
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Plant Sci. , 06 July 2011
Sec. Plant Physiology
Volume 2 - 2011 | https://doi.org/10.3389/fpls.2011.00028
 Govindjee1,2* Dmitriy Shevela3
Govindjee1,2* Dmitriy Shevela3
Cyanobacteria, or the blue-green algae as they used to be called until 1974, are the oldest oxygenic photosynthesizers. We summarize here adventures with them since the early 1960s. This includes studies on light absorption by cyanobacteria, excitation energy transfer at room temperature down to liquid helium temperature, fluorescence (kinetics as well as spectra) and its relationship to photosynthesis, and afterglow (or thermoluminescence) from them. Further, we summarize experiments on their two-light reaction – two-pigment system, as well as the unique role of bicarbonate (hydrogen carbonate) on the electron-acceptor side of their photosystem II, PSII. This review, in addition, includes a discussion on the regulation of changes in phycobilins (mostly in PSII) and chlorophyll a (Chl a; mostly in photosystem I, PSI) under oscillating light, on the relationship of the slow fluorescence increase (the so-called S to M rise, especially in the presence of diuron) in minute time scale with the so-called state-changes, and on the possibility of limited oxygen evolution in mixotrophic PSI (minus) mutants, up to 30 min, in the presence of glucose. We end this review with a brief discussion on the position of cyanobacteria in the evolution of photosynthetic systems.
“The Sun energizes the plants with six tastes, which constitute food for all living beings on the Earth. Thus, the food that supports the lives of creatures is filled with solar energy and the Sun is the father of all living beings”.
(Mahabharata, 2.600 BC)
Stanier and Van Niel (1962) were the first to have the opinion that blue-green algae must be classified under bacteria. This led Roger Stanier to write a letter to Peter Raven in 1970 (November 5, 1970, National Archives of Canada, MG 31, accession J35, vol. 6) where he used the term “cyanobacteria” as a replacement for “blue-green algae.” By 1974, cyanobacteria became “official” since it was included as such in Bergey’s Manual of Bacteriology (Buchanan and Gibbons, 1974). It took several more years before the name “blue-green algae” was abandoned, in favor of cyanobacteria, in the photosynthesis literature (for historical issues, see Sapp, 2005a,b).
It was about 2.4 billion (Ga) years ago that oxygen began to accumulate in significant amounts on our Earth and oxygenic life became possible leading to the evolution of life, as we know it today (Falkowski, 2006; Kump, 2008). However, there is ample evidence that almost a billion years earlier, morphological forms of cyanobacteria were present in stromatolites (Allwood et al., 2006; Bosak et al., 2009; Figure 1). In all likelihood, oxygen produced by those cyanobacteria was sequestered in iron deposits, and thus not released into the atmosphere. There is also the possibility that some of the earliest cyanobacteria behaved like anoxygenic photosynthetic bacteria. A major difference between cyanobacteria and algae is that the former are prokaryotic (i.e., they lack membranes around their organelles) and that respiratory and photosynthetic complexes share a common membrane. Almost all oxygenic photosynthesizers, with the exception of Acaryochloris, a cyanobacterium, use chlorophyll (Chl) a (see an overview by Björn et al. (2009) on why Chl a was the chosen one). For evolutionary diversity of cyanobacteria, see Tomitani et al. (2006) and for images of various cyanobacteria, search for “cyanobacteria, images” using “Google.”
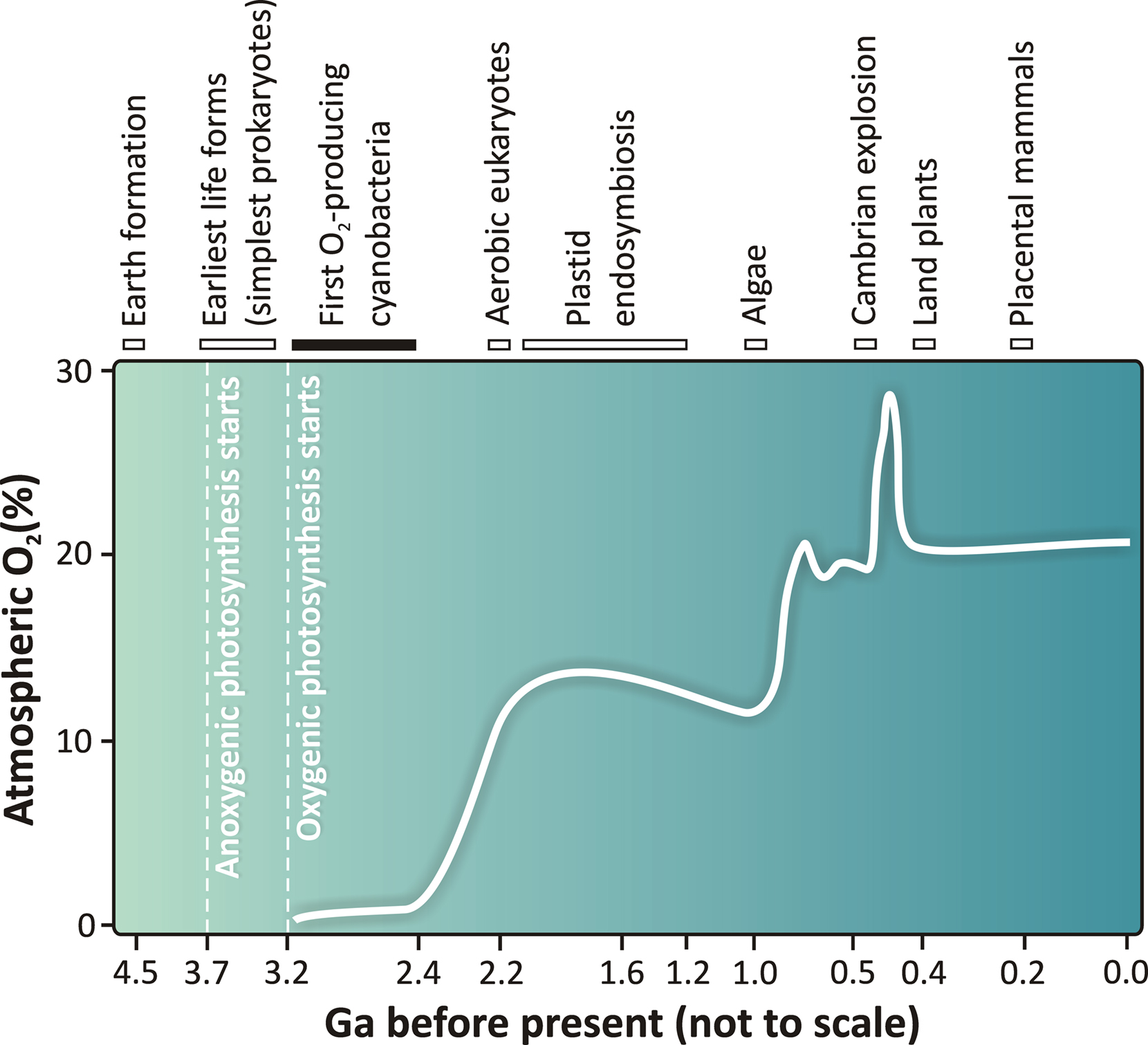
Figure 1. A schematic view of the changing concentration of atmospheric oxygen as a function of geological time in billions of years (Ga). Correlation between the estimated oxygen concentration changes and the major evolutionary events on Earth are based on data presented by Falkowski (2006; 2011), Tomitani et al. (2006), Kump (2008), Blankenship (2002; 2010), and Hohmann-Marriott and Blankenship (2011). The dates in the figure are not to scale and are approximations. Uncertainties of the selected events in the evolution of life are depicted with bars. A number of hypotheses exist as to when oxygenic photosynthesis was invented, by primitive cyanobacteria-like organisms. In this scheme, we have summarized currently available data and depict (by a black bar) that the first cyanobacteria could have evolved as early as 3.2 Ga or as late as 2.4 Ga ago. Therefore, an upper boundary of 3.2 Ga for the age of oxygenic photosynthesis and the initial rise of atmospheric oxygen in the atmosphere as indicated in this figure is still under discussion in the literature (see e.g., Tomitani et al., 2006; Allen and Martin, 2007; Blankenship et al., 2007; Buick, 2008). The figure is modified and adapted from Shevela and Messinger (2011).
Most cyanobacteria, in addition to some carotenoids, contain Chl a, phycocyanin and allophycocyanin as pigments for photosynthesis (Govindjee and Mohanty, 1972; Govindjee and Braun, 1974; Papageorgiou, 1996; Govindjee, 1999; Figure 2). Several cyanobacteria, in addition, contain phycoerythrin (Figure 2). Some (e.g., Prochlorococcus) contain Chl b, and Acaryochloris sp. are known to contain Chl d (Swingley et al., 2008). Further, some Chl a-containing cyanobacteria can be transformed to contain Chl b as well (Vavilin et al., 2003). This shows the great flexibility of cyanobacteria. Cyanobacteria have a unique position in life as they are the precursors of chloroplasts of algae, and, thus, of all plants (Björn and Govindjee, 2009). An interesting phenomenon, recently discovered, is that certain viruses carry either photosystem I (PSI) genes (Sharon et al., 2009) or photosystem II (PSII) genes (Mann et al., 2003; Lindell et al., 2005). This may have interesting implications for the past as well as future evolution. In addition, some differences between cyanobacteria and higher plants are also found on the water-oxidizing side (oxygen-evolving complex, OEC) of PSII. Thus, the water-splitting Mn4CaO5 cluster (Umena et al., 2011) is stabilized by three extrinsic proteins on the lumenal side of PSII. In the case of higher plants, these extrinsic proteins are PsbO (33 kDa), PsbP (23 kDa), and PsbQ (17 kDa) (Figure 3A); in cyanobacteria (Figure 3B), the two smaller proteins are substituted by cytochrome (Cyt) c550 (PsbV, 17 kDa) and PsbU (12 kDa) (Roose et al., 2007).
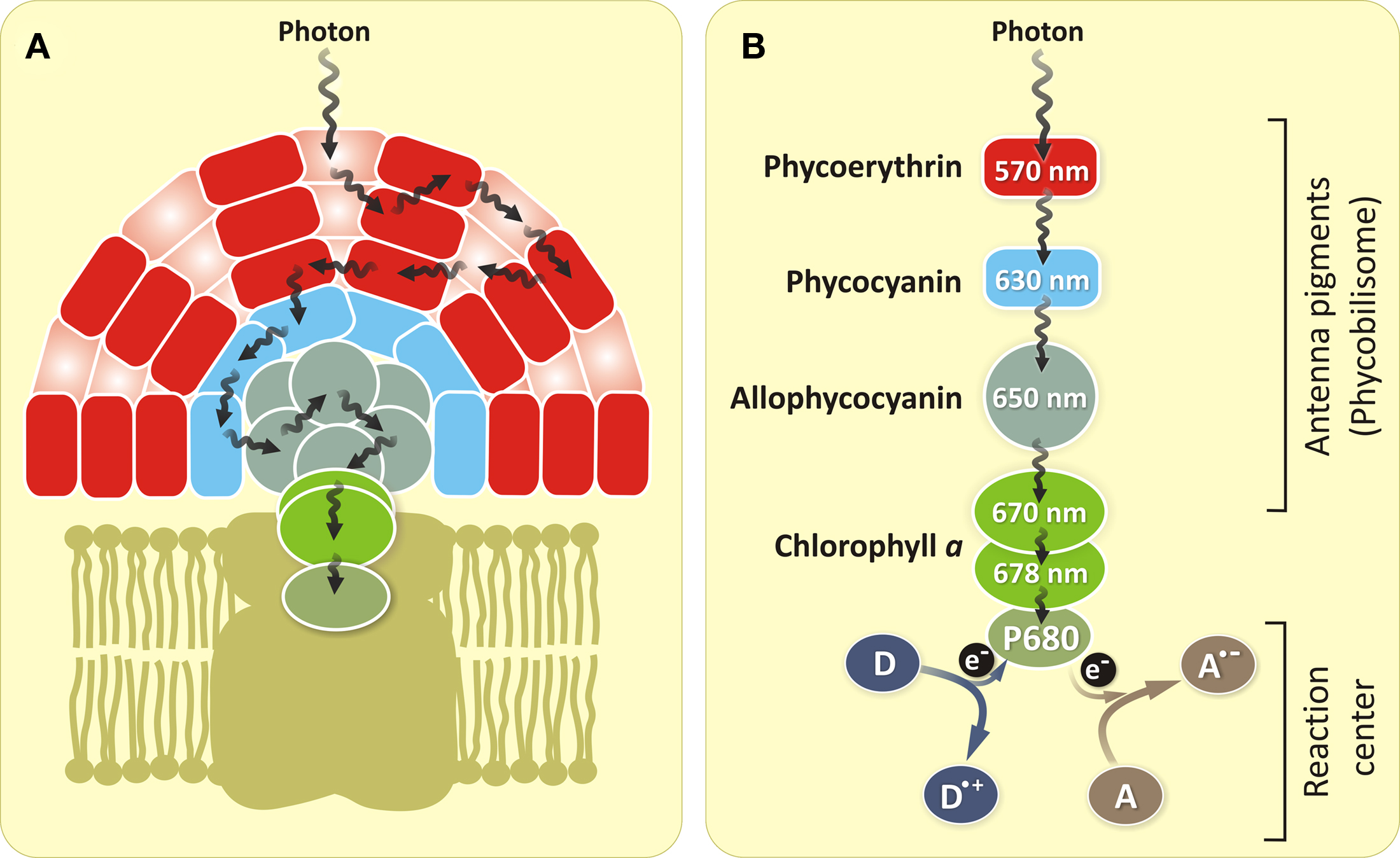
Figure 2. Structural organization of the antenna system of PSII for red algae and cyanobacteria (A) and energy transfer steps including charge separation (photochemical reaction) at the PSII RC (B) for cyanobacteria. The energy of absorbed photons is passed through a number of antenna molecules [phycoerythrin (absent in most cyanobacteria) → phycocyanin → allophycocyanin] until it reaches the RC Chl a (P680). The excited P680 donates its electron, which is in the excited state of the molecule, to an electron acceptor (A). The electron vacancy of the Chl a is filled by the electron from an electron donor (D). The wavelength numbers (nm) inside the circles represent pigments corresponding to the long wavelength absorption maxima of these pigments.
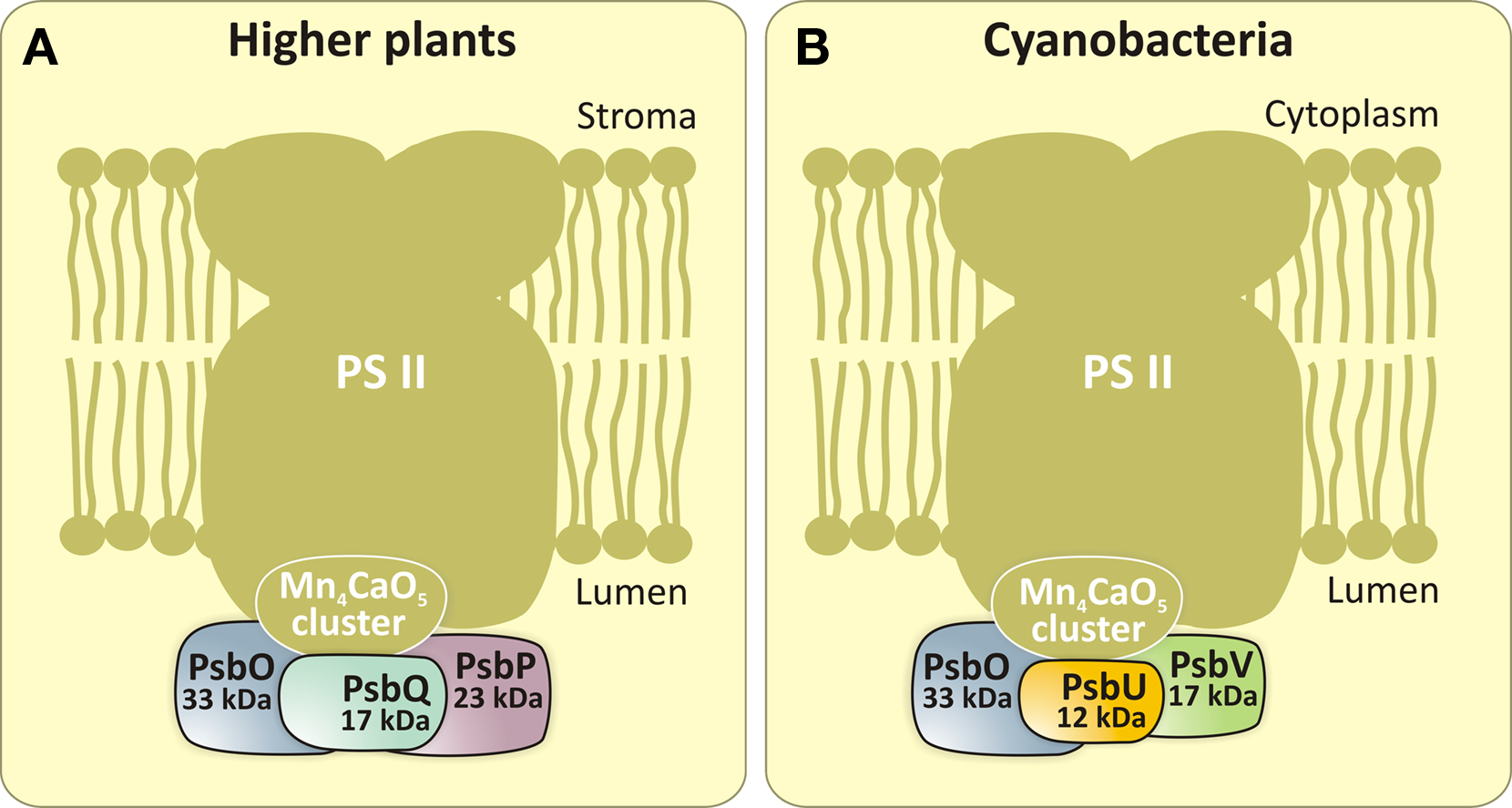
Figure 3. Extrinsic proteins on the lumenal side of PSII in higher plants (A) and cyanobacteria (B). PsbO (33 kDa), PsbQ (17 kDa) and PsbP (23 kDa) are the extrinsic proteins found in higher plants. In cyanobacteria, two of these proteins (PsbP and PsbQ) are replaced by PsbU (12 kDa) and PsbV (Cyt c550, 17 kDa). For a discussion of lipidated extrinsic proteins in cyanobacteria (CyanoP, CyanoQ, and Psb27), see recent review by Fagerlund and Eaton-Rye (2011). Both in higher plants and cyanobacteria, these proteins stabilize the Mn4CaO5 cluster and optimize its water-splitting reactivity.
As mentioned above, cyanobacteria are prokaryotes. Like photosynthetic eukaryotes (algae and plants), they transfer electrons, linearly, from water to NADP, utilizing PSII and PSI, and a Cyt b6f complex that sits in – between these two photosystems. The result is evolution of oxygen, reduction of NADP, and the formation of ATP. In addition, they also cycle electrons around PSI to drive extra ATP production (Bendall and Manasse, 1995; Van Thor et al., 2000).
Literature on cyanobacteria is extensive. We refer the readers to books edited by Desikachary (1972), Carr and Whitton (1982), Packer and Glazer (1988), Bryant (1994), Herrero et al. (2008), Gault and Marler (2009), and Peschek et al. (2011) and to several web sites:
• Wikipedia: http://en.wikipedia.org/wiki/Cyanobacteria;
• At Berkeley: http://www.ucmp.berkeley.edu/bacteria/cyanointro.html;
• At Purdue: http://www-cyanosite.bio.purdue.edu/;
• At Hawaii: http://www.botany.hawaii.edu/faculty/webb/bot311/cyanobacteria/cyanobacteria.htm;
• The following two sites have links to many other sites:
• http://tolweb.org/Cyanobacteria/2290;
• http://www.microbiologybytes.com/video/Cyanobacteria.html;
• Finally, there is also a site for harmful cyanobacteria, from the Center of Disease Control: http://www.cdc.gov/hab/cyanobacteria/facts.htm
For basic information on photosynthesis, we refer the readers to books by Rabinowitch and Govindjee(1969; available at http://www.life.illinois.edu/govindjee/photosynBook.html), Ke (2001), Blankenship (2002), and by Falkowski and Raven (2007). In addition, Scientific American articles are useful in understanding the basic steps (Rabinowitch and Govindjee, 1965; Govindjee and Govindjee, 1974; Govindjee and Coleman, 1990).
In this personal perspective, we will focus mainly on the adventures of one of us (G) that relate to his work on cyanobacteria with many graduate students, postdoctoral associates and researchers around the World. A perspective of this nature is essential to appreciate the Frontiers in Plant Physiology. We present here a summary of discoveries on specific cyanobacteria that will include new spectral forms of Chl a and P750 (in Anacystis (A.) nidulans), the two-light effect and the two-pigment system, excitation energy transfer that seems to follow Förster’s Resonance Energy Transfer theory (from phycobilins to Chl a), the first observation of fast fluorescence changes, the first demonstration of the role of bicarbonate on the electron-acceptor side of PSII, the relationship of slow fluorescence changes with changes that are unrelated to the quencher “QA,” but related to “conformational” changes and/or “state changes” (energy distribution between the two photosystems), and regulation of this energy distribution by various factors, and most recently, the observation of light-induced oxygen evolution in PSI-minus mutants in mixotrophic cultures and in the presence of glucose that demonstrates the flexibility of cyanobacteria.
