 Magda Correia1
Magda Correia1 Bruno Bernardes de Jesus
Bruno Bernardes de Jesus Sandrina Nóbrega-Pereira
Sandrina Nóbrega-Pereira- 1Department of Medical Sciences and Institute of Biomedicine – iBiMED, University of Aveiro, Aveiro, Portugal
- 2Instituto de Medicina Molecular João Lobo Antunes, Faculdade de Medicina, Universidade de Lisboa, Lisbon, Portugal
Heart disease is the leading cause of mortality in developed countries. The associated pathology is typically characterized by the loss of cardiomyocytes that leads, eventually, to heart failure. Although conventional treatments exist, novel regenerative procedures are warranted for improving cardiac regeneration and patients well fare. Whereas following injury the capacity for regeneration of adult mammalian heart is limited, the neonatal heart is capable of substantial regeneration but this capacity is lost at postnatal stages. Interestingly, this is accompanied by a shift in the metabolic pathways and energetic fuels preferentially used by cardiomyocytes from embryonic glucose-driven anaerobic glycolysis to adult oxidation of substrates in the mitochondria. Apart from energetic sources, metabolites are emerging as key regulators of gene expression and epigenetic programs which could impact cardiac regeneration. Long non-coding RNAs (lncRNAs) are known master regulators of cellular and organismal carbohydrate and lipid metabolism and play multifaceted functions in the cardiovascular system. Still, our understanding of the metabolic determinants and pathways that can promote cardiac regeneration in the injured hearth remains limited. Here, we will discuss the emerging concepts that provide evidence for a molecular interplay between lncRNAs and metabolic signaling in cardiovascular function and whether exploiting this axis could provide ground for improved regenerative strategies in the heart.
Introduction
The World Health Organization (WHO) has been reporting every year that cardiovascular diseases (CVD) are the leading cause of death in the world. Although currently there are large range of pharmaceutical drugs and surgical options that prevent further deterioration or restore function to the failing heart, for end-stage heart failure, the only long-term selection is heart transplantation which presents several limitations (Hudson and Porrello, 2013). Therefore, the development of improved cardiac regenerative strategies is an area of growing interest.
Subsequent to cardiac injury, cardiomyocytes undergo necrotic and apoptotic cell death and cardiac fibroblasts are activated to produce collagen and other extracellular matrix components, leading to fibrosis and harmed cardiac function (Song et al., 2012; Hashimoto et al., 2018). The main goal of regenerative cardiovascular medicine is to repair injured hearts by replacing cardiomyocytes and diminishing fibrosis. In order to suppress the outcomes of heart failure several regenerative strategies have been proposed, including post-injury activation of cardiomyocyte proliferation, recruitment of stem cells or progenitor cells, delivery of de novo cardiomyocytes from iPSCs, and direct reprogramming of resident cardiac fibroblasts (CFs) into induced cardiac-like myocytes (iCLMs) (Ieda et al., 2010; Song et al., 2012; Ghiroldi et al., 2018; Hashimoto et al., 2018). Although most strategies aim at recovering cardiac function post-injury, approaches that target mechanisms of heart regeneration at pre-injury and during injury events can also potentially be conceived (Uygur and Lee, 2016; Galdos et al., 2017; Tzahor and Poss, 2017).
Besides holding great promise, most cardiac regenerative strategies still lack effective clinical outcomes (Ghiroldi et al., 2017; Hashimoto et al., 2018). Therefore understanding the molecular mechanisms and players governing cardiac regeneration in the injured hearth is warranted for improving the efficiency of cardiac regenerative strategies. In this context, long non-coding RNAs (lncRNAs), a class of > 200 nucleotides-long ribonucleic acid sequences, are abundantly expressed in the cardiovascular system and are part of a complex regulatory network governing cardiovascular function in health and disease (Bär et al., 2016; Das et al., 2018; Hobuß et al., 2019). Essential roles for some lncRNAs in heart development have been described (Matkovich et al., 2014; Bär et al., 2016; Haemmig et al., 2017) and exploring the role of lncRNAs in cardiovascular function may facilitate the development of new therapeutics for treating cardiovascular disease (Bär et al., 2016; Hobuß et al., 2019).
Although the adult mammalian heart has limited regenerative capacity, with estimation of only ≈1% de novo cardiomyogenesis per year (Neidig et al., 2018), the neonatal heart is capable of substantial regeneration but this capacity is lost by postnatal day (P) 7 (Soonpaa et al., 1996). Interestingly, this lost in proliferative potential is accompanied by a shift in the main energetic metabolic pathway and fuels preferentially used by cardiomyocytes from embryonic glucose-driven anaerobic glycolysis to adult oxygen-dependent oxidative phosphorylation (OXPHOS) of pyruvate and fatty acids (FAs) in the mitochondria (Lopaschuk et al., 1992; Lehman and Kelly, 2002). Apart from energetic sources, metabolites are key regulators of gene expression programs by acting as essential substrates or cofactors for chromatin-modifying enzymes (Intlekofer and Finley, 2019). Interestingly, lncRNAs are emerging as master regulators of cellular and systemic carbohydrate and lipid metabolism with clear implications for cardiovascular function (Zhao, 2015; van Soligen, 2018; Mongelli et al., 2019), opening the possibility for a two-way communication between metabolism and lncRNAs in cardiac heart regeneration.
Here, we will discuss the emerging insights in the molecular interplay between lncRNAs and metabolism in the heart highlighting evidence for the impact of metabolic signaling in cardiac regeneration. Also, particular emphasis will be given to those lncRNAs regulating metabolic targets in the cardiovascular system and the potential modulation of the lncRNAs-metabolic axis for the development of innovative regenerative strategies.
Can an Embryonic-Like Metabolic Program Promote Heart Regeneration?
