 Pura Bolaños
Pura Bolaños Juan C. Calderón
Juan C. Calderón- 1Laboratory of Cellular Physiology, Centre of Biophysics and Biochemistry, Venezuelan Institute for Scientific Research (IVIC), Caracas, Venezuela
- 2Physiology and Biochemistry Research Group-PHYSIS, Faculty of Medicine, University of Antioquia, Medellín, Colombia
The excitation–contraction coupling (ECC) in skeletal muscle refers to the Ca2+-mediated link between the membrane excitation and the mechanical contraction. The initiation and propagation of an action potential through the membranous system of the sarcolemma and the tubular network lead to the activation of the Ca2+-release units (CRU): tightly coupled dihydropyridine and ryanodine (RyR) receptors. The RyR gating allows a rapid, massive, and highly regulated release of Ca2+ from the sarcoplasmic reticulum (SR). The release from triadic places generates a sarcomeric gradient of Ca2+ concentrations ([Ca2+]) depending on the distance of a subcellular region from the CRU. Upon release, the diffusing Ca2+ has multiple fates: binds to troponin C thus activating the contractile machinery, binds to classical sarcoplasmic Ca2+ buffers such as parvalbumin, adenosine triphosphate and, experimentally, fluorescent dyes, enters the mitochondria and the SR, or is recycled through the Na+/Ca2+ exchanger and store-operated Ca2+ entry (SOCE) mechanisms. To commemorate the 7th decade after being coined, we comprehensively and critically reviewed “old”, historical landmarks and well-established concepts, and blended them with recent advances to have a complete, quantitative-focused landscape of the ECC. We discuss the: 1) elucidation of the CRU structures at near-atomic resolution and its implications for functional coupling; 2) reliable quantification of peak sarcoplasmic [Ca2+] using fast, low affinity Ca2+ dyes and the relative contributions of the Ca2+-binding mechanisms to the whole concert of Ca2+ fluxes inside the fibre; 3) articulation of this novel quantitative information with the unveiled structural details of the molecular machinery involved in mitochondrial Ca2+ handing to understand how and how much Ca2+ enters the mitochondria; 4) presence of the SOCE machinery and its different modes of activation, which awaits understanding of its magnitude and relevance in situ; 5) pharmacology of the ECC, and 6) emerging topics such as the use and potential applications of super-resolution and induced pluripotent stem cells (iPSC) in ECC. Blending the old with the new works better!
1 Introduction
By the 1940s, some evidence was published considering the existence of a link between the excitation and the contraction of the skeletal muscle. However, it was very scarce, fraught with technical limitations and in some cases speculative, although considered “plausible”. Alexander Sandow stated in 1952 that the muscle “dependence of contraction on excitation indicates that there must be some process that is initiated in the excited membrane and which by moving inward makes contact with the contractile elements so as to initiate contraction. We shall designate the entire sequence of reactions -excitation, inward acting link, and activation of contraction- by the term excitation-contraction (E-C) coupling” (Sandow, 1952). This topic was so interesting by that time among researchers, that less than 20 years later the nature of the “inward acting link” was clear.
Since then, we have seen the development of a huge amount of technology with increasingly improved temporal and spatial resolution, which has been applied to gain insight into the excitation-contraction coupling (ECC) in skeletal muscle and have helped us outline the current landscape of this phenomenon.
Here, we present the basics of the ECC in skeletal muscle under physiological conditions, highlighting recent exciting conceptual advances and technical developments for its study. We present information gathered in vertebrate models but focused on mammalian ECC. Blending the old with the new allows us to commemorate 7 decades of hard work by honoring notable, classical contributions from many researchers in the field, but also by integrating new advances performed by intrepid young generations. We believe this is the best way to yield the most comprehensive possible picture of the ECC in skeletal muscle.
2 Ultrastructure, molecular machinery, and events of the excitation–contraction coupling mechanism in skeletal muscle
2.1 Historical landmarks
The events responsible for the abrupt muscle transition from rest to contraction occupied famous physiologists back in the 40 s of the 20th Century. Two well accepted concepts by that time were that the sarcolemma had excitatory properties (assumed to be limited only to the surface of the sarcolemma) and that soon after excited, the active state of the muscle was established (Kuffler, 1947; Hill, 1948; Hill, 1949). Moreover, there was a causal relationship because the kinetics of the contractile responses were modulated by the excitation (Kuffler, 1947; Sandow, 1952; Huxley and Taylor, 1958; Caputo, 2011). The activation seemed not to follow directly the excitation, because a longitudinal current and its associated electric field failed to induce a contractile response (Kuffler, 1947; Sten-Knudsen, 1954). An “inward acting link” (Sandow, 1952) between both, the excitation and the contraction, seemed thus necessary for many authors (Kuffler, 1947; Hill, 1948), mainly because they happen in parts of the cell separated by microns. Also, relevant information about a sizeable, yet manipulable, milliseconds delay between excitation and contraction, when several phenomena could be measured (Hill, 1949; Sandow, 1952; Weber and Portzehl, 1954), further suggested that a real link should exist. The duration of that window time was temperature-dependent, for instance, and there was a heat associated to activation which appeared after that latent period, before the shortening heat itself was measured (Hill, 1949; Sandow, 1952). A chemical or energetical link seemed feasible, but “to resolve its occurrence in times of the order of a few milliseconds would be one of fantastic difficulty” (Hill, 1949).
Simple ionic, radial diffusion from the sarcolemma caused by the longitudinal field of the action potential was not the link responsible for the activation of the whole fibre (Hill, 1948; Sten-Knudsen, 1954). Hill did his calculations with Ca2+ probably influenced by the results of Heilbrunn and Wiercinski (Heilbrunn and Wiercinski, 1947), who were “interested in knowing which substances will cause a shortening or contraction of the living protoplasm inside the muscle cell” (Heilbrunn and Wiercinski, 1947). According to their results, Ca2+ but neither Na+, K+, nor Mg2+, induced a notable muscle shortening when injected (Heilbrunn and Wiercinski, 1947).
It then became obvious that the link and its working mechanism was something else than the simple diffusion of a “contractile substance” released from the sarcolemma. Since the activation followed the points where the action potential passed (Sandow, 1952; Sten-Knudsen, 1954), the link should be a more complex process or may have a structural component. As most of the study models employed by that time were not mammalian, that structural element was associated to the Z lines at the I bands (Huxley and Taylor, 1958). Finishing the 50 s, and before the T-tubules (TT) and the triads were fully acknowledged (they were not unambiguously defined in the first images, and sometimes were thought to be part of the sarcoplasmic reticulum -SR-), the ECC was proposed to involve the excitation at the sarcolemma, then somehow the excitation penetration radially along a structure that, like the SR, should have a network nature mainly located at the Z lines. The resulting changes of charge density in those periodic places would lead to the release of Ca2+ from unknown resources (Csapo and Suzuki, 1957; Porter and Palade, 1957; Huxley and Taylor, 1958). This decade confirmed that Ca2+ was the activator of the contraction (Heilbrunn and Wiercinski, 1947; Weber and Portzehl, 1954; Niedergerke, 1955; Ebashi and Endo, 1968), probably by modulating the adenosine triphosphatase (ATPase) activity of the contractile machinery (Weber, 1959), but did not make clear what its origin was.
The latter model proved to be, in general terms, qualitatively correct when the 60 s well recognized the TT and its continuity with the sarcolemma (Endo, 1964; Franzini-Armstrong and Porter, 1964; Huxley, 1964) and the inward spread of the action potential through them (Ebashi and Endo, 1968). Also, the triad evolved from “two vesicles with the intervening space” (Porter and Palade, 1957) to the complex formed when the “T system is bordered on both sides by the terminal sacs of the SR” (Franzini-Armstrong and Porter, 1964). Subsequent work unveiled the main intracellular reservoir of Ca2+ and evidenced that the diffusion of Ca2+ starts when released from such reservoirs at periodic places which coincided with the terminal cisternae of the triadic regions (Hasselbach and Makinose, 1962; Huxley, 1964; Winegrad, 1965; Jöbsis and O’Connor, 1966; Ridgway and Ashley, 1967; Ashley and Ridgway, 1968; Ebashi and Endo, 1968; Winegrad, 1968; Winegrad, 1970), although the exact release mechanism was not fully understood. Huxley envisioned this in 1964 as: “It seems much more likely that the depolarization of the central element of the triad triggers off the release of calcium from the side elements … and that the specialized junctional regions seen are involved in this transmission process” (Huxley, 1964). The inward spread of excitation and the transient apparition of Ca2+ in the sarcoplasm constituted the long sought “inward acting link”. The discovery of a dual effect of adenosine triphosphate (ATP), which led to the proposal of a relaxing, “Marsh-Bendall” factor in muscle homogenates, and demonstrated to be the non-soluble, vesicle-located, ATP-Mg2+ activated sarcoendoplasmic reticulum Ca2+ pump (SERCA) (Kielley and Meyerhof, 1948; Marsh, 1951; Weber and Portzehl, 1954; Kumagai et al., 1955; Ebashi and Lipmann, 1962; Hasselbach and Makinose, 1962; Hasselbach, 1964; Ebashi and Endo, 1968; Caputo, 2011), closed the basic cycle of release and reuptake of Ca2+ from and to the SR.
Finally, Ebashi and Endo (Ebashi and Endo, 1968), who identified the molecular link between the sarcoplasmic Ca2+ and the activation of the contractile machinery, and also participated in the “relaxing factor” work, delineated the basic ECC sequence pretty much as we know it now: “The processes which would bring the contractile elements to the active state may be listed as follows: Action potential and its inward spread through the T-system … the sarcoplasmic reticulum is the site of linkage between excitation and contraction, i.e., Ca ion associated with a certain part of the sarcoplasmic reticulum is released by the influence of the electrical current field, induced by depolarization of the surface membrane, and Ca ion thus released subsequently activates the contractile system”, then, the “sarcoplasmic reticulum, exerts its relaxing effect by removing Ca ion from the contractile system in vivo”. Saul Winegrad complemented: “it is likely that … the longitudinal tubules and the intermediate cisternae … contain the calcium-sequestering system that is believed to operate during relaxation. The calcium taken up by these structures presumably then moves more slowly to the terminal cisternae, the main storage site in the muscle which has completely recovered from mechanical activity” (Winegrad, 1968).
Successive research enriched that sequence with structural and functional details, mainly pertaining to the specific issue of the Ca2+ release from the triad and added regulators and more precise quantitative data to the whole process. Also, demonstrated that all these events are extremely coupled. The readers are referred to the Historical Compendium of Muscle Physiology, for further historical details (Caputo, 2011).
2.2 The sequence of events and the molecular machinery involved in the excitation–contraction coupling
The ECC mechanism in skeletal muscle depicts a fast communication between electrical events taking place in the sarcolemma and the muscle contraction, through a cascade of global and locally restricted Ca2+ transients. The sequence of events entails: 1) initiation and propagation of an action potential (AP) along the plasma membrane, 2) inward, radial spread of the depolarization along the tubular system, 3) dihydropyridine receptors (DHPR)-mediated sensing of changes in the membrane potential, 4) allosteric interaction of the DHPR with the SR Ca2+ release channels (ryanodine receptors, RyR), 5) rapid release of Ca2+ from the triadic regions of the SR and transient increase of Ca2+ concentration ([Ca2+]) in the myoplasm, 6) transient activation of the contractile apparatus and the myoplasmic Ca2+ buffering system, 7) activation of the membranous-linked Ca2+ buffering and transporting system, which comprises the SERCA and the Na+/Ca2+ exchanger (NCX), and 8) appearance of the cascade of secondary mitochondria-restricted and tubular-restricted Ca2+ transients, the latter reflecting the recycling of Ca2+ through the store-operated Ca2+ entry (SOCE) mechanism.
Physiologically, the activation of the muscle fibre is modulated by the depolarization of the sarcolemma, including the tubular system (Kuffler, 1947; Hodgkin and Horowicz, 1960a). Under resting conditions, the fibre is polarized between –70 (Head, 1993) and –83 mV (Luff and Atwood, 1972; Wang et al., 2022), at 22 and 37°C, respectively. Upon binding of acetylcholine (ACh) to the motor end plate, the inward sarcolemmal conductance to Na+ rapidly increases, bringing about an AP. In most experiments, performed between 15 and 30°C, the AP depolarizes the fibre and then slightly polarizes it to positive values between +25 and +35 mV, which seems to be a safety factor for a successful AP conduction and SR Ca2+ release (Wang et al., 2022). The AP spike has 1.5–2.5 ms of duration at half-maximum amplitude and spreads along the sarcolemma over both sides of the motor plate with a propagation velocity of 0.4–1.9 m/s, depending on the measurement technique, the temperature of the experiment and the muscle studied (Luff and Atwood, 1972; Delbono and Stefani, 1993; DiFranco et al., 2008; Pedersen et al., 2011; Banks et al., 2018). The tension-sarcolemmal potential relationship is sigmoidal, with a threshold for activation at about −54–58 mV (Hodgkin and Horowicz, 1960a; Caputo, 2011).
During its sarcolemmal travel, the AP enters sarcolemmal invaginations known as TT. These are periodic, radially (on a transversal section) or transversally (on a longitudinal section) oriented membranous structures (80–100 × 35–40 nm wide), which conduct the AP at about 1 cm/s in a Na+ dependent, regenerative way (Huxley, 1964; González-Serratos, 1971; Bezanilla et al., 1972; Edwards et al., 2012). The so-called TT are actually a tubular network which comprises transverse (∼75%), diagonal (∼10%) and longitudinal (∼15%) tubules (Huxley, 1964; Jayasinghe et al., 2013; Jayasinghe and Launikonis, 2013). This structural arrangement, as well as its electrical properties, secures the rapid delivery of the AP to the interior of the fibre (Bezanilla et al., 1972; Fraser et al., 2011; Pedersen et al., 2011; Edwards et al., 2012), a crucial step for the uniform release of Ca2+ and the subsequent rapid contraction.
The incoming excitation reaches the triadic regions, where a TT is surrounded by two radially dilated portions of the SR, called “terminal cisternae” (Porter and Palade, 1957; Franzini-Armstrong and Porter, 1964; Huxley, 1964). The junctional parts of the terminal cisternae (jSR), and the TT, of the triadic regions, house a bunch of proteins involved in the regulation of the release of Ca2+ from the SR. The central actors in this process are the DHPR anchored to the TT and the RyR1 anchored to the SR membrane, which constitute the Ca2+ release units (CRU). DHPR (L-type Ca2+ channel, CaV1.1) are heteropentamers formed by subunits α1 (transmembrane), γ (transmembrane), β1 (intracellular), α2 (extracellular), and δ1 (extracellular), whose function is regulated by the membrane potential. The cryoelectron microscopy (cryo-EM) reconstructions at 2.7–3.6 Å confirmed that the α1 subunit of the channel has the typical 6 × 4 structure of many voltage-gated channels, i.e., four homologous domains (DI-IV), each with six transmembrane helices (Wu et al., 2016; Zhao et al., 2019). The α1 subunit houses the dome, the pore domain (PD), the selectivity filter (SF), and the voltage-sensing domain (VSD). The dome is a negatively charged, progressively narrowing region mainly shaped by extracellular loops above the SF of the PD, to which it guides Ca2+. The PD is made up of the S5, S6 and P helices of each domain, and several loops stabilized by multiple disulfide bonds, which create a permeation path for Ca2+ of about 60 Å in length. The SF is a specialized, narrow region, predominately formed by negatively charged aminoacids: E292 and G293 of DI, E614 and D615 of DII, E1014 and G1015 of DIII, and E1323 and A1324 of DIV. Since N617 of DII seems also to be particularly important for Ca2+ permeation (Dayal et al., 2017; Idoux et al., 2020; Dayal et al., 2021), the SF is likely more complex than initially proposed (Wu et al., 2016). The auxiliary subunits have a regulatory role on the expression, localization and function of the channel (Gregg et al., 1996).
Since the DHPR carries a Ca2+ current under voltage-clamp protocols in intact fibres (Skoglund et al., 2014; Dayal et al., 2017; Banks et al., 2021), it is expected to function the same as a response to an AP, highlighting its nature as a voltage-gated channel. The activated Ca2+ inward current is slower, and with a slightly lower amplitude compared to the Cav1.2 present in the heart, however, the influx of Ca2+ through this channel is not necessary for the skeletal muscle ECC and contraction (Caputo and Gimenez, 1967; Armstrong et al., 1972; Dayal et al., 2017; Idoux et al., 2020). Instead, the ability of the DHPR to sense the AP is particularly important for the skeletal muscle ECC. The S4 transmembrane helices of the α1 subunit constitute the voltage sensors (VSDI to VSDIV), which decode the information of the tubular excitation and translates it into a signal for the RyR1. The voltage sensing function depends on the S4 enrichment in the positively charged aminoacids arginine and lysine. Their movement during the VSD operation produces a small, yet measurable, voltage-dependent intramembrane charge movement, i.e., a current, which precedes the activation of the Ca2+ release from the SR (Schneider and Chandler, 1973; Rios and Brum, 1987; Banks et al., 2021). The peak of the charge movement time course follows the peak of the AP by 1.5 ms (Banks et al., 2021). It is intriguing why there are sizeable differences in the amplitudes, voltage-dependence and time courses of the VSDI-IV movements (Banks et al., 2021; Savalli et al., 2021), and whether they actually tune in any way the Ca2+ release from the SR. For instance, the VSDII and VSDIV seem to be the first ones activated, but the VSDI is so slowly activated that it seems not to be directly involved in the Ca2+ release activation; contradictory results have been reported regarding the activation kinetics of VSDIII (Banks et al., 2021; Savalli et al., 2021). In any case, the activation of either one or several of the DHPR´s VSD likely leads to a conformational change that gates the opening of the RyR1 in a cooperative way (Schneider and Chandler, 1973; Rios and Brum, 1987; Ríos et al., 1993).
Functional experiments with molecularly engineered DHPR suggested that its loop DII-III, close to the VSDII, is fundamental to ECC (Tanabe et al., 1990). However, the 3D structures available just lack the region between residues 687–789 (Wu et al., 2016; Zhao et al., 2019), which corresponds to the loop DII-III, precluding a conclusion about if it is long enough to clearly reach and interact with the RyR. If not directly, this loop may still interact with the RyR through the SH3 and cysteine-rich domain containing (STAC3) protein (Rufenach and Van Petegem, 2021; Shishmarev et al., 2022), something which awaits to be confirmed as a step to prove if STAC3 mediates the DHPR-RyR coupling relevant for a successful ECC.
Alternatively, the loop I-II-AID-β1 complex (Wu et al., 2016), also close to the VSDII, protrudes from the DHPR to the myoplasm and may directly or indirectly (i.e., the discovery of accessory β1-binding proteins such as Rem opens this possibility) interact with the RyR, explaining early functional observations according to which the absence of the β1 subunit eliminates the ECC (Gregg et al., 1996; Beqollari et al., 2015).
The history has shown that solving this issue is particularly difficult, but to fully understand the ECC mechanism, it is necessary first to make clear if, and how, the DHPR gates the RyR1 through a direct DHPR-RyR interaction or requires one or several accessory proteins (e.g., STAC3, Rem). Afterwards, it is crucial to have the complete, atomic resolution structure of the DHPR-RyR or DHPR-accessory proteins-RyR complexes under different conformations, to unambiguously assign the domains that mediate their interaction and shed light on the gating mechanism. Using purified complexes or native membranes would be, at least theoretically, possible approaches to address this problem.
Supramolecularly, the DHPR are arranged in groups of four, called tetrads, which alternately face the highly ordered RyR1, filling the 15–25 nm TT-SR gap (Huxley, 1964; Block et al., 1988; Franzini-Armstrong and Jorgensen, 1994; Franzini-Armstrong et al., 1998; Franzini-Armstrong, 1999).
The RyR1 are mushroom-like (from its lateral view), four-leaf clover shaped (from the sarcoplasmic view), Ca2+ channels with a big cytoplasmic moiety and a transmembrane region, inserted in rows in the jSR (Fleischer et al., 1985; Imagawa et al., 1987; Block et al., 1988; Saito et al., 1988). The effort of many laboratories worldwide, and the gain in resolution in cryo-EM recent reconstructions (between 3.8 and 6.1 Å) (Efremov et al., 2015; Yan et al., 2015; Zalk et al., 2015), compared to the first structures (over 9 Å) (Block et al., 1988; Saito et al., 1988; Wagenknecht et al., 1989; Ludtke et al., 2005; Samsó et al., 2005), reached a point at which a model of six transmembrane segments for each of the four monomers (6 × 4) that ensemble the functional channel appears reasonable. Thus, the transmembrane region of each monomer looks alike other ion channels: four transmembrane α-helices (S1 to S4) surround the S5 and S6 pore-forming helices. The luminal loops, the S6 and the P-segments constitute an extended permeation pathway of about 80 Å in length, which includes a 10 Å long SF, which drains into a 15 Å long hydrophobic cavity. A motif enriched in glycine residues of the S6 along this pathway is particularly important for gating and Ca2+ permeation in this channel (Efremov et al., 2015; Mei et al., 2015; Yan et al., 2015; Zalk et al., 2015).
The cytoplasmic moiety comprises about 80% of the bulk of the protein and is a complex network of tens of α-helices, surrounding one central spot (SPRY domains) enriched in β-sheets, which conform up to 20 domains (Efremov et al., 2015; Chen and Kudryashev, 2020), mainly involved in binding and transducing the signaling of many ligands to the pore region. Among those domains rich in α-helices, the EF-hand motifs, and the repeat 3-4 highlight: the EF-hands are in the lower face of the cytoplasmic moiety, while the repeats 3-4 are on top and at the corners of the cytoplasmic moiety, where they are involved in Ca2+ sensing and DHPR-RyR interaction, respectively. These structures seem to be the responsible for two putative modes of activation of the RyR1: mediated by the DHPR in those RyR1 coupled to tetrads, and by Ca2+ in those RyR1 not coupled to tetrads. From the nice images published (Samsó et al., 2009; Efremov et al., 2015; des Georges et al., 2016), we can say that the conformational change observed during opening of the channel resemble the flowering of a rose: a central twist and dilation, accompanied by a notorious change in the periphery of the structure which move outwards and downwards. This seems to be associated with an increase in curvature in the SR membrane when observed in native membranes by cryo-electron tomography (Chen and Kudryashev, 2020).
Upon opening, the RyR1 allows a rapid, massive, highly regulated release of Ca2+ from the SR to the myoplasm. From the peak of the AP, the peak of the release of Ca2+ is delayed by about 2–3 ms in most experiments performed between 15 and 25°C (Delbono and Stefani, 1993; Banks et al., 2021). This time window encompasses the charge movement in the DHPR, the RyR1 gating and opening, and the Ca2+ diffusion from the terminal cisternae to the myoplasm. Although differences among fibre types have been recognized (Section 3.1), the peak of the release of Ca2+ is attained within 1.8 ms in most fibres (Calderón et al., 2009; Calderón et al., 2010; Calderón et al., 2014a; Calderón et al., 2014b; Rincón et al., 2021). Beyond the DHPR, a handful of endogenous regulators of the Ca2+-release function of the RyR1, acting either from the myoplasmic or the SR luminal side, have been described: ATP and other purines, Ca2+, Mg2+, reactive oxygen species (ROS) and reactive nitrogen species, redox state, phosphorylation/dephosphorylation status, calmodulin, S100A1, FK 506 binding protein 12 (FKBP12 or calstabin-1), triadin (Trisk-95 and Trisk-51), junctin, homer-1, calumenin-2 and calsequestrin (CASQ) (Imagawa et al., 1987; Lai et al., 1988; Hidalgo et al., 2005; Butanda-Ochoa et al., 2006; Jung et al., 2006; Wei et al., 2006; Goonasekera et al., 2007; Feng et al., 2008; Prosser et al., 2008; Wei et al., 2009; Boncompagni et al., 2012; Wium et al., 2012; Marty, 2015; Meissner, 2017; Ogawa et al., 2021; Woll and Van Petegem, 2022).
Although the SR protein-27 (SRP-27) and junctophilin (JPH)-1 interact with the RyR1 (Phimister et al., 2007; Bleunven et al., 2008), it is not clear yet if they actually regulate the channel. There is debate on whether JPH-2 interacts or not with RyR1 (Phimister et al., 2007; Nakada et al., 2018). The ability of JPH-1 and -2 to regulate Ca2+ release in myotubes likely relies on their DHPR-binding ability and their TT-jSR tethering properties, which also mediate the precise localization of CRU (Nakada et al., 2018; Perni, 2022).
First isolated from rabbit muscle, CASQ is a ∼44-kDa Ca2+ binding protein highly expressed in the lumen of the jSR (MacLennan and Wong, 1971; Franzini-Armstrong et al., 1987; Perni et al., 2013), where it undergoes a [Ca2+]-dependent cooperative and reversible polymerization, forming oligomers with both low and high affinity sites for Ca2+ (Park et al., 2003; Sanchez et al., 2012). When [Ca2+] approaches 1 mM, the three thioredoxin-similar domains which surround a hydrophilic core fold, exposing numerous negatively charged aspartate and glutamate residues, that stack front-to-front forming dimers, which in turn stack back-to-back and continue stacking as [Ca2+] increases, to form a ribbon-like polymers that can ramify as a tree, finally forming a mesh with multiple nodes (Wang et al., 1998; Park et al., 2003; Park et al., 2004; Sanchez et al., 2012; Kumar et al., 2013; Perni et al., 2013; Wang and Michalak, 2020). Those “branches” anchor directly, or through triadin and junctin, to the RyR1, forming a complex that modulates the SR Ca2+ release (Guo and Campbell, 1995; Zhang et al., 1997; Wei et al., 2006; Goonasekera et al., 2007; Wei et al., 2009; Boncompagni et al., 2012; Sanchez et al., 2012; Wang and Michalak, 2020). Its ability to bind up to 80 ions per molecule explains why CASQ1 keeps the total SR Ca2+ as high as 35–175 mM (Royer and Ríos, 2009; Wang and Michalak, 2020). The electrostatic binding of Ca2+ to CASQ, together with its low affinity sites, favors the rapid unbinding and release of Ca2+.
