
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Physiol., 21 June 2024
Sec. Avian Physiology
Volume 15 - 2024 | https://doi.org/10.3389/fphys.2024.1392968
This article is part of the Research TopicNutritional Impacts the Health and Physiology of the Avian Gastro-intestinal TractView all 12 articles
 Elham Atallah1Sabrina Trehiou1Valérie Alquier-Bacquie1Frédéric Lasserre1Julien Arroyo2Caroline Molette2
Elham Atallah1Sabrina Trehiou1Valérie Alquier-Bacquie1Frédéric Lasserre1Julien Arroyo2Caroline Molette2 Hervé Remignon1,3*
Hervé Remignon1,3*Male and female mule ducks were subjected to a force-feeding diet to induce liver steatosis as it is generally done only with male ducks for the production of foie gras. The different biochemical measurements indicated that the course of hepatic steatosis development was present in both sexes and associated with a huge increase in liver weight mainly due to the synthesis and accumulation of lipids in hepatocytes. In livers of male and female ducks, this lipid accumulation was associated with oxidative stress and hypoxia. However, certain specific modifications (kinetics of lipid droplet development and hepatic inflammation) indicate that female ducks may tolerate force-feeding less well, at least at the hepatic level. This is in contradiction with what is generally reported concerning hepatic steatosis induced by dietary disturbances in mammals but could be explained by the very specific conditions imposed by force-feeding. Despite this, force-feeding female ducks seems entirely feasible, provided that the final quality of the product is as good as that of the male ducks, which will remain to be demonstrated in future studies.
Foie gras is a typically renowned French dish and is part of French gastronomy included on the UNESCO’s list of intangible heritage of humanity since 2010. It comes from the force-feeding of waterfowls, mainly ducks. Force-feeding is carried out over a short period (around 10 days) but includes very large quantities of a very caloric meal, almost exclusively composed of corn, which quickly induces hepatic steatosis. Indeed, under these conditions, the liver of birds quickly directs its metabolism toward a very intense production of lipids which are only partially exported and therefore accumulated in the hepatocytes (Heraut et al., 2010; Bax et al., 2012; Bonnefont et al., 2019; Lo et al., 2020). The result of the force-feeding of male ducks is similar, at least at the hepatic level, to what is also described in humans for non-alcoholic fatty liver disease (NAFLD) and/or non-alcoholic steatohepatitis (NASH) (Raza et al., 2021).
For the production of duck foie gras, only male ducks are generally used due to the sexual dimorphism that exists in birds (Brun et al., 2015): male ducks are larger than female ducks and are therefore easier to force-feed. Male ducks are also said to be calmer than female ducks and therefore easier to handle during periods of rearing and force-feeding (Basso et al., 2014). In addition, a significant portion (20%–40%) of female ducks has a liver with a large and superficial venous network, which ultimately makes the product unattractive to consumers (Marie-Etancelin et al., 2015). The origin of this visual defect is not yet documented even if it seems to be of genetic origin and independent of the force-feeding operations themselves.
In recent years, global poultry production has been faced with numerous successive avian influenza crises which have had a significant impact on production and breeding flocks (Spackman, 2020). Among all the consequences of this massive destruction of several million wild and domestic birds, the reduced availability of male ducks for the production of foie gras has sparked interest in the possible substitute use of female ducks. However, until now, no specific information has been published on possible differences in the development of steatotic liver in male or female ducks, while it has been widely documented in mammals of various species (Lornardo et al., 2019; Lefebvre and Staels, 2021).
The aim of this article was therefore to describe separately the evolution of hepatic steatosis in male and female ducks subjected to two different force-feeding programs adapted to their original body sexual dimorphism.
A flock of approximately 1,000, 50% males + 50% females, mule ducks (Caïrina moschata x Anas platyrhynchos) was reared, on the same farm, for 12 weeks, from hatching, according to usual commercial rules. At 12 weeks of age, 12 birds from both sexes (live weights similar to the average live weight of their respective breeding flocks) were randomly selected and slaughtered to constitute the group of ducks (0 meal) before the start of force-feeding. At each stage of sampling, venous livers (exclusively seen in female ducks at a percentage of 20%–30%) were discarded to retain only the non-venous livers from both sexes. The remaining birds were subjected to two different gender-specific force-feeding programs of 21 meals (twice daily for 10 days) using moistened corn flour (97.5% corn) on independent farms and poultry houses. At the start of the force-feeding period, male and female ducks received an amount of 225 g of dry flour/meal, which was gradually, but differently, increased in both sexes to reach a final value of 510 g and 480 g for the last meal in male and female ducks, respectively. In total, during the entire force-feeding period, male and female ducks were forced to ingest different quantities of feed (8.4 kg and 8.0 kg, respectively) depending on their different body sizes. Therefore, this experiment is not a true comparison of the effect of a given force-feeding program on the development of fatty livers in male and female ducks. Rather, it is an analysis of the development of hepatic steatosis in male and female ducks in response to two force-feeding programs best suited to their respective sizes. After 8 and 16 meals of force-feeding, 12 animals of both sexes were randomly selected (in the same manner as described above for ducks at 0 meals) and slaughtered to constitute the 8- (8 meals of force-feeding) and 16-meal groups (16 meals of force-feeding). The remaining birds continued the force-feeding programs for five additional meals. At this time, all the birds were slaughtered, and 24 of them (12 males and 12 females) were selected to constitute the 21-meal group (end of the force-feeding period). At this final stage, because the data provided by the slaughterhouse allowed it, the selection of livers after 21 force-feeding meals was carried out according to the average weights of the livers observed for each sex in the remaining birds (mean ± SD = 566 g ± 93 and 517 g ± 92 for males and females, respectively). At each stage (0, 8, 16, and 21 meals), the ducks were slaughtered 12 h h after the last meal in a commercial slaughterhouse according to its standardized slaughter operations (electronarcosis, bleeding, scalding, and plucking). At the end of the slaughtering line, 20 min post mortem, the livers were harvested and weighed. Then, 50 g of tissue was collected from the median lobe and directly frozen in liquid nitrogen before storage at -80°C. Another piece of the liver was also collected from the same location and stored in paraformaldehyde (4%) buffer for histological observations.
All biochemical measures were performed in duplicate after grinding the tissues in liquid nitrogen.
The dry matter (DM) content was determined by drying the ground liver in an oven at 105°C for 24 h. The total lipid content was measured according to Folch et al. (1957) after extraction with chloroform:methanol (2:1). The total protein content was determined according to the procedure described by the manufacturer (Pierce™ BCA Protein Assay Kit) after an extraction with a phosphate-buffered saline solution. The hydroxyproline (OH-Pro) content was determined according to Woessner (1961) on the delipidated and dry residue obtained after the extraction of the total lipids.
GSH/GSSG analysis: The reduced glutathione/oxidized glutathione (GSH/GSSG) ratio was determined according to the protocol described by the manufacturer (catalog #: 239709, Abcam, Cambridge, United Kingdom).
The activities of the enzymes superoxide dismutase (SOD, catalog #: 19160, Sigma, St Louis, MO, United States) and catalase (Cat, catalog #: KB03012, BioQuoChem, Llanera-Asturias, Spain) were determined according to the procedures described by the manufacturers.
The results are expressed in U/mg of proteins.
The contents of hypoxia-inducible factor 1 alpha (HIF1α) and hypoxia-inducible factor 2 alpha (HIF2α) were determined with ELISA tests on the proteins extracted from livers by using assay kits from MyBioSource (San Diego, CA, United States) according to the manufacturer’s protocols.
Results are expressed in pg/mg of proteins.
Paraformaldehyde-fixed and paraffin-embedded liver tissue sections (3 µm) were stained with hematoxylin and eosin (H&E) for histopathological analysis. The stained liver sections were analyzed (magnification ×100) blindly according to a score ranging from 0 = no visible lipid droplets, 1 = only small lipid droplets, 2 = majority of small lipid droplets, 3 = majority of large lipid droplets, to 4 = almost only large lipid droplets in hepatocytes. Mean score values were obtained from three independent, trained observers.