Unlike anoxygenic photosynthetic bacteria (Blankenship et al., 1995; Hunter et al., 2009), cyanobacteria, like algae and plants, perform oxygenic photosynthesis (Mimuro et al., 2008). For that they use two-pigment systems and two-light reactions. Emerson and Chalmers (1958) discovered that, in A. nidulans, light absorbed in phycobilins enhanced the yield of oxygen evolution when added on top of light absorbed in Chl a. Thus, they proposed that in cyanobacteria, the “short-wave” photosystem is run by phycobilins and the “long-wave” photosystem is run by Chl a (also see Emerson and Rabinowitch, 1960). Kok (1959) showed that the long-wave (red light) system oxidized P700 (P700+; a special Chl a dimer; the reaction center, RC, of PSI), whereas the short-wave (orange light) system reduced P700+ and he related these results to Emerson’s 2-light effect. Govindjee and Rabinowitch (1960a) found that a small peak for Chl a was present in the same system that phycobilins were. This solved the dilemma how phycobilins could run the primary photochemical reactions themselves since Duysens (1952) had found that, in cyanobacteria, phycobilins transferred energy very efficiently to Chl a. Cederstrand et al. (1966) discovered, using an integrating dodecahedron, at least two spectral absorption bands in the Chl a region: Chl a 670 and Chl a 680: the idea of several different Chl a spectral absorption bands was already known, from derivative absorption spectroscopy (see Brown and French, 1961). In the green alga Chlorella and in the diatom Navicula, Chl a 670 belonged to the same system as to where Chl b (or Chl c) belonged (Govindjee and Rabinowitch, 1960b).
Amesz and Duysens (1962) provided experimental data on redox changes in the Cyt and NADP that supported the following picture of electron transfer path in cyanobacteria:
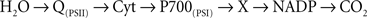
(Scheme 1)
Here, Q refers to a plastoquinone QA, P700 to RC of PSI, and X to ferredoxin.
For a historical perspective on the evolution of the Z-Scheme of electron transport in photosynthesis, see Govindjee and Björn (2011). In addition to the linear electron transport from water to NADP, there is also a cyclic electron transport around PSI. See Figure 4 for a current version of the Z-scheme of electron transport in cyanobacteria. Early measurements on ATP production in cyanobacteria, using the luciferin–luciferase assay, were made by Bedell and Govindjee (1973) in A. nidulans and by Lubberding and Schroten (1984) in Synechococcus sp.
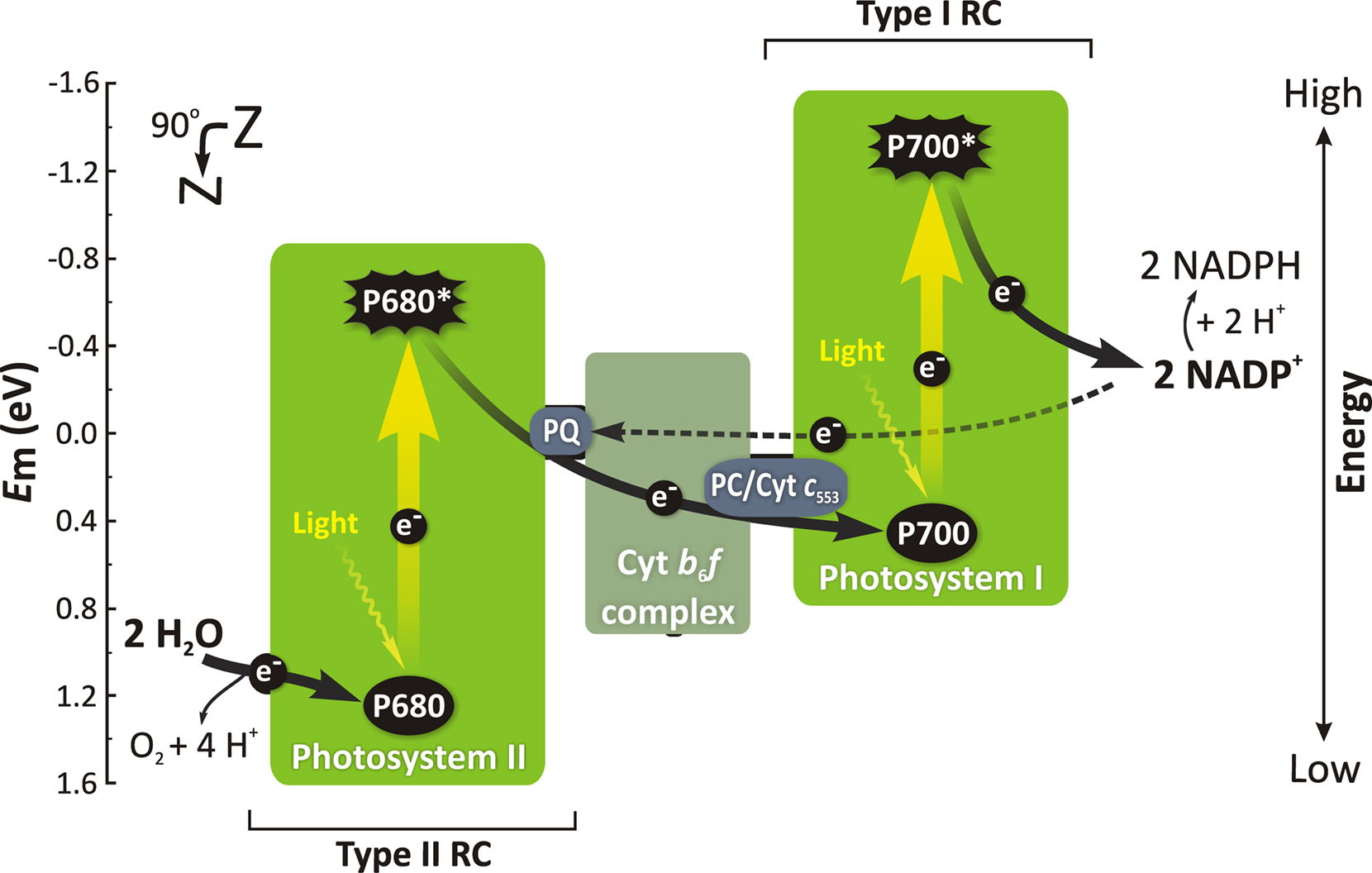
Figure 4. A simplified Z-scheme representing the linear (black solid arrows) and a cyclic (black dashed arrow) electron transport in cyanobacteria. Straight yellow arrows are for excitation of RC Chl a molecules that leads to electrons in the ground state to be raised into higher (singlet) excited state (P680 in PSII and P700 in PSI) in response to the absorption of light quanta (wavy arrows) via the antenna system (see Figure 2). Other abbreviations: Em (eV), redox potential at pH 7 (in electron volts); e−, electron; PQ, plastoquinone; Cyt b6f, cytochrome b6f complex; Cyt c553, cytochrome c553; PC, plastocyanin (for details on Cyt c553 and PC, see Zhang et al., 1992); the numbers 680 and 700 are the wavelengths of the absorption maxima, in the first excited state, for the Chl a RC molecules in PSII and PSI, respectively. Modified and adapted from Govindjee et al. (2010) and Shevela and Messinger (2011).
The flexibility of cyanobacteria was demonstrated recently (see an abstract by Govindjee et al., 2011; Q. J. Wang, A. Singh, H. Li, L. Nedbal, L. A. Sherman, J. Whitmarsh and Govindjee, unpublished) when cells of a PSI-minus mutant of mixotrophically grown Synechocystis sp. PCC 6803 were shown to evolve oxygen (∼25% of that of wild type) for 30 min in the presence of glucose. Studies are under way to find the mechanism for this observation. Perhaps, it involves uphill electron transfer using Δ pH made by PSII.
In addition to the Emerson enhancement effect, an unusual observation was that in several algae and in A. nidulans, far red light (720–760 nm range) caused inhibition (de-enhancement) of photosynthesis when added to short-wavelength light (Govindjee and Rabinowitch, 1960; Govindjee et al., 1960; Rabinowitch et al., 1960). Soon thereafter, Owens and Hoch (1963) using 18O-labeling and mass spectrometry showed that this de-enhancement was due to the effect of light on respiration (O2 uptake).
In search of pigments, which absorb in this wavelength region, Govindjee et al. (1961) discovered a new pigment, in A. nidulans, that had an absorption band at 750 nm (also, see Govindjee, 1963a; Figure 5). P750 had an emission band at 760 nm (Govindjee, 1963b) that was confirmed later by Goedheer and Hammans (1975). Unfortunately, no correlation of this band with photosynthesis was found (Gassner, 1962). This is an example of dead-end research as far as the laboratory of one of us (G) was concerned; however, Van Baalen (1965) found that it was involved in photo-oxidation of uric acid in A. nidulans; and Fischer and Metzner (1969) stated that P750 may be an open chain tetrapyrrole, but it not a bacteriopheophytin, and not a chlorin.
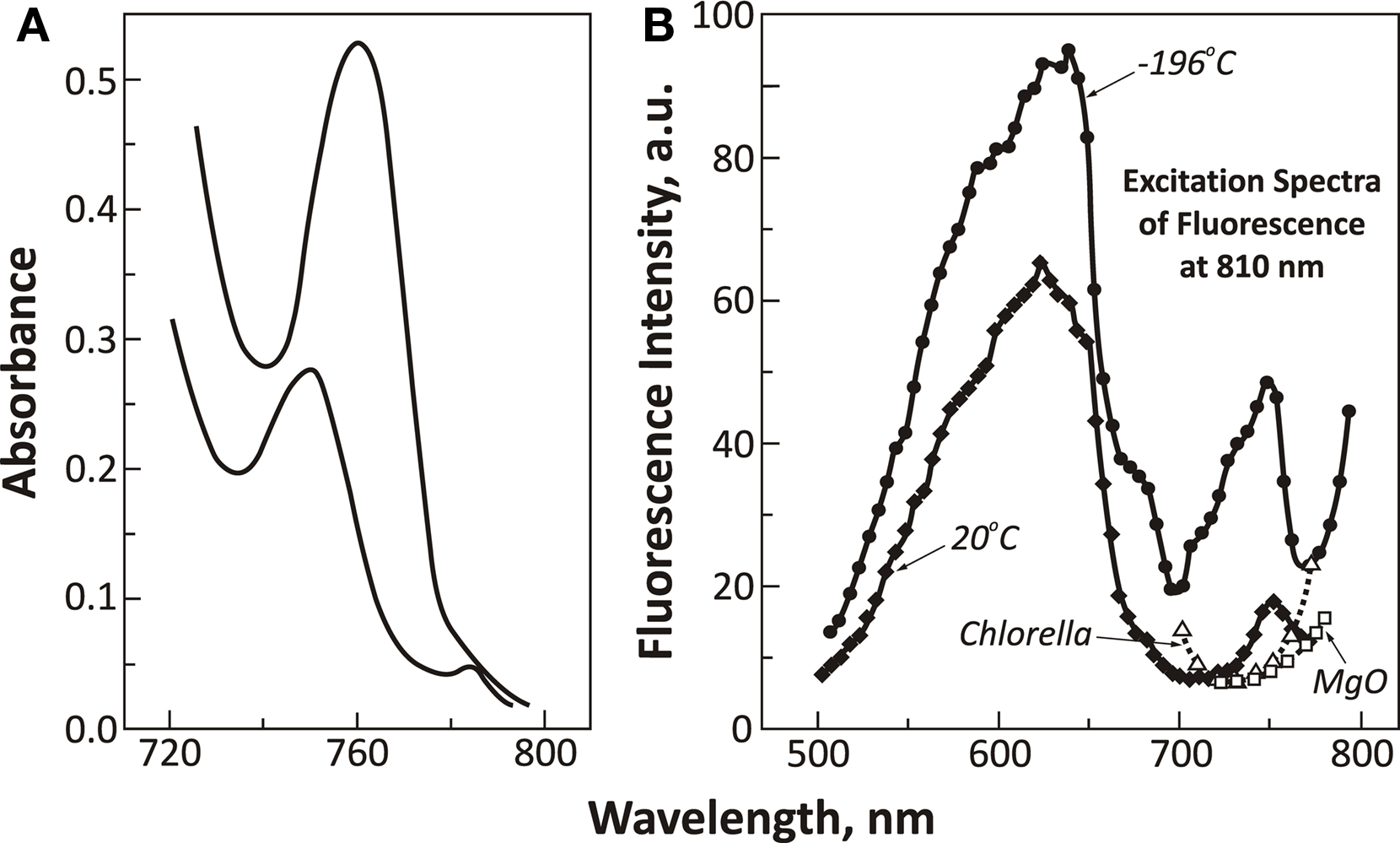
Figure 5. Spectroscopic data on P750 in Anacystis nidulans. (A) Absorbance spectra of two different concentrations of suspensions of A. nidulans cells showing the presence of a band in the 750–760 nm range (called P750; modified and reproduced from Govindjee et al., 1961). (B) Excitation spectra of fluorescence of P750 (measured at its red-most end, at 810 nm) of A. nidulans at 77 K (−196°C) and at 293 K (20°C). A portion of excitation spectrum of the green alga Chlorella is reproduced (open triangles) to show the absence of the excitation band at 750 nm. A curve for MgO is shown (open squares) to point out that the rising part of the curve beyond 760 nm is an artifact due to scattering (reproduced from Govindjee, 1963b).
Öquist (1974) found that iron-deficiency in A. nidulans increased fluorescence at 755 nm, perhaps, due to an increase in the concentration of P750. Goedheer and Hammans (1975) observed excitation energy transfer from several pigments to P750; and Hammans et al. (1977), however, established that P750 actually initiated oxygen uptake in A. nidulans. Further, Hammans (1978) showed that P750 sensitized photo-oxidation of several endogenous reductants in A. nidulans; however, these reductants did not serve as electron donors to PSI, nor were they oxidized in any Chl-sensitized reactions; there was, however, a hint that excitation of both P750 and Chl may cause the formation of singlet oxygen. Murata et al. (1981) showed that P750 was present in the cell envelope, not in the thylakoids, where there was also a small amount of pheophytin (Pheo)-like pigment with absorption maximum at 673 nm, and even some carotenoids. This observation explained why P750 had nothing to do with photosynthesis. Nultsch et al. (1983) found that P750 was absent in Anabaena and that it was not involved in phototaxis. Worcester et al. (1986) related an in vitro micellar system of aggregated Chls to P750; a correlation was suggested with the chlorosomes of the green bacteria. Later, Gombos et al. (1987) showed that P750, an aggregated form of Chl a, was formed during nitrogen starvation of cyanobacteria at ∼39°C, and that this process was reversible. Interestingly, however Shubin et al. (1991) found that, in Spirulina platensis, redox titration shows that F758 (fluorescence band at 758 nm) intensity has the same midpoint potential as P700; they concluded that the fluorescence quantum yield of F758 is proportional to the concentration of P700. Thus, in contrast, to other observations, they suggested a link of P750 to photosynthesis, at least in this organism. Further research is needed to understand fully the role of P750, i.e., where exactly it occurs in the cyanobacterial cells, what function or functions it performs, and possibly its importance in the evolutionary tree.
Chl a fluorescence (FL) and its relation to photosynthesis has been reviewed by many (see chapters in Govindjee et al., 1986, and in Papageorgiou and Govindjee, 2004). For historical perspectives on this subject, see Govindjee (1995) and Govindjee (2004). In order to fully appreciate this topic, we refer the readers to Wydrzynski and Satoh (2005) and Govindjee et al. (2010) to obtain a background on PSII since it is the system that has the most relationship with Chl a FL. For PSI, see Golbeck (2006).
A major use of FL has been in the use of detecting excitation energy transfer from one pigment (D for donor) to another (A for acceptor). If we excite D and all fluorescence appears from A, we know that there has been 100% energy transfer from D to A. However, if fluorescence appears from both D and A, we know that there has been a partial energy transfer (Duysens, 1952). This is the well-known “sensitized FL” method. Arnold and Oppenheimer (1950) were the first to show energy transfer in the cyanobacterium Chroococcus from phycocyanin to Chl a, and to provide a theory for excitation energy transfer. Duysens (1952) showed efficient energy transfer from phycobilins to Chl a in the cyanobacterium Oscillatoria. A powerful method to show energy migration among the same pigment is depolarization of FL, as was first used by Arnold and Meek (1956) for green algae. Mar and Govindjee (1972) showed that addition of 3-(3,4-dichlorophenyl)-1,1-dimethylurea (diuron; or DCMU), that blocks electron flow, led to increased depolarization of FL, thereby indicating increased energy migration in the antenna system. For a review on energy transfer in photosynthesis, see Goedheer (1972), Mimuro (2004), and Clegg et al. (2010). The general path of excitation energy transfer in most cyanobacteria is:
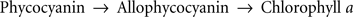
(Scheme 2)
A pathway of excitation energy transfer in cyanobacteria is shown in Figure 2B. Excitation energy migration and transfer, among the antenna molecules, is necessary for efficient photosynthesis since there are comparatively fewer RC molecules that are engaged in photosynthesis as compared to the number of antenna molecules that absorb the incoming light. Measurement of the action (or excitation) spectra of Chl FL is the method of choice to witness this phenomenon. By 1967, the following was known: (1) There is a “red drop” in FL yield, as was obtained through absorption spectra, and the application of the Stepanov relationship to A. nidulans (Szalay et al., 1967); data showed here that the red drop in FL began already around 640 nm in contrast to green algae where the red drop began at 685 nm. It confirmed the concept that in cyanobacteria most of Chl a is weakly fluorescent; (2) Spectral (FL) characteristics of various membrane fragments from Anacystis cells showed that Chl a of PSII was in a different fraction than Chl a of PSI (Shimony et al., 1967).
With this background, Govindjee’s group was ready to make measurements on excitation energy transfer at different temperatures. Cho and Govindjee (1970) measured both the excitation and emission spectra as a function of temperature in A. nidulans down to 4 K (liquid helium temperature; Figure 6). They made several observations: (i) large proportions of Chl a 670, Chl a 678 and even Chl a 705 are present in PSI, and that PSII contains mainly phycocyanin, allophycocyanin and only some Chl a 670 and Chl a 678; (ii) phycocyanin (with absorption bands at 580, 625, and 634–637 nm) and allophycocyanin (at 650 nm) are responsible for the broad fluorescence band at 653–655 nm; Chl a 670 is responsible for fluorescence band at 680 nm (F680), and Chl a 678 is responsible for fluorescence band at 685 nm (F685); (iii) the ratio of FL band at 715 nm (F715) to those at 685 and 695 nm (F685 and F695) is greater in PSI than in PSII; (iv) the efficiency of excitation energy transfer from Chl a 670 to Chl a 678, that approaches 100%, even at 4 K, is independent of temperature suggesting strong coupling between the Chl a-protein complexes; perhaps, here an “exciton” migration occurs as opposed to “hopping” of excitation energy as suggested for energy transfer from phycocyanin and allophycocyanin to Chl a; and (v) transfer efficiency from phycocyanin to Chl a is temperature dependent; it is lower at 4 K than at 77 K suggesting that here Förster resonance energy transfer may be taking place.
