
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Physiol., 01 September 2021
Sec. Craniofacial Biology and Dental Research
Volume 12 - 2021 | https://doi.org/10.3389/fphys.2021.723105
This article is part of the Research TopicCleft Lip and Palate Anomalies/SyndromesView all 11 articles
 Erika Calvano Küchler1,2*
Erika Calvano Küchler1,2* Caio Luiz Bitencourt Reis2
Caio Luiz Bitencourt Reis2 Alice Corrêa Silva-Sousa3
Alice Corrêa Silva-Sousa3 Guido Artemio Marañón-Vásquez4
Guido Artemio Marañón-Vásquez4 Mirian Aiko Nakane Matsumoto2Aline Sebastiani5
Mirian Aiko Nakane Matsumoto2Aline Sebastiani5 Rafaela Scariot5Eva Paddenberg1
Rafaela Scariot5Eva Paddenberg1 Peter Proff1
Peter Proff1 Christian Kirschneck1*
Christian Kirschneck1*Tooth agenesis is a common congenital anomaly in humans and is more common in oral cleft patients than in the general population. Many previous studies suggested that oral cleft and tooth agenesis share a similar genetic background. Therefore, this study explored the association between isolated tooth agenesis and genetic polymorphisms in genes that are crucial for craniofacial and tooth development. Panoramic radiographs, anamnesis, and genomic DNA from 273 patients were included. Patients were classified as tooth agenesis present, when at least one permanent tooth was congenitally missing. Patients with syndromes and oral cleft were excluded. Only unrelated patients were included. The genetic polymorphisms in BMP2 (rs235768 and rs1005464), BMP4 (rs17563), RUNX2 (rs59983488 and rs1200425), and SMAD6 (rs3934908 and rs2119261) were genotyped by real-time polymerase chain reaction. Genotype and allele distributions were compared between the tooth agenesis phenotypes and controls by Chi-square test. Haplotype and diplotype analysis were also performed, in addition to multivariate analysis (alpha of 0.05). A total of 86 tooth agenesis cases and 187 controls were evaluated. For the rs235768 in BMP2, patients carrying TT genotype have higher chance to present tooth agenesis [p < 0.001; prevalence ratio (PR) = 8.29; 95% confidence interval (CI) = 4.26–16.10]. The TT genotype in rs3934908 (SMAD6) was associated with higher chance to present third molar agenesis (p = 0.023; PR = 3.25; 95% CI = 1.17–8.99). BMP2 was also associated in haplotype and diplotype analysis with tooth agenesis. In conclusion, genetic polymorphisms in BMP2 and SMAD6 were associated with isolated tooth agenesis.
Isolated tooth agenesis (or congenitally missing teeth) is one of the most common congenital defects in humans, which affects approximately 20% of the average worldwide population (Vastardis, 2000). Tooth agenesis can be classified into two main types: non-syndromic and syndromic. Non-syndromic tooth agenesis involves a congenitally missing permanent or primary tooth or teeth in an isolated form without any other major birth defects, such as oral cleft and syndromes. Isolated tooth agenesis occurs in both aches (maxilla and mandible) and can affect any type of teeth, although the most commonly affected teeth are third molars, maxillary lateral incisors, and premolar (Küchler et al., 2008a,b). Syndromic tooth agenesis refers to congenitally missing teeth associated with syndromes and oral clefts (such as cleft lip, clef palate, and cleft lip with palate) (Lu et al., 2016). Tooth agenesis is often observed in individuals with oral clefts (in the cleft area and also in non-cleft areas) and their non-affected family members (Küchler et al., 2011). Several observational epidemiological and genetic studies suggest that oral clefts and isolated tooth agenesis share a similar genetic background (Phan et al., 2016).
The bone morphogenetic protein (BMP) family, comprising an extensive group of phylogenetically conserved growth factors, such as BMP2 and BMP4, which plays an important role during tooth development (Zhang et al., 2005; Saadi et al., 2013; Taşlı et al., 2014; Yuan et al., 2015). Interestingly, genetic polymorphisms in BMP2 and BMP4 have been associated with both, isolated tooth agenesis (Antunes et al., 2012; Gong et al., 2015) and oral clefts (Antunes et al., 2013; Saket et al., 2016; Bahrami et al., 2020). Mothers against decapentaplegic homolog 6 (SMAD6) belongs to the SMAD family, which are important signaling pathway proteins during craniofacial development (Estrada et al., 2011; Suzuki et al., 2020). SMAD6 is known to inhibits BMP signaling in the nucleus by interacting with transcription repressors (Wu et al., 2016). Another molecule with a crucial role in craniofacial development is Runt-related transcription factor (RUNX2). This gene has been identified as essential for tooth formation (Camilleri and McDonald, 2006). Therefore, this study explored the association between isolated permanent tooth agenesis and genetic polymorphisms in genes that are crucial for tooth and craniofacial development.
The study protocol was reviewed and approved by the local Ethics Committee (no. 01451418.3.0000.5419). Informed consent was obtained from all participating individuals or parents/legal guardians during the dental appointment and the assent was also obtained from children. The guideline Strengthening the Reporting of Genetic Association (STREGA) was followed for this study (Little et al., 2009).
Pre-dental treatment records including anamnesis and panoramic radiographs from patients undergoing dental treatment in universities and private clinics in Curitiba, Paraná state and Ribeirão Preto, São Paulo state (both cities located in Brazil) were evaluated. The sample consisted of biologically unrelated individuals aged 8–43 years old. The exclusion criteria included patients younger than 7 years of age, patients with syndromes, oral clefts, history of facial trauma or facial surgery, and records with missing radiographs in the tooth agenesis and control group.