Total cellular RNA was extracted from liver samples using the TRI reagent (Molecular Research Center Inc., Cincinnati, Ohio, United States). RNA was quantified using a NanoPhotometer (N60, Implen). Total RNA samples (2 µg) were reverse-transcribed using the High-capacity cDNA Reverse Transcription Kit (Applied Biosystems, Foster City, California, United States) for real-time quantitative polymerase chain reaction (qPCR) analyses. Primers were designed in two consecutive exons to avoid amplification of genomic DNA using PrimerQuest™ Tool (Integrated DNA Technologies, Coralville, Iowa, USA), and primers for SYBR Green assays are presented in Supplementary Table S1. Amplifications were performed on the AriaMx Real-time PCR System (Agilent, Santa Clara, California, United States). RT-qPCR data were normalized to the level of the GAPDH (glyceraldehyde-3 phosphate dehydrogenase) messenger RNA (mRNA) and analyzed by LinRegPCR (v2021.2). This program determines the PCR efficiency per sample and accounts for it in a linear regression approach to correct the cycle threshold value for mRNA level quantification. The initial concentration (N0) for each sample is calculated using N0 = threshold/(Effmean x Cq), with Effmean representing the mean PCR efficiency and Cq representing the quantification cycle.
For each analyzed transcript, the value at the beginning of the force-feeding period was set to 1, in each sex, to facilitate the comparisons.
Statistical analyses were performed with SAS software, version 9.4, of the SAS System for Windows. Analysis of variance were performed with the general linear model (Proc GLM) completed with the Student–Newman–Keuls post hoc test to compare the means obtained after each meals in each of the two groups (sexes) independently. Where necessary, to satisfy normality and homoscedasticity conditions, variables were transformed before analysis (Log2 for RT-PCR and OH-Pro analysis). The percentages of the respective scores from the histological analysis were compared independently for each gender according to Fischer’s exact test. Values are expressed as the means ± standard deviation (SD). We set the significant level at p < 0.05.
Throughout the experiment, total mortality (number of animals dead at the end of the rearing–feeding periods/number of animals at the beginning) was less than 1.5% in both sexes, indicating that almost all ducks were able to withstand the rearing conditions imposed. As expected, the force-feeding programs induced a huge increase in liver weight (Table 1). If, at the beginning of the force-feeding period, the weight of the liver of all the birds was approximately 125 g, nevertheless with a slightly higher value (+2.3%) in male ducks, after 21 meals, the weight of the livers increased by 4.51 and 4.23 in male and female ducks, respectively. These values were identical to those expected, at least in male ducks, for which references under close force-feeding conditions are available (Gabarrou et al., 1996, 695 g for 12.5 d of force-feeding; Bax et al., 2012, 660 g for 12 d of force-feeding; and Bonnefont et al., 2019, 600 g for 10 d of force-feeding). In both sexes, lipid levels increased (from 5.0% to 5.3% at meal 0 in males and females, respectively, to 58.3% and 60.5% at meal 21), while protein levels decreased throughout the overfeeding period. The percentage of lipids was then multiplied by more than 11 between 0 and 21 forced meals in both sexes. This indicates a spectacular accumulation of lipids and, therefore, the development of clear hepatic steatosis. This is due to the rapid transformation by the liver of the large quantities of carbohydrates provided by the successive corn meals (containing 62% starch) imposed by the force-feeding programs. It must therefore be concluded that male and female ducks both retain this capacity to transform sugars from the diet into lipids, which accumulate in the liver during the force-feeding period. This ability to accumulate lipids under these particular conditions has already been described in several experiments conducted with male ducks (Herault et al., 2010; Lo et al., 2020; Pioche et al., 2020; Tavernier et al., 2020). The liver’s hydroxyproline (OH-Pro) content reflects its capacity to develop connective tissues, and this indicator has previously been used (Arai et al., 2022; Montefusco et al., 2022) to illustrate the shift from the simple steatosis associated with non-alcoholic fatty liver disease (NAFLD) to non-alcoholic steatohepatitis (NASH). In mice, an increase in the liver’s OH-Pro content was reported by Matsumoto et al. (2013) and Hartimath et al. (2019) and considered a sign of the development of fibrosis characteristic of the NASH condition. In the present experiment, we report a significant increase (+179% and +236%, p < 0.05, in males and females, respectively) in the OH-Pro contents of livers only between 16 and 21 meals of force-feeding. This indicates that only at the very end of the force-feeding period, a fibrogenesis process could take place in both sexes. This is, however, contradictory to what had previously been reported in male ducks by Remignon and Burgue (2023), who had not observed such an increase. However, this last observation was the first published in force-fed ducks, and it is therefore difficult to recognize whether the current results are atypical or not, even if the presence of hepatic inflammation attested by an increase in its fibrogenesis seems rather logical in light of what is generally observed in severe cases of hepatitis steatosis in mammals (Schuppan et al., 2018; Schwabe et al., 2020).
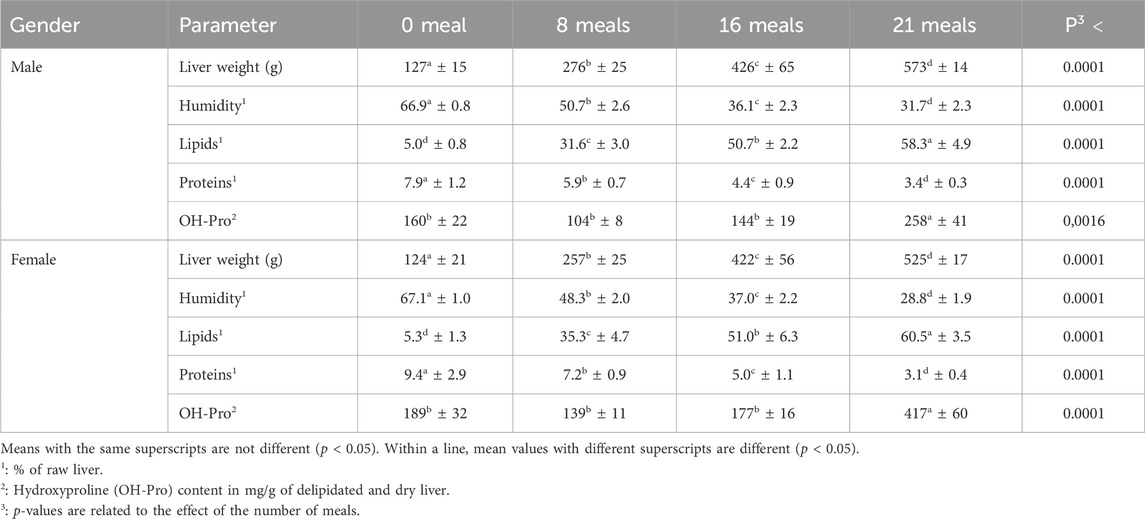
Table 1. Chemical composition of the livers according to the sex of the mule ducks and the number of meals during the force-feeding period (n = 12/sex/stage). Values are the means ± SD.
Histological observations (Figure 1) confirm the biochemical measurements regarding lipid accumulation in hepatocytes. In both sexes, lipids were observed to accumulate in hepatocytes in the form of increasingly present and larger droplets throughout the force-feeding period. However, this accumulation seems to be more gradual and less intense in female ducks. At the final stage (after 21 meals of force-feeding), female ducks still presented a low percentage (10%) of hepatocytes with a majority of small droplets (score 2), while male ducks presented only liver cells with a majority of large lipid droplets (score 3 = 90%) or with almost only large lipid droplets (score 4 = 10%). In mice subjected for long periods to unbalanced diets (high-fat, Western, cafeteria, and fructose-enriched diets), male ducks were also found to be more susceptible to hepatic steatosis and had higher histological scores values than female ducks (Spruss et al., 2012; Gasparin et al., 2018; Smati et al., 2022). According to these authors, the prevention of the development hepatic steatosis development in female ducks could be linked to their particular hormonal status, which could also be the case in female ducks submitted to force-feeding (Tramunt et al., 2021).
