHS-INRA, Beaucouzé, France
- 5Bioprotection Technologies, The New Zealand Institute for Plant and Food Research Limited, Mount Albert Research Centre, Auckland, New Zealand
Upon host penetration, fungal pathogens secrete a plethora of effectors to promote disease, including proteases that degrade plant antimicrobial proteins, and protease inhibitors (PIs) that inhibit plant proteases with antimicrobial activity. Conversely, plants secrete proteases and PIs to protect themselves against pathogens or to mediate recognition of pathogen proteases and PIs, which leads to induction of defense responses. Many examples of proteases and PIs mediating effector-triggered immunity in host plants have been reported in the literature, but little is known about their role in compromising basal defense responses induced by microbe-associated molecular patterns. Recently, several reports appeared in literature on secreted fungal proteases that modify or degrade pathogenesis-related proteins, including plant chitinases or PIs that compromise their activities. This prompted us to review the recent advances on proteases and PIs involved in fungal virulence and plant defense. Proteases and PIs from plants and their fungal pathogens play an important role in the arms race between plants and pathogens, which has resulted in co-evolutionary diversification and adaptation shaping pathogen lifestyles.
Introduction
For successful infection of host plants and establishment of disease, fungal pathogens need weaponry to facilitate penetration, host colonization and uptake of nutrients for growth and reproduction, and at the same time to protect themselves against host defense responses. On the other hand, plants have developed surveillance systems to recognize and defend themselves against invading pathogens. Plant immune receptors recognize conserved microbe-associated molecular patterns (MAMPs) like chitin oligomers released from fungal cell walls during infection. This recognition leads to MAMP-triggered immunity (MTI) and initiates basal defense responses including the activation of structural and (bio)chemical barriers (Jones and Dangl, 2006; Spoel and Dong, 2012). However, adapted plant pathogens have gained the ability to overcome MTI by producing effector molecules that suppress or compromise MTI responses, thereby facilitating effector-triggered susceptibility (ETS; Stergiopoulos and de Wit, 2009). In response, plants have developed an additional layer of defense that enables them to recognize pathogen effectors or effector-modified host targets leading to effector-triggered immunity (ETI; Jones and Dangl, 2006).
Proteases and protease inhibitors (PIs) secreted by pathogens or their host plants have been extensively studied and have been demonstrated to play an important role in ETS and ETI (van der Hoorn, 2008). However, little is known about their role in MTI and related plant basal defense responses. Plant basal defense responses include the induction of pathogenesis-related proteins (PRs) such as antimicrobial chitinases, β-1,3-glucanases and proteases that hydrolyse the fungal cell wall components chitin, glucans, and polypeptides, respectively. The induction of these PR proteins upon plant infection, their antifungal activity, as well as their exploitation in engineering resistance in transgenic plants are very well documented (Wubben et al., 1996; Sels et al., 2008; Balasubramanian et al., 2012; Cletus et al., 2013). An early report in the literature suggested that pathogens might overcome the deleterious effects of plant chitinases by secreting proteases that modified them (Lange et al., 1996; Sela-Buurlage, 1996). This was further supported by recent studies, which indicate that chitinases are targeted by pathogen proteases and protected by PIs (Naumann et al., 2011; Slavokhotova et al., 2014). This encouraged us to review the recent advances on proteases and PIs that play a role in the arms race between plants and their fungal and oomycete pathogens.
Plant Proteases and Protease Inhibitors Involved in Basal Defense
Most PR proteins exhibit direct antimicrobial activities, such as chitinases that degrade chitin present in fungal cell walls. PR proteins play a role in both constitutive and induced basal defense responses (Avrova et al., 2004; Shabab et al., 2008; van Esse et al., 2008). For example, tomato and potato contain basal levels of proteases in their apoplast, including serine proteases like P69, and papain-like cysteine proteases (PLCPs) like Rcr3, which are required for resistance of tomato against Cladosporium fulvum (Song et al., 2009), as well as Pip1 (Phytophthora inhibited protease 1; Tian et al., 2007; Shabab et al., 2008) and C14, which play a role in the resistance of potato against Phytophthora infestans (Kaschani et al., 2010; Bozkurt et al., 2011). After being challenged by pathogens, proteases are induced both locally (Tian et al., 2005) and systemically in the apoplast (Tian et al., 2007; Shabab et al., 2008; Song et al., 2009), which suggests that their activity affects pathogen growth directly or indirectly. Deletion or silencing of genes encoding these proteases enhanced the susceptibility of plants to pathogens, supporting their role in defense responses. Deletion of Rcr3 increased the susceptibility of tomato to the late blight pathogen P. infestans (Song et al., 2009), to the leaf mold pathogen C. fulvum (Dixon et al., 2000), and also to the potato cyst nematode Globodera rostochiensis (Lozano-Torres et al., 2012). Likewise, silencing of C14 in Nicotiana benthamiana significantly increased their susceptibility to P. infestans (Kaschani et al., 2010). These findings suggest that proteases have a determinative role in the execution of defense against plant pathogens.
