
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Physiol., 25 February 2021
Sec. Invertebrate Physiology
Volume 12 - 2021 | https://doi.org/10.3389/fphys.2021.642237
This article is part of the Research TopicSystemic Coordination of Invertebrate HomeostasisView all 10 articles
 Michele Martins1†
Michele Martins1† Luis Felipe Costa Ramos1†
Luis Felipe Costa Ramos1† Jimmy Rodriguez Murillo2
Jimmy Rodriguez Murillo2 André Torres3
André Torres3 Stephanie Serafim de Carvalho1
Stephanie Serafim de Carvalho1 Gilberto Barbosa Domont1
Gilberto Barbosa Domont1 Danielle Maria Perpétua de Oliveira1
Danielle Maria Perpétua de Oliveira1 Rafael Dias Mesquita1
Rafael Dias Mesquita1 Fábio César Sousa Nogueira1
Fábio César Sousa Nogueira1 Rafael Maciel-de-Freitas4*
Rafael Maciel-de-Freitas4* Magno Junqueira1*
Magno Junqueira1*Zika virus (ZIKV) is a global public health emergency due to its association with microcephaly, Guillain-Barré syndrome, neuropathy, and myelitis in children and adults. A total of 87 countries have had evidence of autochthonous mosquito-borne transmission of ZIKV, distributed across four continents, and no antivirus therapy or vaccines are available. Therefore, several strategies have been developed to target the main mosquito vector, Aedes aegypti, to reduce the burden of different arboviruses. Among such strategies, the use of the maternally-inherited endosymbiont Wolbachia pipientis has been applied successfully to reduce virus susceptibility and decrease transmission. However, the mechanisms by which Wolbachia orchestrate resistance to ZIKV infection remain to be elucidated. In this study, we apply isobaric labeling quantitative mass spectrometry (MS)-based proteomics to quantify proteins and identify pathways altered during ZIKV infection; Wolbachia infection; co-infection with Wolbachia/ZIKV in the A. aegypti heads and salivary glands. We show that Wolbachia regulates proteins involved in reactive oxygen species production, regulates humoral immune response, and antioxidant production. The reduction of ZIKV polyprotein in the presence of Wolbachia in mosquitoes was determined by MS and corroborates the idea that Wolbachia helps to block ZIKV infections in A. aegypti. The present study offers a rich resource of data that may help to elucidate mechanisms by which Wolbachia orchestrate resistance to ZIKV infection in A. aegypti, and represents a step further on the development of new targeted methods to detect and quantify ZIKV and Wolbachia directly in complex tissues.
- The abundance of ZIKV polyprotein is reduced in the presence of Wolbachia.
- Shotgun proteomics quantifies ZIKV and Wolbachia proteins directly in tissues.
- Wolbachia regulates proteins involved in ROS production.
- Wolbachia regulates humoral immune response and antioxidant production.
- Metabolism and detoxification processes were associated with mono infections.
Zika virus (ZIKV) is a single-stranded RNA virus that belongs to the Flavivirus genus and Flaviviridae family. Originally discovered in a primate from the Zika forest in Uganda in 1947, remained unimportant for public health until the beginning of the 20th century (Dick et al., 1952). In 2014, ZIKV emerged in the Pacific islands and a few years later invaded the Americas, leading the World Health Organization to claim a global public health emergency due to its association with microcephaly in newborns and Guillain-Barré syndrome, neuropathy, and myelitis in children and adults (Campos et al., 2015; Barreto et al., 2016). As of May 2017, around 75% of inhabitants of the Yap Islands were infected with ZIKV, but no microcephaly was recorded. Whilst in Brazil more than 220,000 cases were recorded and more than 2,600 newborns had birth defects like microcephaly and other neuropathies (Baud et al., 2017).
The mosquito Aedes aegypti is the primary vector of ZIKV worldwide (Chouin-Carneiro et al., 2016; Ferreira-de-Brito et al., 2016). To date, no antivirus therapy or vaccines are available to mitigate ZIKV transmission, which increases the urgency for effective methods targeting the mosquito vector to reduce the burden of arboviruses as ZIKV. Among such strategies, the use of the maternally inherited endosymbiont Wolbachia pipientis has gained attention recently. The bacterium Wolbachia is naturally found in around 60% of arthropod species. Although Wolbachia does not naturally infect A. aegypti, some strains were transinfected by microinjection into this mosquito species with the further discovery of their pathogen-blocking properties (Moreira et al., 2009; Aliota et al., 2016; Dutra et al., 2016). The rationale underlying a mass release of Wolbachia infected mosquitoes is to replace the local A. aegypti population. Usually, it has high vector competence to arboviruses, like ZIKV, while the Wolbachia transinfected mosquitoes have reduced susceptibility and decreased transmission (Hoffmann et al., 2011). This approach is currently ongoing in at least 12 countries and the first communications showing its impact on disease transmission decrease are being revealed (Nazni et al., 2019; Indriani et al., 2020; Ryan et al., 2020).
The mechanisms which are orchestrated by Wolbachia, conferring resistance to arboviruses infection remain to be elucidated, however, it has been assigned to the activation of the mosquito immune system, down-regulation of genes that encode proteins or receptors necessary to the virus binding and depletion of host’s resources essentials to the virus life cycle (Ford et al., 2020). When infected with Wolbachia, the mosquito Toll pathway is activated through the increase of reactive oxygen species (ROS), resulting in the production of antimicrobial peptides like cecropin (Pan et al., 2012). The gene expression modulation of tubulin, insulin receptors, and other genes seems to hinder the virus infection (Haqshenas et al., 2019; Paingankar et al., 2020). Also, it has been proposed that the lipids are relevant to Wolbachia infection. While one study implies it competes with the host’s lipid resources (mainly cholesterol), others suggest it modulates the lipid production which can have a role blocking virus replication (Geoghegan et al., 2017; Koh et al., 2020; Manokaran et al., 2020).
Protein quantification on a genome-wide scale allows a systemic bird’s-eye view of protein variation and pathway regulation under different conditions and samples (Choudhary and Mann, 2010). Therefore, it helps to highlight some features and emergent properties of complex systems, which could be obscured when the analysis is performed only from a reductionist point of view. In this way, mass spectrometry (MS)-based proteomics, together with robust bioinformatic tools, allows the formulation of more ambitious hypotheses due to the possibility of generating large and detailed datasets. Reliable large-scale protein quantification can be achieved using isobaric labeling and shotgun proteomics (Pappireddi et al., 2019). Several quantitative methods have already been successfully applied in insects to access differentially regulated proteins (De Mandal et al., 2020; García-Robles et al., 2020; Serteyn et al., 2020). The A. aegypti infection with Mayaro virus or microsporidian parasites have already been studied (Duncan et al., 2012; Vasconcellos et al., 2020) and a previous descriptive work was based on comparing the proteome of A. aegypti female heads during blood and sugar meals conditions (Nunes et al., 2016).
Here, we use an isobaric labeling-based quantitative proteomic strategy to interrogate the interaction between the Wolbachia wMel strain and ZIKV infection in A. aegypti heads and salivary glands. Wolbachia is present in several tissues and organs in A. aegypti mosquitoes, with higher density in ovaries and tissues on the head and anterior part of the thorax, like the ommatidia cells, brain, salivary glands, and cardia (Moreira et al., 2009). To the best of our knowledge, this is the most complete proteome of A. aegypti reported so far, with a total of 3,790 proteins identified, corresponding to 25.75% of total protein-coding genes. This work includes the identification and quantification of several peptides from the ZIKV and 323 Wolbachia proteins in a complex sample tissue, paving the way to the development of sensitive approaches to detect and quantify the ZIKV and Wolbachia directly in the A. aegypti tissue via MS. Moreover, we describe and discuss proteins and pathways altered in A. aegypti during ZIKV infections, Wolbachia infections, co-infection Wolbachia/ZIKV, and compared with no infection.
To access the effects of ZIKV and Wolbachia infection on A. aegypti proteome, we used mosquitoes from two different sites in Rio de Janeiro (Rio de Janeiro State, Brazil): Tubiacanga (22°47′06″S; 43°13′32″W) and Porto (22°53′43″S, 43°11′03″W), distant 13 km apart from each other. Tubiacanga was selected to represent an area in which mosquitoes have Wolbachia (wMel strain) since it is the first site in Latin America with an established invasion (>90% frequency) (Garcia et al., 2019). Porto represents a Wolbachia free area. Eggs were collected through 80 ovitraps (Codeço et al., 2015) installed every 25–50 m each other in both sites. Ovitraps were placed over an extensive geographic area to ensure we captured the local A. aegypti genetic variability, collecting at least 500 eggs per site. The eggs were hatched and the mosquitoes were maintained at the insectary under a relative humidity of 80 ± 5% and a temperature of 25 ± 3°C, with ad libitum access to a 10% sucrose solution. The experimental infection was performed with mosquitoes from the F1 generation.
Aedes aegypti females were orally infected with the ZIKV strain Asian genotype isolated from a patient in Rio de Janeiro (GenBank accession number KU926309). Previous reports have shown high infectivity of this ZIKV isolate to Rio de Janeiro A. aegypti populations, including Porto, which had 100% infected females after receive an infective blood meal (Fernandes et al., 2016; da Silveira et al., 2018; Petersen et al., 2018). Viral titers in supernatants were previously determined by serial dilutions in Vero cells, expressed in plaque-forming units per milliliters (PFU/ml). All the assays were performed with samples containing 3.55 × 106 PFU/ml. Viral stocks were maintained at −80°C until its use.
Sugar supply was removed 36-h before mosquitoes were challenged with the infective blood meal to increase female’s avidity. Six–seven days old inseminated A. aegypti females from each of the two populations (Tubiacanga and Porto) were separated in cylindrical plastic cages for blood-feeding. Oral infection procedures were performed through a membrane feeding system (Hemotek, Great Harwood, United Kingdom), adapted with a pig-gut covering, which gives access to human blood. The infective blood meals consisted of 1 ml of the supernatant of infected Vero cell culture, 2 ml of washed rabbit erythrocytes, and 0.5 mM of adenosine triphosphate (ATP) as phagostimulant. The same procedure and membrane feeding apparatus were used to feed control mosquitoes, i.e., control insects received a noninfectious blood meal, with 1 ml of Vero cell culture medium replacing the viral supernatant (therefore, with no ZIKV), 2 ml of washed rabbit erythrocytes, and 0.5 mM of ATP as phagostimulant. After the experimental infection, we had a total of 152 A. aegypti females.
The maintenance of A. aegypti colonies with Wolbachia in the lab were achieved by offering blood obtained from anonymous donors from the blood bank of the Rio de Janeiro State University, whose blood bags would be discarded due to small volume. We have no information on donors, including sex, age, and clinical condition, but the blood bank discards those bags positive for Hepatitis B, Hepatitis C, Chagas disease, syphilis, human immunodeficiency virus, and human T-cell lymphotropic virus. Before offering the blood for mosquitoes, it was screened for Dengue virus (DENV) using the Dengue NS1 Ag STRIP (Bio-Rad). The use of human blood was approved by the Fiocruz Ethical Committee (CAAE 53419815.9.0000.5248).
Proteins from a total of 152 A. aegypti females were processed, of which 39 were coinfected with Wolbachia and ZIKV (WZ); 36 non-infected (A); 35 infected with Wolbachia (W) only and 42 infected with ZIKV (Z). On 14 days post-infection, each mosquito head plus the salivary gland was separated from the body using needles and a scalpel blade which were sterilized after every single use (Schmid et al., 2017). Proteins were extracted by lysis with a buffer containing 7 M urea, 2 M thiourea, 50 mM HEPES pH 8, 75 mM NaCl, 1 mM EDTA, 1 mM PMSF, and protease/phosphatase inhibitor cocktails (Roche). Then, samples were vortexed, sonicated in a cold bath for 10 min and sonicated by the probe for 10 min. Lysates were centrifuged at 10,000 g for 10 min at 4°C and the supernatants were carefully transferred to new tubes for further steps.