The fetal heart’s environment is low in oxygen and FAs, thus fetal cardiomyocytes are highly dependent on glycolysis for ATP production (Lopaschuk et al., 1992). The heart suffers a major metabolic alteration driven by the physiological changes at postnatal stages, as enhanced workload and the demand for growth, that cannot be supported by glucose and lactate metabolism (Malandraki-Miller et al., 2018). The mammalian heart has to contract constantly thus, the need for an optimal energy fuel is imperative. During the early postnatal period, the number of mitochondria in cardiomyocytes increases dramatically (Mayor and Cuezva, 1985; Attardi and Schatz, 1988). Mitochondria is the organelle that coordinates the energy transduction function and it is responsible to produce more than 95% of ATP utilized by the heart (Doenst et al., 2013). Additionally, mitochondria regulates intracellular calcium homeostasis, signaling and apoptosis (Kolwicz et al., 2013). As a result, mammalian cardiomyocytes undergo extensive metabolic remodeling after birth. In order to adapt to the high-energetic demands of the postnatal life, cardiomyocytes suffer a metabolic switch and produce their energy via mitochondrial OXPHOS, a more efficient process than glycolysis (Lehman and Kelly, 2002; Vivien et al., 2016). Postnatal cardiomyocytes also revealed a shift in the energetic substrate utilization from pyruvate to FAs that are energetically more favorable (Lopaschuk et al., 1992; Lehman and Kelly, 2002). As the neonatal mammalian heart regenerative capacity is lost by P7, which corresponds with cardiomyocyte binucleation and cell-cycle arrest (Soonpaa et al., 1996; Porrello et al., 2011), one could hypothesize that the “fetal metabolic shift” would have a role in suppressing cardiomyocyte proliferation and heart repair (Martik, 2020). Currently many studies are focusing in understanding the role of mitochondrial metabolism in regulating cell-cycle arrest in postnatal cardiomyocytes with future implications in regenerative strategies.
Heart regeneration in zebrafish is incredibly effective and relies on the proliferation of pre-existing cardiomyocytes. Apart from cardiomyocytes, other cell types (such as epicardial, endocardial, immune cells and fibroblasts) respond to the heart injury and contribute for the healing process (Vivien et al., 2016; Honkoop et al., 2019). Cardiomyocytes from highly regenerative species such as zebrafish have a preference for glycolysis and increase OXPHOS activity promotes cardiomyocyte maturation and reduces the proliferative capacity (Vivien et al., 2016; Honkoop et al., 2019; Fukuda et al., 2019). Although the “fetal switch” to mitochondrial respiration has been associated to loss of the regenerative potential (Malandraki-Miller et al., 2018), the role of bioenergetics in regulating cardiogenesis remains unclear. Recent evidence suggest that hypoxia inducible factor 1 (HIF1) signaling, an important inducer of aerobic glycolysis and the Warburg effect in cancer cells (Kroemer and Pouyssegur, 2008), controls the embryonic switch toward oxidative metabolism in the developing mouse heart (Menendez-Montes et al., 2016). In midgestational mouse heart, the compact myocardium downregulates HIF1α and switches toward oxidative metabolism. Deletion of the E3 ubiquitin ligase von Hippel-Lindau (VHL) results in HIF1α hyperactivation, blocking the midgestational metabolic shift and impairing cardiac maturation and function (Menendez-Montes et al., 2016). This study highlights the VHL-HIF-mediated metabolic program as an important axis for myocardial differentiation and its potential relevance for cardiac regeneration.
The adult mammalian heart cannot regenerate lost or damaged myocardium although it does present a limited myocyte turnover that reveals insufficient for restoring contractile dysfunction. The brief window of regenerative response following injury seems to be also driven by proliferation of pre-existing cardiomyocytes (Porrello et al., 2011; Elhelaly et al., 2019). Strikingly, increase production of mitochondrial-derived reactive oxygen species (ROS) and DNA oxidation leads to cell-cycle arrest in mouse postnatal cardiomyocytes through the activation of DNA damage response pathways (Puente et al., 2014). FAs oxidation is directly linked to high ROS production and cardiomyocyte cell-cycle arrest (Cardoso et al., 2020). Moreover, the constant use of FAs as an energetic fuel provokes a dependency on this substrate as the acetyl-CoA produced from FAs oxidation inhibits the mitochondrial enzyme pyruvate dehydrogenase (PDH) and therefore the reliance on glucose and its metabolites for energy (Rindler et al., 2013). Still, whether modulating substrate utilization can directly impact DNA damage and promote cardiomyocytes cell-cycle re-entry needs further clarification. Supplementation of FAs-depleted diets to mice prolongs the postnatal window for cardiomyocyte proliferation; however, it is associated with a marked hepatomegaly and steatosis due to a compensatory increase in hepatic de novo fatty-acid synthesis (Cardoso et al., 2020). Moreover, deletion of the dehydrogenase kinase isoform 4 (PDK4) in adult cardiomyocytes, the main enzyme responsible for PDH inhibition and FAs usage, results in a marked shift in myocardial substrate utilization with decrease FAs and enhanced pyruvate-driven glucose oxidation, resulting in less DNA damage and increased cardiomyocyte proliferation (Cardoso et al., 2020). Pharmacological activation of PDH through administration of dichloroacetate in mice also resulted in improved glucose utilization and cardioprotective features (Cardoso et al., 2020).
In sum, recent studies are beginning to dissect the intricate relationship between the “fetal metabolic switch” and loss of cardiomyocyte proliferation where several molecular axis (as HIF signaling, ROS and bioenergetic fuels) are emerging as key regulators. This raises important questions and opportunities in the field. For instance, cardiac regenerative strategies based on the generation of induced cardiac-like myocytes (iCLMs) from iPSCs or resident cardiac fibroblasts (CFs) (Hashimoto et al., 2018), could be improved by metabolic modulation and induction of the “fetal switch”. Moreover, systemic metabolic shifts, as nutritional stages and diets, may impact cardiac regeneration post-injury in the mammalian heart (Malandraki-Miller et al., 2018). Further investigation in this field is warranted and may provide unique opportunities to boost cardiac regeneration and repair.
LncRNAs Controlling Metabolic Pathways in the Heart
LncRNAs represent one of the most prominent but least understood transcriptome in the heart. Thousands of lncRNAs have been identified to be dynamically transcribed during development, differentiation, and maturation of cardiac myocytes (Devaux et al., 2015; He et al., 2016; Li et al., 2017; Beermann et al., 2018). Due to their unique regulatory action and tissue-specific expression, lncRNAs are attractive candidates for diagnosis of cardiovascular pathologies and regenerative strategies using several lncRNA-based therapeutic approaches (Bär et al., 2016; Bernardes de Jesus et al., 2018; Hobuß et al., 2019). lncRNAs are localized in the nucleus or the cytoplasm where they may regulate gene expression at transcriptional or posttranscriptional level, respectively, through diverse mechanisms; including epigenetic remodeling, transcriptional activation or repression (signal, decoy, guide, scaffold, or enhancer lncRNAs), formation and maintenance of sub-nuclear domains, posttranscriptional regulation and modulation of protein activity (Schonrock et al., 2012; Kornfeld and Brüning, 2014; Devaux et al., 2015; Thum and Condorelli, 2015; Muret et al., 2019).
