 Urszula Malinowska
Urszula Malinowska Jakub Wojciechowski
Jakub Wojciechowski Marek Waligóra2
Marek Waligóra2 Jacek Rogala
Jacek Rogala
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Physiol. , 22 October 2024
Sec. Computational Physiology and Medicine
Volume 15 - 2024 | https://doi.org/10.3389/fphys.2024.1457371
This article is part of the Research Topic Silicon Revolution in Healthcare View all 4 articles
Nowadays studies using Virtual Reality (VR) are gaining high popularity due to VR being a better approximation of the ecological environment for visual experiments than standard 2D display settings. VR technology has been already applied in medicine in the therapy of mental disorders, neurorehabilitation, and neurofeedback. However, its effectiveness compared to the standard 2D procedure is still not fully documented and limited information about the neurophysiological underpinnings of VR is provided. In this study, we tested participants’ performance during several sessions of the computer game in two different environments, VR vs. 2D monitor display. Participants performed three 25 min gaming sessions of adapted Delay Match-To-Sample task during EEG recording. The results showed that the VR group outperformed the 2D display group in the first session and then maintained its performance level throughout the remaining two sessions while the 2D group increased performance in each session eventually leveling up in the last one. Also group differences in the EEG activity were most profound only in the first session. In this session, the VR group was characterized by stronger and more synchronized neuronal activity, especially in delta, theta, and gamma bands. The VR group was less impacted by visual arousals as indicated by the theta/beta2 ratio in parietal electrodes.
Improving the effectiveness of treatments for mental disorders has been and continues to be a key issue in medicine, especially with the significant increase in the prevalence of nervous system dysfunction, not only due to dementia, trauma, or neurodegenerative diseases but also disorders such as Attention deficit hyperactivity disorder (ADHD), which is increasingly prevalent in younger people (Oehrlein et al., 2016). The use of new technological advances may allow for better outcomes while reducing the duration or number of cumbersome therapy sessions. With the rapid development of Virtual Reality (VR) technology, there are more and more reports on research and clinical applications (Wright, 2014; Parsons et al., 2013). Researchers have demonstrated the effectiveness of VR in therapeutic applications, and VR systems have been used in the treatment of many psychiatric disorders, pain management, and neurorehabilitation for several years (Bohil et al., 2011). Studies on brain function in VR environments mostly related to passive watching of 3D videos indicate that the fully immersive 3D-enriched environment requires the allocation of more brain and sensory resources for cognitive/motor control than 2D presentations (Slobounov et al., 2015). In another EEG-VR study (Malik et al., 2015), Malik et al. reported higher EEG absolute power across various brain regions during 3D video watching, especially including the occipital region in the theta band and the frontal and parietal regions in the alpha band, while in fMRI, Chen et al. (2015) observed increased visual fatigue caused by watching 3D TV compared to 2D. Convergent results on increased load in VR settings are also shown by studies using interactive game or simulator environments. For example, Kakkos and colleagues’ findings (Kakkos et al., 2019) demonstrated significant alterations of alpha and theta band power, indicating increased workload (Hogervorst et al., 2014; Mühl et al., 2014; Brouwer et al., 2012) in a VR environment. Therefore, although these studies are valuable, they do not allow for the evaluation of the effectiveness of multisession training protocols. Another issue related to evaluating the effectiveness of cognitive training based on video games is their complexity due to the involvement of multiple cognitive and motor tasks, making it difficult to isolate the impact of the environment on a single task. To address the above issues, we designed an exploratory investigation aiming at a progress comparison of a gamified working memory task in a 2D computer screen and in head-mounted display virtual reality environments during three consecutive training sessions spread over 1 week. We monitored the course of neural, behavioral, and neuronal processing using EEG across the three sessions, including classical spectral and time-frequency analyses, spectral entropy, and neural connectivity estimated by Phase Locking Value. Additionally, we used theta to beta ratio to monitor changes of cognitive load over the sessions. Finally, we monitored behavioral performance using reaction times and accuracy of test performance.
Experiments for both groups were conducted in the same dimly lit room, at room temperature. For the 2D display group we used a standard
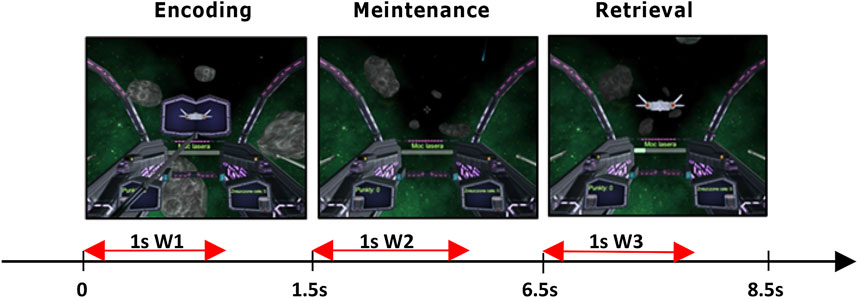
Figure 1. Delayed Match-To-Sample task: 1.5 s encoding phase, next 5 s to maintain the stimulus in working memory, and 2 s of a retrieval phase when participants pressed a button corresponding to the stimulus (left/right ctrl) that matched stimulus presented during encoding. In red are marked 1 s windows selected for EEG analysis.