The control and tooth agenesis cases were identified by the assessment of panoramic radiographs and treatment records by two trained dentists. All panoramic radiographs were examined using the same protocol and in all cases tooth agenesis was clearly evident from the panoramic radiographs alone (Küchler et al., 2008a,b). The inclusion criterion in the tooth agenesis group was that at least one permanent tooth was affected. Tooth agenesis was defined based on the age of subjects and when initial tooth formation should be visible in the radiographs (Küchler et al., 2008a,b). All controls had all permanent teeth, including third molars. Patients with tooth extraction were excluded.
Tooth agenesis cases were also divided into third molar agenesis and other permanent tooth agenesis subgroups for the analysis.
The selection of the genes was initially based on the screening of previously published studies suggesting that these genes are involved in the maxilla, mandible, and tooth development phenotypes. Then, potentially functional genetic polymorphisms were screened from the dbSNP database1 and SNPinfo,2 based on the following criteria: minor allele frequency (MAF) ≥10% in the global population, and classification of the genetic polymorphisms as potentially functional (for altering amino acid sequence of the protein product, or for occurring in the proximal promoter of the gene and potentially influencing gene expression, or previously associated with craniofacial phenotypes). The characteristics of the selected genetic polymorphisms are presented in Table 1.
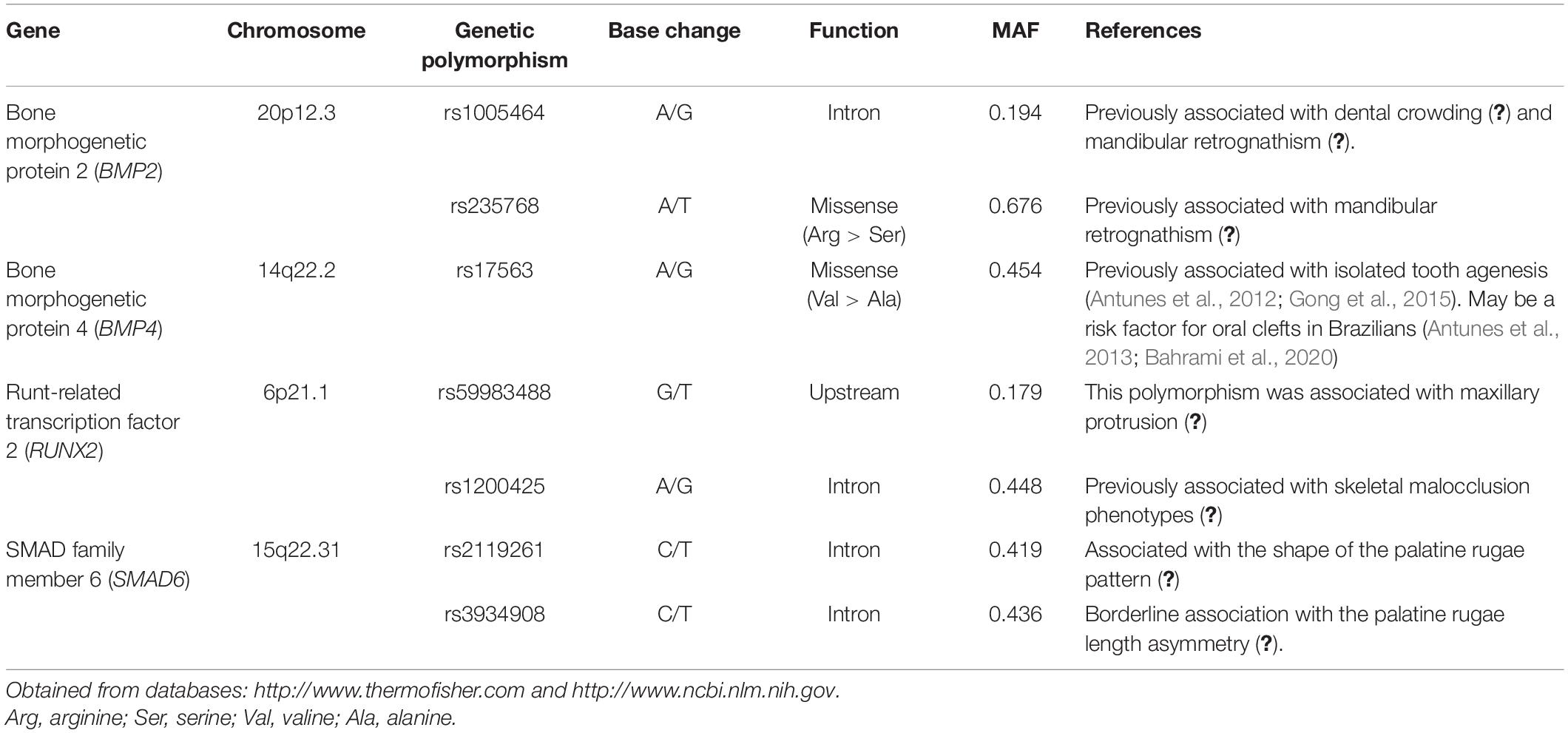
Table 1. Characteristics of the selected genetic polymorphisms and previously reported associations with oral phenotypes.
For the genotyping analysis genomic DNA was used. The DNA was isolated from buccal epithelial cells by a rinse of saline solution. Briefly, the tubes with saliva were centrifuged and supernatant was discarded. Extraction solution (Tris–HCl 10 mmol/L, pH 7.8; EDTA 5 mmol/L; SDS 0.5%, 1 mL) and proteinase K (100 ng/mL) were added to the tube. Ammonium acetate also was added to remove non-digested proteins and the solution centrifuged. DNA was precipitated with isopropanol and washed with ethanol. DNA was then resuspended and quantified by spectrophotometry (NanoDrop 1000; Thermo Scientific, Wilmington, DE, United States) (Küchler et al., 2012).