Plant PIs have also been reported to play a role in plant immunity, through the inhibition of pathogen proteases, or the regulation of endogenous plant proteases (Ryan, 1990; Mosolov et al., 2001; Valueva and Mosolov, 2004; Kim et al., 2009). This has been shown for PIs from barley (Hordeum vulgare) against proteases from Fusarium culmorum (Pekkarinen et al., 2007), as well as for PIs from broad bean (Vicia faba), which inhibited the mycelial growth of several pathogens (Ye et al., 2001). The A. thaliana unusual serine protease inhibitor (UPI) was shown to play a role in defense against the necrotrophic fungi Botrytis cinerea and Alternaria brassicicola (Laluk and Mengiste, 2011). The UPI protein strongly inhibited the serine protease chymotrypsin but also affected the cysteine protease papain (Laluk and Mengiste, 2011). Plants harboring a loss-of-function UPI allele displayed enhanced susceptibility to B. cinerea and A. brassicicola, but not to the bacterium Pseudomonas syringae. Also, hevein-like antimicrobial peptides from wheat (WAMPs) were shown to inhibit class IV chitinase degradation by fungalysin, a metalloprotease secreted by Fusarium verticillioides (Slavokhotova et al., 2014). WAMPs bind to fungalysin, but are not cleaved by the enzyme due to the presence of a Ser residue between the Gly and Cys residues where cleavage of class IV chitinase by fungalysin normally takes place (Naumann et al., 2011; Slavokhotova et al., 2014). Adding equal molar quantities of WAMP and chitinase to fungalysin was sufficient to completely inhibit fungalysin activity suggesting a higher affinity of the protease to the WAMP than to the chitinase.
Interestingly, some pathogens can also manipulate the transcription of plant PIs to inhibit deleterious effects of plant proteases in their favor. For example, production of maize cysteine proteases is induced during infection by Ustilago maydis, but at the same time the fungus induces the production of maize cystatin CC9 that inhibits cysteine proteases to facilitate infection (van der Linde et al., 2012b; Mueller et al., 2013). This suggests an evolutionary arms race in which the infection strategy of the pathogen benefits from the host’s antimicrobial defense to suppress its defense responses.
Fungal Proteases Targeting Host Defense Proteins
The arms race between pathogens and their hosts is often explained by recognition of MAMPs or effectors through pattern recognition receptors or resistance proteins, which results in MTI or ETI (Jones and Dangl, 2006). However, several components of basal defense are both constitutive and induced after interaction between MAMPs/effectors and immune receptors. PR proteins provide an excellent example of this. PR proteins are generally stable proteins that often exhibit a basal level of expression, but are also strongly induced after infection (Sels et al., 2008). PR proteins and their antifungal activity have been exploited to improve broad-spectrum resistance in plants. Plants such as tobacco, tomato, potato, peanut, and cacao have been engineered to over-express chitinases alone (Schickler and Chet, 1997; de las Mercedes Dana et al., 2006; Maximova et al., 2006; Iqbal et al., 2012; Cletus et al., 2013) or in combination with other PR proteins in pea and rice (Sridevi et al., 2008; Amian et al., 2011), and showed enhanced resistance to fungal pathogens.
Plant chitinases and especially chitin-binding domain (CBD)-containing chitinases play an important role in defense against pathogenic fungi (Iseli et al., 1993; Suarez et al., 2001). Some fungal pathogens such as C. fulvum secrete chitin-binding effector proteins like CfAvr4 into the colonized extracellular space of tomato leaves to protect themselves against the antifungal activity of apoplastic plant chitinases (van den Burg et al., 2006). Indeed, CfAvr4 binds to chitin of fungal cell walls, making chitin inaccessible to plant chitinases, thereby preventing hydrolysis by these enzymes (van den Burg et al., 2006). Functional homologs of CfAvr4 have been identified in other Dothideomycete plant pathogens, in which they likely also protect the fungal cell wall against plant chitinases (Stergiopoulos et al., 2010; de Wit et al., 2012; Mesarich et al., 2015). However, many fungal pathogens do not carry homologs of the CfAvr4 gene in their genome. It appears that some fungi secrete proteases that cleave CBD-chitinases. For example, F. solani f. sp. phaseoli is able to modify chitinases during infection of bean to facilitate host colonization (Lange et al., 1996). Also an extracellular subtilisin protease from F. solani f. sp. eumartii was reported to modify chitinases and β-1,3-glucanases present in intercellular washing fluids of potato (Olivieri et al., 2002). More recently, it was shown that F. verticillioides and other maize pathogens, including Bipolaris zeicola and Stenocarpella maydis, secrete two types of proteases that truncate maize class IV CBD-chitinases (Naumann, 2011). A fungalysin metalloprotease of F. verticillioides was found to cleave within the CBD domain between conserved Gly and Cys residues (Naumann et al., 2011), while a novel polyglycine hydrolase present in many fungi belonging to the family of Pleosporineae cleaved within the polyglycine linker present in the hinge domain of class IV chitinases (Naumann et al., 2014, 2015). In another recent study it was shown that the fungal tomato pathogens B. cinerea, V. dahliae, and F. oxysporum f. sp. lycopersici secrete proteases that modify tomato CBD-chitinases (Karimi Jashni et al., 2015). For F. oxysporum f. sp. lycopersici, the synergistic action of a serine protease, FoSep1, and a metalloprotease, FoMep1 (the ortholog of fungalysin from F. verticillioides), was required for cleavage and removal of the CBD from two tomato CBD-chitinases (Karimi Jashni et al., 2015). Removal of the CBD from two tomato CBD-chitinases by these two enzymes led to a reduction of their chitinase and antifungal activity. In addition, mutants of F. oxysporum f. sp. lycopersici lacking both FoSep1 and FoMep1 exhibited reduced virulence on tomato, confirming that secreted fungal proteases are important virulence factors by targeting CDB-chitinases to compromise an important component of plant basal defense (Karimi Jashni et al., 2015).
