 Emile Levy
Emile Levy Jean François Beaulieu4
Jean François Beaulieu4 Schohraya Spahis
Schohraya Spahis
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Physiol., 27 January 2021
Sec. Gastrointestinal Sciences
Volume 12 - 2021 | https://doi.org/10.3389/fphys.2021.629222
This article is part of the Research TopicAdvances in Dietary Fat AbsorptionView all 6 articles
During the last two decades, a large body of information on the events responsible for intestinal fat digestion and absorption has been accumulated. In particular, many groups have extensively focused on the absorptive phase in order to highlight the critical “players” and the main mechanisms orchestrating the assembly and secretion of chylomicrons (CM) as essential vehicles of alimentary lipids. The major aim of this article is to review understanding derived from basic science and clinical conditions associated with impaired packaging and export of CM. We have particularly insisted on inborn metabolic pathways in humans as well as on genetically modified animal models (recapitulating pathological features). The ultimate goal of this approach is that “experiments of nature” and in vivo model strategy collectively allow gaining novel mechanistic insight and filling the gap between the underlying genetic defect and the apparent clinical phenotype. Thus, uncovering the cause of disease contributes not only to understanding normal physiologic pathway, but also to capturing disorder onset, progression, treatment and prognosis.
Intestinal fat transport is a prerequisite process to deliver alimentary lipids to the bloodstream for subsequent metabolism and peripheral energy homeostasis. Several biochemical, physiological and morphological requirements must be met to allow intraluminal digestion and intracellular transport of triglycerides (TG), phospholipids (PL), and cholesterol (CHOL). Digestive processing and mucosal transport represent the essential steps to warrant optimal lipid absorption. However, their abnormalities result in intestinal fat malabsorption not only of lipids, but also of fat-soluble vitamins, concomitantly with gastrointestinal (GI) symptoms along with steatorrhea, nutritional fatty acid (FA) and vitamin deficiency, and substantial extra-intestinal disorders. The objective of the present review is to focus on congenital disorders of intestinal lipid absorption, highlighting the molecular genetics and pathophysiological mechanisms while describing the clinical manifestations and management of patients.
Lipid digestion involves breakdown of TGs into FA and 2-monoglyceride (MG) by lipase, hydrolysis of cholesteryl ester (CE) into free CHOL and FA by CHOL esterase, and decomposition of PL into lysoPL and FA by phospholipase A2 in the intestinal lumen (Levy et al., 2007). The three enzymes are synthesized by the pancreas and are delivered through its exocrine acinar cells. Importantly, an alkaline mixture composed of water and bicarbonate is released by pancreatic ductal cells into duodenum to maintain the ideal pH for enzyme activity. Finally, bile salts that are concentrated in gallbladder stores also flow into the duodenum. Their detergent action assists in breaking down large fat globules into small droplets, and in solubilizing the lipids by forming micelles, thereby allowing the enzymes to get to emulsified lipid droplets. Impairment of enzymatic activity, bicarbonate supply or bile acid output leads to malabsorption symptoms, including steatorrhea, deficiency of essential FAs (EFA) and fat-soluble vitamins (A, D, E, and K), weight loss, abdominal discomfort, and abdominal bloating. Various causes and conditions may be implicated such as chronic pancreatitis, pancreatic duct obstruction, pancreatic cancer, diabetes mellitus, partial or total pancreatectomy, cystic fibrosis, inflammatory bowel diseases, small bowel resections, or bariatric intervention.
Following intraluminal fat digestion, the lipolytic products must cross small intestinal brush border to form lipid-carrying lipoproteins (Figure 1). The access of FAs to enterocytes can be achieved by passive diffusion displaying a “flip-flop” pattern down a favorable concentration gradient (Mashek and Coleman, 2006; Storch and Thumser, 2010). A second FA uptake process involves protein-facilitated FA transfer in view of its saturation nature as exemplified by FA transfer proteins (Storch and Thumser, 2010). Apical microvillus membrane proteins such as fatty acid transport protein 4 (FATP4) and CD36 mediate the FA transfer inside the enterocyte (Stahl et al., 1999; Nassir et al., 2007). FATP4 is highly expressed in villus enterocytes, functions in FA incorporation, and traps FA through their conversion into CoA derivative given its endogenous acyl CoA synthetase activity (Milger et al., 2006). For its part, CD36 is also abundant in the small intestine, facilitates FA uptake, and exhibit various functions in lipid absorption such as fat test perception and food intake (Laugerette et al., 2005; Martin et al., 2011; Pepino et al., 2012).
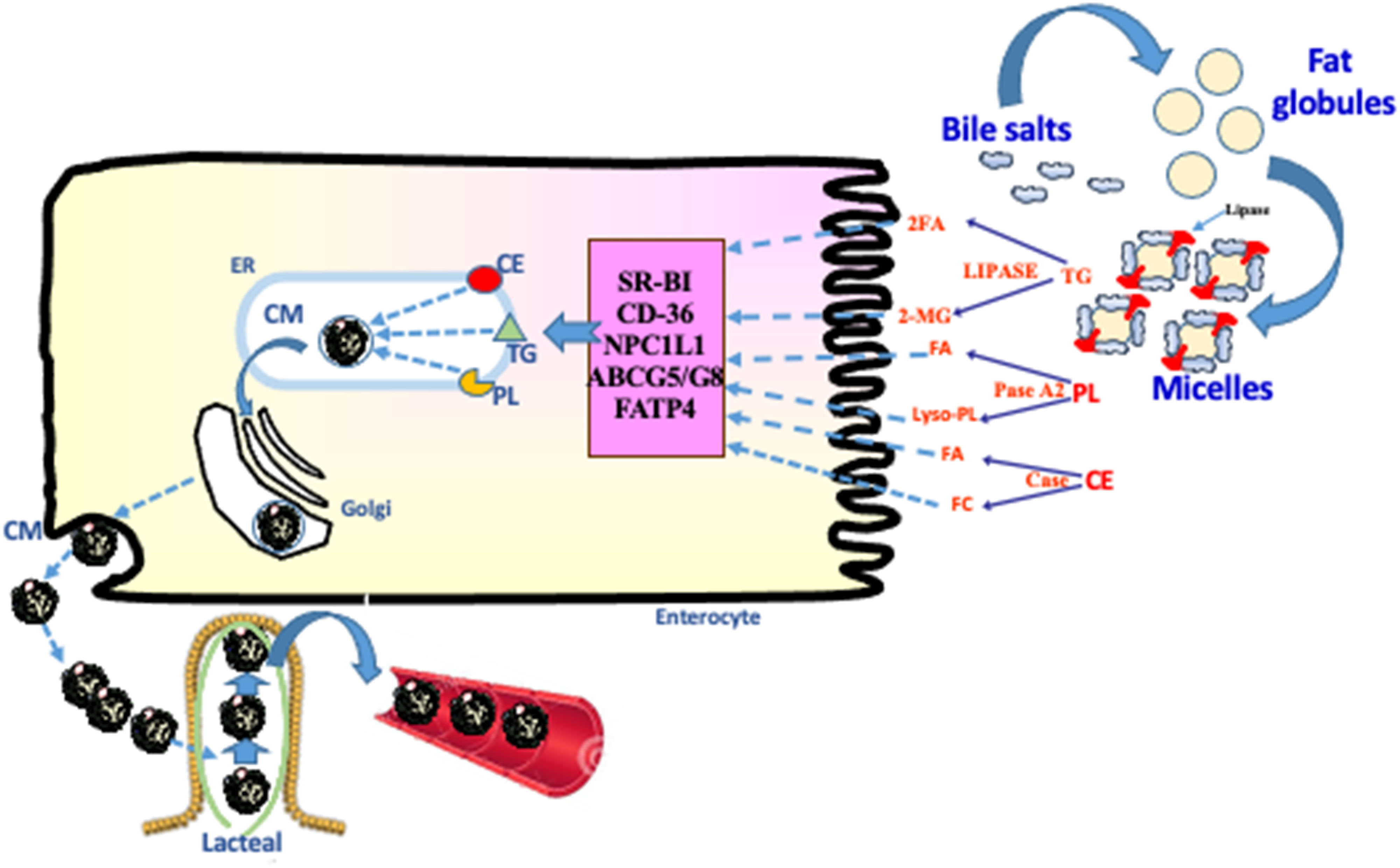
Figure 1. Dietary lipid digestion and absorption. Digestion of dietary fat requires bile acids, digestive enzymes and bicarbonate. Dietary lipids, mostly triacylglycerols (TG) but also cholesteryl ester (CE) and phospholipids (PL), are initially emulsified by bile acids and then hydrolyzed by pancreatic lipase, cholesterol esterase and phospholipase A2. The lipolytic products fatty acids (FA), 2-monoglyceride (2-MG), free cholesterol (FC), and lyso PL form micelles by the action of bile acids in the proximal small bowel. The micelles release the lipolytic products near to the microvillus membrane to allow their uptake by lipid transporters: scavenger receptor BI (SR-BI), fatty acid translocase (CD-36), Niemann-Pick C1-Like 1 (NPC1L1), ATP-binding cassette G5/G8 (ABCG5/G8), and fatty acid transport protein 4 (FATP4). Following their uptake, the lipolytic products are deposited in the endoplasmic reticulum (ER) by L- and I- fatty acid binding proteins (L- and I-FABP), for esterification, assembly with apolipoproteins, packaging into chylomicrons (CM) and transport to bloodstream via the lymphatics.
For the intestinal transport of alimentary CHOL, various microvillus transporters have been described, including Niemann-Pick C1-Like 1 (NPC1L1), scavenger receptor-BI (SR-BI), and CD36. While these transporters regulate CHOL influx, the heteromeric complex ATP-binding cassette G5 (ABCG5)/G8 on villus brush-border membrane is in charge of CHOL efflux directly toward intestinal lumen. Besides, ABCA1 is a basolateral efflux pump that transfers CHOL to apolipoprotein (Apo) A-I, thereby contributing to high-density lipoprotein (HDL) particles (Figure 2).