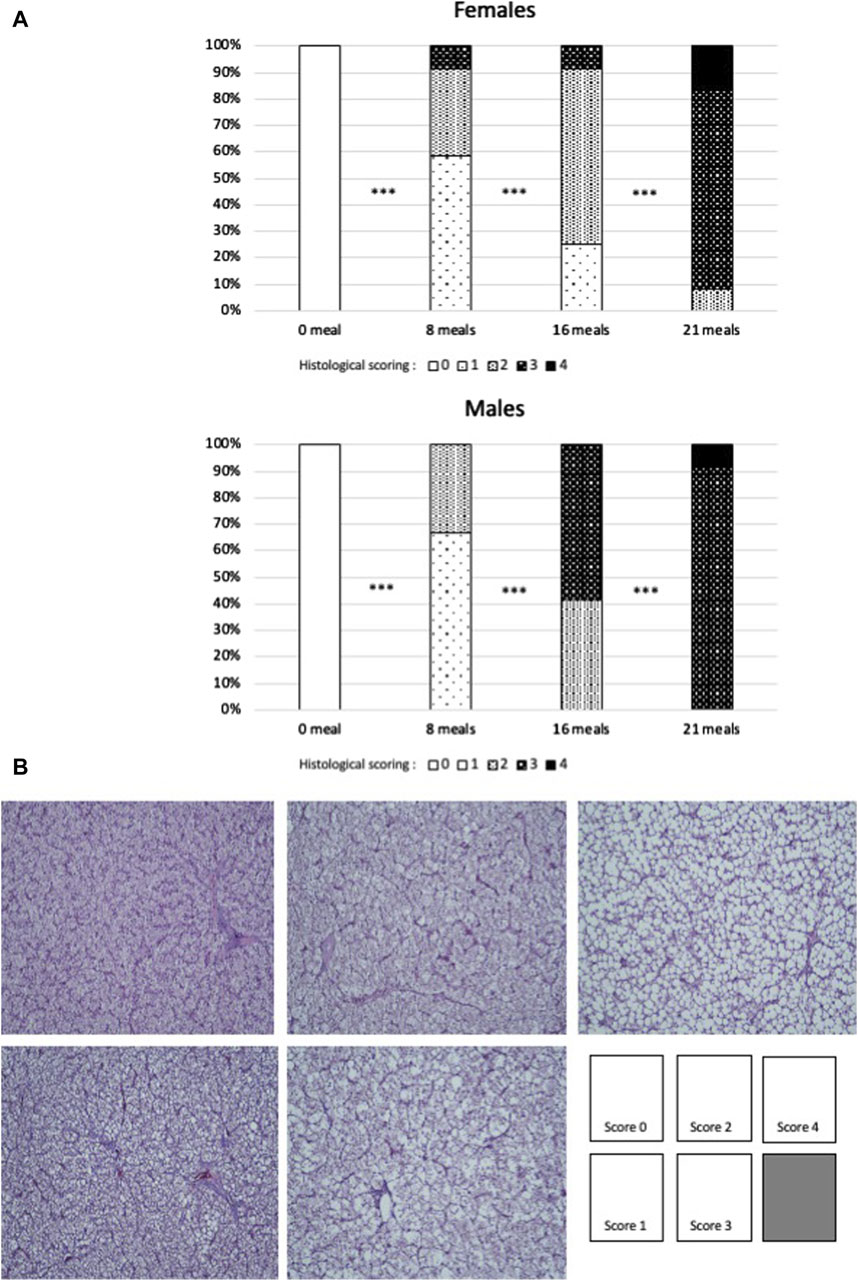
Figure 1. (A) Percentages of each score of histological sections of livers of female or male mule ducks according to the number of meals during the force-feeding period. (n = 12/sex/stage). Significance between two consecutive stages drawn according to Fisher’s exact test (*** = p < 0.001). (B) Examples of scoring grades (hematoxylin–eosin staining), with the same magnification for all pictures.
Corn force-feeding, containing more than 60% starch, induces significant intakes of carbohydrates which mainly arrive in the liver, where they are transformed into lipids. These lipids are initially dedicated to export by lipoproteins (Hermier et al., 2003; Tavernier et al., 2018). However, due to the very large quantities of lipids synthetized, the storage capacity of peripheral tissues, mainly adipose tissues, is quickly exceeded, leading to lipid accumulation in the liver itself. There is therefore an enormous accumulation of complex lipids and free fatty acids in the hepatocytes. This abnormal accumulation of lipids has been described in mammals (Chen et al., 2019) as favoring lipotoxic species such as reactive oxygen species (ROS), which are highly toxic to the cell. However, cells have different mechanisms for neutralizing these ROS, among which the coupling activity of the enzymes superoxide dismutase (SOD) and catalase (Cat) is very effective (Perlemuter et al., 2005) in neutralizing the superoxide ion O2−, one of the most common ROS. In the present study, SOD activities increased rapidly from eight meals and beyond throughout the force-feeding period in both sexes (Figure 2). On the contrary, Cat activities decrease slightly during the same period in both sexes. An increase in SOD and Cat activities has often been described under different conditions (Zeng et al., 2014; Liu et al., 2015; Du et al., 2017; Zhang et al., 2017; Remignon and Burgues, 2023) to be associated to fight against oxidative stress due to the accumulation of lipids in hepatocytes. However, in mice fed a cafeteria diet, Gasparin et al. (2018) reported decreased Cat activity in the liver and indicated that this suggests impaired H2O2 neutralization that could contribute to lipid peroxidation. In the present study, this possible accumulation of H2O2 is highlighted by the decrease in the CAT/SOD ratio (Figure 2) during the force-feeding period in both sexes.
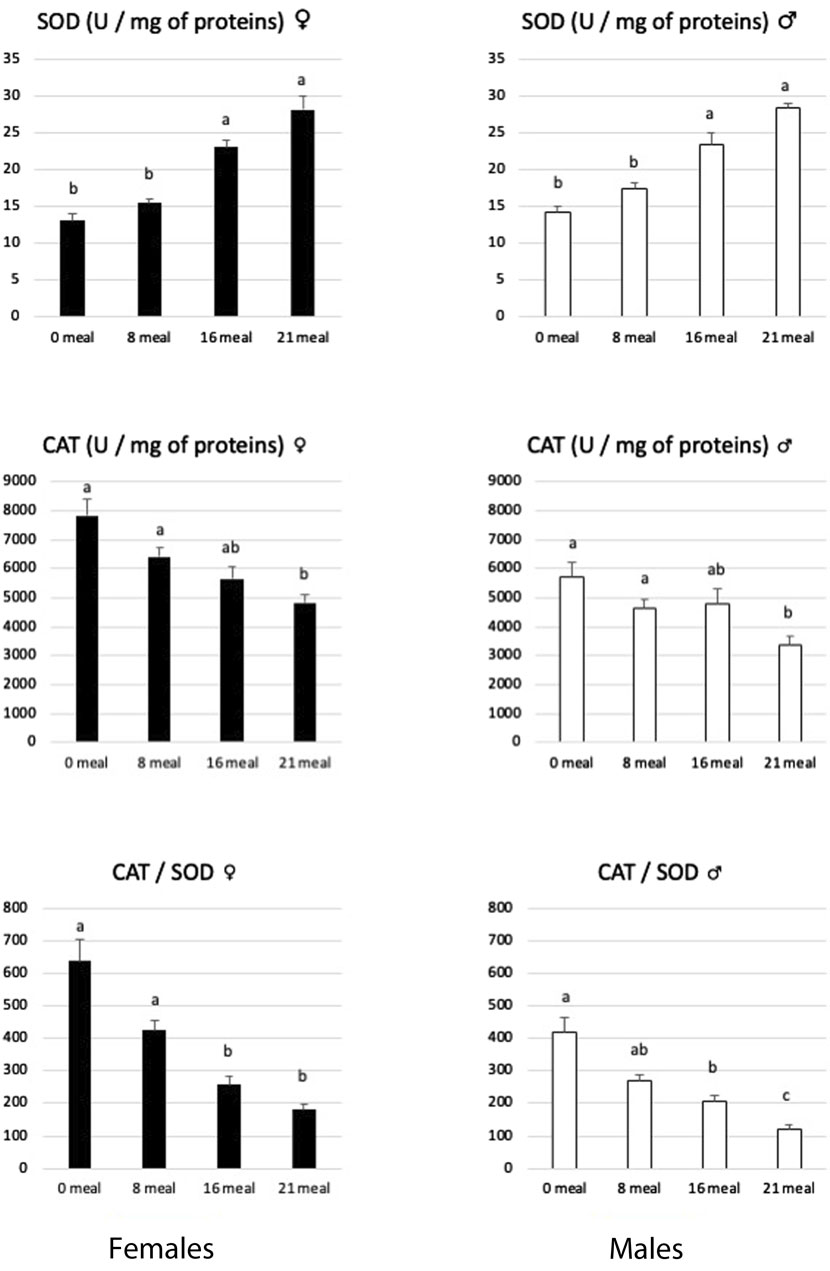
Figure 2. Activities of superoxide dismutase (SOD) and catalase (CAT) enzymes in livers of female and male mule ducks according to the number of meals during the force-feeding period. (n = 12/sex/stage). Values are the means ± SD. Values with different superscripts are different (p < 0.05).
