 Iyyakkannu Sivanesan
Iyyakkannu Sivanesan Se Won Park
Se Won Park
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Plant Sci. , 21 October 2014
Sec. Plant Nutrition
Volume 5 - 2014 | https://doi.org/10.3389/fpls.2014.00571
This article is part of the Research Topic MicroRNAs and their role in plant nutrient stress response. View all 6 articles
Growth and morphogenesis of in vitro cultures of plant cells, tissues, and organs are greatly influenced by the composition of the culture medium. Mineral nutrients are necessary for the growth and development of plants. Several morpho-physiological disorders such as hooked leaves, hyperhydricity, fasciation, and shoot tip necrosis are often associated with the concentration of inorganic nutrient in the tissue culture medium. Silicon (Si) is the most abundant mineral element in the soil. The application of Si has been demonstrated to be beneficial for growth, development and yield of various plants and to alleviate various stresses including nutrient imbalance. Addition of Si to the tissue culture medium improves organogenesis, embryogenesis, growth traits, morphological, anatomical, and physiological characteristics of leaves, enhances tolerance to low temperature and salinity, protects cells and against metal toxicity, prevents oxidative phenolic browning and reduces the incidence of hyperhydricity in various plants. Therefore, Si possesses considerable potential for application in a wide range of plant tissue culture studies such as cryopreservation, organogenesis, micropropagation, somatic embryogenesis and secondary metabolites production.
Plant tissue culture is a collection of experimental procedures for aseptic culture of isolated plant cells, tissues or organs on nutrient media under controlled environmental conditions. Growth and morphogenesis of in vitro cultures of plant cells, tissues and organs are greatly influenced by the composition of the culture medium. The composition of a culture medium has often been modified to stimulate the growth of particular plant material. In general, plant tissue culture medium composed of inorganic nutrients, organic supplements, carbon source, plant growth regulators and a solidifying agent. Mineral nutrients are necessary for growth and development of plants. The optimization of inorganic nutrients in the culture medium improves growth and morphogenesis of plant cells, tissues and organs in vitro. Several physiological disorders such as hooked leaves, hyperhydricity, fasciation and shoot tip necrosis are often associated with the concentration of inorganic nutrients in the culture medium (Reed et al., 2013).
Silicon (Si) is the most abundant mineral element in the soil (Epstein, 1999). Numerous studies have shown that Si treatment improves the growth and yield of various plants, particularly when they are subjected to both abiotic and biotic stresses (Ma, 2004). Several researchers have reviewed the role Si on plant tolerance to abiotic (Balakhnina and Borkowska, 2013; Zhu and Gong, 2014) and biotic stresses (Van Bockhaven et al., 2013). The availability of Si in hydroponic and substrate plant production system is restricted. Addition of Si to the nutrient solution or soilless substrate enhanced growth traits, yield and quality of several crops (Voogt and Sonneveld, 2001). The promoting effects of Si on plants might be due to increasing nutrient uptake and photosynthetic activity. Though Si is a ubiquitous contaminant, the use of silicon-free containers and double distilled water restricts its availability. Furthermore, Si has not been included in any commercial tissue culture media formulation. The inclusion of Si to the culture medium improved the morphogenetic potential of plant cells, tissues and organs. Several studies have shown that the inclusion of Si to the tissue culture medium enhances callus growth, shoot regeneration, and root induction and stimulates somatic embryogenesis, and improve morphological, anatomical and physiological characteristics of plantlets. In addition, Si treatment prolongs the longevity of calli and organs with a potential for plant regeneration. The inclusion of Si to the tissue culture medium also enhances tolerance to low temperature, metal toxicity and salinity. Si enhancing tolerance of plants to various stresses by altering activity of antioxidant enzymes, cation binding capacity of the cell walls, endogenous plant hormone level, increasing production of chitinase, glucanse, lignin, phenolics, and phytoalexins, nutrient uptake, improving strength of cell and plant, maintaining the structure of stomata, relative water content, and reducing uptake of heavy metals. This review concentrates the potential roles of Si in plant tissue culture.
