 Francisco J. Chorro1,2,3*
Francisco J. Chorro1,2,3* Luis Such-Miquel2,4Samuel Cuñat5Oscar Arias-Mutis2,3Patricia Genovés2,3
Luis Such-Miquel2,4Samuel Cuñat5Oscar Arias-Mutis2,3Patricia Genovés2,3 Manuel Zarzoso2,4Antonio Alberola2,5Luis Such-Belenguer2,5
Manuel Zarzoso2,4Antonio Alberola2,5Luis Such-Belenguer2,5 Irene Del Canto2,6*
Irene Del Canto2,6*- 1Department of Medicine, University of Valencia, Valencia, Spain
- 2Biomedical Research Center Network - Cardiovascular Diseases (CIBERCV), Carlos III Health Institute, Madrid, Spain
- 3Research Institute, Valencia Clinic Hospital (INCLIVA), Valencia, Spain
- 4Department of Physiotherapy, University of Valencia, Valencia, Spain
- 5Department of Physiology, University of Valencia, Valencia, Spain
- 6Department of Electronic Engineering, University of Valencia, Burjassot, Spain
Purpose: Myocardial stretch is a proarrhythmic factor. Eleclazine (GS6615) is a late sodium current (INaL) inhibitor that has shown protective effects against arrhythmias in various experimental models. Data on its effects during myocardial stretch are lacking. The aim of this study was to investigate the electrophysiological modifications induced by eleclazine basally and during acute ventricular stretch.
Methods: Left ventricular stretch was induced at baseline and during perfusion with eleclazine in 26 Langendorff rabbit heart preparations. Programmed stimulation and high-resolution mapping techniques were applied using multiple epicardial electrodes.
Results: At baseline, both the ventricular refractory period measured at a fixed cycle length (250 m) and its surrogate obtained during ventricular fibrillation (VF) decreased significantly during stretch (baseline 128 ± 15 vs. stretch 110 ± 14 m; n = 15; p < 0.001, and baseline 52 ± 13 vs. stretch 44 ± 9 m; n = 11; p < 0.05), while the VF dominant frequency (DF) increased significantly (DF baseline 13 ± 3 vs. stretch 17 ± 5Hz; n = 11; p < 0.01). Eleclazine 1.4 μM prolonged refractoriness, diminished both DF and conduction velocity during the arrhythmia, and avoided the stretch induced variations in refractoriness (baseline 148 ± 19 vs. stretch 150 ± 23 m; n = 15; ns, and baseline 73 ± 15 vs. stretch 77 ± 15 m; n = 11; ns) and in DF (baseline 12 ± 5 vs. stretch 12 ± 3 Hz; ns). The VF complexity index was inversely related to refractoriness (r = −0.64; p < 0.001). Under eleclazine perfusion, the VF activation patterns were less complex, and the arrhythmia stopped in 6 out of 11 experiments (55%; p < 0.05 vs. baseline).
Conclusion: Eleclazine (GS6615) reduced the proarrhythmic electrophysiological changes induced by myocardial stretch and slowed and simplified activation patterns during VF in the experimental model used.
1 Introduction
Cardiac arrhythmias represent a major public health concern worldwide. Related clinical manifestations such as sudden cardiac death constitute an important challenge, with the need to develop lines of research aimed at better understanding the underlying causes, mechanisms and the most effective preventive and therapeutic approaches (Marijon et al., 2023; Lynge et al., 2021; Lynge et al., 2023; Jain et al., 2024; Chatterjee, 2023). Its importance is reflected in initiatives such as that of the Lancet commission, seeking to reduce the global burden of sudden cardiac death, which highlights the need for multidisciplinary approaches (Marijon et al., 2023). The mechanisms involved in the appearance of cardiac arrhythmias are diverse and are related, among other factors, to modifications of both the substrate that supports them and the presence of triggering factors (Chatterjee, 2023; Lerman et al., 2024; Packer, 2020). Mechanical stretch acts on both components, and its effects are part of so-called mechanoelectric feedback (Quinn and Kohl, 2021; Sutherland, 2017; Johnson and Antoons, 2018; Gottlieb et al., 2023). Mechanical forces modify cellular electrophysiology through the activation and/or modulation of transmembrane ionic channels and alterations in intracellular calcium handling (Quinn and Kohl, 2021; Ravelli and Allessie, 1997; Chorro et al., 2005; Chorro et al., 2009; Reiter et al., 1988; Prosser et al., 2013). These effects favor the production of afterpotentials and the induction of reentrant activations responsible for the appearance of arrhythmias, as extensively reviewed by several authors (Quinn and Kohl, 2021; Sutherland, 2017; Johnson and Antoons, 2018).
Previous studies have shown the attenuation of the electrophysiological effects induced by mechanical stretch by Na+/H+ exchanger inhibitors such as EIPA (Chorro et al., 2015a), Na+/Ca2+ inhibitors such as KB-R7943 (Chorro et al., 2009) or betablockers such as propranolol (Chorro et al., 2009). Eleclazine (GS6615) is an INaL inhibitor (Rajamani et al., 2016; Zablocki et al., 2016; Liu et al., 2023) that has shown protective effects against arrhythmias in various experimental models (Rajamani et al., 2016; Zablocki et al., 2016; Liu et al., 2023; Fuller et al., 2016; Bacic et al., 2017; Caves et al., 2020; Shen et al., 2022). However, a reduction in arrhythmic complications has not been demonstrated in clinical studies (Shen et al., 2022; Olivotto et al., 2016). Thus, in patients with ventricular tachycardia/ventricular fibrillation treated with implantable cardioverter-defibrillators, eleclazine failed to show a significant reduction in appropriate interventions, and the clinical trials designed to evaluate the effect of eleclazine on symptoms and exercise capacity in patients with hypertrophic cardiomyopathy and to evaluate the safety and tolerability of the drug in patients with long QT syndrome type 3 were discontinued. Experimental data on its effects in other proarrhythmogenic contexts as myocardial stretch are lacking and its analysis can provide data of interest, since other INaL inhibitors such as Ranolazine and GS967 (Del-Canto et al., 2020; Chorro et al., 2015b; Milberg et al., 2013; Del Canto et al., 2018) attenuate these effects. The mechanisms involve both the inhibitory action on INaL and their actions on the rapid component of the sodium current (Burashnikov and Antzelevitch, 2017) and other ionic currents, such as the rapidly activated delayed rectifier potassium current (IKr) (Antzelevitch et al., 2004). Eleclazine is a more potent and selective INaL inhibitor and the study of its actions in this context can provide interesting data on the regulation of the electrophysiological effects produced by mechanical stretch. Myocardial stretch is characterized by an increase in Na + inflow, the activation of the reverse mode of the Na+/Ca2+ exchanger and an increase in intracellular Ca2+ that, together with changes in the oxidative balance, contribute to the activation of INaL. The activity of the sarcoplasmic reticulum, its Ca2+ content and the probability of opening the Ryanodine receptor are also altered, favoring the spontaneous release of Ca2+. Thus, in order to analyze whether a protective role is present in these settings the present study examines the effects of eleclazine in an experimental model in which the modifications of ventricular refractoriness, conduction velocity, wavelength of the activation process and activation patterns during ventricular fibrillation (VF) are analyzed both basally and during the application of acute mechanical stretch in the left ventricle.
2 Materials and methods
The present study followed European Union guidelines (2010/63) and Spanish standards (RD53/2013) on the protection of animals used for scientific purposes. The work was approved by the local Institutional Committee for Animal Care and Use (2015/VSC/PEA/00233 type 2).
2.1 Experimental protocol
We used isolated and perfused heart preparations from New Zealand rabbits with a mean weight of 4.5 ± 0.4 kg (Langendorff technique) for the experiments (n = 26). Following the intravenous administration of heparin and thiopental (60 mg per kg), the heart was removed, submerged in cold Tyrode, and subsequently connected to the system via the aorta. The Tyrode was perfused at 37°C ± 0.5°C, the pH was maintained at 7.4, and oxygenation and pH support of the solution were carried out with carbogen (O2: 95% and CO2: 5%). Mean pressure was set at 70 mmHg. A MapTech device (Waalre, Netherlands) and multiple electrode (121 unipolar electrodes, spacing = 1 mm) located on the surface of the anterior wall of the left ventricle were used to record electrophysiological signals. The indifferent electrode was placed over the aorta. Electrical stimuli (duration = 2 m; double intensity of the diastolic threshold) were applied with a Grass S88 stimulator). In order to induce local stretching of the left ventricular free wall, an L-shaped device was introduced into this camera through the left atrium (Chorro et al., 2009; Chorro et al., 2015a). The device consisted of a hollow tube measuring 3.5 mm in diameter, through which a stem 1.5 mm in diameter could be advanced. The distal end of the stem protruded from the L-shaped device and consisted of a circular platform measuring 7.5 mm in diameter, with which controlled stretching of a circumscribed zone of ventricle wall could be induced. A 12% increase in the vertical and transverse axes of the stretched area was obtained by moving (6 mm) the support of the circular platform located in the ventricular cavity. The L-shaped device was introduced before the stabilization period prior to the electrophysiological protocol.