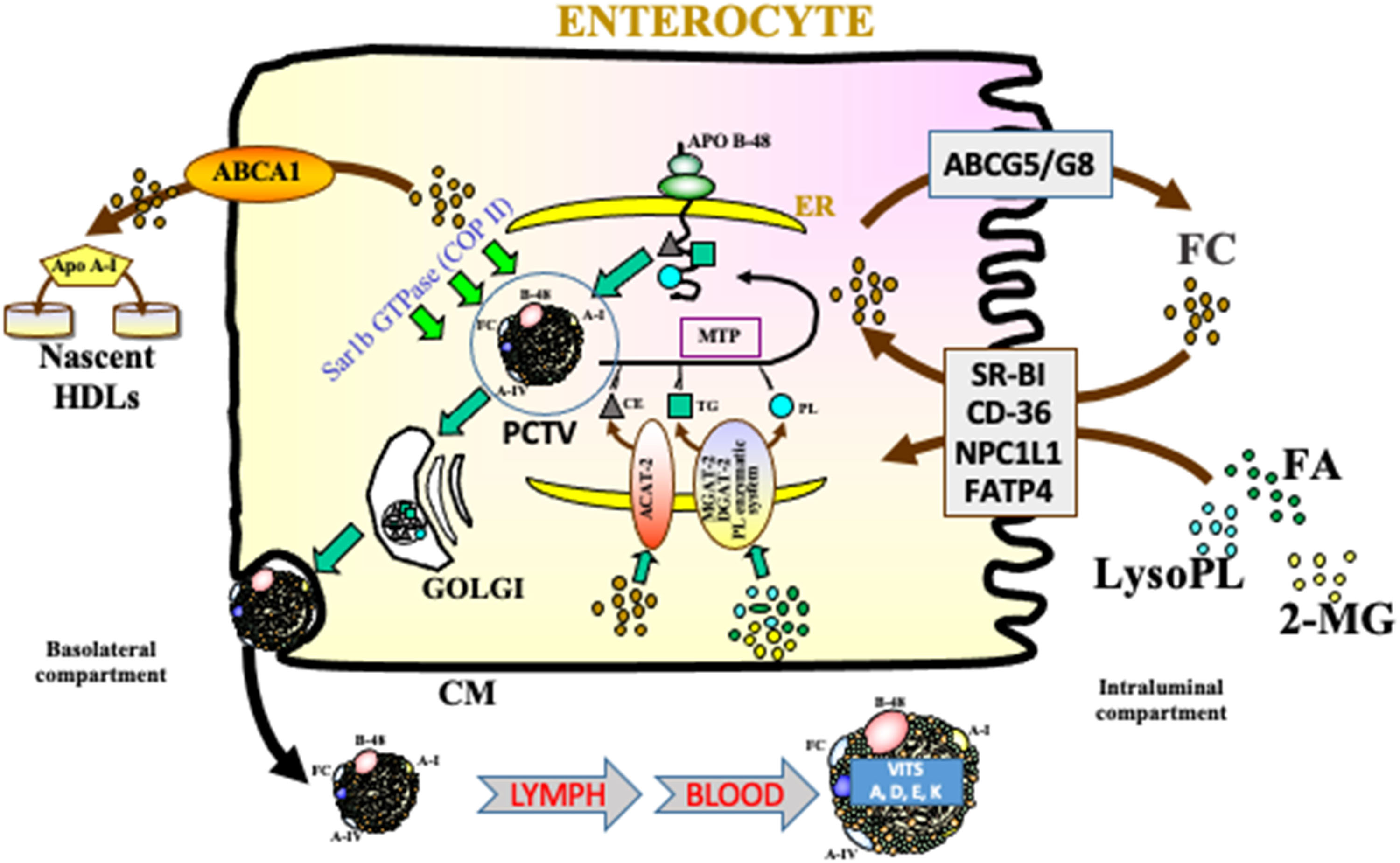
Figure 2. Intracellular network required for lipid transport and lipoprotein assembly. Free cholesterol (FC), monoacylglycerols (MG), lysophospholipids (LysoPL), and fatty acids (FA), carried by bile salt mixed micelles in the intestinal lumen, cross the unstirred water layer and are captured by specific proteins within brush border membranes of the jejunum considered as the optimal segment for lipid absorption. Whereas NPC1L1, viewed as the most putative transporter, mediated the uptake of digestive FC, physiologically excessive amounts are secreted back to the intestinal lumen via the apical membrane of the enterocyte by the ABCG5/G8 heterodimer. Similarly, ABCA1, localized on the basolateral surface contributes to cholesterol homeostasis by promoting cholesterol efflux to plasma apo A-I, which enhances the formation of nascent HDL. Following the transfer of lipolytic products to the endoplasmic reticulum (ER), local acyl-CoA:cholesterol O-acyltransferase 2 (ACAT-2) catalyzes the esterification of FC, while monoacylglycerol acyltransferase (MGAT)-diacylglycerol acyltransferase (DGAT) complex recycles FA and MG into triacylglycerols (TG), and phosphoglyceide-synthesizing enzymes intervene in phospholipids (PL) formation. Under the action of microsomal triglyceride transfer protein (MTP), lipids are then assembled with newly synthesized apo B-48 to generate chylomicrons (CM). These lipoprotein particles are then conveyed in specialized vesicles from the ER to the Golgi apparatus with the crucial participation of Sar1b GTPase. They are finally exocytosed after the fusion of Golgi vesicles and basolateral membrane.
Through the action of these transporters, lipolytic products move into the enterocyte. With the assistance of cytosolic binding proteins such as intestinal fatty acid-binding protein (I-FABP) and Liver-FABP (L-FABP), they are directed to the membrane of the endoplasmic reticulum (ER), where they are re-esterified. The 2-MG and FAs are reconstituted to form TG by the MG pathway involving monoacylglycerol transferase (MGAT) and diacylglycerol transferase (DGAT) (Bell and Coleman, 1980). Sequentially, MGAT catalyzes the formation of diglyceride (DG) whereas DGAT catalyzes the final reaction leading to TG. Diglycerides can also be synthesized by a secondary glycerol phosphate pathway (Lehner and Kuksis, 1996). For their part, CHOL and PL are re-esterified in their original forms by acylcholesterol acyltransferase-2 (ACAT2) and biosynthetic pathways (e.g., lyso-phosphatidylcholine acyltransferases), respectively (Buhman et al., 2000; Li et al., 2015). Thereafter, microsomal triglyceride transfer protein (MTTP) shuttles TG, CE, and PL to the structural Apo B-48 to promote chylomicron (CM) assembly (Black, 2007; Levy et al., 2011). While MTTP appears as an essential protein to uphold CM biogenesis by lipidation of the critical Apo B-48, Sar1B GTPase is another crucial component of COPII vesicles that buds from the ER to transport pre-CM to the Golgi apparatus (Levy et al., 2011). In this intracellular compartment, CM particles fuse into another transport vesicle and are vectorially transported to the basolateral membrane for secretion into the circulation via the lymphatic system (Lo and Coschigano, 2020) (Figure 1).
In this section, we will review how genetic abnormalities may hamper intracellular lipid movement and CM assembly/output.
Abetalipoproteinemia (ABL) is a homozygous autosomal recessive disorder caused by mutations of the MTTP gene (Wetterau et al., 1992; Shoulders et al., 1993) (Table 1). The MTTP is in fact a soluble microsomal heterodimer consisting of a unique large 97 kDa protein conferring lipid transfer activity, and the multifunctional 58 kDa protein disulfide isomerase necessary to maintain the catalytically active structure (Wetterau and Zilversmit, 1984). MTTP resides in the ER lumen and orchestrates the transfer of TG, CE, and PL onto Apo B-48 to produce pre-CM particles. Very often, Apo B-48 is not detected in the intestinal tissue of ABL patients probably due to the limiting availability of lipids, which normally protect it from proteasome degradation (Boren et al., 1992; McLeod et al., 1994; Rustaeus et al., 1995). In fact, the addition of a core lipid to the nascent Apo B-48 in the ER takes place during its translation and translocation, which prevents its degradation and allows the protein to grow and translocate completely into the lumen (Boren et al., 1994; Swift, 1995).
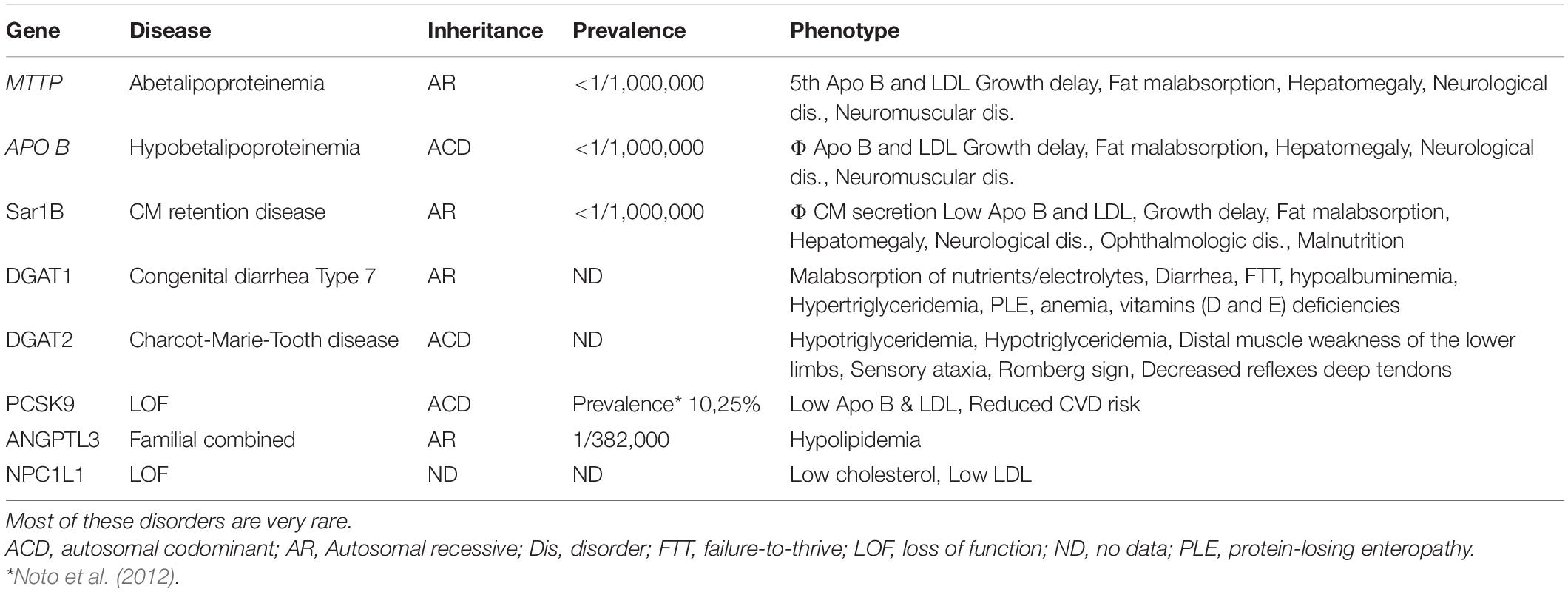
Table 1. Genetic defects of genes associated with intestinal malabsorption and lipid dysmetabolism along with related complications.
Apart from the MTTP residence in the ER, the occurrence of membrane-associated Apo B in the Golgi, coupled with its interaction with active MTTP, suggests an important role for the Golgi in the biogenesis of Apo B-containing lipoproteins (Levy et al., 2002). As the liver proceeds in the same way to assemble very low-density lipoprotein (VLDL), the genetic mutations of MTTP affects Apo B-100-containing hepatic derived lipoproteins. It is important to remember that the two Apo B translation products (B-100 and B-48) are produced by the same gene, but intestinal Apo B-48 is generated through mRNA editing mechanism employing the catalytic deaminase APOBEC1 (Teng et al., 1990, 1993; Anant et al., 1995).
In view of the aberrations of MTTP, there is a total absence of circulating CM (following fat feeding) and Apo B-containing lipoproteins along with extremely low level of plasma TG, total CHOL, and Apo B-100 (Raabe et al., 1998). Young children present with diarrhea, malabsorption, and severe steatorrhea, with additional features such as deficiency of EFAs and fat-soluble vitamins, red blood cell acanthocytosis, retinal degeneration, and neurological dysfunction (likely due to the paucity of vitamins A, D, E, K), and steatohepatitis (Black et al., 1991).
Given the failure to thrive and abnormal clinical features such as neurological sequelae in early childhood, genetic testing should rapidly validate the diagnosis. ABL patients necessitate permanent maintenance on a low-fat diet. Although long-term high-dose supplementation with vitamins A (10–15,000 IU/day) and E (100 mg/kg/day) improve retinal and neurological functions (Granot and Kohen, 2004), their plasma levels seldom return to the normal range (Traber, 2013).
Familial hypobetalipoproteinemia (FHBL) is a monogenic, inherited disorder, which closely resembles ABL, but is essentially caused by genetic defects of the APOB gene on chromosome 2 (Whitfield et al., 2003) (Table 1). As a result of non-sense, frame shift and splicing mutations in the APOB gene, prematurely truncated Apo B forms (i.e., smaller proteins than Apo B) are associated with the total absence of circulating CMs and Apo B-48 in response to fat meals (Levy et al., 1994). Moreover, the liver is unable to secrete VLDL, and extremely low plasma concentrations of low-density lipoprotein (LDL), TG and total CHOL are common in FHBL (Di Leo et al., 2008; Buonuomo et al., 2009). Sometimes, search of mutations uncovered truncated APOB forms of various lengths, ranging from APOB-6.46 to APOB-89. The truncated forms are characterized by the missing carboxyl-terminal portion, which must have interfered with the translation of full-length APOB (Wang et al., 2018).
Homozygous FHBL presents in infancy or early childhood with variable clinical manifestations, including failure to thrive, steatorrhea, undetectable fat-soluble vitamins, EFA deficiency, acanthocytosis, and neurologic deficits with macular degeneration (Lee and Hegele, 2014). In adulthood, FHBL may be affected by hepatic steatosis (Heeks et al., 2013). As can be seen, the phenotype is similar to that of ABL, and the same goes for management. Unfortunately, homozygous FHBL may be accompanied by cirrhosis (Bonnefont-Rousselot et al., 2009; Florkowski et al., 2010) and hepatocellular carcinoma (Di Leo et al., 2008; Cefalu et al., 2013). It is therefore recommended that patients undergo hepatic evaluation regularly.