The protein concentration was determined by the Qubit Protein Assay Kit® fluorometric (Invitrogen), following the manufacturer’s instructions. A total of 100 μg of proteins from each condition was processed. Protein samples were incubated with dithiothreitol (DTT; GE Healthcare) at 10 mM and 30°C for 1 h. Subsequently, iodoacetamide solution (GE Healthcare) was added, with a final concentration of 40 mM. The reaction was performed at room temperature, in the dark, for 30 min. The samples were diluted 10× with 50 mM HEPES buffer to reduce the concentration of urea/thiourea. The solution was incubated with trypsin (Promega) in a 1:50 (w/w, enzyme/protein) ratio at 37°C for 18 h. The resulting peptides were desalted with a C-18 macro spin column (Harvard Apparatus) and then vacuum dried. The dried peptides were labeled with isobaric tags for relative and absolute quantitation (iTRAQ) 4-plex (ABSciex). The labeling process was performed according to Murillo et al. (2017). Briefly, dried peptides were dissolved in 50 mM TEAB and iTRAQ tags were dissolved in dry ethanol, labeling was performed by mixing peptide and tag solutions for 1 h at room temperature. The four tag reactions were combined in a single tube (ratio 1:1:1:1) and dried in a speed vac prior to fractionation. Each condition was labeled as follows: (i) Tag 114 corresponds to sample W (Wolbachia infected); (ii) Tag 115 to sample A (none infection); (iii) Tag 116 to sample WZ (Wolbachia and ZIKV co-infection); and (iv) Tag 117 to sample Z (ZIKV infection).
The iTRAQ labeled peptide mixture was fractionated off-line by HILIC (hydrophilic interaction liquid chromatography) before LC-MS/MS analysis. The dried samples were resuspended in acetonitrile (ACN) 90% / trifluoroacetic acid (TFA) 0.1% and injected into Shimadzu UFLC chromatography using a TSKGel Amide-80 column (15 cm × 2 mm i.d. × 3 μm – Supelco), flow rate 0.2 ml/min; mobile phases A (ACN 85%/TFA 0.1%) and B (TFA 0.1%); gradient: 0% of phase B in 5 min; 0–10% of phase B from 5 to 10 min; 10–20% of phase B from 10 to 55 min; 20–100% of phase B from 55 to 60 min; and 100–0% of phase B from 60 to 65 min. For each pooled sample, eight fractions were collected and combined according to the separation and intensity of the peaks. The pools of fractions were dried in a speed vac and resuspend in 0.1% formic acid (FA). The samples were analyzed in Easy-nLC 1000 coupled to a Q-Exactive Plus mass spectrometer (Thermo Fisher Scientific). For each fraction (or pool of fractions), a linear gradient of 5–40% of B em 105 min, 40–95% in 8 min and 95% B in 5 min was performed in 120 min; phase A (0.1% FA), B (ACN 95%, 0.1% FA). Ionization was performed in an electrospray source with the acquisition of spectra in positive polarity by data-dependent acquisition (DDA) mode, spray voltage of 2.5 kV, and temperature of 200°C in the heated capillary. The acquisition was set as follows: full scan or MS1 in the range of 375–1,800 m/z, resolution of 70,000 (m/z 200), fragmentation of the 10 most intense ions in the HCD collision cell, with standardized collision energy (NCE) of 30, resolution of 17,000 in the acquisition of MS/MS spectra, the first mass of 110 m/z, isolation window of 2.0 m/z and dynamic exclusion of 45 s.
The raw data were processed using Proteome Discoverer 2.4 Software (Thermo Fisher Scientific). Peptide identification was performed with the Sequest HT search engine against A. aegypti (genome version/assembly ID: INSDC: GCA_002204515.1), ZIKV, and Wolbachia databases provided by VectorBase (Giraldo-Calderón et al., 2015), ViPR (Pickett et al., 2012), and UniProt (The Uniprot Consortium, 2019), respectively. Searches were performed with peptide mass tolerance of 10 ppm, MS/MS tolerance of 0.1 Da, tryptic cleavage specificity, two maximum missed cleavage sites allowed, fixed modification of carbamidomethyl (Cys), variable modification of iTRAQ 4-plex (Tyr, Lys, and peptide N-terminus), phosphate (Ser, Thr, and Tyr), and oxidation (Met); peptides with high confidence were considered. False discovery rates (FDR) were obtained using Target Decoy PSM selecting identifications with a q-value equal or to less than 0.01. Data of all technical replicates were log2-transformed and normalized by subtracting the column median. Relative iTRAQ quantification analysis was carried out in the Perseus software, version 1.6.12.0 (Tyanova et al., 2016), based on the intensity of the fragmented reporter peaks in MS/MS.
For ZIKV peptides abundance comparison, statistical analysis was performed in experimental groups using T-test analysis (95% confidence values, p < 0.05), meanwhile for differential proteins was used one-way analysis of variance (ANOVA) and Tukey’s multiple comparisons post-test (95% confidence values, p < 0.05) in Graphpad Prism 8.0.0 for Windows, GraphPad Software, San Diego, CA, United States, www.graphpad.com. Biological processes enrichment for differentially expressed proteins were performed using Fisher’s Exact Test (p < 0.05) in VectorBase website (bit.ly/38OmEX0) (Ashburner et al., 2000; Giraldo-Calderón et al., 2015). The resulting Gene Ontology (GO) lists were summarized and represented in a network interaction format in REVIGO (Supek et al., 2011).
Proteomics has improved in recent years, especially considering its coverage and sensitivity, which offer more opportunities to study changes in the global proteome of complex tissues (Aebersold and Mann, 2016). Therefore, the use of isobaric-labeled quantitative proteomics measurements provides efficiency and increased depth coverage of proteomics analysis (Rauniyar and Yates, 2014). Peptides from the four experimental groups were labeled with iTRAQ 4-plex, enabling multiplexing of samples prior to off-line fractionation and LC-MS/MS analysis (Figure 1). We were able to identify 4,117 protein groups (Table 1), 26,898 peptides and 909,046 MS/MS spectra. The protein search was performed using A. aegypti, ZIKV, and Wolbachia databases simultaneously. Of all identified proteins, 3,790 belong to A. aegypti, 323 to Wolbachia and three unique peptides with a total of 10 PSMs to the ZIKV polyprotein (Figure 2 and Supplementary Figures 1, 2).
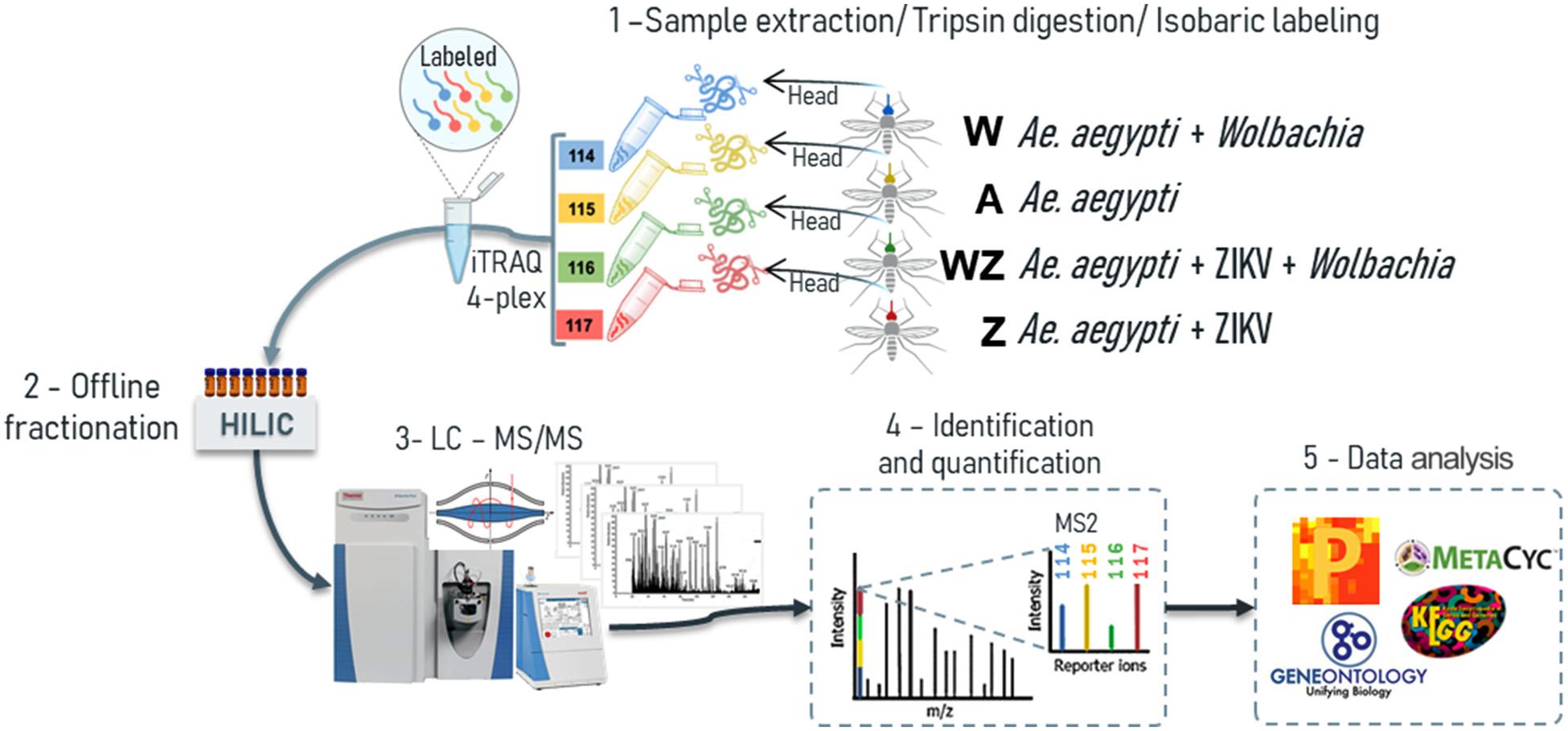
Figure 1. The proteomics workflow carried out in the head + salivary gland of A. aegypti females used samples from four different conditions: A, non-infected mosquitoes; W, Wolbachia infected; Z, ZIKV infected; and WZ, Wolbachia and ZIKV infection. 1 – The samples were subjected to lysis and, subsequently, proteins were reduced, alkylated, and digested using trypsin. The resulting peptides were labeled with iTRAQ-4plex and combined in 1:1:1:1 ratio; 2 – labeled peptides were fractionated offline using HILIC chromatography; 3 – pool of fractions were analyzed by nLC–MS/MS in Q-Exactive Plus mass spectrometer; 4 – fragmentation of the attached tag release a low m/z reporter ions used for relative quantification of the peptides/proteins from which they originated; 5 – statistical analysis were performed in Perseus and bioinformatics analysis used Gene Ontology (GO) in the enrichment of proteins differentially regulated.

Table 1. Proteins with significative diferencial expression by ANOVA test (p < 0.05) and each respective pair analyzed by Tukey test.