2.2 Electrophysiological protocol and parameters analyzed
After 30 min of stabilization, programmed stimulation protocols were applied at baseline, during the perfusion of eleclazine at three concentrations (in a range around the IC50 value for late Na + current blockade [0.35 μM, 0.7 μM, and 1.4 μM]) (Rajamani et al., 2016), and after drug washout. The concentrations were chosen based on the information provided by other studies such as that of Rajamani et al. (2016) who in rabbit ventricular myocytes reported an IC50 value of 0.72 ± 0.06 μM at a holding potential of −120 mV and 0.26 ± 0.01 μM at a holding potential of −80 mV. In the study by Bacic et al. (2017) in which a porcine model was used, they observed that eleclazine administered at a dose of 0.3 mg/kg significantly reduced the incidence of VT in isolated hearts and the plasma levels of the drug were around 0.5 μM. The extrastimulus test was applied to obtain ventricular refractory periods during a fixed cycle length (250 m) in a series of 15 experiments. Ventricular pacing was performed at increasing frequencies to induce ventricular fibrillation in another series of 11 experiments. Coronary perfusion was maintained during the arrhythmia. In each phase, the parameters to be studied were obtained before stretch, 3 min after the start of its application and after stretch suppression.
The parameters analyzed were ventricular refractory periods, conduction velocity, mean and fifth percentile of VV intervals during VF (P5), VF dominant frequency (DF) and spectral concentration (SpC), and characteristics of the activation maps during arrhythmia. The DF is the greatest amplitude in the spectral power distribution of the signal, and SpC is the percentage of the spectrum area in the DF ± 0.5 Hz range. Activation times in each electrode were determined by identifying the moment of maximum negative slope of the electrograms. The minimum threshold for dV/dt to be judged as a local deflection was a percentage (20%) of the maximal negative slope in each channel. The fibrillation interval (VV) histograms and the mean of the consecutive VV intervals were determined during 2-s time windows. Spectral parameters, the VV intervals histograms and the mean value of the consecutive VV intervals were determined for the recordings obtained with all 121 unipolar electrodes of the multiple electrode, during time windows of 2 s. The activation maps during VF were constructed after determining the local activation times and corresponding isochrones during time windows of 1 s. These maps were classified by complexity into types I (a single activation front without lines of conduction block), II (two fronts or one with block lines), and III (three or more fronts with block lines). The conduction velocity (CV) was determined by dividing the distance between two electrodes (spaced 5 mm apart) by the difference between the activation times (average of 5 determinations). During VF, this parameter was determined on the wavefronts where both arrival and exit were recorded in the analyzed area, and to determine the wavelength, the conduction velocity was multiplied by P5 (Chorro et al., 2005; Chorro et al., 2015a).
2.3 Statistical analysis
Continuous variables were reported as the mean ± standard deviation (SD), and categorical variables were reported as percentages. The general linear model was used to compare the differences in repeated measures, using Bonferroni’s correction in multiple comparisons. Differences with p < 0.05 values were considered to be statistically significant. The differences between qualitative variables were analyzed using contingency tables and Fisher’s exact test when indicated. Calculations were made using the SPSS version 22.0 statistical package.
3 Results
3.1 Ventricular refractoriness
The ventricular effective refractory period (VERP) (n = 15) obtained prior to, during, and following myocardial stretch are displayed in Figure 1. Stretch caused a significant decrease in VERP at baseline (pre 128 ± 15 vs. stretch 110 ± 14 m, p < 0.001) and during eleclazine perfusion at 0.35 μM (pre 130 ± 15 vs. stretch 119 ± 18 m, p < 0.05), while the variation did not reach statistical significance at concentrations of 0.7 μM (pre 138 ± 25 vs. stretch 135 ± 32 m, ns) or 1.4 μM (pre 148 ± 19 vs. stretch 150 ± 23 m, ns) (Figure 2). A significant increase in this parameter was observed compared with baseline during perfusions of 0.7 μM and 1.4 μM. In the final experimental phase (after drug washout), refractoriness was reduced compared with the baseline values, and stretch again caused a significant decrease in VERP (pre 110 ± 15 vs. stretch 99 ± 14 m, p < 0.05).
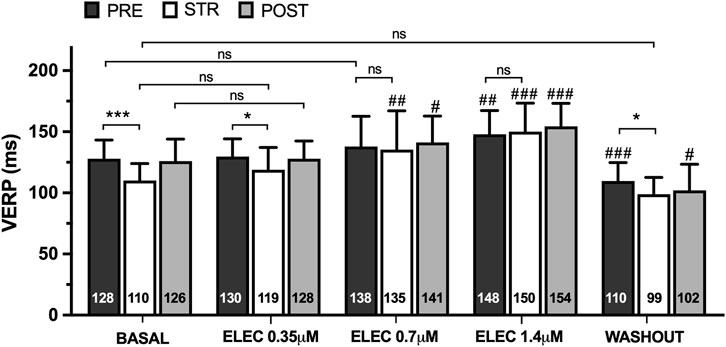
Figure 1. Mean ± standard deviation of ventricular effective refractory period (VERP) obtained at baseline, during the perfusion of eleclazine (ELEC) 0.35 μM, 0.7 μM and 1.4 μM, and after drug washout before (PRE), during (STR) and after (POST) stretch. PRE vs. STR differences: *p < 0.05; ***p < 0.001. Differences vs. baseline: #p < 0.05; ##p < 0.01; ###p < 0.001.
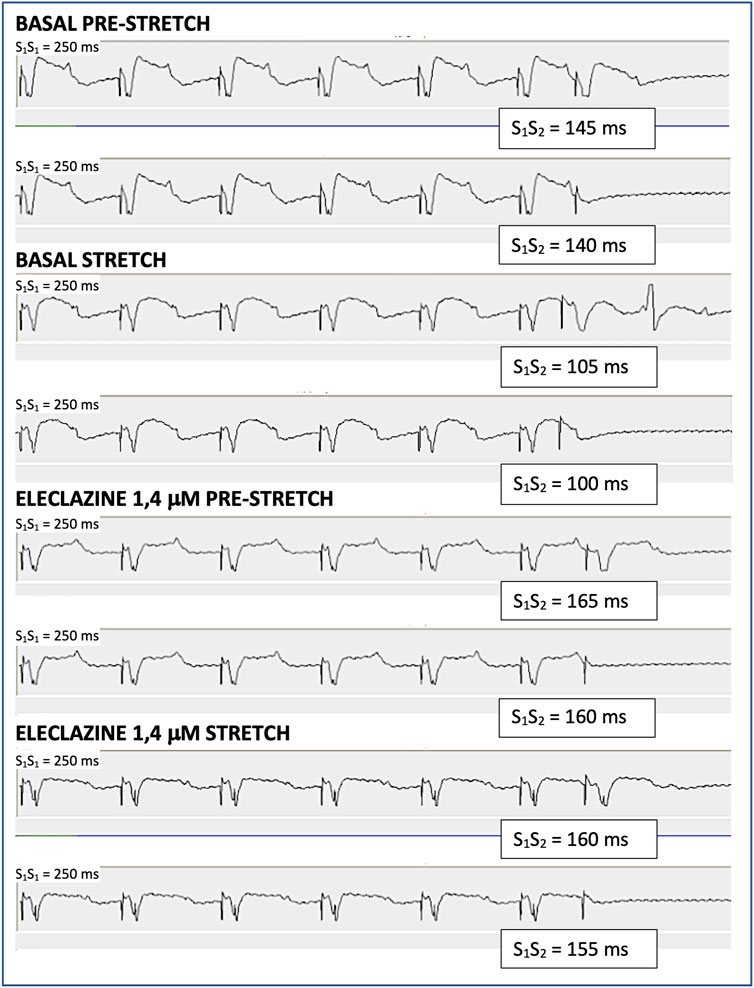
Figure 2. Determination of the ventricular effective refractory period through the extrastimulus test in one of the experiments. S1 are the stimulus of the basic train and S2 is the extrastimulus. The maximum S1S2 interval that does not capture is the refractory period. During baseline myocardial stretch shortens the refractory period by 40 m, while during the perfusion of eleclazine 1.4 μM the refractory period increases and the shortening produced by myocardial stretch is 5 m.