CRD is another congenital malabsorption disorder that highlights the obligatory trafficking of nascent CM between ER and Golgi in intestinal absorptive cells (Levy et al., 1987; Roy et al., 1987) (Table 1). Mutations of SAR1-ADP ribosylation factor, type B (SAR1B) prevent the conveyance of CM-containing vesicles through the early secretory pathway, leading to the accumulation of pre-CM (Jones et al., 2003; Charcosset et al., 2008). In fact, SAR1B, belonging to the Ras superfamily of guanosine triphosphatases (GTPases), is essential for the coatomer COPII that transports proteins from the rough ER to the Golgi apparatus, a process requiring the small Sar1b GTPase for the exchange of GDP for GTP. Important studies have shown the ability of SAR1B to initiate vesicle formation by recruiting first the inner COPII coat components (Sec23 and Sec24) and subsequently the components of the outer flexible coat (Sec13/Sec31) (Barlowe et al., 1994). It has been proposed that CMs of large size move from the ER to Golgi, probably inside the pre-CM transport vesicle (PCTV) (Siddiqi et al., 2003, 2006). From where we stand at present, SAR1B aberrations affect the transport of pre-CM from the ER to the Golgi in PCTV, including their fusion with the cis-Golgi. Nevertheless, further studies are explicitly required to improve our vision of the mechanisms implicated in CRD pathogenesis.
Chronic diarrhea, vomiting, abdominal distension, and failure to thrive are among the most frequent and earliest symptoms affecting CRD patients. Incapacity to export CM in CRD impairs the intestinal transport of fat-soluble vitamins and the status of EFA. Furthermore, plasma levels of CHOL, PL, LDL, HDL, and Apos (B, A-I) are usually below 50% of control values (Peretti et al., 2009, 2010). Additional clinical findings comprise ophthalmologic (micronystagmus, mild deficit in the perception of the blue yellow axis and delayed dark adaptation) and neurological complications (areflexia, proprioceptive aberrancy, ataxia, myopathy, and sensory neuropathy), which are of lesser importance compared with those of ABL and FHPL. Elevated creatine kinase and cardiomyopathy have also been reported along with muscular abnormality. Also noteworthy are the inadequate mineralization and retarded bone maturation. While moderate hepatomegaly and macrovesicular steatosis are detected, steatohepatitis, and cirrhosis remain rare (Peretti et al., 2010).
Although clinical examination and biological evaluations centring on nutrition growth, GI, liver and neurological manifestations may help in the diagnosis, genetic testing (identifying Sar1B mutations) remains the most accurate and reliable tool. Importantly, the signs of chronic diarrhea, fat malabsorption, fat-laden enterocytes, atypical lipid, and vitamin profile are suggestive of CRD.
Management of these patients consists in recommending a fat-free diet, enriched in EFA, medium-chain TGs and liposoluble vitamins, including 50 UI/kg/day vitamin E, 15,000 IU/day vitamin A, 15 mg/week vitamin K, and 800–1,200 UI/kg/day or 100,000 IU/2 months vitamin D if younger than 5 years old and 600,000 IU/2 months if older than 5 years old (Peretti et al., 2010). Recently, a study evaluated the efficacy of fat-soluble vitamin E acetate and tocofersolan (a water-soluble derivative of RRR-α-tocopherol) by evaluating the ability of each formulation to restore vitamin E storage after 4 months of treatment (Cuerq et al., 2018). While in patients with ABL, tocofersolan and α-tocopherol acetate bioavailability was extremely low (2.8 and 3.1%, respectively), bioavailability was higher in patients with CRD (tocofersolan, 24.7%; α-tocopherol acetate, 11.4%).
FA translocase or CD36 is a class B scavenger receptor, which is anchored in the membrane by transmembrane domains, and is largely involved in high affinity FA uptake in several tissues (Yamashita et al., 2007). In the small intestine, CD36 is localized in the villus membrane of the jejunum (Poirier et al., 1996; Chen et al., 2001; Lobo et al., 2001). Knockout (KO) of CD36 did not disturb the intestinal uptake of FA, and no impaired FA absorption was seen in CD36-KO mice (Goudriaan et al., 2002). Later, it was discovered a reduced lipid output in the lymph of CD36-null mice (Drover et al., 2005). Probably, the reason for this inconsistency was that CD36 ablation also impacts on CM clearance by affecting its size (Drover et al., 2005), resulting in circulating CM build-up, thereby concealing the lessened lipid output from the enterocytes to the lymphatic system. The presence of CD36 is therefore necessary for the delivery of lipids from the gut. Accordingly, patients with CD36 deficiency exhibited increased levels of plasma TG, free FA, CM remnants, and Apo B-48 because of an enhanced production of smaller lipoproteins than CM in the intestine (Masuda et al., 2009).
To elucidate the role of FATP4 in intestinal FA uptake, Fatp4 knockdown is performed in primary mouse small intestinal enterocytes. FATP4 deficiency reduces FA uptake (Stahl et al., 1999). Seemingly, this downregulation is not dependent on the transport function of the FATP4 protein but rather on its enzymatic activity conducting to FA esterification with coenzyme A for TG and phospholipid biosynthesis in the gut (Milger et al., 2006; Ko et al., 2020). If heterozygous Fatp4+/– mice displays reduced long-chain FA uptake (Gimeno et al., 2003), Fatp4–/– knockdown mice mates display similar food intake, growth, weight gain, intestinal TG absorption and fecal fat loss on either low or high-fat diets (Shim et al., 2009). Although serum CHOL concentrations were lower in Fatp4–/– mice, the authors conclude that intestinal FATP4 has no physiological part in dietary lipid absorption in mice. However, in the presence of bacterial infection causing intestinal nutrient malabsorption in piglets, a synthetic antimicrobial peptide KR-32 alleviates malabsorption by improving the expression of FABP4 (Liu et al., 2019).
As G/A polymorphism in exon 3 of the FATP4 genes rise to a Gly209Ser substitution with potential structural-functional implications, a group of researchers investigates whether variation within the FATP4 gene influences fasting and postprandial lipid and lipoprotein variables along with markers of insulin resistance (IR) in healthy, middle-aged Swedish men (Gertow et al., 2004). Their hypothesis turns out to be correct in view of the negative association of the FATP4 variant with metabolic syndrome components, including IR, TGs, postprandial lipemia, and HDL-CHOL.
The intestinal form, I-FABP, is encoded by the FABP2 gene, and is expressed exclusively in the proximal intestine where the bulk of fat absorption occurs (Thumser and Storch, 2000; Levy et al., 2009). Apparently, this protein is involved in intracellular targeting of FA given its involvement in FA transfer between membranes to allow FA metabolism and processing of dietary long-chain FA into CMs (Thumser and Storch, 2000; Hussain et al., 2005; Montoudis et al., 2008), while being regulated by lipids, hormones, and cytokines (Dube et al., 2001; Carrier et al., 2004). Accordingly, a reduction of body weight was noticed in I-FABP–/– mice (Gajda et al., 2013), suggesting possible fat malabsorption. When I-FABP KO mice were challenged with high-fat diet, total fecal excretion per gram of food intake was increased concomitantly with decreased energy absorption (Lackey et al., 2020). It finally appeared that intestinal transit and motility are stimulated by I-FABP deletion as a consequence of altered vagal tone induced by reduced cannabinoid receptor 1 activation, thereby affecting nutrient and lipid absorption (Lackey et al., 2020). Despite all these interesting data, I-FABP overexpression in normal molecularly modified normal human intestinal epithelial cells is not related to lipid esterification, Apo synthesis and lipoprotein assembly, which therefore excludes its role in intestinal fat transport (Montoudis et al., 2006). Likewise, although I-FABP shows a high affinity for long-chain FAs and has been suggested to be involved in enterocyte FA uptake (Murphy et al., 1996; Alpers et al., 2000), animal models lacking I-FABP did not exhibit impaired FA uptake (Vassileva et al., 2000; Lagakos et al., 2011; Gajda et al., 2013). On the other hand, decreased amounts of I-FABP are observed in patients with ABL and CRD in link with the pathological intracellular accumulation of lipid structures in the enterocytes, leading the investigators to hypothesize that I-FABP acts as a lipid sensor to prevent the intracellular esterification of FA into TGs which would otherwise lead to further additional intestinal injury (Guilmeau et al., 2007).
Importantly, some studies have shown that variations in FABP2 gene may influence both intestinal lipid absorption and metabolism. The G-to-A substitution at codon 54 of the FABP2 gene, which results in an alanine-to-threonine substitution at amino acid 54 (Ala54Thr) of I-FABP, has been reported to be associated with increased intestinal fat absorption (Agren et al., 1998; Levy et al., 2001), as well as FA oxidation, IR, and diabetes (Baier et al., 1995). Furthermore, it has been proposed that the effects of FABP2 allelic variations on lipid traits are context dependent, indicating that this variant may play an important role in cardiovascular pathogenesis in the presence of IR and dyslipidemia (Stan et al., 2005). In the light of all these observations, I-FABP is central in intestinal physiology and metabolic disorders, but additional efforts are needed to precise its specific functions.
L-FABP is abundantly detected in the small intestine and has broad FA binding specificity with high affinity for long-chain polyunsaturated FAs though it may bind to CHOL, acyl-CoA, bile acid, and phytanic acid (Gordon et al., 1985; Lowe et al., 1987; Thumser and Wilton, 1996; Wolfrum et al., 1999). In addition, it is associated with the ER membrane of enterocytes where it plays a role as a budding initiator protein for PCTV, indicating its influence on CM synthesis/secretion (Neeli et al., 2007; Siddiqi et al., 2010). Accordingly, L-FABP null mice were protected against diet-induced obesity and hepatic steatosis (Newberry et al., 2006). On the other hand, investigators did not record any overt growth delay or failure to gain weight in chow-fed L-Fabp–/– mice (Martin et al., 2003; Newberry et al., 2003). More startling is the finding that female L-Fabp–/– mice develop a striking obesity phenotype in administrating a semisynthetic diet supplemented with CHOL (Martin et al., 2006). To resolve these conflicting data, different fat diets are administered to female L-Fabp–/– mice (Newberry et al., 2008). In contrast to high-polyunsaturated FA, high-saturated fat dramatically protected against obesity and hepatic steatosis, which is indicative that L-FABP functions as a metabolic sensor depending on the type of FA. Later, it has been demonstrated that L-FABP–/– mice are characterized by a modest MG trafficking defect and defective mucosal FA oxidation (Gajda et al., 2013). A recent study has shown that low L-FABP expression compromises initial uptake rate of FA and also reduces basolateral TG secretion (Rodriguez Sawicki et al., 2017).
Noteworthy, L-FABP gene polymorphisms are also associated with FA metabolism particularly in the liver (Richieri et al., 1994). For example, a negative association has been detected between FABP1 T94A and plasma TG levels, probably as a consequence of negative interference of the T94A variant with FA binding in humans (Fisher et al., 2007; Gao et al., 2010). In line with this assumption, T94A substitution markedly altered the human L-FABP structure and stability, along with conformational and functional response to fibric acid derivatives (fibrates), a medication lowering blood TG levels and reducing the liver VLDL production (Martin et al., 2013). It is important to point out the binding of L-FABP with PPARα, resulting in ligand transfer and PPARα transcription of multiple proteins in FA metabolism in mouse primary hepatocytes, with a net effect of lessening plasma TG (Velkov, 2013). Whether similar actions of T94A variant apply to the small intestine is not known despite the abundant L-FABP content in the small intestine.
A few groups have detected an association of intestinal lipid absorption with SR-BI, a cell-surface glycoprotein expressed in the apical microvillus membrane. SR-BI knockdown results in decreased FA influx and CM export in Caco-2 cells (Levy et al., 2004), whereas SR-BI overexpression has led to enhanced dietary CHOL absorption in mice (Bietrix et al., 2006). Other studies have reported raised apical CHOL uptake by Caco-2 cells using SR-BI-blocking antibody and by small inhibitory RNA (Cai et al., 2004). However, these negative findings should be considered with caution since the extracellular loops of SR-BI are efficient receptors for intestinal mixed micelles, and the properties and composition of micellar solution represent a key factor governing micelle interactions with intestinal SR-BI (Goncalves et al., 2015). Precautions do apply since recent works underline that intestinal SR-BI is a critical regulator of CM transport (Lino et al., 2015) and liposoluble vitamins (Mardones et al., 2002; Reboul et al., 2005; van Bennekum et al., 2005). In fact, acute administration of BLT-1, an SR-BI inhibitor, to hamsters and rats significantly lowers postprandial plasma TGs and CM without effect on CHOL accumulation (Lino et al., 2015). Therefore, the authors suggest that intestinal SR-BI is more involved in postprandial TG handling than in CHOL uptake.
