 Jin’e Li1,2
Jin’e Li1,2 Yinghong Liu
Yinghong Liu Jianping Liu
Jianping Liu
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Physiol. , 25 September 2023
Sec. Renal Physiology and Pathophysiology
Volume 14 - 2023 | https://doi.org/10.3389/fphys.2023.1220450
This article is part of the Research Topic Advances and Insights in Peritoneal Dialysis: A Physiological Perspective View all 10 articles
Peritoneal dialysis (PD) is an effective alternative treatment for patients with end-stage renal disease (ESRD) and is increasingly being adopted and promoted worldwide. However, as the duration of peritoneal dialysis extends, it can expose problems with dialysis inadequacy and ultrafiltration failure. The exact mechanism and aetiology of ultrafiltration failure have been of great concern, with triggers such as biological incompatibility of peritoneal dialysis solutions, uraemia toxins, and recurrent intraperitoneal inflammation initiating multiple pathways that regulate the release of various cytokines, promote the transcription of fibrosis-related genes, and deposit extracellular matrix. As a result, peritoneal fibrosis occurs. Exploring the pathogenic factors and molecular mechanisms can help us prevent peritoneal fibrosis and prolong the duration of Peritoneal dialysis.
The peritoneum is made up of a single layer of mesoderm-derived mesothelial cells and a thin layer of connective tissue consisting of a dense subepithelial band. The dense band, composed mainly of collagen fibre bundles, along with some lymphocyte tubes, fibroblasts, mast cells, macrophages, and capillaries, plays a crucial role in maintaining the function and structure of the peritoneum (Bajo et al., 2017). The peritoneal natural semipermeable membrane is responsible for ultrafiltration and solute diffusion during dialysis by converting solutes and water and removing metabolites while maintaining water and electrolyte balance.
Peritoneal dialysis is a cost-effective method of dialysis (Li et al., 2017) that is gaining international attention. However, various nonphysiological factors in the dialysate, such as hyperosmolarity, hyperglycaemia, low pH, glucose degradation products (GDPs), and advanced glycosylation end products (AGEs), can lead to chronic stimulation and damage of the peritoneum in PD patients, causing peritoneal sclerosis. Peritoneal fibrosis has always been a hot topic of research. Here, we provide an overview of how factors such as inflammation, oxidative stress, glucose metabolism, and hypoxia mediate peritoneal fibrosis through cytokine generation and molecular pathway activation.
Epithelial-mesenchymal transition (EMT) is a complex biological process characterized by the gradual loss of epithelial-specific markers, such as E-cadherin and zonula occludens-1, and the acquisition of a fibroblast phenotype expressing fibroblast-specific protein 1 (FSP1) and α-smooth muscle actin (α-SMA). This process is accompanied by changes in cellular behaviour and the production of extracellular matrix (ECM) (Kalluri and Neilson, 2003).
The exact mechanisms underlying the EMT of peritoneal mesothelial cells remain unclear, but they are thought to involve the interaction of cytokines, inflammatory factors, and transcription regulators. Evidence suggests that Smad and non-Smad signalling pathways induced by TGF-β1 play a dominant role in the EMT of peritoneal fibrosis (Lho et al., 2021). In the early phase of fibrosis, glucose, GDPs, and advanced glycosylation end products (AGEs) can upregulate type I and type II TGF-β receptors in mesothelial cells (Suryantoro et al., 2023) by activating protein kinase C-α (PKC-α) (Wang et al., 2016). TGF-β1 signalling activates the phosphorylation of Smad2 and Smad3 via type I TGF-β receptors, and Smad2/Smad3 are transported to the nucleus, where they directly bind to DNA and regulate the transcription of target genes, including Snail, collagen, α-SMA, fibronectin, CTGF, β-catenin, plasminogen activator inhibitor-1 (PAI-1), and matrix metalloproteinase-2 (MMP2), promoting fibrosis (Derynck and Zhang, 2003; Hirahara et al., 2009; Xiao et al., 2010; Lei et al., 2012; Zhang et al., 2022a; Huang et al., 2022; Masola et al., 2022) (Figure 1). Smad1/5/8 proteins activated by ALKs in response to BMP (bone morphogenetic proteins) one to four or other ligands are also transported to the nucleus to regulate the transcription of target genes (Balzer, 2020). In addition to the Smad-dependent signalling pathway, there are also various non-Smad signalling pathways involved in the process of fibrosis, such as PI3K/Akt, c-Jun N-terminal kinases (JNK), Wnt/β-catenin, and ERK/NF-κB (Liu et al., 2012; Jang et al., 2013; Zhang et al., 2013; Lamouille et al., 2014).
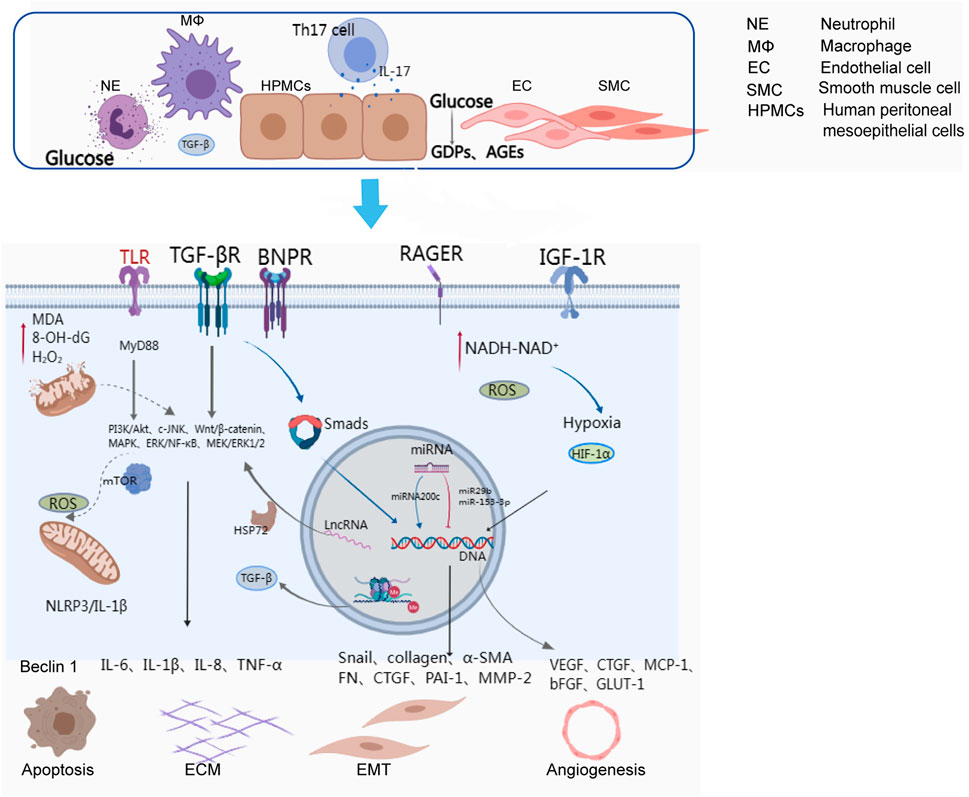
FIGURE 1. Cells and activated molecular pathways involved in peritoneal ultrafiltration failure.
