 Ruth Grene1*
Ruth Grene1* Curtis Klumas1,2
Curtis Klumas1,2 Haktan Suren2,3
Haktan Suren2,3 Kuan Yang1,2 Eva Collakova1 Elijah Myers2,4 Lenwood S. Heath4
Kuan Yang1,2 Eva Collakova1 Elijah Myers2,4 Lenwood S. Heath4 Jason A. Holliday3
Jason A. Holliday3- 1Department of Plant Pathology, Physiology, and Weed Science, Virginia Tech, Blacksburg, VA, USA
- 2Genetics, Bioinformatics and Computational Biology Program, Virginia Tech, Blacksburg, VA, USA
- 3Department of Forest Resources and Environmental Conservation, Virginia Tech, Blacksburg, VA, USA
- 4Department of Computer Science, Virginia Tech, Blacksburg, VA, USA
Microarray gene expression profiling is a powerful technique to understand complex developmental processes, but making biologically meaningful inferences from such studies has always been challenging. We previously reported a microarray study of the freezing acclimation period in Sitka spruce (Picea sitchensis) in which a large number of candidate genes for climatic adaptation were identified. In the current paper, we apply additional systems biology tools to these data to further probe changes in the levels of genes and metabolites and activities of associated pathways that regulate this complex developmental transition. One aspect of this adaptive process that is not well understood is the role of the cell wall. Our data suggest coordinated metabolic and signaling responses leading to cell wall remodeling. Co-expression of genes encoding proteins associated with biosynthesis of structural and non-structural cell wall carbohydrates was observed, which may be regulated by ethylene signaling components. At the same time, numerous genes, whose products are putatively localized to the endomembrane system and involved in both the synthesis and trafficking of cell wall carbohydrates, were up-regulated. Taken together, these results suggest a link between ethylene signaling and biosynthesis, and targeting of cell wall related gene products during the period of winter hardening. Automated Layout Pipeline for Inferred NEtworks (ALPINE), an in-house plugin for the Cytoscape visualization environment that utilizes the existing GeneMANIA and Mosaic plugins, together with the use of visualization tools, provided images of proposed signaling processes that became active over the time course of winter hardening, particularly at later time points in the process. The resulting visualizations have the potential to reveal novel, hypothesis-generating, gene association patterns in the context of targeted subcellular location.
Introduction
Annual oscillations between growth and dormancy are unique features of woody perennials adapted to life in temperate and boreal regions worldwide (Rohde and Bhalerao, 2007). The timing of these transitions is driven by day length and temperature, and underlies the basic physiological principle that growth and substantial freezing tolerance are not compatible (Weiser, 1970). While cold and freezing tolerance are sometimes confounded in the literature, there are important differences between these two processes. Many annual plants have gene regulatory networks that respond to low, above-freezing temperatures (chilling), but because extra- and intra-cellular ice formation presents a suite of different physiological challenges compared to those presented by chilling, many of these same plants are killed by temperatures much below freezing. While much is known about the physiological basis for cold and freezing injury and tolerance in both annuals and perennials, and substantial effort has been devoted to uncovering the genes involved in transient cold tolerance in annuals (Smallwood and Bowles, 2002), much less is known about the genetic and genomic bases for acclimation to freezing in perennials. The distinction between the two is important for two reasons. First, as mentioned above, freezing is a fundamentally different stress than chilling, and second, plant species that have adopted the perennial habit do not wait for cold temperatures to acclimate. Instead, they use annual environmental cues, most often a critical night length, to begin acclimating to impending freezing temperatures well in advance of the need for freezing tolerance (Weiser, 1970). In this way, both the upstream regulatory network and downstream molecular physiologies of seasonal freezing tolerance are different than the cognate processes for cold tolerance in annuals.
In addition to the different metabolic remodeling involved in chilling and freezing tolerance in annuals and perennials, respectively, a distinction can be made between deciduous and evergreen perennials. Several studies have characterized the transcriptional and metabolic remodeling that occurs in angiosperm trees, mostly in poplar (Populus) species, during the transition from growth to dormancy. Ruttink et al. (2007) monitored gene expression and soluble metabolites in hybrid poplar (Populus tremulax P. alba) vegetative buds for 6 weeks following the transition to short days and proposed a model for bud dormancy acquisition in which the initial long night signal progresses through ethylene and abscisic acid (ABA)-mediated processes finally leading to endodormancy. As vegetative buds are the structure that perceive and regulate dormancy, the transcriptome and metabolome changes described in that study comprise both processes related to bud dormancy as well as freezing acclimation. Druart et al. (2007) took a similar approach to characterizing molecular events of the growth and dormancy cycle in the cambial meristem of European aspen (P. tremula), and identified several distinct stages of transcriptional and metabolic remodeling associated with freezing acclimation (Uemura et al., 2006). Many canonical chilling tolerance-related transcripts were induced well before the onset of freezing temperatures, providing a link to phenotypic measurements in trees showing that deep freezing tolerance is typically acquired before it is needed in most years.
Whereas the primary role of freezing tolerance in angiosperm trees is to protect the cambial meristem and vegetative buds, in evergreen species, which do not shed their leaves in winter, freezing tolerance involves both acclimation of the cambial and shoot meristems, as well as the leaves. Most angiosperm perennials that inhabit cold climates have broad leaves, and few retain them during winter. In contrast to angiosperms, nearly all gymnosperms from cold climates retain their leaves (needles) year-round (with the notable exception of larch species (Larix spp.). In addition, gymnosperm trees tend to be ecologically dominant in very cold climates, and understanding the molecular events that allow such species to retain their photosynthetic tissue (although photosynthesis may be greatly reduced) in the face of extreme cold is a basic biological question that cannot be addressed in angiosperms such as Populus spp. Several studies have used gene expression microarrays to study the transition to dormancy in gymnosperms in both the Picea and Pinus genera. In the first such study, Joosen et al. (2006) (Ruonala et al., 2008; Mohamed et al., 2010) employed a 1.5 k cDNA microarray to identify a number of candidate genes for freezing tolerance. More recently, white spruce (Picea glauca) stems were sampled at multiple time points following the transition to short days and quantified for both transcript (11 k cDNA microarray) and protein (2D-PAGE) levels (Galindo Gonzalez et al., 2012). Interestingly, several putative spruce orthologs of the Arabidopsis flowering-time pathway were differentially expressed, strengthening the link between this pathway and light-mediated dormancy signaling in perennials (Bohlenius et al., 2006). To date, the only study of transcriptome remodeling in photosynthetic tissue during freezing tolerance acquisition in a gymnosperm was in Sitka spruce (Picea sitchensis), for which five time points from late summer to early winter were sampled and gene expression monitored using a 21.8 k array (Holliday et al., 2008). Holliday et al. focused primarily on downstream cryoprotective genes, lipids, soluble carbohydrates, and upstream signaling components (e.g., calcium, daylength perception) related to freezing acclimation. However, they did not discuss in detail genes assigned to several “gene ontology (GO)” terms that were overrepresented, including “transport,” “transporter activity,” “endoplasmic reticulum,” and “golgi apparatus.” As little is known about the role of the endomembrane system with regard to the process of freezing acclimation, we sought to more deeply probe expression patterns and possible function of the relevant genes. In the current paper, we employ several novel analytical tools to dissect the role of the secretory system, and its relationship to cell wall remodeling, with respect to cold hardiness in spruce.
Methods
Plant Material, Microarray Analysis, and Metabolite Profiling
The plant material and resulting gene expression data for this study are described in Holliday et al. (2008). Briefly, a common garden comprising Sitka spruce originating from 17 geographic populations was grown in an outdoor, raised-bed common garden at Vancouver, BC, Canada. In the fourth season of growth, plants from a population originating near the center of the species range were sampled for RNA extraction and phenotyped by an artificial freeze test that uses electrolyte leakage as a proxy for cell death in sectioned needles frozen at several test temperatures. In this case, the index of injury (It) at −10°C was used. Needle tissue was sampled at five time points: August 30, October 18, November 22, December 1, and December 13 (2004). Holliday et al. (2008) showed that using this freeze test, the plants began acclimating to freezing after October 18, and reached maximal levels of hardiness on the fourth time point (December 1). Hereafter, expression changes associated with these time points are referred to as TP1 for the ratio of October 18: August 30, TP2 for the ratio of November 22: August 30, and so on. RNA was extracted from these needles, followed by cDNA synthesis and hybridization to a 21.8 k spruce cDNA microarray. Slides were scanned and spot intensity quantified using ImaGene software (BioDiscovery, Inc., El Segundo, CA). Data were background-subtracted, normalized by variance stabilizing normalization (VSN) (Huber et al., 2002), and significant changes in gene expression were identified using a linear mixed-effects model as described in Holliday et al. (2008). P-values for each gene-by-treatment effect were adjusted for the false discovery rate (FDR) (Storey and Tibshirani, 2003). In addition to transcriptome analysis, data on soluble metabolites quantified by GC-MS analysis were taken from Dauwe et al. for the same samples (Dauwe et al., 2012). All statistical analyses were carried out using the R statistical package (www.r-project.org).