The selected genetic polymorphisms were blindly genotyped via real-time polymerase chain reaction (PCR) StepOneTM using TaqManTM technology (Applied Biosystems). The TaqMan technology uses extremely sensitive allele-specific probes (VICTM and FAMTM dyes were used for the alleles). A negative control template was included in every reaction plate. In addition, 10% of samples were randomly selected for repeated analysis and the results showed 100% concordance. DNA samples that failed to be genotyped were excluded from further analyses. The success rates were as follows: rs235768 (BMP2) = 86.0%; rs1005464 (BMP2) = 84.9%; rs17563 (BMP4) = 84.2%; rs59983488 (RUNX2) = 85.5%; rs1200425 (RUNX2) = 83.8%; rs3934908 (SMAD6) = 84.9%; rs2119261 (SMAD6) = 84.2%.
Hardy–Weinberg Equilibrium (HWE) was assessed for each genetic polymorphism by Chi-square test.3 Chi-square test was also used to compare the allele and genotype distribution according to tooth agenesis phenotypes. Prevalence ratio (PR) and 95% confidence intervals (CI) were obtained. PLINK4 was used to compare haplotype frequencies between groups using Fisher’s exact test.
Multivariate Poisson regression adjusted by gender and ethnicity was done to evaluate genotypes in the co-dominant model and also diplotypes. Diplotype is a combination of two haplotypes that may be evaluated by an interaction term (Gatlin et al., 2009; Guo et al., 2017). Poisson regression was performed using SPSS Statistics Version 25.0 (IBM Corp., New York, NY, United States).
The significance level was determined as p < 0.05.
A total of 273 individuals (116 males and 157 females) was included. Eighty-six were included in the tooth agenesis group and 187 in the control group. In the tooth agenesis group, 53 individuals presented third molar agenesis (61.6%) and 42 (48.4%) presented other types of missing teeth. Gender and ethnicity were not associated with tooth agenesis (p > 0.05) (Supplementary Table 1).
Table 2 shows the allele distribution according to the groups. The allele T of the rs235768 in BMP2 was associated with higher chance to present tooth agenesis in comparison with control group (p < 0.001; PR = 3.45; 95% CI = 2.54–4.70). In the subgroup analysis, the T allele was also associated with higher chance to present third molar agenesis (p < 0.001; PR = 4.30; 95% CI = 2.83–6.52) and other agenesis (p < 0.001; PR = 4.48; 95% CI = 2.78–7.21). The allele C of the rs3934908 in SMAD6 was associated only with higher chance to present third molar agenesis (p = 0.036; PR = 1.43; CI 95% = 1.00–2.09).
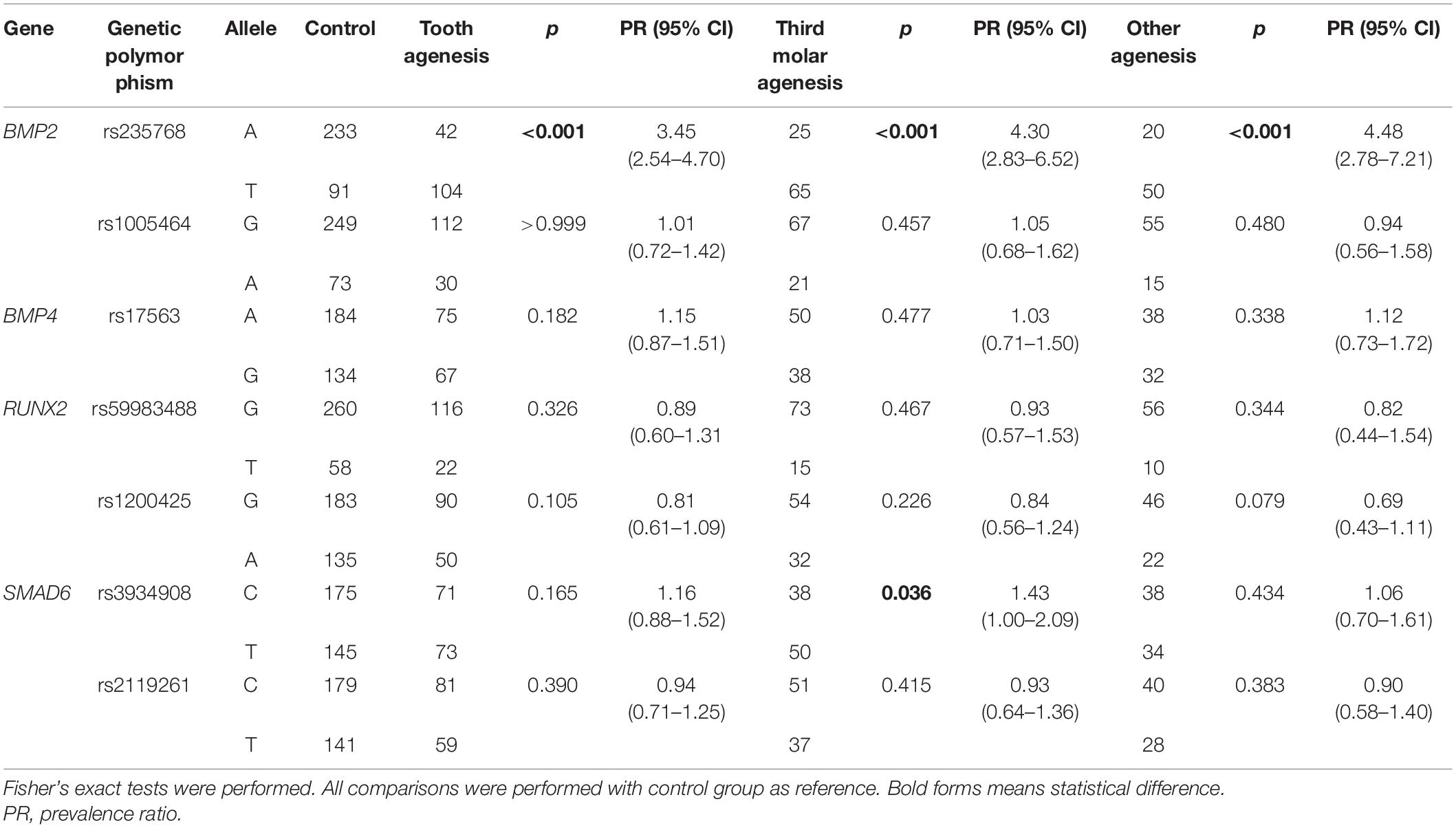
Table 2. Allelic distribution between groups.