For the cognitive procedure, we used the Delay Match-To-Sample (DMTS) task (Blough, 1959), which allows for studying working memory and attention in human subjects. We chose DMTS because it involves multiple memory functions, i.e., encoding, maintenance, and retrieval of stimulus representations in sequential order which allow for their individual analyses unlike the n-back task which requires only constant information updating (Ribeiro et al., 2019). In its basic version, the DMTS task comprised three phases: sample, delay, and choice. Each trial starts with the presentation of a sample to be memorized; next a delay was introduced. During the delay, participants were asked to maintain a representation of the sample. When the delay was over another stimulus or multiple stimuli were simultaneously presented to the participant. Participants were then asked to make a decision about does this stimulus match the sample presented in the first phase or does not. Our version of DMTS (Żygierewicz et al., 2022) started with a 2 s familiarization period, where no action from participants was required, followed by the sample encoding phase lasting 1.5 s, and then the animated silhouette of the spaceship was presented to participants. The delay phase–maintenance of the sample in memory, where no stimulus was presented lasted 5 s. During the last, 2 s long retrieval–choice phase, participants were presented with the spacecraft silhouette matching or not the sample. For analyses we chose 1 s window in each of the task phases. In the encoding and retrieval phases the window began at the time of stimulus appearance. In the maintenance phase window began 0.5 s after the end of the stimulus presentation to allow for fade-out of visual processing, which could interfere with memory retention processing readings. Window length was set to 1 s to cover the whole period of stimulus presentation. The duration of each phase was established experimentally during pilot trials to achieve the average performance of 70% accuracy. The screenshots and times for displaying individual trials are given in Figure 1. The entire experiment included 50 experimental trials involving matching and not matching the sample and were randomly intervened with 50 control trials that did not require attention. The times for displaying individual trials are given in Figure 1. To mitigate shooting to all targets, participants earned one point for each correct trial and lost one point for each incorrect one. Subjects from both groups performed the task in three repeated sessions (S1, S2, S3) on different days, 2–3 days apart.
Twenty eight participants (13 women), aged 26–40 years who were not experienced game players were recruited through advertisements at local universities and employment agencies. The participants were randomized into VR and 2D display groups and performed three sessions of the adapted DMTS task during EEG recording. The EEG signal was recorded using Digitrack software (Elmiko l.t.d.) with 21 electrodes arranged in a 10–20 system, referenced to the right ear. All subjects provided informed written consent in accordance with the Helsinki Declaration as well as Institutional Review Board requirements.
EEG signals were recorded at 500 Hz sampling rate. Artifact removal procedure was described in detail in (Żygierewicz et al., 2022). Preprocessing steps contained semi-automated methods, which included 0.5 Hz high-pass and 45 Hz low-pass filtering, baseline correction, exclusion of trials containing muscle artifacts, and independent component analysis (ICA). The identified eye movement and muscle artifacts components were removed. Next, for each channel and trial, we extracted three 1 s long windows (marked on Figure 1 as W1, W2, and W3) in order to investigate EEG signals properties in the encoding, maintenance, and retrieval phases of each task trial. If we set the beginning of the windows in each trial at time 0, the time spans of the three windows will encompass correspondingly: W1 [0, 1000 m], W2 [2000 m, 3000 m], W3 [6500 m, 7500 m]. In the last step, we extracted only epochs with correct responses, which were used in all subsequent analyses. Analysis was performed in the following classical EEG bands: delta (
Signal complexity: Complexity was evaluated by means of Spectral Entropy (SpEn) – theoretical information measure that provided an estimation of EEG regularity. SpEn definition was based on the formulation of Shannon’s entropy (Shannon, 1948), where the probability was replaced by the estimated power of spectral density PSD, Equation 1:
where L was the number of spectral components.
Connectivity was assessed by Phase Locking Value (PLV) – a measure of phase dependence (Lachaux et al., 1999), here estimated between pairs of sites measured in separate frequency bands (defined above
where
Attention and processing capacity: As the measure of cognitive processing capacity we used the
The VR group scored higher in the task compared to the 2D monitor display group, but only in the first game session Figure 2A. The average number of correct answers in the first session in the VR group was 38.1 vs. 34.5 in the 2D monitor group (p = 0.05). In the subsequent sessions, 2D monitor players gradually increased their scores to 37.2 in the second session and 38.5 in the third one, leveling up with results of VR players who maintained their results from the first session in the following ones at a relatively stable level (37.9 in session 2 and 38.2 in session 3). An average reaction time in session one showed a weak trend of faster correct answers for the VR group, but the results did not reach significance (p
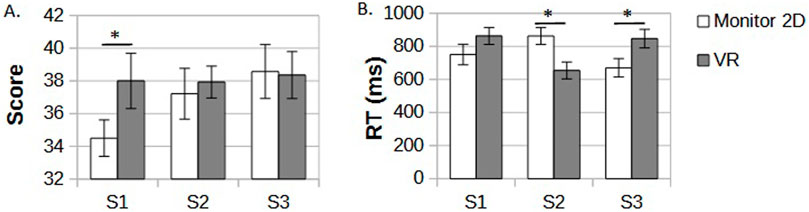
Figure 2. (A) An average number of correct answers in DMTS task in three consecutive sessions (S1-S3) in 2D monitor display and VR groups. (B) plot of average reaction time in the three sessions and separate 2D monitor display and VR groups. Significant differences are marked by black stars.