The apparition of Ca2+ in the myoplasm shows microdomains with an up to 20-fold gradient of [Ca2+], which depends on the distance of a subcellular region from the CRU (Escobar et al., 1994; Baylor and Hollingworth, 2007; DiFranco et al., 2008; Hollingworth et al., 2012; Holash and MacIntosh, 2019). The average of those variable Ca2+ microdomains generates a global, positive, myoplasmic Ca2+ transient. The amount of Ca2+ released from the SR is enough to rise the resting cytoplasmic free [Ca2+] from 45–106 nM (Williams et al., 1990; Westerblad and Allen, 1991; Head, 1993; Konishi, 1998) to a fibre-type dependent averaged value of 7–30 µM (Hollingworth et al., 1996; Baylor and Hollingworth, 2003; Baylor and Hollingworth, 2007; Hollingworth et al., 2012; Milán et al., 2021; Rincón et al., 2021) (Section 3.1). In turn, this global, master Ca2+ transient associates to a cascade of locally restricted Ca2+ transients, from which a diversity of phenomena is activated (contraction, metabolism, heat, etc), such as the positive mitochondrial Ca2+ transients, and the negative SR and tubular Ca2+ transients. Also, as soon as the Ca2+ appears in the myoplasm, several Ca2+ buffering mechanisms are activated. Troponin C (TnC), parvalbumin (PV), ATP and the Ca2+ indicators rapidly bind Ca2+. Each of these Ca2+ binding mechanisms produces its own Ca2+ transient. Subsequently, the NCX, the mitochondria and the SERCA deal with Ca2+ with slower kinetics and the SOCE machinery recycles part of the Ca2+ extruded through the NCX. Each mechanism will be further developed in the coming paragraphs.
Troponins are a family of proteins attached to the thin filaments, from which TnC binds several Ca2+ ions with moderate affinity and are the molecular link between the cytosolic Ca2+ raise and the activation of the contraction (Ebashi et al., 1969). The beginning of the contraction shows a delay of 2–3 ms with respect to the beginning of the Ca2+ release, at room temperature, reflecting the diffusion time and the binding to TnC.
Muscle PV is a low molecular weight protein of the ɑ-sublineage (ɑ-PV), particularly abundant in muscle fibres type II, in which it can reach ∼1,000 µM. Two, high affinity, EF-hand, Ca2+ binding sites, are the responsible for its role as a Ca2+ buffer important in muscle relaxation, as it was recognized long time ago (Gillis et al., 1982; Heizmann et al., 1982; Leberer and Pette, 1986; Füchtbauer et al., 1991; Permyakov and Uversky, 2022). The binding of Ca2+ to TnC and PV explains most of the heat produced during muscle activation but is PV the responsible of the heat absorption observed several milliseconds after the Ca2+ release (Barclay and Launikonis, 2021). Given its differential concentration, and its high affinity when present, PV also has an important role in shaping the different morphologies of the single and tetanic Ca2+ transients obtained in different fibre types (Calderón et al., 2014a) (Section 3.1).
ATP reaches concentrations even higher than PV, which give it importance as a Ca2+ buffer, however, its lower Ca2+ affinity gives it less total capacity for Ca2+ binding than TnC and PV (Baylor and Hollingworth, 2003; Rincón et al., 2021). Under experimental conditions, the Ca2+ dyes also buffer Ca2+, within a variable range of kinetics that depend on the concentration and the intrinsic properties of the indicator molecule (Section 3.1).
Once the unbound Ca2+ returns to the myoplasm, relaxation proceeds. Ca2+ is definitively removed by the mechanisms responsible to extrude it from the myoplasm. The first mechanism activated is the NCX. This protein is located mainly in the tubular network, where it extrudes Ca2+ from the sarcoplasm, with a low total capacity and rapid saturation. Given this kinetics, its role is more evident during tetanic than during single stimulation (Balnave and Allen, 1998; Calderón et al., 2014a; Rincón et al., 2021). Part of this extruded Ca2+ is recycled back to the myoplasm during and after each twitch through Orai1 and likely the transient receptor potential canonical (TRPC) channels, reflecting a phasic activation of the SOCE mechanism (Section 3.3). This NCX-SOCE coupling likely reflects part of the bidirectional SR-TT exchange of Ca2+ proposed back in the 70 s of the last Century (Winegrad, 1968; Winegrad, 1970).
The uptake of Ca2+ by the mitochondria also removes Ca2+ from the myoplasm. The mitochondrial Ca2+ transients follow the cytosolic one with a ∼10 ms delay. Actively studied during the last two decades, at least two lines of evidence suggest that its sizeable buffering capacity shapes the cytosolic Ca2+ transient helping the muscle relax, further carrying metabolic consequences. On one side, dual mitochondrial and myoplasmic Ca2+ measurements demonstrated larger cytosolic Ca2+ transients in regions with polarized vs. depolarized mitochondria in intact flexor digitorum brevis (FDB) fibres during a single twitch, which allowed to estimate a rather large capacity of 10–18% to buffer the cytosolic Ca2+ transient (Yi et al., 2011). On the other side, poisoning different fibre types with FCCP (Section 3.4) induced a reversible lengthening of the decay phase of single and tetanic Ca2+ transients (Caputo and Bolaños, 2008; Calderón et al., 2014a). Recent knowledge about the fine structure of the molecular machinery involved in Ca2+ transport into the mitochondria, as well as the determination of the precise amount of Ca2+ released from the SR and the peak Ca2+ concentration reached in the sarcoplasm, allows us now to describe how and how much Ca2+ enters the mitochondria during ECC in different fibre types (Section 3.2).
The final mechanism responsible for restoring the sarcoplasmic resting [Ca2+] and keeping it low is the SERCA. SERCA is a high molecular weight, highly regulated pump, enriched in the longitudinal region of the SR and the non-junctional membrane of the terminal cisternae, which transports Ca2+ into the SR against its concentration gradient (Hasselbach, 1964; Jorgensen and Jones, 1986; Hasselbach, 1998; Odermatt et al., 1998; Periasamy and Kalyanasundaram, 2007; Rathod et al., 2021). This protein has three large cytoplasmic domains, N, P and A, attached to a domain consisting of 10 hydrophobic trans-SR-membrane helices (M1 to M10) (MacLennan et al., 1985; Toyoshima and Mizutani, 2004). ATP and Mg2+ binding to the more peripheral N domain activates its large movement towards the more central P domain and the turning of the A domain over itself. The resulting movements of some transmembrane helices, particularly M1, M2 and M4, occlude two Ca2+ ions within the transmembrane region and then release them inside the SR (Toyoshima and Mizutani, 2004). Even when a single Ca2+ transient is only ∼4–12 ms width, depending on the type of the fibre, SERCA pumping remains active for more than 60 ms, while dealing with the Ca2+ 1) being unbound from the TnC and PV, 2) leaving the mitochondria and 3) entering the fibre via SOCE.
The last decade was particularly fruitful to take to the next level observations performed in the “old”, past century: 1) researchers finally obtained structures of the DHPR and RyR at near-atomic resolution, helping explain the gating of both channels and mapping several regulators; 2) the structural and functional importance of the new main triadic regulators (JPH, triadins, junctin and STAC3) was acknowledged; 3) the peak myoplasmic [Ca2+] was reliably quantified (Section 3.1); 4) evidence consolidated the importance of mitochondria in Ca2+ handling in skeletal muscle (Section 3.2); 5) the ECC is ultimately a cascade of global and restricted Ca2+ transients associated to the fibre excitation whose concerted action activates the contractile machinery and other functions of the skeletal muscle. Figure 1 presents a complete, updated structural and functional model of the ECC in mammalian skeletal muscle.
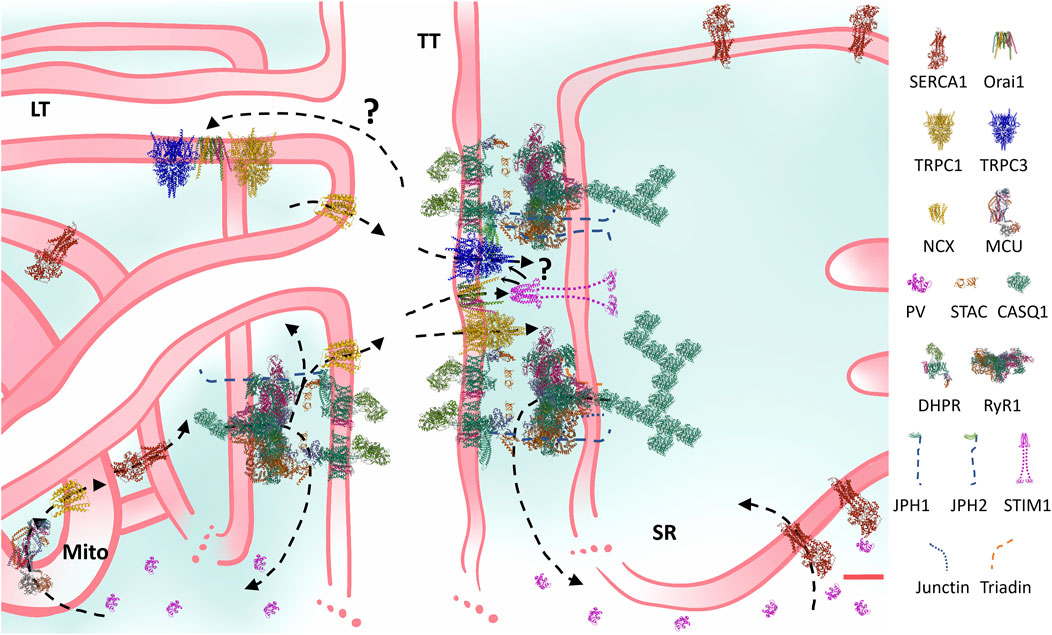
FIGURE 1. Structural and functional comprehensive model of the Ca2+ handling machinery and Ca2+ fluxes of the mammalian skeletal muscle ECC. Constructed at scale based on microscopy and protein measurements, the cartoon considers many current observations and addressed the question if there is space for all ECC proteins in the triadic space. For instance, JPH-1 coprecipitates with RyR1 more than JPH-2, also, super-resolution observations suggest that JPH-1 locates more closely to RyR1 than JPH-2. Regarding the SOCE machinery, some TRPC members have been shown to precipitate with DHPR and Orai. The model shows that a TRPC-Orai-TRPC cluster explaining these observations fits among DHPR-RyR1-empty squares. STIM location and size enables its binding to the TRPC-Orai-TRPC cluster. Also, this pattern may explain how TRPC may be a mediator of the RyR1 involvement in SOCE regulation. It is clear that preassembled STIM-Orai clusters present a solution to the problem of severe movement restrictions in this region. Arrows indicate Ca2+ fluxes. The internal equilibrium of Ca2+ entails short (on the right: sarcoplasmic reticulum (SR) release, myoplasmic buffering and SERCA reuptake) and long (on the left: SR release, myoplasmic and mitochondrial (Mito) buffering and activation, and SERCA uptake) routes. The external equilibrium entails the Ca2+ exit through the NCX and the entry through the SOCE channels, located in the transverse (TT), and probably in the longitudinal tubules (LT). Question marks indicate probable routes or mechanisms, which have not been “seen” yet. Although the high amount of PV was meant to represent a type II fibre, in general terms, the structure and the direction of the fluxes presented remain qualitatively the same in all fibre types. In the few cases in which the structure of the muscle isoforms have not been determined, homologous were used (e.g., non-mammalian NCX or STIM1 were used instead of NCX1-3 or STIM1L). Key to the figure: ECC: excitation-contraction coupling; SOCE: store-operated Ca2+ entry; SERCA: sarcoendoplasmic reticulum Ca2+ adenosine triphosphatase; TRPC: transient receptor potential canonical; NCX: Na+/Ca2+ exchanger; MCU: mitochondrial Ca2+ uniporter; PV: parvalbumin; STAC: SH3 and cysteine-rich domain containing protein; CASQ: calsequestrin; DHPR: dihydropyridine receptor; RyR: ryanodine receptor; JPH: junctophilin; STIM: stromal interaction molecule. Protein data bank structures: 1RTP, 1VFP, 2K60, 2MAJ, 3J8H, 3TEQ, 5GJV, 5JDG, 5KN1, 5ZBG, 6BBF, 6K7Y, 6UY7, 7RW4, 7RXQ. Red calibration bar: 10 nm.
3 Focus on
Here, we will focus on selected topics which have received particular attention during the last decade and whose results can be articulated with previous observations to reach stronger conclusions.
3.1 Assigning reliable numbers to the Ca2+ fluxes and concentrations in mammalian muscle fibres
How much Ca2+ is released from the SR and with which kinetics? Which free [Ca2+] is reached in the myoplasm? Is this Ca2+ enough to activate contraction? What is the kinetics of the Ca2+ reuptake? These questions have been addressed for at least 5 decades, when pioneer work demonstrated the transient increase in myoplasmic Ca2+ associated to the membrane depolarization in amphibia and arthropods muscles using murexide and aequorin as Ca2+ indicators (Jöbsis and O’Connor, 1966; Ridgway and Ashley, 1967; Ashley and Ridgway, 1968; Ebashi and Endo, 1968). The researchers then focused on amphibia until the early 90 s (Gillis et al., 1982; Baylor and Hollingworth, 1988; Caputo and Bolaños, 1994; Escobar et al., 1994), and finally moved to mammalian muscles.
Three attainments coincided at the time at which the field focused on Ca2+ measurements in mammalian muscle models: 1) availability of relevant biochemical information on the ECC and fibre types, 2) the outburst of fluorescent Ca2+ indicators and 3) improvements in calibration, mathematical and computational modelling.
Here, we will focus on how researchers integrated these three attainments to finally find a solution to the challenge of putting reliable numbers to the ECC Ca2+ fluxes and concentrations in mammalian muscle, especially considering the existence of at least four fibre types.
3.1.1 Biochemical information relevant to the mammalian excitation–contraction coupling in fibre types
The existence of muscles with different biochemical and dynamic properties was formally acknowledged long time ago (Ranvier, 1873; Close, 1972). The study of their particularities first focused on the biochemical differences of their fibres (Dubowitz and Pearse, 1960; Engel, 1962; Brooke and Kaiser, 1970; Barnard et al., 1971; Peter et al., 1972). By the 90s, a wealth of information confirmed the presence of at least four phenotypes in the muscles of the mammalian extremities, as recognized by the presence of the isoforms of the myosin heavy chain (MHC): I (slow twitch), and IIA, IIX/D and IIB (fast twitch) (Schiaffino and Reggiani, 2011).
Soon after the ECC phenomenon was demonstrated to be mediated by Ca2+, many researchers presented data concerning biochemical differences in the molecular machinery involved in Ca2+ release and reuptake using the dichotomic model of slow-twitch vs. fast-twitch fibres. Later, the studies were extended to the four fibre types. Details about the molecular and biochemical differences that underlie the quantitative differences in ECC among fibre types have been presented previously and we refer the readers to those papers and the literature cited therein (Bottinelli and Reggiani, 2000; Calderón et al., 2010; Calderón et al., 2014b; Rincón et al., 2021).
We can summarize the most relevant information as follows: 1) there are between twofold and threefold more CRU in the fast, compared to the slow fibres, with no difference in the isoforms. There are only about 1.5 times more triadic accessory proteins (e.g., triadin, JPH) in the fast compared to the slow muscles. Fast fibres only express CASQ1, while slow fibres have both CASQ1 and CASQ2 at a 3:1 ratio, but with a total amount of CASQ, and a total Ca2+ buffering capacity, somewhat lower than in fast fibres. Despite this, there seems to be only a ∼10% difference in SR free Ca2+ content between both types of fibres; 2) TnC isoforms differ between slow and fast fibres, explaining the presence of almost twofold more Ca2+ binding sites in fibres types IIA, IIX/D and IIB, compared to type I; 3) there is a continuum increase in PV content across the four fibre types such that the fastest fibres (type IIB) have up to 300 times more than the slowest fibres (type I). Fibres IIA have about tenfold more PV than type I; 4) the differential ATP content explains the about 50% higher amount of ATP Ca2+ binding sites in the fast fibres compared to slow ones; 5) SERCA different isoforms and content among fibres result in a twofold (for IIA) or up to fivefold (for IIX/D and IIB) larger maximum Ca2+ reuptake flux rate in fast fibres compared to fibres type I; 6) up to twofold higher mitochondrial volume and a larger maximum flux rate of the mitochondrial Ca2+ uniporter (MCU) explain the threefold to fourfold higher capacity of this mechanism in fibres type I compared to fibres type II; 7) NCX1 is more abundant in fibres type I, but the capacity of the NCX3, present in fibres type II, is slightly higher; 8) STIM1 is about 1.5 times more abundant in slow compared to fast fibres, however, the SOCE total capacity seems to be higher in the latter.
Short (FDB) and large (extensor digitorum longus, EDL, and soleus) fibres, either in fascicles, manually isolated or enzymatically dissociated, intact or nude, have been the most used models to study Ca2+ kinetics in different fibre types. Dynamical and molecular markers have been used to identify different fibre types in those preparations (Baylor and Hollingworth, 2003; Calderón et al., 2009; Calderón et al., 2010; Hollingworth et al., 2012; Calderón, 2013; Calderón et al., 2014a).
3.1.2 Using Ca2+ dyes that reliable track the Ca2+ transients
Foremost researchers popularized the use of fluorescent intracellular dyes for determining the dynamic concentration of intracellular Ca2+ in different cell types (Tsien, 1980; Tsien et al., 1982; Grynkiewicz et al., 1985; Minta et al., 1989). A variety of dyes was then developed and many used in skeletal muscle (Raju et al., 1989; Delbono and Stefani, 1993; Hollingworth et al., 1996; Gee et al., 2000; Katerinopoulos and Foukaraki, 2002; Woods et al., 2004) (Table 1). It became clear soon that slow dyes well measured resting Ca2+ but did not render trustable measurements of peak [Ca2+] and Ca2+ kinetics (Baylor and Hollingworth, 1988; Berlin and Konishi, 1993; Hollingworth et al., 1996; Wokosin et al., 2004). This is because of their slow rate of detachment from Ca2+ and their low Ca2+ Kd (below 1 µM in vitro), limiting the range of concentrations at which the dye responds before significantly buffering Ca2+ and becoming saturated. Moreover, the calibration of non-ratiometric, slow dyes is fraught with difficulties, for instance, large errors in the Fmin estimations are common, and they may induce up to a 17% error in the [Ca2+] (Mejía-Raigosa et al., 2021). As a result, a wealth of qualitatively relevant information was generated by using these slow dyes, however, quantitative reliable information was still lacking.
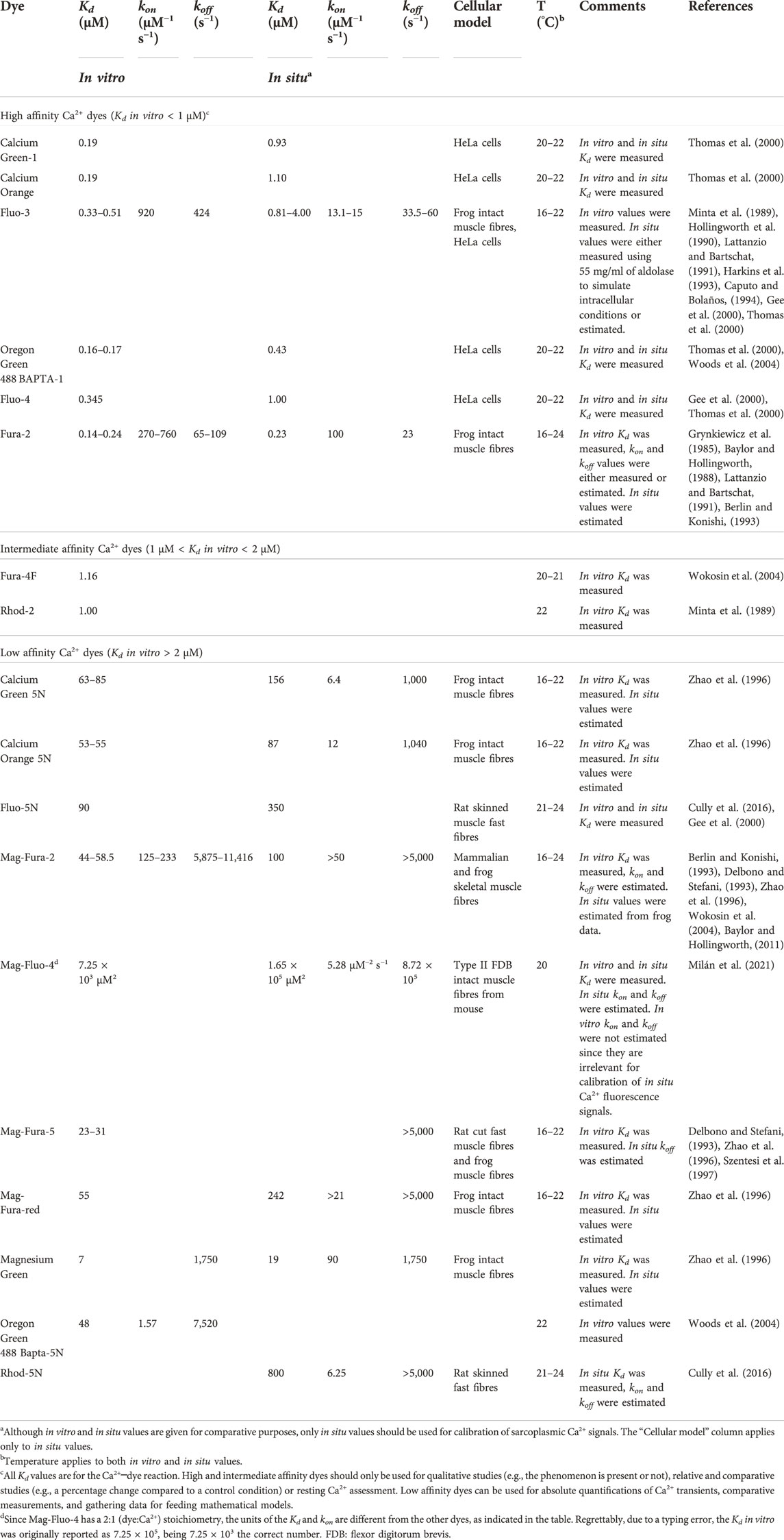
TABLE 1. Best Ca2+ dyes to study ECC in skeletal muscle, with their affinity and rate constants relevant for calibration of their fluorescence signals.
Fast Ca2+ dyes such as Fluo-5N, Mag-Fluo-4, Mag-Fura-2 (Furaptra) and Rhod-5N, typically have an in vitro Kd between 20 and 100 µM, which rise to about 700–1,000 µM in situ, i. e, in the myoplasm or in a medium resembling the myoplasm. This very low affinity in situ is the property that makes them reliably track Ca2+ kinetics in skeletal muscle, as demonstrated because these dyes resolve every single peak of a high-frequency tetanus, and their fluorescence signals superimpose the actual Ca2+ transients (Baylor and Hollingworth, 2003; Baylor and Hollingworth, 2007; Calderón et al., 2014a; Rincón et al., 2021). Moreover, this property makes them be far from saturation, be less prone to buffer the Ca2+ transients, and during calibration, their Fmin is less susceptible to errors because their fluorescence in presence of the resting [Ca2+] is already very low (Hollingworth et al., 1996; Baylor and Hollingworth, 2003; Milán et al., 2021).
Of the above mentioned, polycarboxylate, fast dyes Mag-Fura-2 and Mag-Fluo-4 are the most trustable to gain quantitative insight into the ECC in different fibre types (Table 1). Mag-Fura-2 was originally developed as a ratiometric Mg2+ dye, with a similar spectra as that of Fura-2, but it also binds Ca2+ with low affinity, and a 1:1 (Ca2+:dye) stoichiometry (Raju et al., 1989). For Ca2+ measurements, it has been typically excited between 350 and 430 nm, and its emission has been collected over 480 nm, either as ratiometric or as non-ratiometric (Hollingworth et al., 1996; Baylor and Hollingworth, 2003). An in situ Kd of 98 µM was estimated at 16°C (Hollingworth et al., 1996; Zhao et al., 1996), but experimental measurements in mammalian fibres are pending. Mag-Fluo-4 is a Fluo-4 derivate, with a 1:2 stoichiometry, with excitation and emission peaks at 493–494 and 515–516 nm, respectively, and a good dynamic range (Gee et al., 2000; Milán et al., 2021). The Mag-Fluo-4 in situ Kd is 1.65 × 105 μM2, measured in fast mice fibres at 20°C, which ensures that, even under short loading times (10 min), less than 5% of the dye is bound to Ca2+ during a twitch, thus being far from saturation (Milán et al., 2021). This dye binds more heavily than Mag-Fura-2 (74 vs. 54% of the indicator molecules bound) to intracellular components.