Islam et al. (2005) investigated the effect of calcium silicate (CaSiO3) on callus induction and plant regeneration from mature seed explants of rice ‘Kalizira’, ‘Lucky’, and ‘Pajam’. The highest frequency of callus induction is achieved on Murashige and Skoog (MS) medium containing CaSiO3. However, plant or root regeneration potential of rice calli is cultivar depended. Similarly, effects of Si on plant or root development depend on reed (Phragmites australis) genotype used for callus induction (Mathe et al., 2012). Addition of Si as sodium silicate (Na2SiO3) to the modified MS medium promotes the growth of calli obtained from stem nodal and root explants of P. australis while its effect on somatic embryogenesis is explants dependent: it stimulates embryogenesis of root calli, but it does not influence this process in stem nodal calli. Soares et al. (2011) evaluated the effect of Si source [potassium silicate (K2SiO3) and Na2SiO3] on shoot multiplication of Cattleya loddigesii. The highest number of shoots is observed on the modified Knudson C medium containing 5.0 mg L-1 K2SiO3. In Ajuga multiflora, addition of Si to MS medium containing 2iP and IAA, enhanced adventitious shoot regeneration (about threefold) by increasing the activity of antioxidant enzymes such as SOD, POD, APX, and CAT (Sivanesan and Jeong, 2014). In addition, the authors observed the Si accumulation in leaves of plants developed in the culture medium with Si, but not in plants developed in the medium without Si by wavelength dispersive X-ray analysis. These studies indicate that the effect of Si on morphogenetic potential of in vitro plant cultures depends on plant species, genotype and concentration of Si in the culture medium. Still further studies are required to better understand the biochemical and molecular mechanism of Si on organogenesis and somatic embryogenesis.
The application of Si has been reported to enhance the growth and development of various plants. Zhou (1995) observed silica bodies in leaf tissues of Phalaenopsis hybrid plantlets grown in Vacin and Went medium supplemented with CaSiO3. Addition of CaSiO3 also increased the leaf length. Soares et al. (2011) reported that the addition of 5.0 mg L-1 K2SiO3 and 20.0 mg L-1 Na2SiO3 to the modified Knudson C medium increased the number of roots and length of aerial part and root in seedlings of C. loddigesii. Subsequently the same group investigated the effect of GA3 and Na2SiO3 on growth and development of C. loddigesii (Soares et al., 2013). It was reported that the combination of GA3 and Na2SiO3 increased the number of leaves and roots than GA3 alone. The optimal concentration of Si varies within the same plant species and or genotype. The inclusion of CaSiO3 at 0.5 and 2.0 mg L-1 to the MS medium stimulates the growth of native (Brassavolva perrinii) and hybrid (Laelia cattleya ‘Culminant Tuilerie’ ×L. cattleya ‘Sons Atout Rotunda’) × Brassolaelia cattleya ‘Startifire Moon Beach’) orchid plants, respectively (Soares et al., 2012). Lim et al. (2012) also reported that the effect of Si (K2SiO3) on the growth traits of begonia ‘Super Olympia Red’ and ‘Super Olympia Rose’ and pansy ‘Matrix White Blotch’ and ‘Matrix Yellow Blotch’ are mainly dependent on the cultivars. Braga et al. (2009) investigated the effect of different Si sources such as CaSiO3, K2SiO3, and Na2SiO3 on the growth and anatomical characteristics of strawberry ‘Oso Grande’ seedlings. The fresh and dry weight of seedlings increased in MS medium containing 1.0 g L-1 Na2SiO3. Seedlings of banana ‘Maca’ cultured in the medium supplemented with CaSiO3 increased the chlorophyll content, whereas those cultured in the medium containing Na2SiO3 increased length, fresh and dry weight of shoots (Asmar et al., 2011).
The morphological and anatomical characteristics of in vitro-grown plantlets are different from the field-grown seedlings. Si inclusion to the rooting medium increased leaf tissue thickness and epicuticular wax deposition in banana (Asmar et al., 2013a) and strawberry (Braga et al., 2009) plantlets. Luz et al. (2012) reported that supplementation of CaSiO3, K2SiO3, or Na2SiO3 to the rooting medium improved leaf anatomy of banana ‘Maca’ plantlets. The inclusion of CaSiO3 to the culture medium also increased photosynthetic rate and chlorophyll content of banana plantlets (Asmar et al., 2013b). In strawberry, light and electron microscopic analysis showed deformation in chlorenchyma and the epidermis of leaves from plantlets grown in the culture medium devoid of Si (Soares et al., 2012). Recently, He et al. (2013) confirmed the deposition of Si within the cell walls of in vitro-cultured rice cells. Si improves the structural stability of cell walls during cell elongation and division and thereby maintained cell shape, which may be important for the function and survival of cells (Table 1).