Table 3 presents the haplotype frequency comparisons between groups. All haplotypes formed by the rs235768 and rs1005464 polymorphisms in BMP2 were associated with tooth agenesis and tooth agenesis subgroups (p < 0.001). The haplotypes T–G and T–A were more frequent in tooth agenesis cases, while the haplotypes A–G and A–A were more frequent in controls.
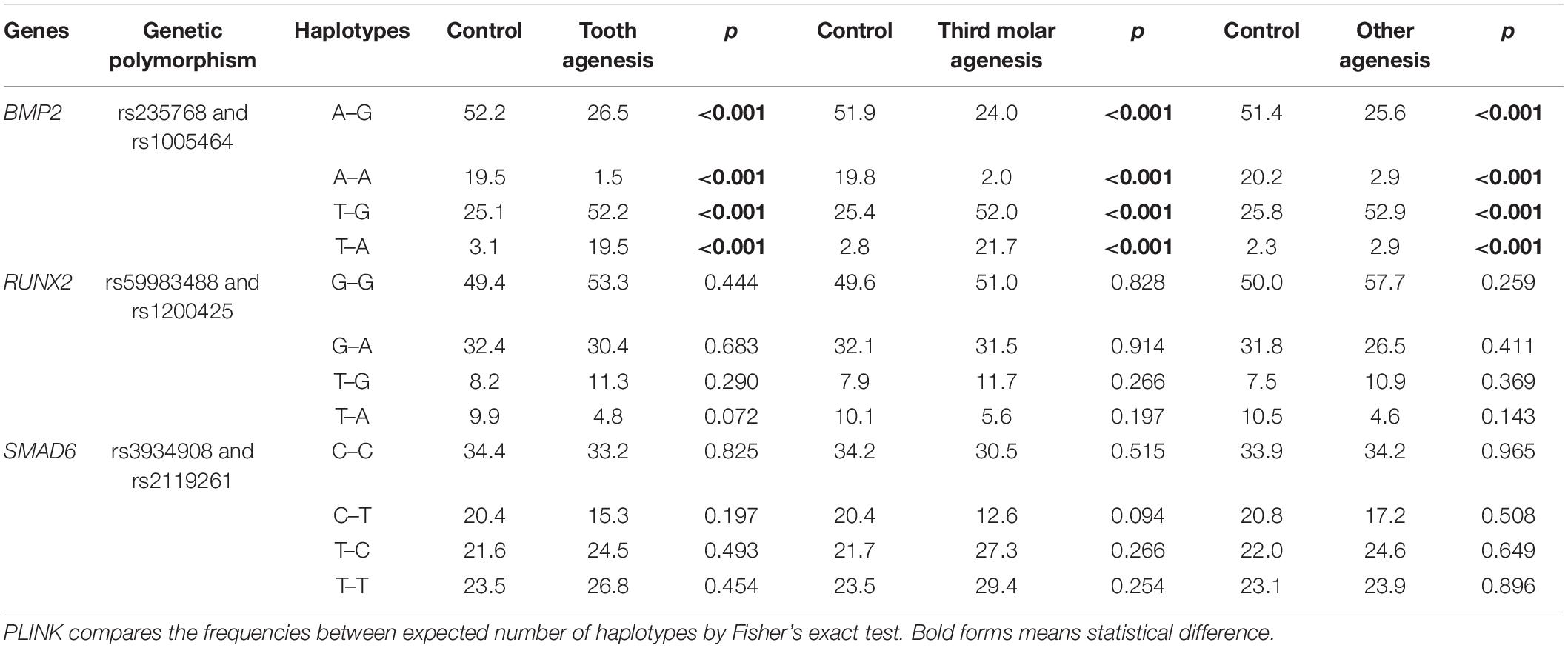
Table 3. Haplotypes frequency comparisons between groups.
Table 4 shows the genotype distribution among groups in the co-dominant model. The rs235768 in BMP2 was associated with tooth agenesis in univariate and multivariate analysis (p < 0.001). The genotype TT in rs3934908 in SMAD6 was associated with an increased chance to present third molar agenesis (p = 0.023; PR = 3.25; CI 95% = 1.17–8.99).