Collectively, the activity of fungal proteases might explain why overexpression of plant chitinases in transgenic plants has not become an effective strategy to obtain durable resistance against fungal pathogens. Secretion of proteases and PIs by pathogens to modify, degrade, or inhibit basal defense proteins might have played an important role during co-evolution with their host plants (Hörger and van der Hoorn, 2013). Therefore, overexpression of chitinases from a heterologous source in transgenic plants might be a more efficient approach to obtain durable resistance against pathogens, as they have not co-evolved with these “foreign” defense proteins.
Fungal Protease Inhibitors Targeting Host Proteases
Plant pathogens also secrete PI effectors to inhibit plant defense proteases and promote disease development. These effectors are targeted to various host compartments (Tian et al., 2009). One such effector, Avr2, secreted by C. fulvum during infection, is required for full virulence of this fungus on tomato (Rooney et al., 2005). Avr2 inhibits the tomato apoplastic PLCPs Rcr3 and Pip1 to support growth of C. fulvum in the apoplast. Also, plants expressing Avr2 showed increased susceptibility to other pathogenic fungi, including B. cinerea and V. dahliae (van Esse et al., 2008). Moreover, A. thaliana plants expressing Avr2 triggered global transcriptional reprogramming, reflecting a typical host response to pathogen attack (van Esse et al., 2008). Two other PI effectors are the cystatin-like proteins EPIC1 (extracellular proteinase inhibitor C1) and EPIC2B (extracellular proteinase inhibitor C2B), whose expression is strongly induced in the oomycete P. infestans during biotrophic growth on tomato leaves (Tian et al., 2007; Song et al., 2009). These PIs selectively target the plant PLCPs Rcr3, Pip1, and C14 in the apoplast of potato and tomato. The EPICs inhibit C14 and possibly other PLCPs over a wider pH range than that observed for Avr2, which only inhibits Pip1 and Rcr3 at pH values occurring in the apoplast where the pathogen grows. In addition, P. infestans secretes two serine PIs (EPI1 and EPI10) that target and inhibit the major apoplastic serine protease P69B, likely to decrease its role in defense (Tian et al., 2004, 2005). It was proposed that EPI1 protects EPIC1 and EPIC2B proteins from degradation by P69B (Tian, 2005). Furthermore, the maize pathogen U. maydis secretes the cysteine PI Pit2 that strongly inhibits three abundant defense-related maize cysteine proteases (CP2 and its two isoforms CP1A and CP1B; Van der Linde et al., 2012a; van der Linde et al., 2012b; Mueller et al., 2013). These findings indicate that cysteine and serine PIs secreted by different groups of filamentous fungal and oomycete pathogens, as well as their activity against plant proteases, can compromise plant basal defense responses. A schematic overview of different types of interactions between pathogen and host proteases and PIs at the plant–pathogen interface is presented in Figure 1.
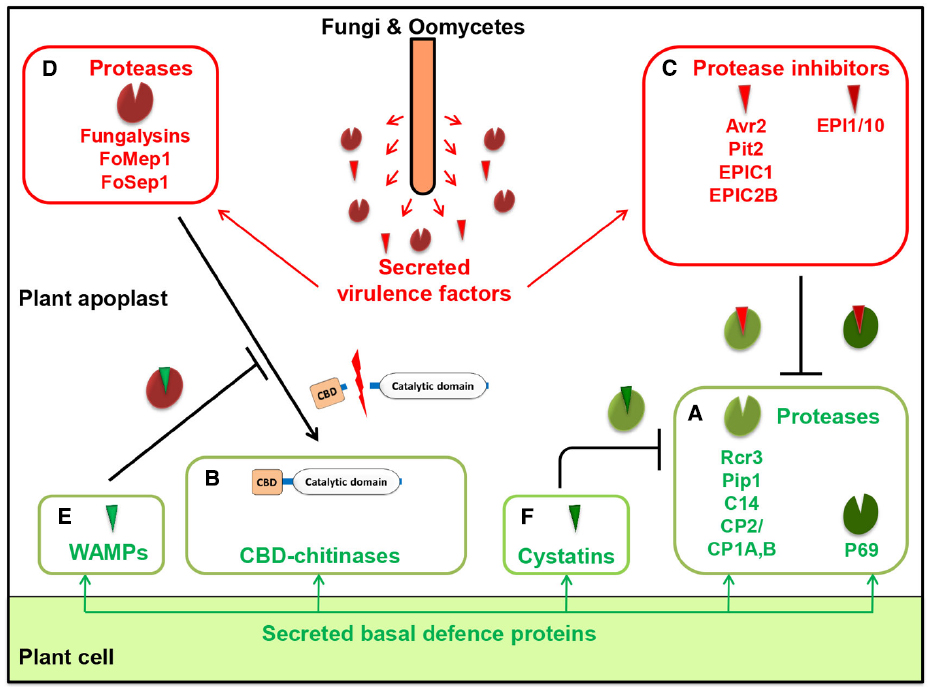
Figure 1. Proteases and protease inhibitors at the plant–pathogen interface. As part of their basal defense response, plants secrete deleterious enzymes such as proteases (A) and chitin-binding domain (CBD)-containing chitinases (B) that target pathogen components. In response, filamentous pathogens secrete protease inhibitors (C) that inhibit plant cysteine or serine proteases. Filamentous pathogens also secrete fungalysin metallo- or serine proteases (D) that process antifungal CBD-chitinases of plants. In response, plants secrete antimicrobial peptides such as hevein-like antimicrobial peptides from wheat (WAMPs) (E) that inhibit fungalysin metalloproteases or cystatins (F) that inhibit endogenous plant cysteine proteases. Examples shown in this figure are discussed in the text.