LncRNAs are emerging as master regulators of cellular and organismal carbohydrate and lipid metabolism in adipose tissue and liver (Kornfeld and Brüning, 2014; Zhao, 2015; van Soligen, 2018; Mongelli et al., 2019; Muret et al., 2019). Alteration in serum lipid levels is one of the most relevant risk factor for CVD (Doggen et al., 2004). In the recent years, several studies have highlighted the complex contribution of lncRNAs in controlling systemic and cell-type-specific cholesterol, FAs, and triglyceride metabolism, with important implications for CVD. For instance, several lncRNAs, including H19, lncRNA HCV regulated 1 (lncHR1), MALAT-1 and lncARSR, have been shown to regulate the expression of the sterol regulatory element binding protein 1c (SREBP-1c), a transcription factor that regulates lipid synthesis and uptake in the liver (Yan et al., 2016; Li et al., 2018; Liu et al., 2018; Zhang et al., 2018). Other examples are the liver-specific triglyceride regulator lncRNA Lancaster (lncLSTR) that regulates triglyceride plasma levels and energy metabolism (Li et al., 2015) and AT102202 that inhibits cholesterol synthesis in the liver by targeting the rate limiting enzyme HMGCR (Liu et al., 2015). Whether lncRNAs-mediated control of systemic lipid metabolism has a direct impact in cardiac regeneration remains to be addressed.
As previously discussed, of particular interest are the lncRNAs controlling the “fetal metabolic switch” from embryonic glycolysis to adult mitochondrial respiration and the preferred usage of FAs as energetic fuel in differentiated cardiomyocytes. Although most of our knowledge in lncRNAs control of metabolism comes from studies in lipogenic tissues and/or cancer energetics (Gomes et al., 2019), some mechanistic insights in cardiac muscle development and function, particularly concerning mitochondrial metabolism, are beginning to arise (Table 1). Due to the implication of mitochondrial-dependent FAs oxidation and ROS production in the loss of cardiomyocyte proliferation (Puente et al., 2014; Cardoso et al., 2020), lncRNAs regulating these pathways are particularly attractive for cardiac regeneration.
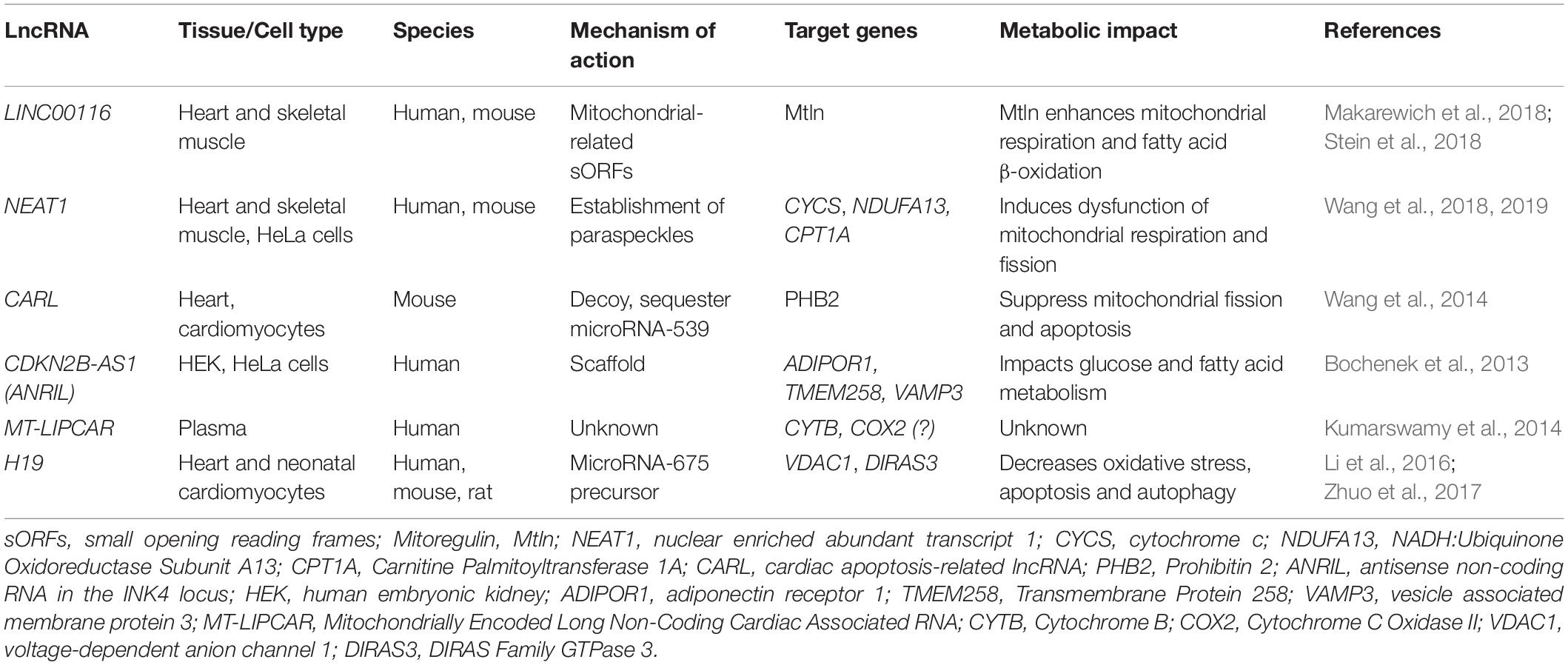
Table 1. LncRNAs with characterized and/or potential metabolic targets in the heart.