Use of MapMan to Reveal Pathways Enriched Over the Hardening Period
The MapMan tool facilitates the classification and statistical analysis of transcripts and metabolites into hierarchical categories (known as bins) in a manner that avoids the redundancy present in other commonly used ontologies (Usadel et al., 2009); therefore, the tool provides an additional level of analysis beyond the functional enrichment of typical GO categories. A further, in–house modification of MapMan was made to allow visual comparisons of changes over time by the inclusion of the ability to view multiple instances of a bin simultaneously. Using this viewpoint, the user is presented with multiple copies of the same bin, where each copy displays the expression of transcripts or metabolites annotated to the bin for the selected condition. This visualization technique, which is similar to Tufte's small multiples concept (Tufte, 1983), is based on the principle that different visualization techniques may be more or less effective depending on the data analysis task that is being performed (Saraiya et al., 2005). As an additional analytical viewpoint, the user is presented with a list of each gene or metabolite annotated to the visualized bin that demonstrates the change in each entity individually.
Functional Gene Ontology Analysis
Functional properties of the differentially expressed clones were categorized based on the GO database (Ashburner et al., 2000) using ontologizer (Bauer et al., 2008). First, putative orthologs of Sitka spruce genes on the microarray were identified using a custom BLAST (Altschul et al., 1997) database comprised of all the Arabidopsis sequences annotated by TAIR (Lamesch et al., 2012) and BLAST executables (http://www.ncbi.nlm.nih.gov) were integrated to automate this process. Over-represented GO terms were identified using the parent-child-union algorithm implemented in Ontologizer. This method is preferred over the other methods because it scores each GO term considering the presence of the parent term rather than taking only the individual GO contributions into account. The Westfall-Young single step error correction was used to adjust for multiple testing in the identification of significant GO terms (Westfall and Young, 1993). An FDR of less than 1% was considered statistically significant.
Automated Gene Association Network Generation Using ALPINE
As genes that showed large and statistically significant changes in expression across time points were identified, the Automated Layout Pipeline for Inferred NEtworks (ALPINE) tool was used to provide context regarding the associations among the identified genes and other genes showing statistically significant changes in expression in the data set. The ALPINE tool, implemented as a plugin for the Cytoscape visualization environment (Shannon et al., 2003), performs the following steps when processing a set of query genes: first, the genes are used as a query set to generate a network of gene associations (co-expression, co-localization, genetic interaction, physical interaction, and more) using the GeneMANIA (Montojo et al., 2010) Cytoscape plugin; second, the resulting gene association network is then filtered to include only those genes that showed a statistically significant expression value in a user-defined minimum number of time points; finally, the filtered network is organized into a layout that uses node color to represent GO molecular function annotations, with nodes drawn over a background image of cellular components using GO cellular component annotations. The annotations and layout are executed using the Mosaic Cytoscape plugin (Zhang et al., 2012). (Specific use of the ALPINE tool is further described in the section “Results”).
Results
GO Enrichment Reveals a Relationship Between Endomembrane Trafficking and Cell Wall Remodeling During Freezing Acclimation
We used Ontologizer software to perform GO enrichment analysis across cell components (Figures 1A,B), which revealed that genes associated with endomembrane function were up regulated during the time course of winter hardening (Figure 1A). Since most cell wall components are synthesized in and transported by the endomembrane secretory system (Carpita, 2011), results for the two broad categories were examined together. In Figure 2, the MapMan bin for each gene and an indication of when a statistically significant change in expression occurred are displayed in heat map form. A distinction between genes annotated as “cellulose synthase” (CES) vs. “cellulose synthase-like” (CSL) enzymes should be made, as CES are involved in the synthesis of cellulose, while CSL are involved in the synthesis of mannan oligomers and xyloglucans (Davis et al., 2010). CESA1 (At4g32410) showed no changes in transcript levels, except up-regulation in TP2 while CESA4 (At5g44030) was down-regulated in TP1, 2, and 4. In contrast, the xylem-specific CES IRX3 (CESA7, At5g17420) was up-regulated in TP1 and 3. Overall, CSL genes showed no changes in expression in TP1 and 2, but CSL12 was up-regulated and CSLG4 down-regulated in TP3 and 4. Both CSLB03 and E1 were consistently down-regulated through TP2 and 4. Cobra-like O-glycosyl hydrolase 4 (At5g15630), involved in xylan and secondary cell wall biosynthesis (Oikawa et al., 2010), exhibited an elevated expression between TP3 and 4. Table 1 summarizes changes in the levels of metabolites relevant to cell wall biosynthesis during winter hardening. Levels of the metabolite glucose-6-phosphate, a precursor of the cellulose biosynthetic pathway, showed an increase over the time course, while glucose-1,6-bisphosphate exhibited slightly higher but not significant levels in TP1 through TP3, followed by a decline at TP4. The levels of arabinose released during cell wall degradation decreased at TP1 and remained low for the remainder of the time course.
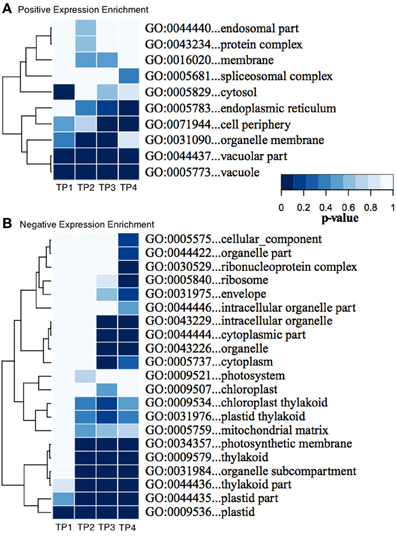
Figure 1. (A,B) Enrichment of GO “Cellular Component” categories across the time series of winter hardening. The GO tool Ontologizer was used to assess statistical enrichment of genes related to the cellular component ontology. Only GO terms with significant enrichment for at least one time point are shown. Colors correspond to probability values for individual terms.
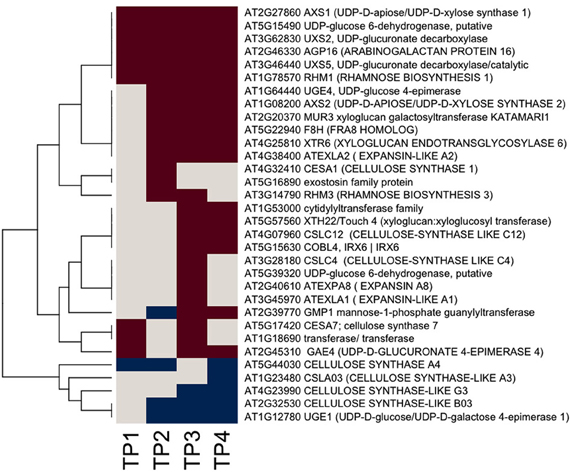
Figure 2. Heat map of significantly responsive cell wall associated genes across the time course of winter hardening. The gene expression levels across the rows are hierarchically clustered. Magenta cells represent significant positive expression changes, blue cells represent significant negative changes, and off-white cells represents no change or no statistically significant change.
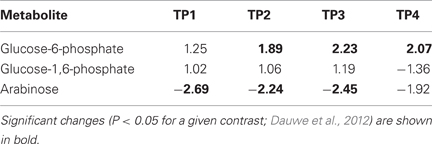
Table 1. Changes in steady-state metabolite levels during winter hardening.