Given their structure, an apparent drawback of both dyes is the possibility of contamination of the Ca2+ signals with Mg2+. With a KdMg in vitro of 5,300 µM for Mag-Fura-2, and a range of KdCa of 44–58.5 µM (Table 1) at ∼20°C, a KdMg/KdCa ratio of ∼90–120 is obtained (Hollingworth et al., 2009). Assuming a KdMg of 6,500 µM (Baylor and Hollingworth, 2011) and a half-fluorescence concentration for Ca2+ of 31.5 µM (Milán et al., 2021), the KdMg/KdCa for Mag-Fluo-4 at ∼20°C is ∼200. Since similar ratios may apply to in situ conditions, and given a resting free [Mg2+] below 1 mM (Westerblad and Allen, 1992), a significant contamination of the Ca2+ signals with Mg2+ in muscle fibres can be ruled out.
3.1.3 Employing a trustable calibration method
The truthful conversion of fluorescence signals into Ca2+ comprises the use of equations and values for parameters of affinity and fluorescence acquired in situ at similar temperatures (Table 1). Several equations have been published for ratiometric and non-ratiometric dyes, as well as for 1:1 or 1:2 stoichiometry (Grynkiewicz et al., 1985; Hollingworth et al., 1996; Zhao et al., 1996; Mejía-Raigosa et al., 2021), including the calibration of Mag-Fura-2 (Hollingworth et al., 1996; Baylor and Hollingworth, 2003) and Mag-Fluo-4 (Milán et al., 2021). For calibrating fast dyes, saponin is a better membrane permeabilizer than ionomycin (Milán et al., 2021).
3.1.4 Then, how much Ca2+ do muscle fibres move?
Two different groups have been devoted to put numbers to the mammalian ECC regarding fibre types, satisfactorily taking into account the three aspects discussed above, and have obtained similar results (Hollingworth et al., 1996; Baylor and Hollingworth, 2003; Baylor and Hollingworth, 2007; Calderón et al., 2009; Calderón et al., 2010; Baylor and Hollingworth, 2011; Calderón et al., 2011; Hollingworth et al., 2012; Calderón et al., 2014a; Calderón et al., 2014b; Milán et al., 2021; Rincón et al., 2021). Their data, acquired using Mag-Fura-2 and Mag-Fluo-4, can be pooled, and presented as the following statements: 1) fibres type I and IIA share the Ca2+ transient kinetics called morphology type I (MT-I), while the fibres IIX/D and IIB share the morphology type II (MT-II); 2) single twitch MT-I signals have rise times between 1.2 and 1.8 ms and decay times of up to 80 ms. They release Ca2+ at rates between 50 and 150 μM/ms, and their peak sarcoplasmic free [Ca2+] ranges from 7 to 13 µM; 3) single twitch MT-II signals typically have rise times between 1.0 and 1.3 ms, with decay times ranging from 13 to 25 ms, being ∼3–4 times narrower than the MT-I signals; 4) MT-II fibres release Ca2+ at a huge rate of 200–250 μM/ms, and their peak sarcoplasmic free [Ca2+] ranges from 15 to 30 µM. For the sake of comparison, the peak SR release rate in mammalian cardiomyocytes ranges from 2.0 to 4.2 μM/ms (Song et al., 1998; Shannon et al., 2000); 5) the total amount of Ca2+ released from the SR in the MT-II is about 350 µM, ∼2.7 times higher than the amount released by the MT-I and about 5 times the amount released in cardiomyocytes (Song et al., 1998). The differential kinetics of the Ca2+ release partially explain the differential kinetics of the contraction in all fibre types (Calderón et al., 2010).
The variability in the values presented above reflects the inherent variability of the skeletal muscle biochemistry and function (Bottinelli and Reggiani, 2000; Bottinelli, 2001), its plasticity, as well as temperature (usually between 16 and 23°C) and sarcomere length differences between papers. Importantly, MT-I values almost never overlap with those of MT-II. Furthermore, these numbers reflect that the skeletal muscle fibre is the cell that deals with the largest and fastest release and reuptake of Ca2+, which, instead of its shortening ability, can be considered its main specialization.
The above numbers have fed increasingly complex mathematical models which have allowed to assign numbers to different compartments and Ca2+ binding mechanisms. A recent comprehensive model simulated the changes in Ca2+ concentrations and fluxes through the sarcomere of the four fibre types, considering classical (Tn, PV, ATP, SERCA, and dye) and new (mitochondria, NCX, and SOCE) Ca2+ binding sites, during single and tetanic stimulation, using Mag-Fluo-4 data (Rincón et al., 2021). The magnitudes of change of the Ca2+-bound forms of the Ca2+ buffers studied follow the order IIB ≥ IIX > IIA > I, except for the mitochondrial peak [Ca2+], which showed the pattern I >> IIA >> IIX ≥ IIB. The kinetics for fibres IIA and IIX proved to be intermediate between I and IIB fibres, supporting dynamic data (Bottinelli et al., 1991; Bottinelli and Reggiani, 2000; Bottinelli, 2001; Rincón et al., 2021).
An important issue is that the peak [Ca2+] described above agree well with several previous observations. For instance, the pCa50 of the Ca2+-induced superprecipitation reactions usually ranged from 5.9 to 5.1 at 23°C, i.e., ∼1–10 µM, in presence of physiological [Mg2+]. Similar observations were done regarding the tension-pCa relationship in skinned fibres, in which the maximum tension required a pCa ∼5.5─5.0, i.e., 3.2─10 µM in all fibre types (Ebashi and Endo, 1968; Bottinelli and Reggiani, 2000). Furthermore, the full activation of the contractile machinery and the appearance of mitochondrial Ca2+ transients in vivo requires [Ca2+] about one order of magnitude higher than 1–2 µM, which is the [Ca2+] that gives the 50% activation of these mechanisms (Ebashi et al., 1969; Sembrowich et al., 1985). The Ca2+ release from loaded SR vesicles, the open probability of the RyR1 and the ryanodine (Ry) binding kinetics, consistently show that the RyR1 half-activates at ∼1–5 µM and peaks between 10 and 30 µM Ca2+ at room temperature (Nagasaki and Kasai, 1983; Fill et al., 1990; Meissner, 2017). Finally, Ca2+ releases and fluxes as large as those reported above are required to account for the heat released during muscle activation, as recently demonstrated (Barclay and Launikonis, 2021).
In conclusion, although a differential biochemical data suggested different Ca2+ transient kinetics for the fibre types, and although biophysical evidence suggested an expected value for the peak sarcoplasmic [Ca2+] over 5 µM, it took a long way to finally assign reliable numbers to this issue: fibres type I and II have peak sarcoplasmic [Ca2+] between 7 and 13 µM, while fibres type IIX/D and IIB have values between 15 and 30 µM. The release rate and the total amount of Ca2+ released in fibres type IIX/D and IIB is ∼2–3 times larger than in fibres type I and IIA. Articulation of old data with data gathered during the last decade has made that coherence becomes now evident across biochemical (e.g., protein isoforms, reaction rates, dependence on Ca2+), dynamical (e.g., tension-pCa relationships) and biophysical (e.g., fluorescence, Ca2+ concentrations and fluxes) measurements and estimations.
3.2 Mitochondria in excitation–contraction coupling
Mitochondria are double membrane organelles, important regulators of cellular Ca2+ homeostasis, signaling, metabolism and energy production in the form of ATP, for which they have been named “the powerhouse of the cell” (Siekevitz, 1957) and “the hub of cellular Ca2+ signaling” (Szabadkai and Duchen, 2008). Moreover, they are highly dynamic, forming networks inside the cells and remodeling their morphology and activity (Anderson et al., 2019).
Recently, several reviews on the skeletal muscle mitochondria dynamics, with strong structural approaches have been published (Anderson et al., 2019; Bloemberg and Quadrilatero, 2019; Li et al., 2020; De Mario et al., 2021; Gherardi et al., 2021; Garbincius and Elrod, 2022). We also reviewed previously their importance for skeletal muscle (Calderón et al., 2014b). However, recent estimates of the rate of increase of Ca2+ and mitochondrial Ca2+ transients in different fibre types provide novel interesting quantitative information, not acknowledged in previous reviews, that fosters us to contribute this section. Thus, here we will center on the research relevant to understand the Ca2+ movements and concentrations into the skeletal muscle mitochondria and their relationship with ECC.
3.2.1 Location and dynamics
The mitochondria inside the mammalian skeletal muscle fibre can be classified as: subsarcolemmal, intermyofibrillar and perinuclear. While the subsarcolemmal and perinuclear have certain mobility, the movements of the intermyofibrillar are more restricted. The first are clustered and less ordered, whilst the latter are packed between the contractile proteins and the SR membranes, or highly ordered within the I-bands by pairs at either side of the Z line, close to the terminal cisternae of the triads, forming a quasi-crystalline structure (Ogata and Yamasaki, 1997; Vendelin et al., 2005; Kuznetsov et al., 2006; Bolaños et al., 2009; Franzini-Armstrong and Boncompagni, 2011; Boncompagni et al., 2020) (Figure 2).
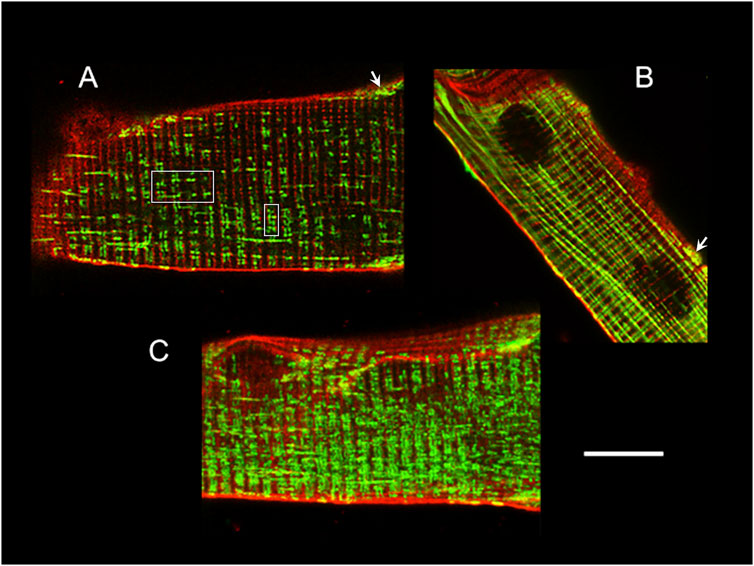
FIGURE 2. Mitochondria location and distribution in muscle fibres. Confocal images of adult mouse FDB fibres loaded with Di-8-Anneps (red) and Mitotracker Green (green) to stain membranes and mitochondria, respectively. The fibre end in (A) shows the T-tubules and intermyofibrillar mitochondria, either paired at both sides of the Z line near the center of the fibre (white squares) or forming elongated structures, which are much more evident in (B), and look like columns parallel to the long axis of the fibre. The arrows in (A) and (B) point to typical clustered, less ordered, subsarcolemmal mitochondria. The mitochondrial network appearance is clearer under the 3D reconstruction shown in (C). Calibration bar: 10 µm, applies to all panels.
Tethers anchor the outer mitochondrial membrane (OMM) to the terminal cisternae opposite to the jSR (Bolaños et al., 2008; Dirksen, 2009b; Boncompagni et al., 2009; Pietrangelo et al., 2015). Those tethers, previously found in liver cells (Mannella et al., 1998; Csordás et al., 2006), are 10 nm long electron-dense structures (Dirksen, 2009b; Boncompagni et al., 2009; Pietrangelo et al., 2015), whose nature remains under debate. They may correspond to the ERMES (Endoplasmic Reticulum-Mitochondria Encounter Structure) complex found in budding yeast (Kornmann et al., 2009), formed by four components (Mmm1, Mdm10, Mdm12, and Mdm34) and several accessory proteins (e.g., Emr1), whose malfunction affects mitochondrial morphology (Rasul et al., 2021). They have also been associated with Mitofusin2 (Mfn2) (de Brito and Scorrano, 2008), whose absence reduces mitochondrial Ca2+ uptake (Ainbinder et al., 2015). This topic awaits further research.
Although previously considered isolated organelles, mitochondria can communicate among them in different cell types (Huang et al., 2013; Lavorato et al., 2017; Vincent et al., 2017; Lavorato et al., 2020). In skeletal muscle by means of fusion-fission, remodeling events or “kissing junctions”, they form elongated structures with narrow connecting ducts, and less frequently nanotunnels, acting as an independent and highly dynamic network which connects the matrixes of non-adjacent mitochondria (Vincent et al., 2016; Vincent et al., 2017; Vincent et al., 2019; Lavorato et al., 2020; Rahman and Quadrilatero, 2021). There has also been described synapses-like structures between adjacent mitochondria (Picard, 2015) that would help integrate information about the network (Picard et al., 2015). Those synapses, connecting ducts and nanotunnels will favor the communication between mitochondria anchored to the SR and the whole network in skeletal muscle and may also provide the structural basis to support the idea of enhanced propagation of intracellular signals (Díaz-Vegas et al., 2019). The “Excitation-Metabolism Coupling” (EMC) term has been put forward to integrate the coupling between the depolarization and the metabolic signaling through the mitochondrial network interconnexions (Díaz-Vegas et al., 2019). If these connecting structures and network somehow directly feedback the ECC, besides having a metabolic role, is an avenue that should be studied in the future.
In a different context, the EMC was previously used referring to the bidirectional communication between SR and mitochondria. Orthograde when the Ca2+ influx activates ATP synthesis and metabolism. Retrograde given that the ATP is also used for the SERCA to reuptake Ca2+ into SR as well as for the inhibition of local SR Ca2+ release regulation. EMC would be more important in mitochondria-enriched slow- and fast-twitch oxidative muscle than in fast-twitch glycolytic muscle (Dirksen, 2009b; Rossi et al., 2009).
Although the structural evidence of the mitochondrial network seems convincing, functional studies demonstrating their importance for the ECC are still lacking.
3.2.2 How and how much Ca2+ enters the mitochondria?
The intermyofibrillar mitochondrial distribution confers them with a privileged position within ∼150 nm from the CRU of the jSR (Boncompagni et al., 2009) (Figure 2). Even in the case of a single twitch or caffeine exposure, mitochondrial Ca2+ can follow, with a short delay, the time course of the cytoplasmic Ca2+ increase (Rudolf et al., 2004; Shkryl and Shirokova, 2006; Bolaños et al., 2009; Yi et al., 2011; Karam et al., 2017), demonstrating that the myoplasmic Ca2+ increase observed during ECC is sensed by the mitochondria.
Given that the mitochondrial affinity for Ca2+ is intermediate, 1.2 µM for slow-twitch and 2 µM for fast twitch fibres (Sembrowich et al., 1985), and the first reported sarcoplasmic peak [Ca2+] were misleadingly low (even below 2 µM), it seemed difficult to explain how Ca2+ entered the mitochondria. However, as discussed (Section 3.1), during the last decade it became clear that all fibres reach between 7 and 30 µM free myoplasmic [Ca2+] just after excitation, making obvious that these values are at least one order of magnitude over the affinity of the mitochondria. Moreover, in compartmentalized models, larger free [Ca2+] between 40 and 60 µM can be attained just between the triads and the Z lines (Baylor and Hollingworth, 2007). This sizeable increase in Ca2+ close to the mitochondria, the higher sensitivity given by the skeletal muscle MICU1.1 variant (Section 3.2) and the large negative mitochondrial potential (ΔΨm, −180 mV) generate a strong electrochemical gradient which favors the diffusion and the Ca2+ transport into the mitochondrial matrix. The maximum flux rate was lately estimated to be 18.2 μM/s for fast and 74.3 μM/s for slow fibres (Rincón et al., 2021). A recent model showed for the first time that the mitochondrial Ca2+ transients reach 0.3 µM in fibres IIX/D and IIB, 0.5 µM in fibres IIA and 1.2 µM in fibres type I (Rincón et al., 2021).
The Ca2+ increase in the mitochondrial matrix enhances the ATP production necessary for muscle contraction, by activating the ATP synthase and enzymes of the tricarboxylic acid cycle and the oxidative phosphorylation (Kavanagh et al., 2000; Finkel, 2011). It also helps shaping the decay phase of the sarcoplasmic Ca2+ transient (Calderón et al., 2014a), likely speeding up the muscle relaxation. Depending on its kinetics, the excess of accumulated Ca2+ can also activate excessive ROS production and programmed dead of the cell through the opening of the mitochondrial permeability transition pore (mPTP) (Biasutto et al., 2016; Li et al., 2020).
3.2.3 Recent advances in Ca2+ handling machinery in mitochondria
The Ca2+ influx from the cytoplasm to the mitochondrial intermembrane space (IMS) occurs through the porine-like, voltage-dependent anion channels (VDAC) placed in the OMM (Colombini, 1980; Colombini, 1983; Colombini, 2012; Shoshan-Barmatz and De, 2017; Shoshan-Barmatz et al., 2018). Out of the three isoforms present in mammalian cells (VDAC1, 2, 3) (Shoshan-Barmatz et al., 2010; Messina et al., 2012; Shoshan-Barmatz and De, 2017), VDAC1 is the most expressed in skeletal muscle (Massa et al., 2000). At low transmembrane voltage the channel is open with high conductance for anions allowing the diffusion of anionic metabolites and adenine nucleotides, being selective for ATP. However, at higher transmembrane voltage the channel close for anions and becomes selective for Ca2+.
From de IMS to the mitochondrial matrix the Ca2+ goes through channels inserted in the inner mitochondrial membrane (IMM): the MCU complex (Kirichok et al., 2004), the Rapid mode (RaM) (Sparagna et al., 1995) and the mRyR1 (Beutner et al., 2001; Ryu et al., 2011).
The MCU is a highly Ca2+ selective channel holocomplex at IMM (Baughman et al., 2011; De Stefani et al., 2011; De Stefani et al., 2016; Mammucari et al., 2016). This complex contains the MCU, the EF-hand Ca2+-binding proteins Mitochondrial Calcium Uptake 1 (MICU1) and MICU2 forming dimers and the Essential MCU Regulator (EMRE) (Sancak et al., 2013). MICU1 acts as a Ca2+-sensing gatekeeper, keeping the channel closed when Ca2+ levels are low and allowing the channel to open in response to transient rises (Perocchi et al., 2010; Mallilankaraman et al., 2012; Csordás et al., 2013; Plovanich et al., 2013; Sancak et al., 2013). In addition, MICU1 is also involved in maintaining the cristae structure and IMM anchorage of MCU (Gottschalk et al., 2019). The MCUb subunit acts as a dominant-negative subunit that reduces the MCU activity (Raffaello et al., 2013).
In skeletal muscle, MICU1.1, a spliced variant of MICU1, forms the heterodimers MICU1.1-MICU2 giving a higher affinity for Ca2+ (Vecellio Reane et al., 2016; Gherardi et al., 2021). Normally, MCU current densities in skeletal muscle reach up to 58 pA/pF (Fieni et al., 2012).
The third channel-forming subunit recently described is EMRE, a transmembrane protein of 10 kDa with a single IMM transmembrane domain (Sancak et al., 2013). EMRE is required for the interaction of MCU with MICU1.1 in skeletal muscle and MICU2. It is essential for in vivo uniporter current given that MCU oligomers alone are not sufficient for in vivo uniporter activity (Sancak et al., 2013). EMRE-dependent regulation requires MICU1.1, MICU2, and cytoplasmic Ca2+; its acidic C-terminal domain functions as a matrix Ca2+ sensor that regulates the MCU activity (Vais et al., 2016). Thus, EMRE acts together with MICU1 as a gatekeeper complex regulating the Ca2+ movements through MCU, able to sense Ca2+ at both sides of IMM and preventing mitochondria both from Ca2+ depletion and overload (Vais et al., 2016).
The Ca2+ release from the mitochondria is under the control of the mitochondrial Na+/Ca2+ exchanger (mNCX, NCLX) (Palty et al., 2010; Palty and Sekler, 2012; Garbincius and Elrod, 2022), the Ca2+/H+ antiporter (mNCH or LETM1 or mHCX) (Jiang et al., 2009; Tsai et al., 2014) and the mPTP (Li et al., 2020; Bernardi et al., 2021). The first two mechanisms, present in the IMM, are capable of reverse function (Garbincius and Elrod, 2022). The Ca2+ efflux by NCLX is slower than the Ca2+ influx by MCU (Rudolf et al., 2004) which may favor Ca2+ overload if cytoplasmic Ca2+ is much increased or NCLX diminished. The nature of the third mechanism, the mPTP, is still unknown, though recently it was proposed to be formed by a Ca2+-dependent conformation of the F-ATP synthase (Li et al., 2020; Bernardi et al., 2021). Physiologically, through transient short openings (flickerings), mPTP may regulate Ca2+ in the mitochondrial matrix (Hüser and Blatter, 1999; Petronilli et al., 1999; Li et al., 2020; Bernardi et al., 2021). Mitochondrial Ca2+ overload, likely reflecting an imbalance between the Ca2+ uptake and release mechanisms, triggers uncontrolled mPTP opening causing the loss of ΔΨm, impairing ATP production, increasing ROS and eventually leading to cell apoptosis (Biasutto et al., 2016; Li et al., 2020; Bernardi et al., 2021), an effect also seen with some protonophores which release mitochondrial Ca2+ (Bolaños et al., 2008; Caputo and Bolaños, 2008). A physiological ECC, associated to regular mitochondrial Ca2+ transients, is essential to keep mPTP closed, preventing these deleterious events (Li et al., 2020). Unfortunately, despite this interesting finding, the topic of the Ca2+ release from muscle mitochondria remains largely unaddressed. The kinetics of this phenomenon, its relationship with the whole ECC machinery, and eventual quantitative or qualitative differences among fibre types should be addressed in the future.
It is satisfactory to see that the last decade finally unveiled the structure of the molecular machinery and the mechanisms involved in the substantial transport of Ca2+ to the skeletal muscle mitochondria and allowed us to estimate the [Ca2+] reached inside their matrix even considering differences among fibre types. Research may now be focused on the mechanisms of muscle mitochondrial Ca2+ exit.
3.3 Store-operated Ca2+ entry in skeletal muscle
SOCE refers to a Ca2+ influx activated in response to the SR depletion and functions in most cells to refill these stores. Since skeletal muscle ECC is independent of extracellular Ca2+ (Section 2.2), the interest in SOCE was low for more than 15 years after its discovery. However, the demonstration of a Ca2+ influx in response to acute SR depletion (Kurebayashi and Ogawa, 2001) independent of any ICa, opened a bulk of work addressing the question of the nature and physiological role of this Ca2+ influx in skeletal muscle. It was proposed that its function could be important in muscle growth, development and contractile function, as well as SR refill to delay fatigue (Stiber et al., 2008; Wei-LaPierre et al., 2013; Sztretye et al., 2017; Michelucci et al., 2018). However, a great depletion of SR may not be necessary to activate SOCE, since it is rapidly activated in response to a single AP (Koenig et al., 2018). Here we will present the main facts that conducted to these conclusions and a brief historical description of this mechanism, to understand its relevance in skeletal muscle.
3.3.1 Basic concepts
SOCE was first described in non-muscle cells, where the depletion of intracellular Ca2+ stores in the continuous presence of inositol triphosphate induced the so called “capacitative Ca2+ entry” (Putney, 1986). That entry was a small rectifying highly Ca2+ selective current, not affected by Ca2+ channel blockers (Hoth and Penner, 1992) called Ca2+ Release Activated Ca2+ Current (ICRAC). Two research groups then identified in 2005 the Stromal Interacting Molecule (STIM) as a single-pass transmembrane EF-hand protein that acts as Ca2+ sensor in the endoplasmic reticulum lumen of many cells with a low affinity of ∼200–600 μM (Liou et al., 2005; Roos et al., 2005; Zhang et al., 2005; Stathopulos et al., 2006; Canato et al., 2010; Friedrich et al., 2010). In 2006 it was confirmed the interaction of STIM with a protein in the plasma membrane called Orai, which constitutes the transmembrane pore of the CRAC complex (Vig et al., 2006b, 2006a; Feske et al., 2006; Prakriya et al., 2006; Soboloff et al., 2006; Hou et al., 2018). STIM-Orai complexes constitute the Ca2+ entry units (CEU). In mammals, two STIM genes, STIM1 and STIM2, and three Orai genes, ORAI1, ORAI2 and ORAI3, have been identified (Zhang et al., 2005; Vig et al., 2006b; Feske et al., 2006). More details about SOCE in non–excitable cells are given elsewhere (Prakriya and Lewis, 2015; Putney, 2017).
3.3.2 Is Store-operated Ca2+ entry relevant to skeletal muscle?
Skeletal muscle highly expresses STIM1 and Orai1 (Stiber et al., 2008; Vig et al., 2008). Both can interact with other channels such as TRPC1 and RyR1 and form complexes that act as the store operated channels complex (Stiber et al., 2008). Also, a longer spliced variant of STIM1, STIM1L, is highly expressed in skeletal muscle, where it colocalizes with Orai1 and binds to actin, forming permanent clusters (Darbellay et al., 2011). The presence of this molecular machinery explains the existence of functional SOCE in adult fibres from skeletal muscle, as first described in mouse EDL bundles after depletion of the SR by repetitive exposure to high K+ in the presence of SERCA inhibitors (Kurebayashi and Ogawa, 2001).