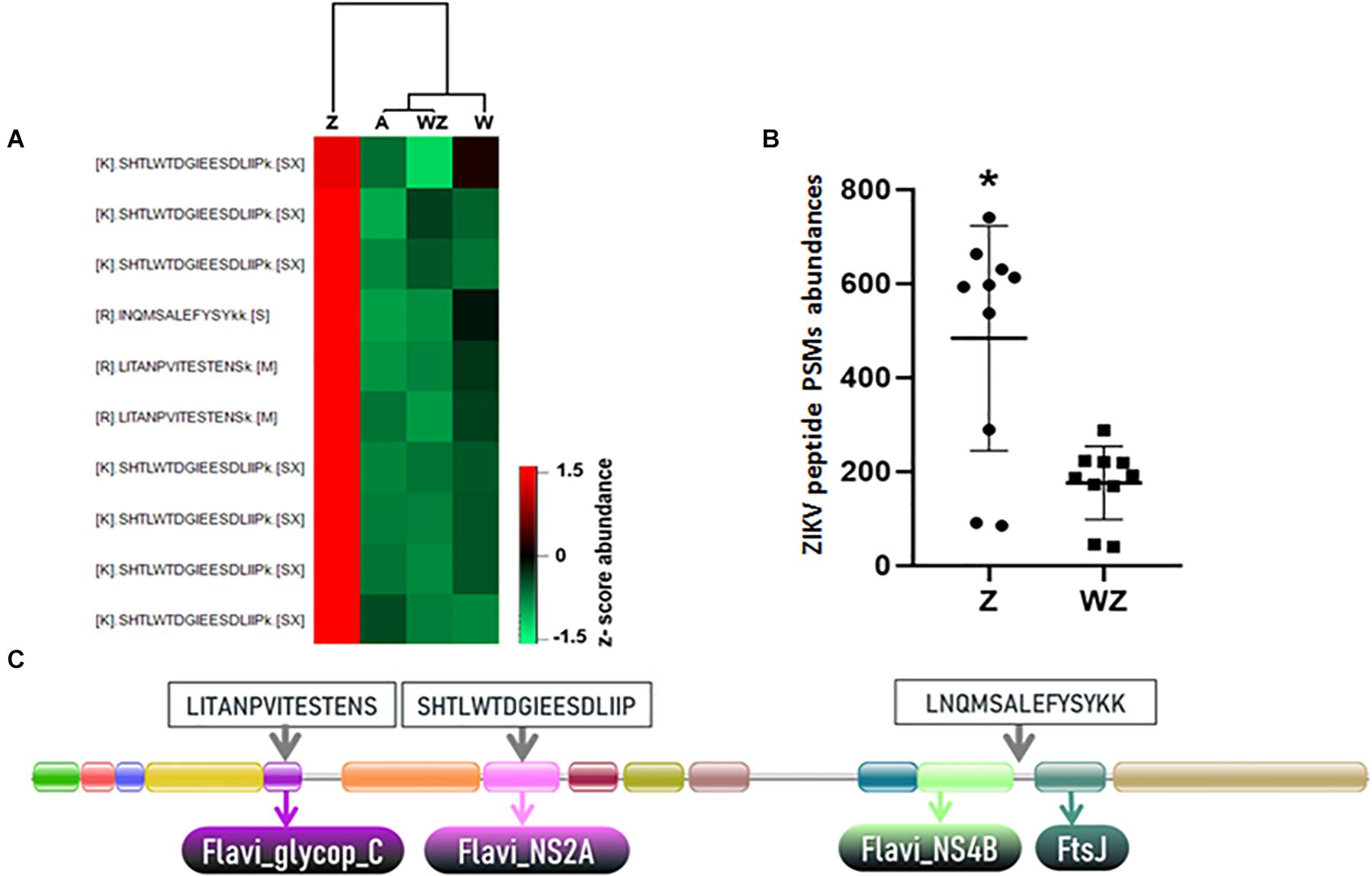
Figure 2. Identification and quantification of ZIKV peptides. (A) Heatmap of ZIKV PSMs identified in the A. aegypti head + salivary gland generated in Perseus software using abundance values transformed by z-score. (B) Total abundance of peptides spectrum matches (PSMs) calculated from report ions intensities from the three ZIKV peptides identified in samples. “*” Means that in Statistical analysis (T-test; *P-value 0.0011), developed by using the software GraphPad Prism 7.0, sample Z has a statistical difference to sample WZ. (C) Domain architecture of the complete ZIKV polyprotein obtained from the PFAM database (PF01570) (El-Gebali et al., 2018). The location within the conserved domains and amino acid sequence of the three unique ZIKV peptides identified in proteomics analysis are highlighted.
The quantitative proteomics analysis was performed by comparing the four experimental groups using the normalized iTRAQ reporter ion intensity of identified peptides. Replicates were highly correlated (Pearson correlation mean of 0.97) and normalization methods were applied (Supplementary Figure 3). The 196 proteins with significant differences were statistically determined by the ANOVA test. Using the Tukey post-test ANOVA, we defined pairs of proteins with significant differences between the groups (Supplementary Table 2). The comparisons between the samples were: single-infected mosquitoes versus non-infected mosquitoes (W versus A and Z versus A); coinfected versus non-infected mosquitoes (WZ versus A); coinfected versus single-infected mosquitoes (WZ versus W and WZ versus Z).
The quantitative analysis using the iTRAQ reporter ions from ZIKV peptides revealed that ZIKV proteins were more abundant in the absence of Wolbachia in contrast to mosquitoes coinfected with ZIKV and Wolbachia (Figure 2). This latter condition showed reporter ions intensity compared to the background signal of uninfected mosquitoes (Figure 2A). This result indicates Wolbachia presence likely reduces virus replication in the female head and salivary gland (Figure 2B). Noteworthy, the presence of the wMel strain in organs like salivary glands and ovaries of A. aegypti females induce the antiviral response that supports Wolbachia deployment to mitigate disease transmission.
The detection of ZIKV in the mosquito head + salivary gland was possible even considering that virus proteins were a minute fraction of all proteins identified. Peptides LITANPVITESTENS, SHTLWTDGIEESDLIIP, and LNQMSALEFYSYKK showed a robust increase in ZIKV infected tissue and this data represents a resource to further develop targeted proteomics methods to quantify ZIKV in infected mosquitoes (MS/MS spectra shown in Supplementary Figure 1).
During a blood meal, the female mosquitoes inject saliva into the host skin. The salivary glands produce a huge amount of molecules to avoid host responses against blood loss and to evade the host immune system (Ribeiro et al., 2016). For transmission of arboviroses, the virus must be secreted with the saliva during a blood meal in a vertebrate host (Linthicum et al., 1996). In this work, we took advantage of the sensitivity of protein detection by MS to detect and quantify proteins from the salivary gland, which are located on the anterior part of the thorax of A. aegypti. We compared the head proteome with two proteomes and one transcriptome (Wasinpiyamongkol et al., 2010; Ribeiro et al., 2016; Dhawan et al., 2017) from salivary glands of female mosquitoes to identify the salivary gland related proteins (Figure 3). We observed that among the 3,790 proteins identified in A. aegypti heads, 830 proteins were previously known to be expressed in the salivary glands. Moreover, we identified 77% of the proteins described by Wasinpiyamongkol et al. (2010) and 65% by Dhawan et al. (2017) in previous salivary gland proteome analysis. The 16 salivary proteins that were detected in all datasets probably are the most expressed and include inhibitors of platelet aggregation (salivary apyrase, AAEL006333-PA; apyrase precursor, AAEL006347PA), clotting inhibitors (SRPN23, AAEL002704-PB; SRPN26, AAEL003182-PA), aegyptins (AAEL010228-PA and AAEL010235-PA), angiopoietins (AAEL000749-PA and AAEL000726-PA), D7 family proteins (AAEL006417-PA and AAEL006424-PA), among others. We compared the 830 proteins previously known to be expressed in the salivary glands with the 196 differentially expressed proteins, to understand if these proteins could play a role during ZIKV and Wolbachia infections and/or co-infections. The modulation of saliva proteins is discussed below where the systemic changes in W, Z, and WZ will be addressed.
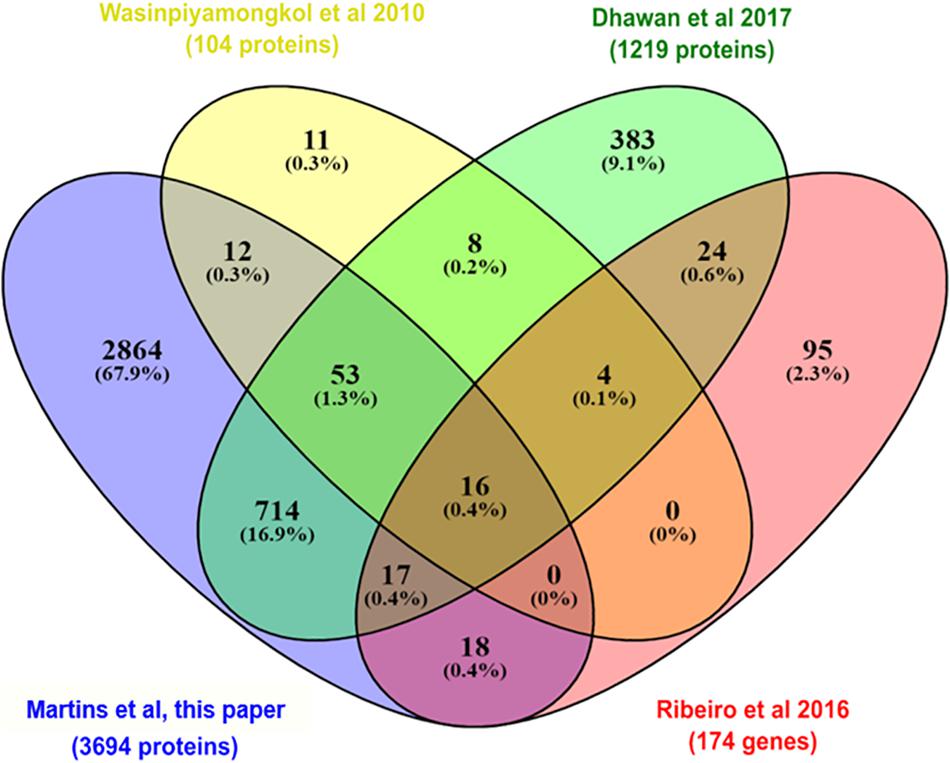
Figure 3. Venn diagram comparing the set of proteins (isoforms were not considered) identified in this work (purple) with two previous proteomes of the salivary glands of A. aegypti (yellow; Wasinpiyamongkol et al., 2010 and green; Dhawan et al., 2017) and one transcriptome (red; Ribeiro et al., 2016) also carried out in salivary glands of A. aegypti, but in non-infected females.
As a class of neuronal signal molecules, neuropeptides are produced by the neurosecretory cells, that are mainly located in the brain, suboesophageal ganglion, among others (Li et al., 2020). In insects, neuropeptides and their receptors play important physiological processes such as development, reproduction, behavior, and feeding (Holt et al., 2002). A previous study that investigated the presence of neuropeptidomics in A. aegypti proteome carried out in the central nervous system (CNS) and midgut samples identified 43 neuropeptides and hormones (Predel et al., 2010). Of these, we were able to identify 13 in our head proteomics analysis (Figure 4): pyrokinin (PK) (AAEL012060), short neuropeptide F (sNPF) (AAEL019691), neuropeptide-like precursor-1(NPLP) (AAEL012640), calcitonin-like diuretic hormone (DH-31) (AAEL008070), SIFamide (AAEL009858), crustacean cardioactive peptide (CCAP) (AAEL000630), allatotropin (AAEL009541), CAPA (AAEL005444), sulfakinin (SK) (AAEL006451), ovary ecdysteroidogenic hormone (OEH; neuroparsin A homolog) (AAEL004155), insulin-like peptide 1 and 7 (AAEL000937; AAEL024251), and adipokinetic hormone 1 (AKH) (AAEL011996). A few neuropeptides and hormones were modulated by co-infection and will be discussed afterward.
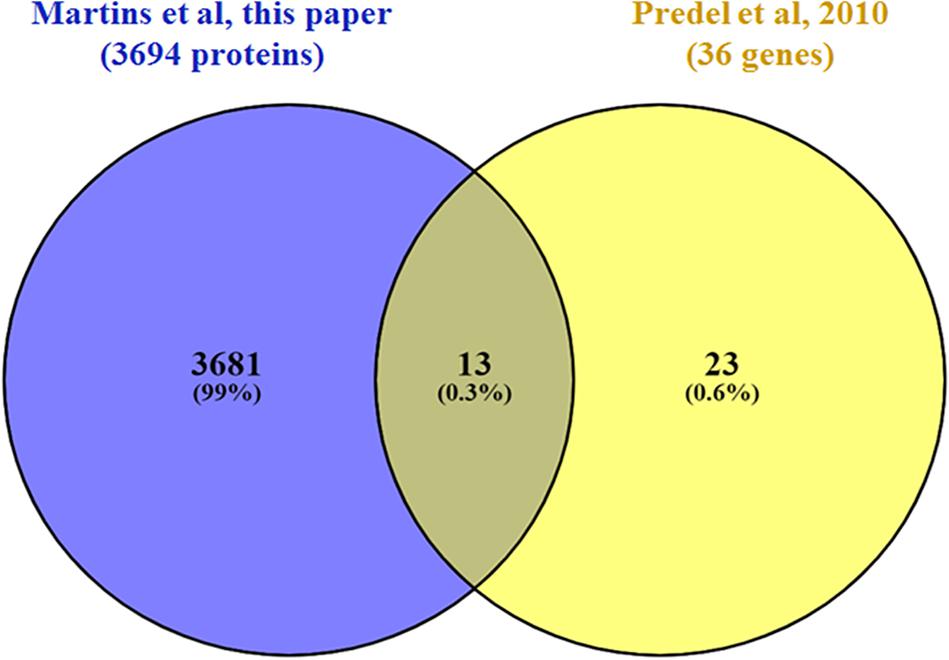
Figure 4. Venn diagram comparing the set of neuropeptides and hormones (isoforms were not considered) identified in this work (purple) with those identified in Predel (yellow; Predel et al., 2010).