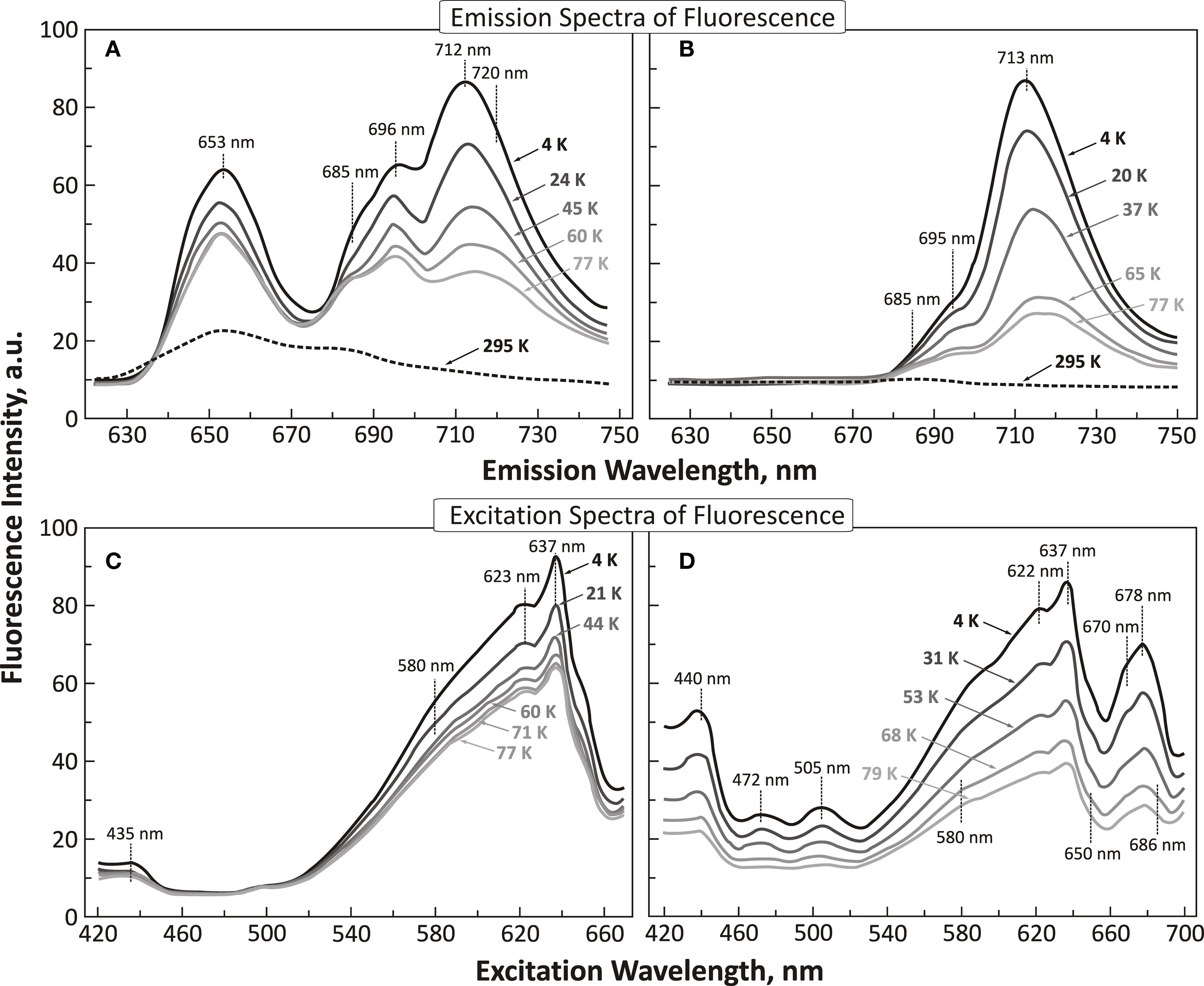
Figure 6. Emission and excitation fluorescence spectra of Anacystis nidulans at low temperatures. (A) Emission spectra of fluorescence in the 4 K (liquid helium temperature) to 77 K (liquid nitrogen temperature) range in cells of A. nidulans; wavelength of excitation was at 620 nm (absorption by phycocyanin; see Figure 2B). For comparison, a room temperature (295 K) spectrum (black dashed curve) is shown. (B) Emission spectra of fluorescence in the 4–77 K range in cells of A. nidulans; wavelength of excitation was at 440 nm (absorption by Chl a; note that cyanobacteria have most of their Chl a in PSI). For comparison, a room temperature (295 K) spectrum is also shown. (C) Excitation (or action) spectra of fluorescence, as a function of temperature (4–77 K) in cells of A. nidulans, measured at 685 nm (mostly PSII fluorescence). (D) Excitation (or action) spectra of fluorescence as a function of temperature (4–79 K) in cells of A. nidulans, measured at 715 nm (mostly PSI fluorescence). All panels of the figure were modified and reproduced from Cho and Govindjee (1970).
The above studies were followed by measurements on the lifetime of FL both at room temperature and at 77 K (Mar et al., 1972). In contrast to green alga Chlorella, Anacystis was calculated to have two times the rate constant of heat loss and had a lower efficiency of energy trapping than in algae. Further, the quantum yield of Chl a FL in Anacystis was about half of that in Chlorella. These observations are in agreement with what is known about photosynthesis measurements in these organisms. Further, both photosynthesis and Chl a FL are not static, but affected by dynamic changes in the environment. Nedbal et al. (2003) discovered a negative feedback regulation of the energetic coupling between phycobilisomes and PSII in the cyanobacterium Synechocystis sp. PCC 6803; they found that dynamic changes in the coupling of the phycobilisomes to PSII are not accompanied by corresponding antiparallel changes in the PSI excitation, suggesting a regulation limited to PSII. On the other hand, Kana et al. (2009) have shown changes in emission spectra, in Synechococcus sp., during the FL transient (for earlier observations in A. nidulans, see Papageorgiou and Govindjee, 1967, 1968). Further, heating of A. nidulans at 60°C for 10 min leads to several changes (Singhal et al., 1981): loss of absorption by phycobilins, and changes in excitation energy transfer from the leftover phycobilins to Chl a of PSI and or conversion of Chl a of PSI to Chl a of PSII.
Maksimov et al. (2011) have recently shown that the effective fluorescence cross section decreases from a value of ∼ 900 Å2 in the wild type Synechococcus sp. PCC 6803 to ∼200 Å2 (in phycocyanin-less mutant) or to ∼80 Å2 (in phycobilisome-less mutant). These mutants are expected to provide further information on excitation energy mechanisms in cyanobacteria.
We know that when green algae and higher plant leaves are illuminated with strong (high-intensity) light, Chl a FL rises from a minimum original low level (O, the origin) to a peak P within 1 s, and then declines to a semi-steady state S within about 10 s; this is followed by a rise to another maximum M and then a decline to a terminal steady state T within about a minute (see e.g., Govindjee and Papageorgiou, 1971; Papageorgiou, 1975; Papageorgiou et al., 2007). When this FL transient is plotted on a logarithmic time scale, two more inflections, called J and I, become obvious at about 2 and 30 ms, respectively (see Strasser et al., 1995; Strasser et al., 2004; Stirbet and Govindjee, 2011). The slow FL transient (minutes) in A. nidulans was first studied by Papageorgiou and Govindjee(1967, 1968), who showed that when phycobilins, but not Chl a, were excited, fluorescence rose from the S level to the M level, within a minute, in both normal and DCMU-treated A. nidulans cells (Figure 7). However, this change was several-fold higher in DCMU-treated cells; no change was observed at 77 K. Interestingly, carbonyl cyanide p-trifluoromethoxy phenylhydrazone (FCCP), which abolishes the pH difference across the thylakoids membrane, prevented the fluorescence increase only in normal, but not in DCMU-treated cells. Phlorizin, a phosphorylase inhibitor, had very little effect. These results suggested that these light-induced changes in Chl a FL yield may be related to the conformational changes which accompany photophosphorylation.
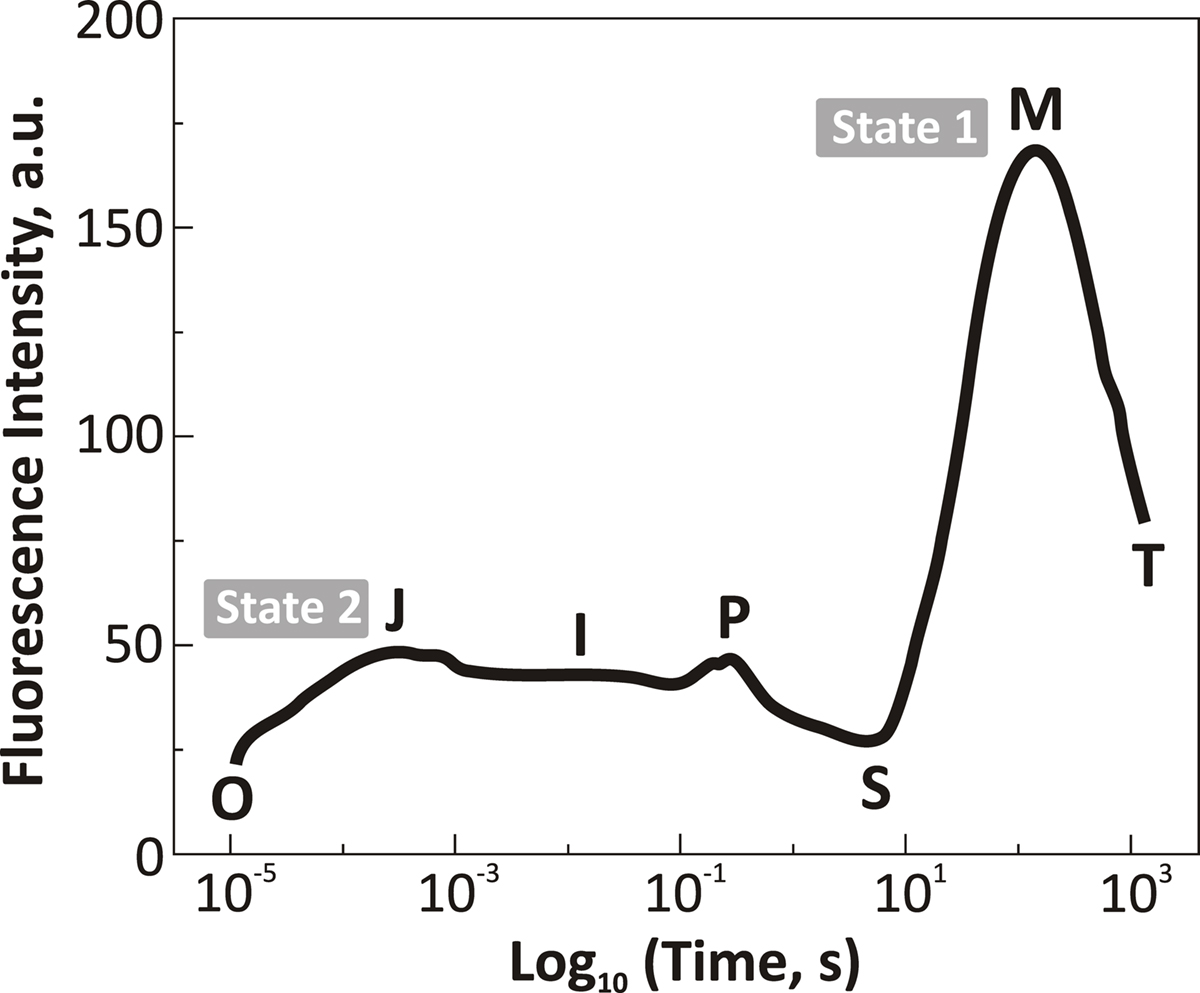
Figure 7. Chl a fluorescence transient of a cell suspension of dark-adapted Synechococcus sp. PCC 7942(Anacystis nidulans) plotted on a logarithmic time scale (from about a few μs to about 1000 s). Exciting light was 650 nm; 3000 μmol photons m-2 s-1. The figure was modified and reproduced from Tsimilli-Michael et al. (2009). Abbreviations: O, initial (minimum) step; J and I, intermediate levels; P, peak level; S, a semi-steady state level; M, a maximum level; and T, the terminal steady state level, not quite reached in this experiment (see Govindjee and Papageorgiou, 1971). This fluorescence transient (or fluorescence induction) is quite different from that observed in green algae and higher plants, where the “OPS” transient dominates. Cyanobacteria in darkness are usually in what is called “state 2” (low fluorescence state) and it takes a long time (almost 100 s) to be transferred to “state 1” (high fluorescence state). For a recent historical review on “state changes,” see Papageorgiou and Govindjee (2011).
Mohanty and Govindjee(1973a,b,c) were the first to study the fast, although of much reduced amplitude, FL transient in A. nidulans. They presented detailed studies with many uncouplers of phosphorylation, including salicylanilides, extending the concept that the slow FL changes were not related to “QA” changes (reviewed in Mohanty and Govindjee, 1974). Recently, using state transition-blocked mutants, Kana et al. (2010) have shown that a large part of the slow S to M rise, in Synechocystis sp. PCC 6803, is due to state 2 to state 1 transition since it is absent in RpaC (minus) mutant that is locked in state 1 (Emlyn-Jones et al., 1999).
When plants, algae, and cyanobacteria are illuminated and then quickly frozen to very low temperatures (e.g., 77 K) in darkness, and then warmed in dark, they emit light, and, this light emission is the afterglow, called thermoluminescence (TL). Arnold and Sherwood (1957) were the first to observe this phenomenon in dried chloroplasts. TL has been used extensively to monitor the water oxidation cycle (referred to as the S-state or the Kok cycle) as well as the redox status of the primary and secondary electron acceptors (QA and QB, respectively) of PSII since TL involves recombination reactions of the electron donor and acceptor sides of PSII. The so-called “B” bands of TL are due to recombination of S2/S3 states with reduced QB, whereas the “Q” band is due to recombination of the S2 state with reduced QA (Sane and Rutherford, 1986; Demeter and Govindjee, 1989; Vass and Govindjee, 1996; Vass, 2005). Excited Chl a (Chl*), which is needed to give light, is produced when electrons on reduced QA and QB move up by thermal energy, and positive charges on the S-states provide the “holes” to the ground state of Chl a, with which electrons can recombine. In their theory for TL, DeVault et al. (1983) and DeVault and Govindjee (1990) included temperature-dependent equilibrium between the various steps in the electron transport pathways and provided a generalized theory to explain the major TL bands in the photosynthetic systems. For TL bands, due to reduced QB, Rose et al. (2008) did show that the theory of DeVault and Govindjee (1990) is essential for explaining all the TL bands, although, for TL band due to reduced QA, simpler models may be sufficient (Rappaport and Lavergne, 2009).
Many investigators have observed TL from cyanobacteria; these have provided important information about their photosynthesis characteristics. Some examples are:
• Govindjee et al. (1985) observed that, in a thermophilic cyanobacterium Synechococcus vulcanus, TL bands were at higher temperatures (Figure 8): The B band was shifted from 25–30°C (in mesophilic organisms) to 50–55°C; and the Q (or D) band from 0–10°C to 35°C; further, the deactivation of the S2/S3 states was observed to slow down from 20–30 to 100–200 s range in this thermophilic cyanobacterium.
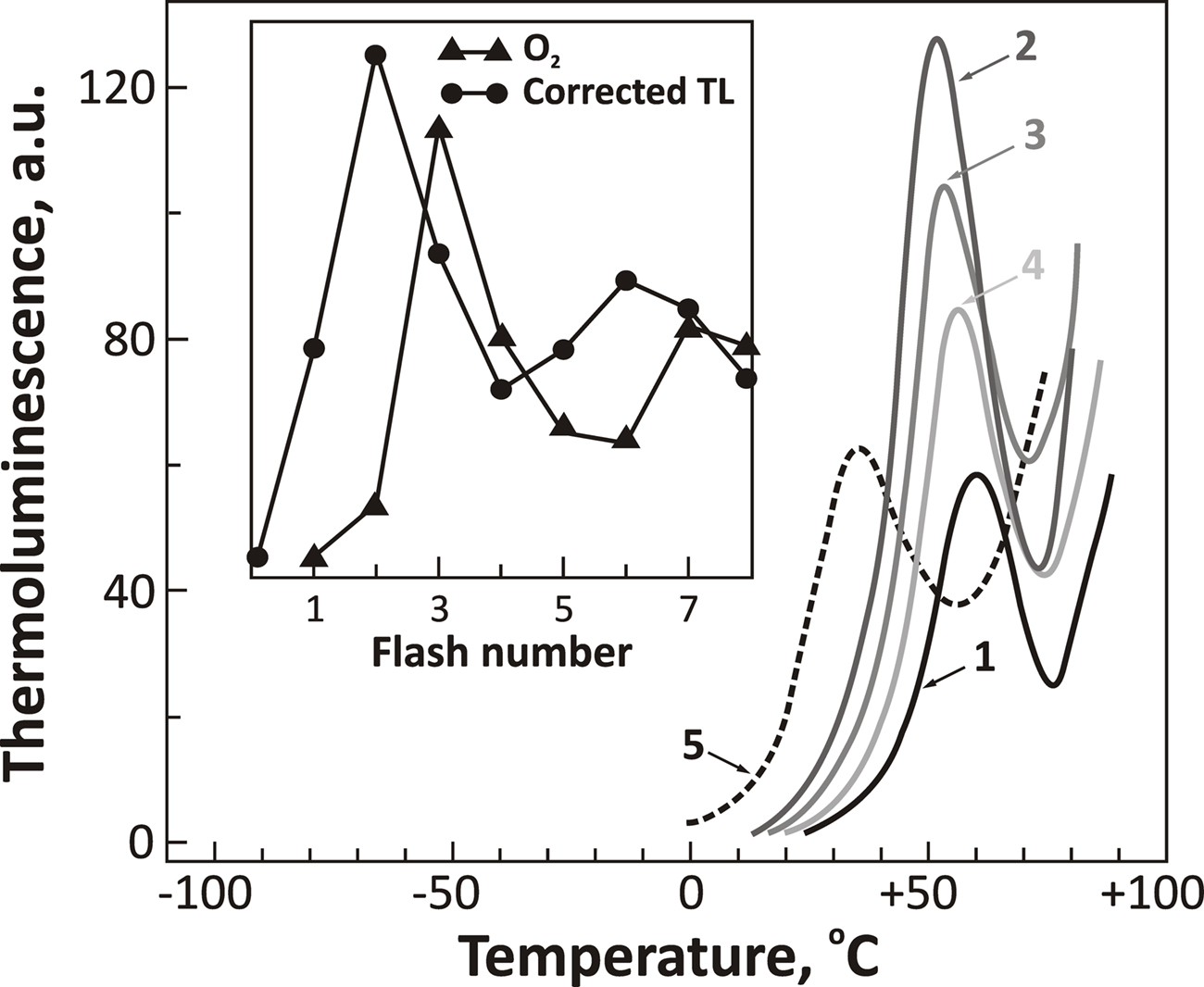
Figure 8. Thermoluminescence intensity from a thermophilic cyanobacterium Synechococcus vulcanus as a function of single-turn over actinic flashes at a flash frequency of 1 Hz (flash numbers correspond to curves 1, 2, 3, and 4, respectively). The dashed curve 5 is the spectrum obtained in the presence of DCMU that blocks electron transport from 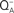 to QB. Insert shows flash number dependence of the 53°C TL band, the B band, due to S2/S3 and
to QB. Insert shows flash number dependence of the 53°C TL band, the B band, due to S2/S3 and 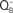 recombination (closed circles), and that of oxygen evolution (closed triangles). The “Q” band, due to S2 and
recombination (closed circles), and that of oxygen evolution (closed triangles). The “Q” band, due to S2 and 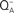 recombination (curve 5) was obtained at 35°C in this organism. The figure was modified and reproduced from Govindjee et al. (1985).
recombination (curve 5) was obtained at 35°C in this organism. The figure was modified and reproduced from Govindjee et al. (1985).
• Burnap et al. (1992) examined a Synechocystis sp. PCC 6803 mutant (Δ PsbO) that lacked the extrinsic manganese-stabilizing protein (MSP) and discovered, from TL emission peak temperatures, that 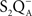 ,
, 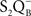 , and
, and 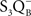 charge pairs were significantly altered and the intensities of the thermoluminescence emissions were significantly reduced in the mutant; thus, TL can be used as a tool to study changes on the donor side of PSII.
charge pairs were significantly altered and the intensities of the thermoluminescence emissions were significantly reduced in the mutant; thus, TL can be used as a tool to study changes on the donor side of PSII.
• Keranen et al. (1998) observed that both the B and the Q thermoluminescence bands, in Synechocystis sp. PCC 6803, are at the same temperatures in an autotrophic and a heterotrophic D1 protein mutant (that are missing part of the cytosolic loop (R225-F239 or R225-V249) between helices IV and V of the D1 protein) implicating a change in the redox potential of 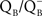 toward a more negative potential than in the wild type, where the B band(s) are at 25–35°C and the Q band at 0–10°C.
toward a more negative potential than in the wild type, where the B band(s) are at 25–35°C and the Q band at 0–10°C.
• Based on several measurements including TL, Meetam et al. (1999) concluded that the PsbY protein in Synechocystis sp. PCC 6803 is not essential for oxygenic photosynthesis and does not provide an important binding site for manganese in the OEC of PSII.
• In a mutant of S. platensis, where electron transport from plastoquinol (PQH2) to Cyt b6f was slowed, Ruengjitchatchawalya et al. (2005) suggested, from TL measurements, the presence of a reversed electron flow via PQ – a decrease in reoxidation of PQH2 seems to explain TL and other data. We note that a TL band at 45°C was present in this mutant, and, that this band was enhanced by trans-thylakoid pH and eliminated by an uncoupler FCCP. Further research is necessary to understand how cyanobacteria – like changes were converted to plant-like changes (see the section above).
• Cser and Vass (2007) made TL measurements on Synechocystis sp. PCC 6803 in which the redox properties of three key PSII components, Pheo, P680, and QA, were modified. A correlation between the redox potential of the primary charge pair (P680Pheo) and the temperature of the TL band was observed. Both the Q and the B TL bands, due to 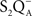 and
and 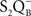 recombination, respectively, were at lower temperatures in the D1Gln130Glu or D1His198Ala mutants, which had their free energy of the primary radical pair shifted to more positive values. In contrast, the opposite was observed for D1Gln130Leu or D1His198Lys, which had free energy level of the primary radical pair shifted to more negative values. In addition, the first group of mutants (with downshifted TL bands) had much decreased TL intensities while the second group (with upshifted TL bands) had highly increased TL intensities.
recombination, respectively, were at lower temperatures in the D1Gln130Glu or D1His198Ala mutants, which had their free energy of the primary radical pair shifted to more positive values. In contrast, the opposite was observed for D1Gln130Leu or D1His198Lys, which had free energy level of the primary radical pair shifted to more negative values. In addition, the first group of mutants (with downshifted TL bands) had much decreased TL intensities while the second group (with upshifted TL bands) had highly increased TL intensities.