Other enzymatic antioxidant systems may act to catalyze the reduction of H2O2 to H2O, as is primarily the case of the glutathione system (for review, see Engin, 2017). Reduced glutathione (GSH) has been described (Forman et al., 2009) to play a very effective role in maintaining redox homeostasis and, therefore, protects cells from oxidative damage (Sahoo et al., 2017). Associated with the action of specific peroxidases, the coupling activities of reduced and oxidized glutathione (GSSG) represent the most powerful cellular antioxidant system: the decrease in the GSSG/GSH ratio is therefore considered a major sign of cellular dysfunctions (Chen et al., 2013; Alkazemi et al., 2021). In the present study (Figure 3), the levels of GSH and GSSG in the liver decreased during the force-feeding period in both sexes. However, the GSSG/GSH ratio remained constant during this period, indicating that the renewal mechanisms of this powerful antioxidant continue to be functional. When large quantities of lipids are synthetized rapidly in hepatocytes, as observed during the development of hepatic steatosis, this also induces an increase in the activity of the ß-oxidation pathway and consequently a great leak of electrons which are at the origin of ROS (Svegliati-Baroni et al., 2019; Geng et al., 2021). If produced in large quantities, these ROS will clearly induce mitochondrial dysfunctions (Guo et al., 2013). To cope with this, hepatocytes will activate specific antioxidant mechanisms involving SOD, Cat, and glutathione, but if the ROS level remains high, these defense mechanisms against oxidative stress may prove insufficient, as illustrated by their strong reductions observed in severe syndromes of NAFLD or NASH (Sumida et al., 2013). In the present study, while the activity level of SOD increased, to neutralize superoxide ions, during the force-feeding period in both sexes, the activity level of CAT, GSH, and GSSG contents decreased, indicating that the liver cells of male and female ducks have to face uncontrolled oxidative stress during the rapid development of hepatic steatosis.
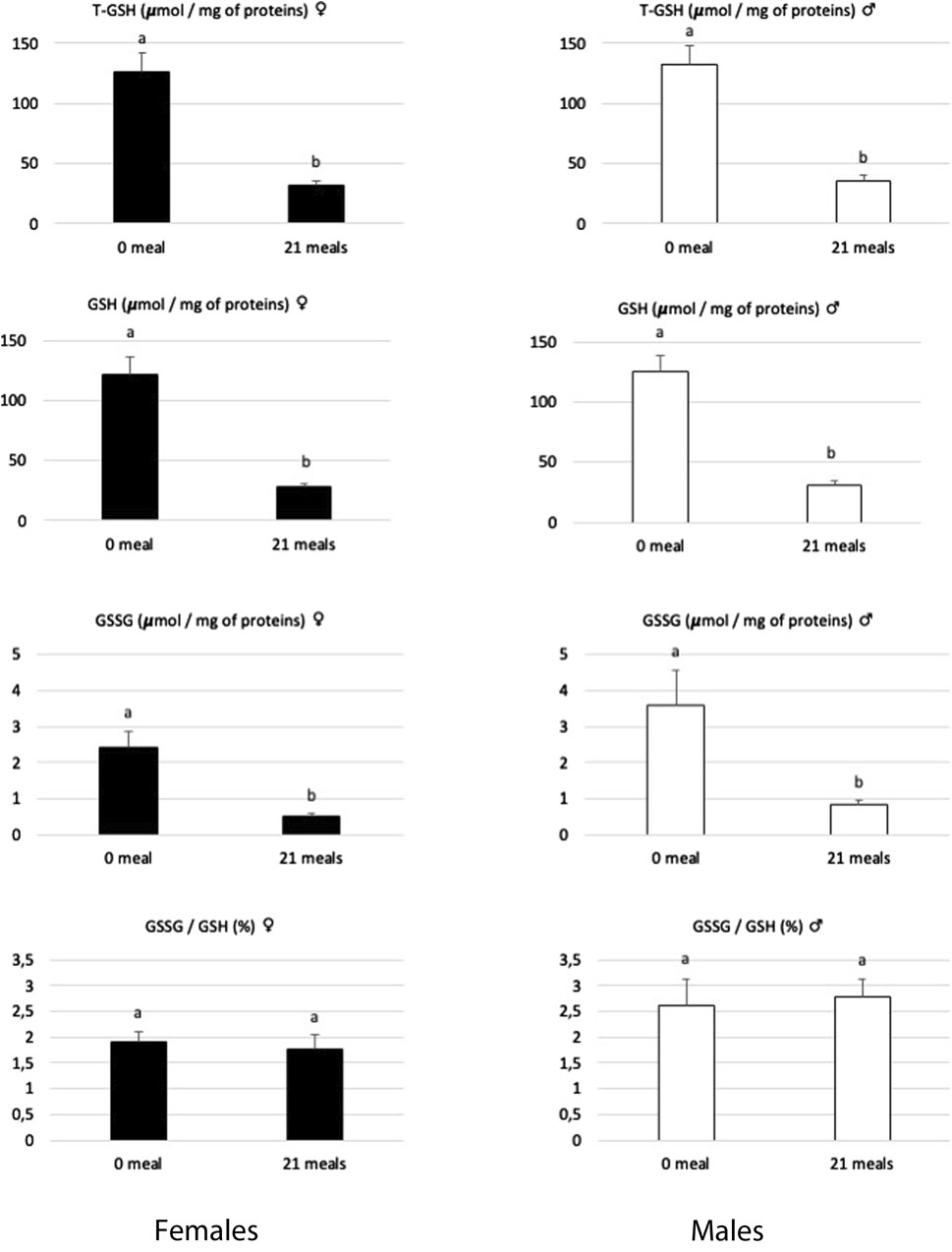
Figure 3. Biochemical contents of total glutathione (T-GSH), reduced glutathione (GSH), oxidized glutathione (GSSG), and the GSSG/GSH ratio in livers of female and male mule ducks before (0 meal, n = 10) and after (21 meals, n = 10 for T-GSH and n = 5 for GSH and GSSG) the force-feeding period. (n = 12/sex/stage). Values are the means ± SD. Values with different superscripts are different (SNK, p < 0.05).