Numerous studies have reported various initiating factors in epithelial-mesenchymal transition (EMT). For instance, caveolin-1 has been proposed to play a critical role in EMT associated with peritoneal fibrosis in patients with Parkinson’s disease (Strippoli et al., 2015). Furthermore, overexpression of insulin-like growth factor 1 receptor (IGF-1R) has been linked to the promotion of EMT in human peritoneal mesothelial cells (Xia et al., 2022). These findings have expanded our understanding of potential strategies for preventing and treating EMT.
Inflammation is a frequent underlying cause of peritoneal fibrosis in patients with PD. While it can be directly induced by pathogenic microorganisms, peritoneal inflammation may also result from the accumulation of uraemia toxins, mechanical stress on blood vessel walls, ageing, and complications of diabetes (Cueto-Manzano et al., 2007). In PD patients, an increase in the number of peritoneal macrophages suggests chronic peritoneal inflammation, even in the absence of acute peritonitis.
The inflammatory response is a complex process involving multiple molecular mechanisms, inflammatory mediators, and signalling pathways. The peritoneum can be damaged by various factors, activating macrophages and neutrophils that mediate NF-κB signalling pathways via Toll-like receptors (TLRs) and release numerous inflammatory cytokines, such as IL-6, IL-1β, IL-8, and TNF-α, leading to peritoneal fibrosis (Kato et al., 2004). TLRs can also activate the downstream molecules of JNK, p38MAPKs, and ERK1/2 through the MyD88 signalling pathway, promoting the expression of proinflammatory cytokines. In addition, the NLRP3 inflammasome, a cellular complex composed of proteins, can activate the cysteine-dependent aspartate-directed proteolytic enzyme and induce the production of IL-1 and IL-18, resulting in inflammatory responses (Hautem et al., 2017) (Figure 1). IL-6 plays a crucial role in regulating inflammation and can be secreted by macrophages, monocytes, and human peritoneal mesothelial cells. Exposure to dialysate and IL-1β can trigger the release of IL-6, which has been reported to increase proportionally with the glucose concentration in the dialysate (Milan Manani et al., 2016; Yang et al., 2020). IL-6, along with soluble IL-6 receptors, can facilitate the synthesis and secretion of MCP-1, MCP-3, and IL-8, as well as adhesion molecules and angiogenesis factors (Salgado et al., 2002; Bellón et al., 2011). IL-17, mainly produced by Th17 cells and γδ T cells, was detected in peritoneal biopsies of PD patients but not healthy subjects (Rodrigues-Díez et al., 2014). IL-17 can promote the production of ELR + CXC chemokines in mesothelial cells, such as CXCL1 and CXCL8. Furthermore, it is believed that IL-17 might directly stimulate mesothelial cells to produce VEGF via an unidentified mechanism (Witowski et al., 2018).
In addition, recent studies have begun to focus on the cyclic GMP-AMP synthase (cGAS)-stimulator of interferon genes (STING) pathway (Decout et al., 2021; Zhang et al., 2022b). Inflammation leads to damage to mitochondrial DNA (mtDNA), which can be released into the cytoplasm under cellular stress and recognized by various DNA sensing mechanisms, including TLR-9, cytoplasmic cGAS-STING signalling, and inflammasome activation, which can result in the development of fibrosis, as well as pathological angiogenesis and endothelial-to-mesenchymal transition (Wang et al., 2022a; Wang et al., 2022b). In the future, there may be more research on peritoneal fibrosis related to peritoneal dialysis that focuses on the cGAS-STING pathway.
The role of epigenetic modifications in fibrosis has garnered increasing recognition. Epigenetic regulation mainly includes DNA methylation, histone modification, and noncoding RNA regulation. It is widely believed that DNA methylation of cytosine-phosphate-guanine (CpG) dinucleotide sites near the gene promoter leads to target gene silencing. In 2017, Kim et al. found that DNMT1 (DNA methyltransferase 1) inhibits RASAL1 protein expression by promoting hypermethylation of RASAL1 (Ras GTPase activating-like protein 1), which in turn upregulates the expression of TGF-β1 and accelerates peritoneal thickening in vivo in experiments with encapsulating peritoneal sclerosis rats (Kim et al., 2014). Histone methylation has also been implicated in PF, as Maeda et al. demonstrated that the expression of H3K9 histone methyltransferase and H3K4 methyltransferase can exacerbate the thickening of the submesothelial compact zone (Maeda et al., 2017). Recent studies have also linked high expression of EZH2 (Zeste homologue enhancer 2) with peritoneal fibrosis, possibly through the activation of profibrotic signalling pathways (Wang et al., 2020; Shi et al., 2022).
Furthermore, there is a growing body of evidence indicating that different types of noncoding RNAs, including microRNAs (miRNAs) and long-chain noncoding RNAs (lncRNAs), are involved in the transcription of genes that regulate peritoneal fibrosis (Liu et al., 2019) (Figure 1). Studies have suggested that certain miRNAs, such as miR-129–5p and miRNA-302c, have a protective effect on the peritoneum during dialysis, while others, such as miRNA-30b, miRNA-23, and miRNA-21, can exacerbate peritoneal fibrosis (Xiao et al., 2015; Morishita et al., 2016a; Hirai et al., 2017; Lopez-Anton et al., 2017; Yanai et al., 2018; Zhang et al., 2019). The functions of miRNAs are diverse and complex, primarily involving the regulation of gene expression through interactions with DNA, RNA, proteins, or their combinatorial interactions involved in transcriptional and posttranscriptional regulation (Guo et al., 2020). Some researchers also believe that lncRNAs, such as BC049991 and AK080622, may participate in peritoneal fibrosis by influencing heat shock protein 72 (HSP72) (Bergmann and Spector, 2014). Additionally, lncRNA AV310809 promotes TGF-β1 by activating the Wnt2/β-catenin signalling pathway, which induces EMT in human peritoneal mesothelial cells (Bergmann and Spector, 2014). While circular RNAs (circRNAs) have been implicated in organ fibrosis, such as cardiac fibrosis, liver fibrosis, and pulmonary fibrosis (Li et al., 2019; Yousefi and Soltani, 2021), relatively little is known about their involvement in peritoneal fibrosis. It is possible that circRNAs may be involved in peritoneal fibrosis by inducing EMT, but further research is needed to determine the specific principles and molecular mechanisms.