• From TL and other measurements, Cser et al. (2008) showed that in Acaryochloris marina, which contains predominantly Chl d, instead of Chl a, the charge recombination energetics is modified: the Em of QA and QB is increased, and the redox gap between RCIIChl* and RCIIChl+ Pheo− is decreased. Further, the free energy span between RCIIChl and RCIIChl* is decreased indicating the involvement of Chl d in primary electron donor activity. Interestingly, in spite of these differences in the energetics on the electron-acceptor side of PSII, the redox potentials and kinetics within the OEC of the Chl d-containing cyanobacterium A. marina closely resembles that of Chl a-containing cyanobacteria and higher plants (see Shevela et al., 2006 and references therein). Therefore, it appears, that the replacement of a large number of Chl a by Chl d in PSII of oxygen-evolving organisms has virtually no effect on the properties of the OEC. Although the redox potentials of QA and Pheo were found to be different in Chl d-containing A. marina and in Chl a-containing Synechocystis sp. PCC 6803, Allakhverdiev et al. (2011) found that the energetics in PSII is conserved, i.e., the difference in the potentials between QA and Pheo is the same.
Cyanobacteria, unlike other anoxygenic photosynthetic bacteria, require bicarbonate anion for their PSII activity. Bicarbonate 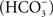 is a traditional term, but “hydrogen carbonate” is now recommended for it since IUPAC uses the latter; we can abbreviate it as HC. For reviews on the bicarbonate effect and PSII, see Govindjee and Van Rensen (1978), Stemler (1982), Vermaas and Govindjee (1982), Blubaugh and Govindjee (1988), Diner et al. (1991), Govindjee (1991, 1993), Govindjee and Van Rensen (1993), Govindjee et al. (1997), Van Rensen et al. (1999), Stemler (2002), Van Rensen and Klimov (2005), and McConnell et al. (2011). For the absence of bicarbonate effect in the photosystems of anoxygenic photosynthetic bacteria, see Shopes et al. (1989), and Wang et al. (1992).
is a traditional term, but “hydrogen carbonate” is now recommended for it since IUPAC uses the latter; we can abbreviate it as HC. For reviews on the bicarbonate effect and PSII, see Govindjee and Van Rensen (1978), Stemler (1982), Vermaas and Govindjee (1982), Blubaugh and Govindjee (1988), Diner et al. (1991), Govindjee (1991, 1993), Govindjee and Van Rensen (1993), Govindjee et al. (1997), Van Rensen et al. (1999), Stemler (2002), Van Rensen and Klimov (2005), and McConnell et al. (2011). For the absence of bicarbonate effect in the photosystems of anoxygenic photosynthetic bacteria, see Shopes et al. (1989), and Wang et al. (1992).
HC has been suggested to function both on the electron acceptor as well as on the electron donor side of PSII; there is no effect on PSI, however (Khanna et al., 1977; Eaton-Rye and Govindjee, 1984). Data on higher plant thylakoids suggest that the diffusing species is CO2 and the binding species is 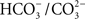 (Sarojini and Govindjee, 1981; Blubaugh and Govindjee, 1986). Using thylakoids, the HC effect has been located on the electron-acceptor side, close to the herbicide-binding side, particularly on the electron transport and protonation reactions at the secondary quinone QB and near the non-heme iron (Govindjee et al., 1976, 1984, 1991a, 1992; Jursinic et al., 1976; Khanna et al., 1980, 1981; Van Rensen and Vermaas, 1981; Eaton-Rye and Govindjee, 1988a,b). Several studies have established that the HC effect can be observed in intact cells and in leaves (Garab et al., 1988; El-Shintinawy and Govindjee, 1990; El-Shintinawy et al., 1990; Govindjee et al., 1993). To study the HC effect, very often formate (
(Sarojini and Govindjee, 1981; Blubaugh and Govindjee, 1986). Using thylakoids, the HC effect has been located on the electron-acceptor side, close to the herbicide-binding side, particularly on the electron transport and protonation reactions at the secondary quinone QB and near the non-heme iron (Govindjee et al., 1976, 1984, 1991a, 1992; Jursinic et al., 1976; Khanna et al., 1980, 1981; Van Rensen and Vermaas, 1981; Eaton-Rye and Govindjee, 1988a,b). Several studies have established that the HC effect can be observed in intact cells and in leaves (Garab et al., 1988; El-Shintinawy and Govindjee, 1990; El-Shintinawy et al., 1990; Govindjee et al., 1993). To study the HC effect, very often formate (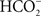 ) is used to obtain
) is used to obtain 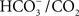 -free system to which
-free system to which 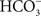 anions are added to measure the HC effect (Figure 9A). Nugent et al. (1988) showed clear effects of removal of HC (by formate addition) directly on the QA-Fe-QB signals in the PSII of the cyanobacterium Phormidium laminosum (Figure 9B). Both the ESR bands related to QA and QB (in the g = 1.8 and g = 1.6 regions) as well as to non-heme region (g = 6 region; shown on spinach samples; Figure 9C) were clearly affected upon the addition of formate (100 mM). The release of CO2 by formate addition was shown first by Govindjee et al. (1991b; 1997) using mass spectrometry and infrared spectroscopy; this has been recently confirmed by Shevela et al. (2008a,b).
anions are added to measure the HC effect (Figure 9A). Nugent et al. (1988) showed clear effects of removal of HC (by formate addition) directly on the QA-Fe-QB signals in the PSII of the cyanobacterium Phormidium laminosum (Figure 9B). Both the ESR bands related to QA and QB (in the g = 1.8 and g = 1.6 regions) as well as to non-heme region (g = 6 region; shown on spinach samples; Figure 9C) were clearly affected upon the addition of formate (100 mM). The release of CO2 by formate addition was shown first by Govindjee et al. (1991b; 1997) using mass spectrometry and infrared spectroscopy; this has been recently confirmed by Shevela et al. (2008a,b).
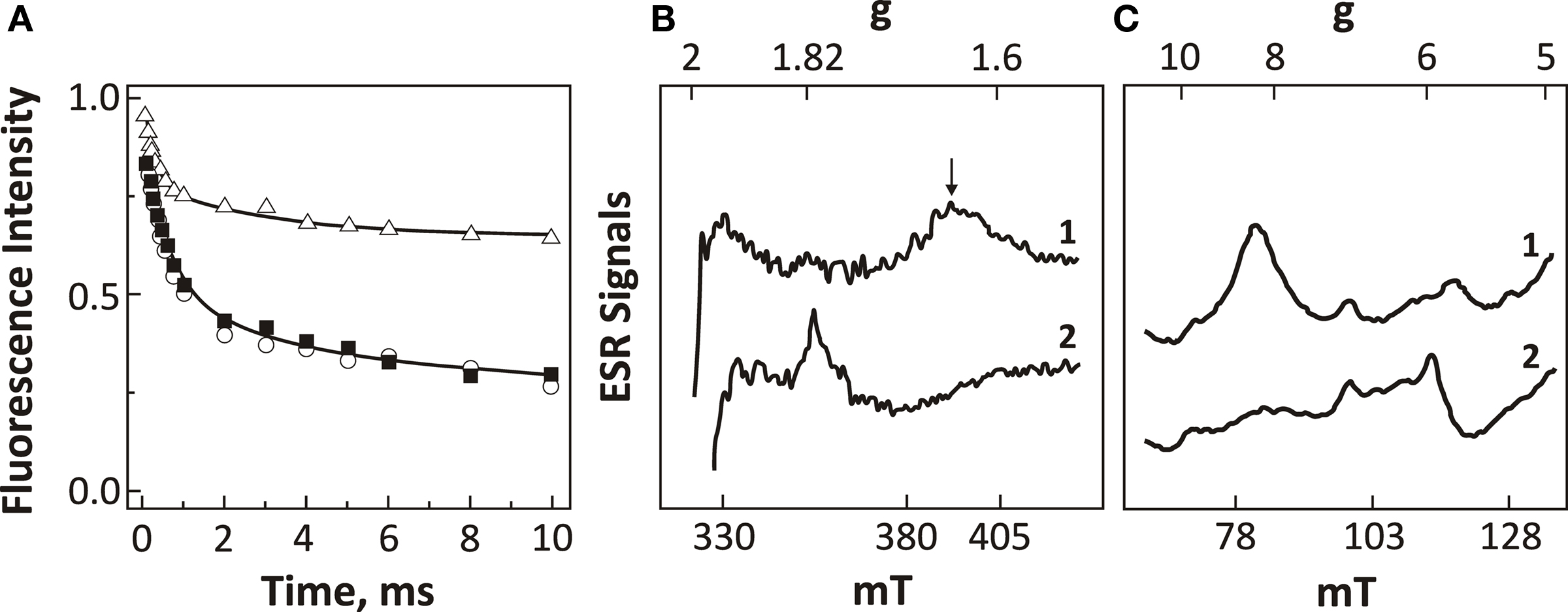
Figure 9. Fluorescence and EPR data representing the effects of HC removal on the electron-acceptor side of PSII. (A) Fluorescence measurements reflecting the decay of 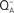 , upon illumination of thylakoids of Synechocystis 6803 after the third flash (repetitive rate of 1 Hz) at pH 7.5 (modified and reproduced from Cao and Govindjee, 1988). The fluorescence decay was measured in control (i.e., HC-containing) samples (closed squares), HC-depleted samples (open triangles), and HC-depleted samples after the re-addition of 2.5 mM
, upon illumination of thylakoids of Synechocystis 6803 after the third flash (repetitive rate of 1 Hz) at pH 7.5 (modified and reproduced from Cao and Govindjee, 1988). The fluorescence decay was measured in control (i.e., HC-containing) samples (closed squares), HC-depleted samples (open triangles), and HC-depleted samples after the re-addition of 2.5 mM 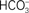 . (B) EPR spectra of the iron–semiquinone in the dark-adapted PSII samples from thermophilic cyanobacterium P. laminosum upon 5 min illumination at 77 K in the absence (spectrum 1) and the presence of 100 mM formate (spectrum 2). The used concentration of formate (100 mM) has been shown to be sufficient for the removal all HC bound to PSII (Govindjee et al., 1991b; Shevela et al., 2008b). (C) EPR spectra of the g = 6 non-heme iron region of the dark-adapted PSII samples from spinach at pH 7.5. The spectra were obtained in the absence (spectrum 1) and the presence of 100 mM formate (spectrum 2). For EPR settings, see Nugent et al. (1988). Abbreviations: g, is the electronic splitting factor (in case of free electron g-factor, it has a value of 2.0023); mT (millitesla), values for the magnetic field. (B,C) were modified and reproduced from Nugent et al. (1988).
. (B) EPR spectra of the iron–semiquinone in the dark-adapted PSII samples from thermophilic cyanobacterium P. laminosum upon 5 min illumination at 77 K in the absence (spectrum 1) and the presence of 100 mM formate (spectrum 2). The used concentration of formate (100 mM) has been shown to be sufficient for the removal all HC bound to PSII (Govindjee et al., 1991b; Shevela et al., 2008b). (C) EPR spectra of the g = 6 non-heme iron region of the dark-adapted PSII samples from spinach at pH 7.5. The spectra were obtained in the absence (spectrum 1) and the presence of 100 mM formate (spectrum 2). For EPR settings, see Nugent et al. (1988). Abbreviations: g, is the electronic splitting factor (in case of free electron g-factor, it has a value of 2.0023); mT (millitesla), values for the magnetic field. (B,C) were modified and reproduced from Nugent et al. (1988).
From the early work of Blubaugh and Govindjee (1988), we have known that there are at least two binding sites of HC. Thus, besides the site at non-heme iron between QA and QB, there has to be another 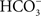 -binding site, which, however, was not seen by the recent crystallographic studies of PSII (Guskov et al., 2010; Umena et al., 2011). Most probably, at the second binding site, may be loosely bound to PSII, and may have escaped detection due possibly to its loss caused by X-ray irradiation as well as by the treatments required during the X-ray measurement.
-binding site, which, however, was not seen by the recent crystallographic studies of PSII (Guskov et al., 2010; Umena et al., 2011). Most probably, at the second binding site, may be loosely bound to PSII, and may have escaped detection due possibly to its loss caused by X-ray irradiation as well as by the treatments required during the X-ray measurement.
In view of a large body of experimental evidence for HC effect on the PSII acceptor side, and the latest PSII crystal structure at the resolution of 1.9 Å that shows the clear presence of HC on the non-heme iron between QA and QB in a thermophilic cyanobacterium Thermosynechococcus vulcanus (Umena et al., 2011), there is a renewed interest in understanding the mechanism of HC effect on the PSII acceptor side. There are clear evidences that HC is not a tightly bound constituent of the OEC of PSII (Aoyama et al., 2008; Shevela et al., 2008b; Ulas et al., 2008; Umena et al., 2011); however, there are numerous indications for the action of HC on the water-splitting side of PSII. For the HC effects on the donor side of PSII, see reviews by Stemler (2002) and Van Rensen and Klimov (2005).
Cao and Govindjee(1988, 1990) were the first to show the effect of HC on PSII in transformable Synechocystis sp. PCC 6803. Govindjee et al. (1993) confirmed this effect in other cyanobacterial cells. Through Chl a FL measurements, the effect was located on the electron-acceptor side of PSII. In view of the fact that in chloroplasts, HC effect was suggested to be near the herbicide-binding site, Govindjee et al. (1990) examined herbicide-resistant D1 mutants of Synechocystis 6714. Chl a FL and O2 evolution measurements revealed a differential sensitivity to 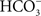 -reversible formate inhibition; resistance to formate treatment followed the order (highest to lowest): [double mutant] A251V/F211S (AZ V) > [single mutant] F211S (AZ 1) > wild type > [single mutant] S264A (DCMU II-A). These results are in agreement with the concept that there may be overlapping herbicide and bicarbonate niches in cyanobacteria as well.
-reversible formate inhibition; resistance to formate treatment followed the order (highest to lowest): [double mutant] A251V/F211S (AZ V) > [single mutant] F211S (AZ 1) > wild type > [single mutant] S264A (DCMU II-A). These results are in agreement with the concept that there may be overlapping herbicide and bicarbonate niches in cyanobacteria as well.
Soon thereafter, the concept arose that even D2 protein may be involved with the HC effect, particularly its specific arginines (Cao et al., 1991). Measurements of oxygen evolution showed that the D2 mutants R233Q (arginine-233 ∼ glutamine) and R251S (arginine-251 ∼ serine) were 10-fold more sensitive to formate than the wild type of Synechocystis sp. PCC 6803, suggesting their possible role in HC binding. However, further, studies on D1-S264 and D1-F255 mutants of Synechococcus sp. PCC 6942 led to the conclusion that the equilibrium dissociation constant for HC is increased in the mutants, while that of the formate remains unchanged. The hierarchy of the equilibrium dissociation constant for HC (highest to lowest, ±2 mM) was: D1-F255L/S264A (46 mM) > D1-F255Y/S264A (31 mM) – D1-S264A (34 mM) – D1-F255Y (33 mM) > wild type (25 mM). These data suggested the importance of D1-S264 and D1-F255 in the 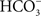 -binding niche. A possible involvement of HC and these two residues in the protonation of
-binding niche. A possible involvement of HC and these two residues in the protonation of 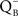 , the reduced secondary plastoquinone of PSII, in the D1 protein was discussed (Cao et al., 1992).
, the reduced secondary plastoquinone of PSII, in the D1 protein was discussed (Cao et al., 1992).
A picture began to emerge that we may not be dealing with specific binding site of HC that is involved in the reactions, but that conformation changes may be responsible for some of our observations in cyanobacteria. This became rather obvious when Vernotte et al. (1995) used several single and double mutants in the d–e interhelical loop of D1 protein in Synechocystis sp. PCC 6714. They showed a differential sensitivity of formate inhibition on the acceptor side of PSII in the following order: S264A-F255L > S264A = N266T > N266T-S264A > F255L = N266D = WT > > N266T-A251V = A251V. On the basis of the absence of additivity of the effects between single and double mutants, Vernotte et al. (1995) concluded that the couples S264 and N266, N266 and A251, and S264 and F255 interact with each other in formate binding. These results are best explained to be due to the modification, to some extent, of the “overall conformation of the D1 protein which, in turn, modified the 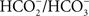 -binding at its site in the same protein.” In a way, “long-distance” (conformational?) effect on the redox properties of the QA and QB seems to support such a thesis since mutations on the d-e loop affect
-binding at its site in the same protein.” In a way, “long-distance” (conformational?) effect on the redox properties of the QA and QB seems to support such a thesis since mutations on the d-e loop affect 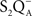 and
and 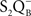 and their recombination (Maenpaa et al., 1995; Mulo et al., 1997). Maenpaa et al. (1995) found that formate which inhibits the QA to QB reaction was several-fold less effective in the CA1 mutant (where three glutamates at 242, 243, and 244 positions were deleted and glutamine 241 was changed to His); on the other hand D1-E229D and D1-E243K mutants were like the wild type Synechocystis sp. PCC 6803. Mulo et al. (1997) demonstrated, also in Synechocystis sp. PCC 6803, that deletion of either the PEST like sequence (ΔR225-F239) or the putative cleavage region (ΔG240-V249, ΔR225-V249) of the D1 protein resulted in severe perturbations on the function of the QB electron acceptor of PSII.
and their recombination (Maenpaa et al., 1995; Mulo et al., 1997). Maenpaa et al. (1995) found that formate which inhibits the QA to QB reaction was several-fold less effective in the CA1 mutant (where three glutamates at 242, 243, and 244 positions were deleted and glutamine 241 was changed to His); on the other hand D1-E229D and D1-E243K mutants were like the wild type Synechocystis sp. PCC 6803. Mulo et al. (1997) demonstrated, also in Synechocystis sp. PCC 6803, that deletion of either the PEST like sequence (ΔR225-F239) or the putative cleavage region (ΔG240-V249, ΔR225-V249) of the D1 protein resulted in severe perturbations on the function of the QB electron acceptor of PSII.
Xiong et al. (1997), considering all the results available till then, presented a model of PSII, based on homology with the bacterial system, and introduced the hypothesis of how 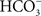 may function in PSII. Obviously, only the basic points are known: HC was suggested in the literature to bind to the non-heme iron that sits between QA and QB. To accommodate other experiments, particularly those on Chlamydomonas mutants (Hutchison et al., 1996; Xiong et al., 1997), specific arginines were implicated as well as conformational changes. What is known from the work of Eaton-Rye and Govindjee(1988a,b) on spinach thylakoids is that HC functions not only in the reduction of QB, but also in its protonation. Recent studies have suggested that D2-K264 (Cox et al., 2009), but more likely D1-Y246 and D2-Y244 (Takahashi et al., 2009) are directly involved in HC binding. Figure 10 shows the binding of
may function in PSII. Obviously, only the basic points are known: HC was suggested in the literature to bind to the non-heme iron that sits between QA and QB. To accommodate other experiments, particularly those on Chlamydomonas mutants (Hutchison et al., 1996; Xiong et al., 1997), specific arginines were implicated as well as conformational changes. What is known from the work of Eaton-Rye and Govindjee(1988a,b) on spinach thylakoids is that HC functions not only in the reduction of QB, but also in its protonation. Recent studies have suggested that D2-K264 (Cox et al., 2009), but more likely D1-Y246 and D2-Y244 (Takahashi et al., 2009) are directly involved in HC binding. Figure 10 shows the binding of 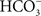 to the electron-acceptor side of PSII from the 1.9 Å resolution structure of PSII (Umena et al., 2011). Here the importance of D1-Y246 and D2-Y244 in the HC effect is clearly obvious. Komenda et al. (2002) discovered that a mutant of Synechocystis sp. PCC 6803, that lacked PsbH protein, had a much weakened binding of HC on the PSII acceptor side; this observation needs to be understood in light of the recent PSII crystal structure (Umena et al., 2011).
to the electron-acceptor side of PSII from the 1.9 Å resolution structure of PSII (Umena et al., 2011). Here the importance of D1-Y246 and D2-Y244 in the HC effect is clearly obvious. Komenda et al. (2002) discovered that a mutant of Synechocystis sp. PCC 6803, that lacked PsbH protein, had a much weakened binding of HC on the PSII acceptor side; this observation needs to be understood in light of the recent PSII crystal structure (Umena et al., 2011).