The presence of SOCE in skeletal muscle was also confirmed in myotubes (Pan et al., 2002; Shin et al., 2003; Cherednichenko et al., 2004; Lyfenko and Dirksen, 2008; Stiber et al., 2008), in mechanically skinned rat EDL and soleus fibres (Launikonis et al., 2003; Launikonis and Ríos, 2007; Cully et al., 2016) and in mouse FDB enzymatically dissociated fibres (González Narváez and Castillo, 2007; Bolaños et al., 2009).
The presence of a permanent, actin stabilized coupling of STIM1-Orai1 at the triad may be puzzling, since it may not be necessary for the activation of a classical SOCE in skeletal muscle. Instead, this organization may be relevant to explain the SOCE rapid activation and deactivation associated with every single AP, which has been more recently characterized (Launikonis and Ríos, 2007; Edwards et al., 2010; Koenig et al., 2018; Koenig et al., 2019). This molecular organization may sense RyR-associated SR depletion microdomains, even when the bulk of the SR is not depleted. Neither ICa blockers nor Ca2+ buffers affect that rapid Ca2+ influx (Koenig et al., 2018). This fast-activated SOCE was named as phasic SOCE (pSOCE) to distinguish it from the slower activated, chronic SOCE (cSOCE) (Koenig et al., 2018; Koenig et al., 2019).
Other authors proposed that a triad SOCE pool permits the pSOCE and a second pool at the level of the longitudinal SR activates the cSOCE (Darbellay et al., 2011; Michelucci et al., 2019). This pool would activate slower after acute SR depletion under SERCA blocking and it could be involved in the CEUs formation after strenuous exercise (Boncompagni et al., 2017; Michelucci et al., 2019). It is still possible that the Orai in the elongated TT or the largely ignored longitudinal tubules, interact with the actin associated STIM1L at the I bands, accounting for the longitudinal SR activating slower SOCE. Another, controversial possibility, for the fast activation is that Orai1 would be activated by a direct conformational coupling to RyR and not to STIM1 (Lyfenko and Dirksen, 2008; Dirksen, 2009a).
In this context, Reddish and coworkers (Reddish et al., 2021) were able to follow local RyR1 Ca2+ release events at level of the jSR in mice FDB fibres expressing the low affinity genetically encoded Ca2+ dyes G-CatchER+ and/or CatchER + -JP45. They found that the jSR local Ca2+ release at RyR1 microdomains was 2.1-fold greater than global SR release with much faster kinetics than the depletion in the bulk SR and that if sensed by STIM1, could quickly activate SOCE locally (Reddish et al., 2021).
The existence of small, fast, repetitive tubular Ca2+ transients associated to SOCE is now convincing. However, its relevance to skeletal muscle function is not clear yet. Based on experimental data previously published, the maximum capacity of this mechanism was recently estimated to be between 3 and 70 nM Ca2+ for fibres type I and IIB, respectively, during a single twitch (Rincón et al., 2021). Remembering that the peak sarcoplasmic Ca2+ ranges from 7 to 30 µM and that the free SR Ca2+ is over 1 mM (Section 3.1), it is difficult to assess what the function of this negligible amount of Ca2+ would be: it is neither relevant for refilling the SR nor for sustaining the sarcoplasmic [Ca2+]. One possible explanation is that some errors in the quantitation of the process have arisen because of underestimation of [Ca2+]. For instance, as discussed (Section 3.1), a peak [Ca2+] of 0.2–1 µM in mammalian fibres (Launikonis et al., 2009; Koenig et al., 2018) is untrue, and an inaccurate TT [Ca2+] calibration was acknowledged when we compare papers in which estimated values of 100 µM (Launikonis et al., 2009) were updated to be over 1 mM (Cully et al., 2016). Another option, if all mentioned SOCE estimates turn true (then, ruling out any role in SR refilling or sarcoplasmic [Ca2+] maintenance), and that the rate of exchange of Ca2+ with the tubules is low (Lamboley et al., 2021), is that SOCE in skeletal muscle may be, under certain conditions, a custodian of the fibre total amount of Ca2+. Although complete models considering the internal equilibrium of Ca2+ have already been presented (Section 3.1), models regarding its external equilibrium remain a pending task. Thus, more quantitative efforts should be done to complement structural studies to better understand the role of SOCE in skeletal muscle. Also, some observations should be reproduced in more physiological models, such as intact fibres, before stronger conclusions about the importance of SOCE in skeletal muscle can be drawn.
The apparent small capacity of this mechanism may explain why, in contrast to non-muscle cells, it has not been possible to electrophysiologically record Icrac in intact skeletal muscle fibres (Allard et al., 2006). The expected small size of the currents and the complex structure and electrical properties of the skeletal muscle fibres further complicate this approach.
Two conditions may highlight the importance of SOCE in intact fibres, the exercise, and the absence of CASQ. Mice subjected to treadmill exercise showed tubule remodeling which helped the SOCE machinery assemble following acute exercise and disassemble during recovery (Boncompagni et al., 2017; Michelucci et al., 2019). Preassembled CEUs are occasionally observed in non-exercised muscle, about 2/100 μm2 in EDL and FDB fibres, while in calsequestrin-null or knockout fibres (nCASQ1), increased to 40 and 17/100 μm2 respectively, probably to compensate the reduced store Ca2+ content due to the absence of CASQ. The associated increase in Ca2+ influx by SOCE observed in these fibres, seems to maintain contractile activation in response to repetitive high frequency stimulation and resistance to fatigue (Michelucci et al., 2020). Unfortunately, the Ca2+ signals in these works were not calibrated, avoiding estimating the quantitative importance of SOCE in those results. An increased SOCE during exercise, a condition expected to increase the exchange of Ca2+ with the exterior, is compatible with the hypothesis according to which SOCE may be a keeper of the total amount of Ca2+ inside the fibre. Also, since CASQ null fibres show a reduced amount of total Ca2+ content (Lamboley et al., 2021), an increased SOCE activation can be reexplained as trying to avoid a further reduction of the fibre’s Ca2+ content.
It has been remarked that there may not be enough space at the triad to accommodate STIM1-Orai1 aggregates together with DHPR, RyR1, plus junctophilin, triadin, calsequestrin, etc. However, the fast activation and deactivation previously shown is consistent with preformed complexes STIM1-Orai1 at the triads, close to the RyR1 release channel, and with limited mobility. Also, given the RyR distribution, at least 40% of the space is free of RyR in the jSR (see for instance (Block et al., 1988; Saito et al., 1988; Chen and Kudryashev, 2020)), which may be occupied by the STIM1-Orai1 clusters, i.e., the latter being surrounded by DHPR-RyR clusters as we propose in Figure 1. Our model shows that the movement restrictions imposed by DHPR-RyR to STIM-Orai explain why the CEU have to be preassembled, and fixed, in the jSR-TT membranes. Future super-resolution studies may shed some light on the actual distribution of all these proteins in the triadic space.
Proteins located at the triadic region are known regulators of SOCE. RyR1 and STIM1 colocalize and Ca2+ microdomains close to it regulate the activation of SOCE locally at jSR level (Stiber et al., 2008; Reddish et al., 2021). Removal of the cytoplasmic amino terminal region of the foot portion of the RyR1 abolishes SOCE (Sampieri et al., 2005). Under resting conditions, healthy muscle fibres show a low RyR1 leak but increased RyR1 leak augments the bidirectional Ca2+ exchange with the TT and mitochondrial metabolism to preserve normal contractile function (Lamboley et al., 2021). This is supported by the fact that SOCE-dependent Ca2+ influx is diminished or inhibited in myotubes lacking RyR1 or blocking the Ca2+ release with 100 µM Ry or azumolene (Pan et al., 2002; Zhao et al., 2006; Yarotskyy and Dirksen, 2012). However, in acute or partial SR-Ca2+ depleted intact dissociated FDB fibres, the pretreatment with 50 μM Ry, prior to SOCE activation, enhanced and maintained SOCE activated and only stopped by removing extracellular Ca2+ or applying SOCE blockers such as 2-APB (Bolaños et al., 2013). It is possible that this apparent contradiction early highlighted the existence of different roles of RyR1 in SOCE. Differential RyR conformational changes may confer it a function either as leak sensor or SR depletion sensor. These changes may be inhibited, potentiated or somehow modified by large concentrations of Ry, as shown in other contexts (Paolini et al., 2004). In any case, RyR1 participates Ca2+ influx by SOCE, an observation which deserves further study.
Given its role inside the SR, CASQ1 is a straightforward candidate to be a SOCE regulator. FDB CASQ1 knock down (CASQ1-null) skeletal muscle fibres diminish SR Ca2+ content, favoring the formation of STIM1 aggregates and their interaction with Orai1 that significantly enhanced SOCE (Zhao et al., 2010; Michelucci et al., 2020). On the other hand, the interaction of STIM1 with CASQ1 prevents its association to Orai1 thus limiting SOCE (Zhang et al., 2016). On the contrary, overexpression of the full length CASQ1 reduced SOCE in myotubes (Shin et al., 2003; Zhao et al., 2010), confirming CASQ as a direct modulator of SOCE.
Triadin and junctophilins could also participate in SOCE regulation, as their knockout or knockdown present alterations in the triad structure and an increase in the TT-SR distance, which could diminish STIM1-Orai1 interaction and SOCE (Hirata et al., 2006; Oddoux et al., 2009; Li et al., 2010; van Oort et al., 2011). Indeed, FDB fibres exposed to hypotonic solutions that could increase that distance show SOCE inhibition (Bolaños et al., 2013).
The TRPC channels subfamily consists of seven isoforms (TRPC1-7), which are expressed in skeletal muscle, though there is controversy about TRPC5-7 (Vandebrouck et al., 2002; Krüger et al., 2008; Zanou et al., 2010; Saüc and Frieden, 2017). TRPC1, TRPC3 and TRPC4 reside in the sarcolemma, where they have been reported to associate with RyR1, STIM1, STIM1L, or Orai1, forming ternary or heteromeric complexes (Vandebrouck et al., 2002; Liao et al., 2007; Ong et al., 2007; Liao et al., 2008; Ong et al., 2016; Antigny et al., 2017; Choi et al., 2020).
TRPC1 was the first and most consistently found to somehow participate in SOCE (Ong et al., 2007). In fact, other authors as well proposed that the influx of Ca2+ through Orai1 channels depends on the recruitment of TRPC1 into the plasma membrane where it is activated by STIM1 (Oláh et al., 2011; Ambudkar et al., 2017). In human skeletal myotubes, TRPC1, TRPC4 and STIM1L interact to sustain Ca2+ entry via SOCE that helps to maintain repetitive Ca2+ transients and differentiation (Antigny et al., 2017). Also, TRPC1 would be important as mediator of the RyR involvement in SOCE regulation (Sampieri et al., 2005). However, opposite evidence has also been provided, since TRPC1−/− mice presented almost normal SOCE, but with a decreased resistance to fatigue and lower Ca2+ transients than TRPC1+/+ fibres (Zanou et al., 2010) and contradictory outcomes about the effect of the overexpression of TRPC1 on SOCE have been observed (Ong et al., 2007; Oláh et al., 2011). The colocalization of TRPC members to the SOCE machinery is acknowledged. However, a precise mechanism by which TRPC members regulate SOCE is still lacking, they may constitute a Ca2+ permeable pathway activated by STIM in parallel to Orai, may be regulators of Orai-mediated Ca2+ entry, or both.
In summary, the current renaissance of SOCE studies doubtlessly demonstrated its existence in skeletal muscle and its activation under several conditions. Preassembled complexes and rapid responses are in tune with the main muscle specialization, i.e., its very fast Ca2+ handling ability. As in other subfields of the ECC, such as the sarcoplasmic peak [Ca2+](Section 3.1) or the mitochondria (Section 3.2), putting reliable numbers to SOCE and SOCE-associated phenomena, would help to better understand its relevance in situ. Given its apparent low importance as keeper of the SR and sarcoplasmic [Ca2+], the possibility that it is a guardian of the total Ca2+ inside a muscle fibre arises. Also, clarification of the mechanism of regulation by neighbor proteins (e.g., RyR, TRPC) is pending. Making clearer the structural organization of the SOCE core and accessory machinery would benefit from super-resolution techniques.
3.4 Pharmacology of the excitation–contraction coupling
Table 2 presents details about the most important molecules that have allowed to study mechanisms relevant to the ECC. It highlights the fact that many of them came out to be less specific than initially thought, making some conclusions in several papers untrustworthy. The search for new, better (e.g., highly specific, less toxic) compounds with ECC applications is utterly encouraged.
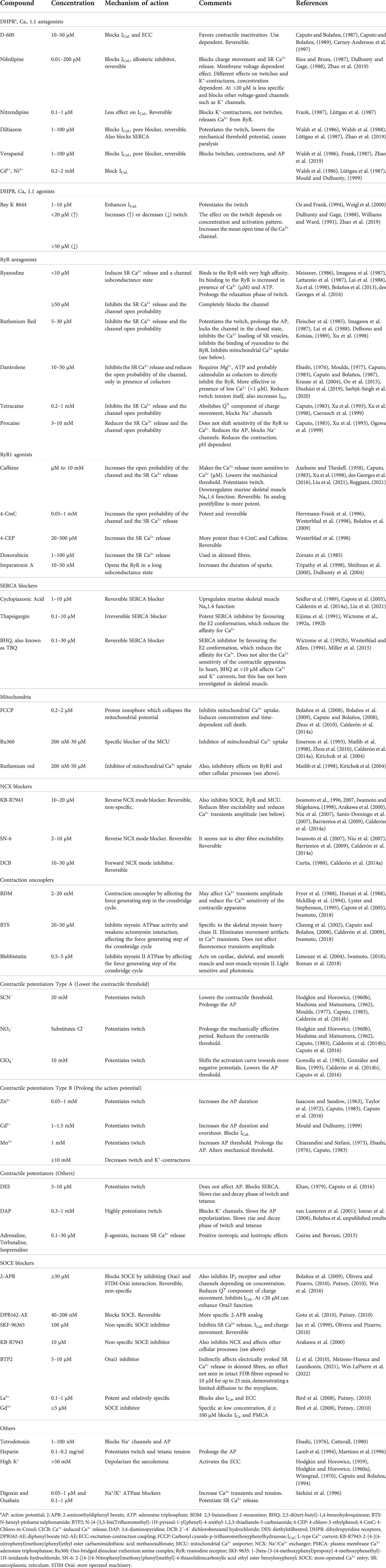
TABLE 2. Pharmacology of the ECC in skeletal muscle.
3.5 Emerging topics
3.5.1 Super-resolution advances and excitation–contraction coupling pioneering studies
Including stimulated emission depletion (STED), structured illumination microscopy (SIM), photo-activated localization microscopy (PALM), stochastic optical reconstruction microscopy (STORM) and their modifications, super-resolution is a novel technique with enormous potential to study skeletal muscle and help solve some issues in ECC. Membrane rearrangements and clustering of proteins, under different experimental conditions, would suitably be studied with up to a 20 nm lateral resolution in fixed and living cells (Jayasinghe et al., 2015; Mishin and Lukyanov, 2019). However, it has been underexploited, probably because of the high costs and still low availability of super-resolution equipment in muscle laboratories.
Monitoring the morphology of the neuromuscular junction using SIM and STORM allowed to propose a new model in which the ACh receptor (AChR) is not located all over the postsynaptic membrane but restricted to the area surrounding the opening of junctional folds (York and Zheng, 2017). Also, STED images showed that width of crests and distance between them with AChR become altered in some neuromuscular diseases (Marinello et al., 2021).
The study of the subsarcolemmal tubular system with STORM revealed an enrichment in longitudinal tubules and branches in different directions (Jayasinghe et al., 2015), being the structural basis of the synchronization of membrane excitation with Ca2+ release from the SR with a safety factor, and probably supporting part of the remodeling potential of the tubular system involved in SOCE. Resolution and quality improvements for STORM images acquisition and ensemble allow to see the boundary membranes and the lumen of the TT (Sun et al., 2014), opening the door for future functional studies restricted to nanoregions.
The approach of pioneers using STORM to see the nanoscale organization of the RyR and CASQ triadic proteins (Jayasinghe et al., 2014), may be extended to simultaneously observe the DHPR-RyR and the STIM-Orai-TRPC clusters. Calculating distances and protein densities in nano- or micro-areas in situ, in different fibre types, would generate a more comprehensive picture of the triadic space. Also, increases in temporal resolution are expected to join the high spatial resolution of nanoscopy to see Ca2+ microdomains, useful for instance in SOCE and mitochondria studies.
3.5.2 Excitation–contraction coupling in induced pluripotent stem cells-derived muscle cells
Notable work demonstrated that mature mammalian cells can be dedifferentiated to render induced pluripotent stem cells (iPSC), which in turn could be redifferentiated to almost any cell of the three germ layers (Takahashi and Yamanaka, 2006; Takahashi et al., 2007). As an example of the potential of iPSC to be differentiated to mesodermic cell types, the authors demonstrated for the first time the generation of iPSC-derived muscle tissue (Takahashi et al., 2007).
iPSC-derived skeletal myocytes obtained in 2D cultures have AP with pretty much the same kinetics as mature muscle fibres. However, although responsive to K+ depolarization in a way non-dependent on external Ca2+, their ECC is very immature, from structural and functional points of view (Skoglund et al., 2014; Lainé et al., 2018). Improved, 3D cultures of induced skeletal muscle bundles showed a greater degree of ECC differentiation, with sizable Ca2+ transients in response to electrical or ACh stimulation (Rao et al., 2018). Nevertheless, the fact that these signals are very slow suggests that the ECC machinery does not reach the maturity of adult muscle fibres in these preparations.
This knowledge, along with its associated technical developments, generates a model suitable for multiple applications in biomedical studies (Andrysiak et al., 2021). Envisioned applications include the understanding of physiological and pathophysiological events and search for new drugs tackling muscle diseases. Although great advances in the methodological protocols have generated quasi-mature muscle cells from iPSC, work is still needed to have fully differentiated fibres to exploit its potential for ECC studies. Also, efforts should be done to have absolute, rather than qualitative (presence or not) or relative (fold change), values of Ca2+ and ECC related variables in iPSC-derived muscle fibres.
4 Conclusion
We have presented the most complete picture of the ECC up to now. Seven decades of exciting research have identified a lot of proteins involved in ECC and a plenty of Ca2+ routes generated by those proteins. Assigning reliable numbers to some of those routes has also been successful. Nonetheless, as of today, two big issues await clarification: what is the exact mechanism of DHPR-RyR coupling and what are the details of the external equilibrium of Ca2+ during ECC. Quantitative approaches, emerging techniques such as super-resolution and iPSC, and finer pharmacology through more specific drugs, may help audacious researchers obtain the answers, hopefully sooner than later.
Author contributions
PB and JC conceived and further developed the idea. Both of them prepared the manuscript, figures and tables. Both read and approved the last version to be submitted and are fully accountable for the content of the work.
Funding
Grants CODI 2020–34909 and CODI 2021–40170 from University of Antioquia (UdeA), to JC. Grants E01708-K and ES03180101 from Planning Office of UdeA, to JC.