As mentioned previously, NPC1L1 mediates CHOL trafficking from the apical microvillus membrane to the ER (Sane et al., 2006; Field et al., 2007; Nakano et al., 2019). Since free and esterified CHOL constitute moieties of CM components, and since NPC1L1 gene expression in the gut is closely correlated with CM-CHOL (Lally et al., 2007), it is believed that NPC1L1-mediated CHOL supply to CM formation plays a role in lipid assimilation process in enterocytes. In line with this assumption, the specific inhibitor of NPC1L1 is able to reduce postprandial Apo B-48 output in hamsters and lessen CM secretion from the intestine of mice fed a Western diet (Sandoval et al., 2010). Interestingly, NPC1L1 impedes CHOL esterification and intracellular CM vesicle trafficking in enterocytes in response to ezetimibe, which suggests that NPC1L1 participates in a control mechanism for competent CM packaging and output by restraining intracellular CHOL movement at a cellular level (Nakano et al., 2020).
Little information is available on the role of ABCG5/G8 in intestinal lipid transport and CM production. However, it is important to note that ABG5/G8 deficiency causes hypertriglyceridemia by increasing intestinal absorption, stimulating hepatic TG production, and lowering plasma TG catabolism in mice with ABCG5/G8 deficiency (Mendez-Gonzalez et al., 2011). In this study, intestinal absorption and secretion of TG were enhanced in ABCG5/G8 null mice, but TG secretion appeared to be greater than TG absorption.
Elegant studies demonstrated that ACAT2, a CHOL-esterifying enzyme residing in the ER membrane, increases CHOL absorption efficiency by providing CE for CM packaging and exocytosis into lymph (Nguyen et al., 2012). However, very poor data are available to draw conclusion on the role of ACAT2 in CM assembly and secretion.
As mentioned before, the DGAT1 gene encodes DGAT1 protein, a microsomal enzyme with an abundant expression particularly in the small intestine (Cases et al., 1998; Haas et al., 2012). In humans, DGAT1 catalyzes the final step in TG synthesis using DG and FA-CoA, supporting lipid absorption (Yen et al., 2008). Given the lack of DGAT2 expression in the human intestine (Haas et al., 2012), mutations in DGAT1 gene may cause conceptually various disorders (Table 1). In fact, patients with molecular aberrations in DGAT1 exhibited protein-losing enteropathy, a congenital diarrheal disorder with failure to thrive in early infancy (Haas et al., 2012; Stephen et al., 2016; Gluchowski et al., 2017; Ratchford et al., 2018; van Rijn et al., 2018; Ye et al., 2019; Xu et al., 2020). Loss-of-function as a consequence of DGAT1 variations may also cause elevated fecal alpha-1-antitrypsin, high TGs, vomiting, low albumin, elevated transaminases, and low IgG (Haas et al., 2012; Stephen et al., 2016; Gluchowski et al., 2017; Schlegel et al., 2018). Although lipotoxicity in the intestinal epithelium leading to mucosal injury may explain clinical features in response to DGAT1mutations, further studies are required to explore the mechanisms. Fat-restricted diet constitutes an appropriate nutrition therapy.
Although DGAT1 and DGAT2 genes in mice are expressed in enterocytes (Buhman et al., 2002; Uchida et al., 2013), there was no evidence of overt fat malabsorption in DGAT1 null mice (Buhman et al., 2002). DGAT1 was not essential for TG absorption and CM synthesis even if a high-fat diet was administered in mice. In fact, DGAT2 may compensate for DGAT1 deficiency. Paradoxically, DGAT1 KO mice displayed resistance to the obesogenic effects of a high-fat diet (Buhman et al., 2002)) and the selective inhibitor JTT-553 of DGAT1 was able to eliminate the rise of plasma TG and CM in rats after olive oil loading (Tomimoto et al., 2015). By contrast, DGAT2–/– mice die within a few hours, likely due to extremely low whole-body TG content and an impaired skin barrier, suggesting a divergent function for the two enzymes (Stone et al., 2004). To determine the specific contribution of each of them on the intestine phenotype, DGAT1 was only expressed in the gut, and its overexpression did not alter TG secretion compared to wild-type mice (Lee et al., 2010). On the other hand, mice with intestine-specific overexpression of DGAT2 have higher intestinal TG (Uchida et al., 2013). Another study was able to demonstrate that DGAT1 and DGAT2 function coordinately to regulate the process of dietary fat absorption by preferentially synthesizing TG for incorporation into distinct subcellular TG pools in enterocytes (Hung et al., 2017). Definitely, significant divergences characterize the human and mouse species, especially taking into account that DGAT1 is central in human intestinal and DGAT2 is the major enzyme of TG synthesis in mice.
As DGAT2 shares no sequence homology with the members of the DGAT1 family, it is important to examine carefully DGAT2 polymorphisms as we have done for DGAT1. When obese children and adolescents and 94 healthy underweight controls were screened for polymorphisms, 15 DNA variants are detected: 4 coding non-synonymous exchanges (p.Val82Ala, p.Arg297Gln, p.Gly318Ser, and p.Leu385Val) and 10 fully synonymous (c.-9447A > G, c.-584C > G, c.-140C > T, c.-30C > T, IVS2-3C > G, c.812A > G, c.920T > C, IVS7+23C > T, IVS7+73C > T, and ∗22C > T) (Friedel et al., 2007). The authors do not find (i) an association between variants or haplotypes and the genomic region of DGAT2, and (ii) an important role of common genetic variation in DGAT2 for the development of obesity. On the other hand, using whole-exome sequencing and biological function examination, an obese subject carried one loss-of-function mutation in FA amide hydrolase and one loss-of-function mutation in DGAT2 (Ning et al., 2017). While inactivation of the former promotes obesity, DGAT2 modification reduced body weight. This is an interesting investigation documenting an interaction model of genetic variants in two distinct genes in relation with obesity modulation. Finally, mutation of DGAT2 leads to Charcot-Marie-Tooth disease, an autosomal-dominant axonal neuropathy with low serum TG concentrations (Hong et al., 2016). Accordingly, mutant DGAT2 overexpression of the mutant DGAT2 overexpression significantly inhibited the proliferation of mouse motor neuron cells (Table 1).
While MGAT1 expression is absent in the small intestine (Cases et al., 1998, 2001; Yen et al., 2002), MGAT2 expression is abundant in mouse gut (Yen et al., 2002) and its activity correlates with the rate of MG absorption (Yen et al., 2015). However, the deletion of MGAT2 did not result in a change in normal quantities of fat absorbed from the small intestine aside from an increased energy expenditure noted in knockout mice (Yen et al., 2009; Nelson et al., 2011). Nevertheless, ablation of MGAT2 specifically in mouse intestine disturbed intestinal TG metabolism and delayed fat absorption (Nelson et al., 2014). In these experimental conditions, the animals were protected against diet-induced weight gain and associated comorbidities.
ANGPTLs constitute a group of proteins, which share structural similarity with angiopoietins, but the absence of the requisite domains do not allow them to bind with the classical angiopoietin receptors, Tie1 or Tie2 (Li and Teng, 2014) (Table 1). The eight members of ANGPTL family play important metabolic roles in diverse biological and pathological processes, including dyslipidemia, IR and wound healing (Basu and Goldberg, 2020). Both ANGPTL3 and ANGPTL4 inhibit lipoprotein lipase (LPL) activity through unfolding and destabilization, leading to its degradation (Sukonina et al., 2006). ANGPTL3 is a 70 kDa protein mostly expressed and secreted by the liver and to a lesser extent by the kidney (Conklin et al., 1999; Romeo et al., 2009), whereas ANGPTL4 glycoprotein is a smaller protein (∼45–65 KDa), which is produced in many cells and tissues, including adipose tissue, liver, intestine, and muscle (Yoon et al., 2000). Although both proteins inhibit LPL activity and raise plasma TG levels, they are regulated by physiological states and different nuclear receptors (Ge et al., 2005). In human, homozygous loss of ANGPTL3 function leads to familial combined hypolipidemia characterized by low plasma levels of TGs, HDL-CHOL, and LDL-CHOL (Shimamura et al., 2007; Xu et al., 2018). Silencing of ANGPTL3 in mouse models and in human hepatoma cells result in reduced output and increased uptake of Apo B-containing lipoproteins (Xu et al., 2018), thereby contributing to low LDL-CHOL observed in mice and humans with genetic ANGPTL3 deficiency (Musunuru et al., 2010). Similarly, ANGPTL4 null mice have decreased plasma TG concentrations, whereas mice overexpressing ANGPTL4 have raised plasma TG levels (Mandard et al., 2006; Lichtenstein et al., 2007). Interestingly, the ablation of ANGPTL4 caused perturbations of intestinal lymphatics, which worsened after feeding a high-fat diet (Desai et al., 2007). Moreover, ANGPTL4 was capable of impeding dietary fat digestion via inhibition of pancreatic lipase whereas its deletion increased fat mass LPL, especially with a ANGPTL4 gene variant for a loss-of-function that leads to hypolipidemia with a reduction of TG-containing lipoproteins (VLDL) and CHOL-carrying lipoproteins such as LDL and HDL (Koishi et al., 2002; Shimizugawa et al., 2002; Teslovich et al., 2010). The complete ANGPTL3 deficiency was associated with a highly reduced postprandial hypertriglyceridemia, probably due to an accelerated catabolism of intestinal derived CM secondary to the increased LPL activity (Minicocci et al., 2016).
Several advances have been made in our understanding of factors responsible for congenital fat malabsorption syndromes. The information currently at hand has led to map out the route of intestinal lipid transport. We certainly appreciate more the mechanisms controlling intra-enterocyte lipid trafficking and CM formation, but there is still a paucity of knowledge related to the processes essential for its extrusion from the Golgi apparatus and absorptive cells. Probably, the delineation of additional genetic defects in the future will lead to a full characterization of the sequential events crucial for CM packaging and output.
EL and SS wrote and edited the manuscript. JFB have made substantial contributions to the manuscript. All authors approved the final version of the manuscript.
This study was supported by the Canadian Institutes of Health Research Grants (PJT 153113), NSERC (RGPIN/04761) and the J. A. DeSève Research Chair in Nutrition.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
ABCG5, ATP-binding cassette G5; ABL, Abetalipoproteinemia; ACAT2, Acylcholesterol acyltransferase-2; ANGPTL4, Angiopoietin-like protein 4; Apo, Apolipoprotein; CE, Cholesteryl ester; CHOL, Cholesterol; CM, Chylomicron; CRD, Chylomicron retention disease; DGAT, Diacylglycerol transferase; EFA, Essential fatty acid; ER, Endoplasmic reticulum; FA, Fatty acid; FATP4, Fatty acid transport protein 4; FHBL, Hypobetalipoproteinemia; GI, Gastrointestinal; GTPase, Guanosine triphosphatase; HDL, High density lipoprotein; I-FABP, Intestinal fatty acid-binding protein; IR, Insulin resistance; KO, Knockout; LDL, Low density lipoprotein; LPL, Lipoprotein lipase; MG, Monoglyceride; MGAT, Monoacylglycerol transferase; MTTP, Microsomal triglyceride transfer protein; NPC1L1, Niemann-Pick C1-Like 1; PCTV, Pre-CM transport vesicle; PL, Phospholipid; SR-BI, Scavenger receptor-BI; TG, Triglyceride; VLDL, Very-low density lipoprotein.