Proteases, PI Effectors, and Their Role in Receptor-Mediated Host Defense Responses
The plant immune system is able to recognize pathogen effectors to mount receptor-mediated defense responses. Although the intrinsic function of protease and PI effectors secreted by some pathogenic fungi promote disease through manipulation of host defense, proteases and PI effectors can also be recognized by host immune receptors mediating defense responses. This adaptation and counter-adaptation reflects the arms race between pathogens and their host plants. A clear example of such an evolutionary arms race are the cysteine PIs Avr2 from C. fulvum and Gr-VAP1 (Globodera rostochiensis Venom Allergen-like Protein) from G. rostochiensis that bind and inhibit the tomato cysteine protease Rcr3pim. The tomato immune receptor protein Cf-2 senses this interaction and mediates the induction of defense responses (Song et al., 2009; Lozano-Torres et al., 2012). Most likely, the interaction causes a conformational change in Rcr3, which is recognized by the Cf-2 receptor (Krüger et al., 2002; Rooney et al., 2005). This hypothesis is supported by the finding that a natural variant of Rcr3 is recognized by Cf-2 in an Avr2-independent manner (Dixon et al., 2000). Moreover, in tomato plants lacking the Cf-2 receptor, targeting of Rcr3 is not sensed and plants are more susceptible to G. rostochiensis (Lozano-Torres et al., 2012).
Co-evolution Between Plants and Their Pathogens is Reflected by the Numerous Variant Proteases and PIs in the Genomes of Both Organisms
The genomes of fungal plant pathogens encode predicted proteases belonging to various subfamilies that vary in number between pathogens with different lifestyles. Generally, hemi-biotrophs and saprotrophs contain higher numbers of secreted proteases than biotrophs (Ohm et al., 2012). However, these predictions are based on gene numbers and may not be supported by their transcription and translation profiles. For example, C. fulvum, which is a biotrophic fungus, has numbers of proteases that are comparable to the phylogenetically closely related hemi-biotroph Dothistroma septosporum (de Wit et al., 2012). However, likely due to its adaptation to a different host and lifestyle, many C. fulvum protease genes are not expressed in planta and some have undergone pseudogenization (van der Burgt et al., 2014). Deletion and duplication of protease genes were reported to occur in the genome of the grass endophytic fungus Epichloë festucae (Bryant et al., 2009) but their biological implications have not yet been studied.
Adaptation of PI effectors from pathogens to inhibit different host proteases has been observed in several cases. The Avr2 PI of C. fulvum, for example, has a high affinity for the host proteases Rcr3 and Pip1 and a low affinity for C14 (Shabab et al., 2008; Hörger et al., 2012). P. infestans EPICs have a high affinity for C14 and a low affinity for Rcr3 and Pip1 (Kaschani et al., 2010). Furthermore, U. maydis Pit2 inhibits the maize cysteine proteases CP1, CP2, and XCP2, but does not inhibit cathepsin CatB (Mueller et al., 2013). Different types of selection pressure may lead to the circumvention of protease inhibition by PIs. For example, purifying or diversifying selection has been reported for the proteases Rcr3, C14, and Pip1, and has been shown to act at their PI binding sites. Sequencing of the tomato proteases Rcr3 and Pip1 across different wild tomato species has shown that these proteins are under strong diversifying selection imposed by Avr2. For instance, one of the variant residues in the binding site of Rcr3 prevented inhibition by Avr2, indicating selection for evasion from recognition by this inhibitor (Shabab et al., 2008). C14 from solanaceous plants is also the target of EPICs secreted by P. infestans and is under diversifying selection in potato and under conservative selection in tomato. This demonstrates that C14 plays an active role in host immunity against this pathogen and variations in the sequence of C14 in natural hosts of P. infestans highlight the co-evolutionary arms race at the plant–pathogen interface (Kaschani et al., 2010).
Evolutionary diversification may vary from point mutation to gene deletion or insertion. EPIC1 and EPIC2 are PIs present in P. infestans, however their orthologs were lost in P. sojae and P. ramorum (Tian et al., 2007). P. mirabilis, a species closely related to P. infestans, is a pathogen of Mirabilis jalapa, and secretes the PI PmEPIC1, an ortholog of EPIC1 that inhibits C14 but not Rcr3 (Dong et al., 2014). However, M. jalapa secretes MRP2, a PLCP homolog of Rcr3, that is more effectively inhibited by PmEPIC1 than by EPIC1 (Dong et al., 2014). Substitution of one amino acid residue in PmEPIC1 and EPIC1 restored the inhibitory function of PmEPIC1 on Rcr3 and of EPIC1 on MRP2, respectively. These results show that proteases and PIs have played important roles in adaptation of the two Phytophthora species to their respective host plants, although the two species diverged only a 1000 years ago (Dong et al., 2014). This is an excellent example for a role of a protease and PI in the arms race between a plant and its pathogen and exemplifies how diversification and adaptation of a protease-PI complex may work at the molecular level.