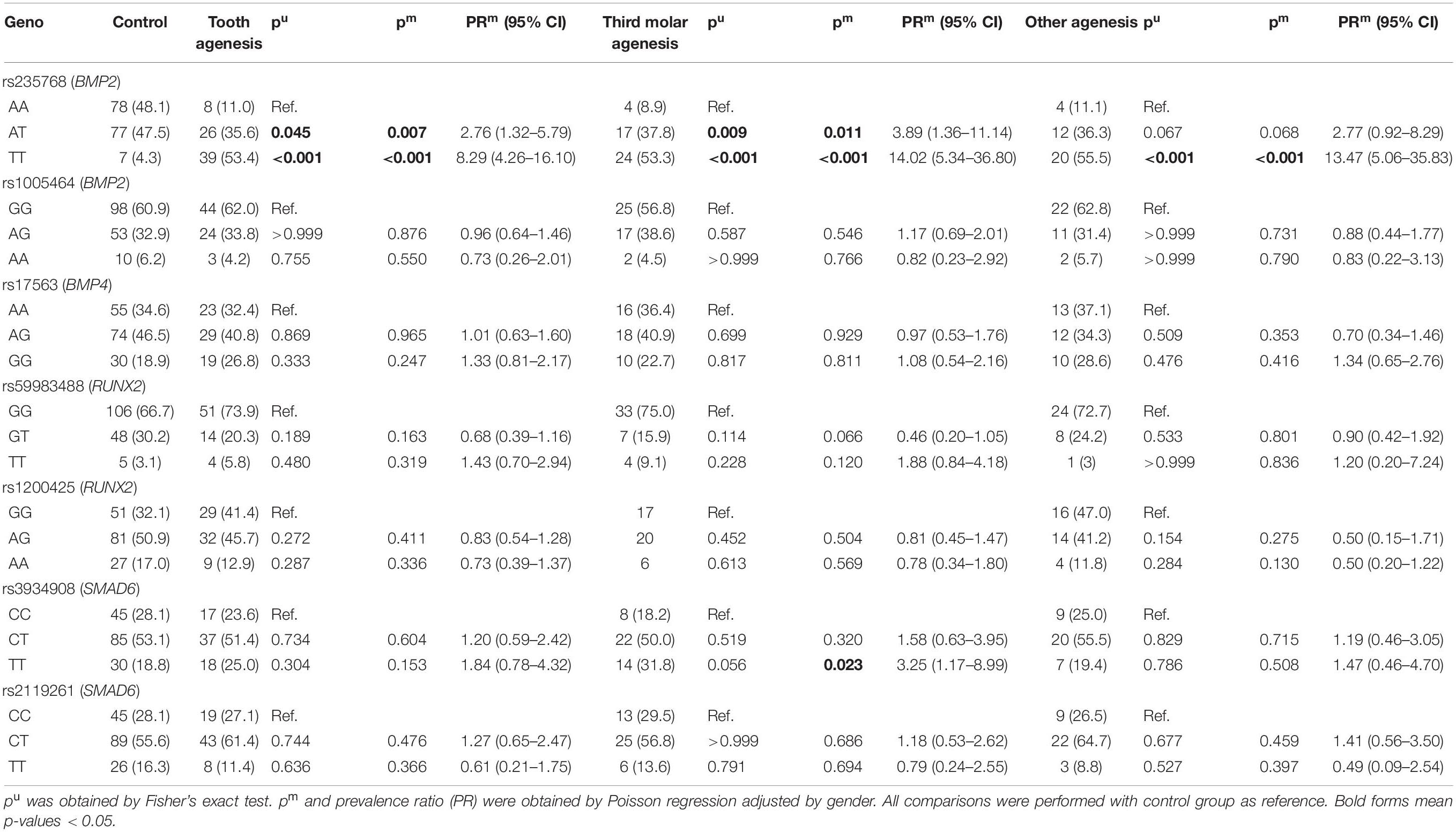
Table 4. Genotypic distribution between groups.
Table 5 shows the diplotype analysis. The diplotype analysis of the studied genetic polymorphisms in BMP2 demonstrated that individuals carrying TT + AA genotypes (rs235768–rs1005464) had a higher chance to present tooth agenesis (p = 0.016; PR = 5.33; 95% CI = 1.36–20.83), third molar agenesis (p = 0.013; PR = 10.15; 95% CI = 1.62–63.29), and other agenesis (p = 0.020; PR = 7.64; 95% CI = 1.38–42.16) than the control individuals.
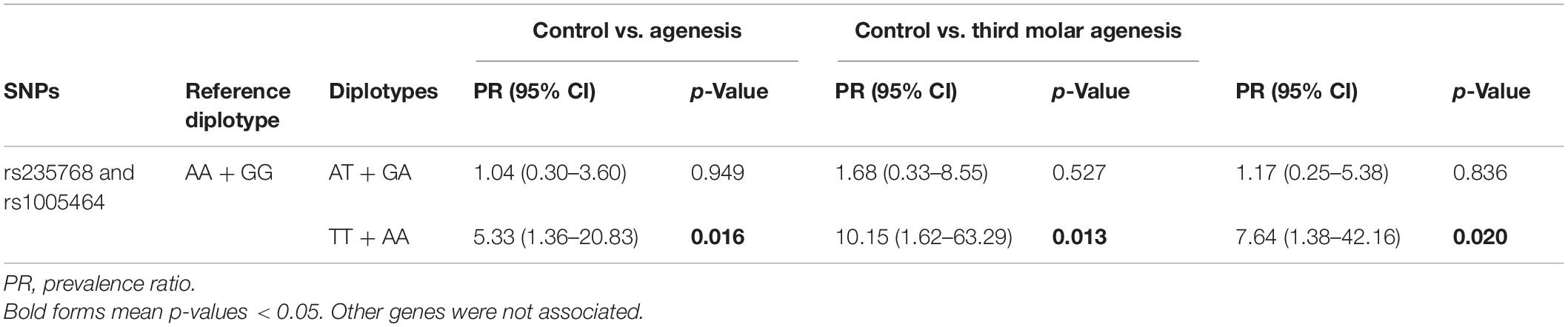
Table 5. Diplotype analysis with BMP2 gene SNPs by Poisson regression adjusted by gender and ethnicity.
Dental development results from several interactions acting synergistically and antagonistically, leading to tooth epithelium and mesenchyme formation. The process is regulated by different mechanisms involving the expression of several genes (Nieminen, 2009). Mutations and/or genetic polymorphisms in one or more genes involved in the earlier stages of dental development could potentially lead to tooth agenesis. Therefore, in the present study, we replicated genotype–phenotype associations previously observed (Antunes et al., 2012; Gong et al., 2015) and also investigated the association of some novel genes and polymorphisms with isolated tooth agenesis.