The development of hepatic steatosis impairs metabolism and tissue remodeling (Suzuki et al., 2014), mainly through severe hypoxia that disrupts hepatic oxygen homeostasis. This activates the production of transcription factors HIF1α and HIF2α, which are involved in the response to initial or acute hypoxia, respectively (Downes et al., 2018). For example, Morello et al. (2018) reported that HIF2 is overexpressed in humans or mice developing hepatic steatosis of various origins, which leads to a hypoxic microenvironment and mitochondrial dysfunctions. Remignon and Burgues (2023) previously reported an increase in HIF1α and HIF2α factors in the livers of force-fed male mule ducks. In the present study, the evolution of these two hypoxic factors follows the same pattern in male and female ducks (Figure 4). These increases indicate that during the force-feeding period, the livers of both male and female ducks experienced hypoxic conditions that could be related to physical disability due to liver enlargement and/or circulatory problems related to the elevated lipemia typically associated with the development of fatty liver. According to McGarry et al. (2018), an increase in ROS may result in a reduction in the efficiency of oxygen delivery. This could be progressively the case throughout the force-feeding period for both sexes and thus explain the development of the cellular oxidative stress observed independently of any sexual determinism.

Figure 4. Biochemical contents of hypoxia-inducible factors 1 and 2 (HIF1α and HIF2α) in livers of female and male mule ducks according to the number of meals during the force-feeding period. (n = 12/sex/stage). Values are the means ± SD. Values with different superscripts are different (SNK, p < 0.05).
According to Medzhitov (2008), inflammation should not be considered solely as a reaction to infection or injury but should be expanded to take into account inflammatory processes induced by other types of adverse conditions. The rapid development of steatosis in the liver, as well as the associated oxidative stress, could therefore induce this type of chronic inflammatory state. TNFα (tumor necrosis factor alpha) and IL18 (interleukin-18) are cytokines which have been shown to be increased in nutritionally induced hepatic steatosis in mice or humans (Henao-Meija et al., 2012; Yamanishi et al., 2016; Tiegs and Horst, 2022; Knorr et al., 2023; Vachliotis and Polyzos, 2023). An increase in the transcription levels of the genes coding for these molecules must therefore be considered a sign of the development of an inflammatory process in the liver. In the present experiment, between the beginning (meal 0) and the end (meal 21) of the force-feeding period, we reported an increase in the transcription level of the TNFα gene in females but not in males and increases in that of IL18 for both sexes (Table 2). This indicates that the inflammatory process was present at the end of the force-feeding period in both sexes, assessed by the increase in the level or transcription of the IL18 gene, but potentially with a higher intensity in females, as illustrated by increases in both levels of transcription IL18 and TNFα genes. This differential level of inflammation in male and female mule ducks during the development of liver steatosis induced by force-feeding has been previously approximated with measurements of OH-Pro levels. On the contrary, Spruss et al. (2012) reported that TNFα levels were altered in male and female mice developing NAFLD.
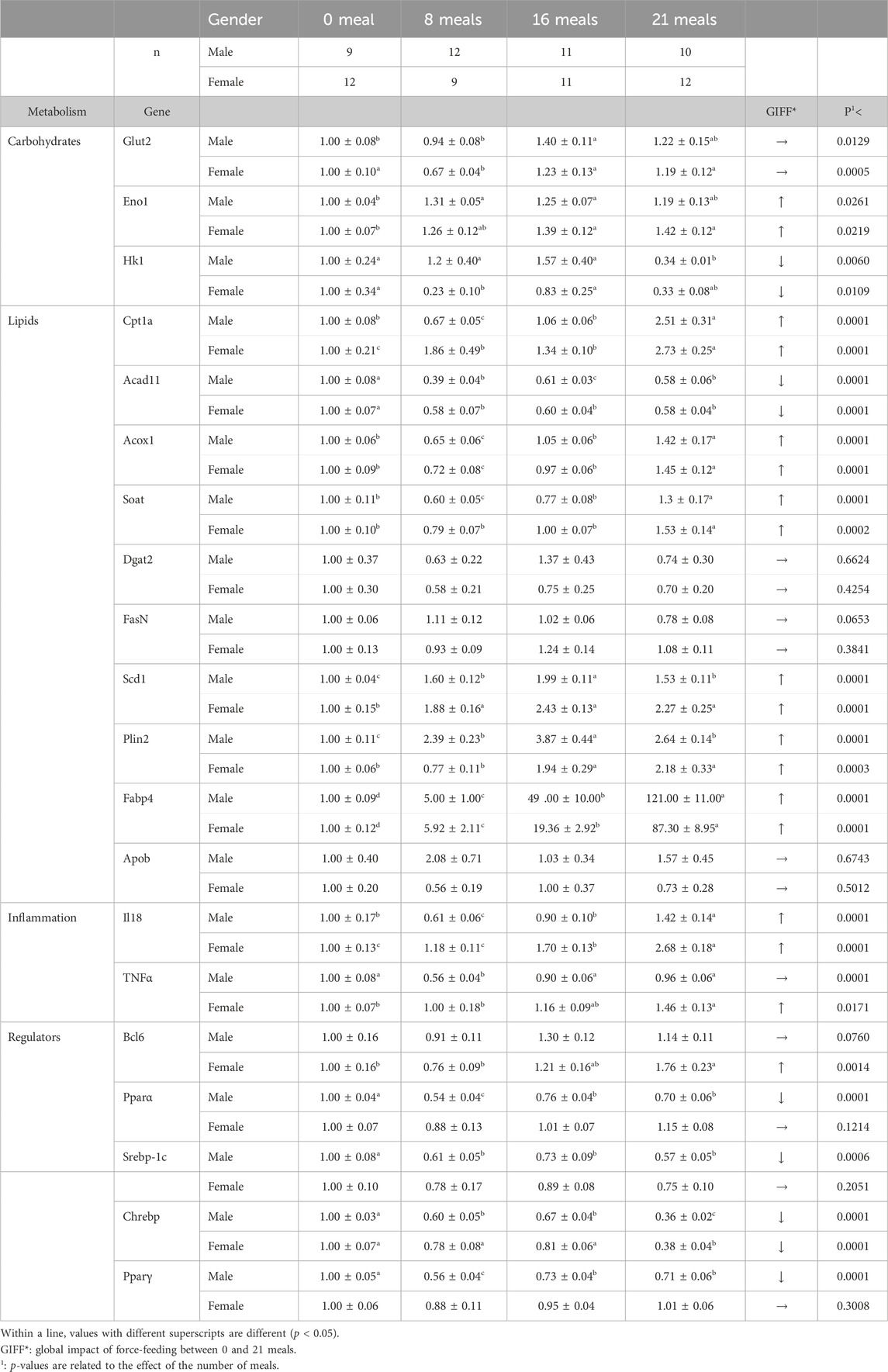
Table 2. Fold-changes (references are mean values for 0 meal) of relative expression of selected genes involved in liver metabolisms of male and female mule ducks during the force-feeding period. Values are the means ± SD.
BCL6 is a transcription factor that plays a key role in determining the active genetic program in male versus female mice and, therefore, their survival under different conditions (Nikkanen et al., 2022). BCL6 is a master transcription factor for the regulation of T follicular helper cells but also acts as a potent antagonist of PPARα-directed gene regulation (Sommars et al., 2019). A high level of hepatic BCL6 in male mice leads to hepatic steatosis and glucose intolerance during dietary excess, while its low level in females may explain better hepatic lipid handling (Salisbury et al., 2021). In the present study, the level of transcription of the BCL6 gene increased only in female ducks during the force-feeding period (Table 2) while it remained constant in males. Since low levels of BCL6 gene transcription in female mice are associated with their increased resistance to the development of hepatic steatosis, we can hypothesize that the observed increase in BCL6 gene transcription in female ducks during the force-feeding period illustrates a decrease in their resistance to hepatic steatosis, which could possibly be linked to their higher inflammatory response. The levels of other transcription factors evaluated (Pparα, Srebp-1c, and Pparγ) decreased in male ducks, while they remained constant in female ducks, during the force-feeding period (Table 2). On the contrary, the transcription level of Chrebp decreased in both sexes. Chrebp (Regnier et al., 2023) and Srebp-1c (Ferré et al., 2021) are involved in lipogenesis pathways when large amounts of glucose are available. In force-fed male ducks, Han et al. (2015) reported that their transcript levels were increased, while we reported them to be downregulated in the present experiment, probably because our birds were fasted for 12 hours before slaughter and, therefore, the level of circulating glucose must have been very low. However, given the enormous amount of lipids accumulated in hepatocytes during force-feeding, this fasting period was not sufficient to increase the level of transcriptional factors such as Pparα or Pparγ, which are generally activated to absorb circulating fatty acids during prolonged periods of fasting (Ruppert and Kersten, 2024).
