 Alyne Goulart Escobar1
Alyne Goulart Escobar1 Danize Aparecida Rizzetti1†
Danize Aparecida Rizzetti1† Janaina Trindade Piagette1
Janaina Trindade Piagette1 Franck Maciel Peçanha1
Franck Maciel Peçanha1 Dalton Valentim Vassallo2
Dalton Valentim Vassallo2 Marta Miguel3
Marta Miguel3 Giulia Alessandra Wiggers1*
Giulia Alessandra Wiggers1*- 1Graduate Program in Biochemistry and Multicentric Graduate Program in Physiological Sciences, Universidade Federal do Pampa, Uruguaiana, Brazil
- 2Department of Physiological Sciences, Universidade Federal do Espírito Santo and School of Medicine of Santa Casa de Misericórdia (EMESCAM), Vitória, Brazil
- 3Bioactivity and Food Analysis Department, Instituto de Investigación en Ciencias de la Alimentación, Campus Universitario de Cantoblanco, Madrid, Spain
Aim: We investigated the antioxidant protective power of egg white hydrolysate (EWH) against the vascular damage induced by mercury chloride (HgCl2) exposure in resistance arteries.
Methods: Male Wistar rats received for 60 days: (I) intramuscular injections (i.m.) of saline and tap water by gavage – Untreated group; (II) 4.6 μg/kg of HgCl2 i.m. for the first dose and subsequent doses of 0.07 μg/kg/day and tap water by gavage – HgCl2 group; (III) saline i.m. and 1 g/kg/day of EWH by gavage – EWH group, or (IV) the combination of the HgCl2 i.m. and EWH by gavage – EWH + HgCl2 group. Blood pressure (BP) was indirectly measured and dose-response curves to acetylcholine (ACh), sodium nitroprusside (SNP), and noradrenaline (NE) were assessed in mesenteric resistance arteries (MRA), as in situ production of superoxide anion, nitric oxide (NO) release, vascular reactive oxygen species (ROS), lipid peroxidation, and antioxidant status.
Results: Egg white hydrolysate prevented the elevation in BP and the vascular dysfunction after HgCl2 exposure; restored the NO-mediated endothelial modulation and inhibited the oxidative stress and inflammatory pathways induced by HgCl2.
Conclusion: Egg white hydrolysate seems to be a useful functional food to prevent HgCl2-induced vascular toxic effects in MRA.
Introduction
The imbalance between the excessive formation of reactive oxygen species (ROS) and limited antioxidant defenses is one of the most harmful mechanisms that induce deleterious cardiovascular effects (Liguori et al., 2018). Oxidative damage has been implicated in vascular injury and endothelial dysfunction (Halliwell, 2007), a proliferation of vascular smooth muscle cells (VSMC), increase in vascular tone, migration of inflammatory mediators, vascular remodeling (Szasz et al., 2007), and hypertension (Touyz and Briones, 2011; Montezano and Touyz, 2014; Montezano et al., 2015; Incalza et al., 2018).
Several studies point out the beneficial effects of consuming different natural antioxidant compounds and bioactive food ingredients, such as protein-derived peptides against oxidative imbalance (Serafini and Peluso, 2016; Peluso and Serafini, 2017; Chikara et al., 2018). Bioactive peptides from egg white hydrolysate (EWH) have shown important antioxidant properties, preventing dysfunction in hypertensive and obese experimental models (Manso et al., 2008; Garces-Rimon et al., 2019). We propose to use EWH obtained after hydrolysis of hen egg white with pepsin for 8 h, which has previously demonstrated antioxidant, anti-inflammatory, and angiotensin-converting enzyme (ACE) inhibitory properties (Miguel et al., 2004; Moreno-Fernandez et al., 2018a,b).
Metals, such as mercury (Hg), are dangerous pollutants in the ecosystem, found in different physical and chemical forms. Their toxic effects are dose and time-dependent and can be characterized as a risk factor for the development of cardiovascular diseases by the promotion of oxidative stress (Genchi et al., 2017). Hg participates in the Fenton reaction, increasing ROS production and also leading to the depletion of important antioxidant enzymes due to their affinity for the sulfhydryl radicals (Valko et al., 2006; Su et al., 2008).
Studies have been shown that 30-day exposure to a low concentration of mercury chloride (HgCl2) induces oxidative stress and activation of cyclooxygenase (COX) and angiotensin II pathways, promoting vascular changes, such as endothelial dysfunction and increased reactivity (Wiggers et al., 2008; Peçanha et al., 2010). It has been found that EWH improves the Hg-induced damage in parameters related to memory deficits (Rizzetti et al., 2016a), peripheral nervous disorders (Rizzetti et al., 2016b), male reproductive dysfunction (Rizzetti et al., 2017b), and conductance arteries injury (Rizzetti et al., 2017a). In aorta of long-term Hg-exposed rats, EWH prevented the high vascular reactivity and endothelial dysfunction promoted by the metal. However, small arteries, such as resistance mesenteric arteries (MRA), represent the primary vessels that are involved in the regulation of arterial blood pressure (BP) as well as blood flow within the organ.
Thus, we investigate whether the antioxidant properties of EWH have a protective power against vascular damage caused by exposure to HgCl2 in MRA and the underlying pathways involved.
Materials and Methods
Animals and Experimental Groups
The experiments were conducted in compliance with the Principles of Laboratory Animal Care (National Institutes of Health publication 80–23, revised 1996) and in agreement with the guidelines by the Brazilian Societies of Experimental Biology and approved by the Ethics Committee on Animal Use Experimentation of the Federal University of Pampa, Uruguaiana, Rio Grande do Sul, Brazil (protocol 05/2014). The rats (Male Wistar rats; age 3 mo; 250–300 g) were kept in controlled light, temperature, and humidity conditions (12/12 h light/dark cycle; 23 ± 2°C; 50 ± 10%, respectively) with free access to food and water and randomly submitted to one of the following protocols for 60 days: (I) intramuscular injections (i.m.) of saline solution 0.9% and tap water by gavage – Untreated group; (II) 4.6 μg/kg of HgCl2 i.m. for the first dose and subsequent doses of 0.07 μg/kg/day, to cover daily loss, using the model described previously (Rizzetti et al., 2017c) and tap water by gavage – HgCl2 group; (III) saline solution 0.9% i.m. and EWH from pepsin for 8 h diluted in tap water (1 g/kg/day), by gavage, according to the model prior reported (Rizzetti et al., 2016a) – EWH group, or (IV) the combination of the HgCl2 i.m. and EWH by gavage – EWH + HgCl2 group. Bodyweight, food, and water intakes were measured once a week.
This experimental model of chronic controlled exposure to a low concentration of Hg has a total metal concentration in the blood at the end of the treatment of 3.04 ng/ml (Rizzetti et al., 2017c), which is within the safety limit established by US Environmental Protection Agency’s (5.8 ng/ml) and similar to the blood Hg concentration of people exposed to the metal by the workplace or through diet (Rice, 2004). Moreover, to make the model more similar to the human exposure condition, we carry out a simultaneous treatment of Hg and EWH, considering that humans are rarely entirely free of any level of exposure to this metal. So our concern was to investigate the benefits of EWH during continuous and concurrent exposure to Hg.
Systolic Blood Pressure
Systolic Blood Pressure (SBP) was recorded once a week by tail plethysmography (ADInstruments Pty Ltd., Bella Vista, NSW, Australia) according to Rizzetti et al. (2017c). The animals were preheating the animals at 37°C for 10 min to make the pulsations of the caudal artery detectable, followed by 10 sequential cycles of tail inflation-deflation. The SBP was considered the average of all measures.