In the past decades, innumerous evidence suggests the association between tooth agenesis and oral clefting [revised by Phan et al. (2016)]. Phan et al. (2016) systematically investigated the currently available literature to investigate co-occurrence of tooth agenesis and oral clefts to gain insight into the molecular mechanisms underlying their dual involvement in the development of teeth and facial primordia. The authors concluded that not only the disrupted gene, but even the location of the mutations within the gene can lead to diverse phenotypes, ranging from the isolated form of tooth agenesis to the syndromic one for of oral clefts. In fact, genes involved in oral clefts and in the syndromic form of tooth agenesis are known as a useful approach to select candidate genes for the isolated forms of tooth agenesis (Vieira, 2003). Another important approach to select candidate genes for isolated tooth agenesis is based on the identification of the genes expressed in dental development. So far, more than 300 genes are listed in the database created to catalog genes expressed in different stages of dental development.5 Therefore, the genes were selected in our study based on their previous associations and their expression and role in dental development.
Genetic polymorphisms are DNA sequence variations occurring in the genome that are characterized by the existence of at least two variants (Botstein and Risch, 2003). They are involved in many phenotypic differences observed in clinical practice. The selection of the genetic polymorphisms studied here was based on their MAF due to our limited sample size. Previous associations with craniofacial phenotypes were also taken into consideration (as shown in Table 1) and also the function of the genetic polymorphism.
Two genetic polymorphisms (rs17563 and rs235768) selected here are missense variations located within the coding region and produce amino acid changes. The rs17563 in BMP4 studied here replaced the amino acid valine by alanine at position 152 of the protein. This genetic polymorphism has been widely investigated in oral cleft research. A recent systematic review and comprehensive meta-analysis investigated case-control studies with 2,058 oral cleft cases and 2,557 controls. In their overall analysis, no significant association between the rs17563 polymorphism and the risk of oral cleft was observed, however, their subgroup analysis demonstrated that rs17563 was associated with oral cleft risk in Chinese and Brazilian populations (Bahrami et al., 2020). The polymorphism rs17563 was also associated with isolated tooth agenesis in Brazilians and Chinese (Antunes et al., 2012; Gong et al., 2015). Although we also investigated Brazilians, the lack of association observed in our study may be explained by the fact that these previous studies excluded third molars. Although we also performed a stratified analysis excluding third molars, the sample size could lead to a false-negative result, once it is well-known that BMP4 is important for tooth development and the BMP4 expression pattern coincides with the bud-to-cap stage transition in tooth development (Saadi et al., 2013).
The other missense variant studied here was the rs235768, which is located in BMP2 resulting in an arginine to serine replacement. BMP2, another important member of the BMP family involved in regulating tooth initiation, can induce human tooth germ cells to differentiate into odontogenic and osteogenic cells (Zhang et al., 2005; Taşlı et al., 2014). In animal models, BMP2 expressed in the presumptive dental epithelium (Neubüser et al., 1997) could result in the arrest of tooth development after knockdown (Yuan et al., 2015). In fact, our study demonstrated interesting results in both studied genetic polymorphisms in BMP2. To carry the T allele increases the risk to present tooth agenesis. The haplotype and diplotype analysis also showed that rs235768 interacts with the intronic variant rs1005464 and is involved in the risk for any type of tooth agenesis, including third molar agenesis.
Third molars are the most common congenitally missing teeth, followed by premolars and maxillary lateral incisors (Polder et al., 2004). Although third molar agenesis is a phenotype highly prevalent in humans, its etiology has been poorly explored so far. A recent study evaluated a large sample of same sex twins (172 monozygotic and 112 dizygotic) and concluded that a dominant factor for third molar agenesis is genetics (Trakinienë et al., 2018). However, the genes involved in third molar agenesis are still unexplored. Our results suggest that genes/genetic polymorphisms involved in the agenesis of other tooth types could be candidate for third molar agenesis studies.
Mothers against decapentaplegic homolog 6 is known to interact with BMP signaling in the nucleus by interacting with transcription repressors (Wu et al., 2016), including BMP2 (Li et al., 2003). SMAD6 is important for craniofacial development (Estrada et al., 2011; Timberlake et al., 2016). A previous study identified rare SMAD6 and common BMP2 alleles involved with craniosynostosis in humans (Timberlake et al., 2016). In our study, genotype and allele distribution of the intronic variant rs3934908 in SMAD6 was associated with third molar agenesis.
Runt-related transcription factor is well known to be involved in tooth development (Camilleri and McDonald, 2006). RUNX2-deficient mice show an arrest of molar tooth development at the early cap stage, suggesting that RUNX2 is required for the progression of tooth development from the cap stage to the bell stage (D’Souza et al., 1999) and therefore is a candidate for isolated tooth agenesis. In our study, some borderline association was observed for the studied genetic polymorphisms in RUNX2, suggesting that future studies should investigate the association of variations in this gene with isolated tooth agenesis in a larger population.
Although our study provides some interesting information regarding the genes involved in the etiology of isolated tooth agenesis, it has some obvious limitations. The limited sample size could lead to a type II error in the analysis of genetic polymorphisms with small effect. Also, the number of selected genetic polymorphisms does not cover the studied genes. Additionally, future studies should also evaluate the role of these genetic polymorphisms in tooth agenesis risk in oral cleft individuals.
In conclusion, our study suggested that genetic polymorphisms in BMP2 and SMAD6 are involved in a higher chance to present isolated tooth agenesis.
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding authors.
The studies involving human participants were reviewed and approved by Local Ethics Committee (no. 01451418.3.0000.5419). Written informed consent to participate in this study was provided by the participants’ legal guardian/next of kin.
EK and CK conceptualize the study. EK, MM, and RS designed and organize the sample recruitment. GM-V and AS performed the sample collection. CR, GM-V, and AS-S performed the laboratory analysis. CR, AS-S, and EP analyzed the data. CR, EK, GM-V, AS-S, and CK wrote the manuscript. EK, PP, and CK funding support. All authors read and approved the final version of the manuscript.
This study was financed in part by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – Brasil (CAPES) – Finance Code 001 and Alexander-von-Humboldt-Foundation (Küchler/Kirschneck accepted in July 4th, 2019). This work was also supported by the São Paulo Research Foundation (FAPESP) (2015/06866-5) and individual scholarships (CAPES).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – Brasil (CAPES), Alexander-von-Humboldt-Foundation, and National Council for Scientific and Technological Development (CNPq).