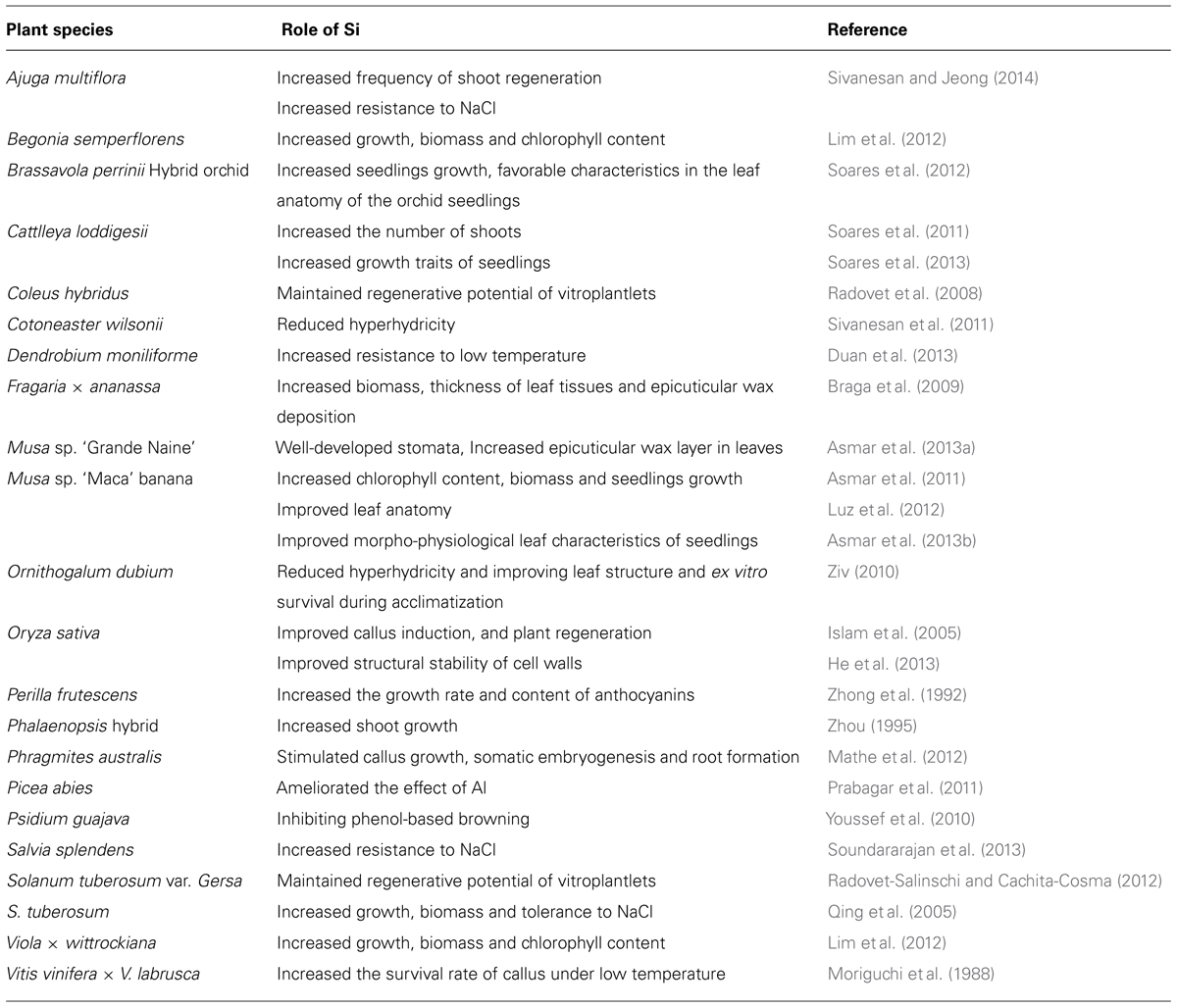
TABLE 1. Role of Si in plant tissue culture.