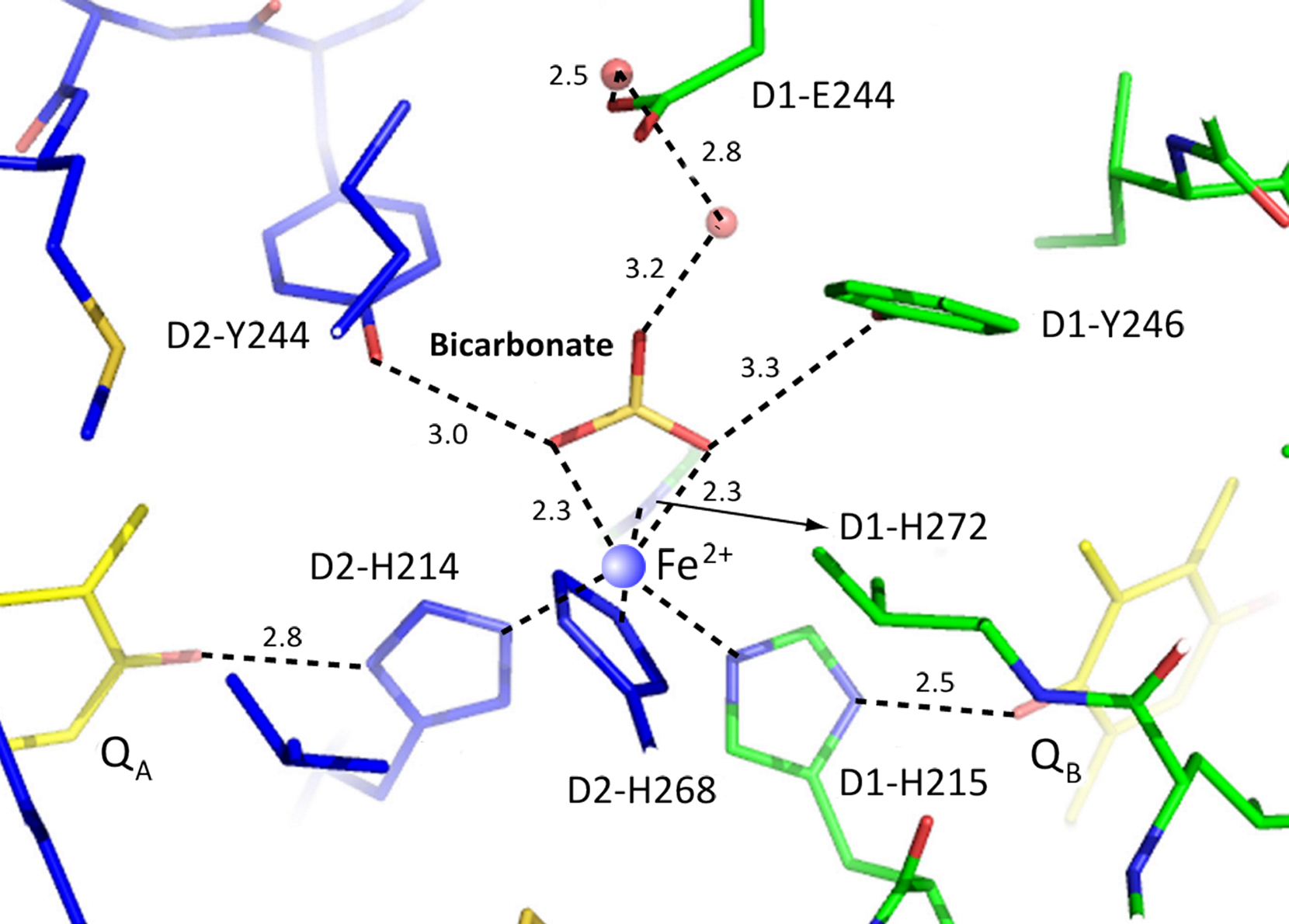
Figure 10. PSII crystal structure from thermophilic cyanobacterium Thermosynechococcus vulcanus (Courtesy of Jian-Ren Shen, personal communication, 2011 to the authors; see Umena et al., 2011). This figure shows the location of HC (bicarbonate) that is bound to non-heme iron (Fe2+; see blue circle in the center). Amino acids within 3.3 Å of HC are two tyrosines and one histidine: D1-Y246; D2-Y244; D1-H272. D2-K264 is not shown. (The concept has evolved in Govindjee’s laboratory, beginning with the work of Alan Stemler, followed by extensive work of Thomas J. Wydrzynski, Paul Jursinic, Rita Khanna, Danny Blubaugh, Julian Eaton-Rye, Sarojini Bose, Fatima El-Shintinawy, Jack J.S. Van Rensen, Wim F.J. Vermaas, Chunhe Xu, Jinchiang Cao, and Jin Xiong (the latter two worked on cyanobacteria) that HC plays a key role on the electron-acceptor side of PSII.) These researchers among many others, have suggested that there is a unique role of HC in the electron flow from reduced QA to  , and proton transport to
, and proton transport to  (Van Rensen et al., 1999). The model shown here provides detailed information that will allow future experimenters to find the molecular mechanism of the HC effect on the PSII.
(Van Rensen et al., 1999). The model shown here provides detailed information that will allow future experimenters to find the molecular mechanism of the HC effect on the PSII.
Answer to the HC problem lies in future studies on the mutants of these and some other amino acids. So, we would say that a new hope has arisen and we can hardly wait to get the final answers!
We end this review with remarks on evolution (for recent reviews, see Buick, 2008; Björn and Govindjee, 2009; Blankenship, 2010; Hohmann-Marriott and Blankenship, 2011). Figure 11 shows a simplified model of the origin and evolution of life (tree of life) based on small-subunit RNA analysis. Despite the simple representation of the origin of the first oxygenic photosynthesizers in this Figure, the earliest steps in the origin of photosynthesis are not known, and are the subject of vivid discussions in the literature (Blankenship et al., 2007; Buick, 2008 and references therein).
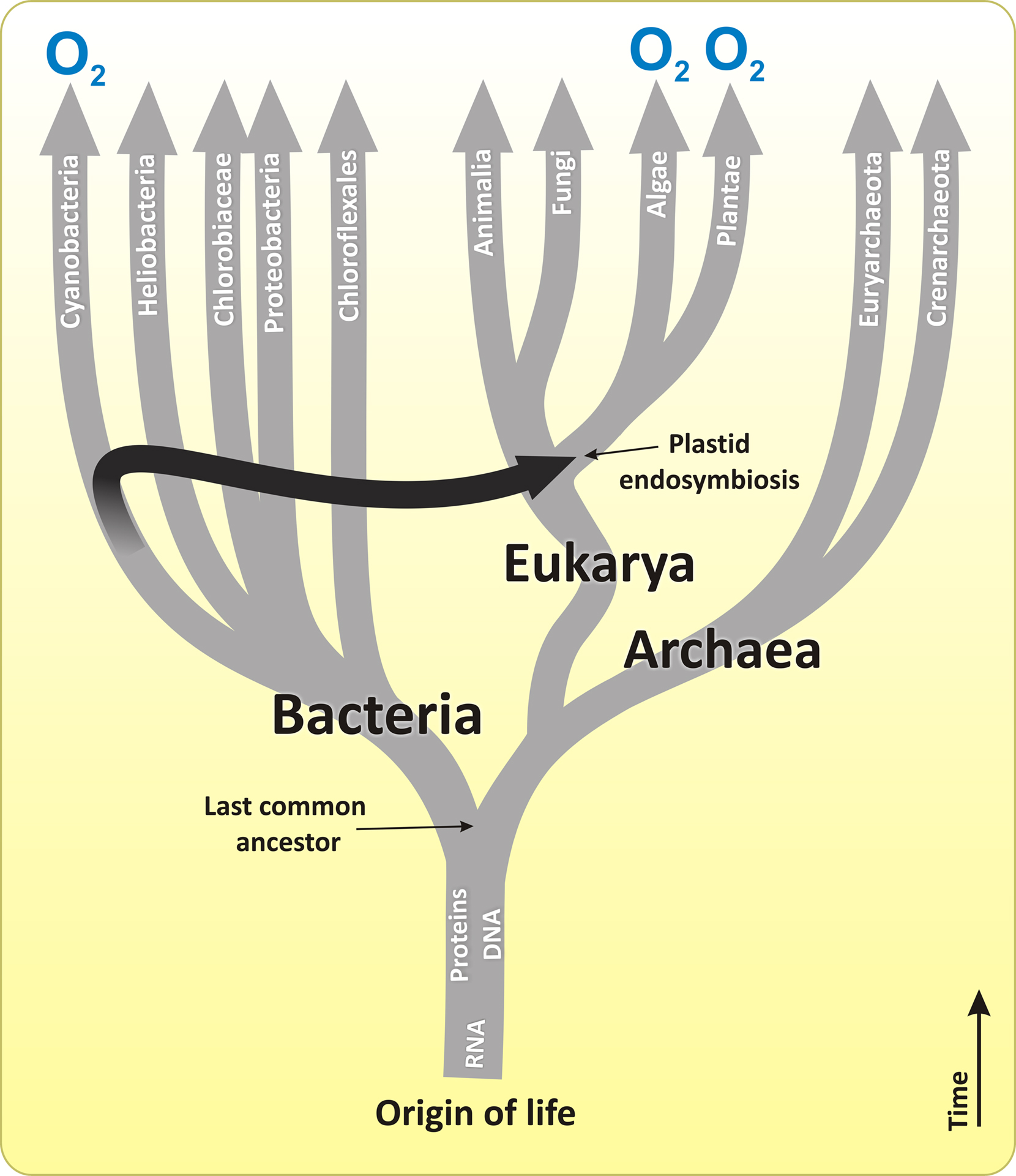
Figure 11. The origin and evolutionary tree of life that is based on small-subunit RNA. The branches that perform oxygenic photosynthesis are labeled with 'O2'. The black arrow indicates the plastid endosymbiotic event that resulted in the origin of eukaryotic photosynthesis from cyanobacteria-like organisms, which ultimately became chloroplasts in algae and later in plants. However, while chloroplasts of the higher plants, glaucophytes, green and red algae are thought to be the result of the plastid (primary) endosymbiosis, all other groups of algae are assumed to have arisen due to the algal (secondary and tertiary) endosymbiosis (not shown), in which one eukaryotic alga was incorporated into another eukaryote. For details, see Olson and Blankenship (2004), Blankenship et al. (2007), Hohmann-Marriott and Blankenship (2011), Kim et al. (2011), Garcia-Mendoza et al. (2011). Only some branches of bacteria, eukarya, and archaea are displayed. Modified and adapted from Blankenship (2002) and Blankenship et al. (2007).
The first appearance of the atmosphere and the oceans on our Earth is dated to about 4.5 Ga ago. It happened soon after the Earth completed its formation stage (Nisbet and Sleep, 2001). Based on the carbon isotope data, the earliest forms of life are thought to have emerged about 3.7–3.8 Ga ago (Mojzsis et al., 1996; Falkowski, 2006). Although photosynthesis was, probably, not the earliest metabolic system on Earth, it, however, arose very early in the history of our planet. It is generally accepted that simple anoxygenic organisms carried out the first photosynthetic activity. These first photobacteria were dependent on the availability of electron donors (reductants) such as H2, H2S, and/or Fe(OH)+ which they used as substrates (Olson, 2006; Blankenship, 2010; Hohmann-Marriott and Blankenship, 2011). However, these substances were much less abundant and thus limited compared to the huge (almost unlimited) H2O pool on the surface of the planet. During the period dated between 3.2 and 2.4 Ga ago, some cyanobacteria-like organisms managed to utilize H2O as a substrate (i.e., as a source of electrons and protons) for the reduction of CO2 using the energy of sunlight to drive this reaction (Oparin, 1965; Xiong and Bauer, 2002; Tomitani et al., 2006; Allen and Martin, 2007; Blankenship et al., 2007; see also Figure 1 and its legend).
Water is a very stable compound. If one considers oxidation of water to O2 and protons, the Em for such a process would be +0.82 V at pH 7 (Atkins and De Paula, 2006; Messinger and Renger, 2008). How did the first cyanobacteria contrive to do it? The following evolutionary developments are thought to have preceded the successful water-splitting by the first O2-producing organisms:
• Origin of RCs and the development of the two types (PSII and PSI) of functionally linked RCs ending up in the same organism was undoubtedly, the key evolutionary step toward the ability by the first cyanobacteria to extract electrons from water. While cyanobacteria (as well as green algae and higher plants) contain two types of RCs (PSII and PSI; Figure 4), all anoxygenic bacteria have only one of the two types: RC1 (pheophytin–quinone type) or RC2 (iron–sulfur type; Figure 12). Therefore, it is generally accepted that the origin of the oxygenic photosynthetic organisms begins with the origin of the linked RCs in a common ancestor of O2-evolving cyanobacteria (for details, see, Xiong and Bauer, 2002; Olson and Blankenship, 2004). Currently two models for the origin of RCs are discussed (for reviews, see Vermaas, 2002; Olson and Blankenship, 2004). According to the selective loss model, the two types of RCs (RC2 and RC1) developed separately in the same organism and later become functionally linked. In this model, the anoxygenic photosynthetic organisms are derived from a primitive cyanobacterium by a loss of either RC1 or RC2. In the fusion model, the RC2 and RC1 developed separately in distinct organisms and later ended up together in one organism by a large-scale lateral gene transfer. For details on the models, see Blankenship (1992), Xiong and Bauer (2002), Blankenship et al. (2007), Hohmann-Marriott and Blankenship (2011). However, whatever model is true, in order to oxidize water to molecular oxygen, two more major changes from the more primitive non-O2-evolving RCs were also required: addition of a charge-accumulating system (i.e., OEC) and that of a RC pigment with a very high oxidizing potential.
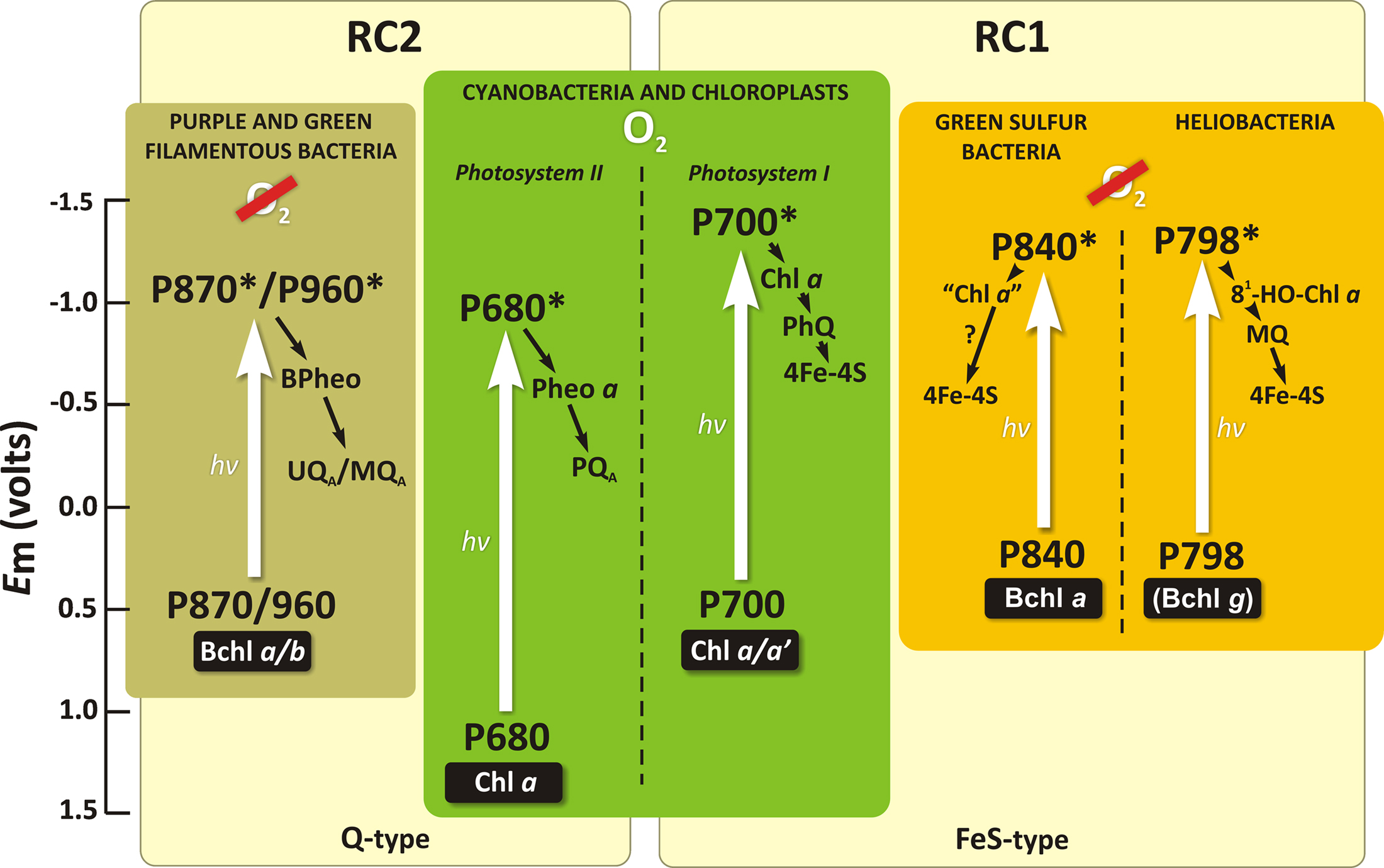
Figure 12. A simplified scheme representing the RCs in various groups of photosynthetic organisms. The initial electron donor (P), as well as the primary and secondary electron acceptors are shown for each RC. Straight white arrows are for excitation of RC molecules that leads to an electron jump from the ground state into the higher excited singlet state, in response to the absorption of light quanta (hv). The number behind P indicates the long wavelength absorption maxima (in nm) of the RC electron donor. Other abbreviations are: BChl, bacteriochlorophyll; BPheo, bacteriopheophytin; Chl, chlorophyll; Fe–S-type, iron–sulfur type of RC; Fe–S, iron–sulfur center; Q-type, pheophytin–quinone type of RC; MQ, menaquinone; PhQ, phylloquinone; PQ, plastoquinone; UQ, ubiquinone. Modified and adapted from Blankenship (2002) and Blankenship (2010).
• The development of the catalytic site of water oxidation (MnCa-protein complex/OEC) that is capable of collecting and storing positive charges was, undoubtedly, a central stage in the transition from anoxygenic bacteria to oxygenic cyanobacteria (Allen and Martin, 2007; Raymond and Blankenship, 2008). The evolutionary origin of the OEC is obscure. Currently, it is thought to have been derived either from Mn catalase enzymes (Blankenship and Hartman, 1998), Mn–HC complexes (Dismukes et al., 2001; Khorobrykh et al., 2008), or Mn(Ca)-containing minerals (Sauer and Yachandra, 2002; Najafpour et al., 2010). Dash et al. (2011) speculated that Cyt c in anoxygenic photosynthetic bacteria might have been replaced by the addition of a Mn-Ca entity by assimilating Mn-Ca minerals like hollandite.
• The evolutionary development of photosynthetic pigments (Chls) and of a strongly oxidizing RC (PSII) most probably preceded or evolved simultaneously with the invention of the OEC (Blankenship and Hartman, 1998; Björn et al., 2009; Blankenship, 2010). A strong oxidant with redox potential greater than +0.82 V was needed to split water into O2 and four protons. We do not know the midpoint redox potential of the RC photoactive pigment in ancient cyanobacteria. In modern water-splitting organisms, the Em value of the RC photoactive Chl a (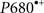 ) is about +1.25 V (Rappaport et al., 2002). Such a high value must be due to a special protein environment (Renger and Holzwarth, 2005). This oxidizing power is about half a volt above the special bacteriochlorophyll (BChl) pair (BChl-L–BChl-M) in the RCs of all anoxygenic bacteria that contain various BChl forms: a, b, or g (Blankenship et al., 1995; Figure 12). BChl a has been suggested to have evolved before Chl a (see e.g., Burke et al., 1993; Raymond et al., 2003; Olson and Blankenship, 2004). In the common ancestor of cyanobacteria, Fe(OH)+ and later H2O2 may have been the electron donors for photosynthetic CO2 fixation until BChl a was replaced by Chl a in the RCs and the OEC was invented (Rutherford and Nitschke, 1996; Blankenship and Hartman, 1998; Borda et al., 2001; Olson, 2006; Raymond and Blankenship, 2008). However, Ohashi et al. (2010) have proposed that the oxygen-sensitive BChl g could be the precursor for Chl a. For many years, Chl a had been thought to be unique and be the essential pigment among the Chl species in photosynthetic O2-evolving organisms. However, with the discovery by Miyashita et al. (1996) of the Chl d-containing cyanobacterium A. marina, the role of Chl a versus Chl d began to be discussed (Akimoto et al., 2006; Kobayashi et al., 2007; Tomo et al., 2007; Cser et al., 2008; Allakhverdiev et al., 2011). Blankenship and Hartman (1998) proposed that Chl d may have been a transitional evolutionary pigment between BChl a-containing RCs of anoxygenic bacteria and Chl a-containing RCs of the first oxygenic cyanobacteria (Blankenship et al., 2007). However, alternatively, it could also well be a more recent adaptation to a particular light environment (Kuhl et al., 2005): the Acaryochloris-like organisms live in environments rich in near-IR light, but show features of adaptation to strong light (Larkum and Kuhl, 2005; Miller et al., 2005). The latter option is supported by the data of Schliep et al. (2010) indicating that Chl a is the ancestor of Chl d.