Autophagy is a double-edged sword. On the one hand, it can be activated by cells to prevent organelle damage caused by reactive oxygen species (ROS) under certain external stimuli. However, it may also induce epithelial apoptosis or EMT. Studies have shown that autophagy initiation can prevent peritoneal tissue damage by blocking NLRP3/IL-1β-mediated inflammasome activation (Wu et al., 2016) (Figure 1).
Normally, autophagy activation and inhibition are in balance, and excessive activation or inhibition can lead to oxidative stress, inflammatory damage, and even fibrosis. In fact, it has been reported that rapamycin, an autophagy inducer, stimulates mTOR signalling in peritoneal mesodermal cells activated by hyperglycaemia, reducing EMT and improving peritoneal fibrosis (Wu et al., 2018). Conversely, high glucose peritoneal dialysate has been shown to induce autophagy and fibrosis in human peritoneal mesothelial cells (Wu et al., 2018). This may be involved in EMT, the proinflammatory response, and angiogenesis by regulating the TGF-β/Smad3, EGFR/ERK1/2, STAT3/NF-κB, and β-catenin axes (Shi et al., 2021), suggesting that autophagy plays a role in peritoneal fibrosis.
Autophagy is reported to contribute to apoptosis as type II programmed cell death via autodigestive cellular progression or extracellular stimulation (Ghavami et al., 2015; Zou et al., 2016). Some researchers have reported that high-glucose peritoneal dialysate induces Beclin 1-dependent autophagy in human peritoneal mesoepithelial cells (HPMCs) and that autophagy inhibition reduces EMT, fibrosis and apoptosis in HPMCs (Wu et al., 2018). It has also been suggested that promoting mitochondrial synthesis and inhibiting apoptosis can improve peritoneal fibrosis (Li et al., 2022).
Angiogenesis is considered to be a significant factor in peritoneal ultrafiltration failure (Schilte et al., 2009). Neovascularization increases the effective surface area for solute exchange and decreases osmotic pressure driven by glucose from the peritoneal dialysate, resulting in a decrease in ultrafiltration volume. Neovascularization is often associated with hypoxia in physiological or pathological states (Lei et al., 2012). Peritoneal mesenchymal cells overmetabolize glucose, leading to pseudohypoxia with increased NADH-NAD+ (oxidized-reduced nicotinamide dinucleotide ratio) in the cytosol (Krediet and Parikova, 2022) (Figure 1). This ratio promotes the expression of hypoxia-inducible factor-1 (HIF-1), which is one of the most important regulators in the hypoxia response. HIF-1 can regulate the transcription of various genes, mediate the occurrence of EMT, and thus participate in peritoneal fibrosis. YANG et al. have suggested that the activation of the HIF-1α/STAT3 signaling pathway is the main contributor to EMT of mesenchymal cells induced by high glucose, and that knockdown of HIF-1α could alleviate the EMT and fibrosis process (Yang et al., 2021). Yoshiyuki et al. found that hypoxia can promote the expression of HIF-1α, Snail-1, VEGF, and MMP-2 in rat mesenchymal cells, which induces EMT, and that HIF-1α inhibitors can diminish fibrosis by suppressing the expression of these factors (Morishita et al., 2016b). VEGF is a key player in peritoneal angiogenesis, and studies have shown that peritoneal dialysis solution with glucose is related to the growth of VEGF (Zweers et al., 2001). GDPs in PD solution can induce the formation of AGEs, and the AGE (RAGE) receptor plays a profibrotic role by mediating the activation of VEGF to induce capillary angiogenesis and promoting TGF-β-induced EMT (De Vriese et al., 2006; Boulanger et al., 2007). Angiogenesis is regulated by cell growth factors such as VEGF and basic fibroblast growth factor (bFGF), as well as statins such as angiostatin and endostatin (ES) (Kakuta et al., 2005; Tanabe et al., 2007; Nakao et al., 2010).
Apart from VEGF, several other factors contribute to peritoneal neovascularization. Aquaporin-1 (AQP 1), which is responsible for fluid transport, plays a vital role in angiogenesis and endothelial cell migration (Saadoun et al., 2005). Angiotensin 2 (Ang-2) is an angiogenic factor that has been found to be involved in peritoneal neovascularization in rats with EPS (Io et al., 2004). Prostaglandin E2 and MCP-1 enhance epithelial cell migration and induce the transcription of angiogenin-related genes, contributing to angiogenesis (Ito et al., 2000; Xiao et al., 2010). Moreover, peritoneal smooth muscle cells are activated after infection and induce the formation of new blood vessels by producing proangiogenic factors such as TGF-β, fibroblast growth factor, VEGF, tumour necrosis factor-α, and IL-8 (Aroeira et al., 2005). Inflammatory factors such as IL-1β and IL-6 may also participate in neovascularization by stimulating endothelial progenitor cell proliferation and inducing VEGF synthesis (Rosell et al., 2009; Catar et al., 2017).
Oxidative stress is a common occurrence in patients with chronic kidney disease (CKD) and its severity increases with the progression of the disease (Dounousi et al., 2006). In patients undergoing peritoneal dialysis, oxidative stress is mainly attributed to the bioincompatibility components of the peritoneal dialysis fluid, such as high glucose concentration, acidic pH, high osmolarity (Liakopoulos et al., 2017) and uraemic toxins (Castoldi et al., 2010). The accumulation of glucose degradation products (GDPs) in the peritoneum triggers the formation of excessive advanced glycation end products (AGEs), reactive oxygen species (ROS), and advanced oxidized protein products (AOPPs). The main pathway of ROS formation is through increased glucose oxidative metabolism triggered by GDPs and AGEs. The accumulation of AGEs in the peritoneal dialysis fluid promotes the expression and accumulation of specific multiligand transmembrane receptors, known as RAGE, in the peritoneum (Figure 1). These receptors induce morphological modifications of intracellular proteins, which alter the structure of the extracellular matrix (ECM) composition and the receptors expressed on the peritoneal cell membrane. Finally, ROS are produced when AGEs and AGE-modified proteins closely integrate with RAGE. Furthermore, ROS mediate the activation of proinflammatory factors, cytokines, transcription factors and growth factors, contributing to downstream aberrant gene transcription and apoptosis (Roumeliotis et al., 2020).