Other transcript levels regarding carbohydrate or lipid metabolisms in the liver presented changes in male and female ducks during the force-feeding period (Table 2). They are close to previous results (Pioche et al., 2020; Tavernier et al., 2020; Massimino et al., 2021; Andrieux et al., 2023) describing the development of hepatic steatosis induced by force-feeding in male mule ducks. Here, we reported, in male and female mule ducks, that force-feeding influences the transcription of genes (Scd1, Soat, FasN, Dgat2, and Plin2) involved in the neo-synthesis of different types of lipids (fatty acids, polar or neutral lipids, and cholesterol-derived lipids) in response to increasing amounts of carbohydrates delivered. Among the transcripts analyzed and related to lipogenesis, only ACAD11 was significantly decreased during the force-feeding period, as observed by Massimino et al. (2021) in force-fed male mule ducks. For genes related to lipid transport, the relative expression of APOB remained constant, while that of FABP4 was increased significantly during the force-feeding period in both sexes. This has already been observed by Massimino et al. (2021), who proposed two ways to explain these opposing regulations: first, depending on the direction of lipid transport through the liver cells (in or out), their expression can be either increased or decreased by force-feeding; second, the timing of RNA collection after the last meal can have a significant impact on the expression level, as demonstrated by a kinetic study of these same genes by Annabelle et al. (2018). The increase in the level of Fabp4 gene transcripts in the liver is spectacular (121 and 87 times higher in males and females during the entire force-feeding period, respectively), as previously reported in Tavernier et al. (2018) and Pioche et al. (2020). In birds, unlike what is observed in mammals, the liver is the main site of lipid biosynthesis (Cui et al., 2018), and the increase in Fabp4 levels observed during the development of liver steatosis reflects simply the increase in hepatic lipogenesis and transport, as occurring in mammalian adipocytes developing NAFLD (Moreno-Vedia et al., 2022).
Our experience does not reveal large differences in the independent development of hepatic steatosis in response to specific force-feeding programs adapted for male and female mule ducks. The overall evolution of the different parameters measured was visible in both sexes despite the existence of a proven sexual body dimorphism in lean animals prior to force-feeding. However, female birds could have a higher level of liver inflammation at the end of the force-feeding period. In mammals, we know that females are more resistant than males to developing hepatic steatosis in response to unbalanced diets (Spruss et al., 2012; Gasparin et al., 2018; Smati et al., 2022) but which are incommensurate with those caused by force-feeding of palmipeds. It would therefore be bold to conclude that male and female birds differ from mammals on this point as the modifications induced by force-feeding are particular with regard to the nature of the food, the quantities delivered, and the duration of the force-feeding period.
Our study demonstrates that the natural propensity of palmipeds to rapidly develop hepatic steatosis in response to an energy-rich diet, delivered in large quantities for a very short time, is not specific to males, although only those are traditionally used for the production of foie gras in France. It is better to conclude that despite their domestication, mule ducks, whatever their genus, have retained a particular capacity to produce and store lipids in large quantities in different body compartments. This allows them to support the large energy expenditure initially associated with long migratory flights.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
Ethical review and approval were not required for the animal study because we collected samples from a commercial slaughterhouse after the regular operations of anesthesia, bleeding, scalding, plucking, and evisceration. We do not manipulate live animals, and no specific experimental manipulations were performed during the times of rearing. All the samples we harvested were commercial ones, ready to be purchased by consumers.
EA: formal analysis, writing–original draft, methodology, and investigation. ST: writing–original draft, methodology, investigation, and formal analysis. VA-B: writing–original draft, methodology, and formal analysis. FL: writing–original draft, methodology, and formal analysis. JA: writing–review and editing and conceptualization. CM: writing–review and editing and conceptualization. HR: writing–original draft, supervision, methodology, investigation, data curation, writing–review and editing, and conceptualization.
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. N. C4697/INRAE.
The authors would like to thank all the technical staff from Euralis Gastronomie for taking care of the animals during the experiment and for facilitating access to the slaughterhouse for sample collection.
Authors JA and CM were employed by Euralis Gastronomie.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors, and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fphys.2024.1392968/full#supplementary-material
Alkazemi D., Rahman A., Habra B. (2021). Alterations in glutathione redox homeostasis among adolescents with obesity and anemia. Sci. Rep. 11, 3034. doi:10.1038/s41598-021-82579-5
Andrieux C., Marchand M., Larroquet L., Veron V., Biasutti S., Barrieu J., et al. (2023). Fasting/refeeding: an experimental model to study the impact of early thermal manipulation on hepatic metabolism in mule ducks. Am. J. Physiology-Regulatory, Integr. Comp. Physiology 324 (1), R45–R57. doi:10.1152/ajpregu.00158.2022
Annabelle T., Karine R., Marie-Dominique B., Karine G., Stéphane D. (2018). Pre- and post-prandial expression of genes involved in lipid metabolism at the end of the overfeeding period of mule ducks. Mol. Cell. Biochem. 438 (1-2), 111–121. doi:10.1007/s11010-017-3118-6
Arai N., Miura K., Aizawa K., Sekiya M., Nagayama M., Sakamoto H., et al. (2022). Probiotics suppress nonalcoholic steatohepatitis and carcinogenesis progression in hepatocyte-specific PTEN knockout mice. Sci. Rep. 12 (1), 16206. doi:10.1038/s41598-022-20296-3
Basso B., Lague M., Guy G., Ricard E., Marie-Etancelin C. (2014). Detailed analysis of the individual feeding behavior of male and female mule ducks. J. animal Sci. 92, 1639–1646. doi:10.2527/jas.2013-7110
Bax M. L., Chambon C., Marty-Gasset N., Remignon H., Fernandez X., Molette C. (2012). Proteomic profile evolution during steatosis development in ducks. Poult. Sci. 91, 112–120. doi:10.3382/ps.2011-01663
Bonnefont C., Molette C., Lavigne F., Manse H., Bravo C., Lo B., et al. (2019). Evolution of liver fattening and foie gras technological yield during the overfeeding period in mule duck. Poult. Sci. 98 (11), 5724–5733. doi:10.3382/ps/pez359