Ziv (2010) investigated the effect of silicon on hyperhydricity in Ornithogalum dubium. Addition of Na2SiO3 to MS liquid medium containing BA, NAA and 6% sucrose in bioreactors, significantly reduced induction of hyperhydric shoots, and increased plant firmness and mechanical strength. Si treatment significantly reduced the content of hydrogen peroxide and activity of oxidative reductive enzymes such as APX, ascorbate oxidase and GPX in leaves of the regenerated shoots of O. dubium when compared with the control (Table 1). Similarly, addition of Si as K2SiO3 to MS medium reduced the hyperhydricity in Cotoneaster wilsonii by decreasing the content of MDA in the regenerated shoots when compared with the control (Sivanesan et al., 2011). The authors observed the presence of Si in the in non-hyperhydric plants, but not in the hyperhydric leaf samples of C. wilsonii by energy dispersive X-ray analysis. Thus, the problem of hyperhydricity can be reduced by the inclusion of Si to both liquid and solid culture medium. Phenolic oxidative tissue browning is one of the bottlenecks in woody plant tissue culture. In guava, tissue browning was completely prevented by sealing the nodal explants cut ends with Si (Youssef et al., 2010) and there was no detrimental effect of Si on the subsequent steps of in vitro propagation. The authors suggested that Si could be used during explants preparation to control phenolic tissue browning in other plants. The morphological, anatomical and physiological characteristics of plantlets can improve in vitro by incorporating Si in the culture medium. However, further studies required to evaluate the effect of different source and concentration of silicon on the growth and development of various plants.
Duan et al. (2013) reported that Si enhance cold resistance of Dendrobium moniliforme by increasing the content of free proline, soluble sugar and soluble protein and decreasing MDA content. Si treatment improved the survival rate of grape ‘Kyoho’ and ‘koshusanjaku’ calli under low temperature by preventing browning (Moriguchi et al., 1988). In vitro storage of Coleus hybridus ‘Jupiter’ and Solanum tuberosum var. Gersa under silicone oil significantly reduced the growth and maintained their regenerative potential (Radovet et al., 2008; Radovet-Salinschi and Cachita-Cosma, 2012). These results reveal that Si can be used as cryoprotectant and included in the cryoprotective mixture for minimizing the toxicity of cryoprotectants. The ameliorating effect of Si on salt stress in vitro has been reported in A. multiflora (Sivanesan and Jeong, 2014), Salvia splendens ‘Hot Jazz’ (Soundararajan et al., 2013) and S. tuberosum (Qing et al., 2005). Si alleviates salt stress in plants by limiting NaCl uptake, maintenance of ultrastructure of stomata, improving photosynthetic activity, reducing free proline content and altering the production of antioxidant enzymes (Qing et al., 2005; Soundararajan et al., 2013; Sivanesan and Jeong, 2014). Prabagar et al. (2011) investigated the effect of Si on aluminium (Al) tolerance in Picea abies suspension cultures. Al toxicity was reduced when the liquid medium was supplemented with Si and the effect was increased at pH 5.0 than pH 4.2. Si supplementation protected P. abies cells and against Al toxicity by reducing the concentration of free Al in the cell wall. Si is also reported to enhance drought tolerance, alleviate lead toxicity and increase resistance to radiation and temperature stresses (Balakhnina and Borkowska, 2013; Zhu and Gong, 2014). The molecular mechanisms of Si on stress tolerance are poorly understood. Thus, more studies are needed to find out the role of Si in abiotic tolerance on various plants.
Recent studies have shown the beneficial effects of Si in plant tissue culture (Table 1). However, further studies on a wide variety of plant species are needed to confirm the role of Si in plant tissue culture. In vitro culture is a useful system for studying physiological and biochemical functions of Si in plants at molecular level. Further, in vitro cell suspension culture systems provide an opportunity to study roles of Si at the single cell level. The inclusion of silicone A to Linsmaier and Skoog liquid medium also enhances cell growth and anthocyanins content in cell suspension culture of Perilla frutescens (Zhong et al., 1992). Thus, Si can also be used for the stimulation of secondary metabolites in the plant cell, tissue and organ cultures. We strongly recommend the inclusion of Si as a beneficial nutrient in the tissue culture medium to solve various micropropagation problems, and to increase tissue culture success.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
This article was supported by the KU Research Professor Program of Konkuk University.
Asmar, S. A., Pasqual, M., de Araujo, A. G., Silva, R. A. L., Rodrigues, F. A., and Pio, L. A. S. (2013a). Morphophysiological characteristics of acclimatized “Grande Naine” banana plants in response to in vitro use of silicon. Cienc. Agrar. 34, 73–82.