Most prominent group differences in spectral analysis were found in the first session, in line with most behavioral differences. In the first session during the sample encoding phase, we observed significantly (p
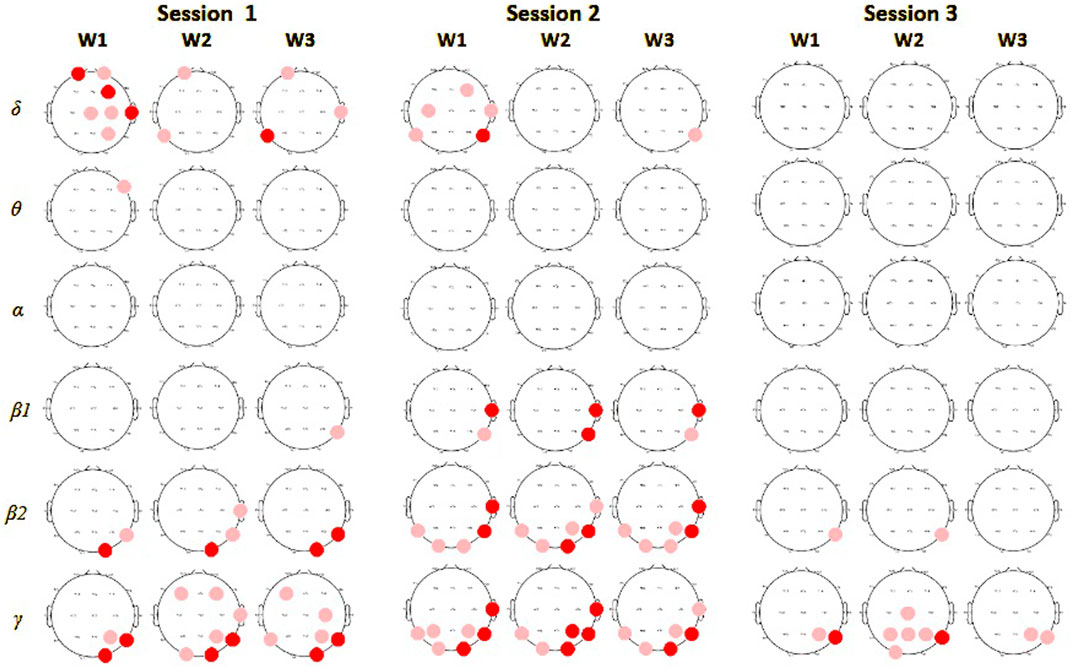
Figure 3. Spectral power comparison between players in monitor display and VR groups in session 1. In columns results for encoding (W1), maintenance (W2), and retrieval phases (W3), in rows for each of the frequency bands. Red circles mark electrodes with significantly higher values of average power in a particular frequency band for subjects in the VR group compared to the monitor display, deep red indicates a significance level at 0.01, and light red at 0.05.
More detailed time-frequency analyses revealed group differences in all EEG bands except for
Interestingly, comparison of EEG spectral changes which took place across sessions within each group revealed opposite patterns. In the
