 Egor Igorevich Kulikov
Egor Igorevich Kulikov Lidia Ivanovna Malakheeva
Lidia Ivanovna Malakheeva Alexey Sergeevich Komarchev
Alexey Sergeevich Komarchev
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
OPINION article
Front. Physiol., 31 January 2024
Sec. Avian Physiology
Volume 15 - 2024 | https://doi.org/10.3389/fphys.2024.1315346
This article is part of the Research TopicThe Role of the Pancreas in PoultryView all 6 articles
In recent years, there have been many studies that have shown the direct and indirect effects of B-cell translocation gene 1 (BTG1) and BTG2 expression levels on insulin secretion and sensitivity of the cell to insulin. These genes belonging to the B-cell translocation gene/transducer of the BTG/TOB family are expressed in different cells and regulate cell cycle processes, such as proliferation, apoptosis, and differentiation (Fletcher et al., 1991; Mauxion et al., 2009; Weisel et al., 2010). They were highly specific among the species of living organisms (Rouault et al., 1993). These studies suggest that there is a direct correlation between the expression levels of the BTG1 and BTG2 genes, standard of insulin, and metabolism in broiler chickens. The study of these genes and their influence on metabolism is, therefore, a promising area of research for physiologists and geneticists.
All metabolic processes occurring in a living organism are based on the use of energy in the form of ATP, synthesized mainly from glucose. As with all metabolic processes, metabolism of glucose is compiled by neuro-humoral regulation and participation of hormones such as insulin and glucagon. Insulin is synthesized in the β-cells of the pancreas (Moore and Cooper, 1991; Norton et al., 2022). Secretion of insulin is related with the level of metabolism in broilers (Vertiprakhov et al., 2023). Tissue metabolomics indicated the unique effects of insulin on amino acid metabolism (Cai et al., 2012).
Insulin is a hormone produced by the pancreas in response to high blood glucose levels. It plays a crucial role in regulating glucose metabolism in the body. In chickens, insulin has been shown to have several effects, including the following:
1) Glucose uptake: insulin stimulates the transfer of glucose from the bloodstream into cells. In chickens, this is particularly important for muscle cells, which use a lot of glucose for producing energy during movement and growth (Qaid et al., 2016).
2) Lipid metabolism: insulin also regulates lipid metabolism in chickens by promoting the storage of excess fat in the adipose tissue and inhibiting the breakdown of stored fat for energy (Saltiel et al., 2001).
3) Growth and development: insulin is a key regulator of growth and development in chickens, particularly during the early stages of life. It promotes cell division and differentiation, leading to increased muscle and bone growth (Hill et al., 1985).
4) Reproduction: insulin also plays a role in reproductive function in birds by regulating the production of steroid hormones and supporting the development of oocytes (Sliwowska et al., 2014).
Overall, insulin is essential for maintaining normal glucose metabolism and promoting growth and development in chickens. Deficiency or imbalance in insulin production can lead to metabolic disorders and other health issues.
As noted above, B-cell translocation gene 1 (BTG1) and BTG2 are two genes that belong to the BTG/Tob family of proteins (see Figure 1). These genes have been shown to play a role in regulating cell growth, differentiation, and apoptosis in various animal species, including chickens.
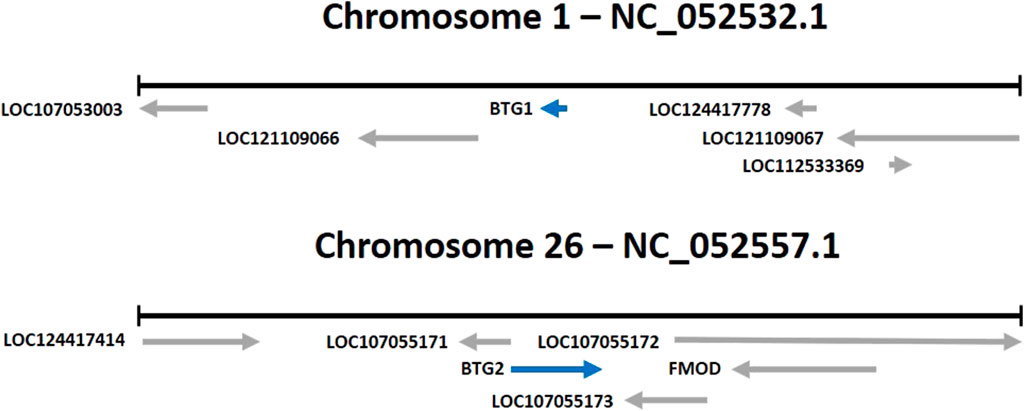
FIGURE 1. Location of the BTG1 and BTG2 genes (Gallus gallus).