Asmar, S. A., Castro, E. M., Pasqual, M., Pereira, F. J., and Soares, J. D. R. (2013b). Changes in leaf anatomy and photosynthesis of micropropagated banana plantlets under different silicon sources. Sci. Hortic. 161, 328–332. doi: 10.1016/j.scienta.2013.07.021
Asmar, S. A., Pasqual, M., Rodrigues, F. A., de Araujo, A. G., Pio, L. A. S., and e Silva, S. O. (2011). Sources of silicon in the development of micropropagated seedlings of banana “Maçã .” Cienc. Rural. 41, 1127–1131. doi: 10.1590/S0103-84782011005000086
Balakhnina, T., and Borkowska, A. (2013). Effects of silicon on plant resistance to environmental stresses: review. Int. Agrophys. 27, 225–232. doi: 10.2478/v10247-012-0089-4
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Braga, F. T., Nunes, C. F., Favero, A. C., Pasqual, M., Carvalho, J. G., and Castro, E. M. (2009). Anatomical characteristics of the strawberry seedlings micropropagated using different sources of silicon. Pesqui. Agropecu. Bras. 44, 128–132. doi: 10.1590/S0100-204X2009000200003
Duan, X., Tang, M., and Wang, W. (2013). Effects of silicon on physiology and biochemistry of dendrobium moniliforme plantlets under cold stress. Agric. Biotechnol. 2, 18–21. doi: 10.1007/s13593-011-0039-8
Epstein, E. (1999). Silicon. Annu. Rev. Plant. Physiol. Plant Mol. Biol. 50, 641–664. doi: 10.1146/annurev.arplant.50.1.641
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
He, C., Wang, L., Liu, J., Liu, X., Li, X., Ma, J.,et al. (2013). Evidence for “silicon” within the cell walls of suspension-cultured rice cells. New Phytol. 200, 700–709. doi: 10.1111/nph.12401
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Islam, M. M., Ahmed, M., and Mahaldar, D. (2005). In vitro callus induction and plant regeneration in seed explants of rice (Oryza sativa L.). Res. J. Agri. Biol. Sci. 1, 72–75.
Lim, M. Y., Lee, E. J., Jana, S., Sivanesan, I., and Jeong, B. R. (2012). Effect of potassium silicate on growth ad leaf epidermal characteristics of begonia and pansy grown in vitro. Kor. J. Hort. Sci. Technol. 30, 579–585.
Luz, J. M. Q., Asmar, S. A., Pasqual, M., de Araujo, A. G., Pio, L. A. S., and Resende, R. F. (2012). Modification in leaf anatomy of banana plants cultivar “Maca” subjected to different silicon sources in vitro. Acta Hort. 961, 239–243.
Ma, J. F. (2004). Role of silicon in enhancing the resistance of plants to biotic and abiotic stresses. Soil Sci. Plant Nutr. 50, 11–18. doi: 10.1080/00380768.2004.10408447
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Mathe, C., Mosolygo, A., Suranyi, G., Beke, A., Demeter, Z., Toth, V. R.,et al. (2012). Genotype and explants-type dependent morphogenesis and silicon response of common reed (Phragmites australis) tissue cultures. Aquat. Bot. 97, 57–63. doi: 10.1016/j.aquabot.2011.11.005
Moriguchi, T., Kozaki, I., Matsuta, N., and Yamaki, S. (1988). Plant regeneration from grape callus stored under a combination of low temperature and silicone treatment. Plant Cell Tissue Organ Cult. 15, 67–71. doi: 10.1007/BF00039890
Prabagar, S., Hodson, M. J., and Evans, D. E. (2011). Silicon amelioration of aluminium toxicity and cell death in suspension cultures of Norway spruce (Picea abies L.). Environ. Exp. Bot. 70, 266–276. doi: 10.1016/j.envexpbot.2010.10.001
Qing, W., Huiying, H., and Jinwen, Z. (2005). Effect of exogenous silicon and proline on potato plantlet in vitro under salt stress. China Vegetables 9, 16–18.
Radovet, D., Cachita-Cosma, D., and Petrus, A. (2008). The micropropagation of Coleus hybridus Jupiter vitrocultures under different paraffin or silicon oil stratums. Studia Universitatis “Vasile Goldiş”, Seria Ştiinte Vieţii (Life Sciences Series) 18, 61–70.
Radovet-Salinschi, D., and Cachita-Cosma, D. (2012). Testing the regenerative capacity of Solanum tuberosum var. Gersa explants after 24 weeks storage in living collection. Analele Universitǎţii din Oradea, Fascicula: Ecotoxicologie, Zootehnie şi Tehnologii de Industrie Alimentarǎ? 11, 423–430.