A somewhat different pattern was observed in the case of genes in the subcategories corresponding to the biosynthesis of cell wall carbohydrate precursors (Figure 2), where an overall up-regulation and fewer decreases in gene expression over the time course were observed in the case of this group. UDP-D-glucose/UDP-D-galactose 4-epimerase 1 (UGE1, At1g12780) converts UDP-galactose to UDP-glucose, and it was the only gene exhibiting a consistent down-regulation from TP2 toTP4. In contrast, UGE4 (At1g64440) showed an opposite trend to UGE1. Most genes encoding different enzymes involved in cellulose biosynthesis showed increases at TP1, with a consistent pattern of increases in transcript levels thereafter over the remainder of the time course. This group included one of the genes encoding UDP-glucose-6-dehydrogenase (At5g15490) that catalyzes the first step in the cellulose biosynthesis pathway, specifically the oxidation of UDP-glucose to UDP-glucuronate, and a number of genes encoding enzymes of the subsequent steps. These enzymes included two UDP-glucuronate decarboxylases (At3g62830 and At3g46440) and UDP-glucuronate 4-epimerase 4 (At2g45310), which was consistently up-regulated except for TP2. Similar trends were observed for genes encoding enzymes that participate in the synthesis of the subsequent carbohydrate precursors of the cell wall from UDP-glucuronate, including UDP-mannose, -rhamnose, -xylose, and -apiose. Genes encoding GDP-mannose 4,6-dehydratase 1 (At5g66280), rhamnose biosynthesis 1 (At1g78570), and UDP-apiose/UDP-xylose synthase 1 (At2g27860) remained increased at and beyond TP1. Other homologous genes encoding these enzymes were up-regulated less consistently during different time points.
Xyloglucans are major components in hemicelluloses in primary cell walls and are synthesized via various glycosyltransferases. The gene encoding xyloglucan:xyloglucosyltransferase (At5g57560, XTH22) that catalyzes the modification of xyloglucan oligomers was up-regulated during TP3 and 4, while xyloglucanendotransglycosylase 6 (XTR6, At4g25810) and xyloglucangalactosyltransferase MUR3 [At2g20370, Golgi (Chevalier et al., 2010)] were also up-regulated at TP2. F8H glycosyltransferase (At5g22940) is involved in the biosynthesis of glucuronoxylans in Arabidopsis that are part of the secondary cell walls in the xylem (Lee et al., 2009). The corresponding gene in Sitka spruce showed up-regulation patterns comparable to XTR6 and MUR3. Some members of the expansin gene family showed up-regulation, e.g., expansin A8 and expansin-like A1 in TP3, while expansin-like A2 from TP2 to 4. The arabinogalactan protein (AGP)-encoding gene family showed strikingly diverse patterns of expression over the time course of winter hardening, with an AGP16 homolog increasing during all time points in winter hardening.
Secretory System
Given that the biosynthesis of cell wall matrix polysaccharides and other cell wall components takes place in the Golgi apparatus (Driouich et al., 2012), an examination of the responsiveness of genes encoding proteins associated with the endomembrane system and associated secretory processes was warranted. The assignment of responsive genes to the “protein targeting secretory pathway” bin in MapMan was taken as a starting point, and the assignment of those genes were further refined using information from PubMed on individual genes, together with information from GO. Results are shown in heat map form in Figure 3. The majority of responsive genes associated with the secretory system were up-regulated during the time course of hardening, suggesting extensive secretory activity. One group of genes was up regulated across the entire time course. This included At3g48570, encoding the gamma subunit of SEC61, a protein transporter, (At2g14740), a vacuolar sorting receptor, and MUR3 (At2g20370). Another group showed up-regulation from TP2 onwards, including At1g11890, encoding SEC22 (El-Kasmi et al., 2011), whose function has been shown to be essential for the integrity of the Golgi (Table 3).
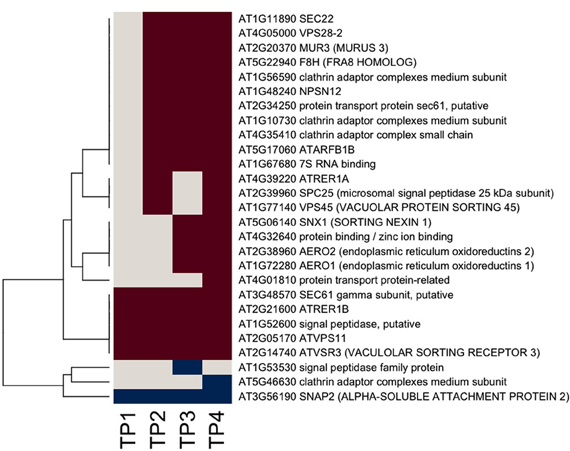
Figure 3. Heat map of significantly responsive endomembrane/secretory pathway associated genes across the time course of winter hardening. The gene expression levels across the rows are hierarchically clustered. Magenta cells represent significant positive expression changes, blue cells represent significant negative changes, and off-white cells represent no change or no statistically significant change.
Genes encoding secretory pathway proteins and cell wall associated functions that were significantly up-regulated at a minimum of one time point are grouped according to compartment, and direction of movement of the protein is indicated, where known (Figure 4; Tables 1 and 2). It is apparent that several genes encoding proteins targeted to the cell membrane and another set destined for the Golgi apparatus responded positively during winter hardening, with comparatively little response among peroxisomal or nuclear-targeted gene products. In the case of the cell membrane, five responsive genes associated with secretion were up-regulated. The other four up-regulated genes encoding homologs of transport/secretion proteins showed an initial change, reflected in the TP2/TP1 comparison, with increases shown over the remaining time points. In the case of proteins localized to the Golgi, ten genes responded positively over the time course. The positive responders included ATRER1A and ATRER1B (At2g21600 and At4g39220, respectively), which facilitate retrograde transport from the Golgi to the ER. Three genes, encoding proteins associated with anterograde transport, from ER to Golgi, were up-regulated, including ATSEC22 (At1g11890). In the case of the other two genes in this category, At4g01810 and At4g32640, an initial decrease was observed at the TP2/TP1 comparison, followed by increases between TP3 and 2. Two genes encoding proteins that are targeted to the trans-golgi network (TGN) were also up-regulated over the time course, including one (At1g77140) that is targeted to the tonoplast. Another gene, At2g14740, is annotated as being part of the “Golgi transport complex,” part of the cell membrane, and also targeted to the vacuole. Such complex annotations may reflect the mobility of proteins within the endomembrane system. In the case of the ER, two genes encoding proteins that catalyze thiol-disulfide exchange were also up-regulated.
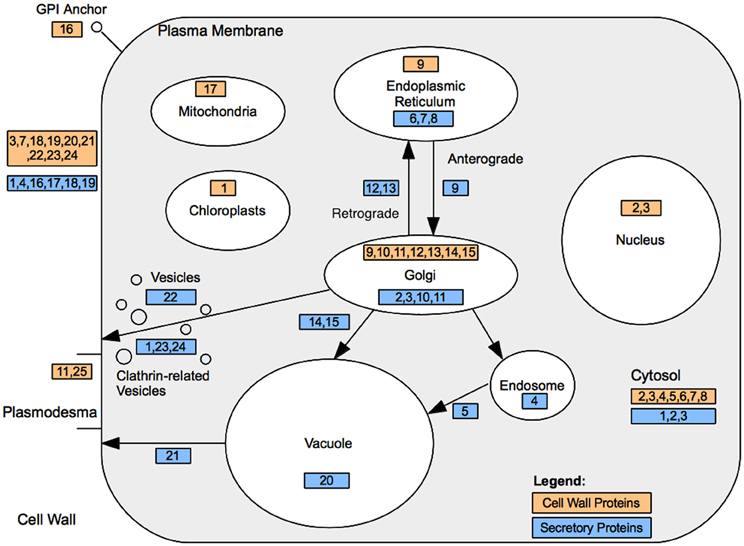
Figure 4. Cell wall and secretory protein localization. Cell wall associated protein locations are denoted by their ID within light orange shaded rectangles. Endomembrane/secretory pathway associated proteins are represented by their ID within light blue rectangles. Further details are presented in Tables 2 and 3.