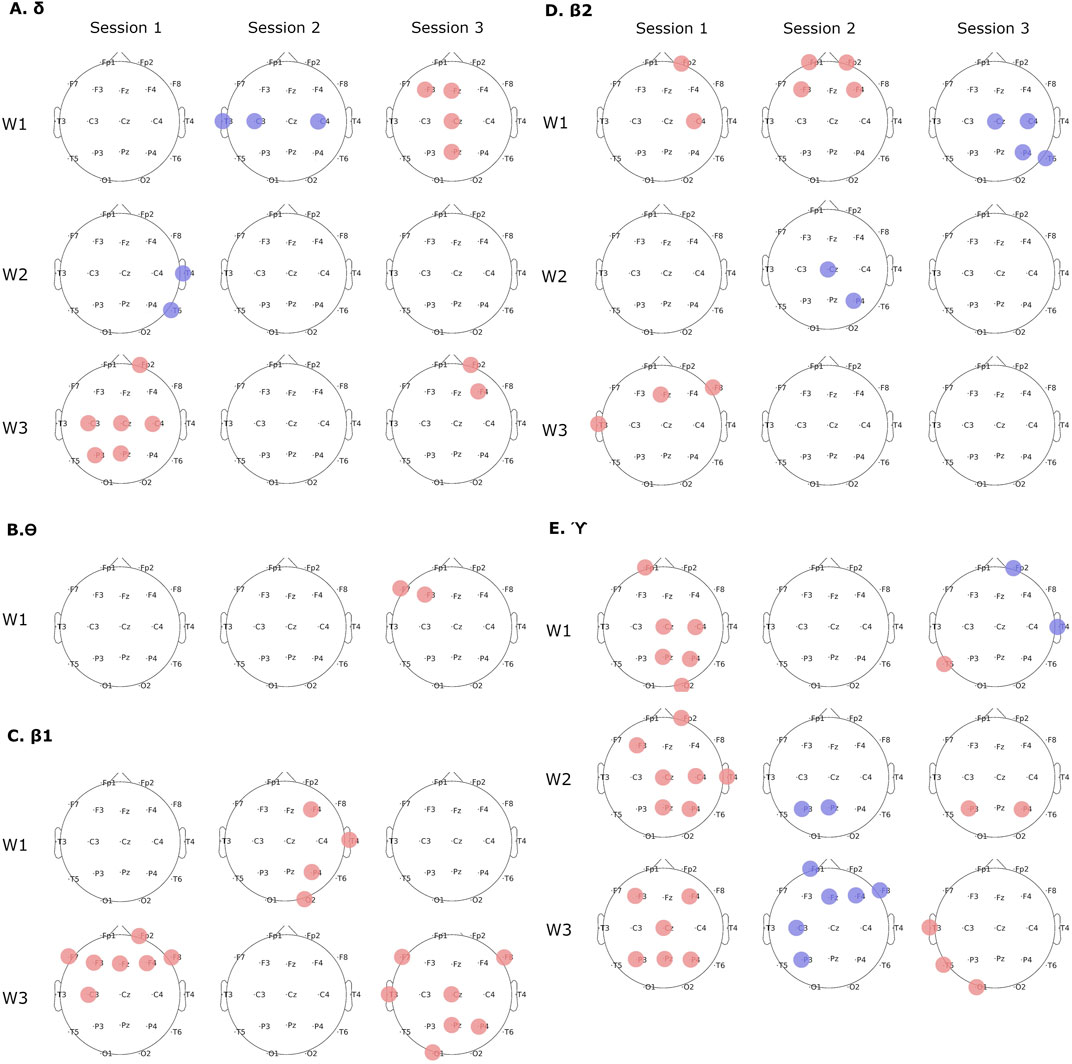
Figure 4. Group differences in time-frequency spectral analyses in subsequent sessions and task phases. (A)
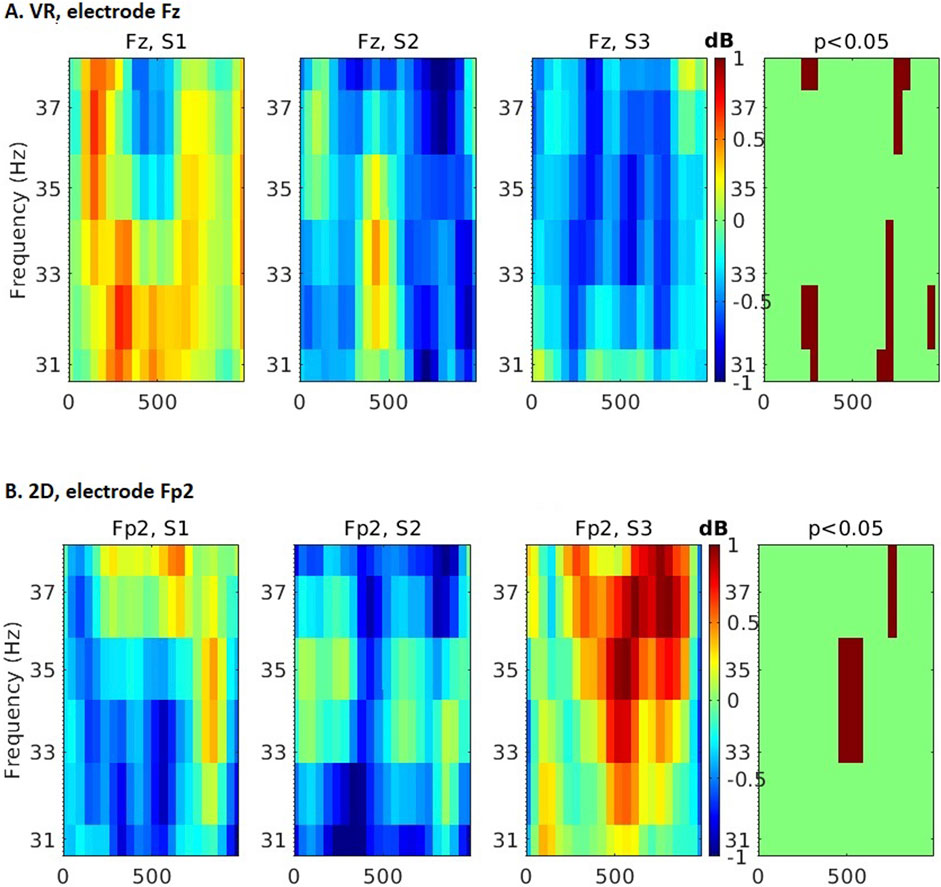
Figure 5. Time-frequency differences across 3 experimental sessions in the
First, we observed smaller EEG signal complexity in the VR group in the first session. Differences between monitor display and VR groups in average spectral entropy from all EEG channels were significant in encoding phase (p = 0.021), but not in maintenance (p = 0.16), and sample matching (p = 0.18) phases during the first session of the DMTS task (Figure 6A). This effect disappeared in sessions 2 (p
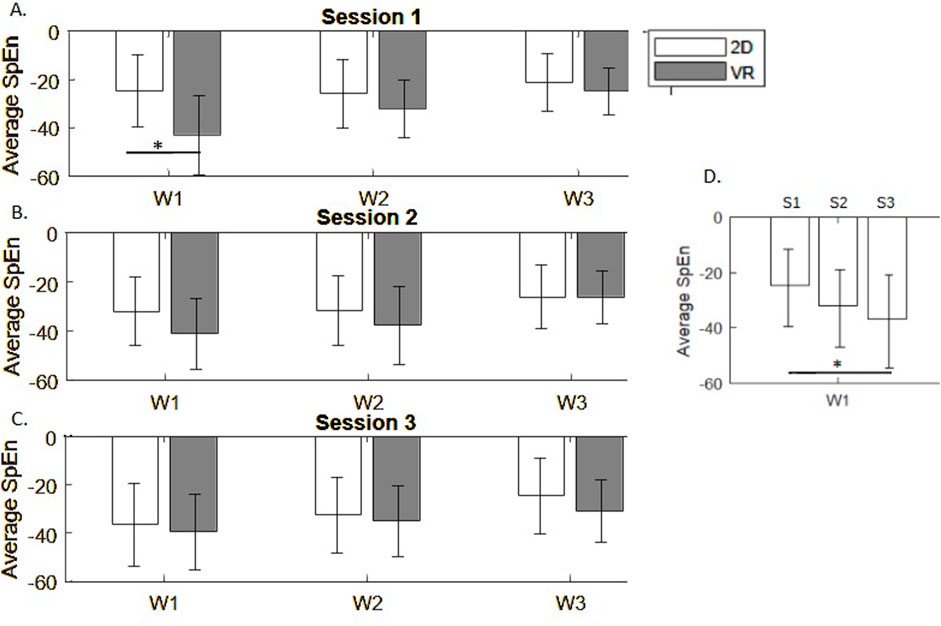
Figure 6. Spectral entropy differences across task phases in 3 experimental sessions (A–C). (D) Average spectral entropy in the monitor display group in repeated sessions S1-S3.