Agren, J. J., Valve, R., Vidgren, H., Laakso, M., and Uusitupa, M. (1998). Postprandial lipemic response is modified by the polymorphism at codon 54 of the fatty acid-binding protein 2 gene. Arterioscler. Thromb. Vasc. Biol. 18, 1606–1610. doi: 10.1161/01.atv.18.10.1606
Alpers, D. H., Bass, N. M., Engle, M. J., and DeSchryver-Kecskemeti, K. (2000). Intestinal fatty acid binding protein may favor differential apical fatty acid binding in the intestine. Biochim. Biophys. Acta 1483, 352–362. doi: 10.1016/s1388-1981(99)00200-0
Anant, S., MacGinnitie, A. J., and Davidson, N. O. (1995). apobec-1, the catalytic subunit of the mammalian apolipoprotein B mRNA editing enzyme, is a novel RNA-binding protein. J. Biol. Chem. 270, 14762–14767. doi: 10.1074/jbc.270.24.14762
Baier, L. J., Sacchettini, J. C., Knowler, W. C., Eads, J., Paolisso, G., Tataranni, P. A., et al. (1995). An amino acid substitution in the human intestinal fatty acid binding protein is associated with increased fatty acid binding, increased fat oxidation, and insulin resistance. J. Clin, Invest. 95, 1281–1287. doi: 10.1172/jci117778
Barlowe, C., Orci, L., Yeung, T., Hosobuchi, M., Hamamoto, S., Salama, N., et al. (1994). COPII: a membrane coat formed by Sec proteins that drive vesicle budding from the endoplasmic reticulum. Cell 77, 895–907. doi: 10.1016/0092-8674(94)90138-4
Basu, D., and Goldberg, I. J. (2020). Regulation of lipoprotein lipase-mediated lipolysis of triglycerides. Curr. Opin. Lipidol. 31, 154–160. doi: 10.1097/mol.0000000000000676
Bell, R. M., and Coleman, R. A. (1980). Enzymes of glycerolipid synthesis in eukaryotes. Annu. Rev. Biochem. 49, 459–487. doi: 10.1146/annurev.bi.49.070180.002331
Bietrix, F., Yan, D., Nauze, M., Rolland, C., Bertrand-Michel, J., Comera, C., et al. (2006). Accelerated lipid absorption in mice overexpressing intestinal SR-BI. J. Biol. Chem. 281, 7214–7219. doi: 10.1074/jbc.m508868200
Black, D. D. (2007). Development and physiological regulation of intestinal lipid absorption. I. Development of intestinal lipid absorption: cellular events in chylomicron assembly and secretion. Am. J. Physiol. Gastrointest. Liver. Physiol. 293, G519–G524.
Black, D. D., Hay, R. V., Rohwer-Nutter, P. L., Ellinas, H., Stephens, J. K., Sherman, H., et al. (1991). Intestinal and hepatic apolipoprotein B gene expression in abetalipoproteinemia. Gastroenterology 101, 520–528. doi: 10.1016/0016-5085(91)90033-h
Bonnefont-Rousselot, D., Condat, B., Sassolas, A., Chebel, S., Bittar, R., Federspiel, M. C., et al. (2009). Cryptogenic cirrhosis in a patient with familial hypocholesterolemia due to a new truncated form of apolipoprotein B. Eur. J. Gastroenterol. Hepatol. 21, 104–108. doi: 10.1097/meg.0b013e3282ffd9f8
Boren, J., Graham, L., Wettesten, M., Scott, J., White, A., and Olofsson, S. O. (1992). The assembly and secretion of ApoB 100-containing lipoproteins in Hep G2 cells. ApoB 100 is cotranslationally integrated into lipoproteins. J. Biol. Chem. 267, 9858–9867. doi: 10.1016/s0021-9258(19)50173-9
Boren, J., Rustaeus, S., and Olofsson, S. O. (1994). Studies on the assembly of apolipoprotein B-100- and B-48-containing very low density lipoproteins in McA-RH7777 cells. J. Biol. Chem. 269, 25879–25888. doi: 10.1016/s0021-9258(18)47328-0
Buhman, K. K., Accad, M., Novak, S., Choi, R. S., Wong, J. S., Hamilton, R. L., et al. (2000). Resistance to diet-induced hypercholesterolemia and gallstone formation in ACAT2-deficient mice. Nat. Med. 6, 1341–1347. doi: 10.1038/82153
Buhman, K. K., Smith, S. J., Stone, S. J., Repa, J. J., Wong, J. S., Knapp, F. F., et al. (2002). DGAT1 is not essential for intestinal triacylglycerol absorption or chylomicron synthesis. J. Biol. Chem. 277, 25474–25479. doi: 10.1074/jbc.m202013200
Buonuomo, P. S., Ruggiero, A., Valeriani, M., and Mariotti, P. (2009). Familial hypobetalipoproteinemia: early neurological, hematological, and ocular manifestations in two affected twins responding to vitamin supplementation. Curr. Opin. Pediatr. 21, 824–827. doi: 10.1097/mop.0b013e32833252f2
Cai, L., Eckhardt, E. R., Shi, W., Zhao, Z., Nasser, M., de Villiers, W. J., et al. (2004). Scavenger receptor class B type I reduces cholesterol absorption in cultured enterocyte CaCo-2 cells. J. Lipid. Res. 45, 253–262. doi: 10.1194/jlr.m300303-jlr200
Carrier, J. C., Deblois, G., Champigny, C., Levy, E., and Giguere, V. (2004). Estrogen-related receptor alpha (ERRalpha) is a transcriptional regulator of apolipoprotein A-IV and controls lipid handling in the intestine. J. Biol. Chem. 279, 52052–52058. doi: 10.1074/jbc.m410337200
Cases, S., Smith, S. J., Zheng, Y. W., Myers, H. M., Lear, S. R., Sande, E., et al. (1998). Identification of a gene encoding an acyl CoA:diacylglycerol acyltransferase, a key enzyme in triacylglycerol synthesis. Proc. Natl. Acad. Sci. U. S. A. 95, 13018–13023. doi: 10.1073/pnas.95.22.13018
Cases, S., Stone, S. J., Zhou, P., Yen, E., Tow, B., Lardizabal, K. D., et al. (2001). Cloning of DGAT2, a second mammalian diacylglycerol acyltransferase, and related family members. J. Biol. Chem. 276, 38870–38876. doi: 10.1074/jbc.m106219200
Cefalu, A. B., Pirruccello, J. P., Noto, D., Gabriel, S., Valenti, V., Gupta, N., et al. (2013). A novel APOB mutation identified by exome sequencing cosegregates with steatosis, liver cancer, and hypocholesterolemia. Arterioscler. Thromb. Vasc. Biol. 33, 2021–2025. doi: 10.1161/atvbaha.112.301101
Charcosset, M., Sassolas, A., Peretti, N., Roy, C. C., Deslandres, C., Sinnett, D., et al. (2008). Anderson or chylomicron retention disease: molecular impact of five mutations in the SAR1B gene on the structure and the functionality of Sar1b protein. Mol. Genet. Metab. 93, 74–84. doi: 10.1016/j.ymgme.2007.08.120
Chen, M., Yang, Y., Braunstein, E., Georgeson, K. E., and Harmon, C. M. (2001). Gut expression and regulation of FAT/CD36: possible role in fatty acid transport in rat enterocytes. Am. J. Physiol. Endocrinol. Metab. 281, E916–E923.
Conklin, D., Gilbertson, D., Taft, D. W., Maurer, M. F., Whitmore, T. E., Smith, D. L., et al. (1999). Identification of a mammalian angiopoietin-related protein expressed specifically in liver. Genomics 62, 477–482. doi: 10.1006/geno.1999.6041
Cuerq, C., Henin, E., Restier, L., Blond, E., Drai, J., Marcais, C., et al. (2018). Efficacy of two vitamin E formulations in patients with abetalipoproteinemia and chylomicron retention disease. J. Lipid. Res. 59, 1640–1648. doi: 10.1194/jlr.m085043
Desai, U., Lee, E. C., Chung, K., Gao, C., Gay, J., Key, B., et al. (2007). Lipid-lowering effects of anti-angiopoietin-like 4 antibody recapitulate the lipid phenotype found in angiopoietin-like 4 knockout mice. Proc. Natl. Acad. Sci. U. S. A. 104, 11766–11771. doi: 10.1073/pnas.0705041104
Di Leo, E., Magnolo, L., Bertolotti, M., Bourbon, M., Carmo Pereira, S., Pirisi, M., et al. (2008). Variable phenotypic expression of homozygous familial hypobetalipoproteinaemia due to novel APOB gene mutations. Clin. Genet. 74, 267–273. doi: 10.1111/j.1399-0004.2008.01023.x
Drover, V. A., Ajmal, M., Nassir, F., Davidson, N. O., Nauli, A. M., Sahoo, D., et al. (2005). CD36 deficiency impairs intestinal lipid secretion and clearance of chylomicrons from the blood. J. Clin. Invest. 115, 1290–1297. doi: 10.1172/jci21514
Dube, N., Delvin, E., Yotov, W., Garofalo, C., Bendayan, M., Veerkamp, J. H., et al. (2001). Modulation of intestinal and liver fatty acid-binding proteins in Caco-2 cells by lipids, hormones and cytokines. J. Cell. Biochem. 81, 613–620. doi: 10.1002/jcb.1090
Field, F. J., Watt, K., and Mathur, S. N. (2007). Ezetimibe interferes with cholesterol trafficking from the plasma membrane to the endoplasmic reticulum in CaCo-2 cells. J. Lipid. Res. 48, 1735–1745. doi: 10.1194/jlr.m700029-jlr200
Fisher, E., Weikert, C., Klapper, M., Lindner, I., Mohlig, M., Spranger, J., et al. (2007). L-FABP T94A is associated with fasting triglycerides and LDL-cholesterol in women. Mol. Genet. Metab. 91, 278–284. doi: 10.1016/j.ymgme.2007.03.002
Florkowski, C., Hedley, J., Bickley, V., Hooper, A. J., Burnett, J. R., and George, P. (2010). Fatty infiltration of the liver in a case of hypobetalipoproteinaemia with a novel mutation in the APOB gene. N. Z. Med. J. 5, 98–100. Epub 2010/04/03.
Friedel, S., Reichwald, K., Scherag, A., Brumm, H., Wermter, A. K., Fries, H. R., et al. (2007). Mutation screen and association studies in the diacylglycerol O-acyltransferase homolog 2 gene (DGAT2), a positional candidate gene for early onset obesity on chromosome 11q13. BMC Genet. 8:17. doi: 10.1186/1471-2156-8-17
Gajda, A. M., Zhou, Y. X., Agellon, L. B., Fried, S. K., Kodukula, S., Fortson, W., et al. (2013). Direct comparison of mice null for liver or intestinal fatty acid-binding proteins reveals highly divergent phenotypic responses to high fat feeding. J. Biol. Chem. 288, 30330–30344. doi: 10.1074/jbc.m113.501676
Gao, N., Qu, X., Yan, J., Huang, Q., Yuan, H. Y., and Ouyang, D. S. (2010). L-FABP T94A decreased fatty acid uptake and altered hepatic triglyceride and cholesterol accumulation in Chang liver cells stably transfected with L-FABP. Mol. Cell Biochem. 345, 207–214. doi: 10.1007/s11010-010-0574-7
Ge, H., Cha, J. Y., Gopal, H., Harp, C., Yu, X., Repa, J. J., et al. (2005). Differential regulation and properties of angiopoietin-like proteins 3 and 4. J. Lipid Res. 46, 1484–1490. doi: 10.1194/jlr.m500005-jlr200
Gertow, K., Bellanda, M., Eriksson, P., Boquist, S., Hamsten, A., Sunnerhagen, M., et al. (2004). Genetic and structural evaluation of fatty acid transport protein-4 in relation to markers of the insulin resistance syndrome. J. Clin. Endocrinol. Metab. 89, 392–399. doi: 10.1210/jc.2003-030682
Gimeno, R. E., Hirsch, D. J., Punreddy, S., Sun, Y., Ortegon, A. M., Wu, H., et al. (2003). Targeted deletion of fatty acid transport protein-4 results in early embryonic lethality. J. Biol. Chem. 278, 49512–49516. doi: 10.1074/jbc.m309759200
Gluchowski, N. L., Chitraju, C., Picoraro, J. A., Mejhert, N., Pinto, S., Xin, W., et al. (2017). Identification and characterization of a novel DGAT1 missense mutation associated with congenital diarrhea. J. Lipid Res. 58, 1230–1237.