Conclusion and Perspective
The recent advances reviewed here exemplify determinative roles of proteases and PIs in shaping plant–pathogen interactions. Analyses of genome databases of both plants and pathogens show that these organisms encode numerous proteases and PIs, of which we are just beginning to understand some of their roles. Advanced transcriptome and proteome tools such as RNA sequencing and protease profiling will facilitate identification of important proteases and PIs for further functional analysis. The redundancy of proteases in pathogens is a technical challenge that has so far hampered defining their biological functions. Targeted deletion of one or even two protease genes failed to change virulence of the plant pathogenic fungi Glomerella cingulata (Plummer et al., 2004) and B. cinerea (ten Have et al., 2010), respectively. Karimi Jashni et al. (2015) could only show decreased virulence of a double protease mutant of the tomato pathogen F. oxysporum by a combined biochemical and genetic approach, and using a defined plant enzyme (CBD-chitinase) as a substrate that was presumed to be involved in plant defense. This indicates that multi-gene targeting of protease and PI genes to identify their role in virulence or avirulence remains a challenge in filamentous fungi. Targeting multiple protease and PI genes might also be hampered by lack of sufficient numbers of selection markers for targeted gene replacement. In the latter case multiple protease and PI genes might be targeted by targeted gene silencing.
Author Contributions
MJ and PdW conceived and wrote the review; JC, RM, and CM critically reviewed the manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
MJ was financed by the Ministry of Science, Research and Technology (MSRT) of Iran; PdW, JC, RM, and CM were financed by Wageningen University and the Royal Netherlands Academy of Arts and Sciences.
References
Amian, A. A., Papenbrock, J., Jacobsen, H. J., and Hassan, F. (2011). Enhancing transgenic pea (Pisum sativum L.) resistance against fungal diseases through stacking of two antifungal genes (chitinase and glucanase). GM Crops 2, 104–109. doi: 10.4161/gmcr.2.2.16125
Avrova, A. O., Taleb, N., Rokka, V. M., Heilbronn, J., Campbell, E., Hein, I., et al. (2004). Potato oxysterol binding protein and cathepsin B are rapidly up-regulated in independent defence pathways that distinguish R gene-mediated and field resistances to Phytophthora infestans. Mol. Plant Pathol. 5, 45–56. doi: 10.1111/j.1364-3703.2004.00205.x
Balasubramanian, V., Vashisht, D., Cletus, J., and Sakthivel, N. (2012). Plant β-1,3-glucanases: their biological functions and transgenic expression against phytopathogenic fungi. Biotechnol. Lett. 34, 1983–1990. doi: 10.1007/s10529-012-1012-6
Bozkurt, T. O., Schornack, S., Win, J., Shindo, T., Ilyas, M., Oliva, R., et al. (2011). Phytophthora infestans effector Avrblb2 prevents secretion of a plant immune protease at the haustorial interface. Proc. Natl. Acad. Sci. U.S.A. 108, 20832–20837. doi: 10.1073/pnas.1112708109
Bryant, M. K., Schardl, C. L., Hesse, U., and Scott, B. (2009). Evolution of a subtilisin-like protease gene family in the grass endophytic fungus Epichloë festucae. BMC Evol. Biol. 9:168. doi: 10.1186/1471-2148-9-168
Cletus, J., Balasubramanian, V., Vashisht, D., and Sakthivel, N. (2013). Transgenic expression of plant chitinases to enhance disease resistance. Biotechnol. Lett. 35, 1719–1732. doi: 10.1007/s10529-013-1269-4
de las Mercedes Dana, M., Pintor-Toro, J. A., and Cubero, B. (2006). Transgenic tobacco plants overexpressing chitinases of fungal origin show enhanced resistance to biotic and abiotic stress agents. Plant Physiol. 142, 722–730. doi: 10.1104/pp.106.086140
de Wit, P. J. G. M, van der Burgt, A., Ökmen, B., Stergiopoulos, I., Abd-Elsalam, K. A., Aerts, A. L., et al. (2012). The genomes of the fungal plant pathogens Cladosporium fulvum and Dothistroma septosporum reveal adaptation to different hosts and lifestyles but also signatures of common ancestry. PLoS Genet. 8:e1003088. doi: 10.1371/journal.pgen.1003088
Dixon, M. S., Golstein, C., Thomas, C. M., van der Biezen, E. A., and Jones, J. D. G. (2000). Genetic complexity of pathogen perception by plants: the example of Rcr3, a tomato gene required specifically by Cf-2. Proc. Natl. Acad. Sci. U.S.A. 97, 8807–8814. doi: 10.1073/pnas.97.16.8807
Dong, S., Stam, R., Cano, L. M., Song, J., Sklenar, J., Yoshida, K., et al. (2014). Effector specialization in a lineage of the Irish potato famine pathogen. Science 343, 552–555. doi: 10.1126/science.1246300
Hörger, A. C., Ilyas, M., Stephan, W., Tellier, A., van der Hoorn, R. A. L., and Rose, L. E. (2012). Balancing selection at the tomato Rcr3 guardee gene family maintains variation in strength of pathogen defense. PLoS Genet. 8:e1002813. doi: 10.1371/journal.pgen.1002813
Hörger, A. C., and van der Hoorn, R. A. L. (2013). The structural basis of specific protease–inhibitor interactions at the plant–pathogen interface. Curr. Opin. Struct. Biol. 23, 842–850. doi: 10.1016/j.sbi.2013.07.013
Iqbal, M., Nazir, F., Ali, S., Asif, M. A., Zafar, Y., Iqbal, J., et al. (2012). Overexpression of rice chitinase gene in transgenic peanut (Arachis hypogaea L.) improves resistance against leaf spot. Mol. Biotechnol. 50, 129–136. doi: 10.1007/s12033-011-9426-2
Iseli, B., Boller, T., and Neuhaus, J. M. (1993). The N-terminal cysteine-rich domain of tobacco class I chitinase is essential for chitin binding but not for catalytic or antifungal activity. Plant Physiol. 103, 221–226. doi: 10.1104/pp.103.1.221
Jones, J. D. G., and Dangl, J. L. (2006). The plant immune system. Nature 444, 323–329. doi: 10.1038/nature05286
Karimi Jashni, M., Dols, H. M. I., Iida, Y., Boeren, S., Beenen, H. G., Mehrabi, R., et al. (2015). Synergistic action of a metalloprotease and a serine protease from Fusarium oxysporum f. sp. lycopersici cleaves chitin-binding tomato chitinases, reduces their antifungal activity and enhances fungal virulence. Mol. Plant Microbe Interact. doi: 10.1094/MPMI-04-15-0074-R [Epub ahead of print].