Si et al. found that PD fluid indeed induces metabolic reprogramming in the mouse peritoneum through conducting gene expression profiling and metabolomics analyses. Specifically, this reprogramming is characterized by a state of hyperglycolysis (Si et al., 2019). Correcting the metabolic state in mesothelial cells may be a therapeutic approach to treating peritoneal fibrosis (Si et al., 2019; Fu et al., 2022). GLUTs and SGLTs are involved in glucose transport and energy metabolism (Onishi et al., 2015; Han et al., 2022). The presence of high glucose levels in peritoneal dialysis fluid can potentially stimulate the expression of GLUT1 and SGLT1, leading to enhanced glucose absorption and a reduction of the osmotic gradient for ultrafiltration (Schröppel et al., 1998; Hong et al., 2016). Unfortunately, this process can ultimately contribute to the development of peritoneal fibrosis. With the continuous emergence of new hypoglycaemic drugs, some have found that SGLT-2 inhibitors reduce the absorption of glucose in peritoneal dialysate by inhibiting the activity of SGLT-2 (Zhou et al., 2019). They also significantly reduce the concentration of TGF-β, peritoneal thickening and fibrosis, and microvascular density, thus improving ultrafiltration (Balzer et al., 2020; Shentu et al., 2021). However, considering that the primary target of SGLT inhibitors in oral formulations is the renal tubules, it may be necessary to create more dosage forms, such as those that can be administered by intraperitoneal injection or added to peritoneal dialysis fluid, to improve peritoneal fibrosis.
In general, there are many factors involved in the development of peritoneal fibrosis in patients with abdominal dialysis, and the molecular signalling pathways involved are also complex, with interactions and influences on each other. Several factors are recognized for their importance in the development of peritoneal fibrosis in patients with PD. The most important is the use of traditional biocompatible peritoneal dialysis, which contains high glucose concentrations and glucose degradation products. Growth factor TGF-β1 is transformed by inducing many profibrotic events, including epithelium-mesenchymal transformation, fibroblast proliferation, and extracellular matrix deposition, playing a central role in peritoneal fibrosis. A solution containing high glucose stimulates the synthesis of TGF-β1 by activating protein kinase C in peritoneal cells and induces TGF-β type I and type II receptors, which induce various fibrosis-related signalling molecular transport pathways, including Smad-dependent and Smad-independent signalling pathways. Increasing studies on epigenetic regulation, hypoxic stimulation, and neovascularization are beginning to confirm their involvement. Peritonitis promotes peritoneal fibrosis through inflammatory factors such as IL-1β and IL-6, exacerbating the chronic induction of TGF-β1 synthesis (Kang et al., 1999), and promoting fibrosis events. Among the many factors involved in peritoneal fibrosis, EMT, the inflammatory response, autophagy, epigenetic regulation, and neovascularization are the dominant factors in fibrosis, of which TGF-β1 plays an important role in the activation of various signalling pathways, the induction of inflammatory factors, and the hypoxia stress response. Correspondingly, many scholars increasingly emphasize the proposed peritoneal mesothelial cell markers that reflect the inflammatory state of the peritoneum and the degree of fibrosis, and the application of clinical evaluation of peritoneal ultrafiltration function is an important part of the evaluation of the treatment effect. Carbohydrate antigen CA125, CTGF, suppression of tumorigenicity 2, MMP-2, and microRNAs (Mizutani et al., 2010; Ge et al., 2012; Barreto et al., 2013; Krediet and Struijk, 2013; Kim et al., 2019) are some of the biomarkers that have been proposed thus far, and the specificity and sensitivity of their diagnosis still need to be confirmed by more studies. Based on the above-related precipitating factors in the final treatment, the proposed peritoneal fibrosis treatments mainly include the use of biocompatible dialysate, tyrosine kinase inhibitors, inflammatory factor blockers, renin-angiotensin system inhibitors, and immunosuppressants, but most of these are still being studied in animal experiments or early clinical studies and are not widely used in clinical practice. In the future, more research is needed to further supplement the research targets and specific and detailed molecular mechanisms related to peritoneal fibrosis, and more clinical therapeutic drug intervention experiments will make greater contributions to delaying peritoneal fibrosis and providing more effective abdominal dialysis therapy for patients with PD.
JL crafted and authored the manuscript, while YL contributed valuable ideas, and JPL meticulously revised and thoroughly reviewed the document. All authors contributed to the article and approved the submitted version.
This work was supported by the National Natural Science Foundation of China (Grant Nos. 82160162, and 81760150) and the Key Project of Jiangxi Provincial Natural Science Foundation (Grant No. 20202ACBL206008).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Aroeira L. S., Aguilera A., Selgas R., Ramírez-Huesca M., Pérez-Lozano M. L., Cirugeda A., et al. (2005). Mesenchymal conversion of mesothelial cells as a mechanism responsible for high solute transport rate in peritoneal dialysis: role of vascular endothelial growth factor. Am. J. kidney Dis. 46, 938–948. the official journal of the National Kidney Foundation. doi:10.1053/j.ajkd.2005.08.011
Bajo M. A., Del Peso G., Teitelbaum I. (2017). Peritoneal membrane preservation. Seminars Nephrol. 37, 77–92. doi:10.1016/j.semnephrol.2016.10.009
Balzer M. S., Rong S., Nordlohne J., Zemtsovski J. D., Schmidt S., Stapel B., et al. (2020). SGLT2 inhibition by intraperitoneal dapagliflozin mitigates peritoneal fibrosis and ultrafiltration failure in a mouse model of chronic peritoneal exposure to high-glucose dialysate. Biomolecules 10, 1573. doi:10.3390/biom10111573
Balzer M. S. (2020). Molecular pathways in peritoneal fibrosis. Cell. Signal. 75, 109778. doi:10.1016/j.cellsig.2020.109778
Barreto D. L., Coester A. M., Struijk D. G., Krediet R. T. (2013). Can effluent matrix metalloproteinase 2 and plasminogen activator inhibitor 1 be used as biomarkers of peritoneal membrane alterations in peritoneal dialysis patients? Perit. Dial. Int. 33, 529–537. journal of the International Society for Peritoneal Dialysis. doi:10.3747/pdi.2012.01063