During pacing-induced ventricular fibrillation (VF) (n = 11), myocardial stretch resulted in a significant decrease in successive activation intervals (VV), both at baseline (pre 90 ± 22 vs. stretch 80 ± 16 m, p < 0.05) and during the perfusion of eleclazine 0.35 μM (pre 96 ± 11 vs. stretch 86 ± 12 m, p < 0.01). Once again, however, this reduction did not reach statistical significance at concentrations of 0.7 μM (pre 106 ± 10 vs. stretch 100 ± 10 m, ns) or 1.4 μM (pre 117 ± 20 vs. stretch 104 ± 10 m, ns) (Figure 3). The same figure shows the variations in fifth percentile of the VV intervals (P5) produced by stretch. This parameter is a surrogate indicator of refractoriness during arrhythmia (Chorro et al., 2005; Chorro et al., 2015a), and differences proved significant at baseline (pre 52 ± 13 vs. stretch 44 ± 9 m, p < 0.05) and under the action of eleclazine 0.35 μM (pre 59 ± 12 vs. stretch 49 ± 7 m, p < 0.05) and 0.7 μM (pre 70 ± 13 vs. stretch 58 ± 11 m, p < 0.01), but failed to reach statistical significance at a concentration of 1.4 μM (pre 73 ± 15 vs. stretch 77 ± 15 m, ns). At this concentration, a significant increase in this parameter with respect to the baseline values was recorded in the three situations studied. After drug washout, P5 decreased, and the reducing effects of stretch on this parameter were once again significant (pre 57 ± 18 vs. stretch 44 ± 7 m, p < 0.05).
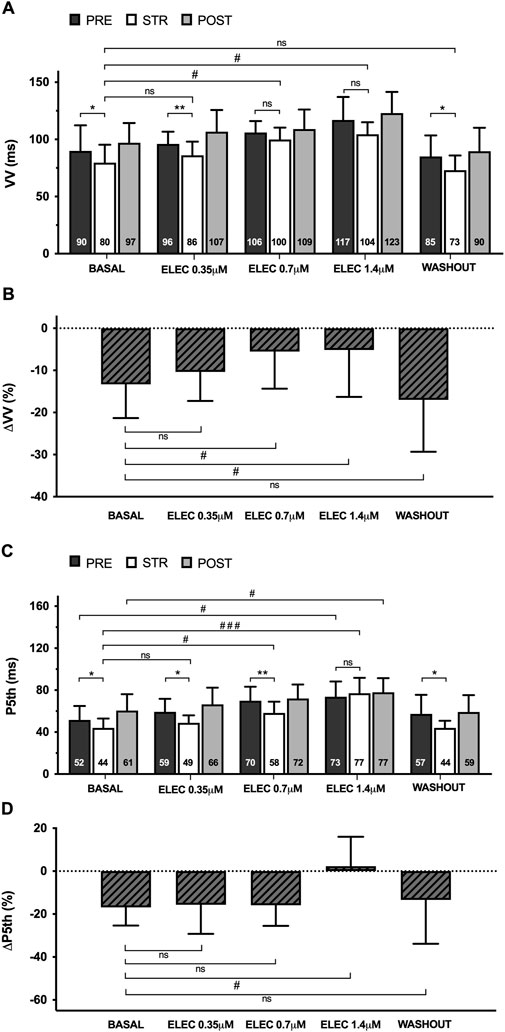
Figure 3. Mean ± standard deviation of the VV (A) and fifth percentile intervals (P5th) (C) during ventricular fibrillation obtained at baseline, during 0.35 μM, 0.7 μM and 1.4 μM eleclazine perfusion (ELEC), and after drug washout before (PRE), during (STR) and after (POST) stretch. The percentage variations of these parameters produced by stretch (ΔVV and ΔP5th) [(B, D) respectively] are also shown. PRE vs. STR differences: *p < 0.05; **p < 0.01. Differences vs. baseline: #p < 0.05; ###p < 0.001.
3.2 VF spectral analysis
Myocardial stretch caused a significant increase in VF dominant frequency (DF) compared to the pre- and post-stretch values, both at baseline (pre 13 ± 3 vs. stretch 17 ± 5 Hz, p < 0.01) and during the perfusion of eleclazine 0.35 μM (pre 13 ± 4 vs. stretch 15 ± 4 m, p < 0.001) and 0.7 μM (pre 12 ± 2 vs. stretch 14 ± 3 m, p < 0.01); the variation was not statistically significant at the 1.4 μM concentration (pre 12 ± 5 vs. stretch 12 ± 3 Hz, ns), however (n = 11) (Figure 4). Under the action of eleclazine (0.7 μM and 1.4 μM), DF was significantly lower than at baseline during stretch. After drug washout, a stretch-related increase in DF was again observed (pre 15 ± 4 vs. stretch 19 ± 3 Hz, p < 0.05). Myocardial stretch also produced a significant decrease in the VF spectral concentration (SpC), both at baseline (pre 31 ± 11 vs. stretch 22 ± 5, p < 0.05) and during 0.35 μM perfusion (pre 30 ± 9 vs. stretch 23 ± 6, p < 0.05), while at higher concentrations the variation did not reach statistical significance (0.7 μM pre 28 ± 7 vs. stretch 25 ± 5, ns; 1.4 μM pre 35 ± 11 vs. stretch 38 ± 12, ns). Spectral concentration during stretch was higher under the action of eleclazine 1.4 μM, and given their direct relationship with SpC, arrhythmia regularity and organization were also greater under the action of the drug.
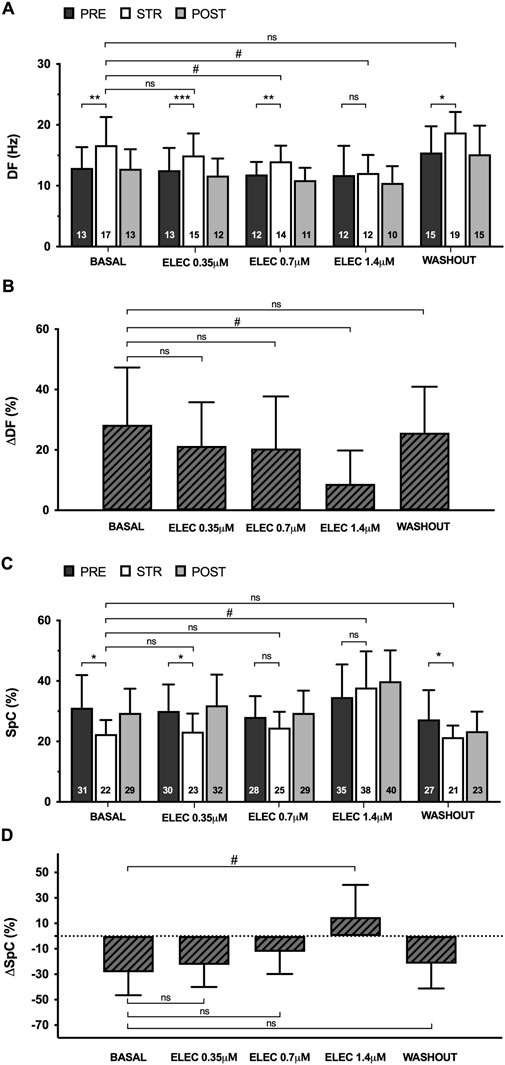
Figure 4. Mean ± standard deviation of dominant frequency (DF) (A) and spectral concentration (SpC) (C) during ventricular fibrillation obtained at baseline, during 0.35 μM, 0.7 μM and 1.4 μM eleclazine perfusion (ELEC) and after drug washout before (PRE), during (STR) and after (POST) stretch. The percentage variations of these parameters produced by stretch (ΔDF and ΔSpC) [(B, D) respectively] are also shown. PRE vs. STR differences: *p < 0.05; **p < 0.01; ***p < 0.001. Differences vs. baseline: #p < 0.05.
3.3 Activation maps during VF
An analysis was made of the VF activation maps obtained at baseline and during perfusion at 1.4 μM - the concentration at which no significant stretch-induced variations in electrophysiological parameters are observed. The stretch-induced increase in complexity of the VF activation patterns observed at baseline was absent under the influence of the drug (n = 11) (Figures 5, 6). In parallel, therefore, the increase in type III maps and the decrease in type I maps observed at baseline during stretch was not found under the action of eleclazine, as reflected by the absence of significant variations in the VF complexity index (Such-Miquel et al., 2013): CI = (number of type I maps x 0.1 + number of type II maps × 1 + number of type III maps × 2)/(number of maps analyzed). Under the action of the drug, the activation maps were simpler than those obtained before, during or following the suppression of myocardial stretch. Complexity was similar to baseline after drug washout, and again the effects of stretch translated into an increase in arrhythmia complexity (baseline: CI pre 1.1 ± 0.2 vs. stretch 1.6 ± 0.1, p < 0.001; 1.4 μM: CI pre 0.7 ± 0.2 vs. stretch 0.6 ± 0.3, ns; and washout: CI pre 1.1 ± 0.2 vs. stretch 1.5 ± 0.2, p < 0.001).