Reed, B. M., Wada, S., DeNoma, J., and Niedz, R. P. (2013). Mineral nutrition influences physiological responses of pear in vitro. In Vitro Cell. Dev. Biol. Plant 49, 699–709. doi: 10.1007/s11627-013-9556-2
Sivanesan, I., and Jeong, B. R. (2014). Silicon promotes adventitious shoot regeneration and enhances salinity tolerance of Ajuga multiflora Bunge by altering activity of antioxidant enzyme. Sci. World J. 2014, 10 doi: 10.1155/2014/521703
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Sivanesan, I., Song, J. Y., Hwang, S. J., and Jeong, B. R. (2011). Micropropagation of Cotoneaster wilsonii Nakai—a rare endemic ornamental plant. Plant Cell Tissue Organ Cult. 105, 55–63. doi: 10.1007/s11240-010-9841-2
Soares, J. D. R., Pasqual, M., de Araujo, A. G., De Castro, E. M., Pereira, F. J., and Braga, F. T. (2012). Leaf anatomy of orchids micropropagated with different silicon concentrations. Acta Sci. Agron. 34, 413–421. doi: 10.4025/actasciagron.v34i4.15062
Soares, J. D. R., Pasqual, M., Rodrigues, F. A., Villa, F., and de Araujo, A. G. (2011). Silicon sources in the micropropagation of the Cattleya group orchid. Acta Sci. Agron. 33, 503–507.
Soares, J. D. R., Villa, F., Rodrigues, F. A., and Pasqual, M. (2013). Concentrations of silicon and GA3 in in vitro propagation of orchids under natural light. Sci. Agrar. Parana. 12, 286–292.
Soundararajan, P., Sivanesan, I., Jo, E. H., and Jeong, B. R. (2013). Silicon promotes shoot proliferation and shoot growth of Salvia splendens under salt stress in vitro. Hort. Environ. Biotechnol. 54, 311–318. doi: 10.1007/s13580-013-0118-7
Van Bockhaven, J., De Vleesschauwer, D., and Höfte, M. (2013). Towards establishing broad-spectrum disease resistance in plants: silicon leads the way. J. Exp. Bot. 64, 1281–1293. doi: 10.1093/jxb/ers329
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Voogt, W., and Sonneveld, C. (2001). “Silicon in horticultural crops grown in soilless culture,” in Silicon in Agriculture, eds L. E. Datnoff, G. H. Snyder, and G. H. Korndorfer (Amsterdam: Elsevier Science), 115–131.
Youssef, M. A., El-Helw, M. R., Taghian, A. S., and El-Aref, H. M. (2010). Improvement of Psidium guajava L. using micropropagation. Acta Hort. 849, 223–230.
Zhong, J. J., Seki, T., Kinoshita, S. I., and Yoshida, T. (1992). Effects of surfactants on cell growth and pigment production in suspension cultures of Perilla frutescens. World J. Microbiol. Biotechnol. 8, 106–108. doi: 10.1007/BF01195826
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Zhou, T. S. (1995). The detection of the accumulation of silicon in Phalaenopsis (Orchidaceae). Ann. Bot. 75, 605–607. doi: 10.1006/anbo.1995.1065
Zhu, Y., and Gong, H. (2014). Beneficial effects of silicon on salt and drought tolerance in plants. Agron. Sustain. Dev. 34, 455–472. doi: 10.1007/s13593-013-0194-1
Keywords: acclimatization, epicuticular wax deposition, hyperhydricity, organogenesis, silicon, stress tolerance
Citation: Sivanesan I and Park SW (2014) The role of silicon in plant tissue culture. Front. Plant Sci. 5:571. doi: 10.3389/fpls.2014.00571
Received: 01 August 2014; Accepted: 04 October 2014;
Published online: 21 October 2014.
Edited by:
Agnieszka Ludwików, Adam Mickiewicz University, PolandReviewed by:
Chanyarat Paungfoo-Lonhienne, The University of Queensland, AustraliaCopyright © 2014 Sivanesan and Park. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Se Won Park, Department of Molecular Biotechnology, Konkuk University, 1 Hwayang-dong, Gwangjin-gu, Seoul 143-701, South Korea e-mail:c2V3cGFya0Brb25rdWsuYWMua3I=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.