Bellón T., Martínez V., Lucendo B., del Peso G., Castro M. J., Aroeira L. S., et al. (2011). Alternative activation of macrophages in human peritoneum: implications for peritoneal fibrosis. Nephrol. Dial. Transplant. 26, 2995–3005. official publication of the European Dialysis and Transplant Association - European Renal Association. doi:10.1093/ndt/gfq771
Bergmann J. H., Spector D. L. (2014). Long non-coding RNAs: modulators of nuclear structure and function. Curr. Opin. Cell. Biol. 26, 10–18. doi:10.1016/j.ceb.2013.08.005
Boulanger E., Grossin N., Wautier M. P., Taamma R., Wautier J. L. (2007). Mesothelial RAGE activation by AGEs enhances VEGF release and potentiates capillary tube formation. Kidney Int. 71, 126–133. doi:10.1038/sj.ki.5002016
Castoldi G., Antolini L., Bombardi C., Perego L., Mariani P., Viganò M. R., et al. (2010). Oxidative stress biomarkers and chromogranin A in uremic patients: effects of dialytic treatment. Clin. Biochem. 43, 1387–1392. doi:10.1016/j.clinbiochem.2010.08.028
Catar R., Witowski J., Zhu N., Lücht C., Derrac Soria A., Uceda Fernandez J., et al. (2017). IL-6 trans-signaling links inflammation with angiogenesis in the peritoneal membrane. J. Am. Soc. Nephrol. 28, 1188–1199. doi:10.1681/ASN.2015101169
Cueto-Manzano A. M., González-Espinoza L., Martin del Campo F., Fortes P. C., Pecoits-Filho R. (2007). Inflammation in peritoneal dialysis: A Latin-American perspective. Perit. Dial. Int. 27, 347–352. journal of the International Society for Peritoneal Dialysis. doi:10.1177/089686080702700326
De Vriese A. S., Tilton R. G., Mortier S., Lameire N. H. (2006). Myofibroblast transdifferentiation of mesothelial cells is mediated by RAGE and contributes to peritoneal fibrosis in uraemia. Nephrol. Dial. Transplant. 21, 2549–2555. official publication of the European Dialysis and Transplant Association - European Renal Association. doi:10.1093/ndt/gfl271
Decout A., Katz J. D., Venkatraman S., Ablasser A. (2021). The cGAS-STING pathway as a therapeutic target in inflammatory diseases. Nat. Rev. Immunol. 21, 548–569. doi:10.1038/s41577-021-00524-z
Derynck R., Zhang Y. E. (2003). Smad-dependent and Smad-independent pathways in TGF-beta family signalling. Nature 425, 577–584. doi:10.1038/nature02006
Dounousi E., Papavasiliou E., Makedou A., Ioannou K., Katopodis K. P., Tselepis A., et al. (2006). Oxidative stress is progressively enhanced with advancing stages of CKD. Am. J. kidney Dis. 48, 752–760. the official journal of the National Kidney Foundation. doi:10.1053/j.ajkd.2006.08.015
Fu J., Li N., He M., Huang D., Zhang P. (2022). STAT3 signaling mediates peritoneal fibrosis by activating hyperglycolysis. Am. J. Transl. Res. 14, 7552–7565.
Ge Y., Xiao L., Chen X., Peng Y., Sun L., Liu F. (2012). MicroRNAs in peritoneal dialysis effluent are promising biomarkers for peritoneal fibrosis in peritoneal dialysis patients. Med. hypotheses 78, 155–156. doi:10.1016/j.mehy.2011.10.015
Ghavami S., Cunnington R. H., Gupta S., Yeganeh B., Filomeno K. L., Freed D. H., et al. (2015). Autophagy is a regulator of TGF-β1-induced fibrogenesis in primary human atrial myofibroblasts. Cell. death Dis. 6, e1696. doi:10.1038/cddis.2015.36
Guo Y., Wang L., Gou R., Tang L., Liu P. (2020). Noncoding RNAs in peritoneal fibrosis: background, mechanism, and therapeutic approach. Biomed. Pharmacother. = Biomedecine Pharmacother. 129, 110385. doi:10.1016/j.biopha.2020.110385
Han L., Qu Q., Aydin D., Panova O., Robertson M. J., Xu Y., et al. (2022). Structure and mechanism of the SGLT family of glucose transporters. Nature 601, 274–279. doi:10.1038/s41586-021-04211-w
Hautem N., Morelle J., Sow A., Corbet C., Feron O., Goffin E., et al. (2017). The NLRP3 inflammasome has a critical role in peritoneal dialysis-related peritonitis. J. Am. Soc. Nephrol. 28, 2038–2052. doi:10.1681/ASN.2016070729
Hirahara I., Ishibashi Y., Kaname S., Kusano E., Fujita T. (2009). Methylglyoxal induces peritoneal thickening by mesenchymal-like mesothelial cells in rats. Nephrol. Dial. Transplant. 24, 437–447. official publication of the European Dialysis and Transplant Association - European Renal Association. doi:10.1093/ndt/gfn495
Hirai K., Yoshizawa H., Imai T., Igarashi Y., Hirahara I., Ookawara S., et al. (2017). Detection of microRNA expression in peritoneal membrane of rats using quantitative real-time PCR. J. Vis. Exp., 55505. doi:10.3791/55505
Hong M., Nie Z., Chen Z., Yu X., Bao B. (2016). High glucose dialysate enhances peritoneal fibrosis through upregulating glucose transporters GLUT1 and SGLT1. Zhejiang da xue xue bao. Yi xue ban = J. Zhejiang Univ. Med. Sci. 45, 598–606. doi:10.3785/j.issn.1008-9292.2016.11.07
Huang Q., Xiao R., Lu J., Zhang Y., Xu L., Gao J., et al. (2022). Endoglin aggravates peritoneal fibrosis by regulating the activation of TGF-β/ALK/Smads signaling. Front. Pharmacol. 13, 973182. doi:10.3389/fphar.2022.973182
Io H., Hamada C., Ro Y., Ito Y., Hirahara I., Tomino Y. (2004). Morphologic changes of peritoneum and expression of VEGF in encapsulated peritoneal sclerosis rat models. Kidney Int. 65, 1927–1936. doi:10.1111/j.1523-1755.2004.00599.x
Ito T., Yorioka N., Yamamoto M., Kataoka K., Yamakido M. (2000). Effect of glucose on intercellular junctions of cultured human peritoneal mesothelial cells. J. Am. Soc. Nephrol. 11, 1969–1979. doi:10.1681/ASN.V11111969
Jang Y. H., Shin H. S., Sun Choi H., Ryu E. S., Jin Kim M., Ki Min S., et al. (2013). Effects of dexamethasone on the TGF-β1-induced epithelial-to-mesenchymal transition in human peritoneal mesothelial cells. Laboratory investigation; a J. Tech. methods pathology 93, 194–206. doi:10.1038/labinvest.2012.166
Kakuta T., Tanaka R., Satoh Y., Izuhara Y., Inagi R., Nangaku M., et al. (2005). Pyridoxamine improves functional, structural, and biochemical alterations of peritoneal membranes in uremic peritoneal dialysis rats. Kidney Int. 68, 1326–1336. doi:10.1111/j.1523-1755.2005.00531.x
Kalluri R., Neilson E. G. (2003). Epithelial-mesenchymal transition and its implications for fibrosis. J. Clin. investigation 112, 1776–1784. doi:10.1172/JCI20530
Kang D. H., Hong Y. S., Lim H. J., Choi J. H., Han D. S., Yoon K. I. (1999). High glucose solution and spent dialysate stimulate the synthesis of transforming growth factor-β1 of human peritoneal mesothelial cells: effect of cytokine costimulation. Perit. Dial. Int. 19, 221–230. journal of the International Society for Peritoneal Dialysis. doi:10.1177/089686089901900307
Kato S., Yuzawa Y., Tsuboi N., Maruyama S., Morita Y., Matsuguchi T., et al. (2004). Endotoxin-induced chemokine expression in murine peritoneal mesothelial cells: the role of toll-like receptor 4. J. Am. Soc. Nephrol. 15, 1289–1299.