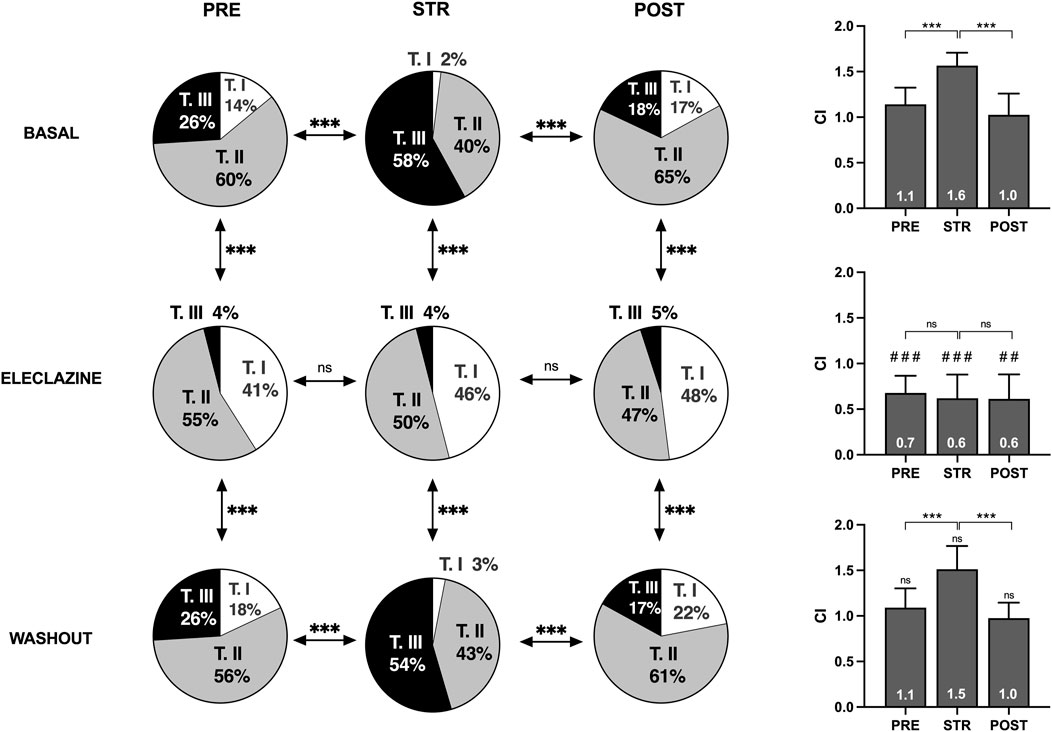
Figure 5. Percentages of activation maps and complexity index (CI) of VF obtained at baseline, during the perfusion of eleclazine 1.4 mM, and after drug washout before (PRE), during (STR) and after (POST) stretch (TI, TII and TIII = types of activation maps classified by complexity). Differences: ***p < 0.001. Differences vs. baseline: ##p < 0.01; ###p < 0.001.
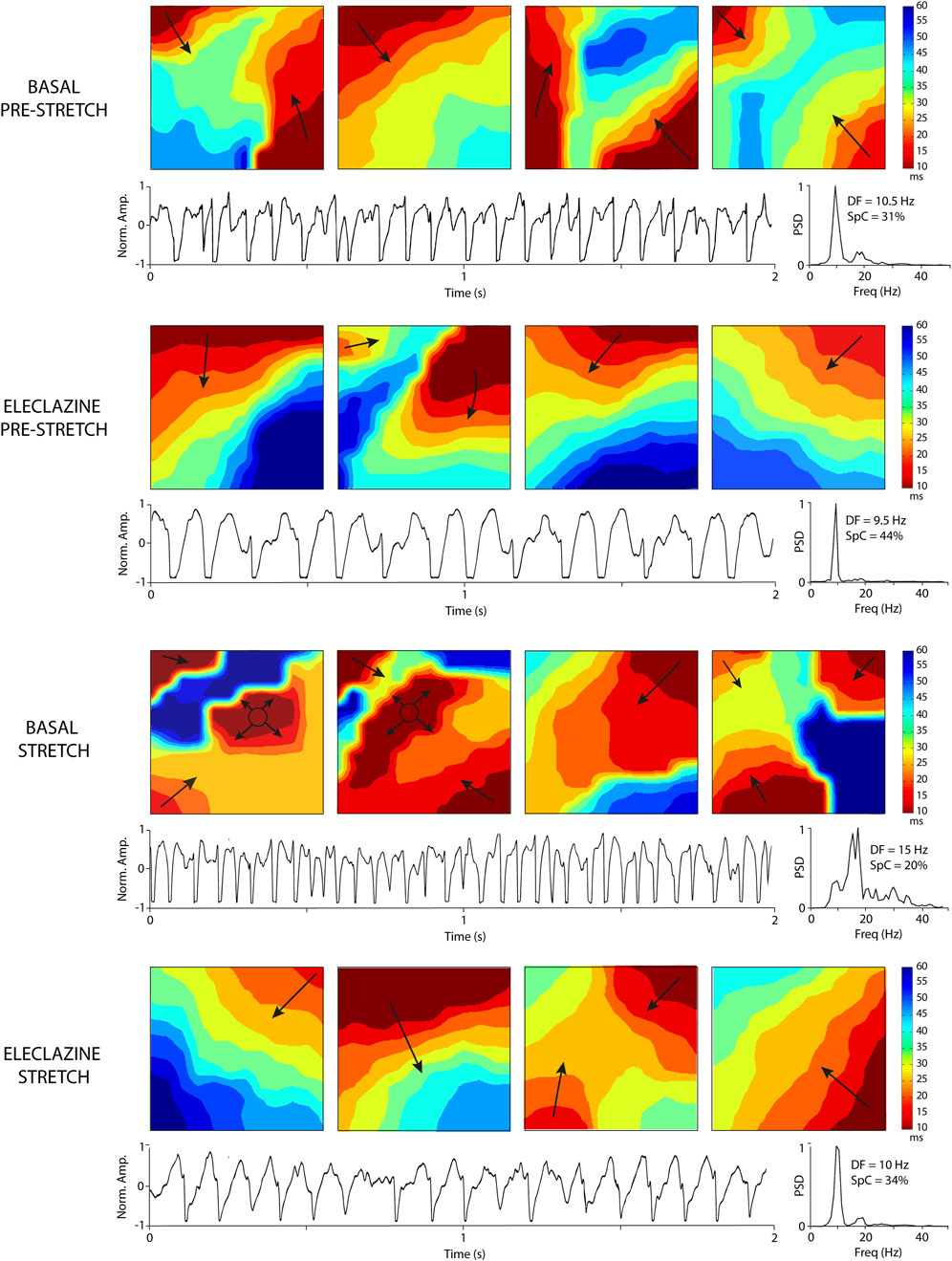
Figure 6. Activation maps and electrograms during VF obtained at baseline and under the perfusion of eleclazine 1.4 mM (before and during stretch). The increase in DF and complexity of the maps produced by stretch is not observed under the action of eleclazine. Abbreviations: DF, dominant frequency; Freq, frequency in hertz; Norm. Amp., normalized amplitude; PSD, Power spectral density; SpC = spectral concentration.
3.4 Determinants of VF complexity
We further analyzed the association between arrhythmia complexity and the electrophysiological parameters obtained during VF. It was found that eleclazine 1.4 μM significantly lowered conduction velocity, while stretch produced no significant variations (Table 1). However, this parameter was not significantly modified under the action of eleclazine when it was determined during ventricular activation at a lower frequency (basic cycle of 250 ms). Correlations between the arrhythmia complexity index and the parameters P5 (r = −0.64), conduction velocity (r = 0.48) and wavelength during VF (r = −0.44) were significant (p < 0.001). The variable entering the equation in the multivariate analysis was P5. During the experimental protocol, VF stopped spontaneously under the action of eleclazine in 6 experiments. Cessation occurred during eleclazine 1.4 μM perfusion in 5 cases and at a 0.7 μM concentration in one case. At baseline, spontaneous cessation of arrhythmia was not observed in any of the cases studied during induced VF (n = 11) (p < 0.05).