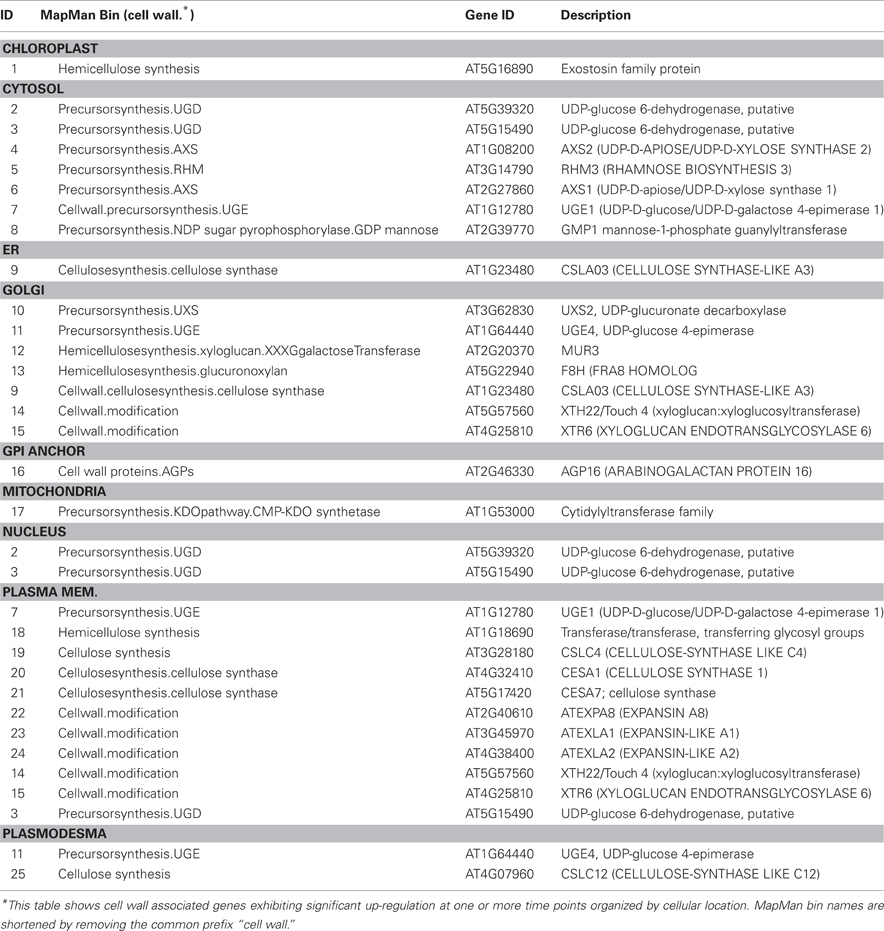
Table 2. Cell wall associated protein localization.
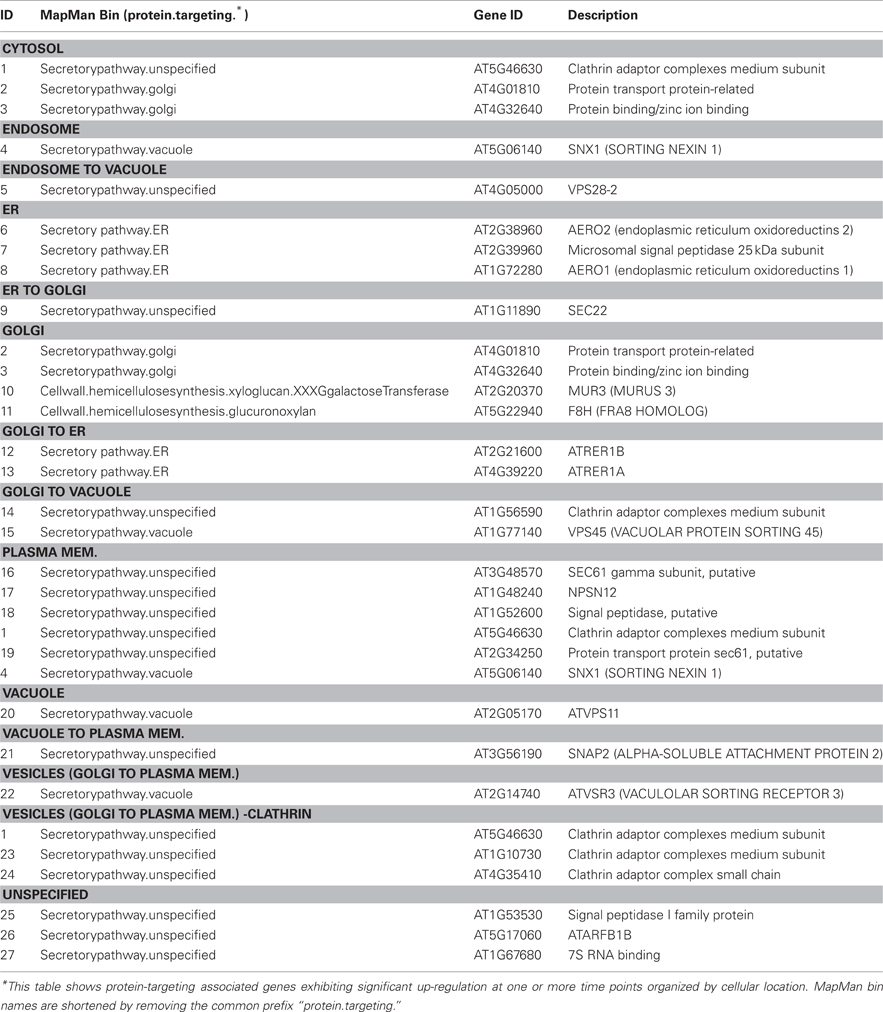
Table 3. Secretory protein localization.
Hormone-Related Responses
Ethylene synthesis and signaling appear to be active throughout the time course (Figure 5). In contrast, fewer ABA-related genes responded over the time course, including two members of the nine cis-epoxycarotenoid dioxygenase (NCED4 and NCED5), which were down regulated, although there is a suggestion that ABA-mediated signaling may have become active (Figure 6). MapMan analysis reflected this, with more responsive genes in the case of the ethylene bin in MapMan than there were for ABA. This is reflected in the up-regulation of five ethylene response factor (ERF) genes (ERF1, 3, 6, 12, and 15) and suggests an increase in ethylene levels up to TP3, followed by maintenance through TP4. In contrast, two genes encoding members of the 2OG-Fe(II) oxygenase gene family, enzymes that degrade ethylene, were down-regulated. Three other ethylene-associated genes that were down-regulated include HOOKLESS1, and two members of the multi-protein bridging factor (MBF) gene family, MBF1B and C.
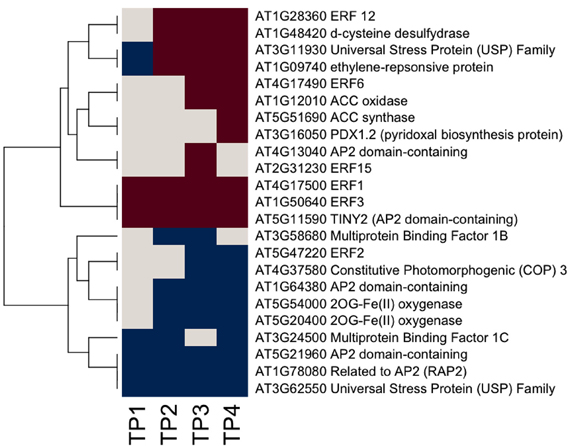
Figure 5. Ethylene-related gene responses across the time course of winter hardening. The gene expression levels across the rows are hierarchically clustered. Magenta cells represent significant positive expression changes, blue cells represent significant negative changes, and off-white cells represent no change or no statistically significant change.

Figure 6. ABA-related gene expression across the time course of winter hardening. The gene expression levels across the rows are hierarchically clustered. Magenta cells represent significant positive expression changes, blue cells represent significant negative changes, and off-white cells represent no change or no statistically significant change.
Genes Associated with Highly Responsive Ethylene Signaling, and Cell Wall Related Genes Later in Winter Hardening
Table 4 lists 13 genes, with their respective MapMan bins, that showed the largest up-regulation over the time course, especially at the later time points. This was true of the gene encoding GPI-anchored AGP16, which may have a role in signaling across the plasma membrane, the transcription factor ERF3 (ETHYLENE RESPONSIVE ELEMENT BINDING FACTOR 3), and TCH4. The ALPINE pipeline was used to visualize documented “guilt by association” relationships for ERF3, reasoning that this strategy might reveal clues concerning major, later, ethylene-related events in winter hardening that are not apparent using our other strategies. Using ERF3 as the input gene, mining gene expression data obtained for TP3 and 4 resulted in the identification of 11 positively expressed genes related to the input gene through “guilt by association.” Figure 7 shows the result, which is based on co-expression, protein–protein interactions, and genetic interactions in the GeneMANIA database only. Table 5 provides a more detailed notes on the genes highlighted in Figure 7. Several genes encoding proteins located in the endomembrane system appeared among the output genes, together with a splicing factor and an RNA-binding protein.