Kim K. H., Ryu H. M., Oh S. H., Oh E. J., Ahn J. S., Lee J. H., et al. (2014). Effect of DNA demethylation in experimental encapsulating peritoneal sclerosis. Ther. Apher. dialysis 18, 628–636. official peer-reviewed journal of the International Society for Apheresis, the Japanese Society for Apheresis, the Japanese Society for Dialysis Therapy. doi:10.1111/1744-9987.12186
Kim Y. C., Kim K. H., Lee S., Jo J. W., Park J. Y., Park M. S., et al. (2019). ST2 blockade mitigates peritoneal fibrosis induced by TGF-β and high glucose. J. Cell. Mol. Med. 23, 6872–6884. doi:10.1111/jcmm.14571
Krediet R. T., Parikova A. (2022). Relative contributions of pseudohypoxia and inflammation to peritoneal alterations with long-term peritoneal dialysis patients. Clin. J. Am. Soc. Nephrol. CJASN 17, 1259–1266. doi:10.2215/CJN.15371121
Krediet R. T., Struijk D. G. (2013). Peritoneal changes in patients on long-term peritoneal dialysis. Nat. Rev. Nephrol. 9, 419–429. doi:10.1038/nrneph.2013.99
Lamouille S., Xu J., Derynck R. (2014). Molecular mechanisms of epithelial-mesenchymal transition. Nat. Rev. Mol. Cell. Biol. 15, 178–196. doi:10.1038/nrm3758
Lei P., Jiang Z., Zhu H., Li X., Su N., Yu X. (2012). Poly(ADP-ribose) polymerase-1 in high glucose-induced epithelial-mesenchymal transition during peritoneal fibrosis. Int. J. Mol. Med. 29, 472–478. doi:10.3892/ijmm.2011.859
Lho Y., Do J. Y., Heo J. Y., Kim A. Y., Kim S. W., Kang S. H. (2021). Effects of TGF-β1 receptor inhibitor GW788388 on the epithelial to mesenchymal transition of peritoneal mesothelial cells. Int. J. Mol. Sci. 22, 4739. doi:10.3390/ijms22094739
Li P. K., Chow K. M., Van de Luijtgaarden M. W., Johnson D. W., Jager K. J., Mehrotra R., et al. (2017). Changes in the worldwide epidemiology of peritoneal dialysis. Nat. Rev. Nephrol. 13, 90–103. doi:10.1038/nrneph.2016.181
Li C., Wang Z., Zhang J., Zhao X., Xu P., Liu X., et al. (2019). Crosstalk of mRNA, miRNA, lncRNA, and circRNA and their regulatory pattern in pulmonary fibrosis. Mol. Ther. Nucleic acids 18, 204–218. doi:10.1016/j.omtn.2019.08.018
Li Z. H., Xu R., Shi J., Yu M. S., Zhong Y., He W. M., et al. (2022). Astragalus total saponins ameliorate peritoneal fibrosis by promoting mitochondrial synthesis and inhibiting apoptosis. Am. J. Chin. Med. 50, 261–274. doi:10.1142/S0192415X22500094
Liakopoulos V., Roumeliotis S., Gorny X., Eleftheriadis T., Mertens P. R. (2017). Oxidative stress in patients undergoing peritoneal dialysis: A current review of the literature. Oxidative Med. Cell. Longev. 2017, 3494867. doi:10.1155/2017/3494867
Liu Q., Zhang Y., Mao H., Chen W., Luo N., Zhou Q., et al. (2012). A crosstalk between the Smad and JNK signaling in the TGF-β-induced epithelial-mesenchymal transition in rat peritoneal mesothelial cells. PloS one 7, e32009. doi:10.1371/journal.pone.0032009
Liu W., Feng R., Li X., Li D., Zhai W. (2019). TGF-β- and lipopolysaccharide-induced upregulation of circular RNA PWWP2A promotes hepatic fibrosis via sponging miR-203 and miR-223. Aging 11, 9569–9580. doi:10.18632/aging.102405
Lopez-Anton M., Lambie M., Lopez-Cabrera M., Schmitt C. P., Ruiz-Carpio V., Bartosova M., et al. (2017). miR-21 promotes fibrogenesis in peritoneal dialysis. Am. J. pathology 187, 1537–1550. doi:10.1016/j.ajpath.2017.03.007
Maeda K., Doi S., Nakashima A., Nagai T., Irifuku T., Ueno T., et al. (2017). Inhibition of H3K9 methyltransferase G9a ameliorates methylglyoxal-induced peritoneal fibrosis. PloS one 12, e0173706. doi:10.1371/journal.pone.0173706
Masola V., Bonomini M., Borrelli S., Di Liberato L., Vecchi L., Onisto M., et al. (2022). Fibrosis of peritoneal membrane as target of new therapies in peritoneal dialysis. Int. J. Mol. Sci. 23, 4831. doi:10.3390/ijms23094831
Milan Manani S., Virzì G. M., Clementi A., Brocca A., de Cal M., Tantillo I., et al. (2016). Pro-inflammatory cytokines: A possible relationship with dialytic adequacy and serum albumin in peritoneal dialysis patients. Clin. kidney J. 9, 153–157. doi:10.1093/ckj/sfv137
Mizutani M., Ito Y., Mizuno M., Nishimura H., Suzuki Y., Hattori R., et al. (2010). Connective tissue growth factor (CTGF/CCN2) is increased in peritoneal dialysis patients with high peritoneal solute transport rate. Am. J. physiology. Ren. physiology 298, F721–F733. doi:10.1152/ajprenal.00368.2009
Morishita Y., Yoshizawa H., Watanabe M., Imai R., Imai T., Hirahara I., et al. (2016a). MicroRNA expression profiling in peritoneal fibrosis. Transl. Res. J. laboratory Clin. Med. 169, 47–66. doi:10.1016/j.trsl.2015.10.009
Morishita Y., Ookawara S., Hirahara I., Muto S., Nagata D. (2016b). HIF-1α mediates Hypoxia-induced epithelial-mesenchymal transition in peritoneal mesothelial cells. Ren. Fail. 38, 282–289. doi:10.3109/0886022X.2015.1127741