In heart and skeletal muscle aged tissue, the lncRNA LINC00116 is among the most significantly downregulated gene (GEO: GSE362 and GSE674). Interestingly, a small region of the most predominant isoform is actively translated in human and mouse muscle and has been found to encode a highly conserved transmembrane microprotein, named mitoregulin (Mtln), also known as Micropeptide regulator of β-oxidation (MOXI), where it associates with the mitochondrial trifunctional protein (MTP), an enzyme complex that plays a critical role in fatty acid β-oxidation (Makarewich et al., 2018; Stein et al., 2018). Isolated heart and skeletal muscle mitochondria from MOXI knockout mice preferentially oxidize carbohydrates over fatty acids, while transgenic MOXI overexpression leads to enhanced β-oxidation. MOXI knockout mice also exhibit a profound reduction in exercise capacity, highlighting the role of MOXI in metabolic control (Makarewich et al., 2018). The impact of Mtln expression in cardiovascular disease and regeneration is still unclear but GTEx portal annotates the existence of common genetic variants that strongly associate with LINC00116 expression in the human heart (Stein et al., 2018). NEAT1 (nuclear enriched abundant transcript 1) is another lncRNA with increase expression in non-regenerative cardiomyocytes (Table 1). In skeletal muscle, NEAT1 modulates myogenesis by accelerating myoblast proliferation and suppressing myoblast differentiation and fusion (Wang et al., 2019). NEAT1 act by recruiting EZH2 to target gene promoters, decreasing the expression of the cyclin-dependent kinase inhibitor p21 and suppressing the myoblast differentiation program. Strikingly, several mitochondrial regulators have been identified to associate to NEAT1 in paraspeckles, a type of nuclear body with multiple roles in gene expression (Wang et al., 2018). Specifically, NEAT1 depletion lead to profound effects on mitochondrial dynamics and function by altering the paraspeckles-specific sequestration of essential mito-mRNAs, including CYCS (cytochrome c), NDUFA13 (NADH:Ubiquinone Oxidoreductase Subunit A13) and CPT1A (Carnitine Palmitoyltransferase 1A) (Wang et al., 2018) and NEAT1-depleted HeLa cells show reduced mitochondrial DNA, ATP production and proliferation rate (Wang et al., 2018).
Cardiac muscle is an extremely metabolically active tissue that undergoes significant changes in energy metabolism in disease. In mouse cardiomyocytes, cardiac apoptosis-related lncRNA (CARL) binds to and sequester microRNA-539, a microRNA found to target the mRNA of the PHB2 sub-unit of prohibitin, a protein localized to the inner mitochondrial membrane that regulates mitochondrial homeostasis (Wang et al., 2014). Downregulation of PHB2 during pathological insults was found to be dependent on upregulation of microRNA-539. CARL acts as the endogenous sponge for this microRNA suppressing mitochondrial fission and cardiomyocyte apoptosis (Wang et al., 2014), highlighting the therapeutic potential of modulating lncRNAs during myocardial infarction. The lncRNA CDKN2B−AS1 (also known as ANRIL) has been described as a genetic risk factor for coronary artery disease (CAD) (Deloukas et al., 2013). ANRIL expression level is associated with left ventricular dysfunction after myocardial infarction (Vausort et al., 2014). Experimental manipulation in several human cell lines (including HEK and HeLa), revealed that ANRIL knock-down decreases the expression of ADIPOR1 (adiponectin receptor 1), TMEM258 (also known as C11ORF10 for chromosome 11 open reading frame 10) and VAMP3 (vesicle associated membrane protein 3), both at the transcript and protein level, which are important genes in the regulation of glucose and fatty-acid metabolism (Bochenek et al., 2013). However, the impact of ANRIL-mediated metabolic regulation in cardiomyocytes remains to be explored. Conversely, in patients with myocardial infarction the levels of the lncRNA hypoxia inducible factor 1A antisense RNA 2 (HIF1A-AS2) was found to be upregulated (Vausort et al., 2014). In humans, the HIF pathway is induced early in acute myocardial and remains activated in chronic human heart failure (Zolk et al., 2008). Due to the role of the HIF signaling in controlling myocardial metabolism and differentiation in the neonatal heart (Menendez-Montes et al., 2016) and the implication of the lncRNA lincRNA-p21 in hypoxia-enhanced glycolysis (Yang et al., 2014), manipulation of the lncRNA/HIF regulatory network might constitute an attractive target to modulate cardiac regeneration.
Type 2 diabetes (T2D) is a multifactorial disorder characterized, among other aspects, by high blood glucose and lipid levels (hyperglycemia and hyperlipidemia) in association with insulin resistance and atherosclerosis (Bornfeldt and Tabas, 2011) and diabetic cardiomyopathy (DCM) is a critical complication of T2D (Jia et al., 2018). Studies suggest that lncRNAs that regulate metabolic targets are aberrantly regulated in DCM, thus targeting lncRNAs could have potential implications for DCM diagnosis and therapy. The mitochondrial long intergenic non-coding RNA predicting cardiac remodeling (MT-LIPCAR) is a lncRNA possibly transcribed from mitochondrial DNA that cross the membrane barrier being released into circulation (Dorn, 2014). Plasma levels of MT-LIPCAR were positively associated with left ventricular diastolic dysfunction in T2D patients with DCM showing prognostic value as an indicator of heart failure and patient mortality. MT-LIPCAR was the first proof that plasma lncRNAs might be used for cardiovascular disease prognostic (Kumarswamy et al., 2014). Despite the invaluable potential as a cardiac biomarker, MT-LIPCAR targets and metabolic impact remains unclear. Evidence suggest that the complete MT-LIPCAR sequence could map to the mitochondrial genes CYTB (Mitochondrially Encoded Cytochrome B) and COX2 (Mitochondrially Encoded Cytochrome C Oxidase II) (Dorn, 2014), raising further questions regarding MT-LIPCAR biogenesis as a mitochondrial or nuclear pseudogene transcript. H19 is a lncRNA transcribed from H19/insulin-like growth factor-II (IGF2) genomic imprinted cluster which accumulates in cardiomyocytes of the mature myocardium in humans and rodents (Pant et al., 2018; Viereck et al., 2020). Decrease expression of cardiac H19 was reported in a rat model of DCM (Li et al., 2016; Zhuo et al., 2017). Overexpression of H19 in myocardial tissue was able to suppress oxidative stress, inflammation and improve left ventricular function leading to DCM amelioration. Mechanistically, H19 serves as template for microRNA-675 expression from its first exon (Zhang et al., 2017; Pant et al., 2018). Since microRNA-675 has multiple biological targets, H19 is able to regulate a number of mitochondrial functions including suppression of apoptosis by targeting voltage-dependent anion channel 1 (VDAC1) (Li et al., 2016), and inhibiting autophagy in cardiomyocytes exposed to high glucose through down-regulation of the GTP- binding protein Di-Ras-3 (DIRAS3) (Zhuo et al., 2017).
In sum, recent work on lncRNAs has started to shed light on their regulatory potential in controlling heart metabolism in health and disease, opening the possibility to explore lncRNAs-mediated metabolic control as a strategy to improve cardiac regeneration and heart function.