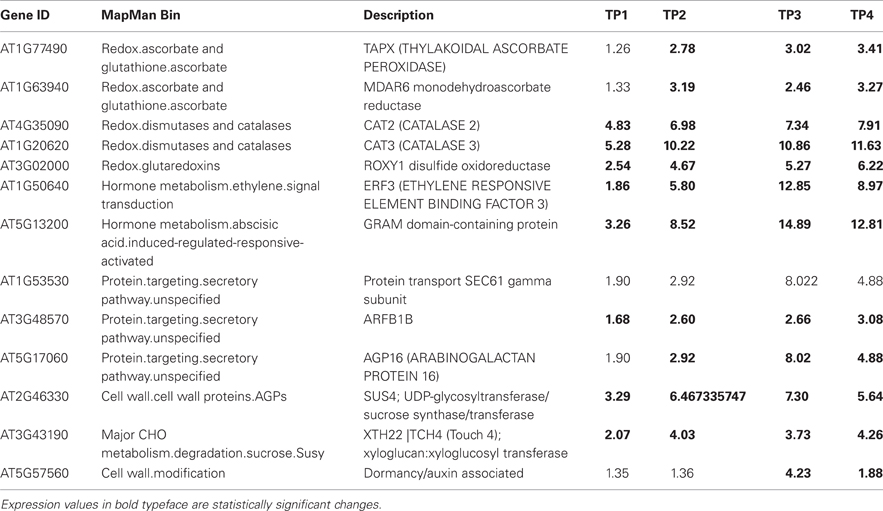
Table 4. Genes with largest up-regulation over the time course of winter hardening.
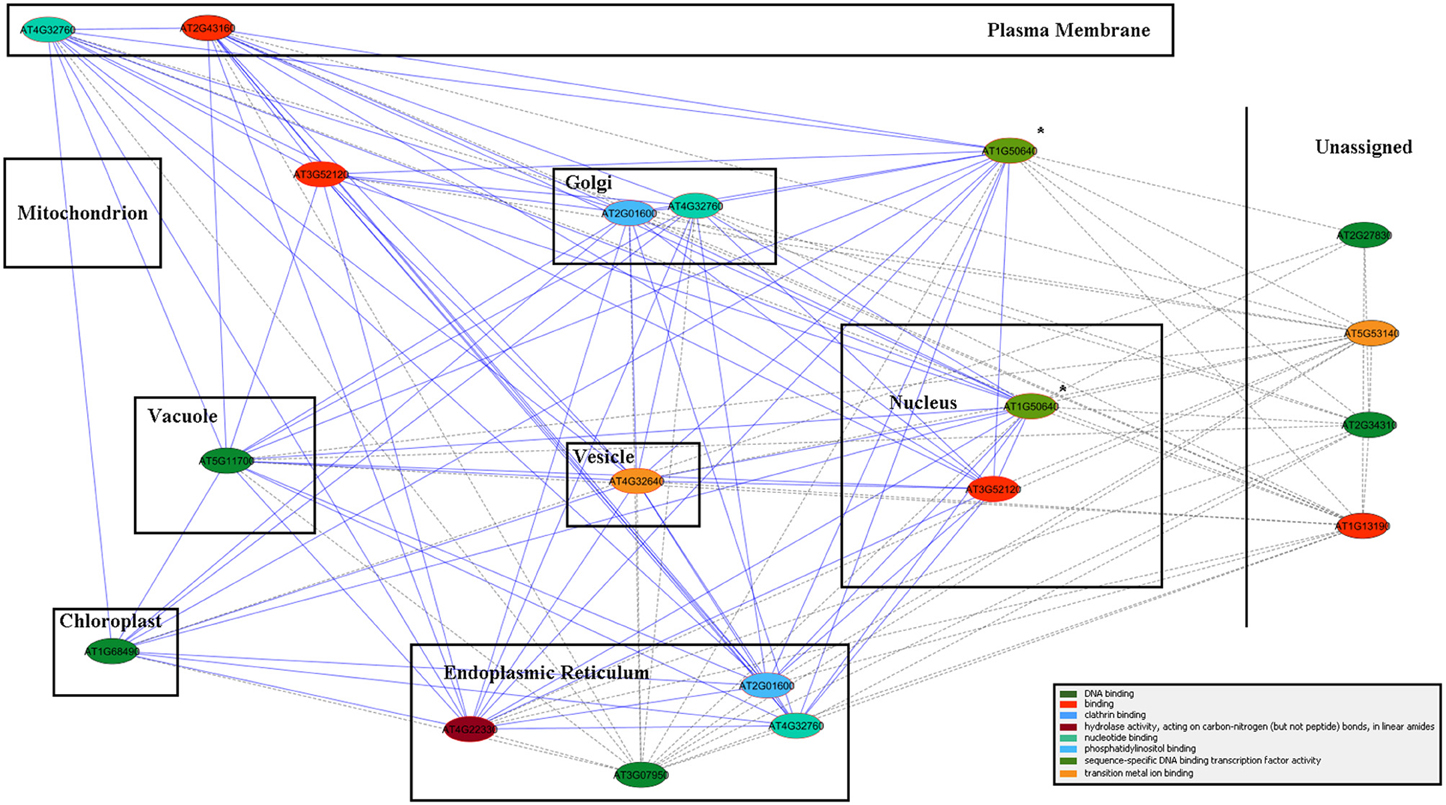
Figure 7. ALPINE gene association network results for ERF3 in a cellular component context. Edges represent “guilt by association” between the two connected genes, and node color represents GO molecular function annotation (the color legend is located in the bottom right corner). Due to a lack of specificity in the version of GO cellular component annotations provided by Mosaic, cellular component assignments, as provided directly by GO, were drawn in place of the default Mosaic background image. The query gene is indicated by the “*” symbol, and genes with multiple cellular component annotations appear multiple times in the network.
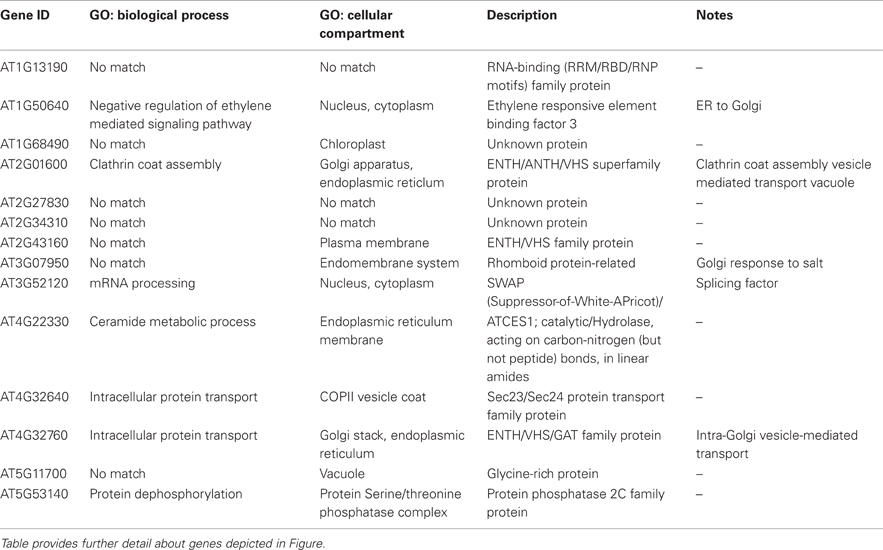
Table 5. Alpine gene association network details.