Studies have shown the expression of BTG1 and BTG2 genes in the brain and nervous system during embryonic development in chickens. This suggests that these genes may be involved in the regulation and function of neural development.
BTG1 has been shown to influence muscle growth in different animals (Rodier et al., 2001; Iwai et al., 2004; Busson et al., 2005; Yuniati et al., 2019). In a study of BTG2 gene expression levels in pigs, an association with inhibition of primary muscle fiber proliferation was found (Feng et al., 2007). The BTG1 gene was identified by genome-wide association studies (GWAS) as a promising marker for muscle development in Brahman cattle (Crispim et al., 2015). High expression of the BTG1 is found in immature muscle cells, especially during active muscle growth. It has been determined that BTG1 stimulates the activity of myogenic factors, in particular the nuclear receptors already known as positive regulators of myogenin, which boosts the transcription by myoblasts and stimulates their differentiation (Busson et al., 2005). In addition, BTG1 and BTG2 have been shown to participate in regulating muscle development and growth in chickens (Mohammadabadi et al., 2021). It was proven that BTG1 and BTG2 are abundantly expressed in skeletal muscles. There were also differences between broilers and laying hens in gene expression levels in the sternum, pancreas, and heart (Zhao et al., 2022). It can be assumed that when analyzing the nucleotide sequence of these genes, differences in polymorphic sites can be determined in meat- and egg-producing hens, which would indicate a direct relationship with productivity and the prospects of these genes as markers affecting metabolism.
Studies have observed that the increasing expression of these genes is related to upregulated muscle regeneration after injury, suggesting that they may be involved in the repair and growth of muscle tissue. Muscle stem cells (MuSCs) are a crucial part in the mechanism of muscle regeneration through paracrine signaling interactions. BTG2 promotes differentiation in hematopoietic and nerve cells by inhibiting Id3 and cyclin D1 to limit cell cycle progression (Yuniati et al., 2019). Scientists hypothesize that BTG2 and Id3 may be markers of resting-state transcription of MuSCs (de Micheli et al., 2020).
Furthermore, BTG1 and BTG2 have been implicated in the regulation of immune function in chickens. Some studies have detected that the high expression of these genes is manifested in response to viral infections, suggesting that they may participate in the host response to viral pathogens. The correlation of BTG2 with immune reactions was confirmed by a group of scientists, who proved that the expression of this gene is significantly correlated with tumor purity (Zhang et al., 2022). Some studies have identified an important role for the BTG1 and BTG2 genes in tumor suppression, i.e., a decreased expression of these genes results in more severe malignant neoplasms, leading to death. These genes may be biomarkers of disease progression (Cho et al., 2004; Kanda et al., 2015).
BTG1 and BTG2 play key roles in a wide range of cellular activities, including cell growth, proliferation, and apoptosis, through modulation of various central biological steps such as transcription, post-transcription, and translation (Kim et al., 2022). The researchers have found that activation of the expression of BTG1 and BTG2 by B-cells at the early stages of their development, especially in living organisms during the embryonic period and the period of postnatal adaptation, is associated with their regulatory functions, particularly their involvement in cell differentiation (Tijchon et al., 2016).
Overall, the BTG1 and BTG2 genes take part in regulating various biological processes in chickens, including neural development, muscle growth and regeneration, and immune function. It has been determined that BTG1 is required to preserve and replenish stem cells in the dentate gyrus and subventricular zone (SVZ) in adults. The reduced number of adult neurons in the dentate gyrus in BTG1-null mice is associated with subtle impairments in hippocampus-dependent learning and memory (Micheli et al., 2015).
Table 1 summarizes the functions of BTG1 and BTG2 that have been previously investigated.