Kaschani, F., Shabab, M., Bozkurt, T., Shindo, T., Schornack, S., Gu, C., et al. (2010). An effector-targeted protease contributes to defense against Phytophthora infestans and is under diversifying selection in natural hosts. Plant Physiol. 154, 1794–1804. doi: 10.1104/pp.110.158030
Kim, J. Y., Park, S. C., Hwang, I., Cheong, H., Nah, J. W., Hahm, K. S., et al. (2009). Protease inhibitors from plants with antimicrobial activity. Int. J. Mol. Sci. 10, 2860–2872. doi: 10.3390/ijms10062860
Krüger, J., Thomas, C. M., Golstein, C., Dixon, M. S., Smoker, M., Tang, S., et al. (2002). A tomato cysteine protease required for Cf-2-dependent disease resistance and suppression of autonecrosis. Science 296, 744–747. doi: 10.1126/science.1069288
Laluk, K., and Mengiste, T. (2011). The Arabidopsis extracellular unusual serine protease inhibitor functions in resistance to necrotrophic fungi and insect herbivory. Plant J. 68, 480–494. doi: 10.1111/j.1365-313X.2011.04702.x
Lange, J., Mohr, U., Wiemken, A., Boller, T., and Vögeli-Lange, R. (1996). Proteolytic processing of class IV chitinase in the compatible interaction of bean roots with Fusarium solani. Plant Physiol. 111, 1135–1144. doi: 10.1104/pp.111.4.1135
Lozano-Torres, J. L., Wilbers, R. H. P., Gawronski, P., Boshoven, J. C., Finkers-Tomczak, A., Cordewener, J. H. G., et al. (2012). Dual disease resistance mediated by the immune receptor Cf-2 in tomato requires a common virulence target of a fungus and a nematode. Proc. Natl. Acad. Sci. U.S.A. 109, 10119–10124. doi: 10.1073/pnas.1202867109
Maximova, S., Marelli, J. P., Young, A., Pishak, S., Verica, J., and Guiltinan, M. (2006). Over-expression of a cacao class I chitinase gene in Theobroma cacao L. Enhances resistance against the pathogen, Colletotrichum gloeosporioides. Planta 224, 740–749. doi: 10.1007/s00425-005-0188-6
Mesarich, C. H., Stergiopoulos, I., Beenen, H. G., Cordovez, V., Guo, Y., Karimi Jashni, M., et al. (2015). A conserved proline residue in Dothideomycete Avr4 effector proteins is required to trigger a Cf-4-dependent hypersensitive response Mol. Plant Pathol. doi: 10.1111/mpp.12265 [Epub ahead of print].