Discussion
With regard to several cell functions, our results show that Sitka spruce needles are surprisingly metabolically active during the fall and early winter, which reflects the need for fundamental remodeling that equips them to survive the stress imposed by freezing temperatures. Numerous cellular processes facilitate this, including the synthesis of cryoprotectant small molecules (di- and oligosaccharides, amino acids, etc.,) and proteins (dehydrins, antifreeze proteins, heat shock proteins). These cryoprotectants are thought to counterbalance the change in osmotic potential between the protoplast and apoplast with extracellular ice formation, to solubilize intra-cellular proteins as the cell dehydrates, and to lower the freezing point of water to prevent intracellular ice formation. However, little is known about the role of cell wall remodeling in this process. As the cell dehydrates and the membrane shrinks correspondingly, it stands to reason that the cell wall must react in a similar way. We previously reported overrepresentation of transcripts in this dataset annotated as involved in cellular transport and trafficking, and sought to better characterize the role that these processes may play in conifer cold acclimation. Although cold acclimation involves substantial metabolic remodeling, which must involve protein and metabolite targeting, little is known about the role of transport in cold acclimation. Studying transcripts in this category in detail led us to conclude that some of this transport activity supports cell wall remodeling, another overlooked area of freezing temperature tolerance. We hypothesized that changes in the cell wall may be required to sustain the severe cellular dehydration accompanying the winter period, and our analysis suggests a coordinated response to do this, from candidate hormone signaling components through synthesis and transport of cell wall substrates.
Cell Walls and Secretory Pathways
Gene expression patterns observed over the time course of hardening are consistent with cellulose biosynthesis, cell wall modification, increased trafficking of cell wall components to the apoplast, and a concomitant increase in associated signaling processes. It appears that processes associated with the synthesis of cell wall polysaccharides, including cellulose, were quite active over the hardening period. Increases in gene expression were observed for the AtCSL (“CSL”) super family, specifically those encoding proteins that do not catalyze actual cellulose synthesis, but encode glycan synthases that catalyze the synthesis of mannan oligomers (AtCSLA gene family), and members of the AtCSLC family that catalyze production of the glucan backbone of xyloglucans present in primary cell walls (Davis et al., 2010). Proteins encoded by members of these two gene families accumulate in the Golgi lumen, as do many other proteins associated with cell wall synthesis (Parsons et al., 2012). An expression increase was also observed for a member of the AtCSLG gene family (AtCSLG3), which has been reported to respond to low temperatures (Provart et al., 2003). In contrast, members of the AtCSLB and AtCSLE subfamilies were down-regulated over the time course, though no function is known for either of these genes as of yet. In addition, At4g32410 and At5g44030, members of the “true” CES family that responded over the time course of hardening, showed up-regulation. Overall, the evidence points to remodeling of the carbohydrate matrix in the cell wall, with a concomitant up-regulation of cellulose biosynthesis itself.
Expression of genes encoding secretory proteins (such as MUR3) show a similar pattern. Interestingly, the protein encoded by MUR3 also plays a role in overall endomembrane organization (Tamura et al., 2005), suggesting that this cell wall precursor bestows integrity on the very secretory pathway that apoplastic substrates follow to their destination. In the case of At1g10730, encoding the medium subunit of a clathrin adaptor, the assignment to either the cell membrane or the TGN was made on the basis of its annotation, since clathrin is known to be part of the recruiting process for vesicles derived from those two subcellular locations (Bassham et al., 2008; Parsons et al., 2012). Signaling processes that regulate cell membrane-cell wall transitions may also have been up-regulated. One of the GPI-anchored AGP genes, AGP16, shows an increase over the entire time course of hardening. Since the AGPs are thought to function as signaling molecules (Zhang et al., 2011) by interacting with proteins within membranes that exist in lipid rafts (Zhang et al., 2011), these different members of the AGP gene family may play roles in coordinating the changes in cell composition and function that accompany the hardening process. Direct evidence exists for a role as co-receptor at the cell membrane for AGP18, another GPI-anchored AGP (Seifert and Roberts, 2007). AGP16 itself was found to interact with the protein encoded by At2g26300 (α-subunit of the G-protein), a GTP-binding/GTPase/channel regulator/signal transducer (Klopffleisch et al., 2011).
Taken together, these data suggest that cell wall functional activity continues until TP4, with alterations in the cell wall matrix figuring prominently in these processes. Cell wall components are known to be synthesized and processed in the endomembrane system (Driouich et al., 2012), and it is interesting to note that many of the gene products for which increases in transcript levels were detected are targeted to the endomembrane system, suggesting substantial cold-induced cellular trafficking. Two notable exceptions are AXS1 and AXS2, which are both predicted to be localized to the nucleus and which show increases at TP3 and 4, respectively. It is possible that these genes “moonlight” as regulators of cell wall function or other processes that underlie the acquisition of cold hardiness.
Role of Phytohormone Signaling in Cell Wall Remodeling
Ethylene-related genes responded positively over the time course of winter hardening. Three genes annotated as related to ethylene biosynthesis and/or regulation of ethylene levels, ACS12, ACC oxidase, and D-cysteine desulfhydrase (D-CDES), were up-regulated. Interestingly, ethylene is known to participate in the flux control of UDP-D-galactose into cell wall polymers (Seifert et al., 2004), providing a causal link between the apparent increase in ethylene levels and the increases in cell wall-related activities over the time course of winter hardening. ACS12 is now known not to have ACS activity but, rather, to function as an aminotransferase (Yamagami et al., 2003). However, ACC oxidase is enzymatically active and is part of the ethylene biosynthesis pathway. D-CDES has been shown to act as a modulator of ethylene levels (McDonnell et al., 2009). In the case of ERF3, which was up-regulated throughout the time course, and substantially up-regulated by TP4, the encoded protein has been shown to interact with histone deacetylase1 and is thought to contribute to the modulation of stress signal transduction (Song and Galbraith, 2006). ERF12 is inferred by virtue of the presence of the same EAR domain present in ERF3 to act as a transcriptional repressor (Fujimoto et al., 2000; Ohta et al., 2001). ERF1 is an activator of gene expression through binding to GCC boxes (Fujimoto et al., 2000). ERF2 acts as a positive regulator of jasmonic acid (JA) signaling (McGrath et al., 2005), and its down-regulation over the time course of hardening suggests repression of that signaling pathway. There is nothing known about the functions of the universal stress proteins, and PDX 1.2, although a member of the pyridoxine biosynthesis family by virtue of its sequence, does not have enzymatic activity and may act as a regulatory protein (Leuendorf et al., 2010).
Ethylene-related genes that were down-regulated throughout the time course of winter hardening included HOOKLESS1, which is thought to act both as a mediator between auxin and ethylene signaling through an unknown mechanism (Chatfield and Raizada, 2008) and to negatively regulate sugar and auxin signaling (Ohto et al., 2006). Two ethylene-responsive members of the MBF gene family, MBF1B and C, were also down-regulated over the entire time course. To date, the evidence suggests that members of the MBF1 gene family are redundant and acting as repressors of ABA signaling (Arce et al., 2010) through ABR1, an ERF gene. ABR1 does not appear to have responded in the current study. Proteins encoded by the NCED gene family catalyze the key step in the ABA biosynthetic pathway, and numerous studies have demonstrated the induction of NCED genes by abiotic stress. NCED5 is known to be part of the ABA biosynthetic pathway, whereas the function of NCED4 has not been established (Nambara and Marion-Poll, 2005), although it was recently localized to plastoglobuli, within the chloroplast (Lundquist et al., 2012). The down-regulation of NCED5 over the time course of winter hardening makes it likely that ABA levels went down over the time course of the study. Despite this, one gene encoding an ABI5-like protein, a positive regulator of ABA signaling, was up-regulated. In addition, a gene annotated as a possibly ABA-responsive GRAM protein was also massively up-regulated, but there appears to be neither any direct information on its function, nor was it co-expressed with known genes, as displayed in GeneMANIA (data not shown).
This inferred decrease in ABA levels may seem surprising in light of the many reports of the induction of ABA biosynthesis when plants are exposed to cold and drought stress (Iuchi et al., 2001; Tan et al., 2003). However, a similar result was reported in a study comparing adaptive changes to mild drought in young Arabidopsis leaves undergoing cell proliferation and expansion (Skirycz et al., 2010; Verelst et al., 2010) with gene expression responses to the same stress in mature leaves. They found that the “classical” and expected ABA-mediated response occurred in the mature leaves only, whereas ethylene-mediated processes took place in the younger tissue, a result that resonates with the data reported here. In the adaptation study, the authors also observed a comparable up-regulation of genes encoding enzymes that catalyze the biosynthesis of polysaccharide primary cell wall components, in agreement with the data presented in this report. However, those authors reported a down-regulation of genes encoding members of the CES gene family, in contrast to the results reported here. In summary, in the case of ABA, the available data strongly suggest a down-regulation of biosynthesis over the time course of winter hardening, whereas ethylene biosynthesis may have been stimulated along with signaling processes, some related to cell wall modification set in motion by that phytohormone. ABA and ethylene are known to have antagonistic roles in the control of plant cell function (Cheng et al., 2009).
