 Jayson R. Gifford
Jayson R. Gifford Christina Blackmon
Christina Blackmon Katelynn Hales1
Katelynn Hales1 Lee J. Hinkle
Lee J. Hinkle
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
PERSPECTIVE article
Front. Physiol. , 20 February 2024
Sec. Exercise Physiology
Volume 15 - 2024 | https://doi.org/10.3389/fphys.2024.1359119
This article is part of the Research Topic Acute and Chronic Cardiovascular Responses to Exercise View all 8 articles
Few formulas have been used in exercise physiology as extensively as the Fick formula, which calculates the rate of oxygen consumption (i.e., V.O2) as the product of cardiac output (Q.) and the difference in oxygen content in arterial and mixed venous blood (Δa
In 1870, physiologist Adolph Fick reasoned one could apply the law of mass conservation to determine cardiac output (i.e., the volume of blood pumped by the heart per unit of time, Q.) by dividing the rate of systemic oxygen consumption (V.O2) by the simultaneously measured difference in oxygen content in arterial (CaO2) and mixed venous blood (
Although the Fick formula has proven a valid and useful way to calculate V.O2 or Q., the assumptions and mathematical context of each variable must be appreciated to reach appropriate conclusions about the complex physiology of V.O2MAX. Unfortunately, the conditions and assumptions required by the Fick formula are often overlooked or oversimplified. These interpretations lead to incomplete or inappropriate conclusions about V.O2MAX physiology. Many of the most-ignored conditions and assumptions of the Fick formula are avoided by understanding the oft-omitted overdot and overline annotations in it. Understanding and recognizing the meaning of these annotations will help physiologists make more appropriate conclusions about V.O2MAX physiology. Therefore, we 1) highlight the conditions and assumptions of the Fick formula by explaining the meaning of the overdot and overline annotations and 2) encourage physiologists to consider these limitations when designing and interpreting studies regarding the physiology of V.O2MAX.
Figure 1 illustrates multiple systems are involved in transporting oxygen from the ambient air to the mitochondria (Richardson, 2003; Wagner, 2008). Although oxygen consumption (V.O2) ultimately occurs within the many mitochondria of the body, total body V.O2 is most often measured by comparing the rate of oxygen inhalation and exhalation at the mouth via indirect calorimetry (Mtaweh et al., 2018).
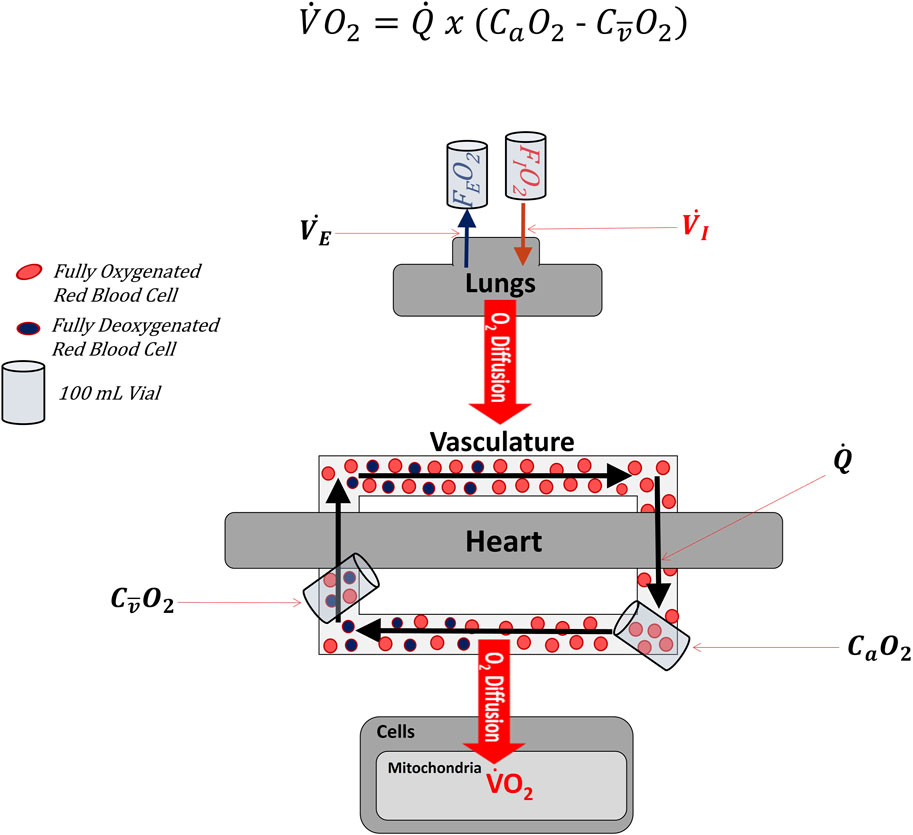