Acknowledgments
We want to acknowledge former students and colleagues, especially Carlo Caputo, for helpful discussions during years.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Ainbinder A., Boncompagni S., Protasi F., Dirksen R. T. (2015). Role of Mitofusin-2 in mitochondrial localization and calcium uptake in skeletal muscle. Cell. Calcium 57, 14–24. doi:10.1016/j.ceca.2014.11.002
Allard B., Couchoux H., Pouvreau S., Jacquemond V. (2006). Sarcoplasmic reticulum Ca 2+ release and depletion fail to affect sarcolemmal ion channel activity in mouse skeletal muscle. J. Physiol. 575, 69–81. doi:10.1113/jphysiol.2006.112367
Ambudkar I. S., de Souza L. B., Ong H. L. (2017). TRPC1, Orai1, and STIM1 in SOCE: Friends in tight spaces. Cell. Calcium 63, 33–39. doi:10.1016/j.ceca.2016.12.009
Anderson A. J., Jackson T. D., Stroud D. A., Stojanovski D. (2019). Mitochondria-hubs for regulating cellular biochemistry: Emerging concepts and networks. Open Biol. 9, 190126. doi:10.1098/rsob.190126
Andrysiak K., Stępniewski J., Dulak J. (2021). Human-induced pluripotent stem cell-derived cardiomyocytes, 3D cardiac structures, and heart-on-a-chip as tools for drug research. Pflugers Arch. 473, 1061–1085. doi:10.1007/s00424-021-02536-z
Antigny F., Sabourin J., Saüc S., Bernheim L., Koenig S., Frieden M. (2017). TRPC1 and TRPC4 channels functionally interact with STIM1L to promote myogenesis and maintain fast repetitive Ca(2+) release in human myotubes. Biochim. Biophys. Acta. Mol. Cell. Res. 1864, 806–813. doi:10.1016/j.bbamcr.2017.02.003
Arakawa N., Sakaue M., Yokoyama I., Hashimoto H., Koyama Y., Baba A., et al. (2000). KB-R7943 inhibits store-operated Ca2+ entry in cultured neurons and astrocytes. Biochem. Biophys. Res. Commun. 279, 354–357. doi:10.1006/bbrc.2000.3968
Armstrong C., Bezanilla F., Horowicz P. (1972). Twitches in the presence of ethylene glycol bis(-aminoethyl ether)-N, N´-tetracetic acid. Biochim. Biophys. Acta 267, 605–608. doi:10.1016/0005-2728(72)90194-6
Ashley C. C., Ridgway E. B. (1968). Simultaneous recording of membrane potential, calcium transient and tension in single muscle fibers. Nature 219, 1168–1169. doi:10.1038/2191168a0
Axelsson J., Thesleff S. (1958). Activation of the contractile mechanism in striated muscle. Acta Physiol. Scand. 44, 55–66. doi:10.1111/j.1748-1716.1958.tb01608.x
Balnave C. D., Allen D. G. (1998). Evidence for Na+/Ca2+ exchange in intact single skeletal muscle fibers from the mouse. Am. J. Physiol. 274, C940–C946. doi:10.1152/ajpcell.1998.274.4.C940
Banks Q., Bibollet H., Contreras M., Bennett D. F., Bannister R. A., Schneider M. F., et al. (2021). Voltage sensor movements of Ca(V)1.1 during an action potential in skeletal muscle fibers. Proc. Natl. Acad. Sci. U. S. A. 118, e2026116118. doi:10.1073/pnas.2026116118
Banks Q., Pratt S., Iyer S., Lovering R., Hernández-Ochoa E., Schneider M. (2018). Optical recording of action potential initiation and propagation in mouse skeletal muscle fibers. Biophys. J. 115, 2127–2140. doi:10.1016/j.bpj.2018.10.026
Barclay C. J., Launikonis B. S. (2021). Components of activation heat in skeletal muscle. J. Muscle Res. Cell. Motil. 42, 1–16. doi:10.1007/s10974-019-09547-5
Barnard R. J., Edgerton V. R., Furukawa T., Peter J. B. (1971). Histochemical, biochemical, and contractile properties of red, white, and intermediate fibers. Am. J. Physiol. 220, 410–414. doi:10.1152/ajplegacy.1971.220.2.410
Barrientos G., Bose D. D., Feng W., Padilla I., Pessah I. N. (2009). The Na+/Ca2+ exchange inhibitor 2-(2-(4-(4-nitrobenzyloxy)phenyl)ethyl)isothiourea methanesulfonate (KB-R7943) also blocks ryanodine receptors type 1 (RyR1) and type 2 (RyR2) channels. Mol. Pharmacol. 76, 560–568. doi:10.1124/mol.109.057265
Baughman J. M., Perocchi F., Girgis H. S., Plovanich M., Belcher-Timme C. A., Sancak Y., et al. (2011). Integrative genomics identifies MCU as an essential component of the mitochondrial calcium uniporter. Nature 476, 341–345. doi:10.1038/nature10234
Baylor S. M., Hollingworth S. (2011). Calcium indicators and calcium signalling in skeletal muscle fibres during excitation-contraction coupling. Prog. Biophys. Mol. Biol. 105, 162–179. doi:10.1016/j.pbiomolbio.2010.06.001
Baylor S. M., Hollingworth S. (1988). Fura-2 calcium transients in frog skeletal muscle fibres. J. Physiol. 403, 151–192. doi:10.1113/jphysiol.1988.sp017244
Baylor S. M., Hollingworth S. (2003). Sarcoplasmic reticulum calcium release compared in slow-twitch and fast-twitch fibres of mouse muscle. J. Physiol. 551, 125–138. doi:10.1113/jphysiol.2003.041608
Baylor S. M., Hollingworth S. (2007). Simulation of Ca2+ movements within the sarcomere of fast-twitch mouse fibers stimulated by action potentials. J. Gen. Physiol. 130, 283–302. doi:10.1085/jgp.200709827
Beqollari D., Romberg C. F., Filipova D., Meza U., Papadopoulos S., Bannister R. A. (2015). Rem uncouples excitation-contraction coupling in adult skeletal muscle fibers. J. Gen. Physiol. 146, 97–108. doi:10.1085/jgp.201411314
Berlin J. R., Konishi M. (1993). Ca2+ transients in cardiac myocytes measured with high and low affinity Ca2+ indicators. Biophys. J. 65, 1632–1647. doi:10.1016/S0006-3495(93)81211-6
Bernardi P., Carraro M., Lippe G. (2021). The mitochondrial permeability transition: Recent progress and open questions. FEBS J. doi:10.1111/febs.16254
Beutner G., Sharma V. K., Giovannucci D. R., Yule D. I., Sheu S. S. (2001). Identification of a ryanodine receptor in rat heart mitochondria. J. Biol. Chem. 276, 21482–21488. doi:10.1074/jbc.M101486200
Bezanilla F., Caputo C., Gonzalez-Serratos H., Venosa R. A. (1972). Sodium dependence of the inward spread of activation in isolated twitch muscle fibres of the frog. J. Physiol. 223, 507–523. doi:10.1113/jphysiol.1972.sp009860
Biasutto L., Azzolini M., Szabò I., Zoratti M. (2016). The mitochondrial permeability transition pore in AD 2016: An update. Biochim. Biophys. Acta 1863, 2515–2530. doi:10.1016/j.bbamcr.2016.02.012
Bird G. S., DeHaven W. I., Smyth J. T., Putney J. W. (2008). Methods for studying store-operated calcium entry. Methods 46, 204–212. doi:10.1016/j.ymeth.2008.09.009
Bleunven C., Treves S., Jinyu X., Leo E., Ronjat M., De Waard M., et al. (2008). SRP-27 is a novel component of the supramolecular signalling complex involved in skeletal muscle excitation-contraction coupling. Biochem. J. 411, 343–349. doi:10.1042/BJ20070906
Block B. A., Imagawa T., Campbell K. P., Franzini-Armstrong C. (1988). Structural evidence for direct interaction between the molecular components of the transverse tubule/sarcoplasmic reticulum junction in skeletal muscle. J. Cell. Biol. 107, 2587–2600. doi:10.1083/jcb.107.6.2587
Bloemberg D., Quadrilatero J. (2019). Autophagy, apoptosis, and mitochondria: Molecular integration and physiological relevance in skeletal muscle. Am. J. Physiol. Cell. Physiol. 317, C111–C130. doi:10.1152/ajpcell.00261.2018
Bolaños P., Guillén A., Dipolo R., Caputo C. (2009). Factors affecting SOCE activation in mammalian skeletal muscle fibers. J. Physiol. Sci. 59, 317–328. doi:10.1007/s12576-009-0039-5
Bolaños P., Guillen A., Gámez A., Caputo C. (2013). Quantifying SOCE fluorescence measurements in mammalian muscle fibres. The effects of ryanodine and osmotic shocks. J. Muscle Res. Cell. Motil. 34, 379–393. doi:10.1007/s10974-013-9360-y
Bolaños P., Guillen A., Rojas H., Boncompagni S., Caputo C. (2008). The use of CalciumOrange-5N as a specific marker of mitochondrial Ca2+ in mouse skeletal muscle fibers. Pflugers Arch. 455, 721–731. doi:10.1007/s00424-007-0312-5
Boncompagni S., Michelucci A., Pietrangelo L., Dirksen R. T., Protasi F. (2017). Exercise-dependent formation of new junctions that promote STIM1-Orai1 assembly in skeletal muscle. Sci. Rep. 7, 14286. doi:10.1038/s41598-017-14134-0
Boncompagni S., Pozzer D., Viscomi C., Ferreiro A., Zito E. (2020). Physical and functional cross talk between endo-sarcoplasmic reticulum and mitochondria in skeletal muscle. Antioxid. Redox Signal. 32, 873–883. doi:10.1089/ars.2019.7934
Boncompagni S., Rossi A. E., Micaroni M., Beznoussenko G. V., Polishchuk R. S., Dirksen R. T., et al. (2009). Mitochondria are linked to calcium stores in striated muscle by developmentally regulated tethering structures. Mol. Biol. Cell. 20, 1058–1067. doi:10.1091/mbc.e08-07-0783
Boncompagni S., Thomas M., Lopez J. R., Allen P. D., Yuan Q., Kranias E. G., et al. (2012). Triadin/junctin double null mouse reveals a differential role for triadin and junctin in anchoring CASQ to the jSR and regulating Ca2+ homeostasis. PLoS One 7, e39962. doi:10.1371/journal.pone.0039962
Bottinelli R. (2001). Functional heterogeneity of mammalian single muscle fibres: Do myosin isoforms tell the whole story? Pflugers Arch. 443, 6–17. doi:10.1007/s004240100700
Bottinelli R., Reggiani C. (2000). Human skeletal muscle fibres: Molecular and functional diversity. Prog. Biophys. Mol. Biol. 73, 195–262. doi:10.1016/s0079-6107(00)00006-7
Bottinelli R., Schiaffino S., Reggiani C. (1991). Force-velocity relations and myosin heavy chain isoform compositions of skinned fibres from rat skeletal muscle. J. Physiol. 437, 655–672. doi:10.1113/jphysiol.1991.sp018617
Brooke M. H., Kaiser K. K. (1970). Three “myosin adenosine triphosphatase” systems: The nature of their pH lability and sulfhydryl dependence. J. Histochem. Cytochem. 18, 670–672. doi:10.1177/18.9.670
Butanda-Ochoa A., Höjer G., Morales-Tlalpan V., Díaz-Muñoz M. (2006). Recognition and activation of ryanodine receptors by purines. Curr. Med. Chem. 13, 647–657. doi:10.2174/092986706776055715
Cairns S. P., Borrani F. (2015). β-Adrenergic modulation of skeletal muscle contraction: Key role of excitation-contraction coupling. J. Physiol. 593, 4713–4727. doi:10.1113/JP270909
Calderón J., Bolaños P., Caputo C. (2011). Kinetic changes in tetanic Ca²⁺ transients in enzymatically dissociated muscle fibres under repetitive stimulation. J. Physiol. 589, 5269–5283. doi:10.1113/jphysiol.2011.213314
Calderón J. C., Bolaños P., Caputo C. (2010). Myosin heavy chain isoform composition and Ca2+ transients in fibres from enzymatically dissociated murine soleus and extensor digitorum longus muscles. J. Physiol. 588, 267–279. doi:10.1113/jphysiol.2009.180893
Calderón J. C., Bolaños P., Caputo C. (2014a). Tetanic Ca2+ transient differences between slow- and fast-twitch mouse skeletal muscle fibres: A comprehensive experimental approach. J. Muscle Res. Cell. Motil. 35, 279–293. doi:10.1007/s10974-014-9388-7
Calderón J. C., Bolaños P., Caputo C. (2014b). The excitation-contraction coupling mechanism in skeletal muscle. Biophys. Rev. 6, 133–160. doi:10.1007/s12551-013-0135-x
Calderón J. C., Bolaños P., Torres S. H., Rodríguez-Arroyo G., Caputo C. (2009). Different fibre populations distinguished by their calcium transient characteristics in enzymatically dissociated murine flexor digitorum brevis and soleus muscles. J. Muscle Res. Cell. Motil. 30, 125–137. doi:10.1007/s10974-009-9181-1
Calderón J. C. (2013). Enzymatic dissociation of long muscles from mice: A model for the study of skeletal muscle fiber types. Iatreia 26, 117–126.
Canato M., Scorzeto M., Giacomello M., Protasi F., Reggiani C., Stienen G. J. M. (2010). Massive alterations of sarcoplasmic reticulum free calcium in skeletal muscle fibers lacking calsequestrin revealed by a genetically encoded probe. Proc. Natl. Acad. Sci. U. S. A. 107, 22326–22331. doi:10.1073/pnas.1009168108
Capote J., Bolaños P., Schuhmeier R. P., Melzer W., Caputo C. (2005). Calcium transients in developing mouse skeletal muscle fibres. J. Physiol. 564, 451–464. doi:10.1113/jphysiol.2004.081034
Caputo C., Bolaños P. (1987). Contractile inactivation in frog skeletal muscle fibers. The effects of low calcium, tetracaine, dantrolene, D-600, and nifedipine. J. Gen. Physiol. 89, 421–442. doi:10.1085/jgp.89.3.421
Caputo C., Bolaños P. (2008). Effect of mitochondria poisoning by FCCP on Ca2+ signaling in mouse skeletal muscle fibers. Pflugers Arch. 455, 733–743. doi:10.1007/s00424-007-0317-0
Caputo C., Bolaños P. (1989). Effects of D-600 on intramembrane charge movement of polarized and depolarized frog muscle fibers. J. Gen. Physiol. 94, 43–64. doi:10.1085/jgp.94.1.43
Caputo C., Bolaños P. (1994). Fluo-3 signals associated with potassium contractures in single amphibian muscle fibres. J. Physiol. 481, 119–128. doi:10.1113/jphysiol.1994.sp020423
Caputo C., Bolaños P., Ramos M., DiFranco M. (2016). Differential effects of contractile potentiators on action potential-induced Ca2+ transients of frog and mouse skeletal muscle fibres. J. Muscle Res. Cell. Motil. 37, 169–180. doi:10.1007/s10974-016-9455-3
Caputo C. (2011). Compendio histórico de fisiología muscular. 1st ed. Caracas, Venezuela: Academia de Ciencias Fisicas, Matematicas y Naturales. Available at: http://obras.acfiman.org/2021/01/11/compendio-historico-de-fisiologia-muscular/.
Caputo C., Gimenez M. (1967). Effects of external calcium deprivation on single muscle fibers. J. Gen. Physiol. 50, 2177–2195. doi:10.1085/jgp.50.9.2177
Caputo C. (1983). “Pharmacological investigations of excitation-contraction coupling,” in Handbook of Physiology - skeletal muscle. Editors L. E. Peachey,, and R. H. Adrian (Bethesda, MD: American Physiological Society.), Chap. 14, 381–415.
Carney-Anderson L., Thompson L. V., Huetteman D. A., Donaldson S. K. (1997). GTP gammaS removal of D-600 block of skeletal muscle excitation-contraction coupling. Am. J. Physiol. 272, C572–C581. doi:10.1152/ajpcell.1997.272.2.C572
Catterall W. A. (1980). Neurotoxins that act on voltage-sensitive sodium channels in excitable membranes. Annu. Rev. Pharmacol. Toxicol. 20, 15–43. doi:10.1146/annurev.pa.20.040180.000311
Chen W., Kudryashev M. (2020). Structure of RyR1 in native membranes. EMBO Rep. 21, e49891. doi:10.15252/embr.201949891
Cherednichenko G., Hurne A. M., Fessenden J. D., Lee E. H., Allen P. D., Beam K. G., et al. (2004). Conformational activation of Ca2+ entry by depolarization of skeletal myotubes. Proc. Natl. Acad. Sci. U. S. A. 101, 15793–15798. doi:10.1073/pnas.0403485101
Cheung A., Dantzig J. A., Hollingworth S., Baylor S. M., Goldman Y. E., Mitchison T. J., et al. (2002). A small-molecule inhibitor of skeletal muscle myosin II. Nat. Cell. Biol. 4, 83–88. doi:10.1038/ncb734
Chiarandini D. J., Stefani E. (1973). Effects of manganese on the electrical and mechanical properties of frog skeletal muscle fibres. J. Physiol. 232, 129–147. doi:10.1113/jphysiol.1973.sp010260
Choi J. H., Jeong S. Y., Oh M. R., Allen P. D., Lee E. H. (2020). TRPCs: Influential mediators in skeletal muscle. Cells 9, 850. doi:10.3390/cells9040850
Close R. I. (1972). Dynamic properties of mammalian skeletal muscles. Physiol. Rev. 52, 129–197. doi:10.1152/physrev.1972.52.1.129
Colombini M. (1983). Purification of VDAC (voltage-dependent anion-selective channel) from rat liver mitochondria. J. Membr. Biol. 74, 115–121. doi:10.1007/BF01870500
Colombini M. (1980). Structure and mode of action of a voltage dependent anion-selective channel (VDAC) located in the outer mitochondrial membrane. Ann. N. Y. Acad. Sci. 341, 552–563. doi:10.1111/j.1749-6632.1980.tb47198.x
Colombini M. (2012). VDAC structure, selectivity, and dynamics. Biochim. Biophys. Acta 1818, 1457–1465. doi:10.1016/j.bbamem.2011.12.026
Csapo A., Suzuki T. (1957). A preliminary note on excitation-contraction coupling. Proc. Natl. Acad. Sci. U. S. A. 43, 278–281. doi:10.1073/pnas.43.3.278
Csernoch L., Szentesi P., Sárközi S., Szegedi C., Jona I., Kovács L. (1999). Effects of tetracaine on sarcoplasmic calcium release in mammalian skeletal muscle fibres. J. Physiol. 515, 843–857. doi:10.1111/j.1469-7793.1999.843ab.x
Csordás G., Golenár T., Seifert E. L., Kamer K. J., Sancak Y., Perocchi F., et al. (2013). MICU1 controls both the threshold and cooperative activation of the mitochondrial Ca2+ uniporter. Cell. Metab. 17, 976–987. doi:10.1016/j.cmet.2013.04.020
Csordás G., Renken C., Várnai P., Walter L., Weaver D., Buttle K. F., et al. (2006). Structural and functional features and significance of the physical linkage between ER and mitochondria. J. Cell. Biol. 174, 915–921. doi:10.1083/jcb.200604016
Cully T. R., Edwards J. N., Murphy R. M., Launikonis B. S. (2016). A quantitative description of tubular system Ca 2+ handling in fast- and slow-twitch muscle fibres. J. Physiol. 594, 2795–2810. doi:10.1113/JP271658
Curtis B. A. (1988). Na/Ca exchange and excitation — Contraction coupling in frog fast fibres. J. Muscle Res. Cell. Motil. 9, 415–427. doi:10.1007/BF01774068
Darbellay B., Arnaudeau S., Bader C. R., Konig S., Bernheim L. (2011). STIM1L is a new actin-binding splice variant involved in fast repetitive Ca2+ release. J. Cell. Biol. 194, 335–346. doi:10.1083/jcb.201012157
Dayal A., Fernández-Quintero M. L., Liedl K. R., Grabner M. (2021). Pore mutation N617D in the skeletal muscle DHPR blocks Ca(2+) influx due to atypical high-affinity Ca(2+) binding. Elife 10, e63435. doi:10.7554/eLife.63435
Dayal A., Schrötter K., Pan Y., Föhr K., Melzer W., Grabner M. (2017). The Ca(2+) influx through the mammalian skeletal muscle dihydropyridine receptor is irrelevant for muscle performance. Nat. Commun. 8, 475. doi:10.1038/s41467-017-00629-x
de Brito O. M., Scorrano L. (2008). Mitofusin 2 tethers endoplasmic reticulum to mitochondria. Nature 456, 605–610. doi:10.1038/nature07534
De Mario A., Gherardi G., Rizzuto R., Mammucari C. (2021). Skeletal muscle mitochondria in health and disease. Cell. Calcium 94, 102357. doi:10.1016/j.ceca.2021.102357
De Stefani D., Raffaello A., Teardo E., Szabò I., Rizzuto R. (2011). A forty-kilodalton protein of the inner membrane is the mitochondrial calcium uniporter. Nature 476, 336–340. doi:10.1038/nature10230
De Stefani D., Rizzuto R., Pozzan T. (2016). Enjoy the trip: Calcium in mitochondria back and forth. Annu. Rev. Biochem. 85, 161–192. doi:10.1146/annurev-biochem-060614-034216
Delbono O., Kotsias B. A. (1989). Ruthenium red effect on mechanical and electrical properties of mammalian skeletal muscle. Life Sci. 45, 1699–1708. doi:10.1016/0024-3205(89)90280-4
Delbono O., Stefani E. (1993). Calcium transients in single mammalian skeletal muscle fibres. J. Physiol. 463, 689–707. doi:10.1113/jphysiol.1993.sp019617
des Georges A., Clarke O. B., Zalk R., Yuan Q., Condon K. J., Grassucci R. A., et al. (2016). Structural basis for gating and activation of RyR1. Cell. 167, 145–157. e17. doi:10.1016/j.cell.2016.08.075
Díaz-Vegas A., Eisner V., Jaimovich E. (2019). Skeletal muscle excitation-metabolism coupling. Arch. Biochem. Biophys. 664, 89–94. doi:10.1016/j.abb.2019.01.037
DiFranco M., Woods C. E., Capote J., Vergara J. L. (2008). Dystrophic skeletal muscle fibers display alterations at the level of calcium microdomains. Proc. Natl. Acad. Sci. U. S. A. 105, 14698–14703. doi:10.1073/pnas.0802217105
Dirksen R. T. (2009a). Checking your SOCCs and feet: The molecular mechanisms of Ca 2+ entry in skeletal muscle. J. Physiol. 587, 3139–3147. doi:10.1113/jphysiol.2009.172148
Dirksen R. T. (2009b). Sarcoplasmic reticulum-mitochondrial through-space coupling in skeletal muscle. Appl. Physiol. Nutr. Metab. 34, 389–395. doi:10.1139/H09-044
Diszházi G., Magyar Z. É., Mótyán J. A., Csernoch L., Jóna I., Nánási P. P., et al. (2019). Dantrolene requires Mg(2+) and ATP to inhibit the ryanodine receptor. Mol. Pharmacol. 96, 401–407. doi:10.1124/mol.119.116475
Dubowitz V., Pearse A. G. (1960). A comparative histochemical study of oxidative enzyme and phosphorylase activity in skeletal muscle. Z. Zellforch. Microsk. Anat. Histochem. 2, 105–117. doi:10.1007/BF00744575
Dulhunty A. F., Curtis S. M., Watson S., Cengia L., Casarotto M. G. (2004). Multiple actions of imperatoxin A on ryanodine receptors: Interactions with the II-III loop "A" fragment. J. Biol. Chem. 279, 11853–11862. doi:10.1074/jbc.M310466200
Dulhunty A. F., Gage P. W. (1988). Effects of extracellular calcium concentration and dihydropyridines on contraction in mammalian skeletal muscle. J. Physiol. 399, 63–80. doi:10.1113/jphysiol.1988.sp017068
Ebashi S., Endo M. (1968). Calcium ion and muscle contraction. Prog. Biophys. Mol. Biol. 18, 123–183. doi:10.1016/0079-6107(68)90023-0
Ebashi S., Endo M., Otsuki I. (1969). Control of muscle contraction. Q. Rev. Biophys. 2, 351–384. doi:10.1017/s0033583500001190
Ebashi S. (1976). Excitation-contraction coupling. Annu. Rev. Physiol. 38, 293–313. doi:10.1146/annurev.ph.38.030176.001453
Ebashi S., Lipmann F. (1962). Adenosine-triphosphate-linked concentration of calcium ions in a particulate fraction of rabbit muscle. J. Cell. Biol. 14, 389–400. doi:10.1083/jcb.14.3.389
Edwards J. N., Cully T. R., Shannon T. R., Stephenson D. G., Launikonis B. S. (2012). Longitudinal and transversal propagation of excitation along the tubular system of rat fast-twitch muscle fibres studied by high speed confocal microscopy. J. Physiol. 590, 475–492. doi:10.1113/jphysiol.2011.221796
Edwards J. N., Murphy R. M., Cully T. R., von Wegner F., Friedrich O., Launikonis B. S. (2010). Ultra-rapid activation and deactivation of store-operated Ca2+ entry in skeletal muscle. Cell. Calcium 47, 458–467. doi:10.1016/j.ceca.2010.04.001
Efremov R. G., Leitner A., Aebersold R., Raunser S. (2015). Architecture and conformational switch mechanism of the ryanodine receptor. Nature 517, 39–43. doi:10.1038/nature13916
Emerson J., Clarke M., Ying W., Sanadi D. (1993). The component of ruthenium red responsible for inhibition of mitochondrial calcium ion transport. Spectra, electrochemistry, and aquation kinetics. Crystal structure of .mu.-O-[(HCO2)(NH3)4Ru]2Cl3. J. Am. Chem. Soc. 115, 11799–11805. doi:10.1021/ja00078a019
Endo M. (1964). Entry of a dye into the sarcotubular system of muscle. Nature 202, 1115–1116. doi:10.1038/2021115b0
Engel W. K. (1962). The essentiality of histo- and cytochemical studies of skeletal muscle in the investigation of neuromuscular disease. Neurology 12, 778–794. doi:10.1212/wnl.51.3.778
Escobar A. L., Monck J. R., Fernandez J. M., Vergara J. L. (1994). Localization of the site of Ca2+ release at the level of a single sarcomere in skeletal muscle fibres. Nature 367, 739–741. doi:10.1038/367739a0
Feng W., Tu J., Pouliquin P., Cabrales E., Shen X., Dulhunty A., et al. (2008). Dynamic regulation of ryanodine receptor type 1 (RyR1) channel activity by Homer 1. Cell. Calcium 43, 307–314. doi:10.1016/j.ceca.2007.06.001