Nakao A., Nakao K., Takatori Y., Kojo S., Inoue J., Akagi S., et al. (2010). Effects of icodextrin peritoneal dialysis solution on the peritoneal membrane in the STZ-induced diabetic rat model with partial nephrectomy. Nephrol. Dial. Transplant. 25, 1479–1488. official publication of the European Dialysis and Transplant Association - European Renal Association. doi:10.1093/ndt/gfp479
Onishi A., Akimoto T., Urabe M., Hirahara I., Muto S., Ozawa K., et al. (2015). Attenuation of methylglyoxal-induced peritoneal fibrosis: immunomodulation by interleukin-10. a J. Tech. methods pathology 95, 1353–1362. doi:10.1038/labinvest.2015.110
Rodrigues-Díez R., Aroeira L. S., Orejudo M., Bajo M. A., Heffernan J. J., Rodrigues-Díez R. R., et al. (2014). IL-17A is a novel player in dialysis-induced peritoneal damage. Kidney Int. 86, 303–315. doi:10.1038/ki.2014.33
Rosell A., Arai K., Lok J., He T., Guo S., Navarro M., et al. (2009). Interleukin-1beta augments angiogenic responses of murine endothelial progenitor cells in vitro. J. Cereb. blood flow metabolism 29, 933–943. official journal of the International Society of Cerebral Blood Flow and Metabolism. doi:10.1038/jcbfm.2009.17
Roumeliotis S., Dounousi E., Salmas M., Eleftheriadis T., Liakopoulos V. (2020). Unfavorable effects of peritoneal dialysis solutions on the peritoneal membrane: the role of oxidative stress. Biomolecules 10, 768. doi:10.3390/biom10050768
Saadoun S., Papadopoulos M. C., Hara-Chikuma M., Verkman A. S. (2005). Impairment of angiogenesis and cell migration by targeted aquaporin-1 gene disruption. Nature 434, 786–792. doi:10.1038/nature03460
Salgado R., Benoy I., Weytjens R., Van Bockstaele D., Van Marck E., Huget P., et al. (2002). Arterio-venous gradients of IL-6, plasma and serum VEGF and D-dimers in human cancer. Br. J. cancer 87, 1437–1444. doi:10.1038/sj.bjc.6600655
Schilte M. N., Celie J. W., Wee P. M., Beelen R. H., van den Born J. (2009). Factors contributing to peritoneal tissue remodeling in peritoneal dialysis. Perit. Dial. Int. 29, 605–617. journal of the International Society for Peritoneal Dialysis. doi:10.1177/089686080902900604
Schröppel B., Fischereder M., Wiese P., Segerer S., Huber S., Kretzler M., et al. (1998). Expression of glucose transporters in human peritoneal mesothelial cells. Kidney Int. 53, 1278–1287. doi:10.1046/j.1523-1755.1998.00899.x
Shentu Y., Li Y., Xie S., Jiang H., Sun S., Lin R., et al. (2021). Empagliflozin, a sodium glucose cotransporter-2 inhibitor, ameliorates peritoneal fibrosis via suppressing TGF-β/Smad signaling. Int. Immunopharmacol. 93, 107374. doi:10.1016/j.intimp.2021.107374
Shi Y., Hu Y., Wang Y., Ma X., Tang L., Tao M., et al. (2021). Blockade of autophagy prevents the development and progression of peritoneal fibrosis. Front. Pharmacol. 12, 724141. doi:10.3389/fphar.2021.724141
Shi Y., Li J., Chen H., Hu Y., Tang L., Wang Y., et al. (2022). Inhibition of EZH2 suppresses peritoneal angiogenesis by targeting a VEGFR2/ERK1/2/HIF-1α-dependent signaling pathway. J. pathology 258, 164–178. doi:10.1002/path.5987
Si M., Wang Q., Li Y., Lin H., Luo D., Zhao W., et al. (2019). Inhibition of hyperglycolysis in mesothelial cells prevents peritoneal fibrosis. Sci. Transl. Med. 11, eaav5341. doi:10.1126/scitranslmed.aav5341
Strippoli R., Loureiro J., Moreno V., Benedicto I., Lozano M. L., Barreiro O., et al. (2015). Caveolin-1 deficiency induces a MEK-ERK1/2-Snail-1-dependent epithelial-mesenchymal transition and fibrosis during peritoneal dialysis. EMBO Mol. Med. 7, 102–123. doi:10.15252/emmm.201404127
Suryantoro S. D., Thaha M., Sutanto H., Firdausa S. (2023). Current insights into cellular determinants of peritoneal fibrosis in peritoneal dialysis: A narrative review. J. Clin. Med. 12, 4401. doi:10.3390/jcm12134401
Tanabe K., Maeshima Y., Ichinose K., Kitayama H., Takazawa Y., Hirokoshi K., et al. (2007). Endostatin peptide, an inhibitor of angiogenesis, prevents the progression of peritoneal sclerosis in a mouse experimental model. Kidney Int. 71, 227–238. doi:10.1038/sj.ki.5002040
Wang L., Balzer M. S., Rong S., Menne J., von Vietinghoff S., Dong L., et al. (2016). Protein kinase C α inhibition prevents peritoneal damage in a mouse model of chronic peritoneal exposure to high-glucose dialysate. Kidney Int. 89, 1253–1267. doi:10.1016/j.kint.2016.01.025
Wang Q., Xu L., Zhang X., Liu D., Wang R. (2020). GSK343, an inhibitor of EZH2, mitigates fibrosis and inflammation mediated by HIF-1α in human peritoneal mesothelial cells treated with high glucose. Eur. J. Pharmacol. 880, 173076. doi:10.1016/j.ejphar.2020.173076
Wang Q., Bu Q., Liu M., Zhang R., Gu J., Li L., et al. (2022a). XBP1-mediated activation of the STING signalling pathway in macrophages contributes to liver fibrosis progression. JHEP Rep. innovation hepatology 4, 100555. doi:10.1016/j.jhepr.2022.100555
Wang L., Zhang Y., Ren Y., Yang X., Ben H., Zhao F., et al. (2022b). Pharmacological targeting of cGAS/STING-YAP axis suppresses pathological angiogenesis and ameliorates organ fibrosis. Eur. J. Pharmacol. 932, 175241. doi:10.1016/j.ejphar.2022.175241