Goncalves, A., Gontero, B., Nowicki, M., Margier, M., Masset, G., Amiot, M. J., et al. (2015). Micellar lipid composition affects micelle interaction with class B scavenger receptor extracellular loops. J. Lipid. Res. 56, 1123–1133. doi: 10.1194/jlr.m057612
Gordon, J. I., Elshourbagy, N., Lowe, J. B., Liao, W. S., Alpers, D. H., and Taylor, J. M. (1985). Tissue specific expression and developmental regulation of two genes coding for rat fatty acid binding proteins. J. Biol. Chem. 260, 1995–1998. doi: 10.1016/s0021-9258(18)89501-1
Goudriaan, J. R., Dahlmans, V. E., Febbraio, M., Teusink, B., Romijn, J. A., Havekes, L. M., et al. (2002). Intestinal lipid absorption is not affected in CD36 deficient mice. Mol. Cell Biochem. 239, 199–202. doi: 10.1007/978-1-4419-9270-3_25
Granot, E., and Kohen, R. (2004). Oxidative stress in abetalipoproteinemia patients receiving long-term vitamin E and vitamin A supplementation. Am. J. Clin. Nutr. 79, 226–230. doi: 10.1093/ajcn/79.2.226
Guilmeau, S., Niot, I., Laigneau, J. P., Devaud, H., Petit, V., Brousse, N., et al. (2007). Decreased expression of Intestinal I- and L-FABP levels in rare human genetic lipid malabsorption syndromes. Histochem. Cell Biol. 128, 115–123. doi: 10.1007/s00418-007-0302-x
Haas, J. T., Winter, H. S., Lim, E., Kirby, A., Blumenstiel, B., DeFelice, M., et al. (2012). DGAT1 mutation is linked to a congenital diarrheal disorder. J. Clin. Invest. 122, 4680–4684. doi: 10.1172/jci64873
Heeks, L. V., Hooper, A. J., Adams, L. A., Robbins, P., Barrett, P. H., van Bockxmeer, F. M., et al. (2013). Non-alcoholic steatohepatitis-related cirrhosis in a patient with APOB L343V familial hypobetalipoproteinaemia. Clin. Chim. Acta. 421, 121–125. doi: 10.1016/j.cca.2013.03.004
Hong, Y. B., Kang, J., Kim, J. H., Lee, J., Kwak, G., Hyun, Y. S., et al. (2016). DGAT2 Mutation in a Family with Autosomal-Dominant Early-Onset Axonal Charcot-Marie-Tooth Disease. Hum. Mutat. 37, 473–480. doi: 10.1002/humu.22959
Hung, Y. H., Carreiro, A. L., and Buhman, K. K. (2017). Dgat1 and Dgat2 regulate enterocyte triacylglycerol distribution and alter proteins associated with cytoplasmic lipid droplets in response to dietary fat. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 1862, 600–614. doi: 10.1016/j.bbalip.2017.02.014
Hussain, M. M., Fatma, S., Pan, X., and Iqbal, J. (2005). Intestinal lipoprotein assembly. Curr. Opin. Lipidol. 16, 281–285. doi: 10.1097/01.mol.0000169347.53568.5a
Jones, B., Jones, E. L., Bonney, S. A., Patel, H. N., Mensenkamp, A. R., Eichenbaum-Voline, S., et al. (2003). Mutations in a Sar1 GTPase of COPII vesicles are associated with lipid absorption disorders. Nat. Genet. 34, 29–31. doi: 10.1038/ng1145
Ko, C. W., Qu, J., Black, D. D., and Tso, P. (2020). Regulation of intestinal lipid metabolism: current concepts and relevance to disease. Nat. Rev. Gastroenterol. Hepatol. 17, 169–183. doi: 10.1038/s41575-019-0250-7
Koishi, R., Ando, Y., Ono, M., Shimamura, M., Yasumo, H., Fujiwara, T., et al. (2002). Angptl3 regulates lipid metabolism in mice. Nat. Genet. 30, 151–157. doi: 10.1038/ng814
Lackey, A. I., Chen, T., Zhou, Y. X., Bottasso Arias, N. M., Doran, J. M., Zacharisen, S. M., et al. (2020). Mechanisms underlying reduced weight gain in intestinal fatty acid-binding protein (IFABP) null mice. Am. J. Physiol. Gastrointest. Liver Physiol. 318, G518–G530.
Lagakos, W. S., Gajda, A. M., Agellon, L., Binas, B., Choi, V., Mandap, B., et al. (2011). Different functions of intestinal and liver-type fatty acid-binding proteins in intestine and in whole body energy homeostasis. Am. J. Physiol. Gastrointest. Liver Physiol. 300, G803–G814.
Lally, S., Owens, D., and Tomkin, G. H. (2007). Genes that affect cholesterol synthesis, cholesterol absorption, and chylomicron assembly: the relationship between the liver and intestine in control and streptozotosin diabetic rats. Metabolism 56, 430–438. doi: 10.1016/j.metabol.2006.10.028
Laugerette, F., Passilly-Degrace, P., Patris, B., Niot, I., Febbraio, M., Montmayeur, J. P., et al. (2005). CD36 involvement in orosensory detection of dietary lipids, spontaneous fat preference, and digestive secretions. J. Clin. Invest. 115, 3177–3184. doi: 10.1172/jci25299
Lee, B., Fast, A. M., Zhu, J., Cheng, J. X., and Buhman, K. K. (2010). Intestine-specific expression of acyl CoA:diacylglycerol acyltransferase 1 reverses resistance to diet-induced hepatic steatosis and obesity in Dgat1-/- mice. J. Lipid. Res. 51, 1770–1780. doi: 10.1194/jlr.m002311
Lee, J., and Hegele, R. A. (2014). Abetalipoproteinemia and homozygous hypobetalipoproteinemia: a framework for diagnosis and management. J. Inherit. Metab. Dis. 37, 333–339. doi: 10.1007/s10545-013-9665-4
Lehner, R., and Kuksis, A. (1996). Biosynthesis of triacylglycerols. Prog. Lipid. Res. 35, 169–201. doi: 10.1016/0163-7827(96)00005-7
Levy, E., Harmel, E., Laville, M., Sanchez, R., Emonnot, L., Sinnett, D., et al. (2011). Expression of Sar1b enhances chylomicron assembly and key components of the coat protein complex II system driving vesicle budding. Arterioscler. Thromb. Vasc. Biol. 31, 2692–2699. doi: 10.1161/atvbaha.111.233908
Levy, E., Marcel, Y., Deckelbaum, R. J., Milne, R., Lepage, G., Seidman, E., et al. (1987). Intestinal apoB synthesis, lipids, and lipoproteins in chylomicron retention disease. J. Lipid. Res. 28, 1263–1274. doi: 10.1016/s0022-2275(20)38593-x
Levy, E., Menard, D., Delvin, E., Montoudis, A., Beaulieu, J. F., Mailhot, G., et al. (2009). Localization, function and regulation of the two intestinal fatty acid-binding protein types. Histochem. Cell Biol. 132, 351–367. doi: 10.1007/s00418-009-0608-y
Levy, E., Menard, D., Delvin, E., Stan, S., Mitchell, G., Lambert, M., et al. (2001). The polymorphism at codon 54 of the FABP2 gene increases fat absorption in human intestinal explants. J. Biol. Chem. 276, 39679–39684. doi: 10.1074/jbc.m105713200
Levy, E., Menard, D., Suc, I., Delvin, E., Marcil, V., Brissette, L., et al. (2004). Ontogeny, immunolocalisation, distribution and function of SR-BI in the human intestine. J. Cell Sci. 117, 327–337. doi: 10.1242/jcs.00856
Levy, E., Roy, C. C., Thibault, L., Bonin, A., Brochu, P., and Seidman, E. G. (1994). Variable expression of familial heterozygous hypobetalipoproteinemia: transient malabsorption during infancy. J. Lipid. Res. 35, 2170–2177. doi: 10.1016/s0022-2275(20)39923-5
Levy, E., Spahis, S., Sinnett, D., Peretti, N., Maupas-Schwalm, F., Delvin, E., et al. (2007). Intestinal cholesterol transport proteins: an update and beyond. Curr. Opin. Lipidol. 18, 310–318. doi: 10.1097/mol.0b013e32813fa2e2
Levy, E., Stan, S., Delvin, E., Menard, D., Shoulders, C., Garofalo, C., et al. (2002). Localization of microsomal triglyceride transfer protein in the Golgi: possible role in the assembly of chylomicrons. J. Biol. Chem. 277, 16470–16477.
Li, Y., and Teng, C. (2014). Angiopoietin-like proteins 3, 4 and 8: regulating lipid metabolism and providing new hope for metabolic syndrome. J. Drug. Target. 22, 679–687. doi: 10.3109/1061186x.2014.928715
Li, Z., Jiang, H., Ding, T., Lou, C., Bui, H. H., Kuo, M. S., et al. (2015). Deficiency in lysophosphatidylcholine acyltransferase 3 reduces plasma levels of lipids by reducing lipid absorption in mice. Gastroenterology 149, 1519–1529. doi: 10.1053/j.gastro.2015.07.012
Lichtenstein, L., Berbee, J. F., van Dijk, S. J., van Dijk, K. W., Bensadoun, A., Kema, I. P., et al. (2007). Angptl4 upregulates cholesterol synthesis in liver via inhibition of LPL- and HL-dependent hepatic cholesterol uptake. Arterioscler. Thromb. Vasc. Biol. 27, 2420–2427. doi: 10.1161/atvbaha.107.151894
Lino, M., Farr, S., Baker, C., Fuller, M., Trigatti, B., and Adeli, K. (2015). Intestinal scavenger receptor class B type I as a novel regulator of chylomicron production in healthy and diet-induced obese states. Am. J. Physiol. Gastrointest. Liver Physiol. 309, G350–G359.
Liu, H., Cao, X., Wang, H., Zhao, J., Wang, X., and Wang, Y. (2019). Antimicrobial peptide KR-32 alleviates Escherichia coli K88-induced fatty acid malabsorption by improving expression of fatty acid transporter protein 4 (FATP4)1. J. Anim. Sci. 97, 2342–2356. doi: 10.1093/jas/skz110
Lo, C. C., and Coschigano, K. T. (2020). ApoB48 as an Efficient Regulator of Intestinal Lipid Transport. Front. Physiol. 11:796. doi: 10.3389/fphys.2020.00796
Lobo, M. V., Huerta, L., Ruiz-Velasco, N., Teixeiro, E., de la Cueva, P., Celdran, A., et al. (2001). Localization of the lipid receptors CD36 and CLA-1/SR-BI in the human gastrointestinal tract: towards the identification of receptors mediating the intestinal absorption of dietary lipids. J. Histochem. Cytochem. 49, 1253–1260. doi: 10.1177/002215540104901007
Lowe, J. B., Sacchettini, J. C., Laposata, M., McQuillan, J. J., and Gordon, J. I. (1987). Expression of rat intestinal fatty acid-binding protein in Escherichia coli. Purification and comparison of ligand binding characteristics with that of Escherichia coli-derived rat liver fatty acid-binding protein. J. Biol. Chem. 262, 5931–5937. doi: 10.1016/s0021-9258(18)45664-5
Mandard, S., Zandbergen, F., van Straten, E., Wahli, W., Kuipers, F., Muller, M., et al. (2006). The fasting-induced adipose factor/angiopoietin-like protein 4 is physically associated with lipoproteins and governs plasma lipid levels and adiposity. J. Biol. Chem. 281, 934–944. doi: 10.1074/jbc.m506519200
Mardones, P., Strobel, P., Miranda, S., Leighton, F., Quinones, V., Amigo, L., et al. (2002). Alpha-tocopherol metabolism is abnormal in scavenger receptor class B type I (SR-BI)-deficient mice. J. Nutr. 132, 443–449. doi: 10.1093/jn/132.3.443
Martin, C., Chevrot, M., Poirier, H., Passilly-Degrace, P., Niot, I., and Besnard, P. (2011). CD36 as a lipid sensor. Physiol. Behav. 105, 36–42. doi: 10.1016/j.physbeh.2011.02.029
Martin, G. G., Atshaves, B. P., McIntosh, A. L., Mackie, J. T., Kier, A. B., and Schroeder, F. (2006). Liver fatty acid binding protein gene ablation potentiates hepatic cholesterol accumulation in cholesterol-fed female mice. Am. J. Physiol. Gastrointest. Liver Physiol. 290, G36–G48.