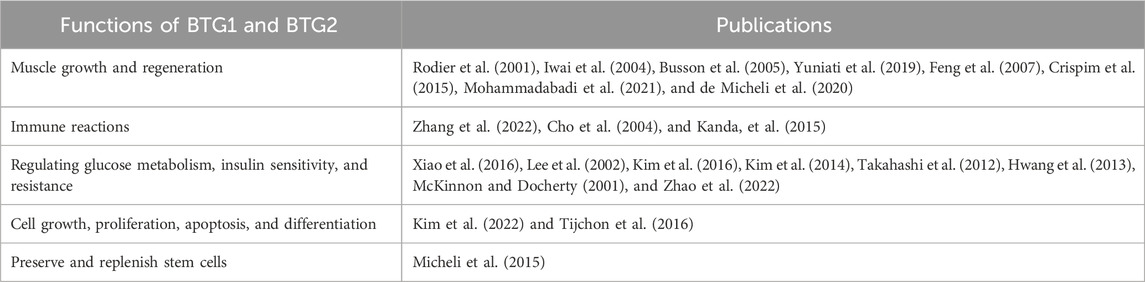
TABLE 1. Functions of the BTG1 and BTG2 genes.
By analyzing the available literature, the possible pathways of the influence of BTG1 and BTG2 genes on insulin levels in chickens were identified.
In humans, insulin is the major anabolic hormone regulating the metabolism of glucose, fats, and proteins. The role of insulin is to maintain body weight either by decreasing catabolism or increasing anabolism (Heslin et al., 1996).
Some serious metabolic diseases are related to loss of insulin sensitivity by cells. This state has been called insulin resistance. Currently, the mechanisms of its development are not very clear and understandable. As stated above, BTG1, as an important cofactor, is involved in many physiological processes, so it may also be related with energy metabolism and exchange of glucose. In vitro, the expression of this gene can influence insulin signaling. So when mice with insulin resistance are infected by adenovirus, the high expression of BTG1 increases the sensitivity of cells to insulin, similar to that observed in wild-type mice. Moreover, transgenic mice with high levels of expression of BTG1 were insensitive to insulin resistance induced by a high-carbohydrate diet (Xiao et al., 2016).
Another study discovered that BTG2 expression was decreased in the adipose tissue of obese humans, proposing that it could be involved in the development of insulin resistance. In an experiment on mice, it was found that the expression level of BTG2 varies depending on the body build, and the knockdown of this gene increased lipid accumulation (Kim et al., 2016).
In addition, a new function of BTG1 in the regulation of insulin sensitivity in the liver cells in mice via activation of transcription factor JUN (c-Jun), which stimulates cell proliferation, has been observed. The study showed that the expression of BTG1 is increased in the rat liver with protein and energy deficiency in the diet (Lee et al., 2002).
Recent work has suggested that members of this family play a role in maintaining blood glucose homeostasis (Takahashi et al., 2012; Kim et al., 2014). High expression of BTG2 by mice accompanied the activation of the gluconeogenic gene in the liver, which leads to an increase in the level of glucose in the bloodstream and, later, a decrease in insulin tolerance (Xiao et al., 2016).
The regulation of insulin secretion also involves glucagon-like peptide-1 (GLP-1), which is synthesized by the endocrine cells of the intestines. This peptide hormone belongs to the incretin family and is an important element in the treatment of type-2 diabetes. Recent research studies established a novel molecular mechanism that proves that GLP-1 stimulates the expression of the insulin gene via BTG2 (Hwang et al., 2013).
The critical stage of pancreatic development is characterized by the high expression levels of pancreatic duodenal homeobox-1 (PDX1), and in the case of the low expression level of PDX1, the pancreas could not progress well. So homozygous mutation of this gene in mice and humans leads to agenesis of the pancreas. In adults, PDX1 manifested by the beta cells of the islets of Langerhans is related to the synthesis of insulin, and that expressed by delta cells activates the secretion of somatostatin, which ultimately leads to decreased glucose levels. Expression of PDX1 is also observed in the dispersed endocrine cells of the duodenum. The reason for some types of diabetes, for example, MODY4 in young people and type II (non-insulin-dependent) diabetes, which usually manifests itself at an older age, may be heterozygous mutations in the PDX1 gene, which can worsen glucose tolerance (McKinnon et al., 2001).
Scientists have found that BTG2 may regulate the expression of PDX1 in pancreatic β-cells, so its stimulation may lead to the secretion of insulin. As noted above, infection by adenovirus leads to a high level of expression of BTG2, which is activated significantly, increasing insulin secretion and expression of the PDX1 gene. More detailed molecular studies showed that BTG2 had a direct influence on PDX1, increasing PDX1 occupancy at the insulin gene promoter. Increased insulin secretion was stimulated by GLP1 or its analog, Exendin-4 (Ex-4), which is used in the treatment of diabetes and was blocked by the small interfering RNA system of BTG2.