By comparing mosquitoes infected and non-infected with ZIKV (Figure 5 and Supplementary Figure 4A), we observe four main enriched processes: nicotinamide nucleotide metabolism, lipid transport, humoral innate response, and carbohydrate catabolism. Nicotinamide mononucleotide (NMN) is known for being an intermediator in NAD+ biosynthesis (Poddar et al., 2019; Hong et al., 2020), which participates actively in the aerobic metabolism process. This enrichment is mediated by the phosphoglycerate kinase (AAEL004988) and transaldolase (AAEL009389) upregulation, leading to carbohydrate catabolism elevation. The pentose phosphate pathway (PPP) was also up-regulated in our study as a response to transaldolase (AAEL009389) protein. The PPP is known to generate NADPH and pentoses (5-carbon sugars) as well as ribose 5-phosphate, a precursor for the synthesis of nucleotides, this led us to relate the activation of PPP with the probation of nucleotide biosynthesis for virus replication (Guo et al., 2019). Both increased fluxes have been observed following Human Cytomegalovirus, Mayaro, and Kaposi-sarcoma viruses in human cells (El-Bacha and Da Poian, 2013).
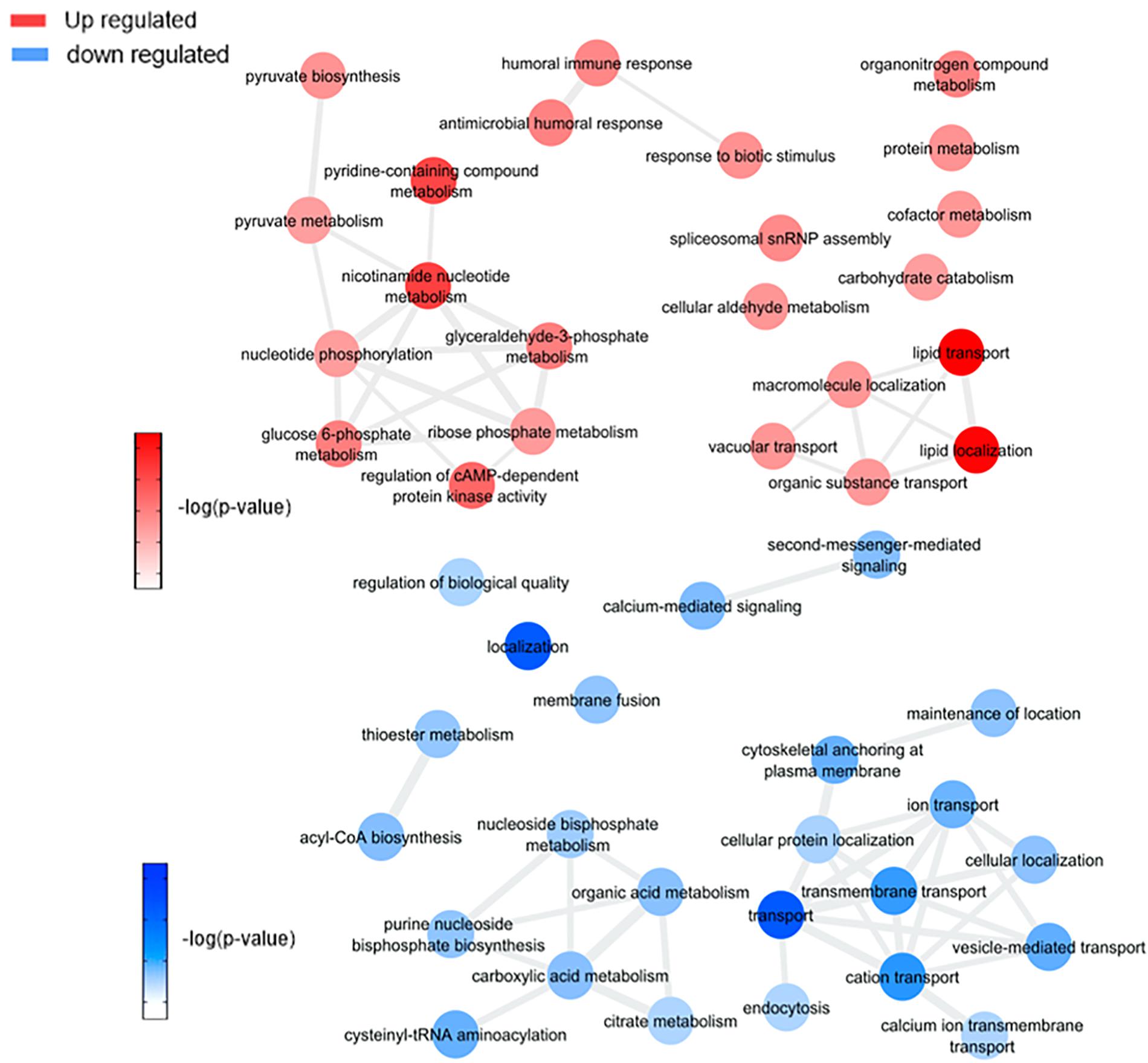
Figure 5. Overview of significantly enriched GO [–log10 (P-value)] terms of ontological interaction network in biological processes of up-regulated (red) and down-regulated (blue) genes in ZIKV infected versus non-infected mosquitoes.
Viruses are capable of modulating mitochondrial bioenergetics aiming to synthesize macromolecular precursors for self-replication, assembly and egress (El-Bacha and Da Poian, 2013). Thus, the enhancement of the lipid transport process should drive a redirection of these energetic resources to the viral capsids formation (Kuzmina et al., 2020), an event that was also described in A. aegypti coinfected with DENV and Wolbachia (Koh et al., 2020). Koh et al. (2020) compared lipid abundances between non-infected and DENV-3 infected mosquitoes and found that most modulated lipids are glycerophospholipids, as described earlier (Perera et al., 2012), with a higher level of unsaturated fatty acids and triacylglycerols. Glycerophospholipids modulation can be related to a virus attempt to build new virus capsids, as these lipids are a compound of this structure (Martín-Acebes et al., 2016). In our results, three genes related to lipid localization and transport were up-regulated: apolipophorin-III (AAEL008789) and two vitellogenin-A1 precursors (AAEL006126 and AAEL006138). Apolipophorins are protein components of lipoprotein particles that are essential for lipid transportation in the insect body, but apolipophorin-III can work as a pattern recognition receptor for insect immune system response (Wang et al., 2019), including in A. aegypti (Phillips and Clark, 2017). Lipid carrier protein lipophorin (Lp) and its lipophorin receptor (LpRfb) were significantly increased in A. aegypti after infections with Gram (+) bacteria and fungi, suggest that lipid metabolism is involved in the mosquito systemic immune responses to pathogens and parasites, as seen in studies before using fat body tissue (Cheon et al., 2006). The apolipophorin-III upregulation can be rolled to lipid transport and also to the immune response as ZIKV is expected to regulate them and the protein participates in both processes.
Another group of up-regulated biological processes includes an innate immune response in host cells due to virus infection (antimicrobial humoral response, humoral immune response, and response to biotic stimulus). Xi et al. (2008) found a significant role for the Toll pathway in DENV resistance regulation, and it was possible to identify differentially expressed antimicrobial peptide cecropin (AAEL029038) related to this pathway in our samples, confirming that Toll-pathway plays a major part in virus response (Xi et al., 2008; Pan et al., 2012; Kingsolver et al., 2013).
We observed down-regulation of tomosyn (AAEL006948), adenylate cyclase (AAEL009314), putative syntaxin-7 (AAEL009398), putative sodium/dicarboxylate cotransporter (AAEL009863), cytochrome c oxidase subunit II (AAEL018664), major facilitator superfamily domain-containing protein (AAEL006002), nuclear pore complex protein Nup188 (AAEL008557). and talin-1 (AAEL024235) related to cation transport processes. Also, three proteins were down-regulated: calcium-transporting ATPase sarcoplasmic/endoplasmic reticulum type (AAEL006582), Na+/K+ ATPase alpha subunit (AAEL012062), and calmodulin (AAEL012326-PA) related to cation transport processes as well. Similarly cytoskeleton, vesicle-mediated transport and ion transport-associated proteins down-regulation were described in CHIKV infection (Cui et al., 2020). Also, a recent study showed that Na+/K+ ATPase inhibition using two distinct drugs in mice helped to block ZIKV replication (Guo et al., 2020), suggesting that this down-regulation could be a mosquito cellular response to fight infection.
Adenosine triphosphate-citrate synthase (AAEL004297), sphingosine phosphate lyase (AAEL003188) and probable cysteine-tRNA synthetase (AAEL022886), proteins associated with amino acids insertion in the citric acid cycle, were also down-regulated and may lead to a decrease in citric acid cycle and oxidative phosphorylation processes. This phenomenon was also observed in A. aegypti salivary glands infected with DENV-2 (Chisenhall et al., 2014; Shrinet et al., 2018), suggesting the occurrence of cellular biomolecule and energy production control by virus infection to avoid mitochondrial exhaustion.
It is possible to notice in our data (Figure 6 and Supplementary Figures 3B, 4B) an enrichment of GDP-mannose metabolism comparing mosquitoes non-infected with Wolbachia-mono infected. The enrichment was based on the upregulation of gdp mannose-4,6-dehydratase (AAEL014961), which converts GDP-mannose to GDP-4-dehydro-6-deoxy-D-mannose, the first of three steps for the conversion of GDP-mannose to GDP-fucose. This result suggests an induction caused by the presence of Wolbachia in insect cells to generate glycoconjugates, as GDP-mannose is a precursor for these molecules (Stick and Williams, 2009; Rexer et al., 2020), that may be used as elements to form new bacterial structures (García-del Portillo, 2020). An important observation was the enrichment of processes downregulated related to the citric acid cycle in mosquito cells infected with Wolbachia. As shown by da Rocha Fernandes et al. (2014), Wolbachia modulated glycogen metabolism in Aedes fluviatilis embryos in favor of its aerobic metabolism (da Rocha Fernandes et al., 2014).
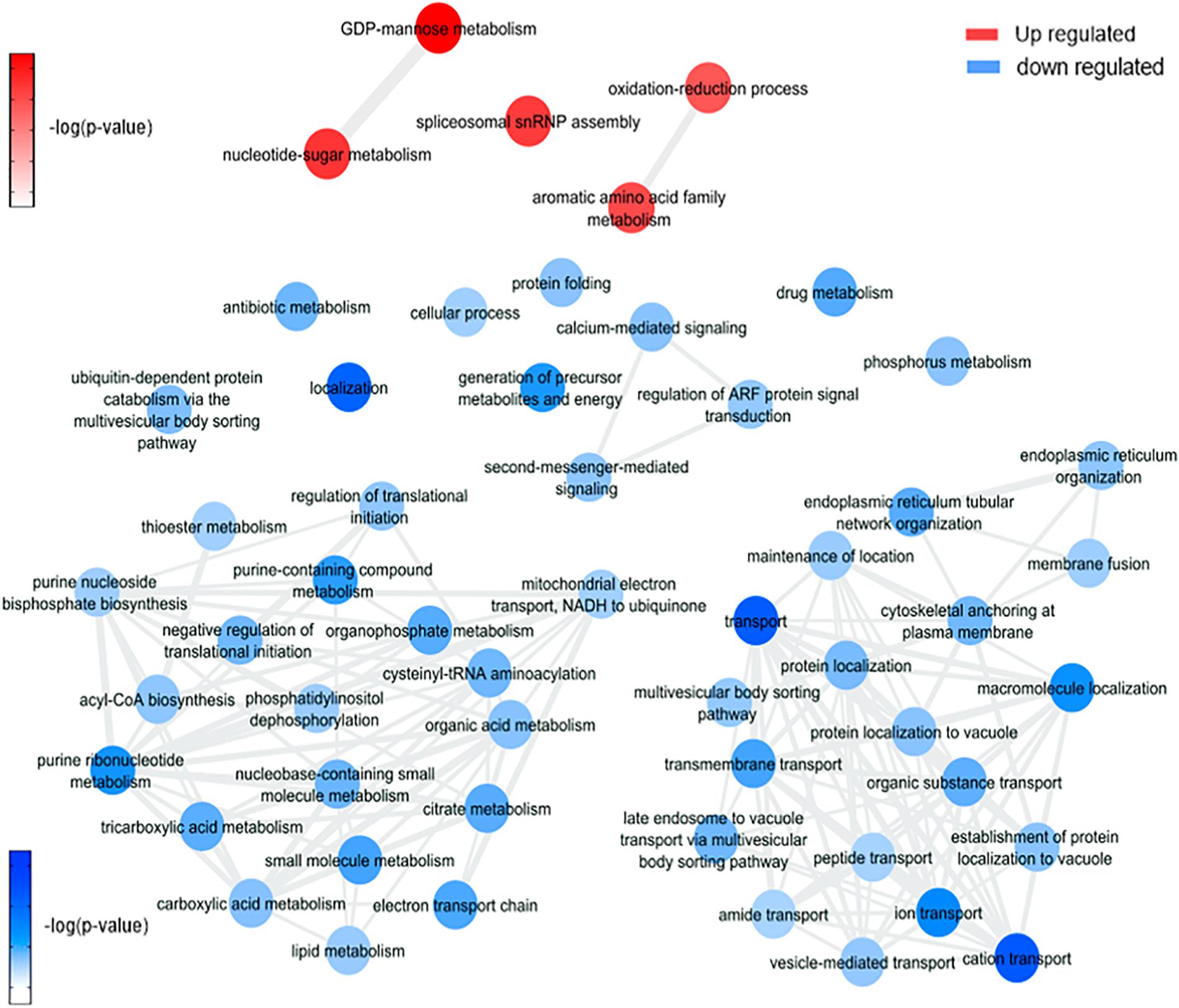
Figure 6. Overview of significantly enriched GO [–log10 (P-value)] terms of ontological interaction network in biological processes of up-regulated (red) and down-regulated (blue) genes in Wolbachia infected versus non-infected mosquitoes.