LncRNAs and Metabolites as Central Epigenetic Players in Gene Expression Regulation
An hallmark function of lncRNAs is their ability to mediate epigenetic regulation and in the heart, lncRNAs have crucial roles in regulating cardiac chromatin structure during development and pathological remodeling (Schonrock et al., 2012). lncRNAs exhibit tissue-specific regulated expression patterns which are frequently lost during disease (Cabili et al., 2011). However, the regulation of lncRNAs expression during different stages of cardiac development and in disease is still under investigation. Strikingly, inhibition of epigenetic modifications was shown to impact the expression pattern of lncRNAs (Schonrock et al., 2012). Metabolites are emerging as key regulators of gene expression programs and epigenetic modifications, acting as essential substrates or cofactors for enzymes that deposit or remove chemical modifications in DNA and/or histones (Intlekofer and Finley, 2019). FAs and cholesterol have been shown to regulate lncRNAs expression in lipogenic tissues placing metabolism as a central regulator of epigenetic-driven lncRNAs transcription. For instance, the expression of the lncRNAs H19 and MALAT1 is upregulated by FAs exposure, coinciding with an increase in (SREBP)-1c in hepatic cells (van Soligen, 2018) and HULC is induced by cholesterol in hepatoma cells via the retinoic receptor RXRA, leading to lipogenesis (Cui et al., 2015). Recent evidence suggests that lipid metabolism also impact lncRNAs expression in the cardiovascular system. For instance, the lncRNA CHROME, a master regulator of cholesterol homeostasis, is upregulated in atherosclerotic vascular disease in non-human primates and conversely, CHROME expression is influenced by dietary and cellular cholesterol (Hennessy et al., 2019). Although evidence for the direct implication of nutritional signals in the epigenetic alterations that govern lncRNAs expression in the heart is still at its early days, it seems clear that lncRNAs and metabolic signaling can engage in a two-way communication road in the control of gene expression that impacts cellular and systemic metabolism. Moreover, nutritional cues have been shown to control the specification of skeletal cell fate, highlighting the possibility for a similar network to take place in cardiomyocyte progenitors. When lipids are scarce, skeletal muscle progenitors activate the expression of forkhead box O (FOXO) transcription factors leading to a Sox9-dependent suppression of FAs oxidation and chondrogenic commitment (van Gastel et al., 2020). Moreover, glucose metabolism is crucial for muscle stem cells (MuSCs) commitment. In proliferating MuSCs, glucose is dispensable for mitochondrial respiration and becomes available for maintaining high histone acetylation via acetyl-CoA, whereas differentiating MuSCs increases glucose oxidation and has consequently reduced acetylation (Yucel et al., 2019). PDH is pivotal for this switch and determines the differentiation potential of myogenic progenitors during muscle regeneration (Yucel et al., 2019). Whether metabolic fuels also directly impinge cardiomyocyte cell fate decisions and dietary cues can modulate cardiac regeneration (for instance, by controlling lncRNAs expression) are exciting possibilities that require further investigation.
Concluding Remarks
Given the emerging regulatory potential of lncRNAs, it is undoubted that these molecules offer potential solutions in the pursuit for cardiac regeneration (Hudson and Porrello, 2013). In the recent years, several lncRNAs with characterized and/or potential metabolic targets in the heart have been identified (Table 1) and a link between metabolic pathways and cardiac proliferative potential has been established. But can we boost cardiac regeneration by modulating the lncRNAs-metabolic axis? Emerging evidence suggests that exploring the two-way communication road between lncRNAs and cardiac (or systemic) metabolism may offer new perspectives and opportunities for increasing the regenerative potential of the injured heart.
Author Contributions
MC, BB, and SN-P planned, wrote, and discussed the manuscript. BB and SN-P revised the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by Fundação para a Ciência e Tecnologia (FCT), and FEDER, LISBOA-01-0145-FEDER-028534, project co-funded by FEDER, through POR Lisboa 2020—Programa Operacional Regional de Lisboa, PORTUGAL 2020.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Attardi, G., and Schatz, G. (1988). Biogenesis of mitochondria. Annu. Rev. Cell Biol. 4, 289–333. doi: 10.1146/annurev.cb.04.110188.001445
Bär, C., Chatterjee, S., and Thum, T. (2016). Long noncoding RNAs in cardiovascular pathology. Diagn. Ther.Circ. 134, 1484–1499. doi: 10.1161/CIRCULATIONAHA.116.023686
Beermann, J., Kirste, D., Iwanov, K., Lu, D., Kleemiß, F., Kumarswamy, R., et al. (2018). A large shRNA library approach identifies lncRNA Ntep as an essential regulator of cell proliferation. Cell Death Differ. 25, 307–318. doi: 10.1038/cdd.2017.158
Bernardes de Jesus, B., Marinho, S. P., Barros, S., Sousa-Franco, A., Alves-Vale, C., Carvalho, T., et al. (2018). Silencing of the lncRNA Zeb2-NAT facilitates reprogramming of aged fibroblasts and safeguards stem cell pluripotency. Nat. Commun. 9:94. doi: 10.1038/s41467-017-01921-6