The level of expression of BTG1 and BTG2 genes in chickens depends on the breeds and type of the tissue. So the expression of BTG genes in thoracic and pancreatic cells was negatively correlated with the glucose level in the bloodstream of chickens (Zhao et al., 2022).
Further research is needed to fully understand the mechanisms of the effects of BTG1 and BTG2 genes on insulin and glucose metabolism.
Based on the analytic results of many sources, we can confidently assume that the use of BTG1 and BTG2 as candidate genes for regulation of the insulin level is promising and can lead to significant results in the poultry sector, where insulin is one of the main hormones responsible for regulation of metabolism, increase in amino acid transport, and stimulation of lipogenesis.
All studies conducted have addressed the effects of the expression of these genes, but they have not analyzed the amino acid sequence or searched for associations with the level of genetic expression.
The study of promoters and coding regions of these genes and the search for their single-nucleotide polymorphisms may provide a more accurate understanding of the mechanisms of their functioning.
For this work, we need to sequence these genes and collect expression data from chickens under equal rearing conditions. Once the associations have been made, we will be able to determine the relationship of the nucleotide sequence with the level of synthesis of the proteins under study.
This work will open the possibility of using these genes as candidates for MAS of chickens.
These research studies can provide a significant boost to the work regarding avian feed conversion, and the obtained data may improve our understanding of the functioning of the relationship between BTG1 and BTG2 genes, insulin levels, and metabolic regulation.
The study of this topic is a promising area for physiologists and geneticists who are researching the functions of the pancreas and insulin.
EK: writing–original draft and writing–review and editing. LM: writing–review and editing. AK: writing–review and editing.
The author(s) declare that no financial support was received for the research, authorship, and/or publication of this article.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors, and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Busson M., Carazo A., Seyer P., Grandemange S., Casas F., Pessemesse L., et al. (2005). Coactivation of nuclear receptors and myogenic factors induces the major BTG1 influence on muscle differentiation. Oncogene 24 (10), 1698–1710. PMID: 15674337. doi:10.1038/sj.onc.1208373
Cai S., Wang O., Wang M., He J., Wang Y., Zhang D., et al. (2012). In vitro inhibitory effect on pancreatic lipase activity of subfractions from ethanol extracts of fermented Oats (Avena sativa L.) and synergistic effect of three phenolic acids. J. Agric. Food Chem. 60 (29), 7245–7251. Epub 2012 Jul 13. PMID: 22765648. doi:10.1021/jf3009958
Cho J. W., Kim J. J., Park S. G., Lee D. H., Lee S. C., Kim H. J., et al. (2004). Identification of B-cell translocation gene 1 as a biomarker for monitoring the remission of acute myeloid leukemia. Proteomics 4 (11), 3456–3463. doi:10.1002/pmic.200400968
Crispim A. C., Kelly M. J., Guimarães S. E. F., Silva F. F. E., Fortes M. R. S., Wenceslau R. R., et al. (2015). Multi-trait GWAS and new candidate genes annotation for growth curve parameters in brahman cattle. PLoS ONE 10, e0139906. doi:10.1371/journal.pone.0139906
De Micheli A., Laurilliard E., Heinke C., Ravichandran H., Fraczek P., Soueid-Baumgarten S., et al. (2020). Single-cell analysis of the muscle stem cell hierarchy identifies heterotypic communication signals involved in skeletal muscle regeneration. Cell Rep. 30, 3583–3595. e5. doi:10.1016/j.celrep.2020.02.067