Feske S., Gwack Y., Prakriya M., Srikanth S., Puppel S.-H., Tanasa B., et al. (2006). A mutation in Orai1 causes immune deficiency by abrogating CRAC channel function. Nature 441, 179–185. doi:10.1038/nature04702
Fieni F., Lee S. B., Jan Y. N., Kirichok Y. (2012). Activity of the mitochondrial calcium uniporter varies greatly between tissues. Nat. Commun. 3, 1317. doi:10.1038/ncomms2325
Fill M., Coronado R., Mickelson J. R., Vilven J., Ma J. J., Jacobson B. A., et al. (1990). Abnormal ryanodine receptor channels in malignant hyperthermia. Biophys. J. 57, 471–475. doi:10.1016/S0006-3495(90)82563-7
Finkel T. (2011). Signal transduction by reactive oxygen species. J. Cell. Biol. 194, 7–15. doi:10.1083/jcb.201102095
Fleischer S., Ogunbunmi E. M., Dixon M. C., Fleer E. A. (1985). Localization of Ca2+ release channels with ryanodine in junctional terminal cisternae of sarcoplasmic reticulum of fast skeletal muscle. Proc. Natl. Acad. Sci. U. S. A. 82, 7256–7259. doi:10.1073/pnas.82.21.7256
Frank G. B. (1987). Pharmacological studies of excitation–contraction coupling in skeletal muscle. Can. J. Physiol. Pharmacol. 65, 711–716. doi:10.1139/y87-117
Franzini-Armstrong C., Boncompagni S. (2011). The evolution of the mitochondria-to-calcium release units relationship in vertebrate skeletal muscles. J. Biomed. Biotechnol. 2011, 830573. doi:10.1155/2011/830573
Franzini-Armstrong C., Jorgensen A. O. (1994). Structure and development of E-C coupling units in skeletal muscle. Annu. Rev. Physiol. 56, 509–534. doi:10.1146/annurev.ph.56.030194.002453
Franzini-Armstrong C., Kenney L. J., Varriano-Marston E. (1987). The structure of calsequestrin in triads of vertebrate skeletal muscle: A deep-etch study. J. Cell. Biol. 105, 49–56. doi:10.1083/jcb.105.1.49
Franzini-Armstrong C., Porter K. R. (1964). Sarcolemmal invaginations constituting the t system in fish muscle fibers. J. Cell. Biol. 22, 675–696. doi:10.1083/jcb.22.3.675
Franzini-Armstrong C., Protasi F., Ramesh V. (1998). Comparative ultrastructure of Ca2+ release units in skeletal and cardiac muscle. Ann. N. Y. Acad. Sci. 853, 20–30. doi:10.1111/j.1749-6632.1998.tb08253.x
Franzini-Armstrong C. (1999). The sarcoplasmic reticulum and the control of muscle contraction. FASEB J. 13, S266–S270. doi:10.1096/fasebj.13.9002.s266
Fraser J. A., Huang C. L.-H., Pedersen T. H. (2011). Relationships between resting conductances, excitability, and t-system ionic homeostasis in skeletal muscle. J. Gen. Physiol. 138, 95–116. doi:10.1085/jgp.201110617
Friedrich O., Fink R. H. A., von Wegner F. (2010). New factors contributing to dynamic calcium regulation in the skeletal muscle triad—A crowded place. Biophys. Rev. 2, 29–38. doi:10.1007/s12551-009-0027-2
Fryer M. W., Neering I. R., Stephenson D. G. (1988). Effects of 2, 3-butanedione monoxime on the contractile activation properties of fast- and slow-twitch rat muscle fibres. J. Physiol. 407, 53–75. doi:10.1113/jphysiol.1988.sp017403
Füchtbauer E. M., Rowlerson A. M., Götz K., Friedrich G., Mabuchi K., Gergely J., et al. (1991). Direct correlation of parvalbumin levels with myosin isoforms and succinate dehydrogenase activity on frozen sections of rodent muscle. J. Histochem. Cytochem. 39, 355–361. doi:10.1177/39.3.1825216
Garbincius J. F., Elrod J. W. (2022). Mitochondrial calcium exchange in physiology and disease. Physiol. Rev. 102, 893–992. doi:10.1152/physrev.00041.2020
Gee K. R., Brown K. A., Chen W. N., Bishop-Stewart J., Gray D., Johnson I. (2000). Chemical and physiological characterization of fluo-4 Ca(2+)-indicator dyes. Cell. Calcium 27, 97–106. doi:10.1054/ceca.1999.0095
Gherardi G., De Mario A., Mammucari C. (2021). The mitochondrial calcium homeostasis orchestra plays its symphony: Skeletal muscle is the guest of honor. Int. Rev. Cell. Mol. Biol. 362, 209–259. doi:10.1016/bs.ircmb.2021.03.005
Gillis J. M., Thomason D., Lefèvre J., Kretsinger R. H. (1982). Parvalbumins and muscle relaxation: A computer simulation study. J. Muscle Res. Cell. Motil. 3, 377–398. doi:10.1007/BF00712090
Gomolla M., Gottschalk G., Lüttgau H. C. (1983). Perchlorate-induced alterations in electrical and mechanical parameters of frog skeletal muscle fibres. J. Physiol. 343, 197–214. doi:10.1113/jphysiol.1983.sp014888
González A., Ríos E., Gonzalez A. (1993). Perchlorate enhances transmission in skeletal muscle excitation-contraction coupling. J. Gen. Physiol. 102, 373–421. doi:10.1085/jgp.102.3.373
González Narváez A. A., Castillo A. (2007). Ca2+ store determines gating of store operated calcium entry in mammalian skeletal muscle. J. Muscle Res. Cell. Motil. 28, 105–113. doi:10.1007/s10974-007-9105-x
González-Serratos H. (1971). Inward spread of activation in vertebrate muscle fibres. J. Physiol. 212, 777–799. doi:10.1113/jphysiol.1971.sp009356
Goonasekera S. A., Beard N. A., Groom L., Kimura T., Lyfenko A. D., Rosenfeld A., et al. (2007). Triadin binding to the C-terminal luminal loop of the ryanodine receptor is important for skeletal muscle excitation contraction coupling. J. Gen. Physiol. 130, 365–378. doi:10.1085/jgp.200709790
Goto J.-I., Suzuki A. Z., Ozaki S., Matsumoto N., Nakamura T., Ebisui E., et al. (2010). Two novel 2-aminoethyl diphenylborinate (2-APB) analogues differentially activate and inhibit store-operated Ca2+ entry via STIM proteins. Cell. Calcium 47, 1–10. doi:10.1016/j.ceca.2009.10.004
Gottschalk B., Klec C., Leitinger G., Bernhart E., Rost R., Bischof H., et al. (2019). MICU1 controls cristae junction and spatially anchors mitochondrial Ca(2+) uniporter complex. Nat. Commun. 10, 3732. doi:10.1038/s41467-019-11692-x
Gregg R. G., Messing A., Strube C., Beurg M., Moss R., Behan M., et al. (1996). Absence of the beta subunit (cchb1) of the skeletal muscle dihydropyridine receptor alters expression of the alpha 1 subunit and eliminates excitation-contraction coupling. Proc. Natl. Acad. Sci. U. S. A. 93, 13961–13966. doi:10.1073/pnas.93.24.13961
Grynkiewicz G., Poenie M., Tsien R. Y. (1985). A new generation of Ca2+ indicators with greatly improved fluorescence properties. J. Biol. Chem. 260, 3440–3450. doi:10.1016/s0021-9258(19)83641-4
Guo W., Campbell K. P. (1995). Association of triadin with the ryanodine receptor and calsequestrin in the lumen of the sarcoplasmic reticulum. J. Biol. Chem. 270, 9027–9030. doi:10.1074/jbc.270.16.9027
Harkins A. B., Kurebayashi N., Baylor S. M. (1993). Resting myoplasmic free calcium in frog skeletal muscle fibers estimated with fluo-3. Biophys. J. 65, 865–881. doi:10.1016/S0006-3495(93)81112-3
Hasselbach W., Makinose M. (1962). ATP and active transport. Biochem. Biophys. Res. Commun. 7, 132–136. doi:10.1016/0006-291x(62)90161-4
Hasselbach W. (1964). Relaxing factor and the relaxation of muscle. Prog. Biophys. Mol. Biol. 14, 167–IN2. doi:10.1016/S0079-6107(64)80006-7
Hasselbach W. (1998). The Ca(2+)-ATPase of the sarcoplasmic reticulum in skeletal and cardiac muscle. An overview from the very beginning to more recent prospects. Ann. N. Y. Acad. Sci. 853, 1–8. doi:10.1111/j.1749-6632.1998.tb08251.x
Head S. I. (1993). Membrane potential, resting calcium and calcium transients in isolated muscle fibres from normal and dystrophic mice. J. Physiol. 469, 11–19. doi:10.1113/jphysiol.1993.sp019801
Heilbrunn L. V., Wiercinski F. J. (1947). The action of various cations on muscle protoplasm. J. Cell. Comp. Physiol. 29, 15–32. doi:10.1002/jcp.1030290103
Heizmann C. W., Berchtold M. W., Rowlerson A. M. (1982). Correlation of parvalbumin concentration with relaxation speed in mammalian muscles. Proc. Natl. Acad. Sci. U. S. A. 79, 7243–7247. doi:10.1073/pnas.79.23.7243
Herrmann-Frank A., Richter M., Sarközi S., Mohr U., Lehmann-Horn F. (1996). 4-chloro-m-cresol, a potent and specific activator of the skeletal muscle ryanodine receptor. Biochim. Biophys. Acta 1289, 31–40. doi:10.1016/0304-4165(95)00131-X
Hidalgo C., Donoso P., Carrasco M. A. (2005). The ryanodine receptors Ca2+ release channels: Cellular redox sensors? IUBMB Life 57, 315–322. doi:10.1080/15216540500092328
Hill A. V. (1948). On the time required for diffusion and its relation to processes in muscle. Proc. R. Soc. B 135, 446–453. doi:10.1098/rspb.1948.0022
Hill A. V. (1949). The onset of contraction. Proc. R. Soc. Lond. B Biol. Sci. 136, 242–254. doi:10.1098/rspb.1949.0023
Hirata Y., Brotto M., Weisleder N., Chu Y., Lin P., Zhao X., et al. (2006). Uncoupling store-operated Ca2+ entry and altered Ca2+ release from sarcoplasmic reticulum through s svilencing of junctophilin genes. Biophys. J. 90, 4418–4427. doi:10.1529/biophysj.105.076570
Hodgkin A. L., Horowicz P. (1960a). Potassium contractures in single muscle fibres. J. Physiol. 153, 386–403. doi:10.1113/jphysiol.1960.sp006541
Hodgkin A. L., Horowicz P. (1960b). The effect of nitrate and other anions on the mechanical response of single muscle fibres. J. Physiol. 153, 404–412. doi:10.1113/jphysiol.1960.sp006542
Hodgkin A. L., Horowicz P. (1959). The influence of potassium and chloride ions on the membrane potential of single muscle fibres. J. Physiol. 148, 127–160. doi:10.1113/jphysiol.1959.sp006278
Holash R. J., MacIntosh B. R. (2019). A stochastic simulation of skeletal muscle calcium transients in a structurally realistic sarcomere model using MCell. PLoS Comput. Biol. 15, e1006712. doi:10.1371/journal.pcbi.1006712
Hollingworth S., Gee K. R., Baylor S. M. (2009). Low-affinity Ca2+ indicators compared in measurements of skeletal muscle Ca2+ transients. Biophys. J. 97, 1864–1872. doi:10.1016/j.bpj.2009.07.021
Hollingworth S., Harkins A., Baylor S. (1990). Absorbance and fluorescence signals from fluo-3 in isolated frog skeletal muscle fibres. J. Physiol. 430, 68P. doi:10.1113/jphysiol.1990.sp018314
Hollingworth S., Kim M. M., Baylor S. M. (2012). Measurement and simulation of myoplasmic calcium transients in mouse slow-twitch muscle fibres. J. Physiol. 590, 575–594. doi:10.1113/jphysiol.2011.220780
Hollingworth S., Zhao M., Baylor S. M. (1996). The amplitude and time course of the myoplasmic free [Ca2+] transient in fast-twitch fibers of mouse muscle. J. Gen. Physiol. 108, 455–469. doi:10.1085/jgp.108.5.455
Horiuti K., Higuchi H., Umazume Y., Konishi M., Okazaki O., Kurihara S. (1988). Mechanism of action of 2, 3-butanedione 2-monoxime on contraction of frog skeletal muscle fibres. J. Muscle Res. Cell. Motil. 9, 156–164. doi:10.1007/BF01773737
Hoth M., Penner R. (1992). Depletion of intracellular calcium stores activates a calcium current in mast cells. Nature 355, 353–356. doi:10.1038/355353a0
Hou X., Burstein S. R., Long S. B. (2018). Structures reveal opening of the store-operated calcium channel Orai. Elife 7, e36758. doi:10.7554/eLife.36758
Huang X., Sun L., Ji S., Zhao T., Zhang W., Xu J., et al. (2013). Kissing and nanotunneling mediate intermitochondrial communication in the heart. Proc. Natl. Acad. Sci. U. S. A. 110, 2846–2851. doi:10.1073/pnas.1300741110
Hüser J., Blatter L. A. (1999). Fluctuations in mitochondrial membrane potential caused by repetitive gating of the permeability transition pore. Biochem. J. 343 Pt 2, 311–317. doi:10.1042/bj3430311
Huxley A. F., Taylor R. E. (1958). Local activation of striated muscle fibres. J. Physiol. 144, 426–441. doi:10.1113/jphysiol.1958.sp006111
Huxley H. E. (1964). Evidence for continuity between the central elements of the triads and extracellular space in frog sartorius muscle. Nature 202, 1067–1071. doi:10.1038/2021067b0
Idoux R., Fuster C., Jacquemond V., Dayal A., Grabner M., Charnet P., et al. (2020). Divalent cations permeation in a Ca(2+) non-conducting skeletal muscle dihydropyridine receptor mouse model. Cell. Calcium 91, 102256. doi:10.1016/j.ceca.2020.102256
Imagawa T., Smith J. S., Coronado R., Campbell K. P. (1987). Purified ryanodine receptor from skeletal muscle sarcoplasmic reticulum is the Ca2+-permeable pore of the calcium release channel. J. Biol. Chem. 262, 16636–16643. doi:10.1016/s0021-9258(18)49303-9
Ionno M., Moyer M., Pollarine J., van Lunteren E. (2008). Inotropic effects of the K+ channel blocker 3, 4-diaminopyridine on fatigued diaphragm muscle. Respir. Physiol. Neurobiol. 160, 45–53. doi:10.1016/j.resp.2007.08.003
Isaacson A., Sandow A. (1963). Effects of zinc on responses of skeletal muscle. J. Gen. Physiol. 46, 655–677. doi:10.1085/jgp.46.4.655
Iwamoto H. (2018). Effects of myosin inhibitors on the X-ray diffraction patterns of relaxed and calcium-activated rabbit skeletal muscle fibers. Biophys. Physicobiol. 15, 111–120. doi:10.2142/biophysico.15.0_111
Iwamoto T., Shigekawa M. (1998). Differential inhibition of Na + /Ca 2+ exchanger isoforms by divalent cations and isothiourea derivative. Am. J. Physiol. 275, C423–C430. doi:10.1152/ajpcell.1998.275.2.C423
Iwamoto T., Watanabe Y., Kita S., Blaustein M. (2007). Na+/Ca2+ exchange inhibitors: A new class of calcium regulators. Cardiovasc. Hematol. Disord. Drug Targets 7, 188–198. doi:10.2174/187152907781745288
Iwamoto T., Watano T., Shigekawa M. (1996). A novel isothiourea derivative selectively inhibits the reverse mode of Na+/Ca2+ exchange in cells expressing NCX1. J. Biol. Chem. 271, 22391–22397. doi:10.1074/jbc.271.37.22391
Jan C. R., Ho C. M., Wu S. N., Tseng C. J. (1999). Multiple effects of 1-[beta-[3-(4-methoxyphenyl)propoxy]-4-methoxyphenethyl]-1H-imidazole hydrochloride (SKF 96365) on Ca2+ signaling in MDCK cells: Depletion of thapsigargin-sensitive Ca2+ store followed by capacitative Ca2+ entry, activation of a direct Ca2+ entry, and inhibition of thapsigargin-induced capacitative Ca2+ entry. Naunyn. Schmiedeb. Arch. Pharmacol. 359, 92–101. doi:10.1007/PL00005336
Jayasinghe I. D., Clowsley A. H., Munro M., Hou Y., Crossman D. J., Soeller C. (2015). Revealing T-tubules in striated muscle with new optical super-resolution microscopy techniquess. Eur. J. Transl. Myol. 25, 4747. doi:10.4081/ejtm.2015.4747
Jayasinghe I. D., Launikonis B. S. (2013). Three-dimensional reconstruction and analysis of the tubular system of vertebrate skeletal muscle. J. Cell. Sci. 126, 4048–4058. doi:10.1242/jcs.131565
Jayasinghe I. D., Lo H. P., Morgan G. P., Baddeley D., Parton R. G., Soeller C., et al. (2013). Examination of the subsarcolemmal tubular system of mammalian skeletal muscle fibers. Biophys. J. 104, L19–L21. doi:10.1016/j.bpj.2013.04.029
Jayasinghe I. D., Munro M., Baddeley D., Launikonis B. S., Soeller C. (2014). Observation of the molecular organization of calcium release sites in fast- and slow-twitch skeletal muscle with nanoscale imaging. J. R. Soc. Interface 11, 20140570. doi:10.1098/rsif.2014.0570
Jiang D., Zhao L., Clapham D. E. (2009). Genome-wide RNAi screen identifies Letm1 as a mitochondrial Ca2+/H+ antiporter. Science 326, 144–147. doi:10.1126/science.1175145
Jöbsis F., O’Connor M. (1966). Calcium release and reabsorption in the sartorius muscle of the toad. Biochem. Biophys. Res. Commun. 25, 246–252. doi:10.1016/0006-291x(66)90588-2
Jorgensen A. O., Jones L. R. (1986). Localization of phospholamban in slow but not fast canine skeletal muscle fibers. An immunocytochemical and biochemical study. J. Biol. Chem. 261, 3775–3781. doi:10.1016/s0021-9258(17)35715-0
Jung D. H., Mo S. H., Kim D. H. (2006). Calumenin, a multiple EF-hands Ca2+-binding protein, interacts with ryanodine receptor-1 in rabbit skeletal sarcoplasmic reticulum. Biochem. Biophys. Res. Commun. 343, 34–42. doi:10.1016/j.bbrc.2006.02.115
Karam C., Yi J., Xiao Y., Dhakal K., Zhang L., Li X., et al. (2017). Absence of physiological Ca(2+) transients is an initial trigger for mitochondrial dysfunction in skeletal muscle following denervation. Skelet. Muscle 7, 6. doi:10.1186/s13395-017-0123-0
Katerinopoulos H. E., Foukaraki E. (2002). Polycarboxylate fluorescent indicators as ion concentration probes in biological systems. Curr. Med. Chem. 9, 275–306. doi:10.2174/0929867023371193
Kavanagh N. I., Ainscow E. K., Brand M. D. (2000). Calcium regulation of oxidative phosphorylation in rat skeletal muscle mitochondria. Biochim. Biophys. Acta 1457, 57–70. doi:10.1016/s0005-2728(00)00054-2
Khan A. R. (1979). Effects of diethyl-stilboestrol on single fibres of frog skeletal muscle. Acta Physiol. Scand. 106, 69–73. doi:10.1111/j.1748-1716.1979.tb06371.x
Kielley W. W., Meyerhof O. (1948). Studies on adenosinetriphosphatase of muscle; a new magnesium-activated a denosinetriphosphatase. J. Biol. Chem. 176, 591–601. doi:10.1016/s0021-9258(19)52676-x
Kijima Y., Ogunbunmi E., Fleischer S. (1991). Drug action of thapsigargin on the Ca2+ pump protein of sarcoplasmic reticulum. J. Biol. Chem. 266, 22912–22918. doi:10.1016/s0021-9258(18)54441-0
Kirichok Y., Krapivinsky G., Clapham D. E. (2004). The mitochondrial calcium uniporter is a highly selective ion channel. Nature 427, 360–364. doi:10.1038/nature02246
Koenig X., Choi R. H., Launikonis B. S. (2018). Store-operated Ca2+ entry is activated by every action potential in skeletal muscle. Commun. Biol. 1, 31. doi:10.1038/s42003-018-0033-7
Koenig X., Choi R. H., Schicker K., Singh D. P., Hilber K., Launikonis B. S. (2019). Mechanistic insights into store-operated Ca2+ entry during excitation-contraction coupling in skeletal muscle. Biochim. Biophys. Acta. Mol. Cell. Res. 1866, 1239–1248. doi:10.1016/j.bbamcr.2019.02.014
Konishi M. (1998). Cytoplasmic free concentrations of Ca2+ and Mg2+ in skeletal muscle fibers at rest and during contraction. Jpn. J. Physiol. 48, 421–438. doi:10.2170/jjphysiol.48.421
Kornmann B., Currie E., Collins S. R., Schuldiner M., Nunnari J., Weissman J. S., et al. (2009). An ER-mitochondria tethering complex revealed by a synthetic biology screen. Science 325, 477–481. doi:10.1126/science.1175088
Krause T., Gerbershagen M. U., Fiege M., Weißhorn R., Wappler F. (2004). Dantrolene? A review of its pharmacology, therapeutic use and new developments. Anaesthesia 59, 364–373. doi:10.1111/j.1365-2044.2004.03658.x
Krüger J., Kunert-Keil C., Bisping F., Brinkmeier H. (2008). Transient receptor potential cation channels in normal and dystrophic mdx muscle. Neuromuscul. Disord. 18, 501–513. doi:10.1016/j.nmd.2008.04.003
Kuffler S. W. (1947). Membrane changes during excitation and inhibition of the contractile mechanism. Ann. N. Y. Acad. Sci. 47, 767–779. doi:10.1111/j.1749-6632.1947.tb31735.x
Kumagai H., Ebashi S., Takeda F. (1955). Essential relaxing factor in muscle other than myokinase and creatine phosphokinase. Nature 176, 166. doi:10.1038/176166a0
Kumar A., Chakravarty H., Bal N. C., Balaraju T., Jena N., Misra G., et al. (2013). Identification of calcium binding sites on calsequestrin 1 and their implications for polymerization. Mol. Biosyst. 9, 1949–1957. doi:10.1039/c3mb25588c
Kurebayashi N., Ogawa Y. (2001). Depletion of Ca2+ in the sarcoplasmic reticulum stimulates Ca2+ entry into mouse skeletal muscle fibres. J. Physiol. 533, 185–199. doi:10.1111/j.1469-7793.2001.0185b.x
Kuznetsov A. V., Troppmair J., Sucher R., Hermann M., Saks V., Margreiter R. (2006). Mitochondrial subpopulations and heterogeneity revealed by confocal imaging: Possible physiological role? Biochim. Biophys. Acta 1757, 686–691. doi:10.1016/j.bbabio.2006.03.014
Lai F. A., Erickson H. P., Rousseau E., Liu Q. Y., Meissner G. (1988). Purification and reconstitution of the calcium release channel from skeletal muscle. Nature 331, 315–319. doi:10.1038/331315a0
Lainé J., Skoglund G., Fournier E., Tabti N. (2018). Development of the excitation-contraction coupling machinery and its relation to myofibrillogenesis in human iPSC-derived skeletal myocytes. Skelet. Muscle 8, 1. doi:10.1186/s13395-017-0147-5
Lamb G. D., Posterino G. S., Stephenson D. G. (1994). Effects of heparin on excitation-contraction coupling in skeletal muscle toad and rat. J. Physiol. 474, 319–329. doi:10.1113/jphysiol.1994.sp020024
Lamboley C. R., Pearce L., Seng C., Meizoso-Huesca A., Singh D. P., Frankish B. P., et al. (2021). Ryanodine receptor leak triggers fiber Ca2+ redistribution to preserve force and elevate basal metabolism in skeletal muscle. Sci. Adv. 7, eabi7166. doi:10.1126/sciadv.abi7166
Lattanzio F. A. J., Bartschat D. K. (1991). The effect of pH on rate constants, ion selectivity and thermodynamic properties of fluorescent calcium and magnesium indicators. Biochem. Biophys. Res. Commun. 177, 184–191. doi:10.1016/0006-291x(91)91966-g
Lattanzio F. A. J., Schlatterer R. G., Nicar M., Campbell K. P., Sutko J. L. (1987). The effects of ryanodine on passive calcium fluxes across sarcoplasmic reticulum membranes. J. Biol. Chem. 262, 2711–2718. doi:10.1016/s0021-9258(18)61565-0
Launikonis B. S., Barnes M., Stephenson D. G. (2003). Identification of the coupling between skeletal muscle store-operated Ca 2+ entry and the inositol trisphosphate receptor. Proc. Natl. Acad. Sci. U. S. A. 100, 2941–2944. doi:10.1073/pnas.0536227100
Launikonis B. S., Ríos E. (2007). Store-operated Ca2+ entry during intracellular Ca2+ release in mammalian skeletal muscle. J. Physiol. 583, 81–97. doi:10.1113/jphysiol.2007.135046
Launikonis B. S., Stephenson D. G., Friedrich O. (2009). Rapid Ca2+ flux through the transverse tubular membrane, activated by individual action potentials in mammalian skeletal muscle. J. Physiol. 587, 2299–2312. doi:10.1113/jphysiol.2009.168682
Lavorato M., Formenti F., Franzini-Armstrong C. (2020). The structural basis for intermitochondrial communications is fundamentally different in cardiac and skeletal muscle. Exp. Physiol. 105, 606–612. doi:10.1113/EP087503
Lavorato M., Iyer V. R., Dewight W., Cupo R. R., Debattisti V., Gomez L., et al. (2017). Increased mitochondrial nanotunneling activity, induced by calcium imbalance, affects intermitochondrial matrix exchanges. Proc. Natl. Acad. Sci. U. S. A. 114, E849–E858. doi:10.1073/pnas.1617788113
Leberer E., Pette D. (1986). Immunochemical quantification of sarcoplasmic reticulum Ca-ATPase, of calsequestrin and of parvalbumin in rabbit skeletal muscles of defined fiber composition. Eur. J. Biochem. 156, 489–496. doi:10.1111/j.1432-1033.1986.tb09607.x
Li A., Yi J., Li X., Zhou J. (2020). Physiological Ca(2+) transients versus pathological steady-state Ca(2+) elevation, who flips the ROS coin in skeletal muscle mitochondria. Front. Physiol. 11, 595800. doi:10.3389/fphys.2020.595800
Li H., Ding X., Lopez J. R., Takeshima H., Ma J., Allen P. D., et al. (2010). Impaired orai1-mediated resting Ca2+ entry reduces the cytosolic [Ca2+] and sarcoplasmic reticulum Ca2+ loading in quiescent junctophilin 1 knock-out myotubes. J. Biol. Chem. 285, 39171–39179. doi:10.1074/jbc.M110.149690