Bochenek, G., Häsler, R., Mokhtari, N. E., El König, I. R., Loos, B. G., Jepsen, S., et al. (2013). The large non-coding RNA ANRIL, which is associated with atherosclerosis, periodontitis and several forms of cancer, regulates ADIPOR1, VAMP3 and C11ORF10. Hum. Mol. Genet. 22, 4516–4527. doi: 10.1093/hmg/ddt299
Bornfeldt, K. E., and Tabas, I. (2011). Insulin resistance, hyperglycemia, and atherosclerosis. Cell Metab. 14, 575–585. doi: 10.1016/j.cmet.2011.07.015
Cabili, M., Trapnell, C., Goff, L., Koziol, M., Tazon-Vega, B., Regev, A., et al. (2011). Integrative annotation of human large intergenic noncoding RNAs reveals global properties and specific subclasses. Genes Dev. 25, 1915–1927. doi: 10.1101/gad.17446611
Cardoso, A. C., Lam, N. T., Savla, J. J., Nakada, Y., Pereira, A. H. M., Elnwasany, A., et al. (2020). Mitochondrial substrate utilization regulates cardiomyocyte cell-cycle progression. Nat. Metab. 2, 167–178. doi: 10.1038/s42255-020-0169-x
Cui, M., Xiao, Z., Wang, Y., Zheng, M., Song, T., Cai, X., et al. (2015). Long noncoding RNA HULC modulates abnormal lipid metabolism in hepatoma cells through an miR-9-mediated RXRA signaling pathway. Cancer Res. 75, 846–857. doi: 10.1158/0008-5472.CAN-14-1192
Das, A., Samidurai, A., and Salloum, F. N. (2018). Deciphering non-coding RNAs in cardiovascular health and disease. Front. Cardiovasc. Med. 5:73. doi: 10.3389/fcvm.2018.00073
Deloukas, P., Kanoni, S., Willenborg, C., Farrall, M., Assimes, T. L., Thompson, J. R., et al. (2013). Large-scale association analysis identifies new risk loci for coronary artery disease. Nat. Genet. 45, 25–33. doi: 10.1038/ng.2480
Devaux, Y., Zangrando, J., Schroen, B., Creemers, E. E., Pedrazzini, T., Chang, C. P., et al. (2015). Long noncoding RNAs in cardiac development and ageing. Nat. Rev. Cardiol. 12, 415–425. doi: 10.1038/nrcardio.2015.55
Doenst, T., Nguyen, T. D., and Abel, E. D. (2013). Cardiac metabolism in heart failure. Circ. Res. 113, 709–724. doi: 10.1161/circresaha.113.300376
Doggen, C. J. M., Smith, N. L., Lemaitre, R. N., Heckbert, S. R., Rosendaal, F. R., and Psaty, B. M. (2004). Serum lipid levels and the risk of venous thrombosis. Arterioscler. Thromb. Vasc. Biol. 24, 1970–1975. doi: 10.1161/01.ATV.0000143134.87051.46
Dorn, G. W. II (2014). LIPCAR: a mitochondrial lnc in the noncoding RNA chain? Circ. Res. 114, 1548–1550. doi: 10.1161/CIRCRESAHA.114.304028
Elhelaly, W. M., Cardoso, A. C., Pereira, A. H. M., Elnawasany, A., Ebrahimi, S., Nakada, Y., et al. (2019). C-Kit cells do not significantly contribute to cardiomyogenesis during neonatal heart regeneration. Circulation 139, 559–561. doi: 10.1161/CIRCULATIONAHA.117.033150
Fukuda, R., Aharonov, A., Ong, Y. T., Stone, O. A., El-Brolosy, M. A., Maischein, H. M., et al. (2019). Metabolic modulation regulates cardiac wall morphogenesis in zebrafish. eLife 8, 1–14. doi: 10.7554/eLife.50161
Galdos, F. X., Guo, Y., Paige, S. L., Vandusen, N. J., Wu, S. M., and Pu, W. T. (2017). Cardiac regeneration: lessons from development. Circ. Res. 120, 941–959. doi: 10.1161/CIRCRESAHA.116.309040
Ghiroldi, A., Piccoli, M., Ciconte, G., Pappone, C., and Anastasia, L. (2017). Regenerating the human heart: direct reprogramming strategies and their current limitations. Basic Res. Cardiol. 112, 1–14. doi: 10.1007/s00395-017-0655-9
Ghiroldi, A., Piccoli, M., Cirillo, F., Monasky, M. M., Ciconte, G., Pappone, C., et al. (2018). Cell-Based therapies for cardiac regeneration: a comprehensive review of past and ongoing strategies. Int. J. Mol. Sci. 19:3194. doi: 10.3390/ijms19103194
Gomes, C. P., Nóbrega-Pereira, S., Domingues-Silva, B., Rebelo, K., Alves-Vale, C., Marinho, S. P., et al. (2019). An antisense transcript mediates MALAT1 response in human breast cancer. BMC Cancer 19:771. doi: 10.1186/s12885-019-5962-0
Haemmig, S., Simion, V., Yang, D., Deng, Y., and Feinberg, M. W. (2017). Long noncoding RNAs in cardiovascular disease, diagnosis, and therapy. Curr. Opin. Cardiol. 32, 776–783. doi: 10.1097/HCO.0000000000000454
Hashimoto, H., Olson, E. N., and Bassel-Duby, R. (2018). Therapeutic approaches for cardiac regeneration and repair. Nat. Rev. Cardiol. 15, 585–600. doi: 10.1038/s41569-018-0036-6
He, C., Hu, H., Wilson, K. D., Wu, H., Feng, J., Xia, S., et al. (2016). Systematic characterization of long noncoding RNAs reveals the contrasting coordination of cis- and trans-molecular regulation in human fetal and adult hearts. Circ. Cardiovasc. Genet. 9, 110–118. doi: 10.1161/CIRCGENETICS.115.001264
Hennessy, E. J., van Solingen, C., Scacalossi, K. R., Ouimet, M., Afonso, M. S., Prins, J., et al. (2019). The long noncoding RNA CHROME regulates cholesterol homeostasis in primates. Nat. Metab. 1, 98–110. doi: 10.1038/s42255-018-0004-9
Hobuß, L., Bär, C., and Thum, T. (2019). Long non-coding RNAs: at the heart of cardiac dysfunction? Front. Physiol. 10:30. doi: 10.3389/fphys.2019.00030
Honkoop, H., de Bakker, D. E. M., Aharonov, A., Kruse, F., Shakked, A., Nguyen, P. D., et al. (2019). Single-cell analysis uncovers that metabolic reprogramming by ErbB2 signaling is essential for cardiomyocyte proliferation in the regenerating heart. eLife 8, 1–27. doi: 10.7554/eLife.50163
Hudson, J. E., and Porrello, E. R. (2013). The non-coding road towards cardiac regeneration. J Cardiovasc Transl Res. 6, 909–923. doi: 10.1007/s12265-013-9486-8
Ieda, M., Fu, J., Delgado-olguin, P., Vedantham, V., Hayashi, Y., Bruneau, B. G., et al. (2010). Direct reprogramming of fibroblasts into functional cardiomyocytes by defined factors. Cell 142, 375–386. doi: 10.1016/j.cell.2010.07.002