FIGURE 1. Simplified diagram lllustrating where variables found in the Fick Formula are measured along the cascade of oxygen from the atmosphere to mitochondria. Oxygen first enters the body through the lungs as a fraction of the inspired air (i.e., FIO2). Oxygen subsequently diffuses across the alveoli into the blood. The heart then pumps blood throughout the body. After leaving the heart, the vascular system constricts and dilates to direct oxygenated blood to active tissues. Once in the active tissues, some oxygen diffuses out of the blood into the distal cells (e.g., skeletal muscle), where it is consumed by the mitochondria in the process of oxidative phosphorylation. Partially deoxygenated blood leaves the active cells and returns to the heart and lungs, where it is ultimately reoxygenated. In the Fick Formula, cardiac output (Q.) represents the volume of blood leaving either side of the heart per minute. In the Fick Formula, arterial oxygen content (CaO2) is measured as the volume of oxygen contained within a fixed volume of arterial blood (typically 100 ml), while venous oxygen content (CO2) is measured as the volume of oxygen contained within a fixed volume of venous blood (typically 100 ml). Importantly, the CO2 used in the Fick Formula must represent the venous blood distal to the active tissues and as close to the heart as possible, so as to represent the average CO2 of the systemic circulation. FEO2: fraction of oxygen in expired air. V.I: Volume of inspired air per unit of time. V.E: Volume of expired air per unit of time.
Because indirect calorimetry often measures V.O2 at the mouth in an exercise physiology setting, one of the two remaining Fick formula variables (Q., Δa
Fick’s formula is often viewed as the gold standard of determining Q. (Hoeper et al., 1999). Nevertheless, exercise physiology labs often do not have the capability of sampling arterial or mixed venous blood, making the Fick-based approach of determining Q. less common in an exercise physiology setting. Alternative methods of estimating Q. have been developed: gas rebreathing, thermodilution, plethysmography and electrical bioimpedance are among those more commonly used in exercise physiology (Montero et al., 2015; Siebenmann et al., 2015). Importantly, each approach is associated with its limitations, shortcomings, and/or inherent errors (Siebenmann et al., 2015).
Some studies have applied the Fick formula to individual limbs or muscle groups to calculate the rate of oxygen consumption in specific regions of the body (Richardson et al., 1993; Boushel et al., 2011; Gifford et al., 2016). In cases of limb-specific V.O2, limb blood flow replaces Q. in the Fick formula and