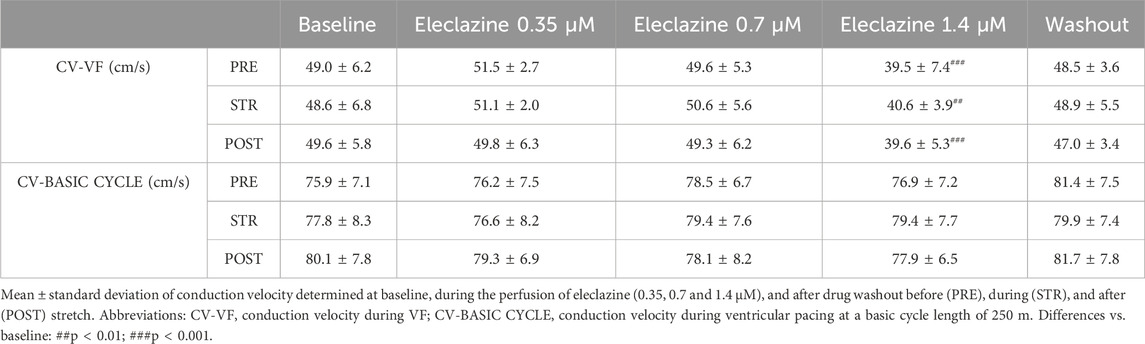
Table 1. Conduction velocity determined at baseline, during the perfusion of eleclazine, and after drug washout.
4 Discussion
The main results of the present study indicate that eleclazine prolongs ventricular refractoriness, shows a frequency-dependent effect on conduction velocity, simplifies the activation patterns and decreases the perpetuation of the arrhythmia. The electrophysiological modifications induced by acute myocardial stretch are attenuated by eleclazine, which avoids the stretch-induced increases in VF activation frequency and complexity related to the stretch-induced shortening of refractoriness (Figure 7).
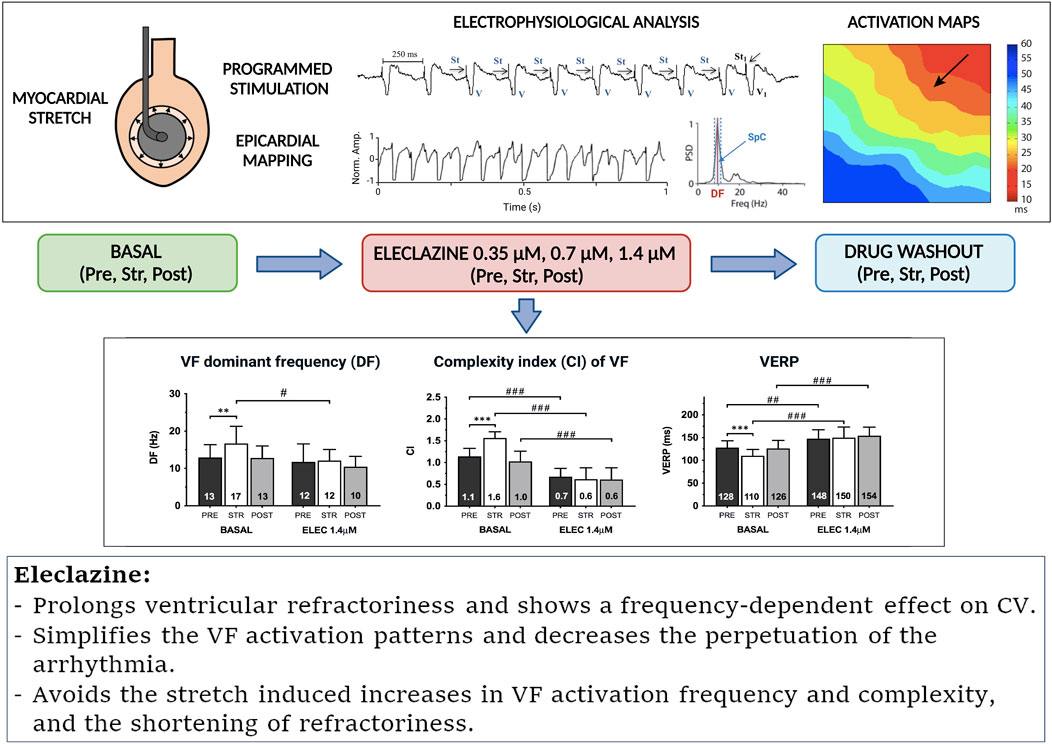
Figure 7. Summary of the methodology used and the effects of eleclazine on the electrophysiological modifications produced by myocardial stretch. Abbreviations: as in previous figures.
4.1 Effects of eleclazine on electrophysiological parameters and VF activation patterns
The effects of the INaL inhibitors ranolazine and GS 967 on basic electrophysiology have been studied in previous reports (Del-Canto et al., 2020; Chorro et al., 2015b; Milberg et al., 2013; Del Canto et al., 2018; Antzelevitch et al., 2004; Potet et al., 2016; Potet et al., 2020). Their actions have been related to the ability to inhibit INaL but also to the effects on the rapid delayed rectifier potassium current IKr and the consequent prolongation of refractoriness (Antzelevitch et al., 2004). On the other hand, the modifications induced in the rapid sodium current have also been implicated in the actions of both ranolazine and the more specific INaL inhibitors GS967 and eleclazine (Burashnikov and Antzelevitch, 2017; Caves et al., 2020; Potet et al., 2016; Potet et al., 2020).
In the present work, eleclazine has shown both a refractoriness-prolonging effect and a conduction velocity-reducing effect during VF. The concentrations used are in a range of the plasma concentrations achieved with therapeutically effective doses (Bacic et al., 2017). Interestingly, the effect on CV has not been observed during stimulation at a cycle length of 250 m and yet it has been significant during VF, when the activation frequency is faster. A use-dependent effect of INaL inhibitors GS-967 and eleclazine has been described in preparations of isolated rat cardiomyocytes (Caves et al., 2020), in heterologously expressed human Nav1.5 (Potet et al., 2016), and in human pluripotent stem-cell derived cardiomyocytes (Potet et al., 2020). Although eleclazine shows a preferential effect for INaL (El-Bizri et al., 2018), it also modifies the rapid component of the sodium current at higher concentrations (Caves et al., 2020; Potet et al., 2020; El-Bizri et al., 2018). In isolated rat myocytes (Caves et al., 2020) the inhibitory effects of eleclazine 10 μM on the rapid component of the sodium current showed a use-dependent effect, and this effect was also shown when using whole rat hearts. A use-dependent block of the rapid INa was also demonstrated in human pluripotent stem-cell derived cardiomyocytes, and the calculated IC50 for the use-dependent block was 0.6 μM at 10 Hz (Potet et al., 2020). The present study has shown a reduction in CV during VF recorded in whole rabbit heart preparations, and significant effects on the perpetuation of the arrhythmia. Both effects are compatible with a use-dependent effect on the rapid component of INa, an effect that at rapid activation frequencies occurs at drug concentrations closer to those at which INaL is inhibited (Burashnikov and Antzelevitch, 2017). The CV reduction in the context of VF has determined that the wavelength of the activation process during the arrhythmia has not been significantly modified during the 1.4 μM concentration despite the concomitant prolongation of ventricular refractoriness. However, the VF activation maps have been simpler and the VF complexity index has decreased, and the multivariate analysis has shown that the main determinant of this simplification of activation maps during the arrhythmia has been the ventricular refractoriness.
4.2 Eleclazine and mechanical cardiac stretch
Myocardial stretch is an arrhythmogenic factor (Quinn and Kohl, 2021; Sutherland, 2017; Johnson and Antoons, 2018; Gottlieb et al., 2023) that has been seen to modify the resting potential, duration of the action potential, myocardial refractoriness, and arrhythmia inducibility in several experimental models (Quinn and Kohl, 2021; Johnson and Antoons, 2018; Ravelli and Allessie, 1997; Chorro et al., 2005; Chorro et al., 2009; Reiter et al., 1988). The myocardial action potential characteristics vary according to transmembrane ionic current modifications (Kistamás et al., 2021; Grandi and Herren, 2014; Hegyi et al., 2018; Wagner et al., 2006), and stretch modifies the balance between inward and outward currents. Stretch increases Na+ and Ca2+ entry into the cardiac myocytes by opening stretch-sensitive channels (both K+ selective and non-cation selective), as well as by modifying the activity of Na+ channels and activating the Na+/Ca2+ exchanger in its reverse mode (Quinn and Kohl, 2021; Prosser et al., 2013; Neves et al., 2016). It modifies Nav1.5 channel function through mechanical action and by increasing reactive oxygen species (ROS) that either directly or with the intervention of protein kinase A and calcium calmodulin kinase II (CaMKII) delay their inactivation (Quinn and Kohl, 2021; Prosser et al., 2013; Ma et al., 2012). CaMKII is also influenced by the increase in Ca2+ and modifies the outflow currents of K+, ICaL, and INaL, contributing to the persistence of INaL along the action potential (Ma et al., 2012; Karagueuzian et al., 2017; Greer-Short et al., 2020; Wei et al., 2017; Shryock et al., 2013).