In session 1 the
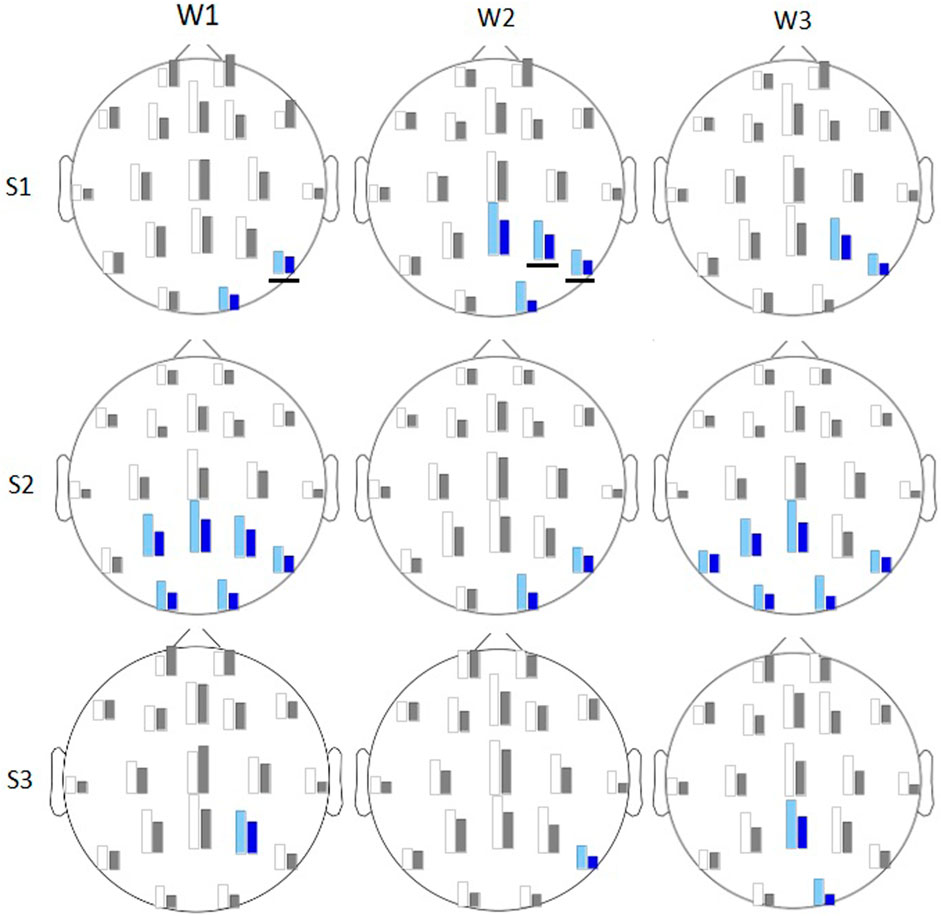
Figure 7. Average
EEG signals interrelations, measured by PLV, revealed in session 1 stronger connectivity in the VR group of subjects compared to the monitor group. That was observed especially in
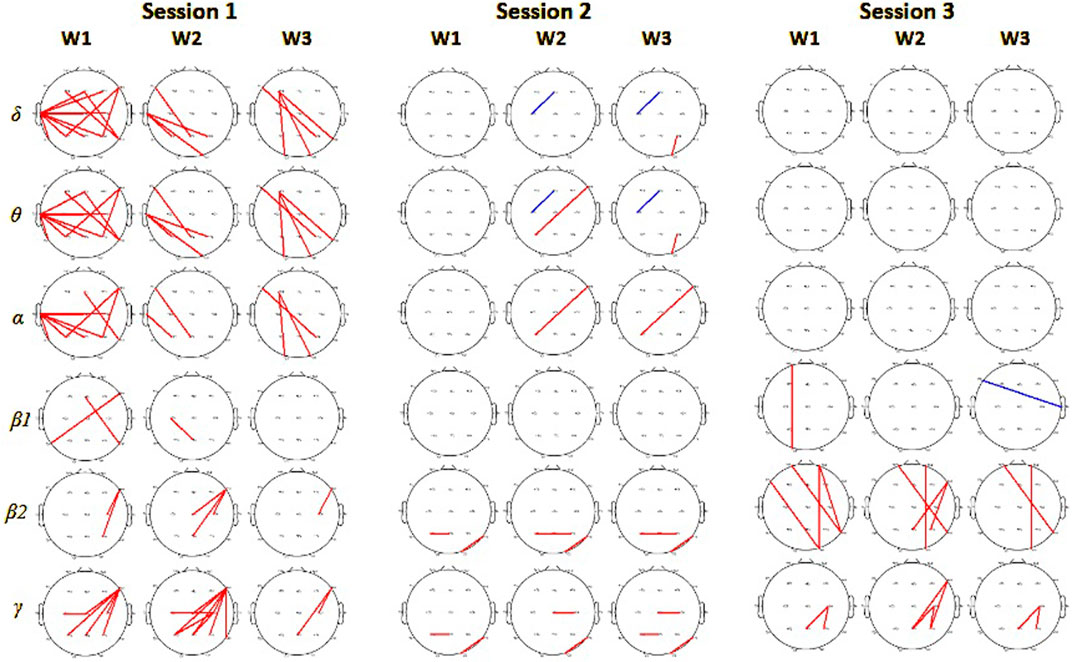
Figure 8. Phase locking value connectivity differences between monitor and VR groups in 3 consecutive sessions. From left for session 1, session 2, and session 3. Red colors mark pairs of electrodes with significantly higher values for subjects in the VR group (on significance level 0.01), and blue for the monitor display group.
Results of our exploratory study comparing 2D monitor display and VR groups in three game sessions showed group differences only in the first session. The VR group members outperformed the monitor display group in the first session and maintained their performance level throughout the remaining two, while the monitor display group increased performance in each session, eventually leveling up with the VR group. The studies comparing behavioral and neurophysiological effects of the VR and computer screen-based tests over multiple training sessions are very rare. One of the very few such investigations is the study performed by Berger and colleagues (Berger et al., 2022), who investigated the effect of neurofeedback training in VR and 2D environments on SMR power. Although their results pointed to a linear increase of the SMR power only in the VR groups, the graphical implementation of the task in both environments was different, therefore one can not exclude that the effect was due to differences in visual stimulus. The study also did not investigate putative differences in cognitive performance. From that perspective more interesting seems to be the experiment conducted by Barrett et al. (2022). In this experiment three groups of participants performed identical categorization tasks also in terms of the visual stimulus implemented in the three environments: VR its computer screen version and the flat version also on a computer screen. The first and second task implementations exactly match the settings of our experiment, however the cognitive task was different. Although this study included only one session, the performance differences in our study were found only in the first session. Interestingly, Barrett et al. (2022) found no behavioral differences between the groups, but the number of fixations in the VR environment was significantly higher than in the 2D one. Also other, previous experiments comparing monitor display and