Liao Y., Erxleben C., Abramowitz J., Flockerzi V., Zhu M. X., Armstrong D. L., et al. (2008). Functional interactions among Orai1, TRPCs, and STIM1 suggest a STIM-regulated heteromeric Orai/TRPC model for SOCE/Icrac channels. Proc. Natl. Acad. Sci. U. S. A. 105, 2895–2900. doi:10.1073/pnas.0712288105
Liao Y., Erxleben C., Yildirim E., Abramowitz J., Armstrong D. L., Birnbaumer L. (2007). Orai proteins interact with TRPC channels and confer responsiveness to store depletion. Proc. Natl. Acad. Sci. U. S. A. 104, 4682–4687. doi:10.1073/pnas.0611692104
Limouze J., Straight A. F., Mitchison T., Sellers J. R. (2004). Specificity of blebbistatin, an inhibitor of myosin II. J. Muscle Res. Cell. Motil. 25, 337–341. doi:10.1007/s10974-004-6060-7
Liou J., Kim M. L., Do Heo W., Jones J. T., Myers J. W., Ferrell J. E., et al. (2005). STIM is a Ca2+ sensor essential for Ca2+-store-depletion-triggered Ca2+ influx. Curr. Biol. 15, 1235–1241. doi:10.1016/j.cub.2005.05.055
Liu S. X., Matthews H. R., Huang C. L.-H. (2021). Sarcoplasmic reticular Ca(2+)-ATPase inhibition paradoxically upregulates murine skeletal muscle Na(v)1.4 function. Sci. Rep. 11, 2846. doi:10.1038/s41598-021-82493-w
Ludtke S. J., Serysheva I. I., Hamilton S. L., Chiu W. (2005). The pore structure of the closed RyR1 channel. Structure 13, 1203–1211. doi:10.1016/j.str.2005.06.005
Luff A. R., Atwood H. L. (1972). Membrane properties and contraction of single muscle fibers in the mouse. Am. J. Physiol. 222, 1435–1440. doi:10.1152/ajplegacy.1972.222.6.1435
Lüttgau H. C., Gottschalk G., Berwe D. (1987). The effect of calcium and Ca antagonists upon excitation-contraction coupling. Can. J. Physiol. Pharmacol. 65, 717–723. doi:10.1139/y87-118
Lyfenko A. D., Dirksen R. T. (2008). Differential dependence of store-operated and excitation-coupled Ca2+ entry in skeletal muscle on STIM1 and Orai1. J. Physiol. 586, 4815–4824. doi:10.1113/jphysiol.2008.160481
Lyster D., Stephenson D. (1995). Contractile activation and measurements of intracellular Ca2+ concentration in cane toad twitch fibres in the presence of 2, 3-butanedione monoxime. Exp. Physiol. 80, 543–560. doi:10.1113/expphysiol.1995.sp003866
MacLennan D. H., Brandl C. J., Korczak B., Green N. M. (1985). Amino-acid sequence of a Ca2+ + Mg2+-dependent ATPase from rabbit muscle sarcoplasmic reticulum, deduced from its complementary DNA sequence. Nature 316, 696–700. doi:10.1038/316696a0
MacLennan D. H., Wong P. T. (1971). Isolation of a calcium-sequestering protein from sarcoplasmic reticulum. Proc. Natl. Acad. Sci. U. S. A. 68, 1231–1235. doi:10.1073/pnas.68.6.1231
Mallilankaraman K., Doonan P., Cárdenas C., Chandramoorthy H. C., Müller M., Miller R., et al. (2012). MICU1 is an essential gatekeeper for MCU-mediated mitochondrial Ca(2+) uptake that regulates cell survival. Cell. 151, 630–644. doi:10.1016/j.cell.2012.10.011
Mammucari C., Raffaello A., Vecellio Reane D., Rizzuto R. (2016). Molecular structure and pathophysiological roles of the mitochondrial calcium uniporter. Biochim. Biophys. Acta 1863, 2457–2464. doi:10.1016/j.bbamcr.2016.03.006
Mannella C. A., Buttle K., Rath B. K., Marko M. (1998). Electron microscopic tomography of rat-liver mitochondria and their interaction with the endoplasmic reticulum. Biofactors 8, 225–228. doi:10.1002/biof.5520080309
Marinello M., Cosette J., Bogni C., Denard J., Stockholm D., Buj-Bello A. (2021). Characterization of neuromuscular junctions in mice by combined confocal and super-resolution microscopy. J. Vis. Exp. 1–21. doi:10.3791/63032
Marsh B. (1951). A factor modifying muscle fibre synaeresis. Nature 167, 1065–1066. doi:10.1038/1671065a0
Martínez M., García M. C., Farías J. M., Cruzblanca H., Sánchez J. A. (1996). Modulation of Ca2+ channels, charge movement and Ca2+ transients by heparin in frog skeletal muscle fibres. J. Muscle Res. Cell. Motil. 17, 575–594. doi:10.1007/BF00124356
Marty I. (2015). Triadin regulation of the ryanodine receptor complex. J. Physiol. 593, 3261–3266. doi:10.1113/jphysiol.2014.281147
Mashima H., Matsumura M. (1962). Roles of external ions in the excitation-contraction coupling of frog skeletal muscle. Jpn. J. Physiol. 12, 639–653. doi:10.2170/jjphysiol.12.639
Massa R., Marliera L. N., Martorana A., Cicconi S., Pierucci D., Giacomini P., et al. (2000). Intracellular localization and isoform expression of the voltage-dependent anion channel (VDAC) in normal and dystrophic skeletal muscle. J. Muscle Res. Cell. Motil. 21, 433–442. doi:10.1023/a:1005688901635
Matlib M. A., Zhou Z., Knight S., Ahmed S., Choi K. M., Krause-Bauer J., et al. (1998). Oxygen-bridged dinuclear ruthenium amine complex specifically inhibits Ca2+ uptake into mitochondria in vitro and in situ in single cardiac myocytes. J. Biol. Chem. 273, 10223–10231. doi:10.1074/jbc.273.17.10223
Mckillop D. F. A., Fortune N. S., Ranatunga K. W., Geeves M. A. (1994). The influence of 2, 3-butanedione 2-monoxime (BDM) on the interaction between actin and myosin in solution and in skinned muscle fibres. J. Muscle Res. Cell. Motil. 15, 309–318. doi:10.1007/BF00123483
Mei Y., Xu L., Mowrey D. D., Mendez Giraldez R., Wang Y., Pasek D. A., et al. (2015). Channel gating dependence on pore lining helix Glycine residues in skeletal muscle ryanodine receptor. J. Biol. Chem. 290, 17535–17545. doi:10.1074/jbc.M115.659672
Meissner G. (1986). Ryanodine activation and inhibition of the Ca2+ release channel of sarcoplasmic reticulum. J. Biol. Chem. 261, 6300–6306. doi:10.1016/s0021-9258(19)84563-5
Meissner G. (2017). The structural basis of ryanodine receptor ion channel function. J. Gen. Physiol. 149, 1065–1089. doi:10.1085/jgp.201711878
Meizoso-Huesca A., Launikonis B. S. (2021). The Orai1 inhibitor BTP2 has multiple effects on Ca2+ handling in skeletal muscle. J. Gen. Physiol. 153, e202012747. doi:10.1085/jgp.202012747
Mejía-Raigosa D., Milán A. F., Giraldo M. A., Calderón J. C. (2021). A new set of equations for the simplified calibration of fluorescence Ca(2+) transients in skeletal muscle fibers. J. Muscle Res. Cell. Motil. 42, 161–168. doi:10.1007/s10974-021-09597-8
Messina A., Reina S., Guarino F., De Pinto V. (2012). VDAC isoforms in mammals. Biochim. Biophys. Acta 1818, 1466–1476. doi:10.1016/j.bbamem.2011.10.005
Michelucci A., Boncompagni S., Pietrangelo L., García-Castañeda M., Takano T., Malik S., et al. (2019). Transverse tubule remodeling enhances Orai1-dependent Ca2+ entry in skeletal muscle. Elife 8, e47576. doi:10.7554/eLife.47576
Michelucci A., Boncompagni S., Pietrangelo L., Takano T., Protasi F., Dirksen R. T. (2020). Pre-assembled Ca2+ entry units and constitutively active Ca2+ entry in skeletal muscle of calsequestrin-1 knockout mice. J. Gen. Physiol. 152, e202012617. doi:10.1085/jgp.202012617
Michelucci A., García-Castañeda M., Boncompagni S., Dirksen R. T. (2018). Role of STIM1/ORAI1-mediated store-operated Ca2+ entry in skeletal muscle physiology and disease. Cell. Calcium 76, 101–115. doi:10.1016/j.ceca.2018.10.004
Milán A. F., Rincón O. A., Arango L. B., Reutovich A. A., Smith G. L., Giraldo M. A., et al. (2021). Calibration of mammalian skeletal muscle Ca2+ transients recorded with the fast Ca2+ dye Mag-Fluo-4. Biochim. Biophys. Acta. Gen. Subj. 1865, 129939. doi:10.1016/j.bbagen.2021.129939
Miller L., Greensmith D. J., Sankaranarayanan R., O’Neill S. C., Eisner D. A. (2015). The effect of 2, 5-di-(tert-butyl)-1, 4-benzohydroquinone (TBQ) on intracellular Ca2+ handling in rat ventricular myocytes. Cell. Calcium 58, 208–214. doi:10.1016/j.ceca.2015.05.002
Minta A., Kao J. P. Y., Tsien R. Y. (1989). Fluorescent indicators for cytosolic calcium based on rhodamine and fluorescein chromophoresfluorescein chromophores. J. Biol. Chem. 264, 8171–8178. doi:10.1016/s0021-9258(18)83165-9
Mishin A. S., Lukyanov K. A. (2019). Live-cell super-resolution fluorescence microscopy. Biochemistry. 84, S19–S31. doi:10.1134/S0006297919140025
Mould J., Dulhunty A. F. (1999). Effects of external cadmium ions on excitation-contraction coupling in rat soleus fibres. Pflugers Arch. 437, 197–203. doi:10.1007/s004240050769
Moulds R. F. W. (1977). A comparison of the effects of sodium thiocyanate and dantrolene sodium on a mammalian isolated skeletal muscle. Br. J. Pharmacol. 59, 129–133. doi:10.1111/j.1476-5381.1977.tb06986.x
Nagasaki K., Kasai M. (1983). Fast release of calcium from sarcoplasmic reticulum vesicles monitored by chlortetracycline fluorescence. J. Biochem. 94, 1101–1109. doi:10.1093/oxfordjournals.jbchem.a134453
Nakada T., Kashihara T., Komatsu M., Kojima K., Takeshita T., Yamada M. (2018). Physical interaction of junctophilin and the Ca(V)1.1 C terminus is crucial for skeletal muscle contraction. Proc. Natl. Acad. Sci. U. S. A. 115, 4507–4512. doi:10.1073/pnas.1716649115
Niedergerke R. (1955). Local muscular shortening by intracellularly applied calcium. J. Physiol. 128, 12P–13P.
Niu C.-F., Watanabe Y., Ono K., Iwamoto T., Yamashita K., Satoh H., et al. (2007). Characterization of SN-6, a novel Na+/Ca2+ exchange inhibitor in Guinea pig cardiac ventricular myocytes. Eur. J. Pharmacol. 573, 161–169. doi:10.1016/j.ejphar.2007.06.033
Oddoux S., Brocard J., Schweitzer A., Szentesi P., Giannesini B., Brocard J., et al. (2009). Triadin deletion induces impaired skeletal muscle function. J. Biol. Chem. 284, 34918–34929. doi:10.1074/jbc.M109.022442
Odermatt A., Becker S., Khanna V. K., Kurzydlowski K., Leisner E., Pette D., et al. (1998). Sarcolipin regulates the activity of SERCA1, the fast-twitch skeletal muscle sarcoplasmic reticulum Ca2+-ATPase. J. Biol. Chem. 273, 12360–12369. doi:10.1074/jbc.273.20.12360
Ogata T., Yamasaki Y. (1997). Ultra-high-resolution scanning electron microscopy of mitochondria and sarcoplasmic reticulum arrangement in human red, white, and intermediate muscle fibers. Anat. Rec. 248, 214–223. doi:10.1002/(SICI)1097-0185(199706)248:2<214::AID-AR8>3.0.CO;2-S
Ogawa H., Kurebayashi N., Yamazawa T., Murayama T. (2021). Regulatory mechanisms of ryanodine receptor/Ca(2+) release channel revealed by recent advancements in structural studies. J. Muscle Res. Cell. Motil. 42, 291–304. doi:10.1007/s10974-020-09575-6
Ogawa Y., Murayama T., Kurebayashi N. (1999). Comparison of properties of Ca2+ release channels between rabbit and frog skeletal muscles. Mol. Cell. Biochem. 190, 191–201. doi:10.1023/a:1006903401123
Oláh T., Fodor J., Ruzsnavszky O., Vincze J., Berbey C., Allard B., et al. (2011). Overexpression of transient receptor potential canonical type 1 (TRPC1) alters both store operated calcium entry and depolarization-evoked calcium signals in C2C12 cells. Cell. Calcium 49, 415–425. doi:10.1016/j.ceca.2011.03.012
Olivera J. F., Pizarro G. (2010). Two inhibitors of store operated Ca2+ entry suppress excitation contraction coupling in frog skeletal muscle. J. Muscle Res. Cell. Motil. 31, 127–139. doi:10.1007/s10974-010-9216-7
Ong H. L., Cheng K. T., Liu X., Bandyopadhyay B. C., Paria B. C., Soboloff J., et al. (2007). Dynamic assembly of TRPC1-STIM1-Orai1 ternary complex is involved in store-operated calcium influx. Evidence for similarities in store-operated and calcium release-activated calcium channel components. J. Biol. Chem. 282, 9105–9116. doi:10.1074/jbc.M608942200
Ong H. L., de Souza L. B., Ambudkar I. S. (2016). Role of TRPC channels in store-operated calcium entry. Adv. Exp. Med. Biol. 898, 87–109. doi:10.1007/978-3-319-26974-0_5
Oo Y. W., Gomez-Hurtado N., Walweel K., van Helden D. F., Imtiaz M. S., Knollmann B. C., et al. (2015). Essential role of calmodulin in RyR inhibition by dantrolene. Mol. Pharmacol. 88, 57–63. doi:10.1124/mol.115.097691
Oz M., Frank G. B. (1994). Effect of the calcium channel agonist Bay K8644 on mechanical and electrical responses of frog skeletal muscle. Can. J. Physiol. Pharmacol. 72, 1220–1225. doi:10.1139/y94-173
Palty R., Sekler I. (2012). The mitochondrial Na(+)/Ca(2+) exchanger. Cell. Calcium 52, 9–15. doi:10.1016/j.ceca.2012.02.010
Palty R., Silverman W. F., Hershfinkel M., Caporale T., Sensi S. L., Parnis J., et al. (2010). NCLX is an essential component of mitochondrial Na+/Ca2+ exchange. Proc. Natl. Acad. Sci. U. S. A. 107, 436–441. doi:10.1073/pnas.0908099107
Pan Z., Yang D., Nagaraj R. Y., Nosek T. A., Nishi M., Takeshima H., et al. (2002). Dysfunction of store-operated calcium channel in muscle cells lacking mg29. Nat. Cell. Biol. 4, 379–383. doi:10.1038/ncb788
Paolini C., Fessenden J. D., Pessah I. N., Franzini-Armstrong C. (2004). Evidence for conformational coupling between two calcium channels. Proc. Natl. Acad. Sci. U. S. A. 101, 12748–12752. doi:10.1073/pnas.0404836101
Park H., Park I. Y., Kim E., Youn B., Fields K., Dunker A. K., et al. (2004). Comparing skeletal and cardiac calsequestrin structures and their calcium binding: A proposed mechanism for coupled calcium binding and protein polymerization. J. Biol. Chem. 279, 18026–18033. doi:10.1074/jbc.M311553200
Park H., Wu S., Dunker A. K., Kang C. (2003). Polymerization of calsequestrin. Implications for Ca2+ regulation. J. Biol. Chem. 278, 16176–16182. doi:10.1074/jbc.M300120200
Pedersen T. H., L-H Huang C., Fraser J. A. (2011). An analysis of the relationships between subthreshold electrical properties and excitability in skeletal muscle. J. Gen. Physiol. 138, 73–93. doi:10.1085/jgp.201010510
Periasamy M., Kalyanasundaram A. (2007). SERCA pump isoforms: Their role in calcium transport and disease. Muscle Nerve 35, 430–442. doi:10.1002/mus.20745
Permyakov E. A., Uversky V. N. (2022). What is parvalbumin for? Biomolecules 12, 656. doi:10.3390/biom12050656
Perni S., Close M., Franzini-Armstrong C. (2013). Novel details of calsequestrin gel conformation in situ. J. Biol. Chem. 288, 31358–31362. doi:10.1074/jbc.M113.507749
Perni S. (2022). The builders of the junction: Roles of Junctophilin1 and Junctophilin2 in the assembly of the sarcoplasmic reticulum-plasma membrane junctions in striated muscle. Biomolecules 12, 109. doi:10.3390/biom12010109
Perocchi F., Gohil V. M., Girgis H. S., Bao X. R., McCombs J. E., Palmer A. E., et al. (2010). MICU1 encodes a mitochondrial EF hand protein required for Ca(2+) uptake. Nature 467, 291–296. doi:10.1038/nature09358
Peter J. B., Barnard R. J., Edgerton V. R., Gillespie C. A., Stempel K. E. (1972). Metabolic profiles of three fiber types of skeletal muscle in Guinea pigs and rabbits. Biochemistry 11, 2627–2633. doi:10.1021/bi00764a013
Petronilli V., Miotto G., Canton M., Brini M., Colonna R., Bernardi P., et al. (1999). Transient and long-lasting openings of the mitochondrial permeability transition pore can be monitored directly in intact cells by changes in mitochondrial calcein fluorescence. Biophys. J. 76, 725–734. doi:10.1016/S0006-3495(99)77239-5
Phimister A. J., Lango J., Lee E. H., Ernst-Russell M. A., Takeshima H., Ma J., et al. (2007). Conformation-dependent stability of junctophilin 1 (JP1) and ryanodine receptor type 1 (RyR1) channel complex is mediated by their hyper-reactive thiols. J. Biol. Chem. 282, 8667–8677. doi:10.1074/jbc.M609936200
Picard M., McManus M. J., Csordás G., Várnai P., Dorn G. W., Williams D., et al. (2015). Trans-mitochondrial coordination of cristae at regulated membrane junctions. Nat. Commun. 6, 6259. doi:10.1038/ncomms7259
Picard M. (2015). Mitochondrial synapses: Intracellular communication and signal integration. Trends Neurosci. 38, 468–474. doi:10.1016/j.tins.2015.06.001
Pietrangelo L., D’Incecco A., Ainbinder A., Michelucci A., Kern H., Dirksen R. T., et al. (2015). Age-dependent uncoupling of mitochondria from Ca2+ release units in skeletal muscle. Oncotarget 6, 35358–35371. doi:10.18632/oncotarget.6139
Plovanich M., Bogorad R. L., Sancak Y., Kamer K. J., Strittmatter L., Li A. A., et al. (2013). MICU2, a paralog of MICU1, resides within the mitochondrial uniporter complex to regulate calcium handling. PLoS One 8, e55785. doi:10.1371/journal.pone.0055785
Porter K. R., Palade G. E. (1957). Studies on the endoplasmic reticulum. III. Its form and distribution in striated muscle cells. J. Biophys. Biochem. Cytol. 3, 269–300. doi:10.1083/jcb.3.2.269
Prakriya M., Feske S., Gwack Y., Srikanth S., Rao A., Hogan P. G. (2006). Orai1 is an essential pore subunit of the CRAC channel. Nature 443, 230–233. doi:10.1038/nature05122
Prakriya M., Lewis R. S. (2015). Store-operated calcium channels. Physiol. Rev. 95, 1383–1436. doi:10.1152/physrev.00020.2014
Prosser B. L., Wright N. T., Hernãndez-Ochoa E. O., Varney K. M., Liu Y., Olojo R. O., et al. (2008). S100A1 binds to the calmodulin-binding site of ryanodine receptor and modulates skeletal muscle excitation-contraction coupling. J. Biol. Chem. 283, 5046–5057. doi:10.1074/jbc.M709231200
Putney J. W. (1986). A model for receptor-regulated calcium entry. Cell. Calcium 7, 1–12. doi:10.1016/0143-4160(86)90026-6
Putney J. W. (2010). Pharmacology of store-operated calcium channels. Mol. Interv. 10, 209–218. doi:10.1124/mi.10.4.4
Putney J. W. (2017). Store-operated calcium entry: An historical overview. Adv. Exp. Med. Biol. 981, 205–214. doi:10.1007/978-3-319-55858-5_9
Raffaello A., De Stefani D., Sabbadin D., Teardo E., Merli G., Picard A., et al. (2013). The mitochondrial calcium uniporter is a multimer that can include a dominant-negative pore-forming subunit. EMBO J. 32, 2362–2376. doi:10.1038/emboj.2013.157
Rahman F. A., Quadrilatero J. (2021). Mitochondrial network remodeling: An important feature of myogenesis and skeletal muscle regeneration. Cell. Mol. Life Sci. 78, 4653–4675. doi:10.1007/s00018-021-03807-9
Raju B., Murphy E., Levy L. A., Hall R. D., London R. E. (1989). A fluorescent indicator for measuring cytosolic free magnesium. Am. J. Physiol. 256, C540–C548. doi:10.1152/ajpcell.1989.256.3.C540
Ranvier L. (1873). Propriétés et structures différentes des muscles rouges et des muscles blancs, chez les Lapins et chez les Raies. Compt Rendus 77, 1030–1034.
Rao L., Qian Y., Khodabukus A., Ribar T., Bursac N. (2018). Engineering human pluripotent stem cells into a functional skeletal muscle tissue. Nat. Commun. 9, 126. doi:10.1038/s41467-017-02636-4
Rasul F., Zheng F., Dong F., He J., Liu L., Liu W., et al. (2021). Emr1 regulates the number of foci of the endoplasmic reticulum-mitochondria encounter structure complex. Nat. Commun. 12, 521. doi:10.1038/s41467-020-20866-x
Rathod N., Bak J. J., Primeau J. O., Fisher M. E., Espinoza-Fonseca L. M., Lemieux M. J., et al. (2021). Nothing regular about the regulins: Distinct functional properties of SERCA transmembrane peptide regulatory subunits. Int. J. Mol. Sci. 22, 8891. doi:10.3390/ijms22168891
Reddish F. N., Miller C. L., Deng X., Dong B., Patel A. A., Ghane M. A., et al. (2021). Rapid subcellular calcium responses and dynamics by calcium sensor G-CatchER. iScience 24, 102129. doi:10.1016/j.isci.2021.102129
Reggiani C. (2021). Caffeine as a tool to investigate sarcoplasmic reticulum and intracellular calcium dynamics in human skeletal muscles. J. Muscle Res. Cell. Motil. 42, 281–289. doi:10.1007/s10974-020-09574-7
Ridgway E. B., Ashley C. C. (1967). Calcium transients in single muscle fibers. Biochem. Biophys. Res. Commun. 29, 229–234. doi:10.1016/0006-291x(67)90592-x
Rincón O. A., Milán A. F., Calderón J. C., Giraldo M. A. (2021). Comprehensive simulation of Ca2+ transients in the continuum of mouse skeletal muscle fiber types. Int. J. Mol. Sci. 22, 12378. doi:10.3390/ijms222212378
Rios E., Brum G. (1987). Involvement of dihydropyridine receptors in excitation-contraction coupling in skeletal muscle. Nature 325, 717–720. doi:10.1038/325717a0
Ríos E., Karhanek M., Ma J., González A., Gonzalez A. (1993). An allosteric model of the molecular interactions of excitation-contraction coupling in skeletal muscle. J. Gen. Physiol. 102, 449–481. doi:10.1085/jgp.102.3.449
Roman B. I., Verhasselt S., Stevens C. V. (2018). Medicinal chemistry and use of myosin II inhibitor ( S )-Blebbistatin and its derivatives. J. Med. Chem. 61, 9410–9428. doi:10.1021/acs.jmedchem.8b00503
Roos J., DiGregorio P. J., Yeromin A. V., Ohlsen K., Lioudyno M., Zhang S., et al. (2005). STIM1, an essential and conserved component of store-operated Ca2+ channel function. J. Cell. Biol. 169, 435–445. doi:10.1083/jcb.200502019
Rossi A. E., Boncompagni S., Dirksen R. T. (2009). Sarcoplasmic reticulum-mitochondrial symbiosis: Bidirectional signaling in skeletal muscle. Exerc. Sport Sci. Rev. 37, 29–35. doi:10.1097/JES.0b013e3181911fa4
Royer L., Ríos E. (2009). Deconstructing calsequestrin. Complex buffering in the calcium store of skeletal muscle. J. Physiol. 587, 3101–3111. doi:10.1113/jphysiol.2009.171934
Rudolf R., Mongillo M., Magalhães P. J., Pozzan T. (2004). In vivo monitoring of Ca(2+) uptake into mitochondria of mouse skeletal muscle during contraction. J. Cell. Biol. 166, 527–536. doi:10.1083/jcb.200403102
Rufenach B., Van Petegem F. (2021). Structure and function of STAC proteins: Calcium channel modulators and critical components of muscle excitation-contraction coupling. J. Biol. Chem. 297, 100874. doi:10.1016/j.jbc.2021.100874
Ryu S.-Y., Beutner G., Kinnally K. W., Dirksen R. T., Sheu S.-S. (2011). Single channel characterization of the mitochondrial ryanodine receptor in heart mitoplasts. J. Biol. Chem. 286, 21324–21329. doi:10.1074/jbc.C111.245597
Saito A., Inui M., Radermacher M., Frank J., Fleischer S. (1988). Ultrastructure of the calcium release channel of sarcoplasmic reticulum. J. Cell. Biol. 107, 211–219. doi:10.1083/jcb.107.1.211
Sampieri A., Diaz-Muñoz M., Antaramian A., Vaca L. (2005). The foot structure from the type 1 ryanodine receptor is required for functional coupling to store-operated channels. J. Biol. Chem. 280, 24804–24815. doi:10.1074/jbc.M501487200
Samsó M., Feng W., Pessah I. N., Allen P. D. (2009). Coordinated movement of cytoplasmic and transmembrane domains of RyR1 upon gating. PLoS Biol. 7, e85. doi:10.1371/journal.pbio.1000085
Samsó M., Wagenknecht T., Allen P. D. (2005). Internal structure and visualization of transmembrane domains of the RyR1 calcium release channel by cryo-EM. Nat. Struct. Mol. Biol. 12, 539–544. doi:10.1038/nsmb938
Sancak Y., Markhard A. L., Kitami T., Kovács-Bogdán E., Kamer K. J., Udeshi N. D., et al. (2013). EMRE is an essential component of the mitochondrial calcium uniporter complex. Science 342, 1379–1382. doi:10.1126/science.1242993
Sanchez E. J., Lewis K. M., Danna B. R., Kang C. (2012). High-capacity Ca2+ binding of human skeletal calsequestrin. J. Biol. Chem. 287, 11592–11601. doi:10.1074/jbc.M111.335075
Sandow A. (1952). Excitation-contraction coupling in muscular response. Yale J. Biol. Med. 25, 176–201.