INaL activity is higher under conditions of oxidative stress or abnormalities in the management of intracellular Ca2+ (Hegyi et al., 2018; Sutanto et al., 2020; Horváth et al., 2023; Horváth et al., 2022; Kiss et al., 2021; Johnson, 2020; Takla et al., 2020; Belardinelli et al., 2015). In turn, the activity of the sarcoplasmic reticulum, its Ca2+ content, and the open probability of Ryanodine channels favoring the spontaneous release of Ca2+, are also dependent on sodium currents and calcium homeostasis (Kistamás et al., 2021; Takla et al., 2020). INaL inhibition (Ma et al., 2012; Karagueuzian et al., 2017) may contribute to a decrease in Na+ and Ca2+ entry, and in turn reduces some of their effects, such as diastolic depolarization, the dispersion of refractoriness, or increased ectopic activity (Kistamás et al., 2021; Karagueuzian et al., 2017; Horváth et al., 2022; Yamazaki et al., 2022). The regulation of the stretch-induced increase in Na + inflow and the activation of the reverse mode of the Na+/Ca2+ exchanger would contribute to attenuate the effects of increased intracellular Ca2+ on CaMKII which in turn phosphorylates the Na + channel and causes an increase in INaL. The reduction in Ca2+ concentration would also influence SERCA and the activation of the Ryanodine receptor, which increases the open probability of this receptor and the Ca2+ release from the sarcoplasmatic reticulum. Similar effects on the stretch-induced modifications of mechanoelectric feedback have been described on modifying Na+ and Ca2+ homeostasis through inhibition of the Na+/Ca2+ exchanger (Chorro et al., 2009) or the Na+/H+ exchanger (Chorro et al., 2015a).
On the other hand, eleclazine also has weak effects on other ion currents, and at a concentration of 14.2 μM it exerts a weak inhibitory effect upon IKr (Rajamani et al., 2016; Caves et al., 2020). As previously commented, eleclazine shows a preferential effect upon INaL (El-Bizri et al., 2018), but also modifies the rapid component of the sodium current especially at high activation frequencies, showing a use-dependent effect (Caves et al., 2020; Potet et al., 2020; El-Bizri et al., 2018). The effects on both components of the Na+ current and on IKr probably play a role in the decreased conduction velocity and increased refractoriness observed under the action of this drug and in the clear attenuation of the stretch-induced effects observed in this study. In this regard, Rajamani et al. (2016) analyzed the magnitude of inhibition of the rapid component of INa by eleclazine 10 μM at two stimulation frequencies in rabbit ventricular myocytes. The inhibitory effect was around 10% at a stimulation frequency of 0.1 Hz and around 20% at a frequency of 3 Hz. They also observed that eleclazine 2 μM, a concentration that is twice the clinical therapeutic concentration, caused less than 10% decrease in the maximum rate of depolarization of ventricular action potential at stimulating frequencies of 60, 180 and 210 bpm. El-Bizri et al. (2018) in human cardiac voltage-gated sodium channels reported an IC50 value of 0.62 ± 0.12 μM to inhibit INaL, with a high selectivity with respect to the effects over the rapid component of INa. Caves et al. (2020) reported an IC50 of 179.9 nM to inhibit the activated INaL in rat ventricular myocytes, and they also observed that eleclazine 10 μM inhibited rapid INa in a use-dependent manner. Potet et al. (2020) reported a reduction of the rapid component of INa in a frequency-dependent manner, and the IC50 determined in human induced pluripotent stem-cell derived cardiomyocytes was 0.6 μM at 10 Hz. They related the use-dependent effect to the high affinity binding to the receptor and a slowed recovery from inactivation of the rapid sodium current. The sodium-channel blocking activity may delay the recovery of excitability prolonging refractoriness. Thus, the inhibition of the rapid component of INa, the reduction of action potential phase 0 slope, the presence of post-repolarization refractoriness and the consequent flattening of the steep initial portion of the restitution curve of refractoriness are factors that may have contributed to simplifying the VF activation patterns.
4.3 Clinical implications
Myocardial fiber stretch and its proarrhythmic action is present in various situations, including regional contraction anomalies secondary to conduction disorders, ischemia or arrhythmias, and mitral prolapse, as well as acute pressure or volume overloads in various clinical contexts (Quinn and Kohl, 2021; Sutherland, 2017; Johnson and Antoons, 2018; Gottlieb et al., 2023; Barber et al., 2020). A reduction in the proarrhythmic effects of stretch via blockade of Na+/Ca2+ and Na+/H+ exchangers or beta-adrenergic receptors has been described in various experimental studies (Chorro et al., 2009; Chorro et al., 2015a). Beneficial effects have also been demonstrated in INaL inhibition with ranolazine in experimental models and in the clinical setting, where atrial and ventricular arrhythmia-reducing effects have been observed (Del-Canto et al., 2020; Chorro et al., 2015b; Younis et al., 2022). However, a reduction in arrhythmic complications has not been demonstrated in clinical studies with eleclazine in patients with Na+ channel alterations such as long QT syndrome type 3 or hypertrophic cardiomyopathy (Shen et al., 2022; Olivotto et al., 2016). The results of the present study show a decrease in the effects of stretch in the experimental model used and suggest a protective role in these settings. Nevertheless, interspecies differences must be taken into account when extrapolating the results obtained to humans. The electrophysiological characteristics of the myocytes of rabbits are closer to those of humans than those of mice and rats. Repolarization involves K+ ionic currents in which expression is similar in the ventricular myocardium of rabbits and humans. On the other hand, the contribution of ICaL and the Na+/Ca2+ exchanger to calcium homeostasis is higher in humans and rabbits than in mice and rats, and also there are differences in the activity of the sarcoplasmic reticulum and the sarcoplasmic reticulum calcium ATPase (SERCA). Differences in calcium handling, action potential restitution and heart size determine a lower susceptibility to electrical alternans phenomena in the experimental models in which mice and rats are used. For these reasons, when considering the clinical implications, it must be kept in mind that experimental models differ from clinical settings. The experimental model used in the present work allows the stretching protocol to be carried out in a reproducible manner and thus study the influence of specific variables. The main objective has been to characterize the modifications of the electrophysiological effects produced by the acute stretching of the myocardium under the action of the drug. This objective is in line with previous studies in which pathophysiological information related to the mechanoelectric feedback has been provided (Chorro et al., 2005; Chorro et al., 2009; Chorro et al., 2015a; Del-Canto et al., 2020; Chorro et al., 2015b; Del Canto et al., 2018).
4.4 Limitations
Interspecies differences must be taken into account when extrapolating the results obtained to the clinical setting. Moreover, the methodology applied in this study involves acute myocardial stretch, and the effects of stretch can give rise to different manifestations in chronic preparations. The effects of stretch in chronic preparations can lead to different manifestations, given the influence of other factors, among them the associated neurohumoral reflexes, the structural changes elicited by the chronic stretch or the electrophysiological remodeling. Neurohumoral reflexes can modulate the direct effects of stretch on the myocardium. On the other hand, the administration of the drug in chronic settings, may add other effects not elicited during the acute administration. In general terms, the acute models facilitate the control of some variables that influence the results, while chronic studies allow us to get closer to the clinical settings.
5 Conclusion
The late sodium current inhibitor eleclazine (GS6615) decreased the proarrhythmic electrophysiological changes induced by myocardial stretch in the experimental model used, and significantly altered activation patterns during VF.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The animal study was approved by the University of Valencia Committee for Animal Care and Use (2015/VSC/PEA/00233 type 2). The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
FC: Conceptualization, Data curation, Formal Analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Validation, Visualization, Writing–original draft, Writing–review and editing. LS-M: Data curation, Formal Analysis, Investigation, Methodology, Visualization, Writing–review and editing. SC: Formal Analysis, Investigation, Methodology, Visualization, Writing–review and editing. OA-M: Formal Analysis, Investigation, Methodology, Software, Supervision, Writing–review and editing. PG: Formal Analysis, Investigation, Methodology, Writing–review and editing. MZ: Formal Analysis, Investigation, Methodology, Software, Validation, Writing–review and editing. AA: Conceptualization, Formal Analysis, Investigation, Methodology, Resources, Supervision, Validation, Writing–review and editing. LS-B: Conceptualization, Investigation, Methodology, Resources, Supervision, Validation, Writing–review and editing. ID: Conceptualization, Data curation, Formal Analysis, Investigation, Methodology, Resources, Software, Supervision, Validation, Visualization, Writing–original draft, Writing–review and editing.
Funding
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This study was supported by the Spanish Society of Cardiology (basic research project 2018); the Carlos III Health Institute (Grant: FIS PI18/01620); and the Generalitat Valenciana (Grant: APOSTD/2018/181).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abbreviations
CaMKII, calcium calmodulin kinase II; CI, ventricular fibrillation complexity index; CV, conduction velocity DF, ventricular fibrillation dominant frequency; IKr, rapidly activating delayed rectifier potassium current; INa, sodium current; INaL, late sodium current; P5, 5th percentile of VV intervals during ventricular fibrillation; ROS, reactive oxygen species; SpC, spectral concentration; VF, ventricular fibrillation.