The results from ALPINE provide circumstantial evidence for a regulatory role for ERF3, involving vesicular trafficking in the endomembrane system and, by inference, stimulating the synthesis and transport of non-cellulosic cell wall components to their targeted destination across the plasma membrane to the apoplast.
Taken together, the transcriptomic and metabolomic data obtained from Sitka spruce needles over the time course of winter hardening suggest that the transition to a hardened state involves an adaptation process, with trafficking through the endomembrane system, cell wall remodeling, and signaling related to ethylene action, as prominent cellular activities. This adaptation mechanism in Sitka spruce appears to be quite distinct from what is currently understood concerning the responses of angiosperms to severe abiotic stress and may be similar to adaptation mechanisms that occur under more mild stress conditions.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This work was supported by the National Science Foundation iPlant Collaborative (DBI-0735191) and NSF ABI1062472.
References
Altschul, S. F., Madden, T. L., Schaffer, A. A., Zhang, J., Zhang, Z., Miller, W., et al. (1997). Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res. 25, 3389–3402.
Arce, D. P., Godoy, A. V., Tsuda, K., Yamazaki, K., Valle, E. M., Iglesias, M. J., et al. (2010). The analysis of an Arabidopsis triple knock-down mutant reveals functions for MBF1 genes under oxidative stress conditions. J. Plant Physiol. 167, 194–200.
Ashburner, M., Ball, C. A., Blake, J. A., Botstein, D., Butler, H., Cherry, J. M., et al. (2000). Gene ontology: tool for the unification of biology. The gene ontology consortium. Nat. Genet. 25, 25–29.
Bassham, D. C., Brandizzi, F., Otegui, M. S., and Sanderfoot, A. A. (2008). The secretory system of Arabidopsis. Arabidopsis Book 6, e0116.
Bauer, S., Grossmann, S., Vingron, M., and Robinson, P. N. (2008). Ontologizer 2.0–a multifunctional tool for GO term enrichment analysis and data exploration. Bioinformatics 24, 1650–1651.
Bohlenius, H., Huang, T., Charbonnel-Campaa, L., Brunner, A. M., Jansson, S., Strauss, S. H., et al. (2006). CO/FT regulatory module controls timing of flowering and seasonal growth cessation in trees. Science 312, 1040–1043.
Carpita, N. C. (2011). Progress in the biological synthesis of the plant cell wall: new ideas for improving biomass for bioenergy. Curr. Opin. Biotechnol. doi: 10.1016/j.copbio.2011.12.003. [Epub ahead of print].
Chatfield, S. P., and Raizada, M. N. (2008). Ethylene and shoot regeneration: hookless1 modulates de novo shoot organogenesis in Arabidopsis thaliana. Plant Cell Rep. 27, 655–666.
Cheng, W. H., Chiang, M. H., Hwang, S. G., and Lin, P. C. (2009). Antagonism between abscisic acid and ethylene in Arabidopsis acts in parallel with the reciprocal regulation of their metabolism and signaling pathways. Plant Mol. Biol. 71, 61–80.
Chevalier, L., Bernard, S., Ramdani, Y., Lamour, R., Bardor, M., Lerouge, P., et al. (2010). Subcompartment localization of the side chain xyloglucan-synthesizing enzymes within Golgi stacks of tobacco suspension-cultured cells. Plant J. 64, 977–989.
Dauwe, R., Holliday, J. A., Aitken, S. N., and Mansfield, S. D. (2012). Metabolic dynamics during autumn cold acclimation within and among populations of Sitka spruce (Picea sitchensis). New Phytol. 194, 192–205.
Davis, J., Brandizzi, F., Liepman, A. H., and Keegstra, K. (2010). Arabidopsis mannan synthase CSLA9 and glucan synthase CSLC4 have opposite orientations in the Golgi membrane. Plant J. 64, 1028–1037.
Driouich, A., Follet-Gueye, M. L., Bernard, S., Kousar, S., Chevalier, L., Vicre-Gibouin, M., et al. (2012). Golgi-mediated synthesis and secretion of matrix polysaccharides of the primary cell wall of higher plants. Front. Plant Sci. 3:79. doi: 10.3389/fpls.2012.00079
Druart, N., Johansson, A., Baba, K., Schrader, J., Sjodin, A., Bhalerao, R. R., et al. (2007). Environmental and hormonal regulation of the activity-dormancy cycle in the cambial meristem involves stage-specific modulation of transcriptional and metabolic networks. Plant J. 50, 557–573.
El-Kasmi, F., Pacher, T., Strompen, G., Stierhof, Y. D., Muller, L. M., Koncz, C., et al. (2011). Arabidopsis SNARE protein SEC22 is essential for gametophyte development and maintenance of Golgi-stack integrity. Plant J. 66, 268–279.
Fujimoto, S. Y., Ohta, M., Usui, A., Shinshi, H., and Ohme-Takagi, M. (2000). Arabidopsis ethylene-responsive element binding factors act as transcriptional activators or repressors of GCC box-mediated gene expression. Plant Cell 12, 393–404.
Galindo Gonzalez, L. M., El Kayal, W., Ju, C. J., Allen, C. C., King-Jones, S., and Cooke, J. E. (2012). Integrated transcriptomic and proteomic profiling of white spruce stems during the transition from active growth to dormancy. Plant Cell Environ. 35, 682–701.
Holliday, J. A., Ralph, S. G., White, R., Bohlmann, J., and Aitken, S. N. (2008). Global monitoring of autumn gene expression within and among phenotypically divergent populations of Sitka spruce (Picea sitchensis). New Phytol. 178, 103–122.
Huber, W., Von Heydebreck, A., Sultmann, H., Poustka, A., and Vingron, M. (2002). Variance stabilization applied to microarray data calibration and to the quantification of differential expression. Bioinformatics 18, S96–S104.
Iuchi, S., Kobayashi, M., Taji, T., Naramoto, M., Seki, M., Kato, T., et al. (2001). Regulation of drought tolerance by gene manipulation of 9-cis-epoxycarotenoid dioxygenase, a key enzyme in abscisic acid biosynthesis in Arabidopsis. Plant J. 27, 325–333.
Joosen, R. V., Lammers, M., Balk, P. A., Bronnum, P., Konings, M. C., Perks, M., et al. (2006). Correlating gene expression to physiological parameters and environmental conditions during cold acclimation of Pinus sylvestris, identification of molecular markers using cDNA microarrays. Tree Physiol. 26, 1297–1313.
Klopffleisch, K., Phan, N., Augustin, K., Bayne, R. S., Booker, K. S., Botella, J. R., et al. (2011). Arabidopsis G-protein interactome reveals connections to cell wall carbohydrates and morphogenesis. Mol. Syst. Biol. 7, 532.
Lamesch, P., Berardini, T. Z., Li, D., Swarbreck, D., Wilks, C., Sasidharan, R., et al. (2012). The Arabidopsis Information Resource (TAIR): improved gene annotation and new tools. Nucleic Acids Res. 40, D1202–D1210.
Lee, C., Teng, Q., Huang, W., Zhong, R., and Ye, Z. H. (2009). The F8H glycosyltransferase is a functional paralog of FRA8 involved in glucuronoxylan biosynthesis in Arabidopsis. Plant Cell Physiol. 50, 812–827.
Leuendorf, J. E., Osorio, S., Szewczyk, A., Fernie, A. R., and Hellmann, H. (2010). Complex assembly and metabolic profiling of Arabidopsis thaliana plants overexpressing vitamin B(6) biosynthesis proteins. Mol. Plant 3, 890–903.
Lundquist, P. K., Poliakov, A., Bhuiyan, N. H., Zybailov, B., Sun, Q., and Van Wijk, K. J. (2012). The functional network of the Arabidopsis plastoglobule proteome based on quantitative proteomics and genome-wide coexpression analysis. Plant Physiol. 158, 1172–1192.