The overdots in the Fick formula are often overlooked, misplaced, or forgotten, but they have important implications for interpreting V.O2 data. Dating back to Isaac Newton (this type of notation is often referred to as “Newton Notation”), “an overdot above a value indicates that the value “is a derivative taken with respect to time” (Weissten, 2019). Thus, the dot above V.O2 indicates it is a measure of the volume of oxygen consumed per unit of time. Cardiac output is denoted with an overdot (Q.) because it is the volume of blood ejected from a ventricle of the heart per unit of time (Brooks et al., 2005). In contrast, Δa
One should not assume, however, because Δa
If the extraction of oxygen is time sensitive, why is there no unit of time in Δa
In this equation, DO2 is the diffusing capacity, measured in ml of oxygen diffused in a capillary per mmHg of pressure of O2 per unit of time (i.e., oxygen diffusion occurs over time). β is a coefficient derived from the slope of the oxyhemoglobin dissociation curve, and Q.cap is the volume of blood flowing through the capillary network per unit of time (Piiper and Scheid, 1981). The length of time which a volume of blood is near the extracting tissue (i.e., red blood cell transit time), which is influenced by Q.cap and Q., influences how much oxygen can be diffused or extracted (Angleys and Østergaard, 2020; Østergaard, 2020). Nevertheless, set up as a quotient, the time units in DO2 and Q.cap cancel each other out, making the final units of oxygen extraction or Δa
Figure 1 illustrates the cardiopulmonary system as though all blood and oxygen went to the same place and experienced the same rates of flow and oxygen extraction. This is an oversimplification. Figure 2 depicts blood and oxygen being delivered to multiple regions of the body (simplified to just 2 different regions in Figure 2) with varying rates of flow and extraction. The overline above the
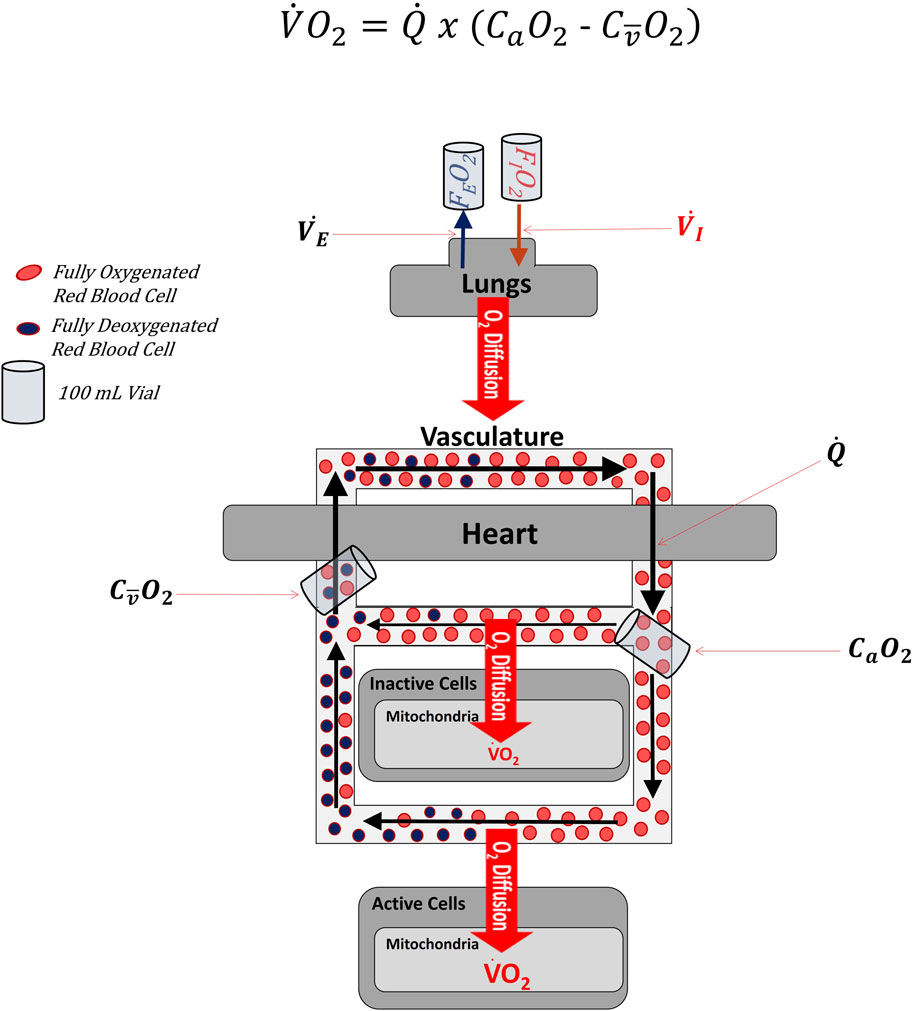
FIGURE 2. simplified diagram of the oxygen cascade illustrating that the venous sample used in the Fick Formula comes from mixed blood that represents the average of venous effluent from all regions of the body, whether active in the exercise or not. Note that the effluent leaving inactive cells is more oxygenated than the effluent leaving active cells. When the two effluents mix, the content of oxygen is no longer representative of the effluent of either the active or inactive cells, but of the average of the two.