References
Antzelevitch C., Belardinelli L., Zygmunt A. C., Burashnikov A., Di Diego J. M., Fish J. M., et al. (2004). Electrophysiological effects of ranolazine, a novel antianginal agent with antiarrhythmic properties. Circulation 110, 904–910. doi:10.1161/01.CIR.0000139333.83620.5D
Bacic D., Carneiro J. S., Bento A. A., Nearing B. D., Rajamani S., Belardinelli L., et al. (2017). Eleclazine, an inhibitor of the cardiac late sodium current, is superior to flecainide in suppressing catecholamine-induced ventricular tachycardia and T-wave alternans in an intact porcine model. Heart rhythm. 14, 448–454. doi:10.1016/j.hrthm.2016.10.021
Barber M., Chinitz J., John R. (2020). Arrhythmias from the right ventricular moderator band: diagnosis and management. Arrhythm. Electrophysiol. Rev. 8, 294–299. doi:10.15420/aer.2019.18
Belardinelli L., Giles W. R., Rajamani S., Karagueuzian H. S., Shryock J. C. (2015). Cardiac late Na⁺ current: proarrhythmic effects, roles in long QT syndromes, and pathological relationship to CaMKII and oxidative stress. Heart rhythm. 12, 440–448. doi:10.1016/j.hrthm.2014.11.009
Burashnikov A., Antzelevitch C. (2017). Effectiveness of late INa versus peak INa block in the setting of ventricular fibrillation. Circ. Arrhythm. Electrophysiol. 10, e005111. doi:10.1161/CIRCEP.117.005111
Caves R. E., Carpenter A., Choisy S. C., Clenell B., Cheng H., McNiff C., et al. (2020). Inhibition of voltage-gated Na+ currents by eleclazine in rat atrial and ventricular myocytes. Heart rhythm. 1, 206–214. doi:10.1016/j.hroo.2020.05.006
Chatterjee N. A. (2023). Prediction of sudden cardiac death: looking beyond ejection fraction. Curr. Cardiol. Rep. 25, 525–534. doi:10.1007/s11886-023-01871-0
Chorro F. J., Del Canto I., Brines L., Such-Miquel L., Calvo C., Soler C., et al. (2015a). Experimental study of the effects of EIPA, Losartan and BQ-123 on electrophysiological changes induced by myocardial stretch. Rev. Esp. Cardiol. 68, 1101–1110. doi:10.1016/j.rec.2014.12.023
Chorro F. J., del Canto I., Brines L., Such-Miquel L., Calvo C., Soler C., et al. (2015b). Ranolazine attenuates the electrophysiological effects of myocardial stretch in Langendorff-perfused rabbit hearts. Cardiovasc Drugs Ther. 29, 231–241. doi:10.1007/s10557-015-6587-4
Chorro F. J., Trapero I., Guerrero J., Such L. M., Canoves J., Mainar L., et al. (2005). Modification of ventricular fibrillation activation patterns induced by local stretching. J. Cardiovasc Electrophysiol. 16, 1087–1096. doi:10.1111/j.1540-8167.2005.40810.x
Chorro F. J., Trapero I., Such-Miquel L., Pelechano F., Mainar L., Cánoves J., et al. (2009). Pharmacological modifications of the stretch-induced effects on ventricular fibrillation in perfused rabbit hearts. Am. J. Physiol. Heart Circ. Physiol. 297, H1860–H1869. doi:10.1152/ajpheart.00144.2009
Del Canto I., Santamaría L., Genovés P., Such-Miquel L., Arias-Mutis O., Zarzoso M., et al. (2018). Effects of the inhibition of late sodium current by GS967 on stretch-induced changes in cardiac electrophysiology. Cardiovasc Drugs Ther. 32, 413–425. doi:10.1007/s10557-018-6822-x
Del-Canto I., Gómez-Cid L., Hernández-Romero I., Guillem M. S., Fernández-Santos M. E., Atienza F., et al. (2020). Ranolazine-mediated attenuation of mechanoelectric feedback in atrial myocyte monolayers. Front. Physiol. 11, 922. doi:10.3389/fphys.2020.00922
El-Bizri N., Xie C., Liu L., Limberis J., Krause M., Hirakawa R., et al. (2018). Eleclazine exhibits enhanced selectivity for long QT syndrome type 3-associated late Na+ current. Heart rhythm. 15, 277–286. doi:10.1016/j.hrthm.2017.09.028
Fuller H., Justo F., Nearing B. D., Kahlig K. M., Rajamani S., Belardinelli L., et al. (2016). Eleclazine, a new selective cardiac late sodium current inhibitor, confers concurrent protection against autonomically induced atrial premature beats, repolarization alternans and heterogeneity, and atrial fibrillation in an intact porcine model. Heart rhythm. 13, 1679–1686. doi:10.1016/j.hrthm.2016.04.015
Gottlieb L. A., Coronel R., Dekker L. R. C. (2023). Reduction in atrial and pulmonary vein stretch as a therapeutic target for prevention of atrial fibrillation. Heart rhythm. 20, 291–298. doi:10.1016/j.hrthm.2022.10.009
Grandi E., Herren A. W. (2014). CaMKII-dependent regulation of cardiac Na(+) homeostasis. Front. Pharmacol. 5, 41. doi:10.3389/fphar.2014.00041
Greer-Short A., Musa H., Alsina K. M., Ni L., Word T. A., Reynolds J. O., et al. (2020). Calmodulin kinase II regulates atrial myocyte late sodium current, calcium handling, and atrial arrhythmia. Heart rhythm. 17, 503–511. doi:10.1016/j.hrthm.2019.10.016
Hegyi B., Bányász T., Izu L. T., Belardinelli L., Bers D. M., Chen-Izu Y. (2018). β-adrenergic regulation of late Na+ current during cardiac action potential is mediated by both PKA and CaMKII. J. Mol. Cell Cardiol. 123, 168–179. doi:10.1016/j.yjmcc.2018.09.006
Horváth B., Kovács Z. M., Dienes C., Óvári J., Szentandrássy N., Magyar J., et al. (2023). Conductance changes of Na+ channels during the late Na+ current flowing under action potential voltage clamp conditions in canine, rabbit, and Guinea pig ventricular myocytes. Pharm. (Basel). 16, 560. doi:10.3390/ph16040560
Horváth B., Szentandrássy N., Almássy J., Dienes C., Kovács Z. M., Nánási P. P., et al. (2022). Late sodium current of the heart: where do we stand and where are we going? Pharm. (Basel) 15, 231. doi:10.3390/ph15020231
Jain V., Rao B., Knijnik L., Shah A. D., Lloyd M. S., El-Chami M. F., et al. (2024). Rising burden of cardiac arrest- and heart failure-related mortality in the United States from 1999 to 2020. Heart rhythm. 5, 254–255. doi:10.1016/j.hroo.2024.03.001
Johnson C. N. (2020). Calcium modulation of cardiac sodium channels. J. Physiol. 598, 2835–2846. doi:10.1113/JP277553
Johnson D. M., Antoons G. (2018). Arrhythmogenic mechanisms in heart failure: linking β-adrenergic stimulation, stretch, and calcium. Front. Physiol. 9, 1453. doi:10.3389/fphys.2018.01453
Karagueuzian H. S., Pezhouman A., Angelini M., Olcese R. (2017). Enhanced late Na and Ca currents as effective antiarrhythmic drug targets. Front. Pharmacol. 8, 36. doi:10.3389/fphar.2017.00036
Kiss D., Horváth B., Hézső T., Dienes C., Kovács Z., Topal L., et al. (2021). Late Na+ current is [Ca2+]i-dependent in canine ventricular myocytes. Pharm. (Basel) 14, 1142. doi:10.3390/ph14111142
Kistamás K., Hézsö T., Horváth B., Nánási P. P. (2021). Late sodium current and calcium homeostasis in arrhythmogenesis. Channels (Austin). 15, 1–19. doi:10.1080/19336950.2020.1854986
Lerman B. B., Markowitz S. M., Cheung J. W., Thomas G., Ip J. E. (2024). Ventricular tachycardia due to triggered activity: role of early and delayed afterdepolarizations. JACC Clin. Electrophysiol. 10, 379–401. doi:10.1016/j.jacep.2023.10.033
Liu X., Ren L., Yu S., Li G., He P., Yang Q., et al. (2023). Late sodium current in synergism with Ca2+/calmodulin-dependent protein kinase II contributes to β-adrenergic activation-induced atrial fibrillation. Philos. Trans. R. Soc. Lond B Biol. Sci. 378, 20220163. doi:10.1098/rstb.2022.0163