Up-regulation of 4-hydroxyphenylpyruvate dioxygenase (AAEL014600) can be related to plastoquinol and vitamin E (tocopherol) synthesis. Both molecules are described in the literature as participants of antioxidant response to different stimuli in organisms (MaitiDutta et al., 2020; Nowicka et al., 2020), including oxidative stress (Kumar et al., 2020; Minter et al., 2020). In this way, we consider that genes correlated to these pathways were regulated during Wolbachia infection because this microorganism can induce the expression of antioxidant proteins (Brennan et al., 2008). Moreover, 4-hydroxyphenylpyruvate dioxygenase is related to L-tyrosine degradation I and L-phenylalanine degradation IV, which are essential traits for blood-feeding arthropods, as they consume an amount of blood proteins and need to deal with a high quantity of free amino acids (Sterkel et al., 2016).
The Wolbachia infection up-regulated 10 salivary gland proteins, such as endoplasmin (AAEL012827-PA), which is required for proper folding of Toll-like receptors in mammals (Fitzgerald and Kagan, 2020), and FKBP12 (AAEL010491-PB) that has a prolyl isomerase activity and bidins to immunosuppressive molecules (Caminati and Procacci, 2020). Twenty salivary proteins were down-regulated by Wolbachia infection. Interestingly, the same ion related proteins down-regulated by ZIKV are also down-regulated by Wolbachia, moreover, proteins related with contraction such as paramyosin (AAEL010975-PA), tropomyosin (AAEL002761-PA), and Actin-1 (AAEL001928-PA) were also down-regulated. Also, a serine protease inhibitor 25 (SRPN25) (AAEL007420-PB) was also identified in our analysis. Its orthologue is Alboserpin, the major salivary gland anticoagulant in Aedes albopictus, which prevents coagulation in an atypical reversible interaction with factor Xa (coagulation activation factor), important for blood intake by female mosquitoes (Stark and James, 1998; Calvo et al., 2011). SRPN25 was up-regulated in W, which may infer that blood takes longer to coagulate, increasing mosquito enzymes access to digest blood. The consequence of these salivary proteins modulation suggests an impairment in salivary gland injection into the host and a possible reduction in the amount of blood ingestion during Wolbachia presence in the mosquito (Turley et al., 2009).
This analysis was based on a comparison of the ZW with Z (Figure 7 and Supplementary Figure 4C) and W (Figure 8 and Supplementary Figure 4D) mosquito heads samples.
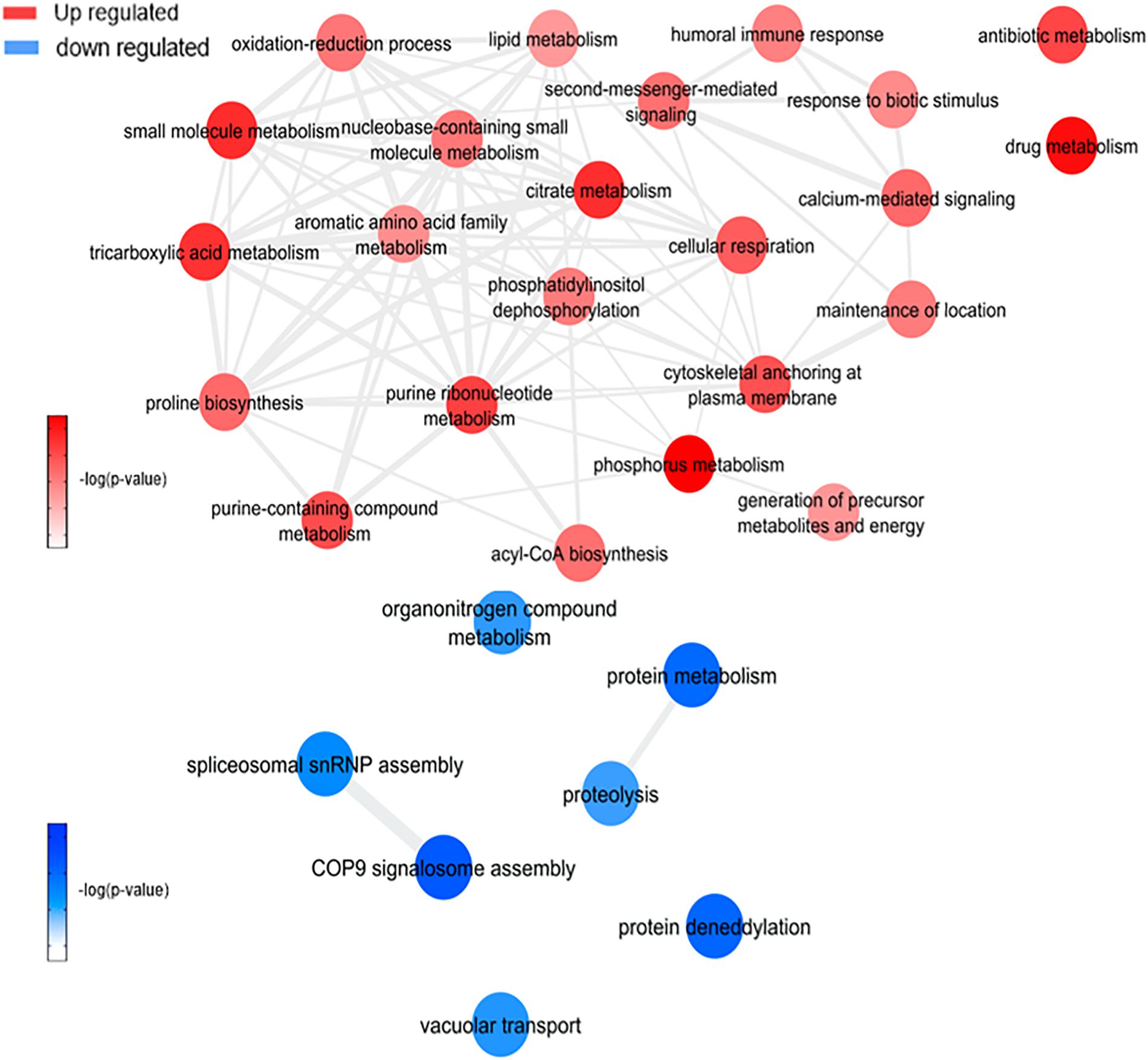
Figure 7. Overview of significantly enriched GO [–log10 (P-value)] terms of ontological interaction network in biological processes of up-regulated (red) and down-regulated (blue) genes in coinfected mosquitoes versus ZIKV infected mosquitoes.
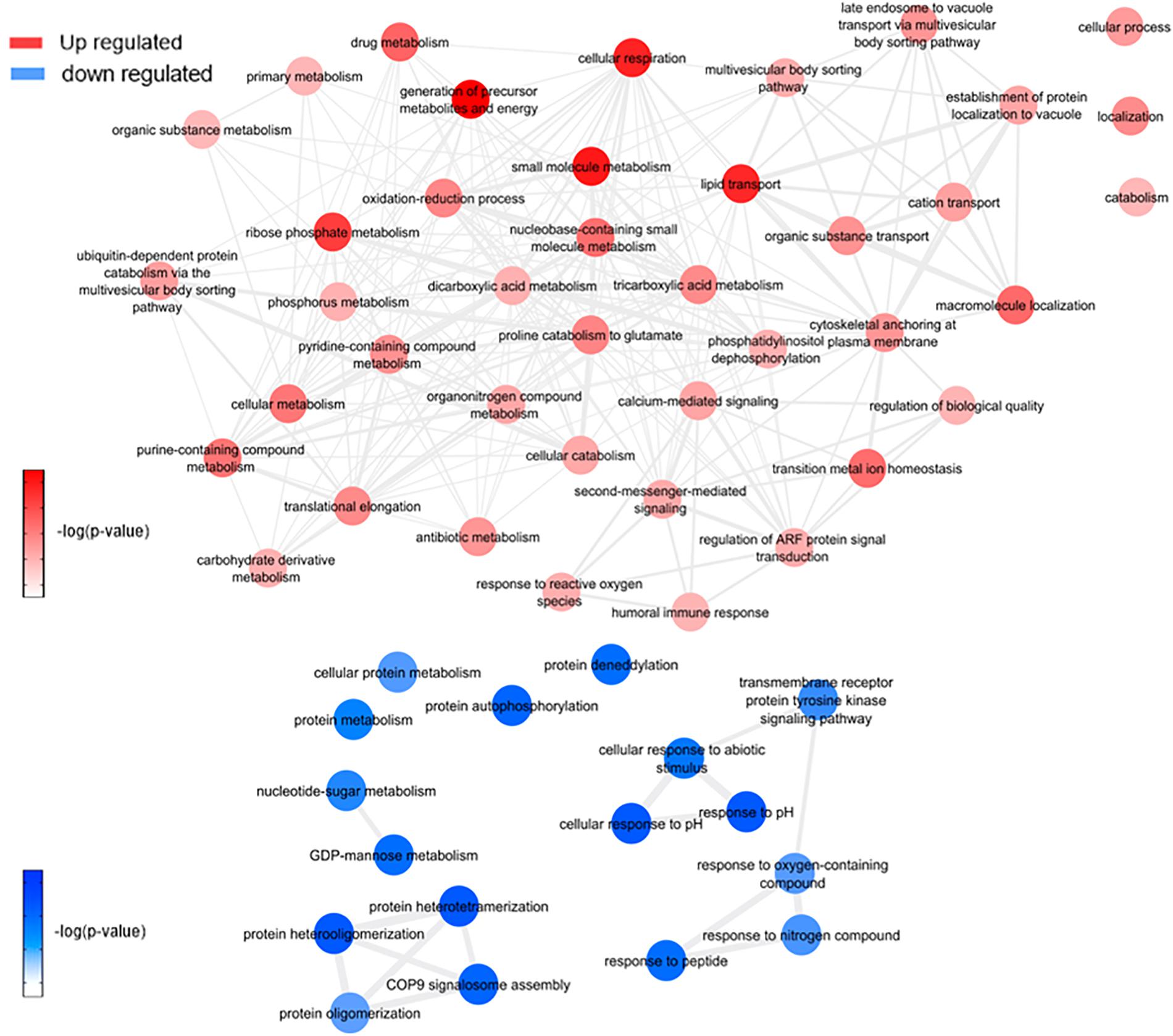
Figure 8. Overview of significantly enriched GO [–log10 (P-value)] terms of ontological interaction network in biological processes of up-regulated (red) and down-regulated (blue) genes in coinfected mosquitoes versus Wolbachia infected mosquitoes.
Zika virus and Wolbachia use mosquitoes cell machinery respectively to synthesize lipids and glycoconjugates molecules for their structural use, as mentioned before. According to WZ results, the enriched lipid synthesis biological processes remain up-regulated despite the Wolbachia presence, similarly to ZIKV infection. Koh et al. (2020) confirmed such behavior during co-infection-induced intracellular events with DENV and Wolbachia, suggesting that they do not compete by lipids in host cell and virus-driven modulation dominates over that of Wolbachia (Koh et al., 2020), which may have occurred as well as in our experiment. In contrast, there is a down-regulation in GDP-mannose proteins genes synthesis, which may suggest that host cells no longer produce glycoconjugates for bacteria use.