Intlekofer, A. M., and Finley, L. W. S. (2019). Metabolic signatures of cancer cells and stem cells. Nat. Metab. 1, 177–188. doi: 10.1038/s42255-019-0032-0
Jia, G., Hill, M. A., and Sowers, J. R. (2018). Diabetic cardiomyopathy: an update of mechanisms contributing to this clinical entity. Circ. Res. 122, 624–638. doi: 10.1161/CIRCRESAHA.117.311586
Kolwicz, S. C., Purohit, S., and Tian, R. (2013). Cardiac metabolism and its interactions with contraction, growth, and survival of cardiomyocytes. Circ. Res. 113, 603–616. doi: 10.1161/CIRCRESAHA.113.302095
Kornfeld, J. W., and Brüning, J. C. (2014). Regulation of metabolism by long, non-coding RNAs. Front. Genet. 5:57. doi: 10.3389/fgene.2014.00057
Kroemer, G., and Pouyssegur, J. (2008). Tumor cell metabolism: cancer’s achilles’. Heel. Cancer Cell 13, 472–482. doi: 10.1016/j.ccr.2008.05.005
Kumarswamy, R., Bauters, C., Volkmann, I., Maury, F., Fetisch, J., Holzmann, A., et al. (2014). Circulating long noncoding RNA, LIPCAR, predicts survival in patients with heart failure. Circ. Res. 114, 1569–1575. doi: 10.1161/CIRCRESAHA.114.303915
Lehman, J. J., and Kelly, D. P. (2002). Transcriptional activation of energy metabolic switches in the developing and hypertrophied heart. Clin. Exp. Pharmacol. Physiol. 29, 339–345. doi: 10.1046/j.1440-1681.2002.03655.x
Li, D., Guo, L., Deng, B., Li, M., Yang, T., Yang, F., et al. (2018). Long non-coding RNA HR1 participates in the expression of SREBP-1c through phosphorylation of the PDK1/AKT/FoxO1 pathway. Mol. Med. Rep. 18, 2850–2856. doi: 10.3892/mmr.2018.9278
Li, P., Ruan, X., Yang, L., Kiesewetter, K., Zhao, Y., Luo, H., et al. (2015). A liver-enriched long non-coding RNA, lncLSTR, regulates systemic lipid metabolism in mice. Cell Metab. 21, 455–467. doi: 10.1016/j.cmet.2015.02.004
Li, X., Wang, H., Yao, B., Xu, W., Chen, J., and Zhou, X. (2016). lncRNA H19/miR-675 axis regulates cardiomyocyte apoptosis by targeting VDAC1 in diabetic cardiomyopathy. Sci. Rep. 6:36340. doi: 10.1038/srep36340
Li, Y., Zhang, J., Huo, C., Ding, N., Li, J., Xiao, J., et al. (2017). Dynamic organization of lncRNA and Circular RNA regulators collectively controlled cardiac differentiation in humans. EBioMedicine 24, 137–146. doi: 10.1016/j.ebiom.2017.09.015
Liu, C., Yang, Z., Wu, J., Zhang, L., Lee, S., Shin, D. -J., et al. (2018). Long noncoding RNA H19 interacts with polypyrimidine tract-binding protein 1 to reprogram hepatic lipid homeostasis. Hepatology. 67, 1768–1783. doi: 10.1002/hep.29654
Liu, G., Zheng, X., Xu, Y., Lu, J., Chen, J., and Huang, X. (2015). Long non-coding RNAs expression profile in HepG2 cells reveals the potential role of long non-coding RNAs in the cholesterol metabolism. Chin. Med. J. (Engl). 128, 91–97. doi: 10.4103/0366-6999.147824
Lopaschuk, G. D., Collins-Nakai, R. L., and Itoi, T. (1992). Developmental changes in energy substrate use by the heart. Cardiovasc. Res. 26, 1172–1180. doi: 10.1093/cvr/26.12.1172
Makarewich, C. A., Baskin, K. K., Munir, A. Z., Bezprozvannaya, S., Sharma, G., Khemtong, C., et al. (2018). MOXI Is a mitochondrial micropeptide that enhances fatty Acid β-Oxidation. Cell Rep. 23, 3701–3709. doi: 10.1016/j.celrep.2018.05.058
Malandraki-Miller, S., Lopez, C. A., Al-Siddiqi, H., and Carr, C. A. (2018). Changing metabolism in differentiating cardiac progenitor cells—can stem cells become metabolically flexible cardiomyocytes? Front. Cardiovasc. Med. 5:119. doi: 10.3389/fcvm.2018.00119
Matkovich, S. J., Edwards, J. R., Grossenheider, T. C., De Guzman Strong, C., and Dorn, G. W. (2014). Epigenetic coordination of embryonic heart transcription by dynamically regulated long noncoding RNAs. Proc. Natl. Acad. Sci. U.S.A. 111, 12264–12269. doi: 10.1073/pnas.1410622111
Mayor, F., and Cuezva, J. M. (1985). Hormonal and metabolic changes in the perinatal period. Biol. Neonate 48, 185–196. doi: 10.1159/000242171
Menendez-Montes, I., Escobar, B., Palacios, B., Gómez, M. J., Izquierdo-Garcia, J. L., Flores, L., et al. (2016). Myocardial VHL-HIF signaling controls an embryonic metabolic switch essential for cardiac maturation. Dev. Cell 39, 724–739. doi: 10.1016/j.devcel.2016.11.012
Mongelli, A., Martelli, F., Farsetti, A., and Gaetano, C. (2019). The dark that matters: long noncoding RNAs as master regulators of cellular metabolism in noncommunicable diseases. Front. Physiol. 10:369. doi: 10.3389/fphys.2019.00369
Muret, K., Désert, C., Lagoutte, L., Boutin, M., Gondret, F., Zerjal, T., et al. (2019). Long noncoding RNAs in lipid metabolism: literature review and conservation analysis across species. BMC Genomics 20:882. doi: 10.1186/s12864-019-6093-3
Neidig, L. E., Weinberger, F., Palpant, N. J., Mignone, J., Martinson, A. M., Sorensen, D. W., et al. (2018). Evidence for minimal cardiogenic potential of stem cell antigen 1-positive cells in the adult mouse heart. Circulation 138, 2960–2962. doi: 10.1161/CIRCULATIONAHA.118.035273
Pant, T., Dhanasekaran, A., Fang, J., Bai, X., Bosnjak, Z. J., Liang, M., et al. (2018). Current status and strategies of long noncoding RNA research for diabetic cardiomyopathy. BMC Cardiovasc. Disord. 18:197. doi: 10.1186/s12872-018-0939-5
Porrello, E. R., Mahmoud, A. I., Simpson, E., Hill, J. A., Richardson, J. A., Olson, E. N., et al. (2011). Transient regenerative potential of the neonatal mouse heart. Science 331, 1078–1080. doi: 10.1126/science.1200708
Puente, B. N., Kimura, W., Muralidhar, S. A., Moon, J., Amatruda, J. F., Phelps, K. L., et al. (2014). The oxygen-rich postnatal environment induces cardiomyocyte cell-cycle arrest through DNA damage response. Cell 157, 565–579. doi: 10.1016/j.cell.2014.03.032