) is about +1.25 V (Rappaport et al., 2002). Such a high value must be due to a special protein environment (Renger and Holzwarth, 2005). This oxidizing power is about half a volt above the special bacteriochlorophyll (BChl) pair (BChl-L–BChl-M) in the RCs of all anoxygenic bacteria that contain various BChl forms: a, b, or g (Blankenship et al., 1995; Figure 12). BChl a has been suggested to have evolved before Chl a (see e.g., Burke et al., 1993; Raymond et al., 2003; Olson and Blankenship, 2004). In the common ancestor of cyanobacteria, Fe(OH)+ and later H2O2 may have been the electron donors for photosynthetic CO2 fixation until BChl a was replaced by Chl a in the RCs and the OEC was invented (Rutherford and Nitschke, 1996; Blankenship and Hartman, 1998; Borda et al., 2001; Olson, 2006; Raymond and Blankenship, 2008). However, Ohashi et al. (2010) have proposed that the oxygen-sensitive BChl g could be the precursor for Chl a. For many years, Chl a had been thought to be unique and be the essential pigment among the Chl species in photosynthetic O2-evolving organisms. However, with the discovery by Miyashita et al. (1996) of the Chl d-containing cyanobacterium A. marina, the role of Chl a versus Chl d began to be discussed (Akimoto et al., 2006; Kobayashi et al., 2007; Tomo et al., 2007; Cser et al., 2008; Allakhverdiev et al., 2011). Blankenship and Hartman (1998) proposed that Chl d may have been a transitional evolutionary pigment between BChl a-containing RCs of anoxygenic bacteria and Chl a-containing RCs of the first oxygenic cyanobacteria (Blankenship et al., 2007). However, alternatively, it could also well be a more recent adaptation to a particular light environment (Kuhl et al., 2005): the Acaryochloris-like organisms live in environments rich in near-IR light, but show features of adaptation to strong light (Larkum and Kuhl, 2005; Miller et al., 2005). The latter option is supported by the data of Schliep et al. (2010) indicating that Chl a is the ancestor of Chl d.
• Finally, the coupling of 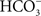 ions to PSII in the first oxygenic photosynthesizers to facilitate quinone reduction could be speculated to be an additional and the latest evolutionary step from anoxygenic/low-efficient-oxygenic to oxygenic/highly efficient oxygenic photosynthesis. The presence of HC bound between QA and QB is unique as it exists only in O2-evolving organisms and is absent in anoxygenic photosynthesizers (Shopes et al., 1989; Wang et al., 1992).
ions to PSII in the first oxygenic photosynthesizers to facilitate quinone reduction could be speculated to be an additional and the latest evolutionary step from anoxygenic/low-efficient-oxygenic to oxygenic/highly efficient oxygenic photosynthesis. The presence of HC bound between QA and QB is unique as it exists only in O2-evolving organisms and is absent in anoxygenic photosynthesizers (Shopes et al., 1989; Wang et al., 1992).
Already ∼2.3 Ga ago, photosynthetic H2O-splitting cyanobacteria started to be the dominant photosynthetic organisms. Geological and geochemical evidence clearly indicates that around this time, free molecular oxygen started to accumulate in significant amounts in the atmosphere (Figure 1; Bekker et al., 2004). Mass spectrometric evidence for a high driving force of O2 evolution by PSII in cyanobacteria and higher plants has been provided by Shevela et al. (2011), confirming earlier indications (Haumann et al., 2008; Kolling et al., 2009). All together, the data obtained clearly indicate that elevated O2 levels (up to 20–50 bars) do not suppress photosynthetic oxygen production. Therefore, it is obvious, that the evolution of oxygenic photosynthesis was not influenced and restricted by the rise of the O2 content in the atmosphere in the past. The rapid development of prokaryotic oxygenic photosynthesis by the first cyanobacteria dramatically changed the Earth by creating an aerobic atmosphere. In turn, this event forced the anaerobic forms of life either to find O2-free niches or to adapt to the high O2 levels by developing protective mechanisms. Those organisms that did not adapt to the presence of O2 faced extinction (Sleep and Bird, 2008). Subsequent evolution of eukaryotic oxygenic photosynthesis due to plastid (primary) endosymbiosis (see Figures 1 and 11), and later due to algal (secondary) endosymbiosis (see Chan and Bhattacharya, 2010; Chan et al., 2011; Hohmann-Marriott and Blankenship, 2011) led to the rise of [O2] to the present day level (∼ 20%). Moreover, the appearance of oxygen levels in high amounts in the atmosphere also led to the formation of the protective ozone layer that absorbs a large part of the UV radiation from the Sun. These new conditions ultimately permitted the development of an aerobic metabolism and more-advanced forms of live. All fossil fuels that we use at this moment are derived from past photosynthesis.
Today, the natural process of oxygenic photosynthesis may serve as a blueprint for developing artificial photosynthesis for human needs (Kawakami et al., 2011; Najafpour and Govindjee, 2011). We need to learn how to transform the unlimited supply of sunlight into a storable and transportable form of energy, such as biomass, or hydrogen that is ecologically a “clean” fuel. This would help us in solving the rising energy demand, taking into consideration the necessity for the supply of “clean” energy. We ask: Can cyanobacteria help us in this regard? And, what can we learn from them? The answer to the first question is “Yes,” but the details of what can we learn from them is being debated around the World.
It is a common knowledge that a large number of cyanobacteria (as well as green algae and plants) use light as the driving force to extract electrons from water in order to generate strong reductants, such as NADPH and reduced ferredoxin, which can, in principle, be utilized as substrates for molecular hydrogen production by either hydrogenases and nitrogenases (for recent reviews, see Lubitz et al., 2008; Ghirardi et al., 2009). Figure 13 shows a hydrogenase-dependent pathway for molecular hydrogen production in cyanobacteria. However, several biochemical barriers exist for the sustained and sufficient H2 production in cyanobacteria. The most important are: (i) the irreversible inhibition of most hydrogenases by O2; (ii) the consumption of photobiologically produced H2 by the bidirectional hydrogenases; (iii) the competition of the Calvin–Benson cycle and H2 production for NADPH (Tamagnini et al., 2007). Because of these limitations, photobiological H2 production occurs only under special conditions and with low yields in the wild type cyanobacteria. Mutagenesis, metabolic engineering, as well as other approaches are being employed to overcome these restrictions (Esper et al., 2006; Ghirardi et al., 2007, 2009; Lubitz et al., 2008), but this unique and complex system, which evolved ∼ 3 Ga ago, has not been successfully used by humanity thus far.
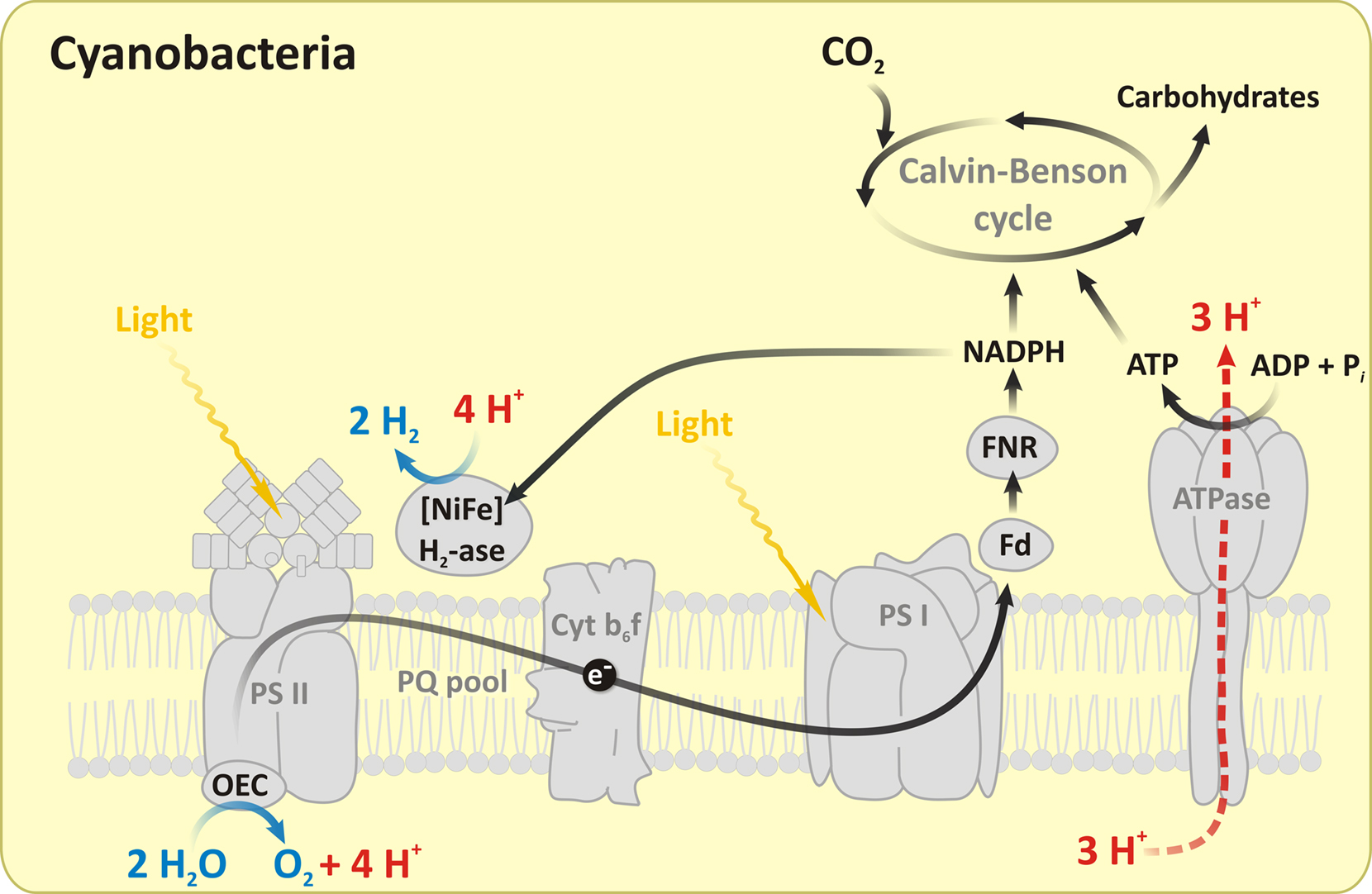
Figure 13. A schematic representation of H2 photoproduction pathway catalyzed by a hydrogenase in cyanobacteria. As in green algae, photosynthetic electron transport delivers electrons after light-induced water-splitting at the OEC to the hydrogenases (H2-ase), leading to photophosphorylation and H2 production. In addition to hydrogenases, nitrogenases are also known to produce H2 in oxygenic cyanobacteria and in anoxygenic purple bacteria. Abbreviations: PQ, plastoquinone; Fd, ferredoxin; FNR, ferredoxin–NADP reductase; ATPase, adenosine triphosphate. The figure is modified and adapted from Shevela and Messinger (2011).
In this respect, Francis Crick was absolutely right, saying that “Evolution is cleverer than you are” (Dennett, 1984). Undoubtedly, in the future, the efforts of the numerous cyanobacteria research groups hold a promise for us all. The adventure with cyanobacteria continues…
The authors stated that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We thank all the co-authors of our past research, as without their participation this perspective of cyanobacterial photosynthesis would not have been possible. Jian-Ren Shen is gratefully acknowledged for providing the figure of his recent PSII crystal structure. We also take this opportunity to refer the readers to a time-line on photosynthesis (Govindjee and Krogmann, 2004), to a web article “Photosynthesis Online” (Orr and Govindjee, 2011): <http://www.life.illinois.edu/govindjee/photoweb/>; and to a conversation between Donald R. Ort and one of us (G): <http://ensemble.atlas.uiuc.edu/app/sites/JZ64_U8FmEmx7Liwjl36Mw.aspx?webSiteID=JZ64>
Akimoto, S., Murakami, A., Yokono, M., Koyama, K., Tsuchiya, T., Miyashita, H., Yamazaki, I., and Mimuro, M. (2006). Fluorescence properties of the chlorophyll d-dominated cyanobacterium Acaryochloris sp. strain Awaji. J. Photochem. Photobiol. A Chem. 178, 122–129.
Allakhverdiev, S. I., Tsuchiya, T., Watabe, K., Kojima, A., Los, D. A., Tomo, T., Klimov, V. V., and Mimuro, M. (2011). Redox potentials of primary electron acceptor quinone molecule (QA¯) and coserved energetics of photosystem II in cyanobacteria with chlorophyll a and chlorophyll d. Proc. Natl. Acad. Sci. U.S.A. 108, 8054–8058.
Allwood, A. C., Walter, M. R., Kamber, B. S., Marshall, C. P., and Burch, I. W. (2006). Stromatolite reef from the Early Archaean era of Australia. Nature 441, 714–718.
Amesz, J., and Duysens, L. N. (1962). Action spectrum, kinetics and quantum requirement of phosphopyridine nucleotide reduction and cytochrome oxidation in the blue-green alga Anacystis nidulans. Biochim. Biophys. Acta 64, 261–278.
Aoyama, C., Suzuki, H., Sugiura, M., and Noguchi, T. (2008). Flash-induced FTIR difference spectroscopy shows no evidence for the structural coupling of bicarbonate to the oxygen-evolving Mn cluster in photosystem II. Biochemistry 47, 2760–2765.
Arnold, W., and Meek, E. S. (1956). The polarization of fluorescence and energy transfer in grana. Arch. Biochem. Biophys. 60, 82–90.
Arnold, W., and Oppenheimer, J. R. (1950). Internal conversion in the photosynthetic mechanism of blue-green algae. J. Gen. Physiol. 33, 423–435.
Arnold, W., and Sherwood, H. K. (1957). Are chloroplasts semiconductors? Proc. Natl. Acad. Sci. U.S.A. 43, 105–114.
Bedell, G. W., and Govindjee, (1973). Photophosphorylation in intact algae: effects of inhibitors, intensity of light, electron acceptor and donors. Plant Cell Physiol. 14, 1081–1097.
Bekker, A., Holland, H. D., Wang, P. L., Rumble, D., Stein, H. J., Hannah, J. L., Coetzee, L. L., and Beukes, N. J. (2004). Dating the rise of atmospheric oxygen. Nature 427, 117–120.
Bendall, D. S., and Manasse, R. S. (1995). Cyclic photophosphorylation and electron transport. Biochim. Biophys. Acta 1229, 23–38.
Björn, L. O., and Govindjee, (2009). The evolution of photosynthesis and chloroplasts. Curr. Sci. 96, 1466–1474.
Björn, L. O., Papageorgiou, G. C., Blankenship, R. E., and Govindjee, (2009). A viewpoint: why chlorophyll a? Photosynth. Res. 99, 85–98.
Blankenship, R. E. (1992). Origin and early evolution of photosynthesis. Photosynth. Res. 33, 91–111.
Blankenship, R. E., and Hartman, H. (1998). The origin and evolution of oxygenic photosynthesis. Trends Biochem. Sci. 23, 94–97.
Blankenship, R. E., Madigan, M. T., and Bauer, C. E. (eds). (1995). Anoxygenic Photosynthetic Bacteria. Dordrecht: Kluwer Academic Publishers.
Blankenship, R. E., Sadekar, S., and Raymond, J. (2007). “The evolutionary transition from anoxygenic to oxygenic photosynthesis,” in Evolution of Aquatic Photoautotrophs, eds P. G. Falkowski and A. N. Knoll (New York: Academic Press), 21–35.
Blubaugh, D. J., and Govindjee, (1986). Bicarbonate, not CO2, is the species required for the stimulation of photosystem II electron transport. Biochim. Biophys. Acta 848, 147–151.
Blubaugh, D. J., and Govindjee, (1988). Kinetics of the bicarbonate effect and the number of bicarbonate-binding sites in thylakoid membranes. Biochim. Biophys. Acta 936, 208–214.
Borda, M. J., Elsetinow, A. R., Strongin, D. R., and Schoonen, M. A. (2001). Mechanism for the production of hydrogen peroxide at metal disulfide surfaces. Abstr. Pap. Am. Chem. Soc. 222, U489.
Bosak, T., Liang, B., Sim, M. S., and Petroff, A. P. (2009). Morphological record of oxygenic photosynthesis in conical stromatolites. Proc. Natl. Acad. Sci. U.S.A. 106, 10939–10943.
Brown, J. S., and French, C. S. (1961). The long wavelength forms of chlorophyll a. Biophys. J. 1, 539–550.
Bryant, D. A. (ed.). (1994). The Molecular Biology in Cyanobacteria. Dordrecht: Kluwer Academic Publishers.
Buchanan, R. E., and Gibbons, N. E. (1974). Bergey’s Manual of Determinative Biology. Baltimore, MD: The Williams and Wilkins Company.
Buick, R. (2008). When did oxygenic photosynthesis evolve? Philos. Trans. R. Soc. Lond. B Biol. Sci. 363, 2731–2743.
Burke, D. H., Hearst, J. E., and Sidow, A. (1993). Early evolution of photosynthesis – Clues from nitrogenase and chlorophyll iron proteins. Proc. Natl. Acad. Sci. U.S.A. 90, 7134–7138.
Burnap, R. L., Shen, J.-R., Jursinic, P. A., Inoue, Y., and Sherman, L. A. (1992). Oxygen yield and thermoluminescence characteristics of a cyanobacterium lacking the manganese stabilizing protein of photosystem II. Biochemistry 31, 7404–7410.
Cao, J., and Govindjee, (1988). Bicarbonate effect on electron flow in cyanobacterium Synechocystis PCC 6803. Photosynth. Res. 19, 277–285.
Cao, J., and Govindjee, (1990). “Anion effects on the electron acceptor side of photosystem II in a transformable cyanobacterium Synechocystis 6803,” in Current Research in Photosynthesis, ed. M. Baltscheffsky (Dordrecht: Kluwer Academic Publishers), 515–518.
Cao, J., Ohad, N., Hirschberg, J., Xiong, J., and Govindjee, (1992). Binding affinity of bicarbonate and formate in herbicide-resistant D1 mutants of Synechococcus sp. PCC 7942. Photosynth. Res. 34, 397–408.
Cao, J. C., Vermaas, W. F. J., and Govindjee, (1991). Arginine residues in the D2 polypeptide may stabilize bicarbonate binding in photosystem II of Synechocystis sp. PCC 6803. Biochim. Biophys. Acta 1059, 171–180.
Carr, N. G., and Whitton, B. A. (eds). (1982). The Biology of Cyanobacteria. Berkeley: University of California Press.
Cederstrand, C. N., Rabinowitch, E., and Govindjee, (1966). Analysis of the red absorption band of chlorophyll a in vivo. Biochim. Biophys. Acta 126, 1–12.
Chan, C. X., Gross, J., Yoon, H. S., and Bhattacharya, D. (2011). Plastid origin and evolution: new models provide insights into old problems. Plant Physiol. 155, 1552–1560.
Cho, F., and Govindjee, (1970). Low-temperature (4-770K) spectroscopy of Anacystis: temperature dependence of energy transfer efficiency. Biochim. Biophys. Acta 216, 151–161.
Clegg, R. M., Sener, M., and Govindjee, (2010). “From Förster resonance energy transfer to coherent resonance energy transfer and back,” in Optical Biopsy VII, ed. R. R. Alfano (SPIE: Bellingham, WA) 7561, 1–21.
Cox, N., Jin, L., Jaszewski, A., Smith, P. J., Krausz, E., Rutherford, A. W., and Pace, R. (2009). The semiquinone-iron complex of photosystem II: structural insights from ESR and theoretical simulation; evidence that the native ligand to the non-heme iron is carbonate. Biophys. J. 97, 2024–2033.
Cser, K., Deák, Z., Telfer, A., Barber, J., and Vass, I. (2008). Energetics of photosystem II charge recombination in Acaryochloris marina studied by thermoluminescence and flash-induced chlorophyll fluorescence measurements. Photosynth. Res. 98, 131–140.
Cser, K., and Vass, I. (2007). Radiative and non-radiative charge recombination pathways in photosystem II studied by thermoluminescence and chlorophyll fluorescence in the cyanobacterium Synechocystis 6803. Biochim. Biophys. Acta 1767, 233–243.
Dash, G. C., Sahu, R. K., Bhoi, S., and Raval, M. K. (2011). “Evolution of photosynthetic reaction center,” in International Conference on Plant Science in Post Genomic Era (Jyoti Vihar, Orissa: School of Life Science, Sambalpur University and The Society for Plant Physiology and Biochemistry). Abstract PSA-047, 86.
Dennett, D. C. (1984). Elbow Room. The Varieties of Free Will Worth Wanting. Cambridge, MA: Bradford Books MIT Press.
Desikachary, T. V. (ed.). (1972). Taxonomy and Biology of Blue-Green Algae. Madras: University of Madras.
DeVault, D., and Govindjee, (1990). Photosynthetic glow peaks and their relationship with the free energy changes. Photosynth. Res. 24, 175–181.
DeVault, D., Govindjee, and Arnold, W. (1983). Energetics of photosynthetic glow peaks. Proc. Natl. Acad. Sci. U.S.A. 80, 983–987.
Diner, B. A., Petrouleas, V., and Wendoloski, J. J. (1991). The iron-quinone electron-acceptor complex of photosystem II. Physiol. Plant. 81, 423–436.
Dismukes, G. C., Klimov, V. V., Baranov, S. V., Kozlov, Y. N., Dasgupta, J., and Tyryshkin, A. (2001). The origin of atmospheric oxygen on earth: the innovation of oxygenic photosynthesis. Proc. Natl. Acad. Sci. U.S.A. 98, 2170–2175.
Duysens, L. N. M. (1952). Transfer of Excitation Energy in Photosynthesis. Doctoral thesis. Utrecht: State University.
Eaton-Rye, J. J., and Govindjee, (1984). A study of the specific effect of bicarbonate on photosynthetic electron trasport in the presence of methyl viologen. Photobiochem. Photobiophys. 8, 279–288.
Eaton-Rye, J. J., and Govindjee, (1988a). Electron transfer through the quinone acceptor complex of photosystem II after one or two actinic flashes in bicarbonate-depleted spinach thylakoid membranes. Biochim. Biophys. Acta 935, 248–257.
Eaton-Rye, J. J., and Govindjee, (1988b). Electron transfer through the quinone acceptor complex of photosystem II in bicarbonate-depleted spinach thylakoid membranes as a function of actinic flash number and frequency. Biochim. Biophys. Acta 935, 237–247.
El-Shintinawy, F., and Govindjee, (1990). Bicarbonate effect in leaf discs from spinach. Photosynth. Res. 24, 189–200.
El-Shintinawy, F., Xu, C., and Govindjee, (1990). A dual bicarbonate-reversible formate effect in Chlamydomonas cells. J. Plant Physiol. 136, 421–428.
Emerson, R., and Chalmers, R. V. (1958). Speculations concerning the function and phylogenetic significance of the accessory pigments of algae. Phycol. Soc. News Bull. 11, 51–56.
Emerson, R., and Rabinowitch, E. (1960). Red drop and role of auxiliary pigments in photosynthesis. Plant Physiol. 35, 477–485.
Emlyn-Jones, D., Ashby, M. K., and Mullineaux, C. W. (1999). A gene required for the regulation of photosynthetic light harvesting in the cyanobacterium Synechocystis 6803. Mol. Microbiol. 33, 1050–1058.
Esper, B., Badura, A., and Rögner, M. (2006). Photosynthesis as a power supply for (bio-)hydrogen production. Trends Plant Sci. 11, 543–549.
Fagerlund, R. D., and Eaton-Rye, J. J. (2011). The lipoproteins of cyanobacterial photosystem II. J. Photochem. Photobiol. B Biol. 104, 191–203.
Falkowski, P. G. (2006). Tracing oxygen’s imprint on Earth’s metabolic evolution. Science 311, 1724–1725.
Falkowski, P. G. (2011). The biological and geological contingencies for the rise of oxygen on Earth. Photosynth. Res. 107, 7–10.
Falkowski, P. G., and Raven, J. A. (2007). Aquatic Photosynthesis. Princeton: Princeton University Press.
Fischer, K., and Metzner, H. (1969). “On chlorophyll and pigment P750 of Anacystis nidulans,” in Progress in Photosynthesis Research, ed. H. Metzner (Tübingen: International Union of Biological Sciences), 547–551.
Garab, G., Rozsa, Z., and Govindjee, (1988). Carbon dioxide affects charge accummulation in leaves: measurements by thermoluminescence. Naturwissenschaften 75, 517–519.
Garcia-Mendoza, E., Ocampo-Alvarez, H., and Govindjee, (2011). Photoprotection in the brown alga Macrocystis pyrifera: evolutionary implications. J. Photochem. Photobiol. B Biol. 104, 377–385.
Gassner, E. B. (1962). On the pigment absorbing at 750 nm occurring in some blue-green algae. Plant Physiol. 37, 637–639.
Gault, P. M., and Marler, H. J. (2009). Handbook on Cyanobacteria: Biochemistry, Biotechnology and Applications. New York, NY: Nova Science Publishers.
Ghirardi, M. L., Dubini, A., Yu, J., and Maness, P.-C. (2009). Photobiological hydrogen-producing systems. Chem. Soc. Rev. 38, 52–61.
Ghirardi, M. L., Posewitz, M. C., Maness, P. C., Dubini, A., Yu, J., and Seibert, M. (2007). Hydrogenases and hydrogen photoproduction in oxygenic photosynthetic organisms. Annu. Rev. Plant Biol. 58, 71–91.
Goedheer, J. H. C. (1972). Fluorescence in relation to photosynthesis. Annu. Rev. Plant Physiol. 23, 87–112.
Goedheer, J. H. C., and Hammans, J. W. K. (1975). Efficiency of light conversion by the blue-green alga Anacystis nidulans. Nature 256, 333–335.
Golbeck, J. H. (ed.). (2006). Photosystem I. The Light-Driven Plastocyanin:Ferredoxin Oxidoreductase. Dordrecht: Springer.
Gombos, Z., Szalontat, B., Suranyi, G., and Csatorday, K. (1987). Selective alteration of the pigment composition og the blue-green alga, Anacystis nidulans. Photochem. Photobiol. 45, 399–402.
Govindjee, (1963a). “Emerson enhancement effect and two light reactions in photosynthesis: dedicated to the memory of late Professor Robert Emerson,” in Photosynthetic Mechanisms of Green Plants, eds B. Kok and A. T. Jagendorf (Washington, DC: National Academy of Sciences – National Research Council), 318–334.
Govindjee, (1991). “A unique role of CO2 in photosystem II,” in Impact of Global Climatic Changes on Photosynthesis and Plant Productivity, eds Y. Abrol, P. N. Wattal, A. Gnanam, Govindjee, D. R. Ort and A. H. Teramura (New Delhi: Oxford/IBH Private Ltd.), 349–369.
Govindjee, (1993). Bicarbonate-reversible inhibition of plastoquinone reductase in photosystem II. Z. Naturforsch. C 48, 251–258.
Govindjee, (1995). Sixty-three years since Kautsky: chlorophyll a fluorescence. Aust. J. Plant Physiol. 22, 131–160.
Govindjee, (1999). “Carotenoids in photosynthesis: an historical perspective,” in The Photochemistry of Carotenoids: Applications in Biology, eds H. A. Frank A. J. Young, G. Britton, and R. J. Cogdell (Dordrecht: Kluwer Academic Publishers), 1–19.
Govindjee, (2004). “Chlorophyll a fluorescence: a bit of basics and history,” in Chlorophyll a Fluorescence: A Probe of Photosynthesis, eds G. C. Papageorgiou and Govindjee (Dordrecht: Kluwer Academic Publishers), 3–40.
Govindjee, Amesz, J., and Fork, D. C. (eds). (1986). Light Emission by Plants and Bacteria. Orlando: Academic Press.
Govindjee, and Björn, L. O. (2011). “Dissecting oxygenic photosynthesis: the evolution of the “Z”-scheme for thylakoid reactions,” in Photosynthesis: Overviews on Recent Progress and Future Perspectives, eds S. Itoh, P. Mohanty and K. N. Guruprasad (New Delhi: IK Publishers), 1–27.
Govindjee, and Braun, B. Z. (1974). “Light absorption, emission and photosynthesis,” in Algal Physiology and Biochemistry, ed. W. D. P. Stewart (Oxford: Blackwell Scientific Publication Ltd.), 346–390.
Govindjee, Cederstrand, C., and Rabinowitch, E. (1961). Existence of absorption bands at 730-740 and 750-760 millimicrons in algae of different divisions. Science 134, 391–392.
Govindjee, Eggenberg, P., Pfister, K., and Strasser, R. J. (1992). Chlorophyll a fluorescence yield decay in herbicide-resistant D1 mutant of Chlamydomonas reinhardtii and the formate effect. Biochim. Biophys. Acta 1101, 353–258.
Govindjee, Kern, J., Messinger, J., and Whitmarsh, J. (2010). “Photosystem II,” in Encyclopedia of Life Sciences (ELS). (Chichester: John Wiley & Sons, Ltd.). doi: 10.1002/9780470015902.a0000669.pub2.
Govindjee, Koike, H., and Inoue, Y. (1985). Thermo-luminescence and oxygen evolution from a thermophilic blue-green-alga obtained after single-turnover light flashes. Photochem. Photobiol. 42, 579–585.
Govindjee, and Krogmann, D. (2004). Discoveries in oxygenic photosynthesis (1727-2003): a perspective. Photosynth. Res. 80, 15–57.
Govindjee, and Mohanty, P. (1972). “Photochemical aspects of photosynthesis in blue-green algae,” in Biology and Taxonomy of Blue-Green Algae, ed. T. Desikachary (Madras: University of Madras), 171–196.
Govindjee, Nakatani, H. Y., Rutherford, A. W., and Inoue, Y. (1984). Evidence from thermoluminescence for bicarbonate action on the recombination reactions involving the secondary quinone electron acceptor of photosystem II. Biochim. Biophys. Acta 766, 416–423.
Govindjee, and Papageorgiou, G. C. (1971). “Chlorophyll fluorescence and photosynthesis: fluorescence transients,” in Photophysiology, ed. A. C. Giese (New York: Academic Press), 1–46.
Govindjee, Pulles, M. P. J., Govindjee, R., Van Gorkom, H. J., and Duysens, L. N. M. (1976). Inhibition of reoxidation of secondary-electron acceptor of photosystem II by bicarbonate depletion. Biochim. Biophys. Acta 449, 602–605.
Govindjee, and Rabinowitch, E. (1960a). Action spectrum of the second Emerson effect. Biophys. J. 1, 73–89.
Govindjee, and Rabinowitch, E. (1960b). Two forms of chlorophyll a in vivo with distinct photochemical function. Science 132, 355–356.
Govindjee, Rabinowitch, E., and Thomas, J. B. (1960). Inhibition of photosynthesis in certain algae by extreme red light. Biophys. J. 1, 91–97.
Govindjee, Schwarz, B., Rochaix, J. D., and Strasser, R. J. (1991a). The herbicide-resistant D1 mutant L275F of Chlamydomonas reinhardtii fails to show the bicarbonate-reversible formate effect on chlorophyll a fluorescence transients. Photosynth. Res. 27, 199–208.
Govindjee, Weger, H. G., Turpin, D. H., Van Rensen, J. J. S., devos, O. J., and Snel, J. F. H. (1991b). Formate releases carbon dioxide/bicarbonate from thylakoid membranes – measurements by mass spectroscopy and infrared gas analyzer. Naturwissenschaften 78, 168–170.
Govindjee, Snel, J. F. H., devos, O. J., and Van Rensen, J. J. S. (1993). Antagonistic effects of light I and light II on chlorophyll a fluorescence yield and P700 turnover as monitors of carbon dioxide depletion in intact algal and cyanobacterial cells. Physiol. Plant. 89, 143–148.
Govindjee, and Van Rensen, J. J. S. (1978). Bicarbonate effects on the electron flow in isolated broken chloroplasts. Biochim. Biophys. Acta 505, 183–213.
Govindjee, and Van Rensen, J. J. S. (1993). “Photosystem II reaction centers and bicarbonate,” in Photosynthetic Reaction Centers, eds J. Deisenhofer and J. R. Norris (Orlando: Academic Press), 357–389.
Govindjee, Vernotte, C., Peteri, B., Astier, C., and Etienne, A. L. (1990). Differential sensitivity of bicarbonate-reversible formate effects on herbicide-resistant mutants of Synechocystis 6714. FEBS Lett. 267, 273–276.
Govindjee, Wang, Q. J., Singh, A., Li, H., Nedbal, L., Sherman, L. A., and Whitmarsh, C. J. (2011). ”Mixotrophically-grown photosystem I deletion mutants* of Synechocystis sp. PCC 6803 are capable of light-induced net oxygen production in the presence of glucose,” in Molecular Bioenergetics of Cyanobacteria: From Cells to Community (Costa Brava: EMBO, European Science Foundation), 40.
Govindjee, Xu, C., and Van Rensen, J. J. S. (1997). On the requirement of bound bicarbonate for photosystem II activity. Z. Naturforsch. 52, 24–32.
Guskov, A., Gabdulkhakov, A., Broser, M., Glöckner, C., Hellmich, J., Kern, J., Frank, J., Mül, F., Saenger, W., and Zouni, A. (2010). Recent progress in the crystallographic studies of photosystem II. Chemphyschem 11, 1160–1171.
Hammans, J. W. K. (1978). P750 sensitized photooxidations in Anacystis nidulans. Plant Physiol. 19, 1457–1463.
Hammans, K. J. W., Hendriks, G. M., and Teerlink, T. (1977). Light dependent oxygen uptake by the blue green alga Anacystis nidulans. Biochem. Biophys. Res. Commun. 74, 1560–1565.
Haumann, M., Grundmeier, A., Zaharieva, I., and Dau, H. (2008). Photosynthetic water oxidation at elevated dioxygen partial pressure monitored by time-resolved X-ray absorption measurements. Proc. Natl. Acad. Sci. U.S.A. 105, 17384–17389.
Herrero, A., Flores, E., and Flores, F. G. (eds). (2008). The Cyanobacteria: Molecular Biology, Genomics, and Evolution. Norwich: Horizon Scientific Press/Caister Academic Press.
Hohmann-Marriott, M. F., and Blankenship, R. E. (2011). Evolution of photosynthesis. Annu. Rev. Plant Biol. 62, 515–548.
Hunter, C. N., Daldal, F., Thurnauer, M. C., and Beatty, J. T. (eds). (2009). The Purple Phototrophic Bacteria. Dordrecht: Springer.
Hutchison, R. S., Xiong, J., Sayre, R. T., and Govindjee, (1996). Construction and characterization of a photosystem II D1 mutant (arginine-269-glycine) of Chlamydomonas reinhardtii. Biochim. Biophys. Acta 1277, 83–92.
Jursinic, P., Warden, J., and Govindjee, (1976). A major site of bicarbonate effect in system II reaction. Evidence from ESR signal II vf, fast fluorescence yield changes and delayed light emission. Biochim. Biophys. Acta 440, 322–330.
Kana, R., Komárek, O., Kotabová, E., Papageorgiou, G. C., Govindjee, and Práśil, O. (2010). “Light-induced state 2-to-state-1 transition is mainly responsible for the S-to-M rise in the chlorophyll a fluorescence induction of the cyanobacterium Synechoccystis sp. PCC 6803,” in 15th International Congress on Photosynthesis (Beijing: Abstract book), 223.
Kana, R., Prasil, O., Komarek, O., Papageorgiou, G. C., and Govindjee, (2009). Spectral characteristic of fluorescence induction in a model cyanobacterium, Synechococcus sp. PCC 7942. Biochim. Biophys. Acta 1787, 1170–1178.
Kawakami, K., Umena, Y., Kamiya, N., and Shen, J.-R. (2011). Structure of the catalytic inorganic core of oxygen evolving photosystem II at 1.9 Å resolution. J. Photochem. Photobiol. B Biol. 104, 9–18.
Ke, B. (2001). Photosynthesis: Photochemistry and Photobiology. Dordrecht: Kluwer Academic Publishers.
Keranen, M., Mulo, P., Aro, E. M., Govindjee, and Tyystjarvi, E. (1998). “Thermoluminescence B and Q bands are the same temperature in an autotrophic and a heterotrophic D1 protein mutant of Synechocystis sp. PCC 6803,” in Photosynthesis: Mechanisms and Effects, ed. G. Garab (Dordrecht: Kluwer Academis Publishers), 1145–1148.
Khanna, R., Govindjee, and Wydrzynski, T. (1977). Site of bicarbonate effect in Hill reaction. Evidence from the use of artificial electron acceptors and donors. Biochim. Biophys. Acta 462, 208–214.
Khanna, R., Pfister, K., Keresztes, A., Van Rensen, J. J., and Govindjee, (1981). Evidence for a close spatial location of the binding sites for CO2 and for photosystem II inhibitors. Biochim. Biophys. Acta 634, 105–116.
Khanna, R., Wagner, R., Junge, W., and Govindjee, (1980). Effects of CO2-depletion on proton uptake and release in thylakoid membranes. FEBS Lett. 121, 222–224.
Khorobrykh, A. A., Terentyev, V. V., Zharmukhamedov, S. K., and Klimov, V. V. (2008). Redox interaction of Mn-bicarbonate complexes with reaction centres of purple bacteria. Philos. Trans. R. Soc. Lond. B Biol. Sci. 363, 1245–1251.
Kim, E., Harrison, J. W., Sudek, S., Jones, M. D. M., Wilcox, H. M., Richards, T. A., Worden, A. Z., and Archibald, J. M. (2011). Newly identified and diverse plastid-bearing branch on the eukaryotic tree of life. Proc. Natl. Acad. Sci. U.S.A. 108, 1496–1500.
Kobayashi, M., Ohashi, S., Iwamoto, K., Shiraiwa, Y., Kato, Y., and Watanabe, T. (2007). Redox potential of chlorophyll d in vitro. Biochim. Biophys. Acta 1767, 596–602.
Kok, B. (1959). Light induced absorption changes in photosynthetic organisms. II. A split-beam difference spectrophotometer. Plant Physiol. 34, 184–192.
Kolling, D. R. J., Brown, T. S., Ananyev, G., and Dismukes, G. C. (2009). Photosynthetic oxygen evolution is not reversed at high oxygen pressures: mechanistic consequences for the water-oxidizing complex. Biochemistry 48, 1381–1389.
Komenda, J., Lupinkova, L., and Kopecky, J. (2002). Absence of the PsbH gene product destabilizes photosystem II complex and bicarbonate binding on its acceptor side in Synechocystis PCC 6803. Eur. J. Biochem. 269, 610–619.
Kuhl, M., Chen, M., Ralph, P. J., Schreiber, U., and Larkum, A. W. D. (2005). A niche for cyanobacteria containing chlorophyll d. Nature 433, 820–820.
Larkum, A. W. D., and Kuhl, M. (2005). Chlorophyll d: the puzzle resolved. Trends Plant Sci. 10, 355–357.
Lindell, D., Jaffe, J. D., Johnson, Z. I., Church, G. M., and Chisholm, S. W. (2005). Photosynthesis genes in marine viruses yield proteins during host infection. Nature 438, 86–89.
Lubberding, H. J., and Schroten, W. (1984). The ATP level in the thermophilic cyanobacterium Synechococcus 6716 during light-dark transition and in the presence of some specific inhibitors. FEMS Microbiol. Lett. 22, 93–96.
Lubitz, W., Reijerse, E. J., and Messinger, J. (2008). Solar water-splitting into H2 and O2: design principles of photosystem II and hydrogenases. Energy Environ. Sci. 1, 15–31.
Maenpaa, P., Miranda, T., Tyystjarvi, E., Tyystjarvi, T., Govindjee, Ducruet, J. M., Etienne, A. L., and Kirilovsky, D. (1995). A mutation in the D-de loop of D-1 modifies the stability of the S2QA- and S2QB- states in photosystem II. Plant Physiol. 107, 187–197.
Maksimov, E. G., Kuzminov, F. I., Konyuhov, I. V., Elanskaya, I. V., and Paschenko, V. Z. (2011). Photosystem 2 effective fluorescence cross-section of cyanobacterium Synechocystis sp. PCC 6803 and its mutants. J. Photochem. Photobiol. B Biol. 104, 285–291.
Mann, N. H., Cook, A., Millard, A., Bailey, S., and Clokie, M. (2003). Marine ecosystems: bacterial photosynthesis genes in a virus. Nature 424, 741–741.
Mar, T., and Govindjee, (1972). “Decrease in the degree of polarization of chlorophyll fluorescence upon addition of DCMU in algae,” in Photosynthesis, Two Centuries After its Discovery by Joseph Priestley, eds G. Forti, M. Avron and A. Melandri (Den Haag: Dr. W. Junk N. V. Publishers), 271–281.
Mar, T., Govindjee, Singhal, G. S., and Merkelo, H. (1972). Lifetime of the excited state in vivo. I. Chlorophyll a in algae, at room and at liquid nitrogen temperatures; rate constants of radiationless deactivation and trapping. Biophys. J. 12, 797–808.
McConnell, I. L., Eaton-Rye, J. J., and Van Rensen, J. J. S. (2011). “Regulation of photosystem II electron transport by bicarbonate,” in Photosynthesis: Plastid Biology, Energy Conversion and Carbon Assimilation, Chapt. 20, eds J. J. Eaton-Rye, B. C. Tripathy and T. D. Sharkey (Dordrecht: Springer), 471–496.
Meetam, M., Keren, N., Itzhak Ohad, I., and Pakrasi, H. B. (1999). The PsbY protein is not essential for oxygenic photosynthesis in the cyanobacterium Synechocystis sp. PCC 6803. Plant Physiol. 121, 1267–1272.
Messinger, J., and Renger, G. (2008). “Photosynthetic water splitting,” in Primary Processes of Photosynthesis, Part 2 Principles and Apparatus, ed. G. Renger (Cambridge: RSC Publishing), 291–351.
Miller, S. R., Augustine, S., Le Olson, T., Blankenship, R. E., Selker, J., and Wood, A. M. (2005). Discovery of a free-living chlorophyll d-producing cyanobacterium with a hybrid proteobacterial/cyanobacterial small-subunit rRNA gene. Proc. Natl. Acad. Sci. U.S.A. 102, 850–855.
Mimuro, M. (2004). “Photon capture, excitation migration and trapping and fluorescence emission in cyanobacteria and red algae,” in Chlorophyll a Fluorescence: A Signature of Photosynthesis, eds G. C. Papageorgiou and Govindjee (Dordrecht: Springer), 173–195.
Mimuro, M., Kobayashi, M., Murakami, A., Tsuchiya, T., and Miyashita, H. (2008). “Oxygen-evolving cyanobacteria,” in Primary Processes of Photosynthesis, Part 1, ed. G. Renger (Cambridge: RSC Publishing), 261–300.
Miyashita, H., Ikemoto, H., Kurano, N., Adachi, K., Chihara, M., and Miyachi, S. (1996). Chlorophyll d as a major pigment. Nature 383, 402–402.
Mohanty, P., and Govindjee, (1973a). Effect of phenazine methosulfate and uncouplers on light-induced chlorophyll a fluorescence yield changes in intact algal cells. Photosynthetica 7, 146–160.
Mohanty, P., and Govindjee, (1973b). Light-induced changes in the fluorescence yield of chlorophyll a in Anacystis nidulans. I. Relationship of slow fluorescence changes with structural changes. Biochim. Biophys. Acta 305, 95–104.
Mohanty, P., and Govindjee, (1973c). Light-induced changes in the fluorescence yield of chlorophyll a in Anacystis nidulans. II. The fast changes and the effect of photosynthetic inhibitors on both fast and slow fluorescence induction. Plant Cell Physiol. 14, 611–629.
Mohanty, P., and Govindjee, (1974). The slow decline and the subsequent rise of chlorophyll fluorescence transients in intact algal cells. Plant Biochem. J. 1, 78–106.
Mojzsis, S. J., Arrhenius, G., Mckeegan, K. D., Harrison, T. M., Nutman, A. P., and Friend, C. R. L. (1996). Evidence for life on Earth before 3,800 million years ago. Nature 384, 55–59.
Mulo, P., Tyystjarvi, T., Tyystjarvi, E., Govindjee, Maenpaa, P., and Aro, E. M. (1997). Mutagenesis of the D-E loop of photosystem II reaction centre protein D1. Function and assembly of photosystem II. Plant Mol. Biol. 33, 1059–1071.
Murata, N., Sato, N., Omata, T., and Kuwabara, T. (1981). Separation and characterization of thylakoid and cell envelope of the blue-green alga (cyanobacterium) Anacystis nidulans. Plant Cell Physiol. 22, 855–866.
Najafpour, M. M., Ehrenberg, T., Wiechen, M., and Kurz, P. (2010). Calcium manganese(III) oxides (CaMn2O4·xH2O) as biomimetic oxygen-evolving catalysts. Angew. Chem. Int. Ed. 49, 2233–2237.
Najafpour, M. M., and Govindjee, (2011). Oxygen evolving complex in photosystem II: better than excellent. Dalton Trans. doi: 10.1039/C1dt10746a
Nedbal, L., Brezina, V., Adamec, F., Stys, D., Oja, V., Laisk, A., and Govindjee, (2003). Negative feedback regulation is responsible for the non-linear modulation of photosynthetic activity in plants and cyanobacteria exposed to a dynamic light environment. Biochim. Biophys. Acta 1607, 5–17.
Nisbet, E. G., and Sleep, N. H. (2001). The habitat and nature of early life. Nature 409, 1083–1091.
Nugent, J. H. A., Corrie, A. R., Demetriou, C., Evans, M. C. W., and Lockett, C. J. (1988). Bicarbonate binding and the properties of photosystem II electron acceptors. FEBS Lett. 235, 71–75.
Nultsch, W., Schuchart, H., and Koenig, F. (1983). Effects of sodium azide on phototaxis of the blue-green alga Anabaena variabilis and consequences to the two-photoreceptor systems-hypothesis. Arch. Microbiol. 134, 33–37.
Ohashi, S., Iemura, T., Okada, N., Itoh, S., Furukawa, H., Okuda, M., Ohnishi-Kameyama, M., Ogawa, T., Miyashita, H., Watanabe, T., Itoh, S., Oh-Oka, H., Inoue, K., and Kobayashi, M. (2010). An overview on chlorophylls and quinones in the photosystem I-type reaction centers. Photosynth. Res. 104, 305–319.
Olson, J. M., and Blankenship, R. E. (2004). Thinking about the evolution of photosynthesis. Photosynth. Res. 80, 373–386.
Oparin, I. A. (1965). “The origin of life and the origin of enzymes,” in Advances in Enzymology and Related Areas of Molecular Biology, ed. F. F. Nord (New York: Interscience Publishers), 347–380.
öquist, G. (1974). Iron deficiency in the blue-green alga Anacystis nidulans: fluorescence and absorption spectra recorded at 77oK. Physiol. Plant. 31, 55–58.
Orr, L., and Govindjee, (2011). Photosynthesis online. Photosynth. Res. 105, 167–200. doi: 10.1007/s11120-010-9570-8
Owens, O. H., and Hoch, G. (1963). Enhancement and de-enhancement effect in Anacystis nidulans. Biochim. Biophys. Acta 75, 183–186.
Papageorgiou, G. C. (1975). “Chlorophyll fluorescence: an intrinsic probe of photosynthesis,” in Bioenergetics of Photosynthesis, ed. Govindjee (New York: Academic Press), 319–372.
Papageorgiou, G. C. (1996). The photosynthesis of cyanobacteria (blue bacteria) from the perspective of signal analysis of chlorophyll a fluorescence. J. Sci. Ind. Res. (C) 55, 596–617.
Papageorgiou, G. C., and Govindjee, (1967). Changes in intensity and spectral distribution of fluorescence. Effect of light treatment on normal and DCMU-poisoned Anacystis nidulans . Biophys. J. 7, 375–389.
Papageorgiou, G. C., and Govindjee, (1968). Light-induced changes in the fluorescence yield of chlorophyll a in vivo. I. Anacystis nidulans. Biophys. J. 8, 1299–1315.
Papageorgiou, G. C., and Govindjee, (eds).(2004). Chlorophyll a Fluorescence: A signature of Photosynthesis. Dordrecht: Springer.
Papageorgiou, G. C., and Govindjee, (2011). Photosystem II fluorescence: slow changes – scaling from the past. J. Photochem. Photobiol. B Biol. 104, 258–270.
Papageorgiou, G. C., Tsimilli-Michael, M., and Stamatakis, K. (2007). The fast and slow kinetics of chlorophyll a fluorescence induction in plants, algae and cyanobacteria: a viewpoint. Photosynth. Res. 94, 275–290.
Peschek, G. A., Obinger, C., and Renger, G. (eds). (2011). Bioenergetic Processes of Cyanobacteria: From Evolutionary Singularity to Ecological Diversity. Dordrecht: Springer.
Rabinowitch, E., and Govindjee, (1965). The role of chlorophyll in photosynthesis. Sci. Am. 213, 74–83.
Rabinowitch, E., Govindjee, and Thomas, J. B. (1960). Inhibition of photosynthesis in some algae by extreme-red light. Science 132, 422–423.
Rappaport, F., Guergova-Kuras, M., Nixon, P. J., Diner, B. A., and Lavergne, J. (2002). Kinetics and pathways of charge recombination in photosystem II. Biochemistry 41, 8518–8527.
Raymond, J., and Blankenship, R. E. (2008). The origin of the oxygen-evolving complex. Coord. Chem. Rev. 252, 377–383.
Raymond, J., Zhaxybayeva, O., Gogarten, J. P., and Blankenship, R. E. (2003). Evolution of photosynthetic prokaryotes: a maximum-likelihood mapping approach. Philos. Trans. R. Soc. Lond. B Biol. Sci. 358, 223–230.
Renger, G., and Holzwarth, A. R. (2005). “Primary electron transfer,” in Photosystem II. The Light-Driven Water:Plastoquinone Oxidoreductase, eds T. J. Wydrzynski and K. Satoh (Dordrecht: Springer), 139–175.
Roose, J. L., Wegener, K. M., and Pakrasi, H. B. (2007). The extrinsic proteins of photosystem II. Photosynth. Res. 92, 369–387.
Rose, S., Minagawa, J., Seufferheld, M., Padden, S., Svensson, B., Kolling, D. R., Crofts, A. R., and Govindjee, (2008). D1-arginine257 mutants (R257E, K, and Q) of Chlamydomonas reinhardtii have a lowered QB redox potential: analysis of thermoluminescence and fluorescence measurements. Photosynth. Res. 98, 449–468.
Ruengjitchatchawalya, M., Kovacs, L., Mapaisansup, T., Sallai, A., Gombos, Z., Ponglikitmongkol, M., and Tanticharoen, M. (2005). Higher plant-like fluorescence induction and thermoluminescence characteristics in cyanobacterium, Spirulina mutant defective in PQH2 oxidation by cytb6/f complex. J. Plant Physiol. 162, 1123–1132.
Rutherford, A. W., and Nitschke, W. (1996). “Photosystem II and the quinone-iron-containing reaction centers: comparison and evolutionary perspectives,” in Origin and Evolution of Biological Energy Conversion, ed. H. Batscheffky (New York: VCH), 143–174.
Sane, P. V., and Rutherford, A. W. (1986). “Thermoluminescence from photosynthetic membranes,” in Light Emission by Plants and Bacteria, eds Govindjee, J. Amesz and D. C. Fork (New York: Academic Press), 329–360.
Sapp, J. (2005a). Microbial Phylogeny and Evolution: Concepts and Controversies. New York: Oxford University Press.
Sapp, J. (2005b). The prokaryote-eukaryote dichotomy: meanings and mythology. Microbiol. Mol. Biol. Rev. 69, 292–305.
Sarojini, G., and Govindjee, (1981). On the active species in bicarbonate stimulation of Hill reaction in thylakoid membranes. Biochim. Biophys. Acta 634, 340–343.
Sauer, K., and Yachandra, V. K. (2002). A possible evolutionary origin for the Mn4 cluster of the photosynthetic water oxidation complex from natural MnO2 precipitates in the early ocean. Proc. Natl. Acad. Sci. U.S.A. 99, 8631–8636.
Schliep, M., Crossett, B., Willows, R. D., and Chen, M. (2010). 18O labeling of chlorophyll d in Acaryochloris marina reveals that chlorophyll a and molecular oxygen are precursors. J. Biol. Chem. 285, 28450–28456.
Sharon, I., Alperovitch, A., Rohwer, F., Haynes, M., Glaser, F., Atamna-Ismaeel, N., Pinter, R. Y., Partensky, F., Koonin, E. V., Wolf, Y. I., Nelson, N., and Beja, O. (2009). Photosystem I gene cassettes are present in marine virus genomes. Nature 461, 258–262.
Shevela, D., Beckmann, K., Clausen, J., Junge, W., and Messinger, J. (2011). Membrane-inlet mass spectrometry reveals a high driving force for oxygen production by photosystem II. Proc. Natl. Acad. Sci. U.S.A. 108, 3602–3607.
Shevela, D., Klimov, V., and Messinger, J. (2008a). “Formate-induced release of carbon dioxide/hydrogencarbonate from photosystem II,” in Photosynthesis. Energy from the Sun: 14th International Congress on Photosynthesis, eds J. F. Allen, E. Gantt, J. H. Golbeck and B. Osmond (Glasgow: Springer), 497–501.
Shevela, D., Su, J. H., Klimov, V., and Messinger, J. (2008b). Hydrogencarbonate is not a tightly bound constituent of the water-oxidizing complex in photosystem II. Biochim. Biophys. Acta 1777, 532–539.
Shevela, D., and Messinger, J. (2011). “Principles of photosynthesis,” in Fundamentals of Materials and Energy and Environmental Sustainability, Chapt. 24, eds D. Ginley and D. Cachen (Cambridge: Cambridge University Press) (in press).
Shevela, D., Nöring, B., Eckert, H. J., Messinger, J., and Renger, G. (2006). Characterization of the water oxidizing complex of photosystem II of the Chl d-containing cyanobacterium Acaryochloris marina via its reactivity towards endogenous electron donors and acceptors. Phys. Chem. Chem. Phys. 8, 3460–3466.
Shimony, C., Spencer, J., and Govindjee, (1967). Spectral characteristics of Anacystis particles. Photosynthetica 1, 113–125.
Shopes, R. J., Blubaugh, D. J., Wraight, C. A., and Govindjee, (1989). Absence of a bicarbonate-depletion effect in electron transfer between quinones in chromatophores and reaction centers of Rhodobacter sphaeroides. Biochim. Biophys. Acta 974, 114–118.
Shubin, V. V., Murthy, S. D. S., Karapetyan, N. V., and Mohanty, P. (1991). Origin of the 77 K variable fluorescence at 758 nm in the cyanobacterium Spirulina platensis. Biochim. Biophys. Acta 1060, 28–36.
Singhal, G. S., Mohanty, P., and Govindjee, (1981). Effects of preheating intact algal cells on pigments revealed by absorption and fluorescence spectra. Z. Pflanzenphysiol. 103, 217–228.
Sleep, N. H., and Bird, D. K. (2008). Evolutionary ecology during the rise of dioxygen in the Earth’s atmosphere. Philos. Trans. R. Soc. Lond. B Biol. Sci. 363, 2651–2664.
Stemler, A. (1982). “The functional role of bicarbonate in photosynthetic light reaction II,” in Photosynthesis, ed. Govindjee (New York: Academic Press), 513–538.
Stemler, A. J. (2002). The bicarbonate effect, oxygen evolution, and the shadow of Otto Warburg. Photosynth. Res. 73, 177–183.
Stirbet, A., and Govindjee, (2011). On the relation between the Kautsky effect (chlorophyll a fluorescence induction) and photosystem II: basics and applications of the OJIP fluorescence transient. J. Photochem. Photobiol. B Biol. 104, 236–257.
Strasser, R. J., Srivastava, A., and Govindjee, (1995). Polyphasic chlorophyll a fluorescence transient in plants and cyanobacteria. Photochem. Photobiol. 61, 32–42.
Strasser, R. J., Tsimilli-Michael, M., and Srivastava, A. (2004). “Analysis of the chlorophyll fluorescence transient,” in Chlorophyll a Fluorescence: A Signature of Photosynthesis, eds G. C. Papageorgiou and Govindjee (Dordrecht: Springer), 321–362.
Swingley, W. D., Chen, M., Cheung, P. C., Conrad, A. L., Dejesa, L. C., Hao, J., Honchak, B. M., Karbach, L. E., Kurdoglu, A., Lahiri, S., Mastrian, S. D., Miyashita, H., Page, L., Ramakrishna, P., Satoh, S., Sattley, W. M., Shimada, Y., Taylor, H. L., Tomo, T., Tsuchiya, T., Wang, Z. T., Raymond, J., Mimuro, M., Blankenship, R. E., and Touchman, J. W. (2008). Niche adaptation and genome expansion in the chlorophyll d-producing cyanobacterium Acaryochloris marina. Proc. Natl. Acad. Sci. U.S.A. 105, 2005–2010.
Szalay, L., Rabinowitch, E., Murty, N., and Govindjee, (1967). Relationship between the absorption and emission spectra and the red drop in the action spectra of fluorescence in vivo. Biophys. J. 7, 137–149.
Takahashi, R., Boussac, A., Sugiura, M., and Noguchi, T. (2009). Structural coupling of a tyrosine side chain with the non-heme iron center in photosystem II as revealed by light-induced Fourier transform infrared difference spectroscopy. Biochemistry 48, 8994–9001.
Tamagnini, P., Leitao, E., Oliveira, P., Ferreira, D., Pinto, F., Harris, D. J., Heidorn, T., and Lindblad, P. (2007). Cyanobacterial hydrogenases: diversity, regulation and applications. FEMS Microbiol. Rev. 31, 692–720.
Tomitani, A., Knoll, A. H., Cavanaugh, C. M., and Ohno, T. (2006). The evolutionary diversification of cyanobacteria: molecular-phylogenetic and paleontological perspectives. Proc. Natl. Acad. Sci. U.S.A. 103, 5442–5447.
Tomo, T., Okubo, T., Akimoto, S., Yokono, M., Miyashita, H., Tsuchiya, T., Noguchi, T., and Mimuro, M. (2007). Identification of the special pair of photosystem II in a chlorophyll d-dominated cyanobacterium. Proc. Natl. Acad. Sci. U.S.A. 104, 7283–7288.
Tsimilli-Michael, M., Stamatakis, K., and Papageorgiou, G. C. (2009). Dark-to-light transition in Synechococcus sp. PCC 7942 cells studied by fluorescence kinetics assesses plastoquinone redox poise in the dark and photosystem II fluorescence component and dynamics during state 2 to state 1 transition. Photosynth. Res. 99, 243–255.
Ulas, G., Olack, G., and Brudvig, G. W. (2008). Evidence against bicarbonate bound in the O2-evolving complex of photosystem II. Biochemistry 47, 3073–3075.
Umena, Y., Kawakami, K., Shen, J.-R., and Kamiya, N. (2011). Crystal structure of oxygen-evolving photosystem II at a resolution of 1.9 Å. Nature 473, 55–60.
Van Baalen, C. (1965). The photooxidation of uric acid by Anacystis nidulans. Plant Physiol. 40, 368–371.
Van Rensen, J. J. S., and Klimov, V. V. (2005). “Bicarbonate interactions,” in Photosystem II. The Light-Driven Water:Plastoquinone Oxidoreductase, eds T. Wydrzynski and K. Satoh. (Dordrecht: Springer), 329–346.
Van Rensen, J. J. S., and Vermaas, W. F. J. (1981). Action of bicarbonate and photosystem 2 inhibiting herbicides on electron-transport in pea grana and in thylakoids of a blue-green-alga. Physiol. Plant. 51, 106–110.
Van Rensen, J. J. S., Xu, C., and Govindjee, (1999). Role of bicarbonate in photosystem II, the water-plastoquinone oxido-reductase of plant photosynthesis. Physiol. Plant. 105, 585–592.
Van Thor, J. J., Jeanjean, R., Havaux, M., Sjollema, K. A., Joset, F., Hellingwerf, K. J., and Matthijs, H. C. (2000). Salt shock-inducible photosystem I cyclic electron transfer in Synechocystis PCC6803 relies on binding of ferredoxin:NADP+ reductase to the thylakoid membranes via its CpcD phycobilisome-linker homologous N-terminal domain. Biochim. Biophys. Acta 1457, 129–144.
Vass, I. (2005). “The history of photosynthetic thermoluminescence,” in Discoveries in Photosynthesis, eds Govindjee, J. T. Beatty, H. Gest and J. F. Allen (Dordrecht: Springer), 665–680.
Vass, I., and Govindjee, (1996). Thermoluminescence from the photosynthetic apparatus. Photosynth. Res. 48, 117–126.
Vavilin, D., Xu, H., Lin, S., and Vermaas, W. (2003). Energy and electron transfer in photosystem II of a chlorophyll b-containing Synechocystis sp. PCC 6803 mutant. Biochemistry 42, 1731–1746.
Vermaas, W. F. J. (2002). “Evolution of photosynthesis,” in Encyclopedia of Life Sciences (ELS) (London: John Wiley & Sons, Ltd), 1–18.
Vermaas, W. F. J., and Govindjee, (1982). “Bicarbonate or CO2 as a requirement for efficient electron transport on the acceptor side of photosystem II,” in Photosynthesis. Development, Carbon Metabolism, and Plant Productivity, ed. Govindjee (New York: Academic Press), 541–558.
Vernotte, C., Briantais, J.-M., Astier, C., and Govindjee, (1995). Differential effects of formate in single and double mutants of D1 in Synechocystis species PCC 6714. Biochim. Biophys. Acta 1229, 296–301.
Wang, X., Cao, J., Maroti, P., Stilz, H. U., Finkele, U., Lauterwasse, C., Zinth, W., Oesterhelt, D., Govindjee, and Wraight, C. A. (1992). Is bicarbonate in photosystem II the equivalent of the glutamate ligand to the iron atom in bacterial reaction centers. Biochim. Biophys. Acta 1100, 1–8.
Worcester, D. L., Michalski, T. J., and Katz, J. J. (1986). Small-angle neutron scattering studies of chlorophyll micelles: models for bacterial antenna chlorophyll. Proc. Natl. Acad. Sci. U.S.A. 83, 3791–3795.
Wydrzynski, T., and Satoh, K. (eds). (2005). Photosystem II. The Light-Driven Water:Plastoquinone Oxidoreductase. Dordrecht: Springer.
Xiong, J., and Bauer, C. E. (2002). Complex evolution of photosynthesis. Annu. Rev. Plant Biol. 53, 503–521.
Xiong, J., Hutchison, R. S., Sayre, R. T., and Govindjee, (1997). Modification of the photosystem II acceptor side function in a D1 mutant (arginine-269-glycine) of Chlamydomonas reinhardtii. Biochim. Biophys. Acta 1322, 60–76.
Keywords: Anacystis nidulans (Synechococcus elongatus – strain PCC 7942), Synechocystis sp. PCC 6803, P750, chlorophyll a fluorescence, red drop, Emerson enhancement effect, photosystem II, photosystem I
Citation: Govindjee and Shevela D (2011) Adventures with cyanobacteria: a personal perspective. Front. Plant Sci. 2:28. doi: 10.3389/fpls.2011.00028
Received: 15 May 2011; Accepted: 21 June 2011;
Published online: 06 July 2011.
Edited by:
Jose Manuel Estevez, University of Buenos Aires and CONICET, ArgentinaReviewed by:
Debashish Bhattacharya, Rutgers, The State University of New Jersey, USACopyright: © 2011 Govindjee and Shevela. This is an open-access article subject to a non-exclusive license between the authors and Frontiers Media SA, which permits use, distribution and reproduction in other forums, provided the original authors and source are credited and other Frontiers conditions are complied with.
*Correspondence: Govindjee, Department of Plant Biology, University of Illinois at Urbana-Champaign, 265 Morrill Hall, 505 South Goodwin Avenue, Urbana, IL 61801, USA. e-mail:Z292QGlsbGlub2lzLmVkdQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.