Brun J.-M., Bernadet M.-D., Cornuez A., Leroux S., Bodin L., Basso B., et al. (2015). Influence of grand-mother diet on offspring performances through the male line in Muscovy duck. BMC Genet. 16 (1), 145. doi:10.1186/s12863-015-0303-z
Chen Y., Dong H., Thompson D., Shertzer H., Nebert D., Vasiliou V. (2013). Glutathione defense mechanism in liver injury: insights from animal models. Food Chem. Toxicol. 60, 38–44. doi:10.1016/j.fct.2013.07.008
Chen Z., Yu Y., Cai J., Li H. (2019). Emerging molecular targets for treatment of nonalcoholic fatty liver disease. Trends Endocrinol. Metabolism 30 (12), 903–914. doi:10.1016/j.tem.2019.08.006
Cui H., Zheng M., Zhao G., Liu R., Wen J. (2018). Identification of differentially expressed genes and pathways for intramuscular fat metabolism between breast and thigh tissues of chickens. BMC Genomics 19 (1), 55. doi:10.1186/s12864-017-4292-3
Downes N. L., Laham-Karam N., Kaikkonen M. U., Ylä-Herttuala S. (2018). Differential but complementary HIF1α and HIF2α transcriptional regulation. Mol. Ther. J. Am. Soc. Gene Ther. 26 (7), 1735–1745. doi:10.1016/j.ymthe.2018.05.004
Du H., Yang J., Bai J., Ming K., Shi J., Yao F., et al. (2017). A flavone-polysaccharide based prescription attenuates the mitochondrial dysfunction induced by duck hepatitis A virus type 1. PloS One 12 (4), e0175495. doi:10.1371/journal.pone.0175495
Engin A. (2017). Non-alcoholic fatty liver disease. Adv. Exp. Med. Biol. 960, 443–467. doi:10.1007/978-3-319-48382-5_19
Ferré P., Phan F., Foufelle F. (2021). SREBP-1c and lipogenesis in the liver: an update1. Biochem. J. 478 (20), 3723–3739. doi:10.1042/BCJ20210071
Folch J., Lee M., Sloane Stanley G. H. (1957). A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 226 (1), 497–509. doi:10.1016/s0021-9258(18)64849-5
Forman H. J., Zhang H., Rinna A. (2009). Glutathione: overview of its protective roles, measurement, and biosynthesis. Mol. aspects Med. 30 (1-2), 1–12. doi:10.1016/j.mam.2008.08.006
Gabarrou J. F., Salichon M. R., Guy G., Blum J. C. (1996). Hybrid ducks overfed with boiled corn develop an acute hepatic steatosis with decreased choline and polyunsaturated fatty acid level in phospholipids. Reproduction, Nutr. Dev. 36, 473–484. doi:10.1051/rnd:19960503
Gasparin F. R. S., Carreño F. O., Mewes J. M., Gilglioni E. H., Pagadigorria C. L. S., Natali M. R. M., et al. (2018). Sex differences in the development of hepatic steatosis in cafeteria diet-induced obesity in young mice. Biochimica Biophysica Acta Mol. Basis Dis. 1864 (7), 2495–2509. doi:10.1016/j.bbadis.2018.04.004
Geng Y., Faber K. N., de Meijer V. E., Blokzijl H., Moshage H. (2021). How does hepatic lipid accumulation lead to lipotoxicity in non-alcoholic fatty liver disease? Hepatol. Int. 15 (1), 21–35. doi:10.1007/s12072-020-10121-2
Guo C., Sun L., Chen X., Zhang D. (2013). Oxidative stress, mitochondrial damage and neurodegenerative diseases. Neural Regen. Res. 8 (21), 2003–2014. doi:10.3969/j.issn.1673-5374.2013.21.009
Han C., Wei S., He F., Liu D., Wan H., Liu H., et al. (2015). The regulation of lipid deposition by insulin in goose liver cells is mediated by the PI3K-AKT-mTOR signaling pathway. PLoS One 10 (5), e0098759. doi:10.1371/journal.pone.0098759
Hartimath S. V., Boominathan R., Soh V., Cheng P., Deng X., Chong Y. C., et al. (2019). Imaging fibrogenesis in a diet-induced model of nonalcoholic steatohepatitis (NASH). Contrast media & Mol. imaging 2019, 6298128. doi:10.1155/2019/6298128
Henao-Mejia J., Elinav E., Jin C., Hao L., Mehal W. Z., Strowig T., et al. (2012). Inflammasome-mediated dysbiosis regulates progression of NAFLD and obesity. Nature 482 (7384), 179–185. doi:10.1038/nature10809
Herault F., Saez G., Robert E., Al Mohammad A., Davail S., Chartrin P., et al. (2010). Liver gene expression in relation to hepatic steatosis and lipid secretion in two duck species. Anim. Genet. 41 (1), 12–20. doi:10.1111/j.1365-2052.2009.01959.x
Hermier D., Guy G., Guillaumin S., Davail S., André J.-M., Hoo-Paris R. (2003). Differential channelling of liver lipids in relation to susceptibility to hepatic steatosis in two species of ducks. Comp. Biochem. Physiology Part B 135, 663–675. doi:10.1016/s1096-4959(03)00146-5
Knorr J., Kaufmann B., Inzaugarat M. E., Holtmann T. M., Geisler L., Hundertmark J., et al. (2023). Interleukin-18 signaling promotes activation of hepatic stellate cells in mouse liver fibrosis. Hepatology 77 (6), 1968–1982. doi:10.1002/hep.32776
Lefebvre P., Staels B. (2021). Hepatic sexual dimorphism - implications for non-alcoholic fatty liver disease. Endocrinology 17 (11), 662–670. doi:10.1038/s41574-021-00538-6
Liu Y., Yuan J. M., Zhang L. S., Zhang Y. R., Cai S. M., Yu J. H., et al. (2015). Effects of tryptophan supplementation on growth performance, antioxidative activity, and meat quality of ducks under high stocking density. Poult. Sci. 94 (8), 1894–1901. doi:10.3382/ps/pev155
Lo B., Marty-Gasset N., Manse H., Bannelier C., Bravo C., Domitile R., et al. (2020). Cellular markers of mule duck livers after force-feeding. Poult. Sci. 99 (7), 3567–3573. doi:10.1016/j.psj.2020.03.048
Lonardo A., Nascimbeni F., Ballestri S., Fairweather D., Win S., Than T. A., et al. (2019). Sex differences in nonalcoholic fatty liver disease: state of the art and identification of research gaps. Hepatol. Baltim.( Md.) 70 (4), 1457–1469. doi:10.1002/hep.30626
Marie-Etancelin C., Retailleau B., Alinier A., Vitezica Z. G. (2015). Sex impact on the quality of fatty liver and its genetic determinism in mule ducks. J. Animal Sci. 93 (9), 4252–4257. doi:10.2527/jas.2015-9121
Massimino W., Andrieux C., Biasutti S., Davail S., Bernadet M. D., Pioche T., et al. (2021). Impacts of embryonic thermal programming on the expression of genes involved in Foie gras production in mule ducks. Front. Physiology 12, 779689. doi:10.3389/fphys.2021.779689
Matsumoto M., Hada N., Sakamaki Y., Uno A., Shiga T., Tanaka C., et al. (2013). An improved mouse model that rapidly develops fibrosis in non-alcoholic steatohepatitis. Int. J. Exp. pathology 94 (2), 93–103. doi:10.1111/iep.12008
McGarry T., Biniecka M., Veale D. J., Fearon U. (2018). Hypoxia, oxidative stress and inflammation. Free Radic. Biol. Med. 125, 15–24. doi:10.1016/j.freeradbiomed.2018.03.042
Medzhitov R. (2008). Origin and physiological roles of inflammation. Nature 454 (7203), 428–435. doi:10.1038/nature07201
Montefusco D., Jamil M., Maczis M. A., Schroeder W., Levi M., Ranjit S., et al. (2022). Sphingosine kinase 1 mediates sexual dimorphism in fibrosis in a mouse model of NASH. Mol. Metab. 62, 101523. doi:10.1016/j.molmet.2022.101523
Morello E., Sutti S., Foglia B., Novo E., Cannito S., Bocca C., et al. (2018). Hypoxia-inducible factor 2α drives nonalcoholic fatty liver progression by triggering hepatocyte release of histidine-rich glycoprotein. Hepatology 67, 2196–2214. doi:10.1002/hep.29754