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fphys.2021.723105/full#supplementary-material
Antunes, L. S., Küchler, E. C., Tannure, P. N., Costa, M. C., Gouvêa, C. V., Olej, B., et al. (2013). BMP4 polymorphism is associated with nonsyndromic oral cleft in a Brazilian population. Cleft Palate Craniofac. J. 50, 633–638. doi: 10.1597/12-048
Antunes, L. S., Küchler, E. C., Tannure, P. N., Lotsch, P. F., Costa, M. C., Gouvêa, C. V., et al. (2012). TGFB3 and BMP4 polymorphism are associated with isolated tooth agenesis. Acta Odontol. Scand. 70, 202–206. doi: 10.3109/00016357.2011.629626
Bahrami, R., Dastgheib, S. A., Niktabar, S. M., Amooee, A., Lookzadeh, M. H., Mirjalili, S. R., et al. (2020). Association of BMP4 rs17563 polymorphism with nonsyndromic cleft lip with or without cleft palate risk: literature review and comprehensive meta-analysis. Fetal Pediatr. Pathol. 40, 305–319. doi: 10.1080/15513815.2019.1707916
Botstein, D., and Risch, N. (2003). Discovering genotypes underlying human phenotypes: past successes for mendelian disease, future approaches for complex disease. Nat. Genet. 33(Suppl), 228–237. doi: 10.1038/ng1090
Camilleri, S., and McDonald, F. (2006). Runx2 and dental development. Eur. J. Oral Sci. 114, 361–373. doi: 10.1111/j.1600-0722.2006.00399.x
D’Souza, R. N., Aberg, T., Gaikwad, J., Cavender, A., Owen, M., Karsenty, G., et al. (1999). Cbfa1 is required for epithelial-mesenchymal interactions regulating tooth development in mice. Development 126, 2911–2920. doi: 10.1242/dev.126.13.2911
Estrada, K. D., Retting, K. N., Chin, A. M., and Lyons, K. M. (2011). Smad6 is essential to limit BMP signaling during cartilage development. J. Bone Miner. Res. 26, 2498–2510. doi: 10.1002/jbmr.443
Gatlin, M. R., Black, C. L., Mwinzi, P. N., Secor, W. E., Karanja, D. M., and Colley, D. G. (2009). Association of the gene polymorphisms IFN-gamma +874, IL-13 -1055 and IL-4 -590 with patterns of reinfection with Schistosoma mansoni. PLoS Negl. Trop. Dis. 3:e375. doi: 10.1371/journal.pntd.0000375
Gong, M., Qian, Y. J., Gu, N., Wang, W., Wang, H., Ma, L., et al. (2015). Association of BMP4 polymorphisms with isolated tooth agenesis in a Chinese Han population: a case-control study. Eur. Rev. Med. Pharmacol. Sci. 19, 2188–2194.
Guo, T., Yin, R. X., Pan, L., Yang, S., Miao, L., and Huang, F. (2017). Integrative variants, haplotypes and diplotypes of the CAPN3 and FRMD5 genes and several environmental exposures associate with serum lipid variables. Sci. Rep. 7:45119. doi: 10.1038/srep45119
Küchler, E. C., Risso, P. A., Costa, M. C., Modesto, A., and Vieira, A. R. (2008a). Assessing the proposed association between tooth agenesis and taurodontism in 975 paediatric subjects. Int. J. Paediatr. Dent. 18, 231–234. doi: 10.1111/j.1365-263X.2007.00876.x
Küchler, E. C., Risso, P. A., Costa, M. C., Modesto, A., and Vieira, A. R. (2008b). Studies of dental anomalies in a large group of school children. Arch. Oral Biol. 53, 941–946. doi: 10.1016/j.archoralbio.2008.04.003
Küchler, E. C., Tannure, P. N., Falagan-Lotsch, P., Lopes, T. S., Granjeiro, J. M., and Amorim, L. M. (2012). Buccal cells DNA extraction to obtain high quality human genomic DNA suitable for polymorphism genotyping by PCR-RFLP and Real-Time PCR. J. Appl. Oral Sci. 20, 467–471. doi: 10.1590/s1678-77572012000400013
Küchler, E. C., Reis, C. L. B., Carelli, J., Scariot, R., Nelson-Filho, P., Coletta, R. D., et al. (2021). Potential interactions among single nucleotide polymorphisms in bone- and cartilage-related genes in skeletal malocclusions. Orthod Craniofac Res. 24, 277–287. doi: 10.1111/ocr.12433
Küchler, E. K., Motta, L. G., Vieira, A. R., and Granjeiro, J. M. (2011). Side of dental anomalies and taurodontism as potential clinical markers for cleft subphenotypes. Cleft Palate Craniofac. J. 48, 103–108. doi: 10.1597/09-159
Li, X., Ionescu, A. M., Schwarz, E. M., Zhang, X., Drissi, H., Puzas, J. E., et al. (2003). Smad6 is induced by BMP-2 and modulates chondrocyte differentiation. J. Orthop. Res. 21, 908–913. doi: 10.1016/S0736-0266(03)00008-1
Little, J., Higgins, J. P., Ioannidis, J. P., Moher, D., Gagnon, F., Von Elm, E., et al. (2009). STrengthening the REporting of Genetic Association Studies (STREGA)—an extension of the STROBE statement. Genet. Epidemiol. 33, 581–598.