Martin, G. G., Danneberg, H., Kumar, L. S., Atshaves, B. P., Erol, E., Bader, M., et al. (2003). Decreased liver fatty acid binding capacity and altered liver lipid distribution in mice lacking the liver fatty acid-binding protein gene. J. Biol. Chem. 278, 21429–21438. doi: 10.1074/jbc.m300287200
Martin, G. G., McIntosh, A. L., Huang, H., Gupta, S., Atshaves, B. P., Landrock, K. K., et al. (2013). The human liver fatty acid binding protein T94A variant alters the structure, stability, and interaction with fibrates. Biochemistry 52, 9347–9357. doi: 10.1021/bi401014k
Mashek, D. G., and Coleman, R. A. (2006). Cellular fatty acid uptake: the contribution of metabolism. Curr. Opin. Lipidol. 17, 274–278. doi: 10.1097/01.mol.0000226119.20307.2b
Masuda, D., Hirano, K., Oku, H., Sandoval, J. C., Kawase, R., Yuasa-Kawase, M., et al. (2009). Chylomicron remnants are increased in the postprandial state in CD36 deficiency. J. Lipid. Res. 50, 999–1011. doi: 10.1194/jlr.p700032-jlr200
McLeod, R. S., Zhao, Y., Selby, S. L., Westerlund, J., and Yao, Z. (1994). Carboxyl-terminal truncation impairs lipid recruitment by apolipoprotein B100 but does not affect secretion of the truncated apolipoprotein B-containing lipoproteins. J. Biol. Chem. 269, 2852–2862. doi: 10.1016/s0021-9258(17)42021-7
Mendez-Gonzalez, J., Julve, J., Rotllan, N., Llaverias, G., Blanco-Vaca, F., and Escola-Gil, J. C. (2011). ATP-binding cassette G5/G8 deficiency causes hypertriglyceridemia by affecting multiple metabolic pathways. Biochim. Biophys. Acta. 1811, 1186–1193. doi: 10.1016/j.bbalip.2011.07.019
Milger, K., Herrmann, T., Becker, C., Gotthardt, D., Zickwolf, J., Ehehalt, R., et al. (2006). Cellular uptake of fatty acids driven by the ER-localized acyl-CoA synthetase FATP4. J. Cell. Sci. 119, 4678–4688. doi: 10.1242/jcs.03280
Minicocci, I., Tikka, A., Poggiogalle, E., Metso, J., Montali, A., Ceci, F., et al. (2016). Effects of angiopoietin-like protein 3 deficiency on postprandial lipid and lipoprotein metabolism. J. Lipid. Res. 57, 1097–1107. doi: 10.1194/jlr.p066183
Montoudis, A., Delvin, E., Menard, D., Beaulieu, J. F., Jean, D., Tremblay, E., et al. (2006). Intestinal-fatty acid binding protein and lipid transport in human intestinal epithelial cells. Biochem. Biophys. Res. Commun. 339, 248–254. doi: 10.1016/j.bbrc.2005.10.202
Montoudis, A., Seidman, E., Boudreau, F., Beaulieu, J. F., Menard, D., Elchebly, M., et al. (2008). Intestinal fatty acid binding protein regulates mitochondrion beta-oxidation and cholesterol uptake. J. Lipid Res. 49, 961–972. doi: 10.1194/jlr.m700363-jlr200
Murphy, E. J., Prows, D. R., Jefferson, J. R., and Schroeder, F. (1996). Liver fatty acid-binding protein expression in transfected fibroblasts stimulates fatty acid uptake and metabolism. Biochim. Biophys. Acta. 1301, 191–198. doi: 10.1016/0005-2760(96)00024-0
Musunuru, K., Pirruccello, J. P., Do, R., Peloso, G. M., Guiducci, C., Sougnez, C., et al. (2010). Exome sequencing, ANGPTL3 mutations, and familial combined hypolipidemia. N. Engl. J. Med. 363, 2220–2227.
Nakano, T., Inoue, I., and Murakoshi, T. (2019). A Newly Integrated Model for Intestinal Cholesterol Absorption and Efflux Reappraises How Plant Sterol Intake Reduces Circulating Cholesterol Levels. Nutrients 1:11.
Nakano, T., Inoue, I., Takenaka, Y., Ito, R., Kotani, N., Sato, S., et al. (2020). Ezetimibe impairs transcellular lipid trafficking and induces large lipid droplet formation in intestinal absorptive epithelial cells. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 1865:158808. doi: 10.1016/j.bbalip.2020.158808
Nassir, F., Wilson, B., Han, X., Gross, R. W., and Abumrad, N. A. (2007). CD36 is important for fatty acid and cholesterol uptake by the proximal but not distal intestine. J. Biol. Chem. 282, 19493–19501. doi: 10.1074/jbc.m703330200
Neeli, I., Siddiqi, S. A., Siddiqi, S., Mahan, J., Lagakos, W. S., Binas, B., et al. (2007). Liver fatty acid-binding protein initiates budding of pre-chylomicron transport vesicles from intestinal endoplasmic reticulum. J. Biol. Chem. 282, 17974–17984. doi: 10.1074/jbc.m610765200
Nelson, D. W., Gao, Y., Spencer, N. M., Banh, T., and Yen, C. L. (2011). Deficiency of MGAT2 increases energy expenditure without high-fat feeding and protects genetically obese mice from excessive weight gain. J. Lipid. Res. 52, 1723–1732. doi: 10.1194/jlr.m016840
Nelson, D. W., Gao, Y., Yen, M. I., and Yen, C. L. (2014). Intestine-specific deletion of acyl-CoA:monoacylglycerol acyltransferase (MGAT) 2 protects mice from diet-induced obesity and glucose intolerance. J. Biol. Chem. 289, 17338–17349. doi: 10.1074/jbc.m114.555961
Newberry, E. P., Kennedy, S. M., Xie, Y., Sternard, B. T., Luo, J., and Davidson, N. O. (2008). Diet-induced obesity and hepatic steatosis in L-Fabp / mice is abrogated with SF, but not PUFA, feeding and attenuated after cholesterol supplementation. Am. J. Physiol. Gastrointest Liver Physiol. 294, G307–G314.
Newberry, E. P., Xie, Y., Kennedy, S., Han, X., Buhman, K. K., Luo, J., et al. (2003). Decreased hepatic triglyceride accumulation and altered fatty acid uptake in mice with deletion of the liver fatty acid-binding protein gene. J. Biol. Chem. 278, 51664–51672. doi: 10.1074/jbc.m309377200
Newberry, E. P., Xie, Y., Kennedy, S. M., Luo, J., and Davidson, N. O. (2006). Protection against Western diet-induced obesity and hepatic steatosis in liver fatty acid-binding protein knockout mice. Hepatology 44, 1191–1205. doi: 10.1002/hep.21369
Nguyen, T. M., Sawyer, J. K., Kelley, K. L., Davis, M. A., and Rudel, L. L. (2012). Cholesterol esterification by ACAT2 is essential for efficient intestinal cholesterol absorption: evidence from thoracic lymph duct cannulation. J. Lipid Res. 53, 95–104. doi: 10.1194/jlr.m018820
Ning, T., Zou, Y., Yang, M., Lu, Q., Chen, M., Liu, W., et al. (2017). Genetic interaction of DGAT2 and FAAH in the development of human obesity. Endocrine 56, 366–378. doi: 10.1007/s12020-017-1261-1
Noto, D., Cefalu, A. B., Valenti, V., Fayer, F., Pinotti, E., Ditta, M., et al. (2012). Prevalence of ANGPTL3 and APOB gene mutations in subjects with combined hypolipidemia. Arterioscler. Thromb. Vasc. Biol. 32, 805–809. doi: 10.1161/atvbaha.111.238766
Pepino, M. Y., Love-Gregory, L., Klein, S., and Abumrad, N. A. (2012). The fatty acid translocase gene CD36 and lingual lipase influence oral sensitivity to fat in obese subjects. J. Lipid. Res. 53, 561–566. doi: 10.1194/jlr.m021873
Peretti, N., Roy, C. C., Sassolas, A., Deslandres, C., Drouin, E., Rasquin, A., et al. (2009). Chylomicron retention disease: a long term study of two cohorts. Mol. Genet. Metab. 97, 136–142. doi: 10.1016/j.ymgme.2009.02.003
Peretti, N., Sassolas, A., Roy, C. C., Deslandres, C., Charcosset, M., Castagnetti, J., et al. (2010). Guidelines for the diagnosis and management of chylomicron retention disease based on a review of the literature and the experience of two centers. Orphanet. J. Rare Dis. 5:24. doi: 10.1186/1750-1172-5-24
Poirier, H., Degrace, P., Niot, I., Bernard, A., and Besnard, P. (1996). Localization and regulation of the putative membrane fatty-acid transporter (FAT) in the small intestine. Comparison with fatty acid-binding proteins (FABP). Eur. J. Biochem. 238, 368–373. doi: 10.1111/j.1432-1033.1996.0368z.x
Raabe, M., Flynn, L. M., Zlot, C. H., Wong, J. S., Veniant, M. M., Hamilton, R. L., et al. (1998). Knockout of the abetalipoproteinemia gene in mice: reduced lipoprotein secretion in heterozygotes and embryonic lethality in homozygotes. Proc. Natl. Acad. Sci. U. S. A. 95, 8686–8691. doi: 10.1073/pnas.95.15.8686
Ratchford, T. L., Kirby, A. J., Pinz, H., and Patel, D. R. (2018). Congenital Diarrhea From DGAT1 Mutation Leading to Electrolyte Derangements, Protein-losing Enteropathy, and Rickets. J. Pediatr. Gastroenterol. Nutr. 66, 82–83e.
Reboul, E., Abou, L., Mikail, C., Ghiringhelli, O., Andre, M., Portugal, H., et al. (2005). Lutein transport by Caco-2 TC-7 cells occurs partly by a facilitated process involving the scavenger receptor class B type I (SR-BI). Biochem. J. 387, 455–461. doi: 10.1042/bj20040554
Richieri, G. V., Ogata, R. T., and Kleinfeld, A. M. (1994). Equilibrium constants for the binding of fatty acids with fatty acid-binding proteins from adipocyte, intestine, heart, and liver measured with the fluorescent probe ADIFAB. J. Biol. Chem. 269, 23918–23930. doi: 10.1016/s0021-9258(19)51026-2
Rodriguez Sawicki, L., Bottasso Arias, N. M., Scaglia, N., Falomir Lockhart, L. J., Franchini, G. R., Storch, J., et al. (2017). FABP1 knockdown in human enterocytes impairs proliferation and alters lipid metabolism. Biochim. Biophys. Acta Mol. Cell. Biol. Lipids. 1862, 1587–1594. doi: 10.1016/j.bbalip.2017.09.006
Romeo, S., Yin, W., Kozlitina, J., Pennacchio, L. A., Boerwinkle, E., Hobbs, H. H., et al. (2009). Rare loss-of-function mutations in ANGPTL family members contribute to plasma triglyceride levels in humans. J. Clin. Invest. 119, 70–79.