Feng Z., Tang Z. L., Li K., Liu B., Yu M., Zhao S. H. (2007). Molecular characterization of the BTG2 and BTG3 genes in fetal muscle development of pigs. Gene 403 (1-2), 170–177. Epub 2007 Aug 25. PMID: 17890019. doi:10.1016/j.gene.2007.08.009
Fletcher B., Lim R. W., Varnum B. C., Kujubu D. A., Koski R., Herschman H. (1991). Structure and expression of TIS21, a primary response gene induced by growth factors and tumor promoters. J. Biol. Chem. 266, 14511–14518. doi:10.1016/S0021-9258(18)98716-8
Heslin M. J. (1996). “Insulin to impact on protein and amino acid metabolism,” in Protein and amino acid metabolism in cancer cachexia. Medical intelligence unit (Berlin, Heidelberg: Springer). doi:10.1007/978-3-662-22346-8_13
Hill D. J., Milner R. D. (1985). Insulin as a growth factor. Pediatr. Res. 19 (9), 879–886. PMID: 2413420. doi:10.1203/00006450-198509000-00001
Hwang S. L., Kwon O., Kim S. G., Lee I. K., Kim Y. D. (2013). B-cell translocation gene 2 positively regulates GLP-1-stimulated insulin secretion via induction of PDX-1 in pancreatic β-cells. Exp. Mol. Med. 45 (5), e25. PMCID: PMC3674408. doi:10.1038/emm.2013.47
Iwai K., Hirata K., Ishida T., Takeuchi S., Hirase T., Rikitake Y., et al. (2004). An anti-proliferative gene BTG1 regulates angiogenesis in vitro. Biochem. Biophys. Res. Commun. 316 (3), 628–635. PMID: 15033446. doi:10.1016/j.bbrc.2004.02.095
Kanda M., Oya H., Nomoto S., Takami H., Shimizu D., Hashimoto R., et al. (2015). Diversity of clinical implication of B-cell translocation gene 1 expression by histopathologic and anatomic subtypes of gastric cancer. Dig. Dis. Sci. 60 (5), 1256–1264. Epub 2014 Dec 9. PMID: 25487193. doi:10.1007/s10620-014-3477-8
Kim S., Hong J. W., Park K. W. (2016). B cell translocation gene 2 (Btg2) is regulated by Stat3 signaling and inhibits adipocyte differentiation. Mol. Cell Biochem. 413, 145–153. doi:10.1007/s11010-015-2648-z
Kim S. H., Jung I. R., Hwang S. S. (2022). Emerging role of anti-proliferative protein BTG1 and BTG2. BMB Rep. 55 (8), 380–388. PMID: 35880434; PMCID: PMC9442347. doi:10.5483/BMBRep.2022.55.8.092
Kim Y. D., Kim S. G., Hwang S. L., Choi H. S., Bae J. H., Song D. K., et al. (2014). B-cell translocation gene 2 regulates hepatic glucose homeostasis via induction of orphan nuclear receptor Nur77 in diabetic mouse model. Diabetes 63 (6), 1870–1880. Epub 2014 Mar 19. PMID: 24647738. doi:10.2337/db13-1368
Lee A. K., Kang K. W., Kim Y. G., Cho M. K., Lee M. G., Shim C. K., et al. (2002). Identification of genes enhanced by protein-calorie malnutrition by differential display polymerase chain reaction (expression of fibrinogen B beta chain, B cell translocation gene 1 and thyroid hormone responsive protein genes). Mol. Cell. Biochem. 231, 163–171. doi:10.1023/a:1014481319278
Mauxion F., Chen C.-Y., Séraphin B., Shyu A.-B. (2009). BTG/TOB factors impact deadenylases. Trends Biochem. Sci. 34, 640–647. doi:10.1016/j.tibs.2009.07.008
McKinnon C. M., Docherty K. (2001). Pancreatic duodenal homeobox-1, PDX-1, a major regulator of beta cell identity and function. Diabetologia 44 (10), 1203–1214. doi:10.1007/s001250100628
Micheli L., Ceccarelli M., Farioli-Vecchioli S., Tirone F. (2015). Control of the normal and pathological development of neural stem and progenitor cells by the PC3/tis21/btg2 and Btg1 genes. J. Cell Physiol. 230 (12), 2881–2890. doi:10.1002/jcp.25038
Mohammadabadi M., Bordbar F., Jensen J., Du M., Guo W. (2021). Key genes regulating SkeletalMuscle development and growth inFarm animals. Animals 11, 835. doi:10.3390/ani11030835
Moore C. X., Cooper G. J. S. (1991). Co-secretion of amylin and insulin from cultured islet beta-cells: modulation by nutrient secretagogues, islet hormones and hypoglycemic agents. Biochem. Biophys. Res. Commun. 179, 1–9. doi:10.1016/0006-291x(91)91325-7