Many physiologists have used the Fick formula to compartmentalize the factors leading to oxygen consumption into two general areas to discern what adaptations are meaningful to V.O2MAX physiology. As discussed, the Fick formula has several assumptions, implied by overdot and overline notations, that are frequently overlooked when interpreting the Fick formula. Overlooking these implications may lead to erroneous or incomplete conclusions about factors affecting V.O2MAX. Interpreting previous and future data through the lens of assumptions may lead to an improved understanding of V.O2MAX physiology and additional advancements in treatments for exercise intolerance.
The overdot above Q. and lack of one above Δa
If no other factors adjust, an increase in Q. will reduce the time during which O2 extraction can occur, thereby decreasing Δa
In contrast, if no other factors adjust, a decrease in Q. will provide a greater amount of time for extraction, resulting in a greater Δa
Clearly, the placement of the overdots in the Fick formula should remind physiologist that Δa
Short-term endurance training typically increases V.O2MAX and Q., while eliciting little-to-no change in Δa
For the sake of simplicity, suppose the heart in Figure 1 pumps 100 mL of blood containing 4 red blood cells (RBC) per second (i.e., Q. = 100 mL/s or 4 RBC/s) and the difference in the number of oxygenated RBC in the arterial and venous circulation (i.e., Δa
The direct contrast of changes in Q. and Δa
At this point relatively little is known about the training-induced adaptations that maintain extraction in the face of increased flow. Some suggest that training-induced increases in vascular function, increased capillary hematocrit, increased capillary density, and decreased flow heterogeneity (Poole et al., 2020) could potentially enhance diffusional conductance by increasing the area of the interface for diffusion. Meanwhile, evidence suggests training-induced increases in capillary density may be sufficient to slow capillary transit time in the face of increased Q. (Saltin, 1985; Kalliokoski et al., 2001). Still others contend that diffusional capacity is in excess to begin with, so adaptations are not necessary (see below). More research investigating the mechanisms responsible for the observed changes—or lack of changes—in Δa
Oxygen must diffuse across the capillary into the muscle mitochondria to be used for oxidative phosphorylation. Nevertheless, even in cases of untapped mitochondrial respiratory capacity (Boushel et al., 2011; Gifford et al., 2016), oxygen is always found in the venous blood (Cardús et al., 1998; Boushel et al., 2011). When venous samples are drawn directly from veins draining a maximally exercising limb, approximately 15% of the oxygen that entered the limb through the arterial circulation is left in the venous blood (Richardson et al., 1993; Lundby and Montero, 2015). Some contend that limitations in muscle oxygen diffusion are the reason for the remaining oxygen in venous blood during maximal exercise (Wagner, 2015b), but others (Lundby and Montero, 2015) suggest this interpretation may ignore or underestimate the mixed nature of venous blood.
The overline above
Using a Fick-Wagner Diagram, several have provided compelling evidence that factors downstream of blood flow, usually identified as diffusional conductance, alter
Although the Fick formula is useful for measuring V.O2, the information it provides about the complex physiology of V.O2MAX is often oversimplified, misinterpreted, and inaccurately stated. The overdot above Q. and the lack of one above Δa
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author.
JG: Conceptualization, Data curation, Formal Analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Validation, Visualization, Writing–original draft, Writing–review and editing. CB: Investigation, Methodology, Validation, Visualization, Writing–original draft, Writing–review and editing. KH: Methodology, Validation, Visualization, Writing–original draft, Writing–review and editing. LH: Methodology, Visualization, Writing–original draft, Writing–review and editing. SR: Methodology, Validation, Visualization, Writing–original draft, Writing–review and editing.
The author(s) declare that no financial support was received for the research, authorship, and/or publication of this article.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acierno L. J. (1999). Adolph Fick: mathematician, physicist, physiologist. Clin. Cardiol. 23, 390–391. doi:10.1002/clc.4960230519
Angleys H., Østergaard L. (2020). Krogh’s capillary recruitment hypothesis, 100 years on: is the opening of previously closed capillaries necessary to ensure muscle oxygenation during exercise? Am. J. Physiol. Heart Circ. Physiol. 318, H425–H447. doi:10.1152/ajpheart.00384.2019
Boushel R., Gnaiger E., Calbet J. A. L., Gonzalez-Alonso J., Wright-Paradis C., Sondergaard H., et al. (2011). Muscle mitochondrial capacity exceeds maximal oxygen delivery in humans. Mitochondrion 11, 303–307. doi:10.1016/j.mito.2010.12.006
Brooks G. A., Fahey T. D., Baldwin K. M. (2005). Exercise physiology: human bioenergetics and its applications. 4th ed. Boston, Ma: McGraw Hill.