Rindler, P. M., Crewe, C. L., Fernandes, J., Kinter, M., and Szweda, L. I. (2013). Redox regulation of insulin sensitivity due to enhanced fatty acid utilization in the mitochondria. Am. J. Physiol. Circ. Physiol. 305, H634–H643. doi: 10.1152/ajpheart.00799.2012
Schonrock, N., Harvey, R. P., and Mattick, J. S. (2012). Long noncoding RNAs in cardiac development and pathophysiology. Circ. Res. 111, 1349–1362. doi: 10.1161/CIRCRESAHA.112.268953
Song, K., Nam, Y. J., Luo, X., Qi, X., Tan, W., Huang, G. N., et al. (2012). Heart repair by reprogramming non-myocytes with cardiac transcription factors. Nature 485, 599–604. doi: 10.1038/nature11139
Soonpaa, M. H., Kim, K. K., Pajak, L., Franklin, M., and Field, L. J. (1996). Cardiomyocyte DNA synthesis and binucleation during murine development. Am. J. Physiol. 271, H2183–H2189. doi: 10.1152/ajpheart.1996.271.5.H2183
Stein, C. S., Jadiya, P., Zhang, X., McLendon, J. M., Abouassaly, G. M., Witmer, N. H., et al. (2018). Mitoregulin: a lncRNA-Encoded microprotein that supports mitochondrial supercomplexes and respiratory efficiency. Cell Rep. 23, 3710.e8–3720.e8. doi: 10.1016/j.celrep.2018.06.002
Thum, T., and Condorelli, G. (2015). Long noncoding RNAs and microRNAs in cardiovascular pathophysiology. Circ. Res. 116, 751–762. doi: 10.1161/CIRCRESAHA.116.303549
Tzahor, E., and Poss, K. D. (2017). Cardiac regeneration strategies: staying young at heart. Science 356, 1035–1039. doi: 10.1126/science.aam5894
Uygur, A., and Lee, R. T. (2016). Mechanisms of cardiac regeneration. Dev. Cell 36, 362–374. doi: 10.1016/j.devcel.2016.01.018
van Gastel, N., Stegen, S., Eelen, G., Schoors, S., Carlier, A., Daniëls, V. W., et al. (2020). Lipid availability determines fate of skeletal progenitor cells via SOX9. Nature 579, 111–117. doi: 10.1038/s41586-020-2050-1
Vausort, M., Wagner, D. R., and Devaux, Y. (2014). Long noncoding RNAs in patients with acute myocardial infarction. Circ. Res. 115, 668–677. doi: 10.1161/CIRCRESAHA.115.303836
Viereck, J., Bührke, A., Foinquinos, A., Chatterjee, S., Kleeberger, J. A., Xiao, K., et al. (2020). Targeting muscle-enriched long non-coding RNA H19 reverses pathological cardiac hypertrophy. Eur. Heart J. 41, 3462–3474. doi: 10.1093/eurheartj/ehaa519
Vivien, C. J., Hudson, J. E., and Porrello, E. R. (2016). Evolution, comparative biology and ontogeny of vertebrate heart regeneration. npj Regen. Med. 1:16012. doi: 10.1038/npjregenmed.2016.12
Wang, K., Long, B., Zhou, L.-Y., Liu, F., Zhou, Q.-Y., Liu, C.-Y., et al. (2014). CARL lncRNA inhibits anoxia-induced mitochondrial fission and apoptosis in cardiomyocytes by impairing miR-539-dependent PHB2 downregulation. Nat. Commun. 5:3596. doi: 10.1038/ncomms4596
Wang, S., Zuo, H., Jin, J., Lv, W., Xu, Z., Fan, Y., et al. (2019). Long noncoding RNA Neat1 modulates myogenesis by recruiting Ezh2. Cell Death Dis. 10:505. doi: 10.1038/s41419-019-1742-7
Wang, Y., Hu, S. B., Wang, M. R., Yao, R. W., Wu, D., Yang, L., et al. (2018). Genome-wide screening of NEAT1 regulators reveals cross-regulation between paraspeckles and mitochondria. Nat. Cell Biol. 20, 1145–1158. doi: 10.1038/s41556-018-0204-2
Yan, C., Chen, J., and Chen, N. (2016). Long noncoding RNA MALAT1 promotes hepatic steatosis and insulin resistance by increasing nuclear SREBP-1c protein stability. Sci. Rep. 6:22640. doi: 10.1038/srep22640
Yang, F., Zhang, H., Mei, Y., and Wu, M. (2014). Reciprocal regulation of HIF-1α and lincRNA-p21 modulates the Warburg effect. Mol. Cell 53, 88–100. doi: 10.1016/j.molcel.2013.11.004
Yucel, N., Wang, Y. X., Mai, T., Porpiglia, E., Lund, P. J., Markov, G., et al. (2019). Glucose metabolism drives histone acetylation landscape transitions that dictate muscle stem cell function. Cell Rep. 27, 3939.e6–3955.e6. doi: 10.1016/j.celrep.2019.05.092
Zhang, L., Zhou, Y., Huang, T., Cheng, A. S. L., Yu, J., Kang, W., et al. (2017). The interplay of LncRNA-H19 and its binding partners in physiological process and gastric carcinogenesis. Int. J. Mol. Sci. 18:450. doi: 10.3390/ijms18020450
Zhang, M., Chi, X., Qu, N., and Wang, C. (2018). Long noncoding RNA lncARSR promotes hepatic lipogenesis via Akt/SREBP-1c pathway and contributes to the pathogenesis of nonalcoholic steatohepatitis. Biochem. Biophys. Res. Commun. 499, 66–70. doi: 10.1016/j.bbrc.2018.03.127
Zhao, X.-Y. (2015). Long noncoding RNAs: a new regulatory code in metabolic control. Trends Biochem. Sci. 40, 586–596.
Zhuo, C., Jiang, R., Lin, X., and Shao, M. (2017). LncRNA H19 inhibits autophagy by epigenetically silencing of DIRAS3 in diabetic cardiomyopathy. Oncotarget 8, 1429–1437. doi: 10.18632/oncotarget.13637
Keywords: lncRNAs, metabolism, mitochondria, regeneration, heart
Citation: Correia M, Bernardes de Jesus B and Nóbrega-Pereira S (2021) Novel Insights Linking lncRNAs and Metabolism With Implications for Cardiac Regeneration. Front. Physiol. 12:586927. doi: 10.3389/fphys.2021.586927
Received: 24 July 2020; Accepted: 15 February 2021;
Published: 10 March 2021.
Edited by:
Bernhard Johannes Haubner, University Hospital Zürich, SwitzerlandReviewed by:
Sang-Bing Ong, The Chinese University of Hong Kong, ChinaCheng-Kai Huang, Hannover Medical School, Germany
Copyright © 2021 Correia, Bernardes de Jesus and Nóbrega-Pereira. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sandrina Nóbrega-Pereira, c2FuZHJpbmEucGVyZWlyYUB1YS5wdA==; Bruno Bernardes de Jesus, YnJ1bm9iLmplc3VzQHVhLnB0