Within the up-regulated genes in WZ compared to Z, ATP synthesis (ATPase subunit O; AAEL010823-PA), and cytoskeleton dynamics like microtubule binding and associated-proteins (AAEL009375-PK, AAEL004176-PB, and AAEL009847-PB) suggest activation of the ubiquitin-proteasome system. Indeed, it was previously reported in Drosophila and mosquito cell lines transfected, that Wolbachia relies on host proteolysis through ubiquitination to acquire essential amino acid for the infection (Fallon and Witthuhn, 2009; White et al., 2017). Flavivirus also need a functioning ubiquitin-proteasome system to complete their life cycle (Choy et al.,2015a,b), however, Ub3881 (AAEL003881), a ubiquitin-protein, is highly down-regulated in mosquitoes infected with DENV and its overexpression was able to control the virus infection (Troupin et al., 2016). In our analysis, we identified a paralog of the Ub3881 ubiquitin protein (AAEL003888), which was up-regulated in WZ versus Z and down-regulated in Z, suggesting that Wolbachia have a role in AAEL003888 expression enhancement which might target the envelope proteins of ZIKV for degradation, therefore hindering the virus assembly.
Exploring neuropeptides and hormones, neuropeptide sulfakinin (SK) (AAEL006451-PB) and insulin-like peptide (ILP) 1 (AAEL000937-PA) were down-regulated comparing WZ with Z. SK are a family of neuropeptides, homologous to mammalian gastrin/cholecystokinin (CCK), responsible for food uptake control in insects (Zels et al., 2015) while ILPs are known for signaling carbohydrates intake in cells (Nässel and Broeck, 2016). A recent study correlated SK influence in ILPs concentration, where a rise in SK concentration promotes ILPs production (Słocińska et al., 2020). SK down-regulation, observed in our data, may decrease in mosquito feeding satisfaction. This fact allied to ILP down-regulation, leading to a decrease in glycolysis cell assimilation, can be understood as a virus-infection influence to increase mosquito blood uptake, elevating ZIKV transmission success.
There are several processes up-regulated after enrichment comparison between WZ and Z. Cellular respiration, aerobic respiration, small molecule metabolic process, tricarboxylic acid cycle, oxidative phosphorylation, ATP synthesis coupled electron transport, respiratory electron transport chain, generation of precursor metabolites and energy, tricarboxylic acid metabolic process, nicotinamide nucleotide metabolic process, and cellular process were enriched up-regulated biological process by proteins genes ATP-citrate synthase (AAEL004297), glutamate semialdehyde dehydrogenase (AAEL006834), ATP synthase delta chain (AAEL010823), phosphatidylinositol-4,5-bisphosphate 4-phosphatase (AAEL011216), NADH-ubiquinone oxidoreductase (AAEL012552), mitochondrial aconitase (AAEL012897), 4-hydroxyphenylpyruvate dioxygenase (AAEL014600), titin (AAEL002565), and myosin (AAEL000596) up-regulation. The increase in aerobic respiration metabolism in Wolbachia infection proved to stimulate ROS production in insect cells as part of the host immune response, but it counterbalances with antioxidant pathways activation (Zug and Hammerstein, 2015). However, it was observed in WZ that the aerobic metabolism increases together with transition metal ions biological process and pathways, mediated by putative secreted ferritin G subunit precursor (AAEL007383) and transferrin (AAEL015458), which can also generate ROS (Liu et al., 2016). ROS likely activates the Toll-immune pathway to confront ZIKV, similar to DENV infection (Selivanov et al., 2008). Also, two eukaryotic elongation factor 1-alpha (eEF1A) proteins (AAEL017096; AAEL017301) were up-regulated comparing WZ and Z. eEF1A increase was described as a response to endoplasmic reticulum stress caused by ROS in Chinese hamster ovary (Borradaile et al., 2005), which may have occurred in the mosquito. Another observed data was SRPN25 down-regulated in WZ, which was up-regulated in W. SRPN25 down-regulation in WZ highlights its function related to the insect immune system. Serpins can block the Toll signaling cascade and β-1,3-glucan-mediated melanin biosynthesis (Jiang et al., 2009), so its downregulation can infer that the insect immune system is activated.
This led us to understand insects defense mechanisms, induced by the presence of endosymbiotic bacteria, that may have been acting against virus infection: melanization activation mediated by SRPN25 down-regulation and Toll-pathway with the influence of SPRN25 down-regulation and the exacerbated production of ROS by aerobic metabolism increase. ROS occurs inside the mitochondria, after the oxidative phosphorylation stage, which has oxygen as the final electron acceptor, forming metabolic water. When oxygen is prematurely and incompletely reduced, it gives rise to superoxide anion, classified as ROS, together with hydrogen peroxide, hypochlorous acid, hydroxyl radical, singlet oxygen, and ozone (Lambert and Brand, 2009). Pan et al. described the relation between Wolbachia presence in A. aegypti and Toll-pathway activation by ROS to control DENV (Pan et al., 2012). This study not only confirms oxidative stress caused by Wolbachia, but also antioxidant genes expression as described earlier in this article. The role of symbiotic microorganisms in arbovirus infection of arthropods vectors is widely discussed, highlighting the importance of a ROS-mediated stimulation of the Toll-pathway and AMPs expression (Yin et al., 2020). As SPRN25 is down-regulated, serpins activity is up-regulated, which lead to serpins key response in the defense mechanism of insects, activating especially the Toll pathway, as well as ROS, and prophenoloxidase (PPO) cascade (Meekins et al., 2017). As for PPO cascade, it mediates melanization immune response in insects, including mosquito vectors (Christensen et al., 2005), It was already investigated that melanization suppression by SRPN activation can lead to a increase in viral infection (Toufeeq et al., 2019), calling attention to SRPN importance in mosquito immune response.
Proteomics analysis of female’s head of A. aegypti during mono-infection with ZIKV or Wolbachia highlighted how those microorganisms use cell host for their benefits. Our data support that ZIKV may induces lipid synthesis and transport allied with glycolysis pathways while Wolbachia increases glycoconjugates production. During co-infection, Wolbachia probably helps A. aegypti to prevent virus infection by stimulating ROS production leading to Toll-pathway humoral immune response, together with antioxidant production to control cell homeostasis. This mechanism seems to be efficient since we have shown that peptides coming from the ZIKV polyprotein are reduced in female’s heads of A. aegypti in the presence of Wolbachia. Our study has provided important insights into the differential manner in which A. aegypti proteome is systemically regulated during mono- and co-infections of ZIKV and Wolbachia. Ultimately, this work represents a rich resource for the insect research community to help elucidate the mechanisms by which Wolbachia orchestrate resistance to ZIKV infection in A. aegypti and it allows the generation of specific hypothesis-driven experiments in future studies.
The data presented in the study are deposited in the Proteome X Change repository, accession number PXD022665.
MM, LR, JM, AT, and SC performed the experiments and data analysis. MM and SC produced manuscript figures. MM, LR, JM, AT, SC, GD, DO, RM, FN, RM-D-F, and MJ wrote the manuscript. RM-D-F and MJ idealized and coordinated the study. All authors approved the manuscript.
The present work was supported by the Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ) and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The authors thank Fabio Mendonça Gomes from Instituto de Biofísica Carlos Chagas Filho – UFRJ and Livia Goto-Silva from D’Or Institute for Research and Education (IDOR) for their critical reading of the manuscript.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fphys.2021.642237/full#supplementary-material
Aebersold, R., and Mann, M. (2016). Mass-spectrometric exploration of proteome structure and function. Nature 537, 347–355. doi: 10.1038/nature19949
Aliota, M. T., Walker, E. C., Yepes, A. U., Velez, I. D., Christensen, B. M., and Osorio, J. E. (2016). The wMel strain of Wolbachia reduces transmission of Chikungunya virus in Aedes aegypti. PLoS Negl. Trop. Dis. 10:e0004677. doi: 10.1371/journal.pntd.0004677
Ashburner, M., Ball, C. A., Blake, J. A., Botstein, D., Butler, H., Cherry, J. M., et al. (2000). Gene ontology: tool for the unification of biology. Nat. Genet. 25, 25–29. doi: 10.1038/75556
Barreto, M. L., Barral-Netto, M., Stabeli, R., Almeida-Filho, N., Vasconcelos, P. F. C., Teixeira, M., et al. (2016). Zika virus and microcephaly in Brazil: a scientific agenda. Lancet 387, 919–921. doi: 10.1016/S0140-6736(16)00545-6
Baud, D., Gubler, D. J., Schaub, B., Lanteri, M. C., and Musso, D. (2017). An update on Zika virus infection. Lancet 390, 2099–2109. doi: 10.1016/S0140-6736(17)31450-2
Borradaile, N. M., Buhman, K. K., Listenberger, L. L., Magee, C. J., Morimoto, E. T. A., Ory, D. S., et al. (2005). A critical role for eukaryotic elongation factor 1A-1 in lipotoxic cell death. MBoC 17, 770–778. doi: 10.1091/mbc.e05-08-0742
Brennan, L. J., Keddie, B. A., Braig, H. R., and Harris, H. L. (2008). The endosymbiont Wolbachia pipientis induces the expression of host antioxidant proteins in an Aedes albopictus cell line. PLoS One 3:e2083. doi: 10.1371/journal.pone.0002083
Calvo, E., Mizurini, D. M., Sá-Nunes, A., Ribeiro, J. M. C., Andersen, J. F., Mans, B. J., et al. (2011). Alboserpin, a factor Xa inhibitor from the mosquito vector of yellow fever, binds heparin and membrane phospholipids and exhibits antithrombotic activity. J. Biol. Chem. 286, 27998–28010. doi: 10.1074/jbc.M111.247924
Caminati, G., and Procacci, P. (2020). Mounting evidence of FKBP12 implication in neurodegeneration. Neural Regen. Res. 15, 2195–2202. doi: 10.4103/1673-5374.284980
Campos, G., Bandeira, A., and Sardi, S. (2015). Zika virus outbreak, Bahia, Brazil. Emerg. Infect. Dis. J. 21:1885. doi: 10.3201/eid2110.150847
Cheon, H.-M., Shin, S. W., Bian, G., Park, J.-H., and Raikhel, A. S. (2006). Regulation of lipid metabolism genes, lipid carrier protein lipophorin, and its receptor during immune challenge in the mosquito Aedes aegypti. J. Biol. Chem. 281, 8426–8435. doi: 10.1074/jbc.M510957200
Chisenhall, D. M., Londono, B. L., Christofferson, R. C., McCracken, M. K., and Mores, C. N. (2014). Effect of dengue-2 virus infection on protein expression in the salivary glands of Aedes aegypti mosquitoes. Am. J. Trop. Med. Hyg. 90, 431–437. doi: 10.4269/ajtmh.13-0412
Choudhary, C., and Mann, M. (2010). Decoding signalling networks by mass spectrometry-based proteomics. Nat. Rev. Mol. Cell Biol. 11, 427–439. doi: 10.1038/nrm2900