Broxterman R. M., Hoff J., Wagner P. D., Richardson R. S. (2020). Determinants of the diminished exercise capacity in patients with chronic obstructive pulmonary disease: looking beyond the lungs. J. Physiol. 598, 599–610. doi:10.1113/JP279135
Calbet J. A. L., Holmberg H. C., Rosdahl H., Van Hall G., Jensen-Urstad M., Saltin B. (2005). Why do arms extract less oxygen than legs during exercise? Am. J. Physiol. - Regul. Integr. Comp. Physiol. 289, 1448–1458. doi:10.1152/ajpregu.00824.2004
Cardús J., Marrades R. M., Roca J., Barberà J. a., Diaz O., Masclans J. R., et al. (1998). Effects of FIO2 on leg V.O2 during cycle ergometry in sedentary subjects. Med. Sci. Sports Exerc. 30, 697–703. doi:10.1097/00005768-199805000-00009
Gifford J. R., Garten R. S., Nelson A. D., Trinity J. D., Layec G., Witman M. A. H. H., et al. (2016). Symmorphosis and skeletal muscle V.O2 max: in vivo and in vitro measures reveal differing constraints in the exercise-trained and untrained human. J. Physiol. 594, 1741–1751. doi:10.1113/JP271229
Heinonen I., Koga S., Kalliokoski K. K., Musch T. I., Poole D. C. (2015). Heterogeneity of muscle blood flow and metabolism: influence of exercise, aging, and disease states. Exerc. Sport Sci. Rev. 43, 117–124. doi:10.1249/JES.0000000000000044
Hirai D. M., Musch T. I., Poole D. C. (2015). Exercise training in chronic heart failure: improving skeletal muscle O2 transport and utilization. Am. J. Physiol. - Hear. Circ. Physiol. 309, H1419–H1439. doi:10.1152/ajpheart.00469.2015
Hoeper M. M., Maier R., Tongers J., Niedermeyer J., Hohlfeld J. M., Hamm M., et al. (1999). Determination of cardiac output by the Fick method, thermodilution, and acetylene rebreathing in pulmonary hypertension. Am. J. Respir. Crit. Care Med. 160, 535–541. doi:10.1164/ajrccm.160.2.9811062
Kalliokoski K. K., Knuuti J., Nuutila P. (2004). Blood transit time heterogeneity is associated to oxygen extraction in exercising human skeletal muscle. Microvasc. Res. 67, 125–132. doi:10.1016/j.mvr.2003.11.004
Kalliokoski K. K., Oikonen V., Takala T. O., Sipilä H., Knuuti J., Nuutila P. (2001). Enhanced oxygen extraction and reduced flow heterogeneity in exercising muscle in endurance-trained men. Am. J. Physiol. Endocrinol. Metab. 280, E1015–E1021. doi:10.1152/ajpendo.2001.280.6.E1015
Levine B. D. (2008). V.O2max: what do we know, and what do we still need to know? J. Physiol. 586, 25–34. doi:10.1113/jphysiol.2007.147629
Lundby C., Montero D. (2015). CrossTalk opposing view: diffusion limitation of O2 from microvessels into muscle does not contribute to the limitation of V.O2 max. J. Physiol. 593, 3759–3761. doi:10.1113/JP270550
Montero D., Diaz-Cañestro C., Lundby C. (2015). Endurance training and V.O2max: role of maximal cardiac output and oxygen extraction. Med. Sci. Sports Exerc. 47, 2024–2033. doi:10.1249/MSS.0000000000000640
Mtaweh H., Tuira L., Floh A. A., Parshuram C. S. (2018). Indirect calorimetry: history, technology, and application. Front. Pediatr. 6, 257–258. doi:10.3389/fped.2018.00257
Østergaard L. (2020). Blood flow, capillary transit times, and tissue oxygenation: the centennial of capillary recruitment. J. Appl. Physiol. 129, 1413–1421. doi:10.1152/JAPPLPHYSIOL.00537.2020