Moreno-Vedia J., Girona J., Ibarretxe D., Masana L., Rodríguez-Calvo R. (2022). Unveiling the role of the fatty acid binding protein 4 in the metabolic-associated fatty liver disease. Biomedicines 10 (1), 197. PMID: 35052876; PMCID: PMC8773613. doi:10.3390/biomedicines10010197
Nikkanen J., Leong Y. A., Krause W. C., Dermadi D., Maschek J. A., Van Ry T., et al. (2022). An evolutionary trade-off between host immunity and metabolism drives fatty liver in male mice. Science 378 (6617), 290–295. doi:10.1126/science.abn9886
Perlemuter G., Davit-Spraul A., Cosson C., Conti M., Bigorgne A., Paradis V., et al. (2005). Increase in liver antioxidant enzyme activities in non-alcoholic fatty liver disease. Liver Int. 25 (5), 946–953. doi:10.1111/j.1478-3231.2005.01126.x
Pioche T., Skiba F., Bernadet M. D., Seiliez I., Massimino W., Houssier M., et al. (2020). Kinetic study of the expression of genes related to hepatic steatosis, glucose and lipid metabolism, and cellular stress during overfeeding in mule ducks. Am. J. Physiology-Regulatory, Integr. Comp. Physiology 318 (2), R453–R467. doi:10.1152/ajpregu.00198.2019
Raza S., Rajak S., Upadhyay A., Tewari A., Anthony Sinha R. (2021). Current treatment paradigms and emerging therapies for NAFLD/NASH. Front. Biosci. (Landmark Ed.) 26 (2), 206–237. doi:10.2741/4892
Régnier M., Carbinatti T., Parlati L., Benhamed F., Postic C. (2023). The role of ChREBP in carbohydrate sensing and NAFLD development. Nat. Rev. Endocrinol. 19 (6), 336–349. doi:10.1038/s41574-023-00809-4
Remignon H., Burgues P. (2023). Evolution of oxidative stress markers in livers of ducks during force-feeding. Sci. Rep. 13 (1), 1046. doi:10.1038/s41598-022-27271-y
Ruppert P. M. M., Kersten S. (2024). Mechanisms of hepatic fatty acid oxidation and ketogenesis during fasting. Trends Endocrinol. metabolism TEM 35 (2), 107–124. doi:10.1016/j.tem.2023.10.002
Sahoo S., Awasthi J. P., Sunkar R., Panda S. K. (2017). “Determining glutathione levels in plants,” in Plant stress tolerance. Editor R. Sunkar (New York, NY: Springer New York), 273–277. doi:10.1007/978-1-4939-7136-7_16
Salisbury D. A., Casero D., Zhang Z., Wang D., Kim J., Wu X., et al. (2021). Transcriptional regulation of N6-methyladenosine orchestrates sex-dimorphic metabolic traits. Nat. Metab. 3 (7), 940–953. doi:10.1038/s42255-021-00427-2
Schuppan D., Surabattula R., Wang X. Y. (2018). Determinants of fibrosis progression and regression in NASH. J. hepatology 68 (2), 238–250. doi:10.1016/j.jhep.2017.11.012
Schwabe R. F., Tabas I., Pajvani U. B. (2020). Mechanisms of fibrosis development in nonalcoholic steatohepatitis. Gastroenterology 158 (7), 1913–1928. doi:10.1053/j.gastro.2019.11.311
Smati S., Polizzi A., Fougerat A., Ellero-Simatos S., Blum Y., Lippi Y., et al. (2022). Integrative study of diet-induced mouse models of NAFLD identifies PPARα as a sexually dimorphic drug target. Gut 71 (4), 807–821. doi:10.1136/gutjnl-2020-323323
Sommars M. A., Ramachandran K., Senagolage M. D., Futtner C. R., Germain D. M., Allred A. L., et al. (2019). Dynamic repression by BCL6 controls the genome-wide liver response to fasting and steatosis. eLife 8, e43922. doi:10.7554/eLife.43922
Spackman E. (2020). A brief introduction to avian influenza virus. Methods Mol. Biol. Clift.( N.J.) 2123, 83–92. doi:10.1007/978-1-0716-0346-8_7
Spruss A., Henkel J., Kanuri G., Blank D., Püschel G. P., Bischoff S. C., et al. (2012). Female mice are more susceptible to nonalcoholic fatty liver disease: sex-specific regulation of the hepatic AMP-activated protein kinase-plasminogen activator inhibitor 1 cascade, but not the hepatic endotoxin response. Mol. Med. 18 (1), 1346–1355. doi:10.2119/molmed.2012.00223
Sumida Y., Niki E., Naito Y., Yoshikawa T. (2013). Involvement of free radicals and oxidative stress in NAFLD/NASH. Free Radic. Res. 47, 869–880. doi:10.3109/10715762.2013.837577
Suzuki T., Shinjo S., Arai T., Kanai M., Goda N. (2014). Hypoxia and fatty liver. World J. Gastroenterology 20, 15087–15097. doi:10.3748/wjg.v20.i41.15087
Svegliati-Baroni G., Pierantonelli I., Torquato P., Marinelli R., Ferreri C., Chatgilialoglu C., et al. (2019). Lipidomic biomarkers and mechanisms of lipotoxicity in non-alcoholic fatty liver disease. Free Radic. Biol. Med. 144, 293–309. doi:10.1016/j.freeradbiomed.2019.05.029
Tavernier A., Davail S., Houssier M., Bernadet M. D., Ricaud K., Gontier K. (2020). Inter genotype differences in expression of genes involved in glucose metabolism in the establishment of hepatic steatosis in Muscovy, Pekin and mule ducks. Mol. Biol. Rep. 47 (2), 1527–1533. doi:10.1007/s11033-019-05182-y
Tavernier A., Ricaud K., Bernardet M.-D., Gontier K., Davail S. (2018). Pre- and post-prandial expression of genes involved in lipid metabolism at the end of the overfeeding period of mule ducks. Mol. Cell. Biochem. 438 (1-2), 111–121. doi:10.1007/s11010-017-3118-6
Tiegs G., Horst A. K. (2022). TNF in the liver: targeting a central player in inflammation. Seminars Immunopathol. 44 (4), 445–459. doi:10.1007/s00281-022-00910-2
Tramunt B., Montagner A., Tan N. S., Gourdy P., Rémignon H., Wahli W. (2021). Roles of estrogens in the healthy and diseased oviparous vertebrate liver. Metabolites 11 (8), 502. doi:10.3390/metabo11080502
Vachliotis I. D., Polyzos S. A. (2023). The role of tumor Necrosis factor-alpha in the pathogenesis and treatment of nonalcoholic fatty liver disease. Curr. Obes. Rep. 12 (3), 191–206. doi:10.1007/s13679-023-00519-y
Woessner J. F. (1961). The determination of hydroxyproline in tissue and protein samples containing small proportions of this imino acid. Archives Biochem. Biophysics 93, 440–447. doi:10.1016/0003-9861(61)90291-0
Yamanishi K., Maeda S., Kuwahara-Otani S., Watanabe Y., Yoshida M., Ikubo K., et al. (2016). Interleukin-18-deficient mice develop dyslipidemia resulting in nonalcoholic fatty liver disease and steatohepatitis. Transl. Res. 173, 101–114. doi:10.1016/j.trsl.2016.03.010
Zeng T., Li J. J., Wang D. Q., Li G. Q., Wang G. L., Lu L. Z. (2014). Effects of heat stress on antioxidant defense system, inflammatory injury, and heat shock proteins of Muscovy and Pekin ducks: evidence for differential thermal sensitivities. Cell Stress Chaperones 19 (6), 895–901. doi:10.1007/s12192-014-0514-7
Keywords: fatty liver, cellular stress, mule duck, sexual dimorphism, liver steatosis
Citation: Atallah E, Trehiou S, Alquier-Bacquie V, Lasserre F, Arroyo J, Molette C and Remignon H (2024) Development of hepatic steatosis in male and female mule ducks after respective force-feeding programs. Front. Physiol. 15:1392968. doi: 10.3389/fphys.2024.1392968
Received: 28 February 2024; Accepted: 24 May 2024;
Published: 21 June 2024.
Edited by:
Krystyna Pierzchała-Koziec, University of Agriculture in Krakow, PolandReviewed by:
Felix Kwame Amevor, Sichuan Agricultural University, ChinaCopyright © 2024 Atallah, Trehiou, Alquier-Bacquie, Lasserre, Arroyo, Molette and Remignon. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Hervé Remignon, aGVydmUucmVtaWdub25AdG91bG91c2UtaW5wLmZy
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.