McDonnell, L., Plett, J. M., Andersson-Gunneras, S., Kozela, C., Dugardeyn, J., Van Der Straeten, D., et al. (2009). Ethylene levels are regulated by a plant encoded 1-aminocyclopropane-1-carboxylic acid deaminase. Physiol. Plant 136, 94–109.
McGrath, K. C., Dombrecht, B., Manners, J. M., Schenk, P. M., Edgar, C. I., Maclean, D. J., et al. (2005). Repressor- and activator-type ethylene response factors functioning in jasmonate signaling and disease resistance identified via a genome-wide screen of Arabidopsis transcription factor gene expression. Plant Physiol. 139, 949–959.
Mohamed, R., Wang, C. T., Ma, C., Shevchenko, O., Dye, S. J., Puzey, J. R., et al. (2010). Populus CEN/TFL1 regulates first onset of flowering, axillary meristem identity and dormancy release in Populus. Plant J. 62, 674–688.
Montojo, J., Zuberi, K., Rodriguez, H., Kazi, F., Wright, G., Donaldson, S. L., et al. (2010). GeneMANIA Cytoscape plugin: fast gene function predictions on the desktop. Bioinformatics 26, 2927–2928.
Nambara, E., and Marion-Poll, A. (2005). Abscisic acid biosynthesis and catabolism. Annu. Rev. Plant Biol. 56, 165–185.
Ohta, M., Matsui, K., Hiratsu, K., Shinshi, H., and Ohme-Takagi, M. (2001). Repression domains of class II ERF transcriptional repressors share an essential motif for active repression. Plant Cell 13, 1959–1968.
Ohto, M. A., Hayashi, S., Sawa, S., Hashimoto-Ohta, A., and Nakamura, K. (2006). Involvement of HLS1 in sugar and auxin signaling in Arabidopsis leaves. Plant Cell Physiol. 47, 1603–1611.
Oikawa, A., Joshi, H. J., Rennie, E. A., Ebert, B., Manisseri, C., Heazlewood, J. L., et al. (2010). An integrative approach to the identification of Arabidopsis and rice genes involved in xylan and secondary wall development. PLoS ONE 5:e15481. doi: 10.1371/journal.pone.0015481
Parsons, H. T., Christiansen, K., Knierim, B., Carroll, A., Ito, J., Batth, T. S., et al. (2012). Isolation and proteomic characterization of the Arabidopsis Golgi defines functional and novel components involved in plant cell wall biosynthesis. Plant Physiol. 159, 12–26.
Provart, N. J., Gil, P., Chen, W., Han, B., Chang, H. S., Wang, X., et al. (2003). Gene expression phenotypes of Arabidopsis associated with sensitivity to low temperatures. Plant Physiol. 132, 893–906.
Rohde, A., and Bhalerao, R. P. (2007). Plant dormancy in the perennial context. Trends Plant Sci. 12, 217–223.
Ruonala, R., Rinne, P. L., Kangasjarvi, J., and Van Der Schoot, C. (2008). CENL1 expression in the rib meristem affects stem elongation and the transition to dormancy in Populus. Plant Cell 20, 59–74.
Ruttink, T., Arend, M., Morreel, K., Storme, V., Rombauts, S., Fromm, J., et al. (2007). A molecular timetable for apical bud formation and dormancy induction in poplar. Plant Cell 19, 2370–2390.
Saraiya, P., Lee, P., and North, C. (2005). “Visualization of graphs with associated timeseries data,” in IEEE Symposium on Information Visualization (INFOVIS 2005). (Minneapolis, MN, USA), 225–232; 23–25 October 2005.
Seifert, G. J., Barber, C., Wells, B., and Roberts, K. (2004). Growth regulators and the control of nucleotide sugar flux. Plant Cell 16, 723–730.
Seifert, G. J., and Roberts, K. (2007). The biology of arabinogalactan proteins. Annu. Rev. Plant Biol. 58, 137–161.
Shannon, P., Markiel, A., Ozier, O., Baliga, N. S., Wang, J. T., Ramage, D., et al. (2003). Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res. 13, 2498–2504.
Skirycz, A., De Bodt, S., Obata, T., De Clercq, I., Claeys, H., De Rycke, R., et al. (2010). Developmental stage specificity and the role of mitochondrial metabolism in the response of Arabidopsis leaves to prolonged mild osmotic stress. Plant Physiol. 152, 226–244.
Smallwood, M., and Bowles, D. J. (2002). Plants in a cold climate. Philos. Trans. R. Soc. Lond. B Biol. Sci. 357, 831–847.
Song, C. P., and Galbraith, D. W. (2006). AtSAP18, an orthologue of human SAP18, is involved in the regulation of salt stress and mediates transcriptional repression in Arabidopsis. Plant Mol. Biol. 60, 241–257.
Storey, J. D., and Tibshirani, R. (2003). Statistical significance for genomewide studies. Proc. Natl. Acad. Sci. U.S.A. 100, 9440–9445.
Tamura, K., Shimada, T., Kondo, M., Nishimura, M., and Hara-Nishimura, I. (2005). KATAMARI1/MURUS3 Is a novel golgi membrane protein that is required for endomembrane organization in Arabidopsis. Plant Cell 17, 1764–1776.
Tan, B. C., Joseph, L. M., Deng, W. T., Liu, L., Li, Q. B., Cline, K., et al. (2003). Molecular characterization of the Arabidopsis 9-cis epoxycarotenoid dioxygenase gene family. Plant J. 35, 44–56.
Tufte, E. (1983). The Visual Display of Quantitative Information, ASIN: 096139210X. Cheshire, CT: Graphics Press.
Uemura, M., Tominaga, Y., Nakagawara, C., Shigematsu, S., Minami, A., and Kawamura, Y. (2006). Responses of the plasma membrane to low temperatures. Physiol. Plantarum 126, 81–89.
Usadel, B., Poree, F., Nagel, A., Lohse, M., Czedik-Eysenberg, A., and Stitt, M. (2009). A guide to using MapMan to visualize and compare Omics data in plants: a case study in the crop species, Maize. Plant Cell Environ. 32, 1211–1229.
Verelst, W., Skirycz, A., and Inze, D. (2010). Abscisic acid, ethylene and gibberellic acid act at different developmental stages to instruct the adaptation of young leaves to stress. Plant Signal. Behav. 5, 473–475.
Weiser, C. J. (1970). Cold Resistance and Injury in Woody Plants: knowledge of hardy plant adaptations to freezing stress may help us to reduce winter damage. Science 169, 1269–1278.
Westfall, P. H., and Young, S. S. (1993). Resampling-Based Multiple Testing: Examples and Methods for P-Value Adjustment. New York, NY: Wiley.
Yamagami, T., Tsuchisaka, A., Yamada, K., Haddon, W. F., Harden, L. A., and Theologis, A. (2003). Biochemical diversity among the 1-amino-cyclopropane-1-carboxylate synthase isozymes encoded by the Arabidopsis gene family. J. Biol. Chem. 278, 49102–49112.
Zhang, C., Hanspers, K., Kuchinsky, A., Salomonis, N., Xu, D., and Pico, A. R. (2012). Mosaic: making biological sense of complex networks. Bioinformatics 28, 1943–1944.
Keywords: microarray, Sitka spruce, carbon metabolism, cell walls, adaptation mechanisms, visualization
Citation: Grene R, Klumas C, Suren H, Yang K, Collakova E, Myers E, Heath LS and Holliday JA (2012) Mining and visualization of microarray and metabolomic data reveal extensive cell wall remodeling during winter hardening in Sitka spruce (Picea sitchensis). Front. Plant Sci. 3:241. doi: 10.3389/fpls.2012.00241
Received: 11 July 2012; Accepted: 10 October 2012;
Published online: 29 October 2012.
Edited by:
Bjoern Usadel, RWTH Aachen University, GermanyReviewed by:
Xinguang Zhu, Chinese Academy of Sciences, ChinaMasanori Arita, University of Tokyo, Japan
Copyright © 2012 Grene, Klumas, Suren, Yang, Collakova, Myers, Heath and Holliday. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits use, distribution and reproduction in other forums, provided the original authors and source are credited and subject to any copyright notices concerning any third-party graphics etc.
*Correspondence: Ruth Grene, Department of Plant Pathology, Physiology, and Weed Science, Virginia Tech, Blacksburg, VA 24061, USA. e-mail:Z3JlbmVAdnQuZWR1