Chouin-Carneiro, T., Vega-Rua, A., Vazeille, M., Yebakima, A., Girod, R., Goindin, D., et al. (2016). Differential susceptibilities of Aedes aegypti and Aedes albopictus from the Americas to Zika Virus. PLoS Negl. Trop. Dis. 10:e0004543. doi: 10.1371/journal.pntd.0004543
Choy, M. M., Sessions, O. M., Gubler, D. J., and Ooi, E. E. (2015a). Production of infectious dengue virus in Aedes aegypti is dependent on the ubiquitin proteasome pathway. PLoS Negl. Trop. Dis. 9:e0004227. doi: 10.1371/journal.pntd.0004227
Choy, M. M., Zhang, S. L., Costa, V. V., Tan, H. C., Horrevorts, S., and Ooi, E. E. (2015b). Proteasome inhibition suppresses dengue virus egress in antibody dependent infection. PLoS Negl. Trop. Dis. 9:e0004058. doi: 10.1371/journal.pntd.0004058
Christensen, B. M., Li, J., Chen, C.-C., and Nappi, A. J. (2005). Melanization immune responses in mosquito vectors. Trends Parasitol. 21, 192–199. doi: 10.1016/j.pt.2005.02.007
Codeço, C. T., Lima, A. W. S., Araújo, S. C., Lima, J. B. P., Maciel-de-Freitas, R., Honório, N. A., et al. (2015). Surveillance of Aedes aegypti: comparison of house index with four alternative traps. PLoS Negl. Trop. Dis. 9:e0003475. doi: 10.1371/journal.pntd.0003475
Cui, Y., Liu, P., Mooney, B. P., and Franz, A. W. E. (2020). Quantitative proteomic analysis of Chikungunya virus-infected Aedes aegypti reveals proteome modulations indicative of persistent infection. J. Proteome Res. 19, 2443–2456. doi: 10.1021/acs.jproteome.0c00173
da Rocha Fernandes, M., Martins, R., Pessoa Costa, E., Pacidônio, E. C., and Araujo et al. (2014). The modulation of the symbiont/host interaction between Wolbachia pipientis and Aedes fluviatilis embryos by glycogen metabolism. PLoS One 9:e98966. doi: 10.1371/journal.pone.0098966
da Silveira, I. D., Petersen, M. T., Sylvestre, G., Garcia, G. A., David, M. R., Pavan, M. G., et al. (2018). Zika virus infection produces a reduction on Aedes aegypti lifespan but no effects on mosquito fecundity and oviposition success. Front. Microbiol. 9:3011. doi: 10.3389/fmicb.2018.03011
De Mandal, S., Lin, B., Shi, M., Li, Y., Xu, X., and Jin, F. (2020). iTRAQ-based comparative proteomic analysis of larval midgut from the beet armyworm, Spodoptera exigua (Hübner) (Lepidoptera: Noctuidae) challenged with the entomopathogenic bacteria Serratia marcescens. Front. Physiol. 11:442. doi: 10.3389/fphys.2020.00442
Dhawan, R., Mohanty, A. K., Kumar, M., Dey, G., Advani, J., Prasad, T. S. K., et al. (2017). Data from salivary gland proteome analysis of female Aedes aegypti Linn. Data Brief. 13, 274–277. doi: 10.1016/j.dib.2017.05.034
Dick, G. W. A., Kitchen, S. F., and Haddow, A. J. (1952). Zika Virus (I). Isolations and serological specificity. Trans. R. Soc. Trop. Med. Hyg. 46, 509–520. doi: 10.1016/0035-9203(52)90042-4
Duncan, A. B., Agnew, P., Noel, V., Demettre, E., Seveno, M., Brizard, J.-P., et al. (2012). Proteome of Aedes aegypti in response to infection and coinfection with microsporidian parasites. Ecol. Evol. 2, 681–694. doi: 10.1002/ece3.199
Dutra, H. L. C., Rocha, M. N., Dias, F. B. S., Mansur, S. B., Caragata, E. P., and Moreira, L. A. (2016). Wolbachia blocks currently circulating Zika virus isolates in Brazilian Aedes aegypti mosquitoes. Cell Host Microbe 19, 771–774. doi: 10.1016/j.chom.2016.04.021
El-Bacha, T., and Da Poian, A. T. (2013). Virus-induced changes in mitochondrial bioenergetics as potential targets for therapy. Int. J. Biochem. Cell Biol. 45, 41–46. doi: 10.1016/j.biocel.2012.09.021
El-Gebali, S., Mistry, J., Bateman, A., Eddy, S. R., Luciani, A., Potter, S. C., et al. (2018). The Pfam protein families database in 2019. Nucleic Acids Res. 47, 427–432. doi: 10.1093/nar/gky995
Fallon, A. M., and Witthuhn, B. A. (2009). Proteasome activity in a naïve mosquito cell line infected with Wolbachia pipientis wAlbB. Vitro Cell. Dev. Biol. Anim. 45, 460–466. doi: 10.1007/s11626-009-9193-6
Fernandes, R. S., Campos, S. S., Ferreira-de-Brito, A., Miranda, R. M., Barbosa, da Silva, K. A., et al. (2016). Culex quinquefasciatus from Rio de janeiro is not competent to transmit the local Zika Virus. PLoS Negl. Trop. Dis. 10:e0004993. doi: 10.1371/journal.pntd.0004993
Ferreira-de-Brito, A., Ribeiro, I. P., Miranda, R. M., Fernandes, R. S., Campos, S. S., Silva, K. A., et al. (2016). First detection of natural infection of Aedes aegypti with Zika virus in Brazil and throughout South America. Memórias Instit. Oswaldo Cruz 111, 655–658. doi: 10.1590/0074-02760160332
Fitzgerald, K. A., and Kagan, J. C. (2020). Toll-like receptors and the control of immunity. Cell 180, 1044–1066. doi: 10.1016/j.cell.2020.02.041
Ford, S. A., Albert, I., Allen, S. L., Chenoweth, S. F., Jones, M., Koh, C., et al. (2020). Artificial selection finds new hypotheses for the mechanism of Wolbachia-Mediated dengue blocking in mosquitoes. Front. Microbiol. 11:1456. doi: 10.3389/fmicb.2020.01456
Garcia, G. A., Sylvestre, G., Aguiar, R., da Costa, G. B., Martins, A. J., Lima, J. B. P., et al. (2019). Matching the genetics of released and local Aedes aegypti populations is critical to assure Wolbachia invasion. PLoS Negl. Trop. Dis. 13:e0007023. doi: 10.1371/journal.pntd.0007023
García-Robles, I., De Loma, J., Capilla, M., Roger, I., Boix-Montesinos, P., Carrión, P., et al. (2020). Proteomic insights into the immune response of the Colorado potato beetle larvae challenged with Bacillus thuringiensis. Dev. Comp. Immunol. 104:103525. doi: 10.1016/j.dci.2019.103525
García-del Portillo, F. (2020). Building peptidoglycan inside eukaryotic cells: a view from symbiotic and pathogenic bacteria. Mol. Microbiol. 113, 613–626. doi: 10.1111/mmi.14452
Geoghegan, V., Stainton, K., Rainey, S. M., Ant, T. H., Dowle, A. A., Larson, T., et al. (2017). Perturbed cholesterol and vesicular trafficking associated with dengue blocking in Wolbachia -infected Aedes aegypti cells. Nat. Commun. 8:526. doi: 10.1038/s41467-017-00610-8
Giraldo-Calderón, G. I., Emrich, S. J., MacCallum, R. M., Maslen, G., Dialynas, E., Topalis, P., et al. (2015). VectorBase: an updated bioinformatics resource for invertebrate vectors and other organisms related with human diseases. Nucleic Acids Res. 43, 707–713. doi: 10.1093/nar/gku1117
Guo, J., Jia, X., Liu, Y., Wang, S., Cao, J., Zhang, B., et al. (2020). Inhibition of Na + /K + ATPase blocks Zika virus infection in mice. Commun. Biol. 3, 380. doi: 10.1038/s42003-020-1109-8
Guo, X., Wu, S., Li, N., Lin, Q., Liu, L., Liang, H., et al. (2019). Accelerated metabolite levels of aerobic glycolysis and the pentose phosphate pathway are required for efficient replication of infectious spleen and kidney necrosis virus in chinese perch brain cells. Biomolecules 9:440. doi: 10.3390/biom9090440
Haqshenas, G., Terradas, G., Paradkar, P. N., Duchemin, J.-B., McGraw, E. A., and Doerig, C. (2019). A role for the insulin receptor in the suppression of dengue virus and Zika virus in wolbachia-infected mosquito cells. Cell Rep. 26, 529.e3–535.e3. doi: 10.1016/j.celrep.2018.12.068
Hoffmann, A. A., Montgomery, B. L., Popovici, J., Iturbe-Ormaetxe, I., Johnson, P. H., Muzzi, F., et al. (2011). Successful establishment of Wolbachia in Aedes populations to suppress dengue transmission. Nature 476, 454–457. doi: 10.1038/nature10356
Holt, R. A., Subramanian, G. M., Halpern, A., Sutton, G. G., Charlab, R., Nusskern, D. R., et al. (2002). The genome sequence of the malaria mosquito Anopheles gambiae. Science 298, 129–149. doi: 10.1126/science.1076181
Hong, W., Mo, F., Zhang, Z., Huang, M., and Wei, X. (2020). Nicotinamide mononucleotide: a promising molecule for therapy of diverse diseases by targeting NAD+ metabolism. Front. Cell Dev. Biol. 8:246. doi: 10.3389/fcell.2020.00246
Indriani, C., Tantowijoyo, W., Rancès, E., Andari, B., Prabowo, E., Yusdi, D., et al. (2020). Reduced dengue incidence following deployments of Wolbachia-infected Aedes aegypti in Yogyakarta, Indonesia: a quasi-experimental trial using controlled interrupted time series analysis. Gates Open Res. 4:50. doi: 10.12688/gatesopenres.13122.1
Jiang, R., Kim, E.-H., Gong, J.-H., Kwon, H.-M., Kim, C.-H., Ryu, K.-H., et al. (2009). Three pairs of protease-serpin complexes cooperatively regulate the insect innate immune responses. J. Biol. Chem. 284, 35652–35658. doi: 10.1074/jbc.M109.071001
Kingsolver, M. B., Huang, Z., and Hardy, R. W. (2013). Insect antiviral innate immunity: pathways, effectors, and connections. J. Mol. Biol. 425, 4921–4936. doi: 10.1016/j.jmb.2013.10.006
Koh, C., Islam, M. N., Ye, Y. H., Chotiwan, N., Graham, B., Belisle, J. T., et al. (2020). Dengue virus dominates lipid metabolism modulations in Wolbachia -coinfected Aedes aegypti. Commun. Biol. 3:e0007443. doi: 10.1038/s42003-020-01254-z
Kumar, A., Prasad, A., Sedlářová, M., Ksas, B., Havaux, M., and Pospíšil, P. (2020). Interplay between antioxidants in response to photooxidative stress in Arabidopsis. Free Radic. Biol. Med. 160, 894–907. doi: 10.1016/j.freeradbiomed.2020.08.027
Kuzmina, N. V., Loshkareva, A. S., Shilova, L. A., Shtykova, E. V., Knyazev, D. G., Zimmerberg, J., et al. (2020). Protein-lipid interactions in formation of viral envelopes. Biophys. J. 118:523a. doi: 10.1016/j.bpj.2019.11.2874
Lambert, A. J., and Brand, M. D. (2009). “Reactive oxygen species production by mitochondria,” in Mitochondrial DNA: Methods and Protocols Methods in Molecular BiologyTM, ed. J. A. Stuart (Totowa, NJ: Humana Press), 165–181. doi: 10.1007/978-1-59745-521-3_11
Li, F., Zhao, X., Zhu, S., Wang, T., Li, T., Woolfley, T., et al. (2020). Identification and expression profiling of neuropeptides and neuropeptide receptor genes in Atrijuglans hetaohei. Gene 743:144605. doi: 10.1016/j.gene.2020.144605
Linthicum, K. J., Platt, K., Myint, K. S., Lerdthusnee, K., Inni, B. L., and Ghn, D. W. V. (1996). Dengue 3 virus distribution in the mosquito Aedes aegypti: an immunocytochemical study. Medic. Vet. Entomol. 10, 87–92. doi: 10.1111/j.1365-2915.1996.tb00086.x
Liu, K., Gao, Y., Liu, J., Wen, Y., Zhao, Y., Zhang, K., et al. (2016). Photoreactivity of metal–organic frameworks in aqueous solutions: metal dependence of reactive oxygen species production. Environ. Sci. Technol. 50, 3634–3640. doi: 10.1021/acs.est.5b06019
MaitiDutta, S., Chen, G., and Maiti, S. (2020). Tocopherol moderately induces the expressions of some human sulfotransferases, which are activated by oxidative stress. Cell Biochem. Biophys. 78, 439–446. doi: 10.1007/s12013-020-00938-x
Manokaran, G., Flores, H. A., Dickson, C. T., Narayana, V. K., Kanojia, K., Dayalan, S., et al. (2020). Modulation of acyl-carnitines, the broad mechanism behind Wolbachia-mediated inhibition of medically important flaviviruses in Aedes aegypti. PNAS 117, 24475–24483. doi: 10.1073/pnas.1914814117
Martín-Acebes, M. A., Vázquez-Calvo, Á, and Saiz, J.-C. (2016). Lipids and flaviviruses, present and future perspectives for the control of dengue, Zika, and West Nile viruses. Prog. Lipid Res. 64, 123–137. doi: 10.1016/j.plipres.2016.09.005
Meekins, D. A., Kanost, M. R., and Michel, K. (2017). Serpins in arthropod biology. Semin. Cell Dev. Biol. 62, 105–119. doi: 10.1016/j.semcdb.2016.09.001