Norton L., Shannon C., Gastaldelli A., Defronzo R. A. (2022). Insulin: the masterregulator of glucose metabolism. Metabolism 129, 155142. doi:10.1016/j.metabol.2022.155142
Qaid M., Abdelrahman M. (2016). Role of insulin and other related hormones in energy metabolism- A review. Cogent Food & Agric. 2. doi:10.1080/23311932.2016.1267691
Rodier A., Rochard P., Berthet C., Rouault J. P., Casas F., Daury L., et al. (2001). Identification of functional domains involved in BTG1 cell localization. Oncogene 20 (21), 2691–2703. doi:10.1038/sj.onc.1204398
Rouault J. P., Samarut C., Duret L., Tessa C., Samarut J., Magaud J. P. (1993). Sequence analysis reveals that the BTG1 anti-proliferative gene is conserved throughout evolution in its coding and 3' non-coding regions. Gene 129 2, 303–306. doi:10.1016/0378-1119(93)90284-A
Saltiel A. R., Kahn C. R. (2001). Insulin signalling and the regulation of glucose and lipid metabolism. Nature 414 (6865), 799–806. doi:10.1038/414799a
Sliwowska J. H., Fergani C., Gawałek M., Skowronska B., Fichna P., Lehman M. N. (2014). Insulin: its role in the central control of reproduction. Physiol. Behav. 133, 197–206. Epub 2014 May 27. PMID: 24874777; PMCID: PMC4084551. doi:10.1016/j.physbeh.2014.05.021
Takahashi A., Morita M., Yokoyama K., Suzuki T., Yamamoto T. (2012). Tob2 inhibits peroxisome proliferator-activated receptor γ2 expression by sequestering Smads and C/EBPα during adipocyte differentiation. Mol. Cell Biol. 32 (24), 5067–5077. Epub 2012 Oct 15. PMID: 23071089; PMCID: PMC3510534. doi:10.1128/MCB.00610-12
Tijchon E., van Emst L., Yuniati L., van Ingen Schenau D., Havinga J., Rouault J. P., et al. (2016). Tumor suppressors BTG1 and BTG2 regulate early mouse B-cell development. Haematologica 101 (7), e272–e276. Epub 2016 Apr 1. PMID: 27036158; PMCID: PMC5004470. doi:10.3324/haematol.2015.139675
Vertiprakhov V., Grozina A. A., Fisinin V., Surai P. (2023). Adaptation of chicken pancreatic secretory functions to feed composition. World's Poult. Sci. J. 79, 27–41. doi:10.1080/00439339.2023.2163042
Weisel F. J., Appelt U. K., Schneider A. M., Ju H., van Rooijen N., Korner H., et al. (2010). Unique requirements for reactivation of virus-specific memory B lymphocytes. J. Immunol. 185 (7), 4011–4021. Epub 2010 Aug 25. PMID: 20739675. doi:10.4049/jimmunol.1001540
Xiao F., Deng J., Yu J., Guo Y., Chen S., Guo F. (2016). A novel function of B-cell translocation gene 1 (BTG1) in the regulation of hepatic insulin sensitivity in mice via c-Jun. FASEB J. 30 (1), 348–359. Epub 2015 Sep 22. PMID: 26396236. doi:10.1096/fj.15-278689
Yuniati L., Scheijen B., Meer LTVD, Leeuwen F. N. V. (2019). Tumor suppressors BTG1 and BTG2: beyond growth control. J. Cell Physiol. 234, 5379–5389. doi:10.1002/jcp.27407
Zhang X. Z., Chen M. J., Fan P. M., Jiang W., Liang S. X. (2022). BTG2 serves as a potential prognostic marker and correlates with immune infiltration in lung adenocarcinoma. Int. J. Gen. Med. 15, 2727–2745. PMID: 35300128; PMCID: PMC8922043. doi:10.2147/IJGM.S340565
Keywords: BTG1 gene, BTG2 gene, insulin, chicken, metabolism
Citation: Kulikov EI, Malakheeva LI and Komarchev AS (2024) The role of BTG1 and BTG2 genes and their effects on insulin in poultry. Front. Physiol. 15:1315346. doi: 10.3389/fphys.2024.1315346
Received: 10 October 2023; Accepted: 15 January 2024;
Published: 31 January 2024.
Edited by:
Vladimir Vertiprakhov, Moscow Timiryazev Agricultural Academy, RussiaReviewed by:
Larisa Ilina, Biotrof Ltd., RussiaCopyright © 2024 Kulikov, Malakheeva and Komarchev. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Egor Igorevich Kulikov, a3VsaWtvdmVnb3IzM0B5YW5kZXgucnU=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.