Lu, Y., Qian, Y., Zhang, J., Gong, M., Wang, Y., Gu, N., et al. (2016). Genetic variants of BMP2 and their association with the risk of non-syndromic tooth agenesis. PLoS One 11:e0158273. doi: 10.1371/journal.pone.0158273
Neubüser, A., Peters, H., Balling, R., and Martin, G. R. (1997). Antagonistic interactions between FGF and BMP signaling pathways: a mechanism for positioning the sites of tooth formation. Cell 90, 247–255. doi: 10.1016/s0092-8674(00)80333-5
Nieminen, P. (2009). Genetic basis of tooth agenesis. J. Exp. Zool. B Mol. Dev. Evol. 15:312B. doi: 10.1002/jez.b.21277
Phan, M., Conte, F., Khandelwal, K. D., Ockeloen, C. W., Bartzela, T., Kleefstra, T., et al. (2016). Tooth agenesis and orofacial clefting: genetic brothers in arms? Hum. Genet. 135, 1299–1327. doi: 10.1007/s00439-016-1733-z
Polder, B. J., Van’t Hof, M. A., Van der Linden, F. P., and Kuijpers-Jagtman, A. M. (2004). A meta-analysis of the prevalence of dental agenesis of permanent teeth. Commun. Dent. Oral Epidemiol. 32, 217–226. doi: 10.1111/j.1600-0528.2004.00158.x
Saadi, I., Das, P., Zhao, M., Raj, L., Ruspita, I., Xia, Y., et al. (2013). Msx1 and Tbx2 antagonistically regulate Bmp4 expression during the bud-to-cap stage transition in tooth development. Development 140, 2697–2702. doi: 10.1242/dev.088393
Saket, M., Saliminejad, K., Kamali, K., Moghadam, F. A., Anvar, N. E., and Khorram Khorshid, H. R. (2016). BMP2 and BMP4 variations and risk of non-syndromic cleft lip and palate. Arch. Oral Biol. 72, 134–137. doi: 10.1016/j.archoralbio.2016.08.019
Silva-Sousa, A. C., Marañón-Vásquez, G. A., Stuani, M. B. S., Proff, P., Andrades, K. M. R., Baratto-Filho, F., et al. (2021). Genetic variants in bone morphogenetic proteins signaling pathway might be involved in palatal rugae phenotype in humans. Sci. Rep. 11:12715. doi: 10.1038/s41598-021-92169-0
Suzuki, Y., Nakajima, A., Kawato, T., Iwata, K., Motoyoshi, M., and Shuler, C. F. (2020). Identification of Smad-dependent and -independent signaling with transforming growth factor-β type 1/2 receptor inhibition in palatogenesis. J. Oral Biol. Craniofac. Res. 10:a022194. doi: 10.1016/j.jobcr.2020.01.002
Taşlı, P. N., Aydın, S., Yalvaç, M. E., and Sahin, F. (2014). Bmp 2 and bmp 7 induce odonto- and osteogenesis of human tooth germ stem cells. Appl. Biochem. Biotechnol. 172, 3016–3025. doi: 10.1007/s12010-013-0706-0
Timberlake, A. T., Choi, J., Zaidi, S., Lu, Q., Nelson-Williams, C., Brooks, E. D., et al. (2016). Two locus inheritance of non-syndromic midline craniosynostosis via rare SMAD6 and common BMP2 alleles. elife 5:e20125. doi: 10.7554/eLife.20125
Ting, T. Y., Wong, R. W. K., and Rabie, A. B. M. (2011). Analysis of genetic polymorphisms in skeletal Class I crowding. Am. J. Orthod. Dentofacial. Orthop. 140, e9–15. doi: 10.1016/j.ajodo.2010.12.015
Trakinienë, G., Šidlauskas, A., Andriuškevièiûtë, I., Šalomskienë, L., Švalkauskienë, V., Smailienë, D., et al. (2018). Impact of genetics on third molar agenesis. Sci. Rep. 8:8307. doi: 10.1038/s41598-018-26740-7
Vastardis, H. (2000). The genetics of human tooth agenesis: new discoveries for understanding dental anomalies. Am. J. Orthod. Dentofacial Orthop. 117, 650–656. doi: 10.1016/s0889-5406(00)70173-9
Vieira, A. R. (2003). Oral clefts and syndromic forms of tooth agenesis as models for genetics of isolated tooth agenesis. J. Dent. Res. 82, 162–165. doi: 10.1177/154405910308200303
Wu, M., Chen, G., and Li, Y. P. (2016). TGF-β and BMP signaling in osteoblast, skeletal development, and bone formation, homeostasis and disease. Bone Res. 4:16009. doi: 10.1038/boneres.2016.9
Yuan, G., Yang, G., Zheng, Y., Zhu, X., Chen, Z., Zhang, Z., et al. (2015). The non-canonical BMP and Wnt/β-catenin signaling pathways orchestrate early tooth development. Development 142, 128–139. doi: 10.1242/dev.117887
Keywords: genetic polymorphisms, craniofacial development, dental anomaly, tooth agenesis, single nucelotide polymorphisms
Citation: Küchler EC, Reis CLB, Silva-Sousa AC, Marañón-Vásquez GA, Matsumoto MAN, Sebastiani A, Scariot R, Paddenberg E, Proff P and Kirschneck C (2021) Exploring the Association Between Genetic Polymorphisms in Genes Involved in Craniofacial Development and Isolated Tooth Agenesis. Front. Physiol. 12:723105. doi: 10.3389/fphys.2021.723105
Received: 10 June 2021; Accepted: 13 August 2021;
Published: 01 September 2021.
Edited by:
Mohammad Khursheed Alam, Al Jouf University, Saudi ArabiaReviewed by:
Rosa Helena Wanderley Lacerda, Federal University of Paraíba, BrazilCopyright © 2021 Küchler, Reis, Silva-Sousa, Marañón-Vásquez, Matsumoto, Sebastiani, Scariot, Paddenberg, Proff and Kirschneck. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Christian Kirschneck, Y2hyaXN0aWFuLmtpcnNjaG5lY2tAa2xpbmlrLnVuaS1yZWdlbnNidXJnLmRl; Erika Calvano Küchler, ZXJpa2FjYWx2YW5vQGdtYWlsLmNvbQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.