Mosolov, V. V., Grigor’eva, L. I., and Valueva, T. A. (2001). Involvement of proteolytic enzymes and their inhibitors in plant protection (review). Appl. Biochem. Microbiol. 37, 115–123. doi: 10.1023/A:1002824527463
Mueller, A. N., Ziemann, S., Treitschke, S., Aßmann, D., and Doehlemann, G. (2013). Compatibility in the Ustilago maydis–maize interaction requires inhibition of host cysteine proteases by the fungal effector Pit2. PLoS Pathog. 9:e1003177. doi: 10.1371/journal.ppat.1003177
Naumann, T. A. (2011). Modification of recombinant maize ChitA chitinase by fungal chitinase-modifying proteins. Mol. Plant Pathol. 12, 365–372. doi: 10.1111/j.1364-3703.2010.00677.x
Naumann, T. A., Naldrett, M. J., Ward, T. J., and Price, N. P. J. (2015). Polyglycine hydrolases: fungal β-lactamase-like endoproteases that cleave polyglycine regions within plant class IV chitinases. Protein Sci. 24, 1147–1157. doi: 10.1002/pro.2705
Naumann, T. A., Wicklow, D. T., and Price, N. P. J. (2011). Identification of a chitinase-modifying protein from Fusarium verticillioides. J. Biol. Chem. 286, 35358–35366. doi: 10.1074/jbc.M111.279646
Naumann, T. A., Wicklow, D. T., and Price, N. P. J. (2014). Polyglycine hydrolases secreted by Pleosporineae fungi that target the linker region of plant class IV chitinases. Biochem. J. 460, 187–198. doi: 10.1042/BJ20140268
Ohm, R. A., Feau, N., Henrissat, B., Schoch, C. L., Horwitz, B. A., Barry, K. W., et al. (2012). Diverse lifestyles and strategies of plant pathogenesis encoded in the genomes of eighteen Dothideomycetes fungi. PLoS Pathog. 8:e1003037. doi: 10.1371/journal.ppat.1003037
Olivieri, F., Eugenia Zanetti, M., Oliva, C. R., Covarrubias, A. A., and Casalongué, C. A. (2002). Characterization of an extracellular serine protease of Fusarium eumartii and its action on pathogenesis related proteins. Eur. J. Plant Pathol. 108, 63–72. doi: 10.1023/A:1013920929965
Pekkarinen, A. I., Longstaff, C., and Jones, B. L. (2007). Kinetics of the inhibition of Fusarium serine proteinases by barley (Hordeum vulgare L.) inhibitors. J. Agric. Food Chem. 55, 2736–2742. doi: 10.1021/jf0631777
Plummer, K. M., Clark, S. J., Ellis, L. M., Loganathan, A., Al-Samarrai, T. H., Rikkerink, E. H. A., et al. (2004). Analysis of a secreted aspartic peptidase disruption mutant of Glomerella cingulata. Eur. J. Plant Pathol. 110, 265–274. doi: 10.1023/b:ejpp.0000019796.78598.8c
Rooney, H. C. E., van’t Klooster, J. W., van der Hoorn, R. A. L., Joosten, M. H. A. J., Jones, J. D. G., and de Wit, P. J. (2005). Cladosporium Avr2 inhibits tomato Rcr3 protease required for Cf-2-dependent disease resistance. Science 308, 1783–1786. doi: 10.1126/science.1111404
Ryan, C. A. (1990). Protease inhibitors in plants: genes for improving defenses against insects and pathogens. Annu. Rev. Phytopathol. 28, 425–449. doi: 10.1146/annurev.py.28.090190.002233
Schickler, H., and Chet, I. (1997). Heterologous chitinase gene expression to improve plant defense against phytopathogenic fungi. J. Ind. Microbiol. Biotechnol. 19, 196–201. doi: 10.1038/sj.jim.2900447
Sela-Buurlage, M. B. (1996). In vitro Sensitivity and Tolerance of Fusarium solani Towards Chitinases and β-1,3-glucanases. Ph.D. thesis, Wageningen University, Wageningen.
Sels, J., Mathys, J., De Coninck, B. M. A., Cammue, B. P. A., and De Bolle, M. F. C. (2008). Plant pathogenesis-related (PR) proteins: a focus on PR peptides. Plant Physiol. Biochem. 46, 941–950. doi: 10.1016/j.plaphy.2008.06.011
Shabab, M., Shindo, T., Gu, C., Kaschani, F., Pansuriya, T., Chintha, R., et al. (2008). Fungal effector protein AVR2 targets diversifying defense-related Cys proteases of tomato. Plant Cell 20, 1169–1183. doi: 10.1105/tpc.107.056325
Slavokhotova, A. A., Naumann, T. A., Price, N. P. J., Rogozhin, E. A., Andreev, Y. A., Vassilevski, A. A., et al. (2014). Novel mode of action of plant defense peptides-hevein-like antimicrobial peptides from wheat inhibit fungal metallo-proteases. FEBS J. 281, 4754–4764. doi: 10.1111/febs.13015
Song, J., Win, J., Tian, M., Schornack, S., Kaschani, F., Ilyas, M., et al. (2009). Apoplastic effectors secreted by two unrelated eukaryotic plant pathogens target the tomato defense protease Rcr3. Proc. Natl. Acad. Sci. U.S.A. 106, 1654–1659. doi: 10.1073/pnas.0809201106
Spoel, S. H., and Dong, X. (2012). How do plants achieve immunity? Defence without specialized immune cells. Nat. Rev. Immunol. 12, 89–100. doi: 10.1038/nri3141
Sridevi, G., Parameswari, C., Sabapathi, N., Raghupathy, V., and Veluthambi, K. (2008). Combined expression of chitinase and β-1, 3-glucanase genes in indica rice (Oryza sativa L.) enhances resistance against Rhizoctonia solani. Plant Sci. 175, 283–290. doi: 10.1016/j.plantsci.2008.04.011
Stergiopoulos, I., and de Wit, P. J. (2009). Fungal effector proteins. Annu. Rev. Phytopathol. 47, 233–263. doi: 10.1146/annurev.phyto.112408.132637
Stergiopoulos, I., van den Burg, H. A., Ökmen, B., Beenen, H. G., van Liere, S., Kema, G. H. J., et al. (2010). Tomato Cf resistance proteins mediate recognition of cognate homologous effectors from fungi pathogenic on dicots and monocots. Proc. Natl. Acad. Sci. U.S.A. 107, 7610–7615. doi: 10.1073/pnas.1002910107
Suarez, V., Staehelin, C., Arango, R., Holtorf, H., Hofsteenge, J., and Meins, F. Jr. (2001). Substrate specificity and antifungal activity of recombinant tobacco class I chitinases. Plant Mol. Biol. 45, 609–618. doi: 10.1023/A:1010619421524
ten Have, A., Espino, J. J., Dekkers, E., van Sluyter, S. C., Brito, N., Kay, J., et al. (2010). The Botrytis cinerea aspartic proteinase family. Fungal. Genet. Biol. 47, 53–65. doi: 10.1016/j.fgb.2009.10.008
Tian, M. (2005). Functional Characterization of Extracellular Protease Inhibitors of Phytophthora infestans. Ph.D. thesis, The Ohio State University, Columbus.