Piiper J., Scheid P. (1981). Model for capillary-alveolar equilibration with special reference to O2 uptake in hypoxia. Respir. Physiol. 46, 193–208. doi:10.1016/0034-5687(81)90121-3
Poole D. C., Behnke B. J., Musch T. I. (2020). The role of vascular function on exercise capacity in health and disease. J. Physiol. 599, 889–910. JP278931. doi:10.1113/JP278931
Richardson R. S. (2003). Oxygen transport and utilization: an integration of the muscle systems. Adv. Physiol. Educ. 27, 183–191. doi:10.1152/advan.00038.2003
Richardson R. S., Poole D. C., Knight D. R., Kurdak S. S., Hogan M. C., Grassi B., et al. (1993). High muscle blood flow in man: is maximal O2 extraction compromised? J. Appl. Physiol. 75, 1911–1916. doi:10.1152/jappl.1993.75.4.1911
Roca J., Agusti a G., Alonso A., Poole D. C., Viegas C., Barbera J. a., et al. (1992). Effects of training on muscle O2 transport at V.O2max. J. Appl. Physiol. 73, 1067–1076. doi:10.1152/jappl.1992.73.3.1067
Saltin B. (1985). Hemodynamic adaptations to exercise. Am. J. Cardiol. 55, 42D–47D. doi:10.1016/0002-9149(85)91054-9
Saltin B., Blomqvist G., Mitchell J., Johnson R., Wildenthal K., Chapman C., et al. (1968). A longitudinal study of adaptive changes in oxygen transport and body composition. Circulation 37/38. doi:10.1161/01.cir.38.5s7.vii-1
Shapiro E. (1972). Adolf Fick-Forgotten genius of cardiology. Am. J. Cardiol. 30, 662–665. doi:10.1016/0002-9149(72)90606-6
Siebenmann C., Rasmussen P., Sørensen H., Zaar M., Hvidtfeldt M., Pichon A., et al. (2015). Cardiac output during exercise: a comparison of four methods. Scand. J. Med. Sci. Sport. 25, e20–e27. doi:10.1111/sms.12201
Wagner P. D. (2000). Diffusive resistance to O 2 transport in muscle. Acta Physiol. Scand. 168, 609–614. doi:10.1046/j.1365-201x.2000.00712.x
Wagner P. D. (2008). Systemic oxygen transport and utilization. J. Breath. Res. 2, 024001–024012. doi:10.1088/1752-7155/2/2/024001
Wagner P. D. (2015a). A re-analysis of the 1968 Saltin et al. “Bedrest” paper. Scand. J. Med. Sci. Sport. 25, 83–87. doi:10.1111/sms.12597
Wagner P. D. (2015b). CrossTalk proposal: diffusion limitation of O2 from microvessels into muscle does contribute to the limitation of V.O2 max. J. Physiol. 593, 3757–3758. doi:10.1113/JP270551
Weisstein E. (2020). Macron. From MathWorld—a wolfram web resource. Available at: https://mathworld.wolfram.com/Macron.html.
Weissten E. W. (2019). Overdot. From MathWorld—a wolfram web resource. Available at: http://mathworld.wolfram.com/Overdot.html.
Keywords: V.O2MAX, cardiac output, arteriovenous oxygen difference, endurance training, muscle oxygen diffusion
Citation: Gifford JR, Blackmon C, Hales K, Hinkle LJ and Richards S (2024) Overdot and overline annotation must be understood to accurately interpret
Received: 20 December 2023; Accepted: 26 January 2024;
Published: 20 February 2024.
Edited by:
Keith George, Liverpool John Moores University, United KingdomReviewed by:
Anton Ušaj, University of Ljubljana, SloveniaCopyright © 2024 Gifford, Blackmon, Hales, Hinkle and Richards. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jayson R. Gifford, amF5c29uZ2lmZm9yZEBieXUuZWR1
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.