Witowski J., Kamhieh-Milz J., Kawka E., Catar R., Jörres A. (2018). IL-17 in peritoneal dialysis-associated inflammation and angiogenesis: conclusions and perspectives. Front. physiology 9, 1694. doi:10.3389/fphys.2018.01694
Wu J., Li X., Zhu G., Zhang Y., He M., Zhang J. (2016). The role of Resveratrol-induced mitophagy/autophagy in peritoneal mesothelial cells inflammatory injury via NLRP3 inflammasome activation triggered by mitochondrial ROS. Exp. Cell. Res. 341, 42–53. doi:10.1016/j.yexcr.2016.01.014
Wu J., Xing C., Zhang L., Mao H., Chen X., Liang M., et al. (2018). Autophagy promotes fibrosis and apoptosis in the peritoneum during long-term peritoneal dialysis. J. Cell. Mol. Med. 22, 1190–1201. doi:10.1111/jcmm.13393
Xia Y., Wan C., Zhang Q., Wang H., Feng Y., Jiang C. (2022). Role of IGF-1R in epithelial-mesenchymal transdifferentiation of human peritoneal mesothelial cells. Clin. Exp. Nephrol. 26, 630–639. doi:10.1007/s10157-022-02209-w
Xiao L., Sun L., Liu F. Y., Peng Y. M., Duan S. B. (2010). Connective tissue growth factor knockdown attenuated matrix protein production and vascular endothelial growth factor expression induced by transforming growth factor-beta1 in cultured human peritoneal mesothelial cells. Ther. Apher. dialysis 14, 27–34. official peer-reviewed journal of the International Society for Apheresis, the Japanese Society for Apheresis, the Japanese Society for Dialysis Therapy. doi:10.1111/j.1744-9987.2009.00701.x
Xiao L., Zhou X., Liu F., Hu C., Zhu X., Luo Y., et al. (2015). MicroRNA-129-5p modulates epithelial-to-mesenchymal transition by targeting SIP1 and SOX4 during peritoneal dialysis. Laboratory investigation; a J. Tech. methods pathology 95, 817–832. doi:10.1038/labinvest.2015.57
Yanai K., Ishii H., Aomatsu A., Ishibashi K., Morishita Y. (2018). MicroRNAs in peritoneal fibrosis: A systematic review. Discov. Med. 26, 271–280.
Yang X., Yan H., Jiang N., Yu Z., Yuan J., Ni Z., et al. (2020). IL-6 trans-signaling drives a STAT3-dependent pathway that leads to structural alterations of the peritoneal membrane. Am. J. physiology. Ren. physiology 318, F338–f353. doi:10.1152/ajprenal.00319.2019
Yang X., Bao M., Fang Y., Yu X., Ji J., Ding X. (2021). STAT3/HIF-1α signaling activation mediates peritoneal fibrosis induced by high glucose. J. Transl. Med. 19, 283. doi:10.1186/s12967-021-02946-8
Yousefi F., Soltani B. M. (2021). Circular RNAs as potential theranostics in the cardiac fibrosis. Heart Fail. Rev. 26, 195–203. doi:10.1007/s10741-019-09908-9
Zhang F., Liu H., Liu F., Peng Y., Chen M., Liu Y., et al. (2013). New insights into the pathogenesis and treatment of peritoneal fibrosis: A potential role of wnt/β-catenin induced epithelial to mesenchymal transition and stem cells for therapy. Med. hypotheses 81, 97–100. doi:10.1016/j.mehy.2013.03.036
Zhang X. W., Wang L., Ding H. (2019). Long noncoding RNA AK089579 inhibits epithelial-to-mesenchymal transition of peritoneal mesothelial cells by competitively binding to microRNA-296-3p via DOK2 in peritoneal fibrosis. FASEB J. 33, 5112–5125. official publication of the Federation of American Societies for Experimental Biology. doi:10.1096/fj.201801111RR
Zhang Y., Feng W., Peng X., Zhu L., Wang Z., Shen H., et al. (2022a). Parthenolide alleviates peritoneal fibrosis by inhibiting inflammation via the NF-κB/TGF-β/Smad signaling axis. Laboratory investigation; a J. Tech. methods pathology 102, 1346–1354. doi:10.1038/s41374-022-00834-3
Zhang D., Liu Y., Zhu Y., Zhang Q., Guan H., Liu S., et al. (2022b). A non-canonical cGAS-STING-PERK pathway facilitates the translational program critical for senescence and organ fibrosis. Nat. Cell. Biol. 24, 766–782. doi:10.1038/s41556-022-00894-z
Zhou Y., Fan J., Zheng C., Yin P., Wu H., Li X., et al. (2019). SGLT-2 inhibitors reduce glucose absorption from peritoneal dialysis solution by suppressing the activity of SGLT-2. Biomed. Pharmacother. = Biomedecine Pharmacother. 109, 1327–1338. doi:10.1016/j.biopha.2018.10.106
Zou M., Wang F., Gao R., Wu J., Ou Y., Chen X., et al. (2016). Autophagy inhibition of hsa-miR-19a-3p/19b-3p by targeting TGF-β R II during TGF-β1-induced fibrogenesis in human cardiac fibroblasts. Sci. Rep. 6, 24747. doi:10.1038/srep24747
Keywords: peritoneal dialysis, peritoneal fibrosis, EMT, TGF-β1, mechanism
Citation: Li J, Liu Y and Liu J (2023) A review of research progress on mechanisms of peritoneal fibrosis related to peritoneal dialysis. Front. Physiol. 14:1220450. doi: 10.3389/fphys.2023.1220450
Received: 10 May 2023; Accepted: 13 September 2023;
Published: 25 September 2023.
Edited by:
Paola Pontrelli, University of Bari Aldo Moro, ItalyReviewed by:
Jianhui Zhou, Chinese PLA General Hospital, ChinaCopyright © 2023 Li, Liu and Liu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jianping Liu, bmRlZnkxNDEwNUBuY3UuZWR1LmNu
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.