Roy, C. C., Levy, E., Green, P. H., Sniderman, A., Letarte, J., Buts, J. P., et al. (1987). Malabsorption, hypocholesterolemia, and fat-filled enterocytes with increased intestinal apoprotein B. Chylomicron retention disease. Gastroenterology 92, 390–399. doi: 10.1016/0016-5085(87)90133-8
Rustaeus, S., Lindberg, K., Boren, J., and Olofsson, S. O. (1995). Brefeldin A reversibly inhibits the assembly of apoB containing lipoproteins in McA-RH7777 cells. J. Biol. Chem. 270, 28879–28886. doi: 10.1074/jbc.270.48.28879
Sandoval, J. C., Nakagawa-Toyama, Y., Masuda, D., Tochino, Y., Nakaoka, H., Kawase, R., et al. (2010). Molecular mechanisms of ezetimibe-induced attenuation of postprandial hypertriglyceridemia. J. Atheroscler. Thromb. 17, 914–924. doi: 10.5551/jat.4929
Sane, A. T., Sinnett, D., Delvin, E., Bendayan, M., Marcil, V., Menard, D., et al. (2006). Localization and role of NPC1L1 in cholesterol absorption in human intestine. J. Lipid. Res. 47, 2112–2120. doi: 10.1194/jlr.m600174-jlr200
Schlegel, C., Lapierre, L. A., Weis, V. G., Williams, J. A., Kaji, I., Pinzon-Guzman, C., et al. (2018). Reversible deficits in apical transporter trafficking associated with deficiency in diacylglycerol acyltransferase. Traffic 19, 879–892. doi: 10.1111/tra.12608
Shim, J., Moulson, C. L., Newberry, E. P., Lin, M. H., Xie, Y., Kennedy, S. M., et al. (2009). Fatty acid transport protein 4 is dispensable for intestinal lipid absorption in mice. J. Lipid. Res. 50, 491–500. doi: 10.1194/jlr.m800400-jlr200
Shimamura, M., Matsuda, M., Yasumo, H., Okazaki, M., Fujimoto, K., Kono, K., et al. (2007). Angiopoietin-like protein3 regulates plasma HDL cholesterol through suppression of endothelial lipase. Arterioscler. Thromb. Vasc. Biol. 27, 366–372. doi: 10.1161/01.atv.0000252827.51626.89
Shimizugawa, T., Ono, M., Shimamura, M., Yoshida, K., Ando, Y., Koishi, R., et al. (2002). ANGPTL3 decreases very low density lipoprotein triglyceride clearance by inhibition of lipoprotein lipase. J. Biol. Chem. 277, 33742–33748. doi: 10.1074/jbc.m203215200
Shoulders, C. C., Brett, D. J., Bayliss, J. D., Narcisi, T. M., Jarmuz, A., Grantham, T. T., et al. (1993). Abetalipoproteinemia is caused by defects of the gene encoding the 97 kDa subunit of a microsomal triglyceride transfer protein. Hum. Mol. Genet. 2, 2109–2116. doi: 10.1093/hmg/2.12.2109
Siddiqi, S., Saleem, U., Abumrad, N. A., Davidson, N. O., Storch, J., Siddiqi, S. A., et al. (2010). A novel multiprotein complex is required to generate the prechylomicron transport vesicle from intestinal ER. J. Lipid Res. 51, 1918–1928. doi: 10.1194/jlr.m005611
Siddiqi, S. A., Gorelick, F. S., Mahan, J. T., and Mansbach, C. M. II (2003). COPII proteins are required for Golgi fusion but not for endoplasmic reticulum budding of the pre-chylomicron transport vesicle. J. Cell. Sci. 116, 415–427. doi: 10.1242/jcs.00215
Siddiqi, S. A., Siddiqi, S., Mahan, J., Peggs, K., Gorelick, F. S., and Mansbach, C. M. II (2006). The identification of a novel endoplasmic reticulum to Golgi SNARE complex used by the prechylomicron transport vesicle. J. Biol. Chem. 281, 20974–20982. doi: 10.1074/jbc.m601401200
Stahl, A., Hirsch, D. J., Gimeno, R. E., Punreddy, S., Ge, P., Watson, N., et al. (1999). Identification of the major intestinal fatty acid transport protein. Mol. Cell. 4, 299–308. doi: 10.1016/s1097-2765(00)80332-9
Stan, S., Lambert, M., Delvin, E., Paradis, G., O’Loughlin, J., Hanley, J. A., et al. (2005). Intestinal fatty acid binding protein and microsomal triglyceride transfer protein polymorphisms in French-Canadian youth. J. Lipid. Res. 46, 320–327. doi: 10.1194/jlr.m400346-jlr200
Stephen, J., Vilboux, T., Haberman, Y., Pri-Chen, H., Pode-Shakked, B., Mazaheri, S., et al. (2016). Congenital protein losing enteropathy: an inborn error of lipid metabolism due to DGAT1 mutations. Eur. J. Hum. Genet. 24, 1268–1273. doi: 10.1038/ejhg.2016.5
Stone, S. J., Myers, H. M., Watkins, S. M., Brown, B. E., Feingold, K. R., Elias, P. M., et al. (2004). Lipopenia and skin barrier abnormalities in DGAT2-deficient mice. J. Biol. Chem. 279, 11767–11776. doi: 10.1074/jbc.m311000200
Storch, J., and Thumser, A. E. (2010). Tissue-specific functions in the fatty acid-binding protein family. J. Biol. Chem. 285, 32679–32683. doi: 10.1074/jbc.r110.135210
Sukonina, V., Lookene, A., Olivecrona, T., and Olivecrona, G. (2006). Angiopoietin-like protein 4 converts lipoprotein lipase to inactive monomers and modulates lipase activity in adipose tissue. Proc. Natl. Acad. Sci. U. S. A. 103, 17450–17455. doi: 10.1073/pnas.0604026103
Swift, L. L. (1995). Assembly of very low density lipoproteins in rat liver: a study of nascent particles recovered from the rough endoplasmic reticulum. J. Lipid. Res. 36, 395–406. doi: 10.1016/s0022-2275(20)39874-6
Teng, B., Burant, C. F., and Davidson, N. O. (1993). Molecular cloning of an apolipoprotein B messenger RNA editing protein. Science 260, 1816–1819. doi: 10.1126/science.8511591
Teng, B., Verp, M., Salomon, J., and Davidson, N. O. (1990). Apolipoprotein B messenger RNA editing is developmentally regulated and widely expressed in human tissues. J. Biol. Chem. 265, 20616–20620.
Teslovich, T. M., Musunuru, K., Smith, A. V., Edmondson, A. C., Stylianou, I. M., Koseki, M., et al. (2010). Biological, clinical and population relevance of 95 loci for blood lipids. Nature 466, 707–713.
Thumser, A. E., and Storch, J. (2000). Liver and intestinal fatty acid-binding proteins obtain fatty acids from phospholipid membranes by different mechanisms. J. Lipid. Res. 41, 647–656.
Thumser, A. E., and Wilton, D. C. (1996). The binding of cholesterol and bile salts to recombinant rat liver fatty acid-binding protein. Biochem. J. 320, 729–733. doi: 10.1042/bj3200729
Tomimoto, D., Okuma, C., Ishii, Y., Akiyama, Y., Ohta, T., Kakutani, M., et al. (2015). Pharmacological characterization of [trans-5’-(4-amino-7,7-dimethyl-2-trifluoromethyl-7H-pyrimido[4,5-b][1,4]oxazin-6 -yl)-2’,3’-dihydrospiro(cyclohexane-1,1’-inden)-4-yl]acetic acid monobenzenesulfonate (JTT-553), a novel acyl CoA:diacylglycerol transferase (DGAT) 1 inhibitor. Biol. Pharm. Bull. 38, 263–269. doi: 10.1248/bpb.b14-00655
Traber, M. G. (2013). Mechanisms for the prevention of vitamin E excess. J. Lipid. Res. 54, 2295–2306. doi: 10.1194/jlr.r032946
Uchida, A., Slipchenko, M. N., Eustaquio, T., Leary, J. F., Cheng, J. X., and Buhman, K. K. (2013). Intestinal acyl-CoA:diacylglycerol acyltransferase 2 overexpression enhances postprandial triglyceridemic response and exacerbates high fat diet-induced hepatic triacylglycerol storage. Biochim. Biophys. Acta. 1831, 1377–1385. doi: 10.1016/j.bbalip.2013.04.013
van Bennekum, A., Werder, M., Thuahnai, S. T., Han, C. H., Duong, P., Williams, D. L., et al. (2005). Class B scavenger receptor-mediated intestinal absorption of dietary beta-carotene and cholesterol. Biochemistry 44, 4517–4525. doi: 10.1021/bi0484320
van Rijn, J. M., Ardy, R. C., Kuloglu, Z., Harter, B., van Haaften-Visser, D. Y., van der Doef, H. P. J., et al. (2018). Intestinal Failure and Aberrant Lipid Metabolism in Patients With DGAT1 Deficiency. Gastroenterology 155:e115.
Vassileva, G., Huwyler, L., Poirier, K., Agellon, L. B., and Toth, M. J. (2000). The intestinal fatty acid binding protein is not essential for dietary fat absorption in mice. FASEB J. 14, 2040–2046. doi: 10.1096/fj.99-0959com
Velkov, T. (2013). Interactions between Human Liver Fatty Acid Binding Protein and Peroxisome Proliferator Activated Receptor Selective Drugs. PPAR Res. 2013:938401.
Wang, L. R., McIntyre, A. D., and Hegele, R. A. (2018). Complex genetic architecture in severe hypobetalipoproteinemia. Lipids. Health Dis. 17:48.
Wetterau, J. R., Aggerbeck, L. P., Bouma, M. E., Eisenberg, C., Munck, A., Hermier, M., et al. (1992). Absence of microsomal triglyceride transfer protein in individuals with abetalipoproteinemia. Science 258, 999–1001. doi: 10.1126/science.1439810
Wetterau, J. R., and Zilversmit, D. B. (1984). A triglyceride and cholesteryl ester transfer protein associated with liver microsomes. J. Biol. Chem. 259, 10863–10866. doi: 10.1016/s0021-9258(18)90593-4
Whitfield, A. J., Marais, A. D., Robertson, K., Barrett, P. H., van Bockxmeer, F. M., and Burnett, J. R. (2003). Four novel mutations in APOB causing heterozygous and homozygous familial hypobetalipoproteinemia. Hum. Mutat. 22:178. doi: 10.1002/humu.9163
Wolfrum, C., Ellinghaus, P., Fobker, M., Seedorf, U., Assmann, G., Borchers, T., et al. (1999). Phytanic acid is ligand and transcriptional activator of murine liver fatty acid binding protein. J. Lipid. Res. 40, 708–714.
Xu, L., Gu, W., Luo, Y., Lou, J., and Chen, J. (2020). DGAT1 mutations leading to delayed chronic diarrhoea: a case report. BMC Med. Genet. 21:239. doi: 10.1186/s12881-020-01164-1
Xu, Y. X., Redon, V., Yu, H., Querbes, W., Pirruccello, J., Liebow, A., et al. (2018). Role of angiopoietin-like 3 (ANGPTL3) in regulating plasma level of low-density lipoprotein cholesterol. Atherosclerosis 268, 196–206. doi: 10.1016/j.atherosclerosis.2017.08.031
Yamashita, S., Hirano, K., Kuwasako, T., Janabi, M., Toyama, Y., Ishigami, M., et al. (2007). Physiological and pathological roles of a multi-ligand receptor CD36 in atherogenesis; insights from CD36-deficient patients. Mol. Cell Biochem. 299, 19–22. doi: 10.1007/s11010-005-9031-4
Ye, Z., Huang, Y., Zheng, C., Wang, Y., Lu, J., Wang, H., et al. (2019). Clinical and genetic spectrum of children with congenital diarrhea and enteropathy in China. Genet. Med. 21, 2224–2230. doi: 10.1038/s41436-019-0488-z
Yen, C. L., Cheong, M. L., Grueter, C., Zhou, P., Moriwaki, J., Wong, J. S., et al. (2009). Deficiency of the intestinal enzyme acyl CoA:monoacylglycerol acyltransferase-2 protects mice from metabolic disorders induced by high-fat feeding. Nat. Med. 15, 442–446. doi: 10.1038/nm.1937
Yen, C. L., Nelson, D. W., and Yen, M. I. (2015). Intestinal triacylglycerol synthesis in fat absorption and systemic energy metabolism. J. Lipid. Res. 56, 489–501. doi: 10.1194/jlr.r052902
Yen, C. L., Stone, S. J., Cases, S., Zhou, P., and Farese, R. V. Jr. (2002). Identification of a gene encoding MGAT1, a monoacylglycerol acyltransferase. Proc. Natl. Acad. Sci. U. S, A. 99, 8512–8517. doi: 10.1073/pnas.132274899
Yen, C. L., Stone, S. J., Koliwad, S., Harris, C., and Farese, R. V. Jr. (2008). Thematic review series: glycerolipids. DGAT enzymes and triacylglycerol biosynthesis. J. Lipid. Res. 49, 2283–2301. doi: 10.1194/jlr.r800018-jlr200
Yoon, J. C., Chickering, T. W., Rosen, Dussault, B., Qin, Y., Soukas, A., et al. (2000). Peroxisome proliferator-activated receptor gamma target gene encoding a novel angiopoietin-related protein associated with adipose differentiation. Mol. Cell Biol. 20, 5343–5349. doi: 10.1128/mcb.20.14.5343-5349.2000
Keywords: fat digestion, lipid absorption, congenital malabsorption syndromes, chylomicron, intestine
Citation: Levy E, Beaulieu JF and Spahis S (2021) From Congenital Disorders of Fat Malabsorption to Understanding Intra-Enterocyte Mechanisms Behind Chylomicron Assembly and Secretion. Front. Physiol. 12:629222. doi: 10.3389/fphys.2021.629222
Received: 13 November 2020; Accepted: 06 January 2021;
Published: 27 January 2021.
Edited by:
Andromeda M. Nauli, Marshall B. Ketchum University, United StatesReviewed by:
Kimberly K. Buhman, Purdue University, United StatesCopyright © 2021 Levy, Beaulieu and Spahis. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Emile Levy, ZW1pbGUubGV2eS5oc2pAc3Nzcy5nb3V2LnFjLmNh
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.