VR environments in a single session showed no benefit of greater immersion experience elicited by VR environment (Pallavicini and Pepe, 2019; Buttussi and Chittaro, 2017; Lugrin et al., 2013). Our seemingly contradictory behavioral performance results may stem from specific game design solely based on a simplified Delay Match-to-Sample task. Games utilizing more selective cognitive functions could be more sensitive to the environmental effects than more complex games, which require more variability and demand on cognitive functions than one function. Another explanation of these contradictions could be possible differences in the interactions between the task at hand and the environment.
To further investigate the effect of the game environment on memory performance we analyzed EEG data. The pattern of behavioral group differences was followed by EEG activity. Analyses of the spectral power differences across sessions revealed two prominent observations: first, higher power of the
Our findings show that the impact of the VR environment may be differentiated for various cognitive functions and the benefits are primarily limited to the learning rate. Therefore, further studies comparing behavioral results and learning rates for different cognitive functions are needed before any clinical applications. Another conclusion that could be drawn from this study is that observed differences in brain activity induced by different task environments do not necessarily result in desired behavioral improvements.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
The studies involving humans were approved by Local Bioethics Committee at Nicolaus Copernicus University in Torun. The studies were conducted in accordance with the local legislation and institutional requirements. The participants provided their written informed consent to participate in this study.
UM: Conceptualization, Formal Analysis, Investigation, Methodology, Software, Visualization, Writing–original draft, Writing–review & editing. JW: Conceptualization, Methodology, Software, Writing–review and editing. MW: Conceptualization, Data curation, Software, Writing–review and editing. JR: Conceptualization, Funding acquisition, Investigation, Methodology, Resources, Supervision, Visualization, Writing–review and editing.
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This work was supported by the National Centre for Research and Development grant POIR 01.01.01-00-178/15.
The authors are grateful to prof. Katarzyna J. Blinowska for helpful discussions on the results and the manuscript.
All authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Alekseichuk I., Turi Z., De Lara G. A., Antal A., Paulus W. (2016). Spatial working memory in humans depends on theta and high gamma synchronization in the prefrontal cortex. Curr. Biol. 26, 1513–1521. doi:10.1016/j.cub.2016.04.035
Barrett R., Poe R., O’Camb J., Woodruff C., Harrison S., Dolguikh K., et al. (2022). Comparing virtual reality, desktop-based 3d, and 2d versions of a category learning experiment. PLoS One 17, e0275119. doi:10.1371/journal.pone.0275119
Berger L. M., Wood G., Kober S. E. (2022). Effects of virtual reality-based feedback on neurofeedback training performance-a sham-controlled study. Front. Hum. Neurosci. 16, 952261. doi:10.3389/fnhum.2022.952261
Blinowska K. J., Kaminski M., Brzezicka A., Kaminski J. (2013). Application of directed transfer function and network formalism for the assessment of functional connectivity in working memory task. Phil. Trans. Roy. Soc. A, 371.
Blough D. S. (1959). Delayed matching in the pigeon. J. Exp. Anal. Behav. 2, 151–160. doi:10.1901/jeab.1959.2-151
Bohil C. J., Alicea B., Biocca F. A. (2011). Virtual reality in neuroscience research and therapy. Psychiatr. Serv. 12, 752–762. doi:10.1038/nrn3122
Brouwer A.-M., Hogervorst M. A., van Erp J. A. F., Heffelaar T., Zimmerman P. H., Oostenveld R. (2012). Estimating workload using eeg spectral power and erps in the n-back task. J. Neural Eng. 9, 045008. doi:10.1088/1741-2560/9/4/045008
Buttussi F., Chittaro L. (2017). Effects of different types of virtual reality display on presence and learning in a safety training scenario. Psychophysiology 24, 1063–1076. doi:10.1109/TVCG.2017.2653117
Chaieb L., Leszczynski M., Axmacher N., Höhne M., Elger C. E., Fell J. (2015). Theta-gamma phase-phase coupling during working memory maintenance in the human hippocampus. Cogn. Neurosci. 6, 149–157. doi:10.1080/17588928.2015.1058254
Chen C., Wang J., Li K., Liu Y., Chen X., Florian S., et al. (2015). Recovering the superficial microvascular pattern via diffuse reflection imaging: phantom validation. Biomed. Eng. Online 14, 87. doi:10.1186/s12938-015-0081-x
Clarke A., Barry R., Karamacoska D., Johnstone S. J. (2019). The EEG Theta/Beta ratio: a marker of arousal or cognitive processing capacity? Appl. Psychophysiol. Biofeedback 44, 123–129. doi:10.1007/s10484-018-09428-6
Delorme A., Makeig S. (2004). EEGLAB: an open source toolbox for analysis of single-trial EEG dynamics including independent component analysis. J. Neurosci. Methods. 134, 9–21. doi:10.1016/j.jneumeth.2003.10.009
Delorme A., Mullen T., Kothe C., Acar Z. A., Bigdely-Shamlo N., Vankov A., et al. (2011). EEGLAB, SIFT, NFT, BCILAB, and ERICA: new tools for advanced EEG processing. Comput. Intell. Neurosci. 2011, 1–12. doi:10.1155/2011/130714
Fernández A., Pinal F., Díaz D., Zurrón M. (2021). Working memory load modulates oscillatory activity and the distribution of fast frequencies across frontal theta phase during working memory maintenance. Neurobiol. Learn. Mem. 183. doi:10.1016/j.nlm.2021.107476
Finley A. J., Angus D., van Reekum C., Davidson R., Schaefer S. (2022). Periodic and aperiodic contributions to theta-beta ratios across adulthood. Psychophysiology 59, e14113. doi:10.1111/psyp.14113
Harmony T., Fernández T., Silva J., Bernal J., Díaz-Comas L., Reyes A., et al. (1996). Eeg delta activity: an indicator of attention to internal processing during performance of mental tasks. Int. J. Psychophysiol. 24, 161–171. doi:10.1016/s0167-8760(96)00053-0
Hogervorst M. A., Brouwer A. M., van Erp J. B. F. (2014). Combining and comparing EEG, peripheral physiology and eye-related measures for the assessment of mental workload. Front. Neurosci. 8, 322. doi:10.3389/fnins.2014.00322
Kakkos I., Dimitrakopoulos G. N., Gao L., Zhang Y., Qi P., Matsopoulos G. K., et al. (2019). Mental workload drives different reorganizations of functional cortical connectivity between 2D and 3D Simulated Flight Experiments. IEEE Trans. Neural. Syst. Rehabi.l Eng. 27, 1704–1713. doi:10.1109/TNSRE.2019.2930082
Kamiński J., Brzezicka A., Wróbel A. (2011). Short-term memory capacity (7 ± 2) predicted by theta to gamma cycle length ratio. Neurobiol. Learn. Mem. 95, 19–23. doi:10.1016/j.nlm.2010.10.001
Ke Y., Chen L., Fu L., Jia P., end Li Y., Zhao X., et al. (2014). Visual attention recognition based on nonlinear dynamical parameters of EEG. Bio-medical Mater. Eng. 24, 349–355. doi:10.3233/BME-130817
Lachaux J.-P., Rodriguez E., Martinerie J., Varela F. J. (1999). Measuring phase synchrony in brain signals. Hum. Brain Mapp. 8, 194–208. doi:10.1002/(sici)1097-0193(1999)8:4<194::aid-hbm4>3.0.co;2-c
Lesenfants D., Francart T. (2020). The interplay of top-down focal attention and the cortical tracking of speech. Sci. Rep. 10, 6922. doi:10.1038/s41598-020-63587-3
Lesenfants D., Habbal D., Chatelle C., Soddu A., Laureys S., Noirhomme Q. (2018). Toward an attention-based diagnostic tool for patients with locked-in syndrome. Clin. EEG Neurosci. 49, 122–135. doi:10.1177/1550059416674842
Lugrin J. L., Cavazza M., Charles F., Le Renard M., Freeman J., Lessiter J. (2013). “Immersive fps games: user experience and performance, 7–12In,” in Proceedings of the 2013 ACM international workshop on immersive media experiences.