Santo-Domingo J., Vay L., Hernández-SanMiguel E., Lobatón C. D., Moreno A., Montero M., et al. (2007). The plasma membrane Na + /Ca 2+ exchange inhibitor KB-R7943 is also a potent inhibitor of the mitochondrial Ca 2+ uniporter. Br. J. Pharmacol. 151, 647–654. doi:10.1038/sj.bjp.0707260
Sarbjit-Singh S. S., Matthews H. R., Huang C. L.-H. (2020). Ryanodine receptor modulation by caffeine challenge modifies Na(+) current properties in intact murine skeletal muscle fibres. Sci. Rep. 10, 2199. doi:10.1038/s41598-020-59196-9
Sárközi S., Szentesi P., Jona I., Csernoch L. (1996). Effects of cardiac glycosides on excitation-contraction coupling in frog skeletal muscle fibres. J. Physiol. 495, 611–626. doi:10.1113/jphysiol.1996.sp021620
Saüc S., Frieden M. (2017). Neurological and motor disorders: TRPC in the skeletal muscle. Adv. Exp. Med. Biol. 993, 557–575. doi:10.1007/978-3-319-57732-6_28
Savalli N., Angelini M., Steccanella F., Wier J., Wu F., Quinonez M., et al. (2021). The distinct role of the four voltage sensors of the skeletal CaV1.1 channel in voltage-dependent activation. J. Gen. Physiol. 153, e202112915. doi:10.1085/jgp.202112915
Schiaffino S., Reggiani C. (2011). Fiber types in mammalian skeletal muscles. Physiol. Rev. 91, 1447–1531. doi:10.1152/physrev.00031.2010
Schneider M. F., Chandler W. K. (1973). Voltage dependent charge movement of skeletal muscle: A possible step in excitation-contraction coupling. Nature 242, 244–246. doi:10.1038/242244a0
Seidler N. W., Jona I., Vegh M., Martonosi A. (1989). Cyclopiazonic acid is a specific inhibitor of the Ca2+-ATPase of sarcoplasmic reticulum. J. Biol. Chem. 264, 17816–17823. doi:10.1016/s0021-9258(19)84646-x
Sembrowich W. L., Quintinskie J. J., Li G. (1985). Calcium uptake in mitochondria from different skeletal muscle types. J. Appl. Physiol. 59, 137–141. doi:10.1152/jappl.1985.59.1.137
Shannon T. R., Ginsburg K. S., Bers D. M. (2000). Potentiation of fractional sarcoplasmic reticulum calcium release by total and free intra-sarcoplasmic reticulum calcium concentration. Biophys. J. 78, 334–343. doi:10.1016/S0006-3495(00)76596-9
Shin D. W., Pan Z., Kim E. K., Lee J. M., Bhat M. B., Parness J., et al. (2003). A retrograde signal from calsequestrin for the regulation of store-operated Ca2+ entry in skeletal muscle. J. Biol. Chem. 278, 3286–3292. doi:10.1074/jbc.M209045200
Shishmarev D., Rowland E., Aditya S., Sundararaj S., Oakley A. J., Dulhunty A. F., et al. (2022). Molecular interactions of STAC proteins with skeletal muscle dihydropyridine receptor and excitation-contraction coupling. Protein Sci. 31, e4311. doi:10.1002/pro.4311
Shkryl V. M., Shirokova N. (2006). Transfer and tunneling of Ca2+ from sarcoplasmic reticulum to mitochondria in skeletal muscle. J. Biol. Chem. 281, 1547–1554. doi:10.1074/jbc.M505024200
Shoshan-Barmatz V., De Pinto V., Zweckstetter M., Raviv Z., Keinan N., Arbel N. (2010). VDAC, a multi-functional mitochondrial protein regulating cell life and death. Mol. Asp. Med. 31, 227–285. doi:10.1016/j.mam.2010.03.002
Shoshan-Barmatz V., De S. (2017). Mitochondrial VDAC, the Na(+)/Ca(2+) exchanger, and the Ca(2+) uniporter in Ca(2+) dynamics and signaling. Adv. Exp. Med. Biol. 981, 323–347. doi:10.1007/978-3-319-55858-5_13
Shoshan-Barmatz V., Krelin Y., Shteinfer-Kuzmine A. (2018). VDAC1 functions in Ca(2+) homeostasis and cell life and death in health and disease. Cell. Calcium 69, 81–100. doi:10.1016/j.ceca.2017.06.007
Shtifman A., Ward C. W., Wang J., Valdivia H. H., Schneider M. F. (2000). Effects of imperatoxin A on local sarcoplasmic reticulum Ca2+ release in frog skeletal muscle. Biophys. J. 79, 814–827. doi:10.1016/S0006-3495(00)76338-7
Siekevitz P. (1957). Powerhouse of the cell. Sci. Am. 197, 131–144. doi:10.1038/scientificamerican0757-131
Skoglund G., Lainé J., Darabi R., Fournier E., Perlingeiro R., Tabti N. (2014). Physiological and ultrastructural features of human induced pluripotent and embryonic stem cell-derived skeletal myocytes in vitro. Proc. Natl. Acad. Sci. U. S. A. 111, 8275–8280. doi:10.1073/pnas.1322258111
Soboloff J., Spassova M. A., Tang X. D., Hewavitharana T., Xu W., Gill D. L. (2006). Orai1 and STIM reconstitute store-operated calcium channel function. J. Biol. Chem. 281, 20661–20665. doi:10.1074/jbc.C600126200
Song L. S., Sham J. S., Stern M. D., Lakatta E. G., Cheng H. (1998). Direct measurement of SR release flux by tracking “Ca2+ spikes” in rat cardiac myocytes. J. Physiol. 512, 677–691. doi:10.1111/j.1469-7793.1998.677bd.x
Sparagna G. C., Gunter K. K., Sheu S. S., Gunter T. E. (1995). Mitochondrial calcium uptake from physiological-type pulses of calcium. A description of the rapid uptake mode. J. Biol. Chem. 270, 27510–27515. doi:10.1074/jbc.270.46.27510
Stathopulos P. B., Li G.-Y., Plevin M. J., Ames J. B., Ikura M. (2006). Stored Ca2+ depletion-induced oligomerization of stromal interaction molecule 1 (STIM1) via the EF-SAM region: An initiation mechanism for capacitive Ca2+ entry. J. Biol. Chem. 281, 35855–35862. doi:10.1074/jbc.M608247200
Sten-Knudsen O. (1954). The ineffectiveness of the `window field’ in the initiation of muscle contraction. J. Physiol. 125, 396–404. doi:10.1113/jphysiol.1954.sp005167
Stiber J., Hawkins A., Zhang Z.-S., Wang S., Burch J., Graham V., et al. (2008). STIM1 signalling controls store-operated calcium entry required for development and contractile function in skeletal muscle. Nat. Cell. Biol. 10, 688–697. doi:10.1038/ncb1731
Sun M., Huang J., Bunyak F., Gumpper K., De G., Sermersheim M., et al. (2014). Superresolution microscope image reconstruction by spatiotemporal object decomposition and association: Application in resolving t-tubule structure in skeletal muscle. Opt. Express 22, 12160–12176. doi:10.1364/OE.22.012160
Szabadkai G., Duchen M. R. (2008). Mitochondria: The hub of cellular Ca2+ signaling. Physiology 23, 84–94. doi:10.1152/physiol.00046.2007
Szentesi P., Jacquemond V., Kovács L., Csernoch L. (1997). Intramembrane charge movement and sarcoplasmic calcium release in enzymatically isolated mammalian skeletal muscle fibres. J. Physiol. 505, 371–384. doi:10.1111/j.1469-7793.1997.371bb.x
Sztretye M., Geyer N., Vincze J., Al-Gaadi D., Oláh T., Szentesi P., et al. (2017). SOCE is important for maintaining sarcoplasmic calcium content and release in skeletal muscle fibers. Biophys. J. 113, 2496–2507. doi:10.1016/j.bpj.2017.09.023
Takahashi K., Tanabe K., Ohnuki M., Narita M., Ichisaka T., Tomoda K., et al. (2007). Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell. 131, 861–872. doi:10.1016/j.cell.2007.11.019
Takahashi K., Yamanaka S. (2006). Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell. 126, 663–676. doi:10.1016/j.cell.2006.07.024
Tanabe T., Beam K. G., Adams B. A., Niidome T., Numa S. (1990). Regions of the skeletal muscle dihydropyridine receptor critical for excitation–contraction coupling. Nature 346, 567–569. doi:10.1038/346567a0
Taylor S. R., Preiser H., Sandow A. (1972). Action potential parameters affecting excitation-contraction coupling. J. Gen. Physiol. 59, 421–436. doi:10.1085/jgp.59.4.421
Thomas D., Tovey S. C., Collins T. J., Bootman M. D., Berridge M. J., Lipp P. (2000). A comparison of fluorescent Ca2+ indicator properties and their use in measuring elementary and global Ca2+ signals. Cell. Calcium 28, 213–223. doi:10.1054/ceca.2000.0152
Toyoshima C., Mizutani T. (2004). Crystal structure of the calcium pump with a bound ATP analogue. Nature 430, 529–535. doi:10.1038/nature02680
Tripathy A., Resch W., Xu L., Valdivia H. H., Meissner G. (1998). Imperatoxin A induces subconductance states in Ca2+ release channels (ryanodine receptors) of cardiac and skeletal muscle. J. Gen. Physiol. 111, 679–690. doi:10.1085/jgp.111.5.679
Tsai M.-F., Jiang D., Zhao L., Clapham D., Miller C. (2014). Functional reconstitution of the mitochondrial Ca2+/H+ antiporter Letm1. J. Gen. Physiol. 143, 67–73. doi:10.1085/jgp.201311096
Tsien R. Y. (1980). New calcium indicators and buffers with high selectivity against magnesium and protons: Design, synthesis, and properties of prototype structures. Biochemistry 19, 2396–2404. doi:10.1021/bi00552a018
Tsien R. Y., Pozzan T., Rink T. J. (1982). Calcium homeostasis in intact lymphocytes: Cytoplasmic free calcium monitored with a new, intracellularly trapped fluorescent indicator. J. Cell. Biol. 94, 325–334. doi:10.1083/jcb.94.2.325
Vais H., Mallilankaraman K., Mak D.-O. D., Hoff H., Payne R., Tanis J. E., et al. (2016). EMRE is a matrix Ca(2+) sensor that governs gatekeeping of the mitochondrial Ca(2+) uniporter. Cell. Rep. 14, 403–410. doi:10.1016/j.celrep.2015.12.054
van Lunteren E., Moyer M., Dick T. E. (2001). Modulation of diaphragm action potentials by K+ channel blockers. Respir. Physiol. 124, 217–230. doi:10.1016/S0034-5687(00)00198-5
van Oort R. J., Garbino A., Wang W., Dixit S. S., Landstrom A. P., Gaur N., et al. (2011). Disrupted junctional membrane complexes and hyperactive ryanodine receptors after acute junctophilin knockdown in mice. Circulation 123, 979–988. doi:10.1161/CIRCULATIONAHA.110.006437
Vandebrouck C., Martin D., Schoor M. C.-V., Debaix H., Gailly P. (2002). Involvement of TRPC in the abnormal calcium influx observed in dystrophic (mdx) mouse skeletal muscle fibers. J. Cell. Biol. 158, 1089–1096. doi:10.1083/jcb.200203091
Vecellio Reane D., Vallese F., Checchetto V., Acquasaliente L., Butera G., De Filippis V., et al. (2016). A MICU1 splice variant confers high sensitivity to the mitochondrial Ca(2+) uptake machinery of skeletal muscle. Mol. Cell. 64, 760–773. doi:10.1016/j.molcel.2016.10.001
Vendelin M., Béraud N., Guerrero K., Andrienko T., Kuznetsov A. V., Olivares J., et al. (2005). Mitochondrial regular arrangement in muscle cells: A “crystal-like” pattern. Am. J. Physiol. Cell. Physiol. 288, C757–C767. doi:10.1152/ajpcell.00281.2004
Vig M., Beck A., Billingsley J. M., Lis A., Parvez S., Peinelt C., et al. (2006a). CRACM1 multimers form the ion-selective pore of the CRAC channel. Curr. Biol. 16, 2073–2079. doi:10.1016/j.cub.2006.08.085
Vig M., DeHaven W. I., Bird G. S., Billingsley J. M., Wang H., Rao P. E., et al. (2008). Defective mast cell effector functions in mice lacking the CRACM1 pore subunit of store-operated calcium release–activated calcium channels. Nat. Immunol. 9, 89–96. doi:10.1038/ni1550
Vig M., Peinelt C., Beck A., Koomoa D. L., Rabah D., Koblan-Huberson M., et al. (2006b). CRACM1 is a plasma membrane protein essential for store-operated Ca2+ entry. Science 312, 1220–1223. doi:10.1126/science.1127883
Vincent A. E., Ng Y. S., White K., Davey T., Mannella C., Falkous G., et al. (2016). The spectrum of mitochondrial ultrastructural defects in mitochondrial myopathy. Sci. Rep. 6, 30610. doi:10.1038/srep30610
Vincent A. E., Turnbull D. M., Eisner V., Hajnóczky G., Picard M. (2017). Mitochondrial nanotunnels. Trends Cell. Biol. 27, 787–799. doi:10.1016/j.tcb.2017.08.009
Vincent A. E., White K., Davey T., Philips J., Ogden R. T., Lawless C., et al. (2019). Quantitative 3D mapping of the human skeletal muscle mitochondrial network. Cell. Rep. 27, 321. doi:10.1016/j.celrep.2019.03.051
Wagenknecht T., Grassucci R., Frank J., Saito A., Inui M., Fleischer S. (1989). Three-dimensional architecture of the calcium channel/foot structure of sarcoplasmic reticulum. Nature 338, 167–170. doi:10.1038/338167a0
Walsh K. B., Bryant S. H., Schwartz A. (1988). Action of diltiazem on excitation-contraction coupling in bullfrog skeletal muscle fibers. J. Pharmacol. Exp. Ther. 245, 531–536.
Walsh K. B., Bryant S. H., Schwartz A. (1986). Effect of calcium antagonist drugs on calcium currents in mammalian skeletal muscle fibers. J. Pharmacol. Exp. Ther. 236, 403–407.
Wang Q., Michalak M. (2020). Calsequestrin. Structure, function, and evolution. Cell. Calcium 90, 102242. doi:10.1016/j.ceca.2020.102242
Wang S., Trumble W. R., Liao H., Wesson C. R., Dunker A. K., Kang C. H. (1998). Crystal structure of calsequestrin from rabbit skeletal muscle sarcoplasmic reticulum. Nat. Struct. Biol. 5, 476–483. doi:10.1038/nsb0698-476
Wang X., Nawaz M., DuPont C., Myers J. H., Burke S. R., Bannister R. A., et al. (2022). The role of action potential changes in depolarization-induced failure of excitation contraction coupling in mouse skeletal muscle. Elife 11, e71588. doi:10.7554/eLife.71588
Weber A. (1959). On the role of calcium in the activity of adenosine 5’-triphosphate hydrolysis by actomyosin. J. Biol. Chem. 234, 2764–2769. doi:10.1016/s0021-9258(18)69777-7
Weber H., Portzehl H. (1954). The transference of the muscle energy in the contraction cycle. Prog. Biophys. Biophys. Chem. 4, 60–111. doi:10.1016/S0096-4174(18)30062-3
Wei L., Gallant E. M., Dulhunty A. F., Beard N. A. (2009). Junctin and triadin each activate skeletal ryanodine receptors but junctin alone mediates functional interactions with calsequestrin. Int. J. Biochem. Cell. Biol. 41, 2214–2224. doi:10.1016/j.biocel.2009.04.017
Wei L., Varsányi M., Dulhunty A. F., Beard N. A. (2006). The conformation of calsequestrin determines its ability to regulate skeletal ryanodine receptors. Biophys. J. 91, 1288–1301. doi:10.1529/biophysj.106.082610
Wei M., Zhou Y., Sun A., Ma G., He L., Zhou L., et al. (2016). Molecular mechanisms underlying inhibition of STIM1-Orai1-mediated Ca(2+) entry induced by 2-aminoethoxydiphenyl borate. Pflugers Arch. 468, 2061–2074. doi:10.1007/s00424-016-1880-z
Wei-LaPierre L., Carrell E. M., Boncompagni S., Protasi F., Dirksen R. T. (2013). Orai1-dependent calcium entry promotes skeletal muscle growth and limits fatigue. Nat. Commun. 4, 2805. doi:10.1038/ncomms3805
Wei-LaPierre L., Groom L., Dirksen R. T. (2022). Acute exposure to extracellular BTP2 does not inhibit Ca2+ release during EC coupling in intact skeletal muscle fibers. J. Gen. Physiol. 154, e202112976. doi:10.1085/jgp.202112976
Weigl L. G., Hohenegger M., Kress H. G. (2000). Dihydropyridine‐induced Ca 2+ release from ryanodine‐sensitive Ca 2+ pools in human skeletal muscle cells. J. Physiol. 525, 461–469. doi:10.1111/j.1469-7793.2000.t01-1-00461.x
Westerblad H., Allen D. G. (1991). Changes of myoplasmic calcium concentration during fatigue in single mouse muscle fibers. J. Gen. Physiol. 98, 615–635. doi:10.1085/jgp.98.3.615
Westerblad H., Allen D. G. (1992). Myoplasmic free Mg2+ concentration during repetitive stimulation of single fibres from mouse skeletal muscle. J. Physiol. 453, 413–434. doi:10.1113/jphysiol.1992.sp019236
Westerblad H., Allen D. G. (1994). The role of sarcoplasmic reticulum in relaxation of mouse muscle; effects of 2, 5-di(tert-butyl)-1, 4-benzohydroquinone. J. Physiol. 474, 291–301. doi:10.1113/jphysiol.1994.sp020022
Westerblad H., Andrade F. H., Islam M. S. (1998). Effects of ryanodine receptor agonist 4-chloro-m-cresol on myoplasmic free Ca2+ concentration and force of contraction in mouse skeletal muscle. Cell. Calcium 24, 105–115. doi:10.1016/S0143-4160(98)90078-1
Wictome M., Henderson I., Lee A. G., East J. M. (1992a). Mechanism of inhibition of the calcium pump of sarcoplasmic reticulum by thapsigargin. Biochem. J. 283, 525–529. doi:10.1042/bj2830525
Wictome M., Michelangeli F., Lee A. G., East J. (1992b). The inhibitors thapsigargin and 2, 5-di(tert-butyl)-1, 4-benzohydroquinone favour the E2 form of the Ca 2+ , Mg 2+ -ATPase. FEBS Lett. 304, 109–113. doi:10.1016/0014-5793(92)80599-C
Williams D. A., Head S. I., Bakker A. J., Stephenson D. G. (1990). Resting calcium concentrations in isolated skeletal muscle fibres of dystrophic mice. J. Physiol. 428, 243–256. doi:10.1113/jphysiol.1990.sp018210
Williams J. H., Ward C. W. (1991). Effects of the calcium channel agonist, Bay K 8644, on the mechanical output of skeletal muscle fibers. Gen. Pharmacol. 22, 735–740. doi:10.1016/0306-3623(91)90088-N
Winegrad S. (1968). Intracellular calcium movements of frog skeletal muscle during recovery from tetanus. J. Gen. Physiol. 51, 65–83. doi:10.1085/jgp.51.1.65
Winegrad S. (1970). The intracellular site of calcium activaton of contraction in frog skeletal muscle. J. Gen. Physiol. 55, 77–88. doi:10.1085/jgp.55.1.77
Winegrad S. (1965). The location of muscle calcium with respect to the myofibrils. J. Gen. Physiol. 48, 997–1002. doi:10.1085/jgp.48.6.997
Wium E., Dulhunty A. F., Beard N. A. (2012). A skeletal muscle ryanodine receptor interaction domain in triadin. PLoS One 7, e43817. doi:10.1371/journal.pone.0043817
Wokosin D. L., Loughrey C. M., Smith G. L. (2004). Characterization of a range of fura dyes with two-photon excitation. Biophys. J. 86, 1726–1738. doi:10.1016/S0006-3495(04)74241-1
Woll K. A., Van Petegem F. (2022). Calcium-release channels: Structure and function of IP(3) receptors and ryanodine receptors. Physiol. Rev. 102, 209–268. doi:10.1152/physrev.00033.2020
Woods C. E., Novo D., DiFranco M., Vergara J. L. (2004). The action potential-evoked sarcoplasmic reticulum calcium release is impaired in mdx mouse muscle fibres. J. Physiol. 557, 59–75. doi:10.1113/jphysiol.2004.061291
Wu J., Yan Z., Li Z., Qian X., Lu S., Dong M., et al. (2016). Structure of the voltage-gated calcium channel Cav1.1 at 3.6 Å resolution. Nature 537, 191–196. doi:10.1038/nature19321
Xu L., Jones R., Meissner G. (1993). Effects of local anesthetics on single channel behavior of skeletal muscle calcium release channel. J. Gen. Physiol. 101, 207–233. doi:10.1085/jgp.101.2.207
Xu L., Tripathy A., Pasek D. A., Meissner G. (1998). Potential for pharmacology of ryanodine receptor/calcium release channels. Ann. N. Y. Acad. Sci. 853, 130–148. doi:10.1111/j.1749-6632.1998.tb08262.x
Yan Z., Bai X., Yan C., Wu J., Li Z., Xie T., et al. (2015). Structure of the rabbit ryanodine receptor RyR1 at near-atomic resolution. Nature 517, 50–55. doi:10.1038/nature14063
Yarotskyy V., Dirksen R. T. (2012). Temperature and RyR1 regulate the activation rate of store-operated Ca²+ entry current in myotubes. Biophys. J. 103, 202–211. doi:10.1016/j.bpj.2012.06.001
Yi J., Ma C., Li Y., Weisleder N., Ríos E., Ma J., et al. (2011). Mitochondrial calcium uptake regulates rapid calcium transients in skeletal muscle during excitation-contraction (E-C) coupling. J. Biol. Chem. 286, 32436–32443. doi:10.1074/jbc.M110.217711
York A. L., Zheng J. Q. (2017). Super-resolution microscopy reveals a nanoscale organization of acetylcholine receptors for trans-synaptic alignment at neuromuscular synapses. eNeuro 4, ENEURO.0232–17.2017. doi:10.1523/ENEURO.0232-17.2017
Zalk R., Clarke O. B., des Georges A., Grassucci R. A., Reiken S., Mancia F., et al. (2015). Structure of a mammalian ryanodine receptor. Nature 517, 44–49. doi:10.1038/nature13950
Zanou N., Shapovalov G., Louis M., Tajeddine N., Gallo C., Van Schoor M., et al. (2010). Role of TRPC1 channel in skeletal muscle function. Am. J. Physiol. Cell. Physiol. 298, C149–C162. doi:10.1152/ajpcell.00241.2009
Zhang L., Kelley J., Schmeisser G., Kobayashi Y. M., Jones L. R. (1997). Complex formation between junctin, triadin, calsequestrin, and the ryanodine receptor. Proteins of the cardiac junctional sarcoplasmic reticulum membrane. J. Biol. Chem. 272, 23389–23397. doi:10.1074/jbc.272.37.23389
Zhang L., Wang L., Li S., Xue J., Luo D. (2016). Calsequestrin-1 regulates store-operated Ca2+ entry by inhibiting STIM1 aggregation. Cell. Physiol. biochem. 38, 2183–2193. doi:10.1159/000445574
Zhang S. L., Yu Y., Roos J., Kozak J. A., Deerinck T. J., Ellisman M. H., et al. (2005). STIM1 is a Ca2+ sensor that activates CRAC channels and migrates from the Ca2+ store to the plasma membrane. Nature 437, 902–905. doi:10.1038/nature04147
Zhao M., Hollingworth S., Baylor S. M. (1996). Properties of tri- and tetracarboxylate Ca2+ indicators in frog skeletal muscle fibers. Biophys. J. 70, 896–916. doi:10.1016/S0006-3495(96)79633-9
Zhao X., Min C. K., Ko J.-K., Parness J., Kim D. H., Weisleder N., et al. (2010). Increased store-operated Ca2+ entry in skeletal muscle with reduced calsequestrin-1 expression. Biophys. J. 99, 1556–1564. doi:10.1016/j.bpj.2010.06.050
Zhao X., Weisleder N., Han X., Pan Z., Parness J., Brotto M., et al. (2006). Azumolene inhibits a component of store-operated calcium entry coupled to the skeletal muscle ryanodine receptor. J. Biol. Chem. 281, 33477–33486. doi:10.1074/jbc.M602306200
Zhao Y., Huang G., Wu J., Wu Q., Gao S., Yan Z., et al. (2019). Molecular basis for ligand modulation of a mammalian voltage-gated Ca2+ channel. Cell. 177, 1495–1506. e12. doi:10.1016/j.cell.2019.04.043
Zhou J., Yi J., Fu R., Liu E., Siddique T., Ríos E., et al. (2010). Hyperactive intracellular calcium signaling associated with localized mitochondrial defects in skeletal muscle of an animal model of amyotrophic lateral sclerosis. J. Biol. Chem. 285, 705–712. doi:10.1074/jbc.M109.041319
Keywords: skeletal muscle, excitation-contraction coupling, Ca2+, fluorescence, Ca2+ channels, ryanodine receptor—RYR1, mitochondria
Citation: Bolaños P and Calderón JC (2022) Excitation-contraction coupling in mammalian skeletal muscle: Blending old and last-decade research. Front. Physiol. 13:989796. doi: 10.3389/fphys.2022.989796
Received: 08 July 2022; Accepted: 08 August 2022;
Published: 02 September 2022.
Edited by:
Enrique Jaimovich, University of Chile, ChileReviewed by:
Gonzalo Pizarro, Universidad de la República, UruguayErick Omar Hernandez-Ochoa, University of Maryland, United States
Copyright © 2022 Bolaños and Calderón. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Juan C. Calderón, amNhbGRlcm9udjAwQHlhaG9vLmNvbQ==
†ORCID: Pura Bolaños, orcid.org/0000-0002-0999-7666; Juan C. Calderón, orcid.org/0000-0002-8695-6972