Minter, B. E., Lowes, D. A., Webster, N. R., and Galley, H. F. (2020). Differential effects of MitoVitE, α-Tocopherol and Trolox on oxidative stress, mitochondrial function and inflammatory signalling pathways in endothelial cells cultured under conditions mimicking sepsis. Antioxidants 9:195. doi: 10.3390/antiox9030195
Moreira, L. A., Iturbe-Ormaetxe, I., Jeffery, J. A., Lu, G., Pyke, A. T., Hedges, L. M., et al. (2009). A Wolbachia symbiont in Aedes aegypti limits infection with dengue, chikungunya, and plasmodium. Cell 139, 1268–1278. doi: 10.1016/j.cell.2009.11.042
Murillo, J. R., Goto-Silva, L., Sánchez, A., Nogueira, F. C. S., Domont, G. B., and Junqueira, M. (2017). Quantitative proteomic analysis identifies proteins and pathways related to neuronal development in differentiated SH-SY5Y neuroblastoma cells. EuPA Open Proteom. 16, 1–11. doi: 10.1016/j.euprot.2017.06.001
Nässel, D. R., and Broeck, J. V. (2016). Insulin/IGF signaling in Drosophila and other insects: factors that regulate production, release and post-release action of the insulin-like peptides. Cell. Mol. Life Sci. 73, 271–290. doi: 10.1007/s00018-015-2063-3
Nazni, W. A., Hoffmann, A. A., NoorAfizah, A., Cheong, Y. L., Mancini, M. V., Golding, N., et al. (2019). Establishment of Wolbachia strain wAlbB in Malaysian populations of Aedes aegypti for dengue control. Curr. Biol. 29, 4241.e5–4248.e5. doi: 10.1016/j.cub.2019.11.007
Nowicka, B., Fesenko, T., Walczak, J., and Kruk, J. (2020). The inhibitor-evoked shortage of tocopherol and plastoquinol is compensated by other antioxidant mechanisms in Chlamydomonas reinhardtii exposed to toxic concentrations of cadmium and chromium ions. Ecotoxicol. Environ. Saf. 191:110241. doi: 10.1016/j.ecoenv.2020.110241
Nunes, A. T., Brito, N. F., Oliveira, D. S., Araujo, G. D. T., Nogueira, F. C. S., Domont, G. B., et al. (2016). Comparative proteome analysis reveals that blood and sugar meals induce differential protein expression in Aedes aegypti female heads. Proteomics 16, 2582–2586. doi: 10.1002/pmic.201600126
Paingankar, M. S., Gokhale, M. D., Deobagkar, D. D., and Deobagkar, D. N. (2020). Drosophila melanogaster as a model host to study arbovirus–vector interaction. bioRxiv doi: 10.1101/2020.09.03.282350
Pan, X., Zhou, G., Wu, J., Bian, G., Lu, P., Raikhel, A. S., et al. (2012). Wolbachia induces reactive oxygen species (ROS)-dependent activation of the Toll pathway to control dengue virus in the mosquito Aedes aegypti. PNAS 109, E23–E31. doi: 10.1073/pnas.1116932108
Pappireddi, N., Martin, L., and Wühr, M. (2019). A review on quantitative multiplexed proteomics. ChemBioChem 20, 1210–1224. doi: 10.1002/cbic.201800650
Perera, R., Riley, C., Isaac, G., Hopf-Jannasch, A. S., Moore, R. J., Weitz, K. W., et al. (2012). Dengue virus infection perturbs lipid homeostasis in infected mosquito cells. PLoS Pathog. 8:e1002584. doi: 10.1371/journal.ppat.1002584
Petersen, M. T., da Silveira, I. D., Tátila-Ferreira, A., David, M. R., Chouin-Carneiro, T., Van den Wouwer, L., et al. (2018). The impact of the age of first blood meal and Zika virus infection on Aedes aegypti egg production and longevity. PLoS One 13:e0200766. doi: 10.1371/journal.pone.0200766
Phillips, D. R., and Clark, K. D. (2017). Bombyx mori and Aedes aegypti form multi-functional immune complexes that integrate pattern recognition, melanization, coagulants, and hemocyte recruitment. PLoS One 12:e0171447. doi: 10.1371/journal.pone.0171447
Pickett, B. E., Sadat, E. L., Zhang, Y., Noronha, J. M., Squires, R. B., Hunt, V., et al. (2012). ViPR: an open bioinformatics database and analysis resource for virology research. Nucleic Acids Res. 40, D593–D598. doi: 10.1093/nar/gkr859
Poddar, S. K., Sifat, A. E., Haque, S., Nahid, N. A., Chowdhury, S., and Mehedi, I. (2019). Nicotinamide mononucleotide: exploration of diverse therapeutic applications of a potential molecule. Biomolecules 9:34. doi: 10.3390/biom9010034
Predel, R., Neupert, S., Garczynski, S. F., Crim, J. W., Brown, M. R., Russell, W. K., et al. (2010). Neuropeptidomics of the mosquito Aedes aegypti. J. Proteome Res. 9, 2006–2015. doi: 10.1021/pr901187p
Rauniyar, N., and Yates, J. R. (2014). Isobaric labeling-based relative quantification in shotgun proteomics. Proteome Res. 13, 5293–5309. doi: 10.1021/pr500880b
Rexer, T., Laaf, D., Gottschalk, J., Frohnmeyer, H., Rapp, E., and Elling, L. (2020). “Enzymatic synthesis of glycans and glycoconjugates,” in Advances in Biochemical Engineering/Biotechnology, (Berlin: Springer), 1–50. doi: 10.1007/10_2020_148
Ribeiro, J. M. C., Martin-Martin, I., Arcà, B., and Calvo, E. (2016). A deep insight into the sialome of male and female Aedes aegypti mosquitoes. PLoS One 11:e0151400. doi: 10.1371/journal.pone.0151400
Ryan, P. A., Turley, A. P., Wilson, G., Hurst, T. P., Retzki, K., Brown-Kenyon, J., et al. (2020). Establishment of wMel Wolbachia in Aedes aegypti mosquitoes and reduction of local dengue transmission in cairns and surrounding locations in northern Queensland, Australia. Gates Open Res. 3:1547. doi: 10.12688/gatesopenres.13061.2
Schmid, M. A., Kauffman, E., Payne, A., Harris, E., and Kramer, L. D. (2017). Preparation of mosquito salivary gland extract and intradermal inoculation of mice. Bio Protocol 7:e2407.
Selivanov, V. A., Zeak, J. A., Roca, J., Cascante, M., Trucco, M., and Votyakova, T. V. (2008). The role of external and matrix pH in mitochondrial reactive oxygen species generation. J. Biol. Chem. 283, 29292–29300. doi: 10.1074/jbc.M801019200
Serteyn, L., Ponnet, L., Saive, M., Fauconnier, M.-L., and Francis, F. (2020). Changes of feeding behavior and salivary proteome of Brown Marmorated Stink Bug when exposed to insect-induced plant defenses. Arthropod-Plant Interact. 14, 101–112. doi: 10.1007/s11829-019-09718-8
Shrinet, J., Bhavesh, N. S., and Sunil, S. (2018). Understanding oxidative stress in aedes during chikungunya and dengue virus infections using integromics analysis. Viruses 10:314. doi: 10.3390/v10060314
Słocińska, M., Chowański, S., and Marciniak, P. (2020). Identification of sulfakinin receptors (SKR) in Tenebrio molitor beetle and the influence of sulfakinins on carbohydrates metabolism. J. Comp. Physiol. B 190, 669–679. doi: 10.1007/s00360-020-01300-6
Stark, K. R., and James, A. A. (1998). Isolation and characterization of the gene encoding a novel factor xa-directed anticoagulant from the yellow fever mosquito, Aedes aegypti. J. Biol. Chem. 273, 20802–20809. doi: 10.1074/jbc.273.33.20802
Sterkel, M., Perdomo, H. D., Guizzo, M. G., Barletta, A. B. F., Nunes, R. D., Dias, F. A., et al. (2016). Tyrosine detoxification is an essential trait in the life history of blood-feeding arthropods. Curr. Biol. 26, 2188–2193. doi: 10.1016/j.cub.2016.06.025
Stick, R. V., and Williams, S. J. (2009). “Chapter 11 - Glycoproteins and proteoglycans,” in Carbohydrates: The Essential Molecules of Life (Second Edition), eds R. V. Stick and S. J. Williams (Oxford: Elsevier), 369–412.
Supek, F., Bošnjak, M., Škunca, N., and Šmuc, T. (2011). REVIGO summarizes and visualizes long lists of gene ontology terms. PLoS One 6:e21800. doi: 10.1371/journal.pone.0021800
The Uniprot Consortium (2019). UniProt: a worldwide hub of protein knowledge. Nucleic Acids Res. 47, D506–D515. doi: 10.1093/nar/gky1049
Toufeeq, S., Wang, J., Zhang, S.-Z., Li, B., Hu, P., Zhu, L.-B., et al. (2019). Bmserpin2 is involved in BmNPV infection by suppressing melanization in Bombyx mori. Insects 10:399. doi: 10.3390/insects10110399
Troupin, A., Londono-Renteria, B., Conway, M. J., Cloherty, E., Jameson, S., Higgs, S., et al. (2016). A novel mosquito ubiquitin targets viral envelope protein for degradation and reduces virion production during dengue virus infection. Biochim. Biophys. Acta Gen. Subj. 1860, 1898–1909. doi: 10.1016/j.bbagen.2016.05.033
Turley, A. P., Moreira, L. A., O’Neill, S. L., and McGraw, E. A. (2009). Wolbachia infection reduces blood-feeding success in the dengue fever mosquito, Aedes aegypti. PLoS Negl. Trop. Dis. 3:e516. doi: 10.1371/journal.pntd.0000516
Tyanova, S., Temu, T., Sinitcyn, P., Carlson, A., Hein, M. Y., Geiger, T., et al. (2016). The Perseus computational platform for comprehensive analysis of (prote)omics data. Nat. Methods 13, 731–740. doi: 10.1038/nmeth.3901
Vasconcellos, A. F., Mandacaru, S. C., de Oliveira, A. S., Fontes, W., Melo, R. M., de Sousa, M. V., et al. (2020). Dynamic proteomic analysis of Aedes aegypti Aag-2 cells infected with Mayaro virus. Parasit. Vect. 13:297. doi: 10.1186/s13071-020-04167-2
Wang, X., Zhang, Y., Zhang, R., and Zhang, J. (2019). The diversity of pattern recognition receptors (PRRs) involved with insect defense against pathogens. Curr. Opin. Insect Sci. 33, 105–110. doi: 10.1016/j.cois.2019.05.004
Wasinpiyamongkol, L., Patramool, S., Luplertlop, N., Surasombatpattana, P., Doucoure, S., Mouchet, F., et al. (2010). Blood-feeding and immunogenic Aedes aegypti saliva proteins. Proteomics 10, 1906–1916. doi: 10.1002/pmic.200900626
White, P. M., Serbus, L. R., Debec, A., Codina, A., Bray, W., Guichet, A., et al. (2017). Reliance of Wolbachia on high rates of host proteolysis revealed by a genome-wide RNAi screen of Drosophila cells. Genetics 205, 1473–1488. doi: 10.1534/genetics.116.198903
Xi, Z., Ramirez, J. L., and Dimopoulos, G. (2008). The Aedes aegypti toll pathway controls dengue virus infection. PLoS Pathog. 4:e1000098. doi: 10.1371/journal.ppat.1000098
Yin, C., Sun, P., Yu, X., Wang, P., and Cheng, G. (2020). Roles of symbiotic microorganisms in arboviral infection of arthropod vectors. Trends Parasitol. 36, 607–615. doi: 10.1016/j.pt.2020.04.009
Zels, S., Dillen, S., Crabbé, K., Spit, J., Nachman, R. J., and Vanden Broeck, J. (2015). Sulfakinin is an important regulator of digestive processes in the migratory locust, Locusta migratoria. Insect Biochem. Mol. Biol. 61, 8–16. doi: 10.1016/j.ibmb.2015.03.008
Keywords: proteome, quantitative, Aedes aegypti, Wolbachia, Zika virus, immune response
Citation: Martins M, Ramos LFC, Murillo JR, Torres A, de Carvalho SS, Domont GB, de Oliveira DMP, Mesquita RD, Nogueira FCS, Maciel-de-Freitas R and Junqueira M (2021) Comprehensive Quantitative Proteome Analysis of Aedes aegypti Identifies Proteins and Pathways Involved in Wolbachia pipientis and Zika Virus Interference Phenomenon. Front. Physiol. 12:642237. doi: 10.3389/fphys.2021.642237
Received: 15 December 2020; Accepted: 04 February 2021;
Published: 25 February 2021.
Edited by:
Jose Luis Ramirez, United States Department of Agriculture (USDA), United StatesReviewed by:
Valérie Choumet, Institut Pasteur, FranceCopyright © 2021 Martins, Ramos, Murillo, Torres, de Carvalho, Domont, de Oliveira, Mesquita, Nogueira, Maciel-de-Freitas and Junqueira. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Magno Junqueira, bWFnbm9qdW5xdWVpcmFAaXEudWZyai5icg==; Rafael Maciel-de-Freitas, ZnJlaXRhc0Bpb2MuZmlvY3J1ei5icg==
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.