Malik A. S., Khairuddin R. N., Amin H. U., Smith M., Kamel N., Abdullah J. M., et al. (2015). EEG based evaluation of stereoscopic 3D displays for viewer discomfort. Biomed. Eng. Online 11, 14–21. doi:10.1186/s12938-015-0006-8
Mathewson K. E., Basak C., Maclin E. L., Low K., Boot W. R., Kramer A. F., et al. (2012). Different slopes for different folks: alpha and delta EEG power predict subsequent video game learning rate and improvements in cognitive control tasks. Exp. Brain Res. 49, 1558–1570. doi:10.1111/j.1469-8986.2012.01474.x
Mühl C., Jeunet C., Lotte F. (2014). EEG-based workload estimation across affective contexts. Front. Neurosci. 8, 114. doi:10.3389/fnins.2014.00114
Oehrlein E. M., Burcu, M. M., Safer D. J., Zito J. M. (2016). National trends in adhd diagnosis and treatment: comparison of youth and adult office-based visits. Psychiatr. Serv. 67, 964—–969. doi:10.1176/appi.ps.201500269
Pallavicini F., Pepe A. (2019). “Comparing player experience in video games played in virtual reality or on desktop displays: immersion, flow, and positive emotions,” in 210In Extended abstracts of the annual symposium on computer-human interaction in play companion extended abstracts, 195.
Parsons T., McPherson S., Interrante V. (2013). Enhancing neurocognitive assessment using immersive virtual reality. Nat. Rev. Neurosci. 35, 27–34. doi:10.1109/vaat.2013.6786190
Pavlov Y. G., Kotchoubey B. (2022). Oscillatory brain activity and maintenance of verbal and visual working memory: a systematic review. Psychophysiology 59, e13735. doi:10.1111/psyp.13735
Ribeiro F. S., Santos F. H., Albuquerque P. B. (2019). How does allocation of emotional stimuli impact working memory tasks? an overview. Adv. Cognitive Psychol. 15, 155–168. doi:10.5709/acp-0265-y
Roux F., Uhlhaas P. (2014). Working memory and neural oscillations: alpha–gamma versus theta–gamma codes for distinct WM information? Trends cognitive Sci. 18, 16–25. doi:10.1016/j.tics.2013.10.010
Sato N., Yamaguchi Y. (2007). Theta synchronization networks emerge during human object–place memory encoding. NeuroReport 18, 419–424. doi:10.1097/WNR.0b013e3280586760
Shannon C. E. (1948). A mathematical theory of communication. Bell. Syst. Tech. J. 27, 623–656. doi:10.1002/j.1538-7305.1948.tb00917.x
Slobounov S. M., Ray W., Johnson B., Slobounov E., Newell K. M. (2015). Modulation of cortical activity in 2D versus 3D virtual reality environments: an eeg study. Int. J. Psychophysiol. 95, 254–260. doi:10.1016/j.ijpsycho.2014.11.003
Summerfield C., Mangels J. A. (2005). Coherent theta-band EEG activity predicts item-context binding during encoding. Neuroimage 24, 692–703. doi:10.1016/j.neuroimage.2004.09.012
Weiss S., Muller H. M., Rappelsberger P. (2000). Theta synchronization predicts efficient memory encoding of concrete and abstract nouns. NeuroReport 11, 2357–2361. doi:10.1097/00001756-200008030-00005
Wright W. G. (2014). Using virtual reality to augment perception, enhance sensorimotor adaptation, and change our minds. Front. Syst. Neurosci. 8, 56–6. doi:10.3389/fnsys.2014.00056
Żygierewicz J., Janik R. A., Podolak I. T., Drozd A., Malinowska U., Poziomska M., et al. (2022). Decoding working memory-related information from repeated psychophysiological EEG experiments using convolutional and contrastive neural networks. J. Neural. Eng. 19, 046053. doi:10.1088/1741-2552/ac8b38
Keywords: virtual reality, EEG, DMTS, cognitive rehabilitation, time-frequency
Citation: Malinowska U, Wojciechowski J, Waligóra M and Rogala J (2024) Performance of game sessions in VR vs standard 2D monitor environment. an EEG study. Front. Physiol. 15:1457371. doi: 10.3389/fphys.2024.1457371
Received: 30 June 2024; Accepted: 04 October 2024;
Published: 22 October 2024.
Edited by:
Toshiyo Tamura, Waseda University, JapanReviewed by:
Seonggwang Yoo, Northwestern University, United StatesCopyright © 2024 Malinowska, Wojciechowski, Waligóra and Rogala. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Urszula Malinowska, dWxhQGZ1dy5lZHUucGw=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.