Tian, M., Benedetti, B., and Kamoun, S. (2005). A second Kazal-like protease inhibitor from Phytophthora infestans inhibits and interacts with the apoplastic pathogenesis-related protease P69b of tomato. Plant Physiol. 138, 1785–1793. doi: 10.1104/pp.105.061226
Tian, M., Huitema, E., da Cunha, L., Torto-Alalibo, T., and Kamoun, S. (2004). A Kazal-like extracellular serine protease inhibitor from Phytophthora infestans targets the tomato pathogenesis-related protease P69b. J. Biol. Chem. 279, 26370–26377. doi: 10.1074/jbc.m400941200
Tian, M., Chaudhry, F., Ruzicka, D. R., Meagher, R. B., Staiger, C. J., and Day, B. (2009). Arabidopsis actin-depolymerizing factor AtADF4 mediates defense signal transduction triggered by the Pseudomonas syringae effector AvrPphB. Plant Physiol. 150, 815–824. doi: 10.1104/pp.109.137604
Tian, M., Win, J., Song, J., van der Hoorn, R., van der Knaap, E., and Kamoun, S. (2007). A Phytophthora infestans cystatin-like protein targets a novel tomato papain-like apoplastic protease. Plant Physiol. 143, 364–377. doi: 10.1104/pp.106.090050
Valueva, T., and Mosolov, V. (2004). Role of inhibitors of proteolytic enzymes in plant defense against phytopathogenic microorganisms. Biochemistry 69, 1305–1309. doi: 10.1007/s10541-005-0015-5
van den Burg, H. A., Harrison, S. J., Joosten, M. H. A. J., Vervoort, J., and de Wit, P. J. (2006). Cladosporium fulvum Avr4 protects fungal cell walls against hydrolysis by plant chitinases accumulating during infection. Mol. Plant Microbe Interact. 19, 1420–1430. doi: 10.1094/MPMI-19-1420
van der Burgt, A., Karimi Jashni, M., Bahkali, A. H., and de Wit, P. J. (2014). Pseudogenization in pathogenic fungi with different host plants and lifestyles might reflect their evolutionary past. Mol. Plant Pathol. 15, 133–144. doi: 10.1111/mpp.12072
van der Hoorn, R. A. L. (2008). Plant proteases: from phenotypes to molecular mechanisms. Annu. Rev. Plant Biol. 59, 191–223. doi: 10.1146/annurev.arplant.59.032607.092835
van der Linde, K., Mueller, A. N., Hemetsberger, C., Kashani, F., van der Hoorn, R. A. L., and Doehlemann, G. (2012a). The maize cystatin CC9 interacts with apoplastic cysteine proteases. Plant Signal. Behav. 7, 1397–1401. doi: 10.4161/psb.21902
van der Linde, K., Hemetsberger, C., Kastner, C., Kaschani, F., van der Hoorn, R. A. L., Kumlehn, J., et al. (2012b). A maize cystatin suppresses host immunity by inhibiting apoplastic cysteine proteases. Plant Cell 24, 1285–1300. doi: 10.1105/tpc.111.093732
van Esse, H. P., van’t Klooster, J. W., Bolton, M. D., Yadeta, K. A., van Baarlen, P., Boeren, S., et al. (2008). The Cladosporium fulvum virulence protein Avr2 inhibits host proteases required for basal defense. Plant Cell 20, 1948–1963. doi: 10.1105/tpc.108.059394
Wubben, J. P., Lawrence, C. B., and de Wit, P. J. (1996). Differential induction of chitinase and 1,3-β-glucanase gene expression in tomato by Cladosporium fulvum and its race-specific elicitors. Physiol. Mol. Plant Pathol. 48, 105–116. doi: 10.1006/pmpp.1996.0010
Keywords: cysteine protease, metalloprotease, serine protease, protease inhibitor, chitinase, defence
Citation: Karimi Jashni M, Mehrabi R, Collemare J, Mesarich CH, and de Wit PJGM (2015) The battle in the apoplast: further insights into the roles of proteases and their inhibitors in plant–pathogen interactions. Front. Plant Sci. 6:584. doi: 10.3389/fpls.2015.00584
Received: 07 May 2015; Accepted: 13 July 2015;
Published: 03 August 2015.
Edited by:
Maryam Rafiqi, Computomics, GermanyReviewed by:
Shunyuan Xiao, University of Maryland, USADavid Jones, The Australian National University, Australia
Copyright © 2015 Karimi Jashni, Mehrabi, Collemare, Mesarich and de Wit. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Pierre J. G. M. de Wit, Laboratory of Phytopathology, Wageningen University and Research Centre, Droevendaalsesteeg 9, Wageningen 6708 PB, Netherlands,cGllcnJlLmRld2l0QHd1ci5ubA==