Vascular Experiments in the Mesenteric Arteries
The MRA from anesthetized (ketamine-xylazine, 87 and 13 mg/kg i.p.) rats (diameter of third-order branch, in μm: Untreated: 262 ± 5.1; HgCl2: 301 ± 4.8; EWH: 254 ± 7.0 and EWH + HgCl2: 318 ± 5.4; p > 0.05) were removed and rings of 2 mm in length were mounted in an isometric small-vessel myograph (Multi Wire Myograph System, DMT620, ADInstruments, Australia) according to Mulvany and Halpern (1977) and connected to an acquisition system (PowerLab 8/35, ADInstruments, Australia). Briefly, the rings were submerged in warmed (37°C) Henseleit solution (KHS, in mM at 37°C: 115 NaCl, 25 NaHCO3, 4.7 KCl, 1.2 MgSO4 7H2O, 2.5 CaCl2, 1.2 KH2PO4, 11.1 glucose, and 0.01 Na2EDTA) continuously bubbled with carbogen (5% CO2 in O2). Initially, segments were contractility contracted with KCl (120 mM) to test the viability of the vessels. The endothelium of some vessels was removed by rubbing the intimal surface. The absence of acetylcholine (ACh – 0.01 nM – 30 mM)-induced relaxation in rings pre-contracted by noradrenaline (NE 10 μM) was taken as an indicator of successful endothelium denudation. Endothelium-independent relaxation were tested by sodium nitroprusside (SNP – 0.1 nM − 3.5 mM) under the same pre-contracted conditions (NE – 10μm). We also evaluated the vascular response to increasing concentrations of NE (10 nM – 30 μM). To evaluate the pathways involved in the contractile responses, the following drugs were added 30 min before the generation of the NE concentration-response curves: nitric oxide synthase (NOS) inhibitor N-nitro-L-arginine methyl ester (L-NAME, 100 mM), a NAD(P)H oxidase inhibitor (VAS2870, 10 μM), an essential cofactor for NO synthesis, tetrahydrobiopterin (BH4, 100 μM), a superoxide dismutase mimetic (TEMPOL, 10 μM), and a non-selective COX inhibitor (Indomethacin, 10 μM).
Vascular in situ Nitric Oxide and Reactive Oxygen Species Detection
Measurements of NO and ROS levels in MRA segments were performed as previously published by Martin et al. (2012) and Avendaño et al. (2014), respectively. Briefly, the MRA rings were incubated for 45 min with 4,5-diaminofluorescein diacetate (DAF-2, 10 μM) to assess the release of NO. The first collection of the medium measured the basal release of NO, and after the segments were incubated with NE 0.1 nM and relaxed with ACh 10 μM, the medium was collected again, and the induced release of NO was measured by fluorescence method (excitation at 492 nm and emission at 515 nm – SpectraMax M5 Molecular Devices, Sunnyvale, CA, United States). The results are expressed as arbitrary units/g tissue as a percentage of fluorescence obtained for Untreated rats. The release of NO is the result of fluorescence evoked by ACh subtracted from the basal release of NO.
For ROS measured, MRA rings were dissected, frozen in OCT solution and then cut in 14-μm-thick sections, placed on a glass slide, and equilibrated in a Krebs-HEPES buffer (in mM: 130 NaCl, 5.6 KCl, 2 CaCl2, 0.24 MgCl2, and 8.3 HEPES), and 11 glucose, pH 7.4. DHE (2 μM, 30 min, 37°C) incubated for 30 min in a dark-chamber and then viewed by a fluorescence microscope (Eclipse 50i55i Epi-fluorescence Nikon, Tokyo, Japan, magnification: × 40). The total ring area was analyzed using Image J version V1.56 (National Institutes of Health, Bethesda, Maryland, United States) software.
Biochemical Assays
The supernatant fraction derived from homogenization (50 mM Tris-HCl, pH 7.4, 1/10, w/v) and centrifugation (2,400 g; 10 min; 4°C) of the MRA was frozen at −80°C. The levels of reactive species (RS) were measured according to Loetchutinat et al. (2005) by a spectrofluorometric method. Briefly, after diluted (1:5 in Tris-HCl 50 mM; pH 7.4), samples received 1 mM of 2', 7'-dichlorofluorescein diacetate (DCHF-DA). The 2', 7'-dichlorofluorescein (DCF) fluorescence intensity emission represents the amount of ROS formed (520 nm emission with 480 nm excitation for 60 min at 15 min intervals – SpectraMax M5 Molecular Devices, CA, United States). The ROS levels were expressed as fluorescence units and as a percentage of those obtained for Untreated rats.
The malondialdehyde (MDA) levels that represent the lipid peroxidation were measured following Ohkawa et al. (1979) protocol with modifications by the colorimetric method. Briefly, thiobarbituric acid (TBA) 0.8%, acetic acid buffer (pH 3.2) plus sodiumduodecilsulphate (SDS) 8% were added to the samples and the color reaction was measured against blanks (532 nm – SpectraMax M5 Molecular Devices, Sunnyvale, CA, United States). The data were expressed as nmol of MDA per gram of tissue.
The total antioxidant capacity was measured by Ferric Reducing/Antioxidant Power (FRAP) assay, according to Benzie and Strain, 1996. Briefly, homogenate of MRA was added to FRAP reagent [acetate buffer pH 3.6, 10 mM; 2,4,6-Tri(2-piridil)-s-triazina – TPTZ, 40 mM HCl; FeCl3-10:1:1; 37°C] and incubated at 37°C for 10 min. The absorbance was read at 593 nm (SpectraMax M5 Molecular Devices, CA, United States). The standard dose-response curve of Trolox (50–1,000 μM) was performed, and results are presented with particular reference to Trolox equivalents.
Non-protein thiols (NPSH) were measured, according to Ellman (1959). MRA tissue was added to a buffer of potassium phosphate (1 M, pH 7.4), and 5,5'-dithio-bis(2-nitrobenzoic acid) (DTNB, 10 mM) and the color reaction was spectrophotometrically read (412 nm – SpectraMax M5 Molecular Devices, CA, United States). The results were expressed as nmol of NPSH per gram of tissue.
Statistical Analyses
Values are expressed as means ± SEM. The results were analyzed using a two-way ANOVA. When two-way ANOVA showed a statistical significance, the Fisher post hoc test was applied (Graph Pad Prism 6.0 Software, San Diego, CA). To compare the effects between groups (endothelium removal and drugs) on the response to NE, some results are expressed as the differences of areas under the concentration-response curves (dAUC) in the control and experimental situations. The differences were expressed as the % of the AUC of the corresponding control situation. The results were considered statistically significant for values of p < 0.05.
Results
The water and food intake (data not shown) as well body weight gain were not modified during the treatment (body weight gain, in g: Untreated: 58.5 ± 4.1; HgCl2: 60.3 ± 4.9; EWH: 59.7 ± 3.8; EWH + HgCl2: 61.1 ± 5.3; n = 8; and p > 0.05). Increased SBP values were observed in HgCl2-treated rats and EWH treatment was able to decrease SBP values in HgCl2-treated rats (SBP values, in mmHg: Untreated: 120.1 ± 1.9; HgCl2: 135.2 ± 2.8*; EWH: 124.5 ± 1.5; EWH + HgCl2: 122.0 ± 2.2#; n = 8; and p < 0.05 – * vs. Untreated and # vs. HgCl2). In MRA reactivity, the maximum response to KCl was similar between groups (in mN/mm, Untreated: 14.1 ± 0.3; HgCl2: 14.2 ± 0.5; EWH: 14.3 ± 0.1; EWH + HgCl2: 14.1 ± 0.2; n = 8; and p > 0.05), demonstrating that neither Hg nor EWH alter vascular integrity of MRA.
Egg white hydrolysate intake prevented the increased contractile responses to NE and the reduced endothelium-dependent vasodilator response to ACh induced by HgCl2 exposure. The endothelium-independent vasodilator response to NPS in MRA was not affected in all treatments (Figures 1A–C).
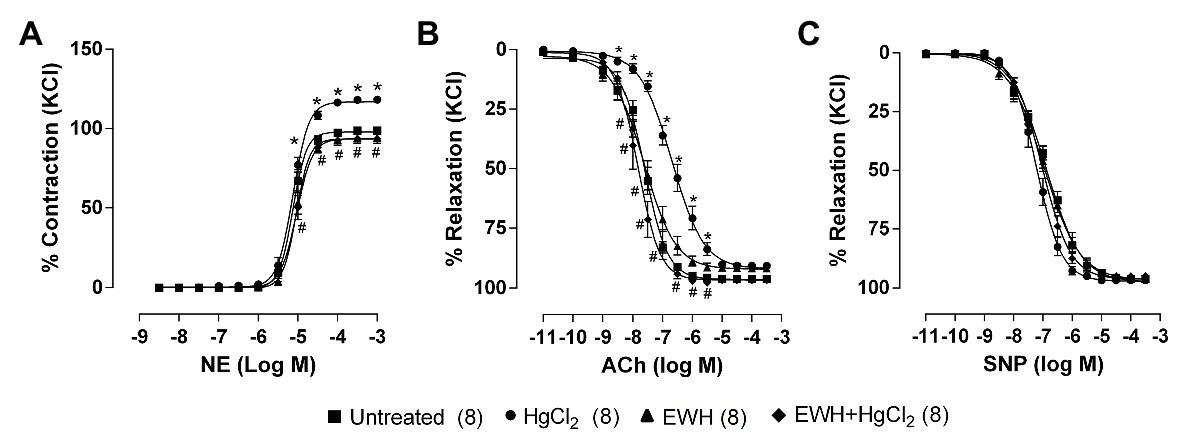
Figure 1. Effect of egg white hydrolysate (EWH) co-treatment in mesenteric resistance arteries (MRA) reactivity of rats exposure to mercury chloride (HgCl2) for 60 days. Concentration–response curves to (A) noradrenaline (NE), (B) acetylcholine (ACh), and (C) sodium nitroprusside (SNP) in MRA segments. The results (mean ± SEM) are expressed as a percentage of the response to 120 mmol/l KCl. Two-Way ANOVA followed by Fisher test; n of each group in parenthesis; *p < 0.05 vs. Untreated and # vs. HgCl2.
Denuded endothelium or NOS inhibitor incubation (L-NAME) increased the contractile response to NE in all groups except in HgCl2 group as evidenced by the dAUC values (Figures 2A–E,a–e). These findings show the absence of endothelial participation in the vasoconstrictor response to NE in this group. MRA segments from HgCl2 exposure animals that received EWH showed similar effects of endothelium removal or L-NAME compared to the Untreated group (Figures 2A–E,a–e), suggesting that EWH prevented this reduced endothelial modulation by NO. In agreement, ACh-induced NO release was lower in MRA from Hg-treated rats. On the other hand, rats receiving EWH alone and those receiving the combination of HgCl2 and EWH had a greater percentage of NO release when compared to the Untreated group (Figure 2a’). This finding suggests that EWH was able to induce NO production by NOS.
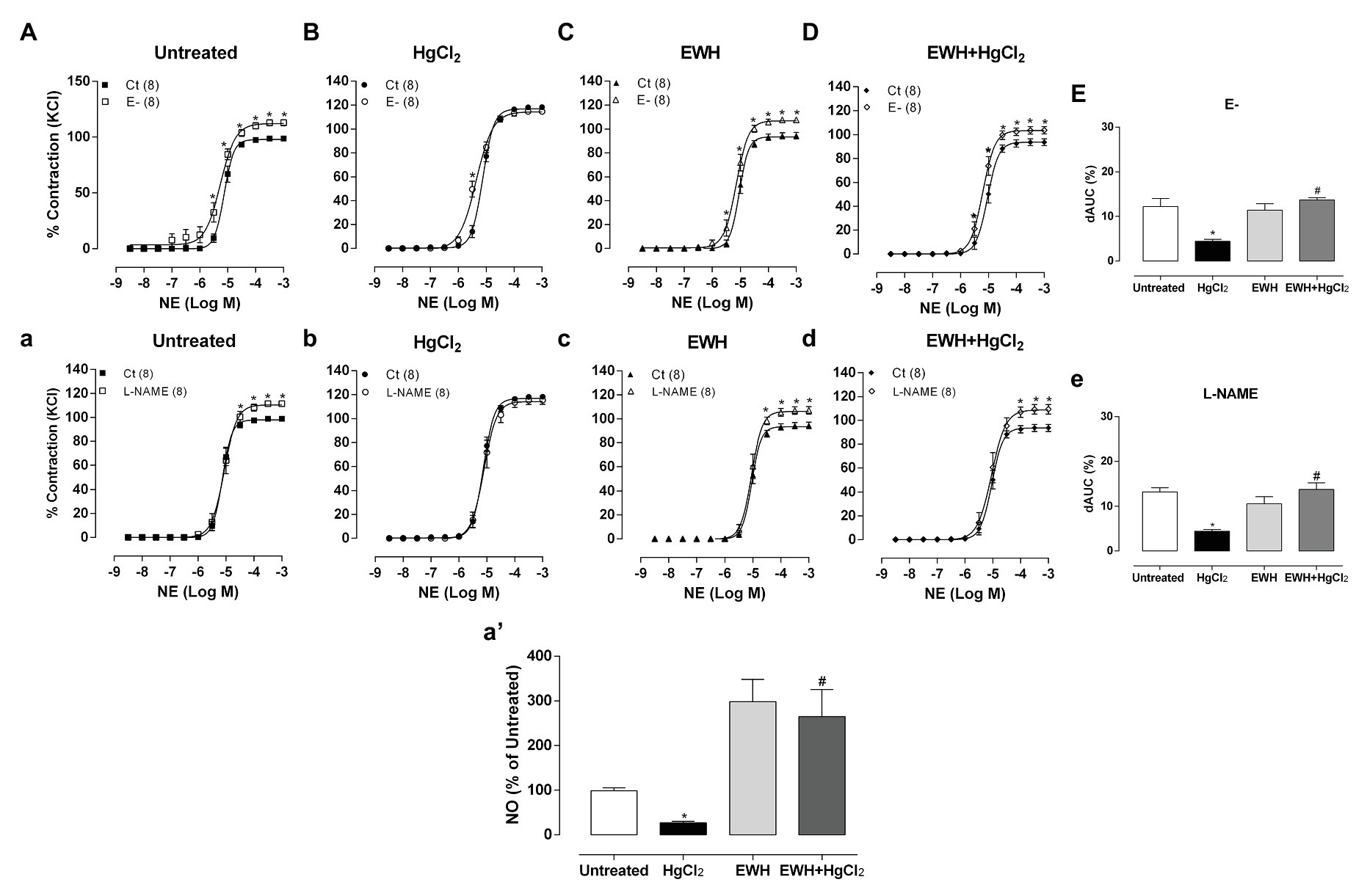
Figure 2. Effects of EWH co-treatment in MRA of rats exposure to HgCl2 for 60 days on the endothelium and nitric oxide (NO) mediated vascular response. Concentration-response curve to NE in the absence of endothelium (E−; A−D) and N-nitro-L-arginine methyl ester (L-NAME; 100 μM; a–d) in mesenteric segments from rats Untreated (A), treated with HgCl2 (B), with EWH (C), and with EWH + HgCl2 (D). The results (mean ± SEM) are expressed as a percentage of the response to 120 mmol/l KCl. Two-Way ANOVA followed by Fisher test; *p < 0.05 vs. Untreated and # vs. HgCl2. Differences in the area under the concentration-response curves (dAUC) in mesenteric segments are represented in (E,e) of the four experimental groups; one-way ANOVA, *p < 0.05 vs. Untreated and # vs. HgCl2. In (a’) represent the NO release in all experimental groups.
The cofactor for NO synthesis BH4 incubation had a smaller contractile response to NE in HgCl2-treated rats, showing lower BH4 bioavailability and, possibly, an uncoupled state of endothelial nitric oxide synthase (eNOS) in this vessel. EWH intake maintained BH4 bioavailability in MRA from HgCl2-treated rats, indicating the improvement of eNOS and NO synthesis (Figures 3A–E).
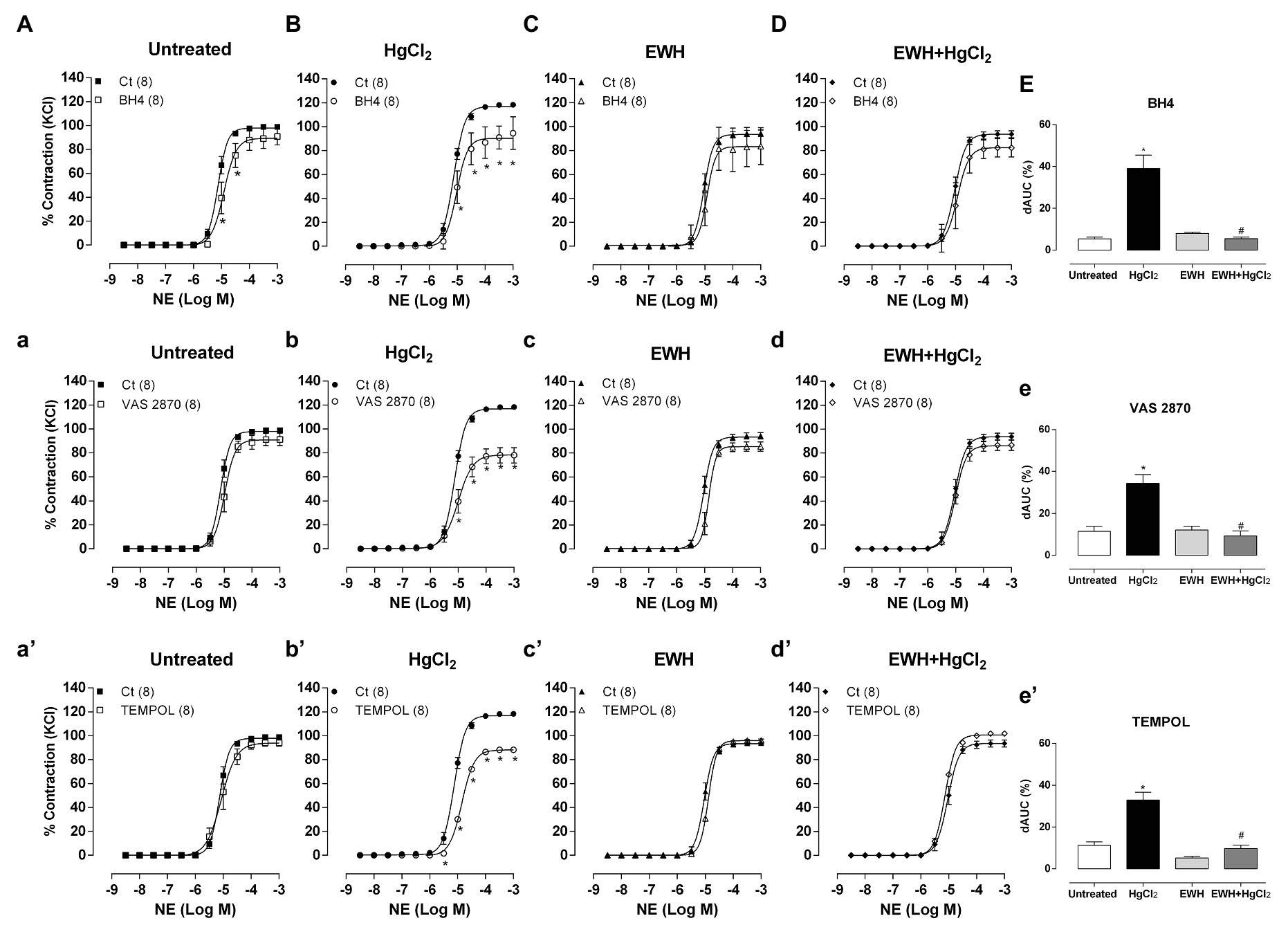
Figure 3. Effects of EWH on NO cofactor modulation, participation of reactive oxygen species (ROS) from NAD(P)H oxidase and participation of superoxide anion in vasoconstrictor responses to NE in MRA from rats exposed to low concentrations of HgCl2 for 60 days. Concentration-response curve to NE in the absence (Ct), presence of the endothelial nitric oxide synthase (eNOS) cofactor (BH4; A–D), the presence of the NAD(P)H synthase inhibitor (VAS2870; a–d) and the presence of the superoxide anion scavenger mimetic (TEMPOL; a’–d’) in mesenteric segments from rats Untreated (A), treated with HgCl2 (B), with EWH (C), and with EWH + HgCl2 (D). The results (mean ± SEM) are expressed as a percentage of the response to 120 mmol/l KCl. Differences in the area under the concentration-response curves (dAUC) in mesenteric segments in the presence and the absence of BH4, VAS2870 and TEMPOL of the four experimental groups (E,e,e’); n of each group in parenthesis, one-way ANOVA, *p < 0.05 vs. Untreated and # vs. HgCl2.
The NAD(P)H oxidase inhibitor VAS2870 reduced the contractile response to NE in MRA only in HgCl2 group (Figures 3a–e). EWH prevented the increased ROS participation from NAD(P)H oxidase on contractile response to NE. The superoxide dismutase mimetic TEMPOL reduced the contractile response to NE in MRA only from HgCl2-exposed rats; thus, the NE responses remained unchanged in the other groups (Figures 3a’–e’). We also observed significantly higher superoxide anion production in arteries of rats exposed to HgCl2; EWH prevented this effect (Figure 4). Chronic Hg treatment for 60 days increased ROS levels and lipid peroxidation in MRA from exposed rats (Figures 5A,B) and reduced the total antioxidant capacity and the NPHS levels in the Hg-exposed rats (Figures 5C,D). EWH was able to prevent the oxidative stress in MRA of HgCl2-treated rats, balancing the pro-oxidant and antioxidant status (Figures 5A–D).
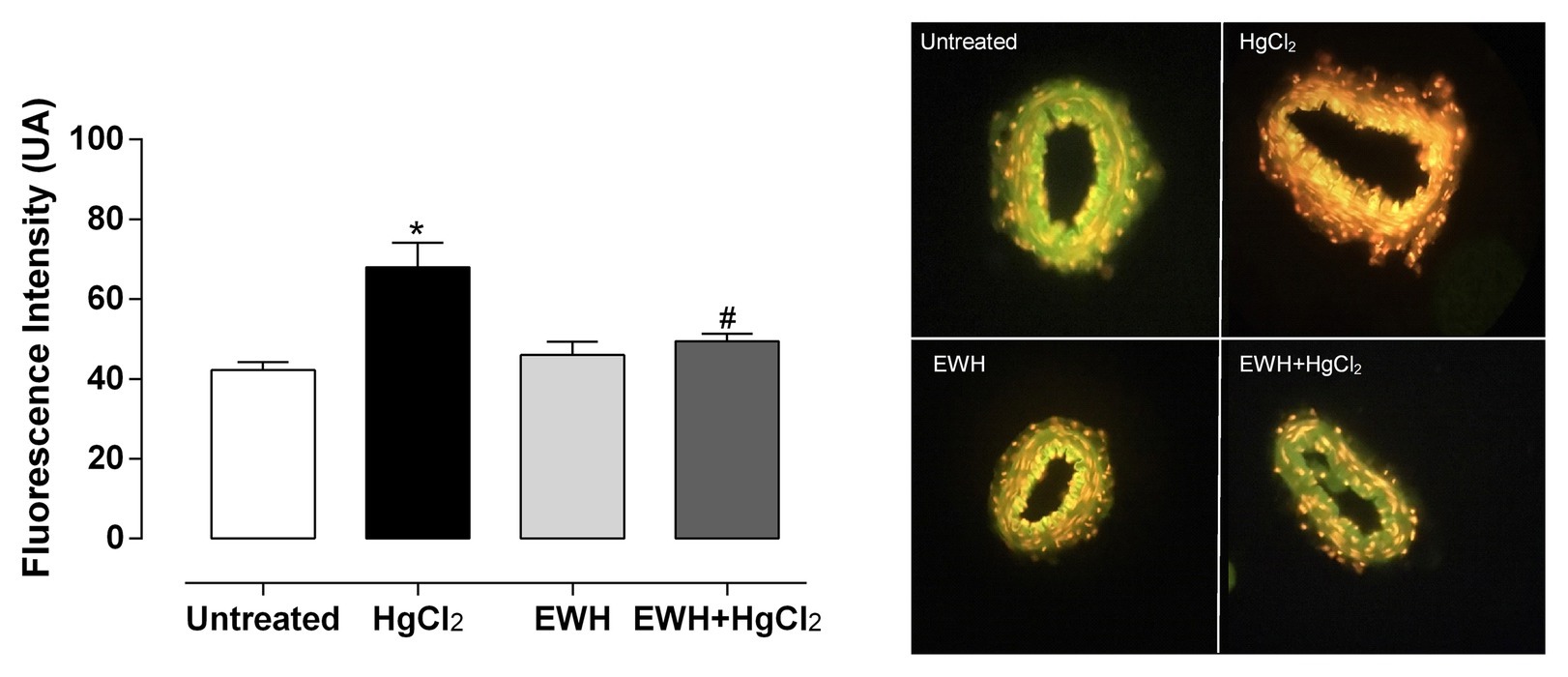
Figure 4. Effects of EWH on local anion superoxide production in mesenteric of rats exposed to low concentrations of HgCl2 for 60 days. Superoxide Anion production in mesenteric from rats Untreated, treated HgCl2, with EWH and with EWH + HgCl2 levels in mesenteric of all groups; n of each group in parenthesis, one-way ANOVA, *p < 0.05 vs. Untreated, # vs. HgCl2.
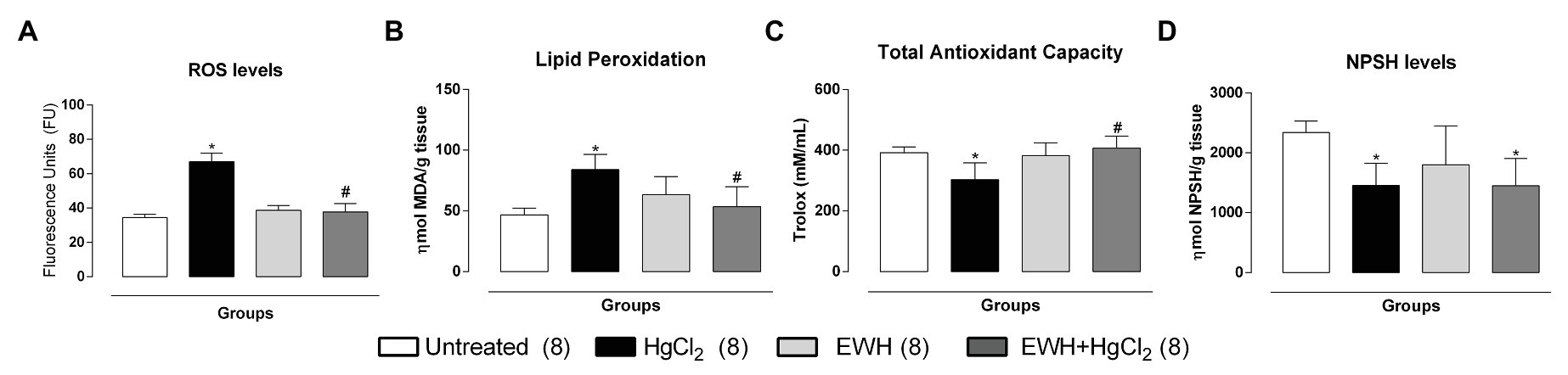
Figure 5. Effects of EWH in the ROS (A), MDA (B), the total antioxidant capacity (C), and NPSH (D) in mesenteric of rats exposed to low concentrations of HgCl2 for 60 days. Data are expressed as mean ± SEM. Number of animals is indicated in parentheses respectively, one-way ANOVA, *p < 0.05 vs. Untreated; # vs. HgCl2.
Indomethacin reduced the response to NE in MRA segments from only in HgCl2-treated rats (Figures 6A–E) indicating that the enhanced COX pathway participation in these responses was prevented by EWH.
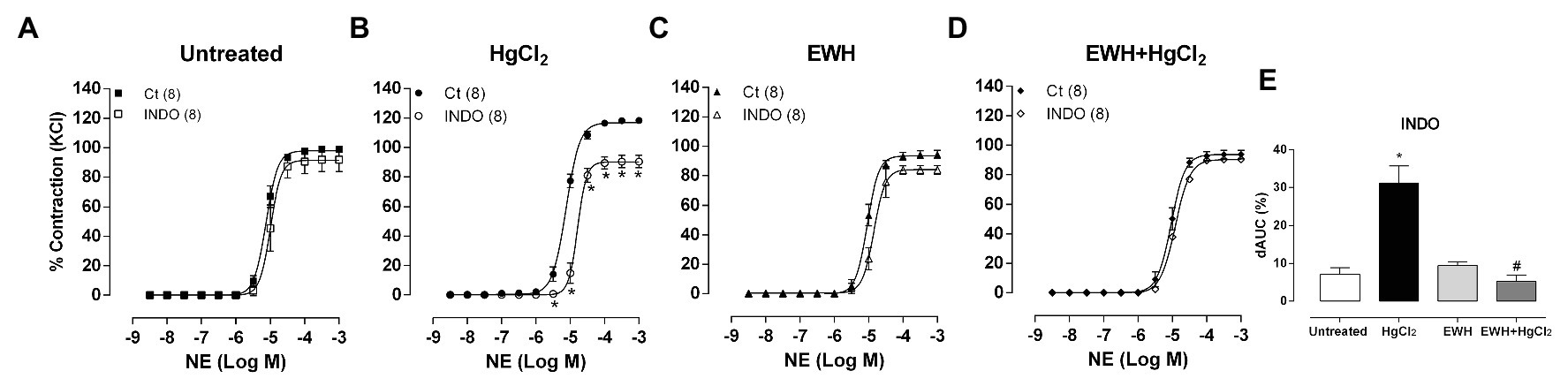
Figure 6. Effects of EWH on contribution of cyclooxygenase (COX) pathway on vasoconstrictor responses to NE in mesenteric of rats exposed to low concentrations of HgCl2 for 60 days. Concentration-response curve to NE in the absence (Ct), the presence of the non-selective COX inhibitor (Indomethacin; A–D) in mesenteric segments from rats Untreated (A), treated with HgCl2 (B), with EWH (C), and with EWH + HgCl2 (D). The results (mean ± SEM) are expressed as a percentage of the response to 120 mmol/l KCl. Two-Way ANOVA followed by Fisher test; n of each group in parenthesis; *p < 0.05 vs. Untreated and # vs. HgCl2. Differences in the area under the concentration-response curves (dAUC) in mesenteric segments are represented in (E) graph (one-way ANOVA).
Discussion
Intake of EWH as a functional food was able to reverse the increase in SBP induced by chronic exposure to HgCl2 at low concentrations, which is related to the reduction of contractile responses and the vascular dysfunction induced by the metal in MRA. These effects were associated, at least in part, with the capacity of EWH to produce NO from eNOS and with its antioxidant and anti-inflammatory properties. The bioactive peptides of EWH protect against high concentrations of ROS from NAD(P)H oxidase and, possibly, activation of inflammatory COX in MRA in HgCl2-treated rats, thus normalizing the NO modulation in the vasculature.
The Hg is a well-known environmental risk factor for cardiovascular diseases (Virtanen et al., 2007). Acute Hg exposure promotes the reduction of myocardial contractility and inhibition of myosin ATPase activity (Vassallo et al., 1999). Moreover, subchronic exposure to Hg, at doses similar to human exposure, increases vascular reactivity of resistance and conductance vessels in rats (Wiggers et al., 2008; Peçanha et al., 2010). A prolonged exposure at low doses of HgCl2 for 60 days increased SBP and vascular dysfunction in conductance arteries; these effects were related to stimulation of ACE activity, NAD(P)H oxidase-mediated oxidative stress and activation of COX-2 inflammatory pathway in these vessels (Rizzetti et al., 2017a,c). Our results demonstrate that resistance arteries are also affected by prolonged exposure to Hg, which could better explain the high SBP values observed, since the main vascular site responsible for vascular resistance and maintenance of BP is the resistance arteries (Oparil et al., 2018). Our purpose was to investigate if the mechanisms involved in this increment of SBP could be blocked by EWH treatment.
The Hg exerts its toxic effects on the cardiovascular system through oxidative stress caused by the production of superoxide anion from NAD(P)H oxidase and, possibly, inflammatory mediators derived from COX. Although we have not verified the specific participation of the COX-2 pathway in the current study, previous work of our group, at the same model, evidenced the participation of this pathway in increasing vascular reactivity in conductance arteries using a selective COX-2 inhibitor (NS 398), and the relationship between ROS and COX-2-derived prostanoids. Moreover, the vascular functional findings showed a reduction in the COX-2 participation in the cardiovascular system of Hg-treated rats after EWH intake, proving its anti-inflammatory property observed in vitro (Rizzetti et al., 2017a). Thus, in this study, we suggest that inflammatory mediators derived from COX-2 are also involved in increasing vascular reactivity in resistance arteries. However, future investigations using selective inhibitors of this pathway may explore these findings.
Besides, we verified for the first time the possible involvement of eNOS uncoupling in HgCl2-induced negative actions on vascular tissue; this could be due to high vascular oxidative stress caused by inflammatory stimuli from the COX pathway. Our findings demonstrated the protective effect of EWH on resistance arteries, which blocked the mechanisms involved in the increment of blood pressure by Hg.
Egg white hydrolysate derived from hydrolysis with pepsin for 8 h has several biological activities, such as antioxidant, free radical scavenger, ACE inhibitor, vascular-relaxing, and anti-inflammatory (Miguel et al., 2004). Fourteen of its constituent peptides have been identified (FRADHPFL, RADHPFL, YAEERYPIL, YRGGLEPINF, ESIINF, RDILNQ, IVF, YQIGL, SALAM, and FSL; Davalos et al., 2004; Miguel et al., 2004; Garcés-Rimón et al., 2016) and their biological actions have been previously reported in vitro or in vivo studies (Miguel et al., 2006, 2007; Garces-Rimon et al., 2019). However, these peptides and amino acids act individually, cooperatively, and synergistically; during their passage through the gastrointestinal tract, some modifications due to new hydrolysis should be considered (Miner-Williams et al., 2014). Thus, we do not know if the effects observed in this study are due to a specific peptide or the sum of all its constituent peptides’ effects. Although most studies conducted in recent years have focused on the isolation of peptide sequences released during hydrolysis, it has recently been proven that the administration of complete hydrolysates could be more relevant than the administration of a single isolated peptide since a more significant biological effect could be achieved (Liu et al., 2017). Moreover, we consider that hydrolysates could be more attractive products for developing functional foods from a technological and organoleptic point of view.
In the present study, we demonstrated improvements in resistance arteries function and, consequently, in SBP levels of Hg-treated rats after the EWH treatment. The mechanisms involved on cardiovascular beneficial effects observed after consumption of EWH are probably due to its vascular-relaxing, antioxidant and anti-inflammatory effects. The antihypertensive capacity of EWH we show here was previously reported in experimental models of spontaneously hypertensive rats (SHR) (Miguel et al., 2006) and was attributed, at least in part, to the vasodilator peptides whose N-terminal position exhibits amino acids Arg or Tyr (Miguel et al., 2004, 2005).
It has been previously reported that alterations in eNOS gene increase the susceptibility to cardiovascular diseases in individuals after Hg exposure by modulating NO levels (de Marco et al., 2012). Exposure to Hg at low concentrations has deleterious vascular effects on aorta, coronary and basilar arteries related to greater vascular reactivity caused by the reduction of NO bioavailability in these vessels (Wiggers et al., 2008; Peçanha et al., 2010; Botelho et al., 2019). Moreover, a study in mesenteric arteries from rats exposed for 30 days to HgCl2 showed eNOS protein expression upregulation possibly due to a compensatory mechanism against metal-induced endothelial dysfunction (Wiggers et al., 2008). Our results indicate that lower NO bioavailability and endothelial dysfunction persists in MRA of long-term Hg exposure. Moreover, we showed that lower NO bioavailability occurs, at least in part, due to less NO release in resistance arteries.
Considering previous and present findings, we could hypothesize that the increased eNOS protein expression in MRA can accompany the reduction of this isoform activity or that its expression and activity may be increased. However, this enzyme could be decoupled and producing ROS, which would support the increase in oxidative stress associated with reducing the NO release observed in this study. In this condition, eNOS shifts from NO production to overproducing superoxide anion, increasing oxidative stress in the vasculature (Antoniades et al., 2006). Here we observed lower BH4 bioavailability in MRA of Hg-treated rats, which represents an important cofactor for the production of NO by this enzyme. This finding suggests that Hg induces eNOS uncoupling by inhibiting its cofactor. In any case, the alteration of eNOS protein expression induced by exposure to Hg increases, oxidative stress, and vascular damage are being protected by the ingestion of EWH.
In this sense, NO bioavailability could also be related to oxidative stress imbalance due to overproduction of vascular ROS, generation of peroxynitrite, and reduction of antioxidant reserves (Faria et al., 2018). Interestingly, oxidative stress has also been reported as an important mechanism responsible for lower NO bioavailability induced by Hg (Wiggers et al., 2008; Rizzetti et al., 2017c); it is also involved in Hg-induced vascular damage in our study. Besides, oxidative stress can directly modify eNOS protein or its cofactor BH4, leading to enzymatic dysfunction in vascular tissue (Dumitrescu et al., 2007).
It is important to emphasize that EWH is composed of several peptides that have vasodilator effects (Miguel et al., 2007). These peptides were related to EWH capacity for increasing NO release by the rise in eNOS activity or the upregulation in protein expression of endothelial cells (Miguel et al., 2020). Our study suggests that EWH was able to increase the NO release possibly by enhancing the eNOS function, which is the major isoform involved in the control of vascular function and blood pressure. The vasodilator power of EWH in SHR models is related to Arg or Tyr peptide content necessary in the N-terminal position for vasodilator activity (Garcia-Redondo et al., 2010).
An alternative mechanism implied on the vasodilator activity of the EWH and derived peptides is its action mediated by NO production via the bradykinin B1 receptor (Miguel et al., 2007). Others authors showed that egg ovotransferrin-derived peptides increased NO-mediated vasodilation in MRA of SHR animals through increasing eNOS expression (Majumder et al., 2013). Future studies are required to elucidate the mechanisms of action by which EWH acts on the expression and activity of eNOS.
In the present study, EWH was able to prevent oxidative stress in MRA of Hg-treated rats, avoiding superoxide anion generation from NAD(P)H oxidase and balancing pro-oxidant and antioxidant status verified by the functional experiments and the oxidative stress biomarkers in vascular tissue. Long-term Hg exposed-rats had lower NOX-4 and p22phox mRNA levels in aorta after co-treatment with EWH; this was related to lower vascular reactivity in conductance vessels (Rizzetti et al., 2017a). A similar effect could be found in MRA in the present study to explain our findings. Previous studies showed that antioxidant EWH properties decreased plasmatic MDA levels and increased the levels of low glutathione in the liver of Zucker obese animals (Garcés-Rimón et al., 2016). Restored plasma antioxidant capacity was found in high-fat/high-dextrose fed rats with metabolic syndrome after EWH intake (Moreno-Fernandez et al., 2018a). Also, normalized oxidative biomarkers were reported in neurological, reproductive and cardiovascular systems from Hg-exposed animals after EWH consumption (Rizzetti et al., 2016a,b, 2017a,b).
In summary, EWH intake promotes protective effects against the endothelial dysfunction in MRA, and consequently, the increase in SBP of long-term HgCl2-exposed rats. EWH benefits could be related to its NO-induced vasodilatation capacity and its antioxidant and anti-inflammatory properties. EWH reduces high ROS generation by NAD(P)H oxidase and, possibly, the activation of inflammatory COX in MRA from Hg-treated rats. Our findings strongly suggest that dietary supplementation with EWH may represent an important strategy for counteracting the effects of cardiotoxicity by pro-oxidant agents such as heavy metals.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
Ethics Statement
The animal study was reviewed and approved by Ethics Committee on Animal Use Experimentation of the Federal University of Pampa, Uruguaiana, Rio Grande do Sul, Brazil (protocol 05/2014).
Author Contributions
DR, FP, DV, MM, and GW conceived and designed the experiments. AE, DR, and JP performed the experiments. AE, DR, FP, and GW analyzed the data. DV, MM, and GW contributed reagents, materials and analysis tools. AE, DR, DV, MM, and GW wrote the paper. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by National Council for Scientific and Technological Development – CNPq [CNPq 307399/2017-6]; Coordenação de Aperfeiçoamento de Pessoal de Nível Superior; Programa Nacional de Cooperação Acadêmica; Pró-reitoria de Pesquisa - Universidade Federal do Pampa [N° 20180615102630]; Fundação de Amparo à Pesquisa do Rio Grande do Sul – FAPERGS/Brazil [PQG:19/2551-0001810-0]; Fundação de Amparo à Pesquisa e Inovação do Espírito Santo – FAPES/CNPq/PRONEX [N° 80598773], and Spanish Goverment (MICINN) [AGL-2017-89213].
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fphys.2020.595767/full#supplementary-material
Abbreviations
ACE, Angiotensin-converting enzyme; ACh, Acetylcholine; BH4, Tetrahydrobiopterin; COX, Cyclooxygenase; DAF, 2-4,5-diaminofluorescein diacetate; dAUC, Areas under the concentration-response curves; DCF, 2', 7'-dichlorofluorescein; DCHF-DA, 2', 7'-dichlorofluorescein diacetate; DTNB, 5,5'-dithio-bis(2-nitrobenzoic acid); EWH, Egg white hydrolysate; Fe2+, Ferrous ion; Fe3+, Ferric ion; FRAP, Ferric Reducing/Antioxidant Power; Hg, Mercury; HgCl2, Mercury chloride; i.m., Intramuscular injections; L-NAME, N-nitro-L-arginine methyl ester; MDA, Malondialdehyde; MRA, Resistance mesenteric arteries; NE, Noradrenaline; NO, Nitric oxide; NOS, Nitric oxide synthase; NPSH, Non-protein thiols; ROS, Reactive oxygen species; RS, Reactive species; SBP, Systolic blood pressure; SDS, Sodiumdodecilsulphate; SHR, Spontaneously hypertensive rats; SNP, Sodium nitroprusside; TBA, Thiobarbituric acid; TPTZ, 2,4,6-Tri(2-piridil)-s-triazina.
References
Antoniades, C., Shirodaria, C., Warrick, N., Cai, S., de Bono, J., Lee, J., et al. (2006). 5-methyltetrahydrofolate rapidly improves endothelial function and decreases superoxide production in human vessels: effects on vascular tetrahydrobiopterin availability and endothelial nitric oxide synthase coupling. Circulation 114, 1193–1201. doi: 10.1161/CIRCULATIONAHA.106.612325
Avendaño, M. S., Lucas, E., Jurado-Pueyo, M., Martinez-Revelles, S., Vila-Bedmar, R., Mayor, F. Jr., et al. (2014). Increased nitric oxide bioavailability in adult GRK2 hemizygous mice protects against angiotensin II-induced hypertension. Hypertension 63, 369–375. doi: 10.1161/HYPERTENSIONAHA.113.01991
Benzie, I. F., and Strain, J. J. (1996). The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: the FRAP assay. Anal. Biochem. 239, 70–76. doi: 10.1006/abio.1996.0292
Botelho, T., Marques, V. B., Simoes, M. R., do Val Lima, P. R., Simoes, F. V., Vassallo, D. V., et al. (2019). Impaired participation of potassium channels and Na(+)/K(+) -ATPase in vasodilatation due to reduced nitric oxide bioavailability in rats exposed to mercury. Basic Clin. Pharmacol. Toxicol. 124, 190–198. doi: 10.1111/bcpt.13113
Chikara, S., Nagaprashantha, L. D., Singhal, J., Horne, D., Awasthi, S., and Singhal, S. S. (2018). Oxidative stress and dietary phytochemicals: role in cancer chemoprevention and treatment. Cancer Lett. 413, 122–134. doi: 10.1016/j.canlet.2017.11.002
Davalos, A., Miguel, M., Bartolome, B., and Lopez-Fandino, R. (2004). Antioxidant activity of peptides derived from egg white proteins by enzymatic hydrolysis. J. Food Prot. 67, 1939–1944. doi: 10.4315/0362-028X-67.9.1939
de Marco, K. C., Antunes, L. M., Tanus-Santos, J. E., and Barbosa, F. Jr. (2012). Intron 4 polymorphism of the endothelial nitric oxide synthase (eNOS) gene is associated with decreased NO production in a mercury-exposed population. Sci. Total Environ. 414, 708–712. doi: 10.1016/j.scitotenv.2011.11.010
Dumitrescu, C., Biondi, R., Xia, Y., Cardounel, A. J., Druhan, L. J., Ambrosio, G., et al. (2007). Myocardial ischemia results in tetrahydrobiopterin (BH4) oxidation with impaired endothelial function ameliorated by BH4. Proc. Natl. Acad. Sci. U. S. A. 104, 15081–15086. doi: 10.1073/pnas.0702986104
Ellman, G. L. (1959). Tissue sulfhydryl groups. Arch. Biochem. Biophys. 82, 70–77. doi: 10.1016/0003-9861(59)90090-6
Faria, T. O., Simoes, M. R., Vassallo, D. V., Forechi, L., Almenara, C. C. P., Marchezini, B. A., et al. (2018). Xanthine oxidase activation modulates the endothelial (vascular) dysfunction related to HgCl2 exposure plus myocardial infarction in rats. Cardiovasc. Toxicol. 18, 161–174. doi: 10.1007/s12012-017-9427-x
Garces-Rimon, M., Gonzalez, C., Hernanz, R., Herradon, E., Martin, A., Palacios, R., et al. (2019). Egg white hydrolysates improve vascular damage in obese zucker rats by its antioxidant properties. J. Food Biochem. 43:e13062. doi: 10.1111/jfbc.13062
Garcés-Rimón, M., Gonzalez, C., Uranga, J. A., Lopez-Miranda, V., Lopez-Fandino, R., and Miguel, M. (2016). Pepsin egg white hydrolysate ameliorates obesity-related oxidative stress, inflammation and steatosis in zucker fatty rats. PLoS One 11:e0151193. doi: 10.1371/journal.pone.0151193
Garcia-Redondo, A. B., Roque, F. R., Miguel, M., Lopez-Fandino, R., and Salaices, M. (2010). Vascular effects of egg white-derived peptides in resistance arteries from rats. Structure-activity relationships. J. Sci. Food Agric. 90, 1988–1993. doi: 10.1002/jsfa.4037
Genchi, G., Sinicropi, M. S., Carocci, A., Lauria, G., and Catalano, A. (2017). Mercury exposure and heart diseases. Int. J. Environ. Res. Public Health 14:74. doi: 10.3390/ijerph14010074
Halliwell, B. (2007). Oxidative stress and cancer: have we moved forward? Biochem. J. 401, 1–11. doi: 10.1042/BJ20061131
Incalza, M. A., D’Oria, R., Natalicchio, A., Perrini, S., Laviola, L., and Giorgino, F. (2018). Oxidative stress and reactive oxygen species in endothelial dysfunction associated with cardiovascular and metabolic diseases. Vascul. Pharmacol. 100, 1–19. doi: 10.1016/j.vph.2017.05.005
Liguori, I., Russo, G., Curcio, F., Bulli, G., Aran, L., Della-Morte, D., et al. (2018). Oxidative stress, aging, and diseases. Clin. Interv. Aging 13, 757–772. doi: 10.2147/CIA.S158513
Liu, F., Ma, C., Gao, Y., and McClements, D. J. (2017). Food grade covalent complexes and their application as nutraceutical delivery systems: a review. Compr. Rev. Food Sci. F. 16, 76–95. doi: 10.1111/1541-4337.12229
Loetchutinat, C., Kothan, S., Dechsupa, S., Meesungnoen, J., Jay-gerin, J. P., and Mankhetkorn, S. (2005). Spectrofluorometric determination of intracellular levels of reactive oxygen species in drug-sensitive and drug-resistant cancer cells using the 2',7'-dichlorofluorescein diacetate assay. Radiat. Phys. Chem. 72, 323–331. doi: 10.1016/j.radphyschem.2004.06.011
Majumder, K., Chakrabarti, S., Morton, J. S., Panahi, S., Kaufman, S., Davidge, S. T., et al. (2013). Egg-derived tri-peptide irw exerts antihypertensive effects in spontaneously hypertensive rats. PLoS One 8:e82829. doi: 10.1371/journal.pone.0082829
Manso, M. A., Miguel, M., Even, J., Hernandez, R., Aleixandre, A., and Lopez-Fandino, R. (2008). Effect of the long-term intake of an egg white hydrolysate on the oxidative status and blood lipid profile of spontaneously hypertensive rats. Food Chem. 109, 361–367. doi: 10.1016/j.foodchem.2007.12.049
Martin, A., Perez-Giron, J. V., Hernanz, R., Palacios, R., Briones, A. M., Fortuno, A., et al. (2012). Peroxisome proliferator-activated receptor-gamma activation reduces cyclooxygenase-2 expression in vascular smooth muscle cells from hypertensive rats by interfering with oxidative stress. J. Hhypertens. 30, 315–326. doi: 10.1097/HJH.0b013e32834f043b
Miguel, M., Lopez-Fandino, R., Ramos, M., and Aleixandre, A. (2005). Short-term effect of egg-white hydrolysate products on the arterial blood pressure of hypertensive rats. Br. J. Nutr. 94, 731–737. doi: 10.1079/BJN20051570
Miguel, M., Lopez-Fandino, R., Ramos, M., and Aleixandre, A. (2006). Long-term intake of egg white hydrolysate attenuates the development of hypertension in spontaneously hypertensive rats. Life Sci. 78, 2960–2966. doi: 10.1016/j.lfs.2005.11.025
Miguel, M., Manso, M., Aleixandre, A., Alonso, M. J., Salaices, M., and Lopez-Fandino, R. (2007). Vascular effects, angiotensin I-converting enzyme (ACE)-inhibitory activity, and antihypertensive properties of peptides derived from egg white. J. Agric. Food Chem. 55, 10615–10621. doi: 10.1021/jf072307o
Miguel, M., Recio, I., Gomez-Ruiz, J. A., Ramos, M., and Lopez-Fandino, R. (2004). Angiotensin I-converting enzyme inhibitory activity of peptides derived from egg white proteins by enzymatic hydrolysis. J. Food Prot. 67, 1914–1920. doi: 10.4315/0362-028X-67.9.1914
Miguel, M., Vassallo, D. V., and Wiggers, G. A. (2020). Bioactive peptides and hydrolysates from egg proteins as a new tool for protection against cardiovascular problems. Curr. Pharm. Des. 26, 3676–3683. doi: 10.2174/1381612826666200327181458
Miner-Williams, W. M., Stevens, B. R., and Moughan, P. J. (2014). Are intact peptides absorbed from the healthy gut in the adult human? Nutr. Res. Rev. 27, 308–329. doi: 10.1017/S0954422414000225
Montezano, A. C., Dulak-Lis, M., Tsiropoulou, S., Harvey, A., Briones, A. M., and Touyz, R. M. (2015). Oxidative stress and human hypertension: vascular mechanisms, biomarkers, and novel therapies. Can. J. Cardiol. 31, 631–641. doi: 10.1016/j.cjca.2015.02.008
Montezano, A. C., and Touyz, R. M. (2014). Reactive oxygen species, vascular Noxs, and hypertension: focus on translational and clinical research. Antioxid. Redox Signal. 20, 164–182. doi: 10.1089/ars.2013.5302
Moreno-Fernandez, S., Garces-Rimon, M., Gonzalez, C., Uranga, J. A., Lopez-Miranda, V., Vera, G., et al. (2018a). Pepsin egg white hydrolysate ameliorates metabolic syndrome in high-fat/high-dextrose fed rats. Food Funct. 9, 78–86. doi: 10.1039/c7fo01280b
Moreno-Fernandez, S., Garcés-Rimón, M., Uranga, J. A., Astier, J., Landrier, J. F., and Miguel, M. (2018b). Expression enhancement in brown adipose tissue of genes related to thermogenesis and mitochondrial dynamics after administration of pepsin egg white hydrolysate. Food Funct. 9, 6599–6607. doi: 10.1039/c8fo01754a
Mulvany, M. J., and Halpern, W. (1977). Contractile properties of small arterial resistance vessels in spontaneously hypertensive and normotensive rats. Circ. Res. 41, 19–26. doi: 10.1161/01.res.41.1.19
Ohkawa, H., Ohishi, N., and Yagi, K. (1979). Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 95, 351–358. doi: 10.1016/0003-2697(79)90738-3
Oparil, S., Acelajado, M. C., Bakris, G. L., Berlowitz, D. R., Cifkova, R., Dominiczak, A. F., et al. (2018). Hypertension. Nat. Rev. Dis. Primers 4:18014. doi: 10.1038/nrdp.2018.14
Peçanha, F. M., Wiggers, G. A., Briones, A. M., Perez-Giron, J. V., Miguel, M., Garcia-Redondo, A. B., et al. (2010). The role of cyclooxygenase (COX)-2 derived prostanoids on vasoconstrictor responses to phenylephrine is increased by exposure to low mercury concentration. J. Physiol. Pharmacol. 61, 29–36.
Peluso, I., and Serafini, M. (2017). Antioxidants from black and green tea: from dietary modulation of oxidative stress to pharmacological mechanisms. Br. J. Pharmacol. 174, 1195–1208. doi: 10.1111/bph.13649
Rice, D. C. (2004). The US EPA reference dose for methylmercury: sources of uncertainty. Environ. Res. 95, 406–413. doi: 10.1016/j.envres.2003.08.013
Rizzetti, D. A., Altermann, C. D., Martinez, C. S., Pecanha, F. M., Vassallo, D. V., Uranga-Ocio, J. A., et al. (2016a). Ameliorative effects of egg white hydrolysate on recognition memory impairments associated with chronic exposure to low mercury concentration. Neurochem. Int. 101, 30–37. doi: 10.1016/j.neuint.2016.10.002
Rizzetti, D. A., Fernandez, F., Moreno, S., Uranga Ocio, J. A., Pecanha, F. M., Vera, G., et al. (2016b). Egg white hydrolysate promotes neuroprotection for neuropathic disorders induced by chronic exposure to low concentrations of mercury. Brain Res. 1646, 482–489. doi: 10.1016/j.brainres.2016.06.037
Rizzetti, D. A., Martin, A., Corrales, P., Fernandez, F., Simoes, M. R., Pecanha, F. M., et al. (2017a). Egg white-derived peptides prevent cardiovascular disorders induced by mercury in rats: role of angiotensin-converting enzyme (ACE) and NADPH oxidase. Toxicol. Lett. 281, 158–174. doi: 10.1016/j.toxlet.2017.10.001
Rizzetti, D. A., Martinez, C. S., Escobar, A. G., da Silva, T. M., Uranga-Ocio, J. A., Pecanha, F. M., et al. (2017b). Egg white-derived peptides prevent male reproductive dysfunction induced by mercury in rats. Food Chem. Ttoxicol. 100, 253–264. doi: 10.1016/j.fct.2016.12.038
Rizzetti, D. A., Torres, J. G., Escobar, A. G., da Silva, T. M., Moraes, P. Z., Hernanz, R., et al. (2017c). The cessation of the long-term exposure to low doses of mercury ameliorates the increase in systolic blood pressure and vascular damage in rats. Environ. Res. 155, 182–192. doi: 10.1016/j.envres.2017.02.022
Serafini, M., and Peluso, I. (2016). Functional foods for health: the interrelated antioxidant and anti-inflammatory role of fruits, vegetables, herbs, spices and cocoa in humans. Curr. Pharm. Des. 22, 6701–6715. doi: 10.2174/1381612823666161123094235
Su, L., Wang, M., Yin, S. T., Wang, H. L., Chen, L., Sun, L. G., et al. (2008). The interaction of selenium and mercury in the accumulations and oxidative stress of rat tissues. Ecotoxicol. Environ. Saf. 70, 483–489. doi: 10.1016/j.ecoenv.2007.05.018
Szasz, T., Thakali, K., Fink, G. D., and Watts, S. W. (2007). A comparison of arteries and veins in oxidative stress: producers, destroyers, function, and disease. Exp. Biol. Med. 232, 27–37.
Touyz, R. M., and Briones, A. M. (2011). Reactive oxygen species and vascular biology: implications in human hypertension. Hypertens. Res. 34, 5–14. doi: 10.1038/hr.2010.201
Valko, M., Rhodes, C. J., Moncol, J., Izakovic, M., and Mazur, M. (2006). Free radicals, metals and antioxidants in oxidative stress-induced cancer. Chem. Biol. Interact. 160, 1–40. doi: 10.1016/j.cbi.2005.12.009
Vassallo, D. V., Moreira, C. M., Oliveira, E. M., Bertollo, D. M., and Veloso, T. C. (1999). Effects of mercury on the isolated heart muscle are prevented by DTT and cysteine. Toxicol. Appl. Pharmacol. 156, 113–118. doi: 10.1006/taap.1999.8636
Virtanen, J. K., Rissanen, T. H., Voutilainen, S., and Tuomainen, T. P. (2007). Mercury as a risk factor for cardiovascular diseases. J. Nutr. Biochem. 18, 75–85. doi: 10.1016/j.jnutbio.2006.05.001
Wiggers, G. A., Pecanha, F. M., Briones, A. M., Perez-Giron, J. V., Miguel, M., Vassallo, D. V., et al. (2008). Low mercury concentrations cause oxidative stress and endothelial dysfunction in conductance and resistance arteries. Am. J. Physiol. Heart Circ. Physiol. 295, H1033–H1043. doi: 10.1152/ajpheart.00430.2008
Keywords: egg white hydrolysate, antioxidant properties, mercury, mesenteric resistance arteries, vascular damage, nitric oxide, oxidative stress
Citation: Escobar AG, Rizzetti DA, Piagette JT, Peçanha FM, Vassallo DV, Miguel M and Wiggers GA (2020) Antioxidant Properties of Egg White Hydrolysate Prevent Mercury-Induced Vascular Damage in Resistance Arteries. Front. Physiol. 11:595767. doi: 10.3389/fphys.2020.595767
Edited by:
Ana Paula Davel, Campinas State University, BrazilReviewed by:
Stêfany Cau, Federal University of Minas Gerais, BrazilEliana Hiromi Akamine, University of São Paulo, Brazil
Copyright © 2020 Escobar, Rizzetti, Piagette, Peçanha, Vassallo, Miguel and Wiggers. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Giulia Alessandra Wiggers, Z2l1bGlhd3BAZ21haWwuY29t; Z2l1bGlhcGVjYW5oYUB1bmlwYW1wYS5lZHUuYnI=
†Present address: Danize Aparecida Rizzetti, Polytechnic School, Federal University of Santa Maria, Santa Maria, Brazil