Lynge T. H., Nielsen J. L., Risgaard B., van der Werf C., Winkel B. G., Tfelt-Hansen J. (2023). Causes of sudden cardiac death according to age and sex in persons aged 1–49 years. Heart rhythm. 20, 61–68. doi:10.1016/j.hrthm.2022.08.036
Lynge T. H., Risgaard B., Banner J., Nielsen J. L., Jespersen T., Stampe N. K., et al. (2021). Nationwide burden of sudden cardiac death: a study of 54,028 deaths in Denmark. Heart rhythm. 18, 1657–1665. doi:10.1016/j.hrthm.2021.05.005
Ma J., Luo A., Wu L., Wan W., Zhang P., Ren Z., et al. (2012). Calmodulin kinase II and protein kinase C mediate the effect of increased intracellular calcium to augment late sodium current in rabbit ventricular myocytes. Am. J. Physiol. Cell Physiol. 302, C1141–C1151. doi:10.1152/ajpcell.00374.2011
Marijon E., Narayanan K., Smith K., Barra S., Basso C., Blom M. T., et al. (2023). The Lancet Commission to reduce the global burden of sudden cardiac death: a call for multidisciplinary action. Lancet 402, 883–936. doi:10.1016/S0140-6736(23)00875-9
Milberg P., Frommeyer G., Ghezelbash S., Rajamani S., Osada N., Razvan R., et al. (2013). Sodium channel block by ranolazine in an experimental model of stretch-related atrial fibrillation: prolongation of interatrial conduction time and increase in post-repolarization refractoriness. Europace 15, 761–769. doi:10.1093/europace/eus399
Neves J. S., Leite-Moreira A. M., Neiva-Sousa M., Almeida-Coelho J., Castro-Ferreira R., Leite-Moreira A. F. (2016). Acute myocardial response to stretch: what we (don’t) know. Front. Physiol. 6, 408. doi:10.3389/fphys.2015.00408
Olivotto I., Hellawell J. L., Farzaneh-Far R., Blair C., Coppini R., Myers J., et al. (2016). Novel approach targeting the complex pathophysiology of hypertrophic cardiomyopathy: the impact of late sodium current Inhibition on exercise capacity in subjects with symptomatic hypertrophic cardiomyopathy (LIBERTY-HCM) Trial. Circ. Heart Fail 9, e002764. doi:10.1161/CIRCHEARTFAILURE.115.002764
Packer M. (2020). What causes sudden death in patients with chronic heart failure and a reduced ejection fraction? Eur. Heart J. 41, 1757–1763. doi:10.1093/eurheartj/ehz553
Potet F., Egecioglu D. E., Burridge P. W., George A. L. (2020). GS-967 and eleclazine block sodium channels in human induced pluripotent stem cell-derived cardiomyocytes. Mol. Pharmacol. 98, 540–547. doi:10.1124/molpharm.120.000048
Potet F., Vanoye C. G., George A. L. (2016). Use-dependent block of human cardiac sodium channels by GS 967. Mol. Pharmacol. 90, 52–60. doi:10.1124/mol.116.103358
Prosser B. L., Khairallah R. J., Ziman A. P., Ward C. W., Lederer W. J. (2013). X-ROS signaling in the heart and skeletal muscle: stretch-dependent local ROS regulates [Ca2⁺]i. J. Mol. Cell Cardiol. 58, 172–181. doi:10.1016/j.yjmcc.2012.11.011
Quinn T. A., Kohl P. (2021). Cardiac Mechano-Electric Coupling: acute effects of mechanical stimulation on heart rate and rhythm. Physiol. Rev. 101, 37–92. doi:10.1152/physrev.00036.2019
Rajamani S., Liu G., El-Bizri N., Guo D., Li C., Chen X. L., et al. (2016). The novel late Na+ current inhibitor, GS-6615 (eleclazine) and its anti-arrhythmic effects in rabbit isolated heart preparations. Br. J. Pharmacol. 173, 3088–3098. doi:10.1111/bph.13563
Ravelli F., Allessie M. A. (1997). Effects of atrial dilatation on refractory period and vulnerability to atrial fibrillation in the isolated Langendorff-perfused rabbit heart. Circulation 96, 1686–1695. doi:10.1161/01.cir.96.5.1686
Reiter M. J., Synhorst D. P., Mann D. E. (1988). Electrophysiological effects of acute ventricular dilatation in the isolated rabbit heart. Circ. Res. 62, 554–562. doi:10.1161/01.res.62.3.554
Shen H., Dong S. Y., Ren M. S., Wang R. (2022). Ventricular arrhythmia and sudden cardiac death in hypertrophic cardiomyopathy: from bench to bedside. Front. Cardiovasc Med. 9, 949294. doi:10.3389/fcvm.2022.949294
Shryock J. C., Song Y., Rajamani S., Antzelevitch C., Belardinelli L. (2013). The arrhythmogenic consequences of increasing late INa in the cardiomyocyte. Cardiovasc Res. 99, 600–611. doi:10.1093/cvr/cvt145
Such-Miquel L., Chorro F. J., Guerrero J., Trapero I., Brines L., Zarzoso M., et al. (2013). Evaluation of the complexity of myocardial activation during ventricular fibrillation. An experimental study. Rev. Esp. Cardiol. 66, 177–184. doi:10.1016/j.rec.2012.08.012
Sutanto H., Lyon A., Lumens J., Schotten U., Dobrev D., Heijman J. (2020). Cardiomyocyte calcium handling in health and disease: insights from in vitro and in silico studies. Prog. Biophys. Mol. Biol. 157, 54–75. doi:10.1016/j.pbiomolbio.2020.02.008
Sutherland G. R. (2017). Sudden cardiac death: the pro-arrhythmic interaction of an acute loading with an underlying substrate. Eur. Heart J. 38, 2986–2994. doi:10.1093/eurheartj/ehw449
Takla M., Huang C. L. H., Jeevaratnam K. (2020). The cardiac CaMKII-Nav1.5 relationship: from physiology to pathology. J. Mol. Cell Cardiol. 139, 190–200. doi:10.1016/j.yjmcc.2019.12.014
Wagner S., Dybkova N., Rasenack E. C. L., Jacobshagen C., Fabritz L., Kirchhof P., et al. (2006). Ca2+/calmodulin-dependent protein kinase II regulates cardiac Na+ channels. J. Clin. Invest 116, 3127–3138. doi:10.1172/JCI26620
Wei X. H., Yu S. D., Ren L., Huang S. H., Yang Q. M., Wang P., et al. (2017). Inhibition of late sodium current suppresses calcium-related ventricular arrhythmias by reducing the phosphorylation of CaMK-II and sodium channel expressions. Sci. Rep. 7, 981. doi:10.1038/s41598-017-01056-0
Yamazaki M., Tomii N., Tsuneyama K., Takanari H., Niwa R., Honjo H., et al. (2022). Rotors anchored by refractory islands drive torsades de pointes in an experimental model of electrical storm. Heart rhythm. 19, 318–329. doi:10.1016/j.hrthm.2021.10.012
Younis A., Goldenberg I., Farooq S., Yavin H., Daubert J., Raitt M., et al. (2022). Reduction in ventricular tachyarrhythmia burden in patients enrolled in the RAID trial. JACC Clin. Electrophysiol. 8, 754–762. doi:10.1016/j.jacep.2022.02.018
Zablocki J. A., Elzein E., Li X., Koltun D. O., Parkhill E. Q., Kobayashi T., et al. (2016). Discovery of dihydrobenzoxazepinone (GS-6615) late sodium current inhibitor (late INai), a phase II agent with demonstrated preclinical anti-ischemic and antiarrhythmic properties. J. Med. Chem. 59, 9005–9017. doi:10.1021/acs.jmedchem.6b00939
Keywords: arrhythmias, myocardial stretch, Eleclazine (GS6615), ventricular fibrillation, cardiac mapping
Citation: Chorro FJ, Such-Miquel L, Cuñat S, Arias-Mutis O, Genovés P, Zarzoso M, Alberola A, Such-Belenguer L and Del Canto I (2025) Effects of Eleclazine (GS6615) on the proarrhythmic electrophysiological changes induced by myocardial stretch. Front. Physiol. 16:1525836. doi: 10.3389/fphys.2025.1525836
Received: 10 November 2024; Accepted: 10 January 2025;
Published: 31 January 2025.
Edited by:
T. Alexander Quinn, Dalhousie University, CanadaReviewed by:
Marie-Louise Ward, The University of Auckland, New ZealandStephan De Waard, University of Bern, Switzerland
Copyright © 2025 Chorro, Such-Miquel, Cuñat, Arias-Mutis, Genovés, Zarzoso, Alberola, Such-Belenguer and Del Canto. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Francisco J. Chorro, RnJhbmNpc2NvLkouQ2hvcnJvQHV2LmVz; Irene Del Canto, aXJlbmUuZGVsY